the journal of vol. 263, 23, 15, pp. 11414-11420,1988 1988 ... · the journal of biological...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1988 by The American Society for Bioehemistry and Molecular Biology, Inc.
Vol. 263, No. 23, Issue of August 15, pp. 11414-11420,1988 Printed in U. S. A.
C-terminal-specific Monoclonal Antibodies against the Human Red Cell Glucose Transporter EPITOPE LOCALIZATION WITH SYNTHETIC PEPTIDES*
(Received for publication, February 9, 1988)
Lars Andersson and Per Lundahl From the Department of Biochemistry, Biomedical Center, University of Uppsala, P. 0. Box 576, S-75123 Uppsalu, Sweden
Mice were immunized with human red cell glucose transporter for production of monoclonal antibodies. Four peptides were synthesized that correspond to rel- atively hydrophilic segments of the human HepG2 glu- cose transporter (Mueckler, M., Caruso, C., Baldwin, S. A., Panico, M., Blench, I., Morris, H. R., Allard, W. J., Lienhard, G. E., and Lodish, H. F. (1986) Science 229,941-945), including a C-terminal segment. After identification of hybridomas that were positive for the red cell glucose transporter, enzyme-linked immuno- sorbent assays were done with the synthetic peptides in solution to detect peptide-binding monoclonals. The very hydrophilic C-terminal peptide 478-492 (P2), but no other peptide, gave strong and selective inhibi- tion of antibody binding to the glucose transporter. Two C-terminal-specific monoclonal antibodies were selected. The binding of these two antibodies to im- mobilized inside-out vesicles of human red cell mem- branes could be inhibited with the peptide P2. The antibodies did not react with right-side-out vesicles. The binding of these C-terminal-specific antibodies to the glucose transporter, to immobilized vesicles, and to the peptide P2 was enhanced by the presence of the peptide 218-232 (Pl), although the peptide P1 alone showed no reaction with these antibodies. This sug- gests that the C terminus and the segment 218-232 of the red cell glucose transporter are exposed at the cytoplasmic face of the membrane and interact in the transporter.
The C-terminal-specific monoclonal antibodies re- acted strongly in Western blotting with the human red cell glucose transporter.
Most mammalian cells use glucose (1). The glucose trans- port into cells is mediated by integral membrane proteins. The human red cell glucose transporter is one of the most studied facilitated diffusion transport proteins (2-4). Second to the anion transporter it is the most abundant transport protein of red cell membranes, with around 10‘ copies per cell (5, 6). The glucose transporter is a glycoprotein, probably with a single, heterogeneous, N-linked oligosaccharide (7, 8)
* This work was supported by the Swedish Natural Science Re- search Council, the 0. E. and Edla Johansson Science Foundation, the Sven and Lilly Lawski Science Foundation, and by a grant to Jan Andersson from the Swedish Technical Board of Development. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
and shows upon SDS’ gel electrophoresis a very broad band with the average apparent M, 55,000. Treatment with endo- P-galactosidase or endoglycosidase F sharpens the band and reduces the apparent M, to 46,000 (9). The glucose transporter co-purifies with small amounts of nucleoside transporter (10, l l ) , which is found in approximately lo4 copies per cell (5,6).
In 1985, Mueckler and co-workers (7) presented the cDNA- deduced amino acid sequence of a glucose transporter from the human hepatoma cell line HepG2. Later, Birnbaum and co-workers (12) deduced the sequence of a rat brain glucose transporter from cDNA. The two sequences are nearly iden- tical. The HepG2 sequence is probably very similar, if not identical, to the sequence of the human red cell glucose transporter (7,13). Mueckler and co-workers (7) also proposed a model for the orientation in the membrane of the 492- residue polypeptide chain, M, 54,117, of the HepG2 trans- porter.
In recent years the use of synthetic peptides has facilitated the localization of epitopes (antigenic determinants) of pro- teins. Synthetic peptides representative of virtually any given part of the surface of a protein can elicit the production of antibodies that react with the protein (14, 15). Synthetic peptides can also be used in immunobinding assays to deter- mine the specificities of monoclonal antibodies obtained from mice immunized with the corresponding protein (16, 17).
Peptide-specific antibodies can be used to study the surface structure of membrane proteins of known sequences. In the present work we have used synthetic peptides for analysis of the location of binding epitopes for monoclonal antibodies against the glucose transporter from human red cells. Davies and co-workers (18) have recently reported results obtained by a different approach with antisera from rabbits immunized with N- and C-terminal peptides of the HepG2 glucose trans- porter.
MATERIALS AND METHODS
Human red cell concentrate (stored 4-5 weeks) was supplied by the Blood Bank at the University Hospital, Uppsala, Sweden. Pig blood was collected, citrated, and used within 1 day. Bovine serum albumin, n-octyl-6-D-glucopyranoside (octyl glucoside), Tris (Trizma base), ethylmercurithiosalicylate (thimerosal), and Tween 20 were obtained from Sigma. Glutardialdehyde (25%, “for electronmicros- copy”) and SDS (90% detergent, 10% inorganic salts) were purchased from Merck. DEAE-cellulose (DE52) was bought from Whatman. Sephadex G-50M, Sepharose CL-4B and CL-GB, SPDP, and a Su-
The abbreviations used are: SDS, sodium dodecyl sulfate; BSA, bovine serum albumin; mAb, monoclonal antibody; SPDP, N-SUC- cinimidyl-3-(2-pyridyldithio)propionate;DMEM,Dulbecco’smodified Eagle’s medium; ELISA, enzyme-linked immunosorbent assay; PAGE, polyacrylamide gel electrophoresis; FCS, fetal calf serum; HAT, hypoxanthine, aminopterin, and thymidine; 1, liter; HEPES, N-2-hydroxyethylpiperazine-N‘-2-ethanesulfonic acid.
11414

Glucose Transporter Epitope Localization 11415
perose 6 column were supplied by Pharmacia LKB Biotechnology Inc. Freund's adjuvants were obtained from Difco Laboratories and poly(ethyleneglyco1) 1,500 from Boehringer Mannheim. ELISA flat- bottomed microtiter plates (Immunoplate I) were obtained from Nunc, Denmark. Tetramethylbenzidine was bought from Fluka, Swit- zerland and HRP Color Development Reagent (4-chloro-1-naphthol) and prestained SDS-PAGE immunoblotting standard proteins from Bio-Rad. Peroxidase-conjugated goat anti-mouse IgG (H+L) anti- serum was purchased from Jackson Immunoresearch Laboratories, West Grove, PA (affinity-purified). Nitrocellulose membranes were type BA 45, 0.45 pm, from Schleicher & Schull, Federal Republic of Germany.
DMEM was purchased from GIBCO and was always supplemented with 10 mM HEPES buffer (pH 7.0 in 2 M stock solution), 2 mM L- glutamine, 0.05 mM P-mercaptoethanol, 120 mg of penicillin sulfate/ 1, and 100 mg of streptomycin sulfate/l (final concentrations). Hy- poxanthine, aminopterin, and thymidine were also bought from GIBCO and were used at 14,0.18, and 3.9 mg/l, respectively, in HAT- DMEM and hypoxanthine-thymidine-DMEM.
For plasma desorption mass spectrometry (19), a Bion 20 instru- ment from Bio-Ion Nordic AB, Uppsala, Sweden, was used. ELISA plate washings were done with a Microwash 2 automatic plate washer from Skatron A/S, Lier, Norway, and ELISA absorbances were measured with a Flow Laboratories (United Kingdom) Titertek Mul- tiskan MC photometer.
Preparation of Integral Membrane Proteins-Integral membrane proteins from red cells were prepared at 6 "C by gel filtration of hemolysate on 5-1 Sepharose CL-4B and CL-GB columns in tandem and subsequent centrifugations (11). Briefly, 200 ml of red cells were hemolysed in 800 ml of 5 mM sodium phosphate buffer (pH 8.0) and applied to the first column in the same buffer. The ghost fraction was transferred to the second column, which had been equilibrated with 5 mM Na2EDTA-NaOH (pH 10.5). The membranes were collected by centrifugation and washed twice with 2 mM Naz EDTA, 15 mM NaOH, 0.2 mM dithioerythritol (pH 12), to remove residual peripheral mem- brane proteins, and once with 50 mM Tris-HC1 (pH 6.8). The material was then suspended in the latter buffer, frozen dropwise in liquid nitrogen, and stored at -70 "C. Integral membrane proteins from pig red cells were prepared as above. Pig and sheep red cell ghosts were prepared in essentially the same way, at pH 8. The sheep and pig red cell membrane preparations were used in immunoblotting only.
Preparation of Glucose Transporter-The red cell glucose trans- porter was purified by anion exchange chromatography (cf. Ref. 20) followed by medium pressure gel filtration chromatography (cf. Refs. 11 and 21). 100 mg of integral membrane protein were vigorously stirred at 2 "C in 70 mM Tris-HC1 (pH 7.0 as measured at 22 "C), 2 mM dithioerythritol, and 50 mM octyl glucoside at a protein concen- tration of 4 g/l and centrifuged at 4 "C for 60 min at 160,000 X g. The glucose transporter was purified at 6 'C by chromatography of the solubilized material on a 13 X 2-cm DEAE-cellulose column in Tris- HCl and octyl glucoside as above with 1 mM dithioerythritol. The detergent was then removed at 6 "C on a 41 X 2-cm Sephadex G50 column in 50 mM Tris-HC1,lOO mM NaCl, and 1 mM Na2EDTA (pH 7.0 at 22 "C). The transporter-lipid vesicles that were formed were sedimented at 160,000 X g for 2 h a t 4 "C and frozen. The next day the material was solubilized in a volume of 0.8 ml in 50 mM Tris-HC1 (pH 7.0 at 22 "C), 400 mM NaCl, 0.75 mM dithioerythritol, and 310 mM octyl glucoside (final concentrations) at 6 "C. The clear solution was centrifuged at 14,500 X g for 20 min at 6 "C and fractionated on a 30 X 1.0-cm Superose 6 small-bead gel filtration column in 70 mM Tris-HC1 (pH 7.0), 500 mM NaCl, 1 mM dithioerythritol, and 50 mM octyl glucoside at 22 "C at a flow rate of 9 ml/h. The glucose trans- porter eluted largely as aggregates (cf. Ref. 21). Fractions were ana- lysed by SDS-PAGE. The main transporter fractions were combined and stored at -70 "C for immunization. The protein concentration was approximately 350 pglml. A second batch, 200 pg of protein/ml, was prepared from 50 mg of integral membrane proteins for ELISA. SDS-PAGE analyses are shown in Fig. 2.
with 50 pl of 4% SDS, 20 mM dithioerythritol, 82 mM Tris, 80 mM Electrophoresis and Siluer Staining-50-pl samples were mixed
borate, 10 mM Na2EDTA, and 10% sucrose (pH 8.6) in conical Eppendorff tubes. The tubes were sealed and immersed in boiling water for 5 min. 10 pl of 0.4 M iodoacetic acid were added at 22 "C. The material was centrifuged and 100-pl samples were applied to the gel. Electrophoresis was run for 12 h at 75 V with the discontinuous buffer system described by Neville (22) in 15 X 15 X 0.15 cm gels including a 2.5-em-high stacking gel. The separation gel concentra- tion was T = 11%, C = 1%.
The gels were silver-stained by a procedure similar to the one described in Ref. 23 (cf. Refs. 13 and 24): The gels were immersed and shaken in 400 ml of 40% methanol/lO% acetic acid (40 m i d , 2 x 1 1 of water (2 X 5 min), 300 ml of 12% glutardialdehyde (practical grade) for 15 min, 4 X 1 1 of water (4 X 10 min), 2 X 500 ml of 20% ethanol (2 X 15 min), and, in subdued light, 200 ml of an alkaline ammonium/silver nitrate solution. This solution was prepared as follows: 400 mg of AgN03 were dissolved in 2 ml of water, mixed with 2 ml of 25% ammonia and then with 400 mg of NaOH in 10 ml of water, and finally diluted with 20% ethanol to a volume of 200 ml. After 15 min in this solution the gels were rinsed 2 X 5 min in 2 X 500 ml of 20% ethanol and developed for 3-5 min with 200 pl of formaldehyde and 50 p1 of 2.3 M citric acid in 200 ml of 20% ethanol. The developer was removed and 300 ml of 20% ethanol with 1% acetic acid and 150 pl of ethanolamine were added. After 1 h in this solution the gels were shrunk at 4 "C in 20% ethanol overnight and dried between cellophane sheets (25).
Peptides-Four polypeptide segments, predicted or presumed not to be embedded in the membrane (Fig. l), were selected from the sequence of the glucose transporter from human HepG2 hepatoma cells (7). The corresponding peptides were synthesized at the Depart- ment of Immunology, Biomedical Center, Uppsala, by solid phase methodology (26), using tertiary butyloxycarbonyl amino acids in the Applied Biosystems model 430A peptide synthesizer. A cysteine res- idue was included at each C terminus to facilitate conjugation of the peptide with a carrier. A tyrosine residue was included in the two peptides that lacked tyrosine to allow their detection by UV absor- bance. The peptides were cleaved from the support with hydrofluoric acid containing 15% anisole and 3% methylethylsulfide and were then washed with ether.
Plasma desorption mass spectrometry (19) of each peptide showed peaks corresponding to the ions [peptide+H]+ and [peptide+Na]+ with the correct masses. Analytical reversed-phase high performance liquid chromatography indicated a peptide purity of 80% or more, sufficient for our purposes.
According to the model of Mueckler and co-workers (7) for the membrane orientation of the glucose transporter (Fig. l), the P1 and P2 sequences are located on the cytoplasmic side of the membrane and presumably on protruding protein parts. The P2 sequence is the C terminus. The P4 and P5 segments are located at the outside of the membrane. P4 is situated close to the proposed glycosylation site and P5 is a short helix-connecting segment (Fig. 1). A fifth peptide, P3, corresponding to the segment 421-430, showed low solubility and was used only in preliminary experiments.
Conjugation of Peptides-The peptides were conjugated to BSA by use of SPDP (27). BSA was activated with SPDP, and the BSA- pyridyldisulfide derivatives formed were reacted at pH 7.5 with the cysteine thiols of the peptides with a 5-lo-fold molar excess of peptides to BSA. 8-Mercaptoethanol was then added to a concentra- tion of 0.5 mM to inactivate nonreacted pyridyldisulfides. SDS-PAGE with BSA and activated BSA as controls showed that nearly all BSA molecules were conjugated to some extent.
Immunization, Cell Fusion, and Hybridoma Selection-3-Month- old female BALB/c mice were immunized intraperitoneally with a mixture of 150 pl of the glucose transporter preparation (50 pg of protein) and 150 p1 of Freund's complete adjuvant, per mouse. After 5 weeks the mice were bled from their tails and the immune responses to the glucose transporter were checked by ELISA. Booster doses of 70 pg of protein from the same transporter preparation as above, with Freund's incomplete adjuvant, were given intraperitoneally. 4 days later one of the mice was killed. The spleen was removed and gently homogenized. Spleen cells were washed and fused with SpZ/O-Ag 14 nonsecreting, HAT-sensitive mouse myeloma cells (28).
For fusion, 90 X lo6 spleen cells were mixed with 35 X lo6 myeloma cells and the cells were sedimented. The tube with the cell pellet was gently tapped while 1 ml of poly(ethyleneglyco1) solution, prewarmed to 37 "C, was added dropwise during 1 min. 50 ml of warm (37 "C) serum-free DMEM was then added, the first 10 ml during 5 min. The cells were gently suspended, kept a t 37 'C for 10 min, and then sedimented and resuspended in 360 ml of DMEM with 10% FCS and fresh rat thymus feeder cells (5 X 106/ml) (11). 290 ml of this suspension were spread on 30 96-well culture plates, 100 p1 per well. This should give few hybridomas per well and thus reduce competition between clones.
The next day HAT-DMEM with 10% FCS was added for selection of fused cells, and hybridomas were grown with regular half-volume replacements of this medium. Cultures, which showed strong antibody binding to the glucose transporter in ELISA, were expanded into

11416 Glucose Transporter Epitope Localization
A OUTSIDE
P5
FIG. 1. Location and sequence of selected segments. A , location of the segments 218-232, 478-492, 51-64, and 292-307 in the model of the HepG2 glu- cose transporter proposed by Mueckler and co-workers (7). These segments were chosen for analysis of the location of binding epitopes for monoclonal anti- bodies against the glucose transporter from human red cells. B , sequences of the synthesized peptides P1, P2, P4, and P5, corresponding to the above seg- ments. A hydropathy plot indicates that P1 and P2 are the most hydrophilic among these peptides. Tyr-Cys or Cys alone was added at the C terminus to simplify detection and conjugation.
1 N
0 P1
6 P1 Arg-Asn-Glu-Glu-Asn-Arg-Ala-Lys-Ser-VaI-Leu-Lys-Lys-Leu-Arg-Tyr-Cys
21 8 232
P2 Thr-Pro-Glu-Glu-Leu-Phe-His-Pro-Leu-Gly-Ala-Asp-Ser-GIn-VaI-Tyr-Cys 478 492
P4 Arg-Tyr-Gly-Glu-Ser-l le-Leu-Pro-Thr-Thr-Leu-Thr-Thr-Leu-Cys 51 64
P5 Tvr-Tvr-Ser-Thr-Ser-l le-Phe-Glu-Lvs-Ala-Glv-Val-Gin-Gln-Pro-Val-Cvs 292 .
1-ml wells in hypoxanthine-thymidine-DMEM with 10% FCS. Cell samples were frozen. The cultures were further expanded in 10-ml volumes in DMEM with 10% FCS. Cell supernatants were made 0.1% in sodium azide and stored at 4 "C.
ELISA-All procedures were carried out at room temperature. Antigens (glucose transporter or peptides) were diluted with 50 mM fresh Tris-HC1 (pH 9.5) (buffer C). A 50-pl aliquot was added per well and the plates were left overnight. The coating solution was flicked out and 150 p1 of blocking solution were added (100 mM sodium phosphate (pH 6.8) with 10 g of BSA/l and 0.25 g of thimer- osal/l). After 2 h, the plates were washed four times with deionized water containing 0.05% Tween 20 in an automatic plate washer. Cell culture supernatants were supplemented with 0.1 volume of 0.5% Tween 20 in buffer D (137 mM NaCl, 3 mM KC1,8.4 mM Na2HPO4, and 1.6 mM KHzPOa (pH 7.4)) or diluted with solution E (100 mM sodium phosphate (pH 6.8), 5 g of BSA/l, 0.25 g of thimerosal/l, and 0.05% Tween 20). Samples were added to the plates (50 pl/well) and the plates were incubated for 2 h. In the inhibition experiments, antibody-containing samples were mixed with competing antigen either directly in the wells of the ELISA plates (see Fig. 4) or in a separate preincubation step before addition to the ELISA plates (see Fig. 3). After four washings, 50 pl of peroxidase-conjugated goat anti- mouse IgG antiserum, diluted 5000 times with solution E, was added to each well. After 1 h the plates were washed four times. For color development, 10 volumes of 2 mM Hz02 in 100 mM citric acid-KOH (pH 4.25) was mixed with 1 volume of 20 mM tetramethylbenzidine in methanol/acetone (9:l) and 200 p1 of this mixture were added to each well. When an appropriate color intensity had developed (usually in less than 10 min), the peroxidase reaction was interrupted by the addition of 100 pl of 1 M sulfuric acid per well. The absorbances at 450 nm were read immediately, since the color intensity increased slowly. The above HzOz and tetramethylbenzidine solutions could be used for several weeks when stored in the dark.
Membrane Vesicles-Human red cell membrane vesicles of differ- ent "sidedness" were prepared and assayed for sidedness as described by Steck and Kant (29) and Steck (30). Acetylcholinesterase and glyceraldehyde-3-phosphate dehydrogenase were used as markers of exterior and interior membrane surfaces, respectively. The protein concentration was determined by light absorbance measurements at 280 and 310 nm after solubilization of the vesicles with SDS (31). The vesicles were stored at 4 "C and were used within 10 days.
The ability of the two kinds of membrane vesicles to adsorb antibodies was determined by measuring the extent of antibody binding to the glucose transporter in ELISA after treating diluted
307
hybridoma cell culture supernatants with vesicles in suspension. Details are given in the legend to Fig. 5.
Immobilized inside-out vesicles were also used as antigen in ELISA (cf. Ref. 32): Suspensions of vesicles were added to ELISA plate wells and the plates were centrifuged for 5 min at 1500 X g. Without emptying the wells, the plates were slowly immersed in cold buffer D containing 0.25% glutardialdehyde. After 10 min the wells were flicked empty, rinsed three times with cold buffer D, blocked with BSA, and used in competitive ELISA. Details are given in the legend to Fig. 4.
Protein Zmmunoblotting-Immunoblotting (Western blotting) was done at room temperature. Human and pig red cell integral membrane proteins as well as pig and sheep red cell ghost proteins were separated by gel electrophoresis in SDS (see above). The proteins were then transferred to nitrocellulose membranes for 1 h at 100 V (14 V/cm) followed by 2 h at 200 V, in a water-cooled Bio-Rad Trans-Blot cell a t 20-30 "C. The transfer solution was 20% methanol, 192 mM glycine, and 50 mM Tris (pH 8.6). Further protein binding to the nitrocellulose membranes was blocked by treatment with 0.1% Tween 20 in buffer D for 2 h. The membranes were then incubated overnight with hybridoma cell culture supernatants diluted with buffer D con- taining 0.05% Tween 20 and 0.1% sodium azide. Finally, the mem- branes were washed 6 X 6 min with buffer D, incubated for 2 h with peroxidase-conjugated goat anti-mouse IgG antiserum in solution E, washed as above, and immersed in a freshly prepared substrate solution: HRP Color Development Reagent, dissolved to 3 g/1 in methanol at 2 "C and mixed with 5 volumes of 6 mM HzOz in buffer D. The color development was interrupted by thorough rinsing with water. The incubations were done with gentle tiltings in a cuvette formed by two horizontal glass plates kept 1 mm apart by two plastic spacers (11).
RESULTS
Glucose Transporter Purity-Electrophoretic analyses of the two glucose transporter preparations that were used for immunization and ELISA showed a high degree of purity (Fig. 2). The transporter occupied a broad zone of apparent M, 38,000-63,000. Lanes with large amounts of transporter showed a minor component of M, 30,000 in the preparation used for ELISA, but not in the material used for immuniza- tion. The tendency of the transporter to aggregate made it difficult to detect high", contaminants, although we used a

Glucose Transporter Epitope Localization 11417
Mr
XI o - ~ 94 - 66 - 60 - 44 - 36 - 29-
20 -
1 2 3 4 5 6 7 8 91011
FIG. 2. Electrophoretic analysis of purified glucose trans- porter and of human red cell integral membrane proteins. The analysis was done as described under “Materials and Methods.” The detergent used was sodium lauryl sulfate (Sigma) containing 68% C,,, 25% CI4, and 6% C16 sulfate. Lane I, calibration proteins: phospho- rylase b (M, 94,000), BSA (66,000), ovalbumin (44,000), carbonic anhydrase (29,000), and soybean trypsin inhibitor (20,000). Lanes 2- 5, glucose transporter preparation used for immunization, approxi- mately 0.6, 4, 4, and 9.5 pg, respectively. Lane 6, human red cell integral membrane proteins, approximately 10 pg. Lane 7, calibration proteins: BSA (66,000), catalase (60,000), and lactate dehydrogenase (36,000). Lanes 8-11, glucose transporter preparation used for ELISA, protein amounts as in lanes 2-5. The samples in lanes 4 and 10 were not treated with dithioerythritol and not heated.
mixture of CI2, CI4, and CI6 alkyl sulfates, which reduces the aggregation (20). In samples not treated with dithioerythritol and not heated, the glucose transporter, monomer as well as dimer, shows a somewhat higher M , than in reduced and heated samples (Fig. 2).
Selection of Hybridomas-After fusion and selection in HAT medium, 70% of the wells in the 30-cell culture plates showed cell growth, which indicates an average of 1.2 hybrid- oma clones per well (Poisson distribution). 315 of the wells that showed cell growth were tested for antibody binding to the glucose transporter. 69 of these were positive. Conse- quently, few wells should have contained more than one transporter-positive clone.
When peptides are added to ELISA plate wells for coating, the extent of peptide adsorption is not easy to determine and, furthermore, the peptides may be adsorbed in such a way that the binding of certain antibodies is prevented. Therefore, we tested all the transporter-positive cultures not only by direct ELISA screening with peptides Pl-P5,1 pg/well, but also by direct ELISA screening with peptides conjugated to BSA, and we also tested selected cultures by competitive ELISA with peptides in solution and transporter-coated wells for the de- termination of antibody selectivity for the peptides. Some supernatants were positive for P1, P4, or P5, but showed unsatisfactory selectivity or weak reactions. Another six su- pernatants showed strong and selective binding to the C- terminal peptide P2. Cells from two of these P2-positive cultures were thawed and cloned by limiting dilution. The new cultures were screened with transporter and P2 as above. Selected clones were tested by direct and competitive assays with peptides P1, P2, P4, and P5. Two P2-positive clones, B82:4 and B315:32, were expanded and tested by direct ELISA and by competitive ELISAs with (a) adsorbed glucose trans- porter competing with free P1, P2, P4, and P5 (see Fig. 3) and (b) glucose transporter in immobilized inside-out vesicles of red cell membranes, competing with free P1 and P2 (see Fig. 4). In addition (c), adsorption of antibodies to inside-out and right-side-out vesicles in suspension was assayed by sim-
ilar procedures (see Fig. 5). Cell culture supernatants, denoted mAb B82:4 and mAb B315:32 were used in these experiments. The mAbs were not purified. The peroxidase-conjugated anti- serum showed no affinity for transporter or peptides. The results and interpretations of the experiments a-c are pre- sented below.
Experiment a-Competitive assays with wells coated with glucose transporter and peptides P1, P2, P4, and P5 as com- peting antigens showed that P2 inhibited binding of mAbs B82:4 and B315:32 to the transporter (Fig. 3). P4 and P5 gave no or only slight inhibition. The largest amount of peptide used, 40 pg/well, corresponds to a 20,000-fold molar excess of peptide over the amount of transporter applied to each well. Unexpectedly, P1 increased the binding of the antibodies (Fig. 3). This enhancing effect of P1 was also seen in competitive assays when we used wells coated with P2 or wells with immobilized inside-out vesicles (not shown). The enhance- ment of antibody binding by P1 may be explained by hypoth- esis (i) or (ii) below.
(i) P1 binds to the ELISA plate plastic when added together with the mAbs, or binds to the blocking BSA. The mAbs or the peroxidase-conjugated antibodies then bind specifically or nonspecifically to P1 in addition to the binding of antibodies caused by the transporter.
(ii) P1 interacts with P2 or with the P2 segment of the transporter in a way that causes some P2 peptides or P2 segments, respectively, to adopt a conformation or confor- mations suitable for binding of the mAbs or, in Pl-P2 com- plexes, P1 possibly participates directly in the binding to the antibody.
To verify or disprove these hypotheses we have used addi- tional ELISA variants, as described below. The results favor hypothesis ii. Direct ELISA showed no binding to P1 or to conjugated P1, which indicates that the antibodies do not react, or react weakly, with P1 alone. Assays as in Fig. 3, but without transporter, showed no enhancement of the signal in the presence of P1. Therefore, binding of P1 to the ELISA
,b* 0.04 0.4 4 40
AMOUNT OF PEPTIDE (pg)
\ 0.04 0.4 4 40
AMOUNT OF PEPTIDE (pg)
FIG. 3. Competitive ELISA with wells coated with glucose transporter and peptides in solution as competing antigens. The monoclonal antibody (hybridoma cell supernatant) was B82:4, diluted 300-fold ( A ) and B315:32, diluted 6,000-fold (B) . The peptide (Pl, P2, P4, or P5) was dissolved in buffer C , mixed with the hybridoma cell culture supernatant diluted with solution E, and incubated for 2 h a t 22 ‘C before transfer of a 50-p1 sample containing the given amount of peptide to a transporter-coated well in an ELISA plate to test antibody binding. The amount of the glucose transporter preparation applied for coating was 50 ng/well. The ELISA signal of samples treated as above, but without peptide, was 100%. The color development time was 3 min. The number of the peptide is indicated at the right of each curve.
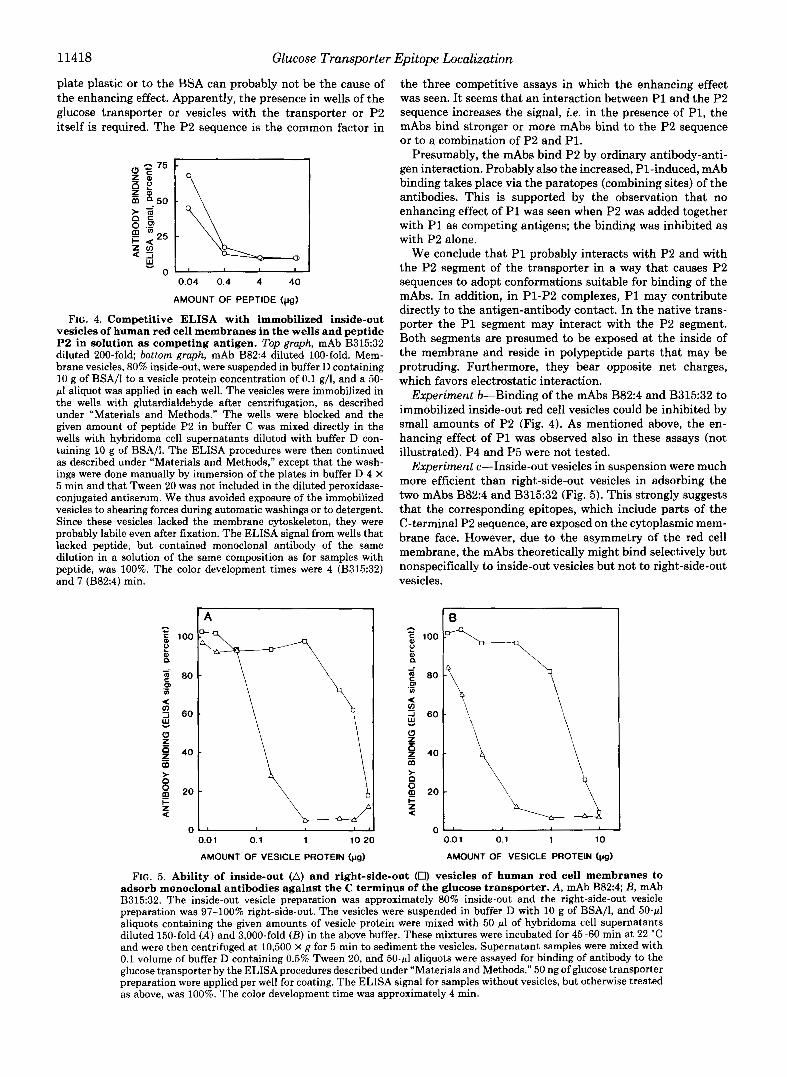
11418 Glucose Transporter Epitope Localization
plate plastic or to the BSA can probably not be the cause of the enhancing effect. Apparently, the presence in wells of the glucose transporter or vesicles with the transporter or P2 itself is required. The P2 sequence is the common factor in
0.04 0.4 4 40
AMOUNT OF PEPTIDE (pg)
FIG. 4. Competitive ELISA with immobilized inside-out vesicles of human red cell membranes in the wells and peptide P2 in solution as competing antigen. Top graph, mAb B315:32 diluted 200-fold bottom graph, mAb B824 diluted 100-fold. Mem- brane vesicles, 80% inside-out, were suspended in buffer D containing 10 g of BSA/l to a vesicle protein concentration of 0.1 g/l, and a 50- pl aliquot was applied in each well. The vesicles were immobilized in the wells with glutardialdehyde after centrifugation, as described under “Materials and Methods.” The wells were blocked and the given amount of peptide P2 in buffer C was mixed directly in the wells with hybridoma cell supernatants diluted with buffer D con- taining 10 g of BSA/I. The ELSA procedures were then continued as described under “Materials and Methods,” except that the wash- ings were done manually by immersion of the plates in buffer D 4 X 5 min and that Tween 20 was not included in the diluted peroxidase- conjugated antiserum. We thus avoided exposure of the immobilized vesicles to shearing forces during automatic washings or to detergent. Since these vesicles lacked the membrane cytoskeleton, they were probably labile even after fixation. The ELISA signal from wells that lacked peptide, but contained monoclonal antibody of the same dilution in a solution of the same composition as for samples with peptide, was 100%. The color development times were 4 (B315:32) and 7 (B82:4) min.
the three competitive assays in which the enhancing effect was seen. It seems that an interaction between P1 and the P2 sequence increases the signal, i.e. in the presence of P1, the mAbs bind stronger or more mAbs bind to the P2 sequence or to a combination of P2 and PI.
Presumably, the mAbs bind P2 by ordinary antibody-anti- gen interaction. Probably also the increased, P1-induced, mAb binding takes place via the paratopes (combining sites) of the antibodies. This is supported by the observation that no enhancing effect of P1 was seen when P2 was added together with P1 as competing antigens; the binding was inhibited as with P2 alone.
We conclude that P1 probably interacts with P2 and with the P2 segment of the transporter in a way that causes P2 sequences to adopt conformations suitable for binding of the mAbs. In addition, in P1-P2 complexes, P1 may contribute directly to the antigen-antibody contact. In the native trans- porter the P1 segment may interact with the P2 segment. Both segments are presumed to be exposed at the inside of the membrane and reside in polypeptide parts that may be protruding. Furthermore, they bear opposite net charges, which favors electrostatic interaction.
Experiment &Binding of the mAbs B82:4 and B315:32 to immobilized inside-out red cell vesicles could be inhibited by small amounts of P2 (Fig. 4). As mentioned above, the en- hancing effect of P1 was observed also in these assays (not illustrated). P4 and P5 were not tested.
Experiment c-Inside-out vesicles in suspension were much more efficient than right-side-out vesicles in adsorbing the two mAbs B82:4 and B315:32 (Fig. 5). This strongly suggests that the corresponding epitopes, which include parts of the C-terminal P2 sequence, are exposed on the cytoplasmic mem- brane face. However, due to the asymmetry of the red cell membrane, the mAbs theoretically might bind selectively but nonspecifically to inside-out vesicles but not to right-side-out vesicles.
0.0 1 0.1 1 10 20
AMOUNT OF VESICLE PROTEIN (pg)
1 0.0 1 0.1 1 10
AMOUNT OF VESICLE PROTEIN (pa)
FIG. 5. Ability of inside-out (A) and right-side-out (0) vesicles of human red cell membranes to adsorb monoclonal antibodies against the C terminus of the glucose transporter. A, mAb B82:4; B, mAb B315:32. The inside-out vesicle preparation was approximately 80% inside-out and the right-side-out vesicle preparation was 97-100% right-side-out. The vesicles were suspended in buffer D with 10 g of BSA/I, and 50-pl aliquots containing the given amounts of vesicle protein were mixed with 50 pl of hybridoma cell supernatants diluted 150-fold ( A ) and 3,000-fold ( B ) in the above buffer. These mixtures were incubated for 45-60 min at 22 “C and were then centrifuged at 10,500 X g for 5 min to sediment the vesicles. Supernatant samples were mixed with 0.1 volume of buffer D containing 0.5% Tween 20, and 5O-pl aliquots were assayed for binding of antibody to the glucose transporter by the ELISA procedures described under “Materials and Methods.” 50 ng of glucose transporter preparation were applied per well for coating. The ELISA signal for samples without vesicles, but otherwise treated as above, was 100%. The color development time was approximately 4 min.

Glucose Transporter Epitope Localization 11419
s 1 2 3 s 1 2 3 Mr
x
120- 102-
52-
38-
30-
21- - FIG. 6. Protein Immunoblotting. A, mAb B82:4, diluted 40-fold;
B, mAb B315:32, diluted 40-fold. The samples for lunes 1-3 and S were 120 pg of pig red cell ghost membrane protein (lane I ) , 60 pg of pig red cell integral membrane protein (lane 2), 60 pg of human red cell integral membrane protein (lane 3), and prestained reduced SDS- PAGE standard proteins (lane S) . Sample preparation and electro- phoresis were done as described under “Materials and Methods.” After protein transfer and blocking, the nitrocellulose membranes were incubated with the diluted hybridoma cell supernatants. The peroxidase-conjugated goat anti-mouse IgG antiserum was diluted 2000-fold. The arrows indicate the top and bottom edges of the separation gel.
We conclude that the mAbs B82:4 and B315:32 are directed toward the C terminus of the glucose transporter and that the C terminus is present on the cytoplasmic face of the human red cell membrane. We do not yet know whether these anti- bodies are also directed toward the nucleoside transporter, which co-purifies with the glucose transporter (10, 11).
Protein Immunoblotting-When human integral membrane proteins were separated by SDS-PAGE and the proteins were transferred to nitrocellulose membranes, mAb B82:4 as well as B315:32 labeled a broad zone from apparent M, 38,000 to 83,000 (Fig. 6). The upper limit of this interval is much higher than that of the zone of the purified glucose transporter observed on silver-stained gels (Fig. 2). This indicates superior sensitivity of our immunostainings compared to the silver staining, or cross-reactions with other membrane compo- nents. The zone boundaries are sharp. The front of the zone shows a distinct Mr-38,000 component. A zone of low color intensity is present a t M, 63,000. The faintly stained zone in the interval M, 95,000 to approximately 160,000 probably corresponds to the transporter dimer.
Red cells from adult pigs contain no functional glucose transporter, but have a nucleoside transporter of apparent M, 45,000-66,000 (33-35). We have tested ghost proteins and integral membrane proteins from pigs by immunoblotting to reveal possible cross-reactions of the C-terminal-specific mAbs with the pig nucleoside transporter. With the pig ghost membrane proteins only two very faint and narrow bands a t M , 44,000 and 78,000 were seen. No bands were detected by immunoblotting of pig integral membrane proteins (Fig. 6). The mAbs B82:4 and B315:32 directed toward the C terminus of the human red cell glucose transporter thus did not stain the pig nucleoside transporter. Another two glucose trans- porter-positive clones, B133:96 and B164:130, showed similar immunoblotting results as above with human and pig integral membrane proteins. They did not react with sheep red cell ghost proteins (not shown). When human glucose transporter was treated with endoglycosidase F (9) to remove the oligo- saccharide and was analyzed by two-dimensional electropho- resis essentially as in Ref. 11, immunoblotting with mAb
B315:32 showed a zone of nonglycosylated transporter mon- omer as expected.’
DISCUSSION
Recently, Davies and co-workers (18) reported that rabbit antisera, specific for the glucose transporter C terminus, showed binding to the cytoplasmic surface of the red cell membrane. Our results with mouse monoclonal antibodies against the C terminus of the human red cell glucose trans- porter are consistent with their result. In addition, we have found evidence for an interaction between the segment 218- 232 (Pl) and the C terminus (P2). This indicates that also the segment 218-232 is present on the inside of the membrane, in agreement with the model of Mueckler and co-workers (7), and that this segment is located adjacent to the C terminus in the native glucose transporter. The antibody-binding epi- topes may include amino acid residues from both segments (cf. Ref. 36). The C-terminal peptide P2 reacted strongly with several anti-transporter antibodies, which indicates that the C terminus is well exposed on the protein surface. The above results, and results in other recent reports (37, 38), support the Mueckler model for the human HepG2 glucose trans- porter. The sequence and the structure of this transporter are probably very similar to those of the human red cell glucose transporter (cf. Ref. 7).
Acknowledgments-We are grateful to Jan Andersson, Gunnar Alm, Lilian Torngren, Britta Cederblad, and Erik Wiersma at the Department of Immunology, Biomedical Center, Uppsala, for help, advice, and access to laboratory facilities. We are also grateful to Ake Engstrom at the above department for synthesis of the peptides and mass spectrometry analyses, and to Anders Larsson and Erik Mascher for comments on the manuscript. We thank Susanne Eriksson for typing the manuscript.
REFERENCES 1. Elbrink, J., and Bihler, I. (1975) Science 188, 1177-1183 2. Wheeler, T. J., and Hinkle, P. C. (1985) Annu. Reu. Physiol. 47,
3. Wheeler, T. J., and Hinkle, P. C. (1982) in Membranes and Transport (Martonosi, A. N., ed) Vol. 2, pp. 161-167, Plenum Press, New York
4. Baldwin, S. A. and Lienhard, G. E. (1981) Trends Biochem. Sci.
5. Kwong, F. Y. P., Baldwin, S. A., Scudder, P. R., Jarvis, S. M., Choy, M. Y. M., and Young, J. D. (1986) Biochem. J. 240,
503-517
6,208-211
349-356 6. Steck, T. L. (1974) J. Cell Bid. 62, 1-19 7. Mueckler, M., Caruso, C., Baldwin, S. A., Panico, M., Blench, I.,
Morris, H. R., Allard, W. J., Lienhard, G. E., and Lodish, H. F. (1985) Science 229,941-945
8. Gorga, F. R., Baldwin, S. A., and Lienhard, G. E. (1979) Biochem. Biophys. Res. Commun. 91,955-961
9. Lienhard, G. E., Crabb, J. H., and Ransome, K. J. (1984) Biochim. Biophys. Acta 769,404-410
10. Wu, J.-S. R., Kwong, F. Y. P., Jarvis, S. M., and Young, J. D. (1983) J. Bid. Chem. 258,13745-13751
11. Lundahl, P., Greijer, E., Cardell, S., Mascher, E., and Andersson, L. (1986) Biochim. Biophys. Acta 855,345-356
12. Birnbaum, M. J., Haspel, H. C., and Rosen, 0. M. (1986) Proc. Natl. Acad. Sci. U. S. A. 83,5784-5788
13. Lundahl, P., Greijer, E., Lindblom, H., and Fagerstam, L. G. (1984) J. Chromatogr. 297, 129-137
14. Lerner, R. A. (1982) Nature 299,592-596 15. van Regenmortel, M. H. V. (1987) Trends Biochem. Sci. 12,237-
240 16. Muller, S., Couppez, M., Briand, J.-P., Gordon, J., Sautikre, P.,
and van Regenmortel, M. H. V. (1985) Biochim. Biophys. Acta
17. Geysen, H. M., Rodda, S. J., Mason, T. J., Tribbick, G., and Schoofs, P. G. (1987) J. Immunol. Methods 102, 259-274
827,235-246
A.-K. Englund, personal communication.

11420 Glucose Transporter Epitope Localization
18.
19.
20.
21.
22. 23.
24.
25.
26.
27.
Davies, A., Meeran, K., Cairns, M. T., and Baldwin, S. A. (1987)
Sundqvist, B., and Macfarlane, R. D. (1985) Mass Spectrom. Rev.
Baldwin, S. A., Baldwin, J. M., and Lienhard, G. E. (1982)
Mascher, E., and Lundahl, P. (1987) J. Chromatogr. 397, 175-
Neville, D. M., Jr. (1971) J. Biol. Chem. 246, 6328-6334 Johansson, S., and Skoog, B. (1987) J. Biochem. Biophys. Methods 14 (suppl.) p. 33
Tunbn, P., and Johansson, K.-E. (1984) J. Biochem. Biophys. Methods 9,171-179
Wallevik, K., and Jensenius, J. C. (1982) J. Biochem. Biophys. Methods 6,17-21
Barany, G., and Merrifield, R. B. (1979) in The Peptides (Gross, E., and Meienhofer, J., eds) Vol. 2, pp. 1-255, Academic Press, New York
Carlsson, J., Drevin, H., and Axin, R. (1978) Biochem. J. 173,
J. Biol. Chem. 262,9347-9352
4,421-460
Biochemistry 2 1,3836-3842
186
723-737
28.
29.
30. 31.
32.
33.
34.
35.
36.
37.
38.
Shulman, M., Wilde, C. D., and Kohler, G. (1978) Nature 276,
Steck, T. L., and Kant, J. A. (1974) Methods Enzymol. 31, 172-
Steck, T. L., (1974) Methods Membr. Biol. 2,245-281 Liljas, L., Lundahl, P., and Hjertin, S. (1974) Biochim. Biophys.
Stocker, J . W., and Heusser, C. H. (1979) J . Immunol. Methods
Young, J. D., Paterson, A. R. P., and Henderson, J. F. (1985)
Kim, H. D., and McManus, T. J. (1971) Biochim. Biophys. Acta
Engelhardt, W. A., and Ljubimowa, M. (1930) Biochem. Z. 227,
Barlow. D. J.. Edwards. M. S.. and Thornton. J. M. (1986) Nature
269-270
180
Acta 362,327-337
26,87-95
Biochim. Biophys. Acta 842, 214-224
230,l-11
6-15
322,'747-i48 . .
Holman. G. D.. and Rees. W. D. (1987) Biochim. Bwuhys. Acta 897,395-405
Cairns, M. T., Alvarez, J., Panico, M., Gibbs, A. F., Morris, H.
Acta 905, 747-748 R., Chapman, D., and Baldwin, S. A. (1987) Biochim. Biophys.
"