the feeding ewlogy and hktt'jlnb tiiuluby of a .apt- vui
TRANSCRIPT

THE FEEDING EWLOGY AND HKtt'JlNb tiIULUbY OF A .APt- VUi MW
COLONY IN THE SOUTHWESTERN CAPE PROVINCE
A l i s t a i r S tuar t Robertson
A d iss e r ta t io n submitted to tne Faculty o f Science, M i .
o f the Witwatersrand, Johannesburg, fo r the degree of
Master of Science.
Johannesburg 1983

. , hen W i t h a sudden l i f t o f t h e one g r e a t pin ion.
Swung p r o u d l y to a c u r v e and f r o m U s h e i g h t ̂
l o o k ha l f a mile of s u n l i g h t In one lo n g sweep.
"Buzzards"Martin Armstrong

abstract
mmM - : . % % = , « . . . - — > - •
I*®< u r r p s s bv a 11 age j r o u p s .

i v
DECLARATION
I declare tha t th i s d i sse r t a t i on is my own work
ind that i t has not been submit ted to any Other Univers i ty .
■

ACKNOWLEDGEMENTS
The o u t l ine of th is study was proposed by Dr Andre' Boshoff
o f the Cape Department o f Nature and Environmental Conservation,
and I was employed by t h a t depar tment f o r two years . In 1983, I
was suppor ted by b u rs a r ie s from the CSIR and the U n i v e r s i t y of
the W i tw a te rs ra n d . I thank numerous members o f the CDNEC, in
p a r t i c u la r Wally du Preez and Kobus Mul le r , fo r t h e i r help wi th
a d m i n i s t r a t i v e m a t te rs . Andre 'gu ided me u n o b s t r u s i v e ly , and I
o f f e r my sincere appreciat ion fo r a l l his encouragement and help
throughout t h i s study.I thank P ro fe sso r Hugh Paterson f o r assuming sup e rv is ion
a f t e r the i n i t i a l s u p e rv is o r l e f t , and f o r c r i t i c i s m o f the
manuscript. I an also gra te fu l to Carl Vernon and Dr Peter Mundy fo r t h e i r enthusiasm in the study, and fo r commenting on d ra f ts
o f the m anusc r ip t . Pete's red pen was much in evidence in the
d ra f ts , and h is a t ten t ion to de ta i l was great ly appreciated. My
thanks to Marc Centner for help w i th s t a t i s t i c s , Neil Caithness
fo r he lp w i t h the word p rocessor . P ro fe sso r M i le s Markus
a n a l y s i s o f the b lood smears and Ms Ansie de Kock of tne
Un ive rs i ty of Port El izabeth fo r analysis of the addled eggs.Cape V u l tu r e s l i v e on c l i f f s and, as a c l im b e r , I was
in t ro d u c e d to them in 1977 by a Be lg ian e x p a t r i a t e , in o rde r to
pa r take in the V u l tu r e Study Group's r i n g i n g a c t i v i t i e s , i
consequen t ly thank Dr John Ledger and o th e r VSG members fo r
those f rag ran t and in s t ru c t iv e t r i p s , as well as the VSG fo r the
loan o f photographic equipment used during th i s study.L i v i n g a lone next to a v u l t u r e co lony was an in tense
experience, and i t would have been a very .onely one were i t not
f o r f r i e n d s such as the Februarys, Pe te r and Jenny Steyn, John
and A l i s o n M ic h le r , and Tony and K i t t y M e tse la a r , a l l o f whom
s u f f e r e d m, i n t r u s i o n s in to son i e t y w i t h a warmth t h a t deeply
impressed me. Mr and Mrs Feb. provided an "al ways-welcome" home
in Cape Town which I great ly appreciated.
I share these remote places with the memory of ny fa tn e r , and my mother cares ; I am p a r t ic u l a r ly gra te fu l to both fur
supporting my education in the direction ot my choice.

TABLE OF CONTENTS
Abstract 111
Declaration iv
Acknowledgements V
Lis t of Tables vi 1 i
Li >t of Figures ix
1. INTRODUCTION ..............
2. STUDY A R E A . . . .
2.1. P o th e r g —
2.1.1. Lo ca t ion
2.1.2. Cl imate
2.1.3. H i s t o r y
2.2. A a s v o g e l v l e i2.2.1. Loca t ion
2.2.2. Cl ima-.e
3. FEEDING ECOLOGY ...............................................................3.1. I n t r o d u c t i o n and methods .......................................
3 .2 . R e s u 1 t s ..........................................................................3.2.1. The foraging range
3.2.2. Nature o f the food source3.2.3. Stock contained w i th in the foraging range 1'
3.2.4. Amount o f food obtained by the vu l tu res 21
.2.5. Foraging behaviour
3.2.6. Contamination of the food source 31
3.3. D i s c u s s i o n .....................................................................3.3.1. The foraging range
3.3.2. Food obtained
3.3.2. Foraging behaviour
3.3.4. Contamination3.3.5. Allegations concerr . sheep attacks 41
4. BREEDING BIOLOGY ......................4.1. I n t r o d u c t i o n and methods
4242

4.2.1. Th< pre-laying pen4.2.2. The incubation period4.2.3. The nestling period4.2.4. The post-fledging dependence period
4.2.5. Breeding success
4.2.6. General behaviour
4.3. D i s c u s s i o n o f p e r i o d s ............................................
4.3.1. The p re- lay ing period
4.3.2. The incubation period
4.3.3. The nes t l ing period4.3.4. The pos t- f ledg ing dependence period
5. POPULATION DYNAMICS ........................................................
5.1. I n t r o d u c t i o n ................................................................5.2. In formation concerning the demographic parameters .
5.2.1. Populat ion size and s t ruc tu re
5.2.2. Deferred matur i ty5.2.3. Frequency of act ive s i te s , and of b r e e d i n g
5.2.4. Age-class survival rates
5.3. D iscuss ion .....................................................................5.3.1. Populat ion s tructure
5.3.2. Deferred m a tu r i ty
5.3.3. S u rv iva l
6. GENERAL DISCUSSION ...........................................................
APPENDIX A
APPENDIX B
APPENDIX C
APPENDIX D
APPENDIX E
REFERENCES

L IS T OF TABLES
Table
1. Breakdown of the posta l survey ................................
2. M a t e r i a l f o u r . ' i ' nest. .................................................3. ivestock total; for the Bredasdorp-Swellendam
a r e a .............................................................................................4 . colony food requirements, based on an average da j
metabolic rate ADMR) of 0.47 g per day(J. Komen per s. comm.) and a actor or ,5
taken from Gessaman (1973) .....................................................
5. Results o f es t imat ions of crop content of
i n d i v i d u a l b i r d s ..................................................................6. The re su l ts o f analys is of eggs co l lec ted during
the study period, expressed as mg/kg wet weight ..........
7. C op u la t io n a t te m p ts observed in the co lony ..............
8. Copulat ion attempts observed per period of
Breeding cyc le ..............................................................................9. The number o f days th a t juve n i les , during the i r
PFDPs, were considered not to have foraged .....................
10. The occurrence of f o o d -s o l i c i t in g behaviour
during the PFDP of f ive juven i les in 1981/2 ................
11. Breeding data fo r 1981 and 1982 atuotberg (PB) and Aasvogelvlei (AW) ...............................
12. P o s s i b l e c a u s e s o f breeding f a i l u r e and their
estimated occurrence at Potberg (PB) andAasvogel vlei (AW) during the study period ................
11. The number of months, in the year af ter fledging,that juveniles were resighted in the kloof .................
14. The number of nestlings that have been metal ringed
•md colour ringed at Potberg (data from Bos io 'f >
Currie 1981, CONEC records), and the number 1h w resubsequently r ' 1 during ■ . ly . eri o ............

Figure
L IS T OF FIGURESPag
41. The study area ....................................................2 . The percentage of r a i n f a l l each month a , a
proport ion of the total annual r a in fa l l , averaged
over a period o f 25 years (1955-19/9 .................................
3. View from the top o' the Potberg ..............................4. Results of the postal survey, giving *:he ocut ion
o f fa rm ers t h a t re po r ted to "never see vuitures ......
5. Results o f the postal survey, giving the location of farmers tha t reported to see vultures once > • ' yf ive years" ( 5 ) , or "once every ten years (6) 1
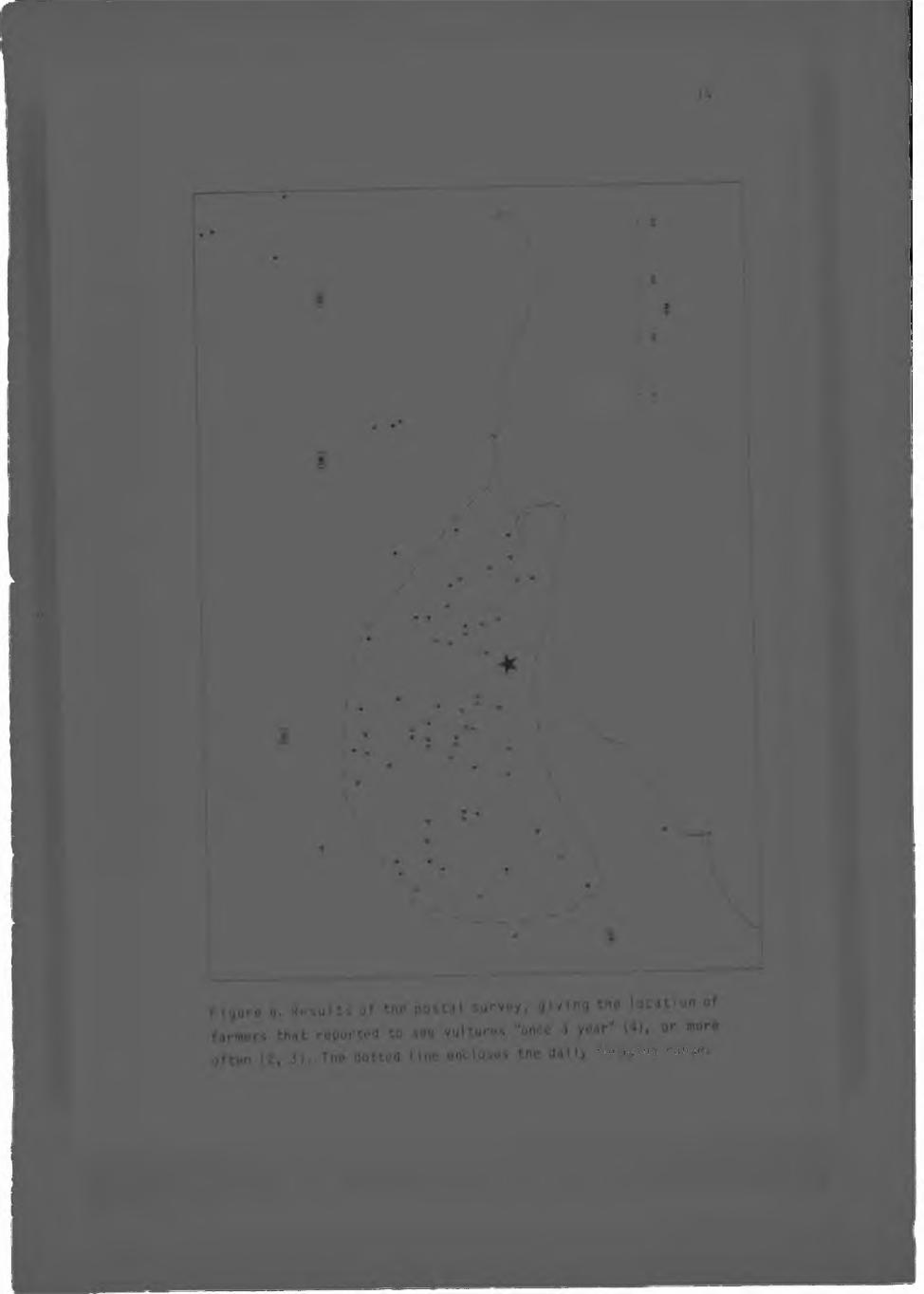
6. Results of the postal survey, g iv ing the local on of farmers that reported to see vu l tu res ' once a year
(4), o r more of ten (2, 3) ...........................................................7. Feeding forays of ind iv idua l vu l tu res , averagej
per calendar month ........................................................................8. Factors in f luenc ing the a v a i l a b i l i t y o f carcasses
to the vul tures ................................................... ..........................9. Dai ly es t imat ions o f the proport ion o f tne colon/
considered to have obtained food in an observation day .
10. Growth of Potberg nes t l ings in body mass, in r e la t io n to the mean curve from the Maga' iesberg
co lonies (Mundy 1982).....................................................................11. Growth of Potberg nes t l ings in wing 'enyth, in
re la t i o n to the mean curve from the M a g a l iesberg
colonies (Mundy 1982).......................................................... .12. The r e l a t i o n s h i p between body mass and wing length
of nestlings13. Monthly foraging pattern of the whole
colony : May 1981 ...............................14. Monthly foraging patterns oi the whole
colony : June - September 1981 ....
15. Monthly foraging patterns of th (hoi. co lony : October 1981 - January 1982
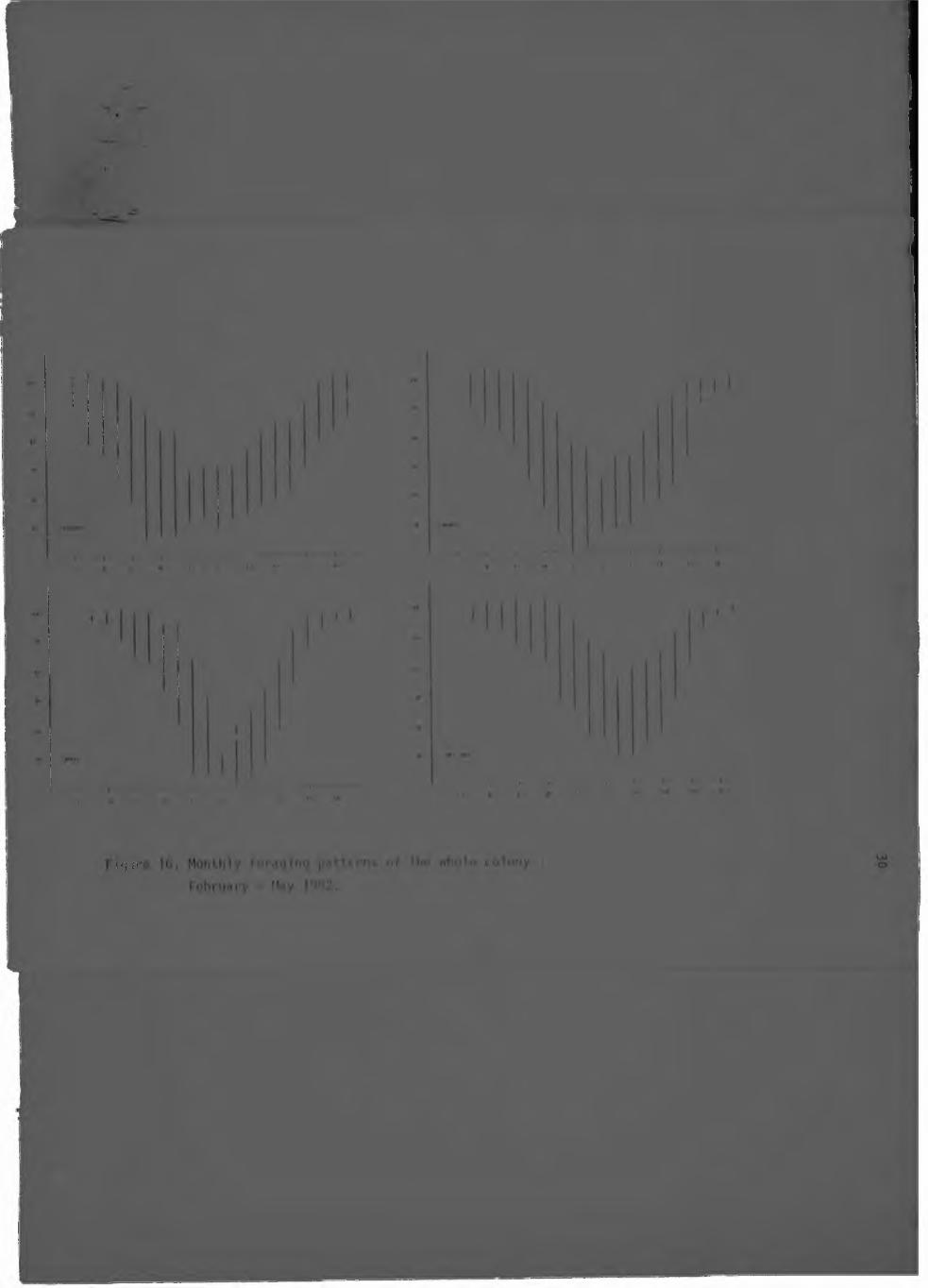
16. Monthly foraging patterns o* the whole
colony : February - May 1(82.............
...........................................................................................

32• • v17. Total c o u n t s o f v u l t u r e s made i n the evening
at P o t b e r g and A a s v o g e l v l e i ( • )
18. NW 1 c l i f f ............................................................................... t j19. NW2 c l i f f ..............................................................................
2 0 . NE c l i f f ......................................................... .....................21. The average level o f occupancy o f nests during
o i . f e r e n t p e r i o d s o f the b reed ing c y c le ................... 4o
22. A view of the NW2 breeding ledge ......................................... b223. Dates o f egg la y in g a t Potberg a n d Aasvogel v l e i ....... 54
61
2 5 . S im u l i id presence re la ted to the amount of
24. S i m u l i i d i n f e s t a t i o n o f a 47-day o ld n e s t l i n g ..........
ra in in the s ix months preceding September ( • ) o l
26. Diagram o f the breeding cycles o f ind iv idua l nests
( numbered on l e f t ) ..................................................................27. Diagram dep ic t ing the periods when a ju ve n i le
(nest no. 6) was at i t s nest during i t s PFDP ................... b6
28. Diagram dep ic t ing the periods when a juve n i le
(nest no. 6) was at i t s nest during i t s PFDP ................... b/
29. Diagram dep ic t ing the periods when a j uven i l e
(nest no. 63) was a t i t s nest during i t s PFDP ................ b0
30. The number o f ac t ive nests a t Potberg (numbered,
and A a s v o g e l v le i ( • ) f o r the l a s t nine vsars ............. 94
31. Diagram depic t ing the length of tune ringed
vu l tu res were resighted in the <loof .................................

1
CHAPTER ONE : INTRODUCTION
The Cape V u l t u r e (Gj^s c op ro the res F o r s t e r 1798) Is the
e a v i e s t m e m b e r o f t h e f a m i l y A c c i p i t n d a e t h a t o c c u r s as an
-ndemic species t h r o u g h o u t s o u t h e r n A f r i c a (Mundy 1982). A s p e . t s
,f u s H f e h i s t o r y i n c l u d e a low b r e e d i n g r a t e , d e f e r r e d
1981). I t b r e e d s in c o l o n i e s on c l i f f f a c e s and i s t h e o n , y
u u l t u r e s p e c i e s t o d o so i n s o u t h e r n A f r i c a (M u n d y
c o m b i n a t i o n o f t h e c o l o n i a l and c l i f f - n e s t i n g h a b i t s t h e n r e n d e r s
t h i s spec s c o n v e n i e n t t o s t u d y t h e c o n s eq u enc es o f l a r g e
and l o w r e p r o d u c t i v e r a t e s on t h e biology o f a v i a n s p e c i e s ; the
r e s u l t s may t h e r e f o r e be a p p l i c a b l e i n g e n e r a l t e r m s to a wi e
assemblage ^ ^ ^ Q t n e r g r i f f o n s p e c i e s d i s t r i b u t e d
th rougho u t the Old w o r ld ( l i s t e d in Brown i Amadon 1968) cape
Vultures are described as s p e c ia l i s t feeders on the carcasses o
large migratory ungulates (Houston 19740. The evo lu t ion of t h i s
e x c lu s iv e scavenging behav iour i s p o s tu la te d to have occurred
under c o n d i t i o n s (Houston 0£. c i U which probab ly no .onger
occu r th roughou t any g r i f f o n species ' range -n A t r i c a (Houston
1974a). Houston's (1972) study o f the feed ing and breed ing
eco logy o f two g r i f f o n species i n h a b i t i n g r e l a t i v e l y n a tu ra l
c o n d i t i o n s in East A f r i c a i s then e s p e c ia l l y re le van - when
considering the congeners th a t range over areas of in d u s tn a
a g r ic u l tu ra l development In southern A fr ica . In p a r t i c u la r h s
s tudy o f M p p e l l ' s G r i f f o n G. r u e p p e i m (Houston 19-4a. - .
1976), t h a t area's e c o lo g ic a l c o u n te rp a r t to G. c o p r o th e r es
(Mundy 1982), provides numerous po ints o f po ten t ia l comparison.In a comparat i ,e study o f the b io lo g y o f the f i v e sou the rn
A fr ican vu l tu re species. Mundy (1982) h igh l igh ted the decl ine of
the Cape Vulture and l i s te d a va r ie ty o f man-induced factors to
account f o r the decl ine in numbers. This decl ine was described as " a c c e le r a t in g " and g i v in g "cause r concern" ( S ie g f r i e d e ta _ l .
1976), and the v u l t u r e is now l i s t e d as "a v u ln e ra b le endemic
species of southern A fr ica which has los t much of i t s populat ion"
(Brooke in press). That rev iew documents e s t im a te d numbers and


CHAPTER ONE : INTROUUC:ION
Tho r , p . Vulture (Gyps c o p ro t i jr .
heaviest member of the fam ily A c c lp U r id a t th t endemic species throughout southern Afr ica (Mundy 1 « ) . A of i t s l i f e h is to ry include a lo« breed ing r a t e , d, rer m a t u r i t y , prolonged parental c a r e , high a d u l t s u r v iv a l and 1o.
juvenile survival rates (Houston 1974b, Mundy 1982, Piper et aj,
1 , 8 1 ). n breeds in colonies on c l i f f faces and i s vu ltu re species to do so In southern A fr ica (Mundy 1982). combination of the colonial and c l i f f -nes t ing habits then render
this species convenient to study the consequences of large s i t
and low reproductive rates on the biology of avian species ; results may therefore be applicable in general terms to a wide-
assemblage of animals.Along w ith the six other g r i f fo n species distr 'DU --
throughout the Old World ( l i s t e d in Brown & Amadon 196b, -*P<
Vultures are described as specialist feeders on the carcasse large migratory ungulates (Houston 1974c). The evolution of this
exclusive scavenging behaviour is postulated to have occurred
under condit ions (Houston 0 £ . c U . ) which probably no longer
occur throughout any g r i f f o n species' range in Afr ica (Hous*
1974a). Houston's (1972) study of the feeding and bree ecology of two g r i f fo n species inhab it ing r e l a t i v e l y nat i
conditions in East A fr ic a is then espec ia l ly relevant w considering the congeners that range over areas o' industna
agricultural development in southern Africa, .n particu e ,study o f Ruppell 's G r i f f o n G. r u e £ £ e n i i (Houston 19 74a,
1976), that area's ecological counterpart to G. co fro j;
(Mundy 1982), provides numerous points of potential compain a comparative study of the biology of the f ive south, -
African vulture species, Mundy (1982) highlighted the decllrv
the Cape Vulture and listed a variety of man-induced ‘ actor
account for the decline in numbers. This decline was desc “accelerating" and giving "cause t concern' (S ieg fr ied
1976), and the vulture is now l is te d as "a vulnerable e in species of southern Africa which has lost much o‘ i ts popu a
(Brooke in press). That review documents estimated numne

2
population t r e „Js, and concludes -1 th a to ta l of some 2u00
p-eeding pairs throughout the subcontinent. In the Cape Provinc
alone, some 65 breeding pairs were estimated ,n 19/9 (Boshc M
Vernon 1980), and the Transvaal Province now constitutes toe ma-n
s t r o n g h o l d (Tarboton S Allan in press). The temporal gradation i n
the degree of man's impact on tne food supply and numbers or the
Cape vulture, including i ts past and present distribution in the
Caoe Province, is described In Boshoff i Vernon (1980).Although the Cape Vulture is one of the best studio o
South Afr ican rare and endangered birds (Brooke in press , c e r ta in aspects of i t s (and raptors in general) l i f e history
remained " t o t a l l y un invest iga ted" , e.g. the p o s t - f l e d g i n g
dependence p e r io d (Mundy 1982:234, Newton 1979). More
importantly, the possibi l i ty of the food supply being the factor
l im it ing the population "remains to be fu l ly investigated IMundy
1982:275). In essence, this was the rationale behind this study . to examine the nature and effects of the current food supply on a p a r t ic u la r colony's reproductive performance, as well as to
determine tne degree of inf luence of various factors tha t nave
been documented as influencing numbers in different areas of tne
species' range. Tne aim, there fo re , was to provide a d e ta i le d , and essentially conservation-oriented, case study of a colony.
I studied the Potberg colony In the southwestern cape
Province, and monitored breeding at another colony 120 km away in
the L i t t l e Karoo. Evidence Indicated that these vultures likely
constituted a discrete group of the species' population (boshoff S Vernon 19»0). Situated in sheep country (J a rv is e t a j . . 19 4 i , the area Including the k lo o f was purchased in 1978 b" the Cape
Department of Nature and Environmental Conservation (CONICI, and
now forms part of the De Hoop Nature Reserve. I t is unique in
that i t is tne only remaining regular breeding colony within the
winter ra in fa l l region, and lies on the periphery of the species' current range. Although the colony had been under sporadic
observation, and r ing ing v i s i t s , since 1951, annual monitoring
began in 1974 with the colour nging of nestl ings. A va i lab le
data for the period 1951 - 1980 are collated In Boshoff 4 Currie
(1981) and Boshoff (1981) My study commenced in March VWl, with the emohasis in the f i r s t year on observing the birds in t h e i r

••.*■»« «■ «


s o u t h e r n s l o p e s and 4 km f r o m i t s w e s t e r n e x t r e m i t v . f i t h i n th e
S S S S : : : :
V : 1 , - L q u a r t z u J T l i ^ of the Table Mountain Series IHeydorn
Tinley 1980) are ,-pproximately 70 m high and the kloof is abou.
km in length. The nesting c l i f f s are some 60 m high.
2.1.2. Climate
The study area is s i tu a te d with in the bi,nodal r a in f a I
r e g i o n as defined by Heydorn « Tinley (1980). close to the
junct ion of th is region with the w in te r r a i n f a l l area.
JAM
FllM W
AVK
MAT
JON
75 Tear A .e ra g r
II,J
51.J SI .8 SM S I . )
HO 81 07S7,».1.8
II).I18.S ?S,I
lb ,4 M .J 7 ) , I 4 1.6 .46.9 1*1,J>4,9 14.9 n.an.a 07.J i *. '
J IN
AUG
SIPOCTMOV
otc
?S i r A » .
57.8 01.0 )».?40.8 71.7 IS ,4
uu 7.8 36,4 40,sa.S8 0 .u
40,1
1),l SI .8 7B.S
8747./»,/> J ,»
10.021.738.1
nn
14 * II 8l-V T f ith
J A
Figure 2. The percentage of rain^ 11 each month as a proport ion
o f the total annual r a in fa l l , averaged over i period ot years (1965-1979). Actual monthly amounts in mm are g iven above v".e
f i jure (J. C. M ich ler pers. comm.).

6
g e n e r a l , the area e x h i b i t s a temperate M ed l te r ranea ,
5 5 = = : = =rear 1981 was excep t iona l ly wet m . t a t i_4 p f e l l than the average obtained tor tne last t >
demonstrated In Heydorn 4 Tln ley (1980). a marked feature
r , i n f a l l in the Cape coastal region Is the strong o r o g r . p t , -
ccntro l . where r a i n f a l l curves fo l low the r e l i e f undula.io,
closely. This influence was observable at the Potberg where, 'numerous occasions, the mountain was covered in cloud wh.lu esurrounding farmlands were r e l a t i v e l y c lear . The Incidence u p re c ip i ta t io n and cloudy condit ions is therefore higher in .e
kloof than in the surrounding area.The strongest winds occur In midsummer, with the au’-unv an
early w in ter being the seasons with the leas t wind (Schu.z ,
1S65). P r e v a i l in g summer winds are soutneafter y
winter northwesterly winds are frequent.In 1.982, the farm O r ie f o n t e .n (14 km W o f tne Potberg,
received an average of 7,2 hours of sunlight per day (range V -
9,6 hrs) : only In A p r i l , June and July did th is f igure H ibelow six hours per day. Between the months of October and Marcn.
the average was not less than seven hours per day. These a r tother data were obtained from the Swellendam Extension 01; , . ' o'A g r i c u l t u r a l Technical Services (ATS) ; where this source is used,1 the te x t , i t i s acknowledged as "ATS-J.P. Piete-sen pers.
rnmm.
2.1.3. History
Before the a r r i v a l o f European s e t t le r s in the southwestern
Cape, bovids th a t frequented the area in r e la t i v e l y large numbers
i n c l u d e d Bontebok OamaI i scus d d f t a s oorcas o ’a . a , . fl. .
Hartebeest Alcelaphus bus.laphus (Pal las) and Eland Taurotragus
oryx (Pal las) (du P lessis 1969, Skead 1580). Being of r e la t i v e l y

7
large size, these would have p rov ided a l i k e l y food *o«rce . « r g r i f f o n v u l t u r e s . Carnivores included Hyaena .p p . , L e o p - , i s
Panthera pardus (L.) as w e l l as L ions P. Jeo " ■ / ;c i t . ) . The vegetat ion type in the s l i g h t l y undulat ing to , ^ s e c t e d lowlands th a t surround the Potoerg on a l l f lanks except the southern is (or ra ther , was) Coastal Rl.enosterbosveld Aco.n.
1975, Dav e t a h 1979). The l im es to ne h i l l s and sand pin ■-
comprising the coastal forelands were, and are largely s t i ’ h t
the Coastal Macchia (Fynoos) vegetat ion type (Day et ah : ‘ ' ■In the late eighteenth century, a group of pure-bred me-
sheep were introduced to the (Bredasdorp) area. Pastures w
introduced as a crop ro ta t ion in the la te 1930's, and sheep
farming intensified. Wheat farming was introduced in the ,• r
nineteenth century and in 1892 the f i r s t bags were transported to
the Cape Town markets (B. Brynard i n m t .h With the except or of the south, the area around the mountain is now a patchwork of
uxhPAtfields and stock pastures (Figure 3).
Figure 3. View from the
top of the Potberg.The Langeberg range

poratea
he CONE
c.c
Tills colony (33 5*!^; 21 Jot) is s i tu d t t formed by the Gourits River, 4 km north of the confluence of the
C ounts and Groot r iv e rs . This posit ion 1s 8 km north of the
Ungeoerg range, and 1s 120 km ( l i n e of s ight) from Potberg, a t
an a l t i t u d e of ca 150 m (Figure 1).The nest s i tes face south-east, i .e . on the western side of
the r i v e r , and are ea s i ly viewed from the eastern bank. This
observation point is reached via the farm "Grootplaas" (resident owner P. Beukes) ; the nesting c l i f f s are included on this farm's
property. The c M i * s are ca 90 m in height and the nests are
located, on average ca 30 m above the water. During the study
period, the nests were spread along some 1,T km o t f t' f a-
2 .2 . 2 .Climate
S i tu a te d in the r a in shadow area of the L i t t l e ta* j o , ttn

<-t
t h e v u i t u r
e r v a t i o n s c o m m en i
l a r d 1 e s s c weatner
, 165 days wer
i , was e r e c t e d a t
Potential food aval labF o u r m e thod s we re
range a r o u n d t h e c o l o m
a. a q u e s t i o n n a i r e , r
was p o s t e d t o t h e f a n
B r e d a s d o r p , S w e l l e n d <
b. Per ,onal i n t e r v i e w s
km r a d i u s o f t h e c o l o r

Informat ion per ta in ing to the a v a i l a b i l i t y o f '.area to
the vul tures was obtained by personal interv iews wi th the f i rme-s
w i t h i n the foraging range. The farmers were questioned on, j n t r
a l i a , sheep m o r t a l i t y r a te s (and temporal v a r i a t i o n s t h e r e o f ) ,
carcass management procedures and t h e i r personal a t t i t udes to tne
vul tures.
Food o b t a i n e d , and f o r a g i n g behaviourTo determine the range o f food used by the colony, the base.*
o f t he b r e e d i n g and r o o s t i n g c l i f f s were sea rched r o r
regurgi tated pe l l e t s and ar te fac ts and farmers were interviewed
in o rder to ob ta i n t h e i r personal observa t i ons . A r t e f a c t s and
o th e r i n d i c a t o r s o f the nature o f the food ob ta i ned we> •-*
co l lec ted during v i s i t s to the nests. The Potberg breeding areas
w e r e v i s i t ed four t imes in 1981 and f i v e t imes in 1982, al though
i n d i v i d u a l nest s i t e s were v i s i t e d no more than three t imes in
any one year . The Aasvogel v l e i s i t e s were v i s i t e d once i n each
y e a r .
A number o f carcass# s consumed by t h e v u l t u r e s w e re l o c a t e d
by s e a r c h i n g t h e f a r m l a n d s close t o t h e c o l o n y .
Var i ous methods were used in an a t t emp t to de termine the
actual amount o f food ob ta i ned by the b i r d s , as we l l as g i ve an
i n d i c a t i o n o f i t s a v a i l a b i l i t y . One o f these, the v i sua l
e s t i m a t i o n o f crop c on te n t o f r e t u r n i n g f o rage rs , has been
attempted before (Houston 19 2, 19 6). ; he crop, a d iver t i cu lum
o f the oesophagus which f u n c t i o n s as an e l a s t i c food s to r e ,
projects beyond the contour feathers when f u l l and is observable
in bot. i f l y i n g and perched b i r d s (e.g. the smal l bulge in the
neck of the vu l tu re <n the f ron t i sp i ece, also Houston 197b:Figure
3). Capt i ve b i r d s were fed measured amounts o f meat and the
resu l tan t crop bulge was photographed in order to tain a visual
es t im a te o f tne degree o f d i s t e ns io n . Dur ing the day- long
obse rv a t i on s , e s t i m a t i o n s were made o f the propot ion o f the
colony cons idered to have ob ta i ned food t h a t day. I n d i v i d u a l
breeding s i tes were monitored each aservat ion day and f he degree
of crop d i s t e n s i o n o f each occupant was scored i n t o oi e of the
f o l lowing three categor ies, whenever possible :
0 : No food ob ta i ned.

1 : F o o d o b t a i n e d , l e s s t h a n c_a 7oug .
i • Food o b t a i n e d , more t h a n ca - UUg.
A t each v i s i t t o t h e n e s t s , n e s t l i n g s - e r e w e ig h e d and t h e i r
l e n g t h s w e re m easu red . As n e a r l y a , I 1931 and 1982 n e s t l i n g s
f r o m P o t h e r e w e r e o f kn ow n age, t h e s e f i g u r e s w e re t h e n d s r e e t l y
r o m p a r a b l e t o t h e c o r r e l a t i o n s o f w e i g h t and g r o w t h o b t a i n e d a t
n u m e r o u s c o l o n i e s d u r i n g t h e s o u t h e r n A f r i c a n r i n g i n g p rog ram m e.
During t h e o b s e r v a t i o n d a y s , c o u n t s w e r e made e v e r y
m m utes o f t h e t o t a l n u m b e r o f b i r d s p r e s e n t i n t h e k l o o t m
order to d o c u m e n t w h o l e c o l o n y f o r a g i n g t i m e s and t h e p r o p o r t i o n
o f t im e s p e n t i n t h e s e a r c h f o r f o o d . E s t i m a t e s o f c l o u d c o v e r
and w i n d s p e e d w e r e made t h r o u g h o u t e a c h d a y . The r e s u l t s w e r e
grouped by c a l e n d a r m o n th , and c o m p a re d by M u l t i p l e Range tests
using a HP-85 c a l c u l a t o r .
3 . 2 . Results
3.2.1.The foraging range
A b r e a k d o w n o f t h e p o s t a l s u r v e y i s g i v e n i n T a o le 1.
Table 1. B r e a k d o w n o f t h e p o s t a : s u r v e y .
Quest ionra ires f a ' ' ed
» returned ( v i a b l e )
» " ( incomplete )
Uns o l i c i t ed - n f o m a t n suppl ied
Total Per cent
1173 (A) -
C.;5 (B) 47 (o f A)
51 4 (o f A)
156 23 (o f B)
The r e s u l t s o f t h e p o s t a l s u r v e y a r e d e p i c t e d i n F i g u r e s 4 , 5 and
6 . V u l t u r e s s i g h t e d by f a r m e r s i n t h e f a r m l a n d s s u r r o u n d i n g th e
P o r o e r g a r e p r e s u m e d t o b e c o l , m e m b e r s . F i g u r e 6 i n c l u d e s
s i g h t i n g s o f b i r d s t h a t a r e p r e s u m e d t o h a v e f o r a g e d f r o m t h e
A a s v o g e l v l e i c o l o n y , as t h e s i g h t i n g s w e r e made I n f a r m l a n d s
c lose to t h a t c o l o n y . F ig u re 6 a l s o i n c l u d e s an o u t l i n e o f the

n of
td r marks the
l e ib e r q , RD-

F ig u re 5. Resu l ts o f the po s ta l survey, g i v i n g the lo c a t io n
fa rmers t h a t re p o r te d to see v u l t u r e s "once every 5 ye a rs ' 151,
or "once every 1U years (6).
foraying range

IS
c u r r e n t fo ra g in g range, an area o f a p p ro x im a te ly 200 000 ha, as
de te rm ined by compar ison o f t h i s F ig u re w i t h F igu re 4. The l im e s to n e r id g e i s u n c u l t i v a te d , suppor ts a c o m p a ra t iv e ly low
number o f stock and is not included w i th in the foraging range in
the ca lcu la t ion s o f po ten t ia l food. This f igu re therefore d i f fe r s
from tha t given in Robertson (1983a). The Oe Hoop Nature Reserve
i s a lso exc luded due to i t s low ungu la te c o n te n t a t the t im e o the s tudy , behav iour o f the rad io -m arked v u l t u r e and me
observation tha t vu l tu res "are never" seen over the i)e Hoop area
(P. van der Uesthu izen pers. comm.). S ig h t in g s of v u l t u r e s
o u ts id e o f t h i s range are documented in cha p te r f i v e (5.1.).
A l though c e r t a i n farms where the r e s id e n ts r e p l i e d t h a t they
never saw vu l tu res could not be located, a l l farms where vultures
were seen "once a yea r" o r mort o f te n were loca ted , and are
included in Figure 6.A l l " feeding" s i tes located during the rad io - t rack ing study
were s i t u a t e d w i t h i n the boundaries o f the o u t l i n e in F igure 6
(Boshoff, Robertson 4 Norton in prep.I.The o v e r a l l average feed ing t im e was 3 hours 42 minutes
( range 52 m inu tes - 7 h e i r s 5 m inu tes , n=323), and these are
presented in Figure 7. Analysis ind icated s ig n i f i c a n t d i f ferences
between various months, in p a r t i c u la r shorter forays fo r months
d u r in g which lambing occu r red , as compared to o u t - o f - l a m b in g
season months (Tukey's M u l t ip le Range tes t , 1% level) . Montns of
lambing are in c lu d e d In 3.2.3. On 28 August 1982, a group ot
v u l t u r e s was observed to feed on a farm 6 km from the k lo o f .
A f t e r fe e d in g , 10 b i r d s (o f 16 p resen t ) lay down and were s t a t i o n a r y f o r 35 m inu tes , some 35 m from tne carcass,
weather was o v e rc a s t and when a breeze p icked up and a l i g h t
d r i z z l e s t a r t e d , a l l f le w o f f ; the o b s e rv a t io n i s inc luded to
i l l u s t r a t e how the feeding times may be inf luenced by a range ot
behavioural land other) factors (see Mundy 1982). From the r a c a -
t rack ing study, a fac to r o f distance to feeding s i te per feeding
t im e was ob ta ined (B osho f f , Robertson 4 Norton in prep.), . s ' " )
t h i s f a c t o r , the above average t ime o f 3 hours 42 minutes corresponds to feeding s i tes located approximately 13 km from the
k lo o f .


17
3.2.^ N . r . ' o f the fooJ source
A rang e o f a r t e f a c t s , bones and bone f r a g m e n t s , p resum ed t o
have been b r o u g h t back t o t h e k l o o f by t h e v u l t u r e s was fo u n d a t
t h e b a s e s o f t h e b r e e d i n g a n d r o o s t i n g c l i ' t s , : ah - ;
r e g u r g i t a t e d p e l l e t s w e re o f s heep 's w o o l . Seven sheep e a r t a g s
w e r e f o u n d a t t h e b a s e o f t h e b r e e d i n g c l i f f s d u r i n g t h e s t u d y .
Table 2. Me' found at nests. Where possible, the bones were
id e n t i f i e d : ver teb-a ( v ) , metatarsal v-i t i or r i b i n .
1981
in ta c tBore
f • .n e tt •- '
j rny t $f n tiv e r in g
. th e r1 - g lass 20x30
R egurg ita te
. )GELVLEi
i nt actBone
fragment ' 3 . ■ - *
3 ■cv. V) dOxtG(r)
■ irn y noof cove ring
Regurg ita te
Al l of the 45 farmers tha t were interv iewed considered that
the v u l t u r e s c u r r e n t l y feed e x c l u s i v e l y on sheep carcasses,
a l though f i v e were o f the o p in io n t h a t dead c a t t l e cou ld w r y
l i k e l y ' supplement th is d iet.A l l c a r c a s s e s v i s i t e d by v u l t u r e s , t h a t w e re l o c a t e d d u r i n g
the r a d i o - t r a c k i n g s t u d y , w e r e o f s h e e p ( B o s h o f f , R o b e r t s o n 4
N o r t o n i n p r e p . ) .

18
3.2.3.Stock contained within the foraying r
A large amount o f s tock , o f which sheep and c a t t l e are the
most numerous, is farmed in the area. Sheep and c a t t l e t o t a l s tor
the la s t six years are presented in ’ able 3.
Table 3. L i v e s t o c k to ta ls f o r t h e B r e d a s d o r p - S w e l 1e n d a m a r e a .
These data were obtained f r o m ATS-J.P. Pietersen p e r s . comm, and
t h e yearly reports of t h e S w e l l endam S t a t e V e t e r i n a r i a n .
, " " " " —---------- — • - ' ---------------------
P e rio d o f census lendam Sredasdoro
c a t t lesheec c a t t le sheep
•981 /2 306091 16230 315020 ’ 9740
' 9 8 0 /' 29730: •5912 33569’ ■3554
•979/80 354868 16449 332230 '9779
'9 7 8 /9 236361 1 6U 5 296974 21660
1977/8 222390 16098 240600 •8800
'976 " 219130 15039 238371 18584
In the c a lcu la t io ns of the theoret ica l amount of food
ava i la b le to the vu ltures , only sheep are considered. This was because evidence obtained during th is study indicated that » e
Potberg vu ltures feed almost exc lus ive ly on sheep carcasses
(3 .2 .2 . ) .The average annual adult sheep mortality estimate obtained
from these interviews was 2,2 pe cent. All farmers Interviewed
considered sheep deaths to be temporally dependant, witn a peak at the time of lambing. In addit ion, in the w in ter r a i n f a l l region an average o> lb ,9 per cent of lambs die between birth and

19
, „ „ 1 n 9 (Lou- 1970). Of 45 fa rmers in te r v ie w e d , the dates , f
1 ambi n ! seasons were ob ta ine d from 38: a , , o f these cons: d e n , , the months of M a r c h - A p r i l to cons t i tu te tne mam lambing seas..,,
w h i le 34 (89%) i n d ic a te d t h a t ap p ro x im a te ly 30 per cent o yea r 's lambing occurs in the September-October pe r iod . Also,
losses to d iseases such as lamb dysentery and, es^ c l 3 l , y ' enterotoxaemta are heaviest during the w in te r months (Monmg S
veldman 1976). F ig u re 8 o u t l i n e s the f a c t o r s i n f l u e n c in g
a v a i l a b i l i t y o f these carcasses to the vu ltures.
SHEEP DEATH
BURIED, BURNT REMOVED
CO,ERED IN POISON
POTENTIAL FOOD
the vultures.
The farms of a l l the farmers that were interviewed were
situated within the boundaries of the foraging range in Figure o.An estimated average of 1,7 sheep per hectare of cultivated
farmland as obtained from these interviews. This is probably a n
underestimate, e.g. on the farm Mopama (35 km N of Potberg), some
8500 sheep are pastured on ca 2000 ha, an average of 4,2 sheep
per ha (M.G. Lourens pers. comm.). The potential number el sheep
deaths that can be expected in one year is then ca lculated as
follows:
Foraging area : 1 ^ !) ha74% under c u l t i v a t i o n : 143 5bt) ha
1,7 sheep per ha : 244 Obr! sheep
M o r ta l i t y est imate : 5369 sheep

20
The f ig u re of 74 per cent of the area surrounding the c o l o n y
being under c u l t i v a t io n .a s obtained from ATS-J. P- ' 'e te rsen
pers. comm. Of the 45 farmers questioned, 29 <621 considered i t
important to remove the carcass from the grazing lands as soon a
i t was found. I f the carcass was reasonably' • ■ . . . . 4a l w a y s be removed, but 16 farmers repl i ed that carcasses in an advanced state of decomposition would ei ther be l e f t or covered
w i t h p o i s o n . I f the factor of two-thirds is applied to the figure r e a c h e d in the above calculations, 1790 carcasses are calculated
as b e i n g th e o r e t i c a l l y av a i l a b le to the vul tures wi thin tne foraging area in one year. This must be considered a minimum
est imate : not a l l carcasses are located and not a l l those that are removed are rendered unavai lable to the vul tures (pers.
ohs.). ,A recent study of the da i ly requirements of Cape Vultures
(Komen 1983) confirmed an ear l i er estimate (Houston 1972, Mundy1982) of 300-500 g of lean meat per day, thus the amount requiredby the whole colony may be estimateu (Table 4). In addition, eachnest l ing requires some 76 kg of lean meat between hatching and
fledging (Komen 1983).
Table 4. Colony food requirements, based on an ave-age dai ly
metabol ic rate (ADMRl of 0,47 g per day (J. Komen pers. comm.) and a factor of 1,5 taken from Gessaman ( 1 9 3 ) . One sheep is
taken as the equivalent of 15 kg of meat (Jarvis et aU 1974).
-----------— — ———— —
Wet'k Year
AOMR 1 .5 x ADMR kg Sheep kg sheep
0 .4 7 0 .7 0 4 . 9 0 . 3 257 .3 17.1
2 8 ,2 4 2 . 3 19 ,7 15439 1920

3.2.4. Amount of food obtained by the vultures
The resu l ts of the visual est imat ion o f the proport ion ot
the colony c o n s i d e r e d to have obtained food during a p a r t i c u la r
observation day, expressed as averages fo r each calendar month,
are presented in Figure 9. The overa l l monthly average of 40 per
cent t r a n s la t e s i n t o an i n d i v i d u a l b i r d feed ing ap p ro x im a te ly
every two and a h a l f days. D i f f i c u l t i e s were exper ienced w i th
t h i s method (3.3.).
6C
4 0
20
m o n t h s
F igu re 9. D a i l y e s t im a t i o n s o f the p ro p o r t i o n o f the co lony
cons idered to have ob ta ined food in an o b s e rva t ion day. he
proport ions are averaged per ca le .dar month. The mean estimate is
i nd ica ted ( x).
A l though members o f a p a i r t h a t have f a i . e u du r i ig breeding cycle may re ta in th e i r nest s i te , the level of occupy
drops a f t e r the p o in t of breed ing f a i l u r e (chapte r fo u r ) , in
not a l l s i te occupants could be constantly monitored throughout
one year . As most birds at nest s i t e s were not I n d i v i d u a l l y
recognisable, only breeding s i tes where a f ledg l ing was raised m the 1981 season were monitored. The resu l ts o f t h i s i n d iv id u a l


Table 5. Results ol i 11on of crop content of ind i v uai d i ruColumn B n ves the number ot column D t number of day
to have obtained food.idered
i t no. crops estimated
16
17
61
63 6 1 , 7MM
Tota l : 15H3 6 9 , 0 17 , 1 6 4 . 4
1

23
r p i ■ n j I e >, wher • in ov*;ra 11
a iv l , i .r p f oi were ob ta ined per nest per day
Tj i 5, 11 umn A/B). 01 - i ' op , recorded, :>3 pe rc e n t were: , d a; empty (C/A) ; -hi ndicates that individuals obtain
• ; i ne of i vei . tw at tempts , he d i f ference between the• er of tim< . bi wer<- recorded > having obtained food,
ired i retur . ig w ti tn empty crop, was not signif icant (X2•0,6-3, f id . f . , p>0,25). Of the b i r d s recorded as having obtained
d , 69 per e n 1 were est imated as having obtained more than
-h, al ly requirement (E/D). On 64 per cent of the observation, rop est imat ions of both members of a pair were obtained
i) and on I 7 per cent of these days, botn birds were concluded
i laving obtained food in the one day. At changeovers during theubation period, the incoming partner had obtained food on 64
r ;ent of the post - l lh30 changeovers (4.2-2.).weights and wing lengths of Potberg nestlings of known age
: presented in Figures 10 and 11, compared to the curve ofmi 3t" measurements made at the Maga 1 iesber g and Botswana
ionies o r ig ina l graph supplied by P.J. Mundy). Figure 12
ocuments the body mass - wing length correlation measured during
the study period at Aasvogelvlei, in relat ion to the mean curve
• Transvaal nest l ing growth. In 1981, four Potberg nest l ings
were v is i t ed twice , and one was weighed once. In 1982, two nestling.; were visi ted three times; the age of one of these was
not [town accurately , but was est imated and included in the
f igure due to i t s exceptional mass. , ae weight of another
es t l ing was not obtained. Thus a l l (but one) nest l ings were
neavier than the mean weight at the same age that was recorded at '.he Magaliesberg colonies, and a l l but two had longer wing
engths for t h e i r age ( in the two "shortens", feathers had not vet appeared). One 1981 and one 1982 nest l ing had except ional ly 'ong wing lengths for their age- nestling periods were 139 days
Hid 124 days (shortest recorded in this study) respectively.Three large nestlings vacated their nests in November 1982,
t t 1 i ke ly the resul t of human disturbance (see chapter four, i. .5.). Two were located, one a l i ve but seriously injured, withi-i i week, of the event and post-mortems on both were carried
it by i vet rinary »urgeon (N. )rager). hi level of fat storage

Figure 10. Grow V of otberg m i t l ings n oody mass,
to the mean cu^V'. f> 1 Maga1 i sber.;; colonies Mund>
;oe .
460 ,
. u *<) 60ACI (iNv
Figure 11. Growth of Potoer ; n< . t l i f i i i a <vrelat ion to the mean cu^vr f n i tfv1 v ig il i < - i c -1
1982).

BODY
MA
SS
(kg
)
25
7 -I
5 -
* 1
3 -
/» *
/
• 19 T , 1979, 1980i 1981
A 1982O A W 1961a AW 1982
— i— 100
— I—
200— l—300
-T400 500 SCO
WING LENGTH m ;
Figure 12. The relationship between oody mass and wing length of nest l ings. This includes nest l ings weighed during the study
period at Potberg and Aasvogel v le i (as indicated) , as wel l as Potberg nestlings weighed in 1977, 1979 and 1980 (data from IDNEC
records). The straight l ine of the relationship obtained from the
Maga1iesberg colonies was supplied by P.J. Mundy.

26
was considerable in both and was clashed as score b and score 7 (highest possible score of 9, see Houston 1976:17). The stomach
of one contained several sticks, the largest measuring 180 x 10
mm, an end of which had caused an ulcer in the pyl m s egion and
most l i k e l y a small rupture in the 1 e f t l i v e r lobe ( local ised
f ibr inous p e r i t o n i t i s seen). No parasi tes were round in the
intest inal tracts of ei ther nestling.Al l farmers more than 15 km from the colony that were
questioned noted that the proport ion of carcasses they located
that had been consumed by the vultures was part icularly small.A l l e g a t i o n s concern ing v u l t u r e at tacks on sheep are
di scussed i n 3.3.
3.2.5.Foraging behaviour
Proportion of time spent foragingThe numbers of bi rds present in the kloof during each
observation day have been averaged for each calendar month and
are presented in Figures 13 - 16, as an indicat ion of the
proportion of each day that the whole colony spent foraging. During the summer months (October - March), at least 50 per cent had l e f t the kloof by 09h30 (3 months) or lOhOO, and the winter
months (May - August) reflected a shi f t of at least half an hour l a t e r in the day. Conversely, 50 per cent or more had returned by
1 3h30 or e a r l i e r ( three months) or 14h00 in the summer months, and by 14h00 ( f ive months) or later in the winter months, at the
lOhOO time count, signi f icant ly more birds were present in May 1982 than during each month from September through to February
i nc lus ive (Tukey's Mu l t i p l e Range test , 5; level ) . The same
situation held for the llhOO count between May ’ 982 and October, November, December and March. ere were no s ign i f i can t
di f ferences at the 5 per cent level between the other hourly counts.

no.
- igure 13. Monthly foraging pattern of the whole colony : May
1981. The means ( is .d. ) of vul tures present in the kloof at each
time interval are averaged per calendar month. Figures 14, 15 and
16 have the same format as this Figure.

rocoMonthly foraging pattern »t Mr wiiolc < Mi my : June
September 1981.Figure

Figure 15. Monthly foraging pattern!, oi the whnl< colony
October 1981 - January 198:.
igut

3 1
Must vul tures re tu rn e d to the k l o o f a f t e r each day s foraging, as establ ished by the ount: made a t the end of each
day (Figure 17). The average of the montnly standard deviat ions
of vul tures present a t the l a t count o f each day was 2,04
i n d i v i d u a l s . This c o n t r a s t s w i th behaviour observed a t
Aasvagelvlei where, at the few times birds were counted, breeding
birds appeared to stay away from the colony overnight (chapter
f i v e , 5.2.).
Vultures at carcassesEight reports of vultures that were counted on the ground at
a sheep carcass were obtained, and I observed a f u r t n e r iO
i n s t a n c e s . The average number of birds on the ground in al l cases
was 9,8 (range 4-21) . At six of the instances that I observed, a p a i r of White-necked Ravens Corvus a 1b i c o11i s ( L a t h a m ) was
present.I located seven vulture-consumed lamb carcasses, iess than
one month old, in September 1982 on the ‘ arm adjacent to the
Potberg. Of these, only one carcass had bones other than leg
bones present and four of the skul ls , or port ions thereof , had
been removed - presumably by the vultures.A feature of the older carcasses which had been eaten by
vul tures was tnat even from nearby they did not appear to have
been fed on at a l l . Closer examination revealed gaps in the base
of the legs and around the poster ior , where the biros had made
t h e i r en t ry . Four of the 10 o l d e r sheep s t i l l conta ined
decomposing viscera. Also, on every carcass (including a donkey
and Sp ingbok Anti dorcas marsupial i s (Zimmerman) that I had
placed) the skin around the lower jaw was missing. A three day
old carcass that had been completely emptied out weighed 18 kg ; i t s l i v e w e igh t (seven months old) was estimated at 60 kg.. Two
four month old lamb carcasses, attended by vultures weighed
kg and 9 kg.
3.2.6.Contamination of the food urce
Two repercussions of food contamination were considered : the ingestion of small amounts of poison that accumulate and

32
$e
n o . v u l C u r e s
JO
16
F ig u re 17. Total counts of vul tures made in the evening at Potberg and Aasvone I vl el (•). Fo t hn Potberg counts, the mean
(+s.d.) f o r each month is gi v , single counts were made at
Aasvoge!v le i .
1 I
| t I M 1 I I

33
subsequent ly a f f e c t an i n d i v i d u a l ' s b reed ing, and the sudden
ingest ion of a le tha l or near lethal dose (a "poisoning event"),
e.g. Ledger (1 9SU) and M o r r is & Mundy (1981).The range o f poisons used in the area on sheep is presented
in appendix e. The modes of app l ica t ion are via dips or d i re c t l y
e x te rna l ly (e.g. in the p reven t ion o f blow f l y m y ias ’ s). Blow f l i e s ( fam i ly C a l1iphoridae) p r im a r i l y breed in car r ion , but also
lay eggs in l i v e sheep , la rvae then d ig e s t meat ex te rna l l y and
i f the sheep remains u n t re a te d , i t may e v e n tu a l l y d ie (Ledge-'
1979). The c h i e f area o f a t tack i s between the h ind legs ( i b i d.
Thus v u l t u r e s e n te r in g a carcass through a t re a te d an a ; area
would come in to d i r e c t contact w i th the poison.The resu l ts o f analysis of eggs fo r pestic ide contaminati n
are presented in Table 6.
Table 6. The r e s u l t s o f a n a ly s is o f eggs c o l l e c t e d du r ing In -
study period, expressed as mg /kg wet weight.
Sample 0C-: C y - bhc p.p'OOE p . p ’ OOO P.P'COT O ie ld H n
P3S7 0,003 6 , CSS 0,03 0.04 Trace 0,002
PB3 0.003 0,02 0.002 0.008 0,02
PBV 0,49 0 . V 0.02 0.U5
AW , 3 - 0 .3 / 0,07 0.02
Another methou o f dimin ishing blow f l y presence is to t re a t
carcasses w i t h po ison, and t h i s i s the o r i g i n o f a p o t e n t i a l
po ison ing event. One such carcass was l o c f e d 30 km NW o f the
kloof. Of 45 farmers interv iewed, i x admitted to th is pract ice,
bu t on ly " in f requen t ly " .Without exception, each farmer considered the White-necked
Raven a pest fo r i t s al leged attacks on young lambs, lambing ewes
and sheep in d i s t r e s s . This species is r a re l y seen in the area

34
(pers. obs.), a l t h o u g h i t was c o n s i d e r e d common i n t h e p a s t . The
o p i n i o n o f 42 o f 45 f a r m e r s was t h a t i t de c l in ed i n s t a t u s as a
d i r e c t r e s u l t of the po i^n i r . j> ' ircas ses i c t out f jr i t .
m o s t cas> th» noi son "OazzeV (ac t ive ingredient Diazinon) was
s a i d t o have been us u. No fa rmer re po r ted ever having found a
d e a d v u l t u r e , and no poisoned v u l t u r e s were ' ;und dur ing t - i i
s t u d y .
/

J . J . l . f h e foraging range
d i tn t h e i r e f f i c i e n t method of soaring, g r i f f o n vul tu res are
.apable of covering vast distances in search of food. Pennycuick
1972) documented an average speed o f 47 km/hr f o r a fo rag ing
tuppeH's Gr i f fon , and a s im i la r speed fo r the Cape Vu'vure was
ob ta ined from the r a d io - t r a c k i n g study (Boshof f , Robertson &
Norton in prep.). G r i f f o n s breed ing in the Serenget i were found
to t ra v e l up to 150 km from t h e i r nest s i t e to reach the
c o n c e n t r a t i o n s o f food (Hous ton 1976, P en nycu ick 1972).
Pennycuick (1972) est imated a foraging radius of some 110 km fo r
breeding G. ru e p p e l l i i in the Serengeti , and Jarv is et a]_. (1974)
ca lcu la ted a theo re t ica l d a i l y foraging radius o f 130-225 km from
the uo tberg co lony. This re s u l te d in a feed ing range, i .e . an
area w i th in which members obtain most or a l l o f t h e i r food, of ca
200 000 sq. km. J a r v i s e t aj_. (1974) concluded t h a t i t was
possible tha t the colony's requirements were met w i th in an area
of 35 km ra d iu s , a l though the study d id not cons ide r t h a t the
Indian Ocean reduces th is po ten t ia l foraging area to one half .The methods used t o d e f in e the s ize o f the c u r re n t d a i l y
foraging range indicated t h a t i t was smaller than a t ie o re t i c a l
naximum :a. As areas of vu l tu re "presence" indicated by the postal survey were surrounded by areas o f "absence", the boundary was defined
w i th r e la t i v e confidence. Also, the area covered by th is survey
was c le a r ly s u f f i c i e n t to encompass the da i ly foraging movements.
wMth a maximum radius of some 40 km, th is boundary is w i th in one hour's f l i g h t of the colony. Ind iv idua ls may occasional ly exceed
the boundar ies, e.g. the rad io -m arked v u l t u r e ro o s t in g on the
angeberg (Bosno f f , Robertson & Norton in prep.), and o th e r
sightings (e.g. Mart in 1963). These extra-feeding range movements
ire documented in chapter f ive (5.1.).b . T he r a d i o - m a r k e d v u l t u r e fo rag in g behaviour was not
onsidered a typ ica l , as i t was found t o obtain food successful ly
is w e l l as forage w i th o th e r members o f the Potberg colony
Boshoff, oher ' n 1 Norton in prep.).

3b
c. I f the factor of "time spent away from the kloof" per distance
to "feeding" s i te is v - . i d (3.2.1. ) , the average of 13 km to " food" sources (as measured by the individual feeding 'or ays
documented in the k l o o f ) i s o b v io u s ly well wi thin a v u l t u r e ’ s
foraging c a p a b i l i t i e s . As the feeding forays were documented for
breed ing b i r d s , t h i s d is tance i s probably longer : a c t i v e breeders would be expected to return to their nests and nest l ings
ooner (Drent & Daan 1980).In essence, the feed ing range c o n s t i t u t e s the summated
e f f e c t o f the i n d i v i d u a l requ i rem ents of colony members :
compar ison o f co lony requ i rem en ts ( "a b le 4) w i th the expecteo
number o f carcasses ava i lab le to the vul tu res in one year w i th in
the range (3.2.3.) tends to r e f l e c t , t h i s . S i m i l a r inverse co r re la t ions are documented between the home ranges o f ind iv idual
r a p to r s and the num er ica l d e n s i ty o f t h e i r prey reviewed -
Schoener 1968).The shape o f the range s probably i n f uenced *.wv
topographical f e a tu r es of the area, v i z . the or ien tat ion an j
elongated shape of the Potberg, and the presence of the limestone
r idg e to the west o f the colony. Winds are st rongest and
sout hea st er l y in summer (Schulze 1965 and th- shape o ' the
mounta in would p r o v i d e c o n s i d e r a b l e wave I f t over the
surrounding northwestern farmlands. According to a farmer whose
farm l i es on the northern slopes of this ridge and 15 km from the
k l o o f , the v ul tu re s use the r idge as a highway and are o f t r seen f l y i n g close to the slopes, in p a r a l l e l wi th the r idge (J. van Eeden pers. comm.). his is consistent with the expectation
of vultures exploi t ing orographic l i f t , in this case ridge M*t . produced by winds incident on the southern side of the r 4dge. For birds with a p a r t ic u la r ly high w i ng- loa di ng (Pennycuick 1972:. orographic- induced assistance in f l i g h t would be expected t<
influence f lying a ct iv i t y .I t would seem l i k e l y t h a t b i r d s from Aasvogelvlei foray
over farmlands south o f the Langeberg range, as indicated by tin
posta l survey re tu rn s . A lso, a ogle a d u l t was observed 46 km
SSE o f t h i s co lony in August 1982 (A.F. Boshof f in 11 t t .). nes* sightings are actual ly very near the Perdeherg location (chapter
f i v e , 5.2.).

3 7
3.3.2.Food obtained
Quantity of foodD i f f i c u l t i e s were experienced w i th the visual est imation o'
the p ro p o r t i o n o f the colony cons idered to have ob ta ined r ood
during a p a r t i c u la r day. Houston (1972) has discussed some of the
in a c c u ra c ie s w i t h t h i s method, v i z . the accuracy o f the v isua l
sc o r in g and the t im e taken f o r food to be d iges ted v i .e. a b i r d
th a t ob ta ined a f u l l crop in the a f te rnoon o f the p rev ious day
would s t i l l d i s p la y a b u lg in g crop). Roost ing b i r d s tended to
perch facing the c l i f f (away from the observer), thus est imations
were made la rge ly on f l y in g b i rds as they returned. Perched birds
would frequent ly leave perch s i tes and c i r c le in the k loo f , thus
ind iv idua ls could be seen more than once. Inaccuracies a1so arose
when c o n s id e r in g b i r d s t h a t have inges ted 500 g or less : the
crop dis tension is d i f f i c u l t to discern when the b i rd i s perched,
and, not being v i s i b l e as a c lea r bulge when in f l i g h t , is eas i ly
missed. A f u l 1 crop may con ta in some 1400 g o f meat and v isce ra
(Houston 1972, pers. obs.). Houston (1972, 1976:Figure 3) ,
acknowledging these d e f i c i e n c ie s in the method, proceeded to
document average crop con ten ts o f r e t u r n in g G. ru e p p e l1 i %
fo rag e rs to an accuracy o f 250 g and g re a te r . Bear ng these
va r ious p o in ts in mind, the r e s u l t o f b i rd s feeding every 2,5
days i s cons idered a minimum f ig u r e . Given the ina ccu rac ie s of
the method, th is overa l l view of the food s i tua t ion to f £a 600 g ob ta ined every 2,5 days) corresponds to some 2^0 g ob ta ined per
day. This approximates to h a l f the d a i ly adu l t requirement (Komen
1983, Mundy 1982).A more t i m e - s p e c i f i c measure o f the frequency o f food
o b t a i n e d was the number o f t im e s an in c o m in g a d u l t was
documented, at a post- l lhJU changeover, as having obtained food
during a p a r t i c u la r period of the breeding cycle (4.2.2.). During
the 1981 In c u b a t ion pe r io d , a t t w o - t h i r d s o f a l l p o s t - fo ra g in g
changeovers, the incoming a d u l t had ob ta ined a t le a s t 500 g of
food. During the nes t l ing period up to an approximate nest l ing
age o f 100 days in each case) t h i s p ro p o r t i o n was f o u r - f i f t h s .
However during the nes t l ing period, parents would be more l i k e ly
to v i s i t the nest a f t e r feed ing ( to feed the n e s t l i n g ) than i f

they had not fed, and th i s f igu re may be skewed as a r e s u l tCorre lat ions of weight and winglength w i th known age provide
an ind ica t ion o f the quant i ty o f food obtained, as well as "chick
c o n d i t i o n " , du r ing t h a t p a r t i c u l a r stage o f the cyc le (Mundy &
Komen 1983, Newton 1980). Cer ta in ly , the overshoot in body mass
th a t i s found in many n e s t l i n g s is p r i m a r i l y a m a t te r o f f a t
d e p o s i t io n (B rya n t & Gard iner 1979), which occurs a f t e r the
pe r iod o f maximal g rowth . F igures 10 and 11 then in d ic a te a
p e r io d o f s u f f i c i e n t food du r ing the e a r l y to mid n e s t l i n g
period. This i s re f le c ted in the enhanced growth rate during the
ear ly p lase of the period (0-70 days), and above-average ( fo r the
M a g a l i i s b e rg ) accum u la t ion o f reserves dur ing the d e p o s i t io n
phase j f 70-100 days (J. Komen pers. comm.). The l a t t e r was
con f i rm ed by r e s u l t s o f the post-mortem. Th is f a t i s a reserve
in tended to ease the t r a n s i t i o n to s e l f - f e e d in g f o l l o w in g
f l e d g in g , as w e l l as to b r id ge gaps in pa ren ta l food d e l i v e r y
tha t may occur during the nes t l ing period (Drent & Daan 1980).
Qual i ty of foodHouston (1978) cons idered t h a t the p a r t i c u l a r l y leng thy
nes t l ing period of g r i f f o n vul tures was re la ted to an aspect o f
the q u a l i t y o f food they obta ined. Parents have d i f f i c u l t y in
meeting the calcium requirements of the nes t l ing , due largely to
t h e i r food source which c o n ta in s a mere 0,01 per cent ca lc ium
(Houston 1972). Parents then meet the requirements by providing
ca lc ium -r ich food i tems, such as bone-chips, to tn e i r nestl ings
(Mundy 1982). In most areas, "natura l" as well as ranching areas
t h a t con ta in la rge an ima ls o n ly , an in te rm e d ia te is needed to
reduce large bones to an edib le size ; where those intermediates
have disappeared, consequent forms o f bone-diseases have been
documented (Mundy 4 Ledger 1976). At the Potberg colony, only one
instance of bore deformation has been documented, and Boshoff &
C u r r ie (1981) concluded i t p o s s ib le t h a t the d i e t o f the b i r d s
was such th a t bones small enough to swal low are obta ined.
Furthermore, approximately one th 1 o f each year's lambs appear
in September, c e r t a i n l y on one la rge farm ad jacen t to the
Potberg. Thus given t h e i r high m o r ta l i t y rate and the observation
t h ' t v u l t u r e s c l e a r l y v i s i t tne lamb carcasses ( t h i s s tudy,

39
Boshoff , Robertson & Norton in prep.), the t.rmng of th is lambing
season enables the parents to supply the calcium requirements of
t h e i r young. On two occas ions in September, and one in October
1982, I observed groups of less than ten v u l t u r e s on the ground
a t old sheep carcasses (where only bleached bones and some wool
rema-ned). This may suggest that vu l tu res r e v i s i t old carcasses
i n o r d e r to o b t a i n bone p i e c e s , a l t h o u g h t h i s needs
substan t ia t ion .
3.3.3.Foraging ;haviuur
The foraging patterns o f the colony as a whole (Figures 13 - 16) are remarkably cons is tent over the observation period, given
the , a l b e i t s l i g h t , change in co lony numbers (F igu re 17), the
va r ia t io n in hours o sun l igh t between w in te r and summer, and the
change in breed ing s ta tu s o f some 36 b i r d s th roughout the ea
( " t y i n g " them to s i t e s ) . Th is i s in marked c o n t r a s t to Rooks
Corvus frugi legus (L.) fo r example, which spent more than 90 per
cent o f the day l igh t feeding during summer when food was scarcest
(Feare 19721. The standard deviat ion of the mean of each month's
da i ly counts provides an ind ica t ion of the v a r i a b i l i t y observed
dur ing individua days : as a general ru le , th is v a r i a b i l i t y was
greater during the morning than i t was i i the afternoon (except
January). This .a r ia t io n l i k e l y r e f le c ts the birds' dependance on
l i f t ( th e rm a ls or o rog raph ic l i f t Induced by wind) : in the
mornings they would general ly congregate on spec i f ic c l i f f s and
" w a i t " f o r s u i t a b l e f l y i n g c o n d i t i o n s . Those c l i f f s ,
in c id e n ta l ly , were hardly ever used in the afternoon.
The shortest proport ion of t ime spent foraging , in terms of
theore t ica l food ava i lab le , would be expected during the lambing
season (March - May), a period o f above average m o r ta l i t y . This
would r e s u l t in a h ighe r p r o b a b i l i t y o f a food i tem o c c u r r in g
w i t h i n a g iven ra d iu s o f the co lony . B i rds re tu rn ed a t very
s i m i l a r t imes du r ing these montns (s tandard d e v ia t i o n o f the
15h30 means o f v u l t u r e s p resen t o 4.2, 2.7 and 3.0). The length
o f the feeding forays documented during these months (Figure 7)
support t h i s i n d i c a t i o n o f increased food a v a i l a b i l i t y . In
con tras t , the v a r i a b i l i t y in numbers re turn ing at the end of the

40
day was g r e a t e s t in January and Fe b ru a ry , and along w i tn the
reasonably lengthy individual feeding forays documented auring
t h e s e months, t h i s p r o b a b l y r e f l e c t s a r e l a t i v e l y lower
a v a i l a b i l i t y of food. S imi l ar l y , the radio-marked vul ture’s mean
fur thest foraging s i te and mean furthest "feeding" si te in summer
were on the average more than t wi ce as f ar as those in w i n t e r
(Boshoff, Robertson & Norton in prep.).I f 10 b i r d s on average v i s i t carcasses (3.2.5.) and each
carcass suppl ies £a 15 kg (Jarvis et aj_. 1974), then i f each b i rd
removes ca 1200 g (Houston 1976), ro food remains. A du l t merino
sheep we igh c j 50 kg (ATS-J.P. P i e te ^ s e n pe rs . comm.) ;
presumably more than 33 per cen t o f t h i s w e igh t is e d ib le to
v u l t u r e s , e.g. 65 per cen t o f an Impa 1 a Aepyceros melampus is
e d ib le to g r i f f o n v u l t u r e s (Mundy e t a_1_. 1983), then a carcass
w i J obviously supply more food. The f igu re o f 15 kg per sheep is thus a mi mi mum estimate, as an adul t merino sheep could supply
some 30 kg o f food.
3 .3 .4 .Contamination
With regard to contain'nation o f the food source, the e f fec ts
of gradual accumulation are considered f i r s t . Only dead eggs were
analysed during th is study : the resu l ts therefore do not r e f l e c t
the le v e ls f o r a l l eggs (or r a t h e r , the le v e ls in females -
Newton 1979:241). No t i s s u e s were examined. A Potberg egg
co l lec ted In 1975 displayed the highest DDE content Ua 3,5 mg/kg
wet we igh t) :.f v u l t u r e eggs th a t had been analysed by t h a t date
(Mundy e_t a_l. 1982). DDE, a s ta b le m e ta b o l i t e o f DOT, causes
shel l thinning at,, egg breakage as well as Increasing m o r ta l i t y
o f embryos in unbroken eggs ( rev iew ed in New ton 19791. The
r e s u l t s in Ta b le 6 a re l i k e l y t o be be l ow the c r i t i c a l
c o n c e n t ra t io n ; in the Peregr ine F,: Icon F a I c o p e re g r ln u s ,
hatching f a i l u r e due to DDE contamir i t io n s ta r t s a t 15 mg/kg wet
we igh t (Peakal 1 & Kemp 1976). The . o f DDT was r e s t r i c t e d in
1970 and withdrawn from a g r ic u l tu ra l use in 1976 ; tnese levels
l i k e l y r e f l e c t continued use on a much reduced scale, possibly as
a msul t of stockpi 11 ng.
The die I dr in level of P1317 (Table 6) is below the threshold

o f 1 ppm above which upsets to b reed ing occu rred in the Golden
E , g , e A a u m c h o s a e t o s (Uockie e t a i . 19b9). The Potber;, egg
co l lec ted in 1975 contained a higher concentrat ion o f ca V PP
(Mundy e t a l . 1982), a l though t h a t study d id not quest-on the
possible"e77e=ts o f th i s level. In the study of organo-chlor,ne
residues in Golden Eagles in Scotland, d ie ld r in (described as the
most tox ic and abundant residue) was concluded as having caused a
substantia l decl ine in breeding success. In a s i tu a t io n sim, ar
to t n a t a t Potberg, the con tam inan t was inges ted as the eagles
fed on sheep carcasses. A f t e r i t was banned from sheep d ips in
1966, the eagles' breeding success consequently improved (Lockie
e t a l . 1969). The ev idence , a l b e i t scan t , then im p l i c a t e s
" d ie ld r i n as a p o s s ib le cause o f reduced ha tch ing ra te s in tne
past.
3.3.5.A11 egat ions concerning sheep attacks
Much has been documented concern ing a l l e g a t i o n s tn a t
vu l tu res attack l i v e sheep, in p a r t i c u la r lambs and lambing ewes,
and th is aspect is reviewed in Boshoff & Vernon (1979, 1980) and
Mundy (1982). None of the 45 farmers interv iewed remembered such
an ins tance in the p rev ious ten years in the study area, and
v i r t u a l l y a l l d i f f e re n t ia te d between vu l tu res and ravens in t ins
regard (3.2.5.). I did not observe lambing ewes fo r any length of t ime, but was assured by the shepherd of a large farm bordering
the Potberg that vultures "leave the sheep alone'. The only time
I observed v u l tu re s ( f rom some 700 m) in sheep graz ing lands
where no carcass was p resen t , they remained perched f o r 20
minutes some 15 m from the nearest sheep before disappearing. I was l e f t with the impression of them "wai t ing in an t ic ipa t io n !

CHAPTER FOUR : BREEDING BIOLOGY
4.1.Introduction and methods
The broad aim here was to study breeding at the Potoerg
colony through a l l i t s stages in order to determine and
unders tand recrui tment to the populat ion. Breeding at the
Aar-vogelvlei colony was also monitored in both years.
Figure 18. NWI cl i f Nests ac* ve during the study period are
numbered.
All active nest sites and roc,t ing 1 :gi were visible from
the observation point, although cer t a in s i t were more eas i ly
observed than others. Thus nests on N * c l i f f were easier to

42
CHAPTER FOUR : BREEDING BIOLOGY
4.1. In t roduct ion and metnods
The broad aim here was to study breeding at trie Potderg
colony through a l l i t s stages in order to determine and
understand recrui tment to the population. Breeding at the
Aasvogelvlei colony was also monitored in both years.
Figure 18. NWl c l i f f . Nests act ive during the study period are
n.jinoered.
All act 've nest sites and roosting ledges were visible from
the observation point, although c e r t a i n s i tes were more eas i ly
observed th.*n others. Thus nests on NW2 c l i f f were easier to

43
r , v $ r L
F ig u re 19. MW2 c l i f f . Nests actve du r ing the study p e r iod are
numbered.
F ig u re 20. NE c l i f f . Nests a c t i v e du r ing the study p e r iod are
numbered.

monitor than those on NE c l i f f (see Figures 18-20). Nonetheless,
an a t te m p t was made to m on i to r events a t a i l s i t e s and, f o r example, the presence of b irds at a l l s i tes and roost ing ledges
was documented every t h i r t y minutes during each observation day.
The proport ion of each class, i.e. one, both or ne i ther occupant
present at each count during an observation day, was determined and subsequently converted to an arcsine transformat ion 'Schef ler
1969). These f i g u r e s were then compared w i t h i n s i t e s (e.g.
proport ion o f counts of a s ingle occupant versus the f 'op o r t io n
of both occupants) as well as between d i f f e re n t s i tes by M u l t ip le Range tes ts , using a Hewlett Packard HP-85 ca lcu la to r . Individual
vu l tu res were age-estimated using the characters given by Mundy
(1982:43-44) . Observat ions began in May 1981, f o r 12 days each
month re g a rd le s s o f weather c o n d i t i o n s , and ended in May 1982. Thus o b s e rv a t io n s inc luded the t a i l - e n d a c t i v i t i e s o f the 1980
breeding season, a l l stages o f the 1981 season and the i n i t i a t i o n
o f the 1982 season. In a d d i t i o n , n ine consecut ive days of
o b s e rva t ion were conducted from 26 J u ly to 3 August 1982. In t o t a l , 155 days were spent a t the o bse rva t ion p o in t , where one
'day' rep re sen ts the t im e p e r i o d Q7h0Q-16h30. Nes ts were
r e g u l a r l y inspec ted in o rder to ob ta in dates o f f l e d g in g end
observations of juveni les .A l l n e s t l i n g s a t Potberg and Aasvoge lv le i were co lo u r
r ing ed , weighed and measured as p a r t o f the n a t io n a l co lo u r
r i n g i n g orogramme on Cape V u l tu re s (Ledger 19/4). Nests were
v is i te d on f ive other occasions in order to c o l le c t addled eggs or s h e l l f ragments and, on 23 June 1981, to rep lace an egg tha t
an adu l t had knocked out of the nest cup. In 1981, blood samples
were ob ta ined from both c o lo n ie s and on both nest v i s i t s at
Potberg. Smears were prepared by the method described in Greiner
& Mundy (1979) and subsequently ^canned fo r the presence o f blocd
parasites by M.8. Markus.Al though most o f the breed ing p a i rs were not i n d i v i d u a l l y
recognisable, there were a number of r inged breeders present and, at nest no. 63 where both wer i lour r inged, strangers alone at
the nest f o r longer than f i v e minutes were seen on on ly th ree
occasions. Thus unless shown to the contrary, vul tu res observed
at a s i te were documented as the occupants.

45
Add Ied eggs were analysed f o r the presence o f p e s t i c id e
res idues by gas chromatography (A.C. de Kock in 11 t t . ) , and the
thickness o f shel l fragments was determined using a Federal Bench
comparator thickness guage (L.F. K i f f j_n 1 i t t . ). A rad io - t rack ing
study o f the p o s t - f l e d g in g dependence pe r io d o f th ree 1982
j u v e n i l e s was a t tem pted ; the r e s u l t s of t h i s study have been w r i t t e n up separately (Boshoff, Robertson & Norton in prep.).
The breed ing c y c le has been d iv id e d i n t o <ts successive
stages o f p r e - l a y i n g , in c u b a t io n , n e s t l i n g , and p o s t - f l e d g in g
dependence p e r i o d s (Newton 1979) and each i s c o n s id e r e d
separately, as is re ten t ion o f a nest s i te from a previous year's
b reed ing , the stages and causes o f f a i l u r e in b reed ing, and
v a r io u s i n t e r - and i n t r a - s p e c i f i c I n t e r a c t i o n s . Unless s ta ted
otherwise, a l l observations were made a t the Potberg colony.
4.2.Results
4.2.1.The pre- laying period
For r a p to r s in genera l , the main a c t i v i t i e s in f pe r iod
preced ing la y in g in c lu d e n e s t - s i t e In s p e c t io n , n e s t - b u i l d i n g ,
various courtship orocedures and copulat ion (Newton 1979). Due to
the leng th o f the breeding c y c le , c e r t a i n nests d isp layed an
o ve r la p o f p o s t - f l e d g in g and p r e - l a y in g pe r iods (see 4.2.4.),
whereas other pa irs tha t fa i le d in breeding retained t h e i r nest
s i te from the date of fa i lu re .
f ig u re 21 depicts the average occupancy o f 10 nests for the
1981 pre- lay ing period. As observations ceased before eggs were
l a i d in 1982, no s i m i l a r f i g u r e s f o r t h a t yea r are a v a i la b le .
C e r ta in s i t e s th a t f a i l e d in 1981 were a c t i v e in 1982 and were occupied to va ry in g degrees in the i n t e r i m per iod . These b i rds
were c le a r ly re ta in ing the s i te , as they were observed to repel in t ruders from the s i te (see 4.2.6,). Nest s i tes were l e f t vacant
f ive per cent of the observation t ne during th is period (n=10),
as In d ic a te d in F igure 21. A na lys is in d ic a te d s i g n i f i c a n t
d i f ferences a t the 0.01 level in the degree o f occupancy between
nes ts , e.g. two nests on the NW2 breeding ledge. However, two
other nests on the same ledge on NW1 c l i f f were occupied by one

1 It
; ^
5
-1'. ■ ei •man.
IKCUBAT IOii
0 2
MSP TNG
Number- o f v u ltu re s a t n e s t s i t e
' 1’ fhe average level of occupancy of nests durino different period- of the breeding cycle. The number of nests used in the
' : ' ^ f 'ons for each period i< indicated.

Per c e n t
occupancy60 -
40 i
20 -

no. nests : 10 ; 5
ICO
80 -Per cent
occupancy60 ..
20 -
V X k X tX 0 1 2
INCUBATION
20 20
POST-FLEDGING
0 2
Numlier of vu ltures a t nest s ite
Figure 21. The average level of occupancy of nests during d i f fe re n t
periods of the breeding cycle. The number of nests used in the
calcu la t ions for each period is indicated.

4 7
or both adul ts fo r s im i la r amounts of time.
Ac t iva t ion of nest s i tesOf the 17 breeding s i tes tha t were ac t ive in 1981, 14 (82 %)
were a c t i v e again in 1982. Of the a d d i t i o n a l fo u r t h a t became
a c t i v e in 1982, one had been a c t i v e in 1980 (see chap te r f i v e ) . Thus on ly th re e s i t e s were i n i t i a t e d du r ing the o b se rva t ion
pe r iod . An occupied s i t e can be d i s t i n g u is h e d from an area used
fo r roosting by in t rude r repu ls ion b e h a . io u r (4.2.6.).A pa i r o f non-ringed birds was observed a t a s i t e , the real
female occupant o f which had a metal r i n g , on 29 October 1981:
both stood very c lose toge th e r and bent over the s i t e , fac ing
inwards and p i c k in g a t s t i c k s on the su r face . Seven minutes
la te r , the femal* occupant landed at the s i te and repulsed both,
who then repeated t h i s behav iour at another (vacant) s i t e . The
impression gained was of a pa ir "prospecting" a possible future
breeding s i te , some seven months before the next season's average
la y in g date.At another s i te , a pa ir comprising an immature male (est. 3
rd yea r) and female (es t . 4 th year) was observed to copu la te 3b
t im e s between 30 June and 30 December 1981. On most occasions
a f t e r the a t te m p t , both b i r d s would bend over and p ick a t the
"nes t " con te n ts : t h i s was examined on 8 October and found to
c o n s i s t o f a few s t i c k s and f r e s h green s p ra y s - an " in te rmediate" nest structure . The f i r s t in t rude r repuls ion event
observed a t t h i s s i t e was on 20 December. Th is s i t e produced a
f l e d g l i n g in 1982/3, thus i t e f f e c t i v e l y was a c t i v a te d in mid-
1981, i f the parents are assumed to be the same ind iv iduals.A probab le mate replacement occurred a t one s i t e in 1983,
where a colour r inged, six year old female was observed at nest
no.61, from la te December 1982. Th is s i t e was a c t i v e in the two
previous years (both partners adul t and non-ringed). The six year old's sex was determined by observation of a copulat ion attempt
w i th an unknown adu l t male a t a perch po int on 18 May 1982. This
female was hatched a t nest n .57 on NW1 (F igure 1«), and was
often sighted perched on NW1 c l i f f during l (j81 and 1982.

48
CopulationC op u la t io n a t tem p ts , which precede the i n i t i a t i o n of nest
bu i ld ing , were eas i ly noted due to the male's d is t in c t i v e hoarse
c a l l as he g r ip s the female 's r u f f w h i le ba lanc ing on her back (as descr ibed f o r the White-backed V u l tu re in Mundy 1982 ; see
a lso F ig u re 22). With l a t e r a l motions o f h is t a i l , the male
knocks the female 's t a i l from s ide to s ide be fore leaning back and pressing h is t a i l down to achieve co i t ion . While on the nest,
both b i rds spend much t ime bending over and pecking at the nest
con ten ts (e.g. F igu re 22) : t h i s would, in e f f e c t , p o s i t i o n the
female's back fo r the male, who would then mount from her side or
re a r . Of 32 a t tem p ts th a t were t imed, the male was on the
female's back fo r 42 seconds (range 22 - 66 seconds), although in
at least 12 other cases, the male stood on the female's back for
longer than th ree minutes be fore being jabbed o f f . Some males
gave the impression of being ' inexperienced' by moving around on
the female's back and not attempting to juxtapose cloacas. Thus
in 17 o f 31 observed a t tem pts (55 £) a t nest no. 5/ in 1981, no
cloacal contact occurred. This was observed to lesser extents at
o th e r s i t e s , and not a t a l l a t some s i t e s , a l though i t is 'o f ten
d i f f i c u l t to judge' (Green 1976).As w i t h the W h i te -b a c k e d V u l t u r e (Houston 1976), no
d i s c e r n i b l e d is p la y occurred a t the nest be fo re or after- c o p u la t io n , a l though p a i rs were occas iona l ly seen to al lopreen
necks g e n t l y , both be fore and a f t e r the event ( f o r the l a t t e r ,
see Mundy 1982). A form o f "scapu la r a c t io n " d is p la y (B e r ru t i
1981) g e n e ra l l y occurs j u s t a f t e r a b i r d has landed at a s i t e ,
where i t tu rns i t s head and places i t s b i l l a t the j u n c t i o n of
i t s wing and body. Less o f t e n , both members o f the p a i r were
engaged in t h i s behav iour, and i t i s o f te n accompanied by
autopreening.In the pe r iod May 1981 - May 1982 ( 154 observa t ion days),
1783 c o p u la t io n a t tem p ts were observed, of which 17U6 (95. / t) occurred a t nes t s i t e s (Table 7). A peak in frequency occurred
j u s t p r i o r to the average lay ing date o f each season, l o r seven
successfu l nests in 1981, 67,9 per cen t o f observed a t tempts occurred du r ing the p r e - la y in g per iod (Table 8) ; t h i s inc ludes
one s i t e (nes t no. 61) where the n e s t l i n g disappeared at 111/2

A aSi te No. be • re T o ta l oN '.ervetl A /e T o ta l observed
egg laid "d> 31-Dec 81 Jan 82-May 82
7 112 113 E 9 9 .1 38
13 31 35 E 88 .6 18
16 42 53 i 79.2 54
6 30 39 E 76.9 34
26 49 61 E 80 .3 53
44 55 E 80 .0 64
63 59 59 E 100 22
55 44 45 £ 97.8 25
17 48 57 E 84 ,2 S '
61 17 30 £ 56.7 54
30 38 49 E 77.5 5
13 - 39 49 E 79.5 48
57 31 31 E 100 46
29 39 57 E 68 ,4 4
45 21 21 E 100 0
3 54 54 E 100 1
2 32 - 30
50 a • - 25
25 24 - 71
64 0 19
8 117 - - 21
57* 34 * -
ROOST 56 21
POiMTSJ * 1079 2 * 04
Table 7. C opu la t ion a t tem p ts observed in the co lony. This Table
also documents the s i tes that were act ive during the study period (E : egg l a i d ) . Nest no. 57* i s a ja c e n t to nest no. 57, bu t has
never known to be act ive in the h is to ry of the colony.

days o f age.Three hundred and seventeen ( 1 \ 8 - i ) a t te m p ts occurred at
s i t e s o r ro o s t p o in ts where no egg was produced ; e.g. a t nest
no. 8. 138 a t te m p ts were observed w i t h o u t an egg appearing
e i th e r year. Eleven attempts (0,6%), where a male tha t had jumped
onto an assumed female's back was aggressively jabbed o f f , that
occu rred on r o o s t p o in t s , were c lassed as " r e s i s t e d mat ings"
(Estep & Bruce 1981). S i m i l a r i n c i d e n t s , a l though not a t the
nesting area, have been observed in White-backed Vultures (Mundy
1982). In s ix o f the e leven cases, one o f tr.e b i r d s was from a
known nest s i t e (4 males, 2 females). These contrasted w i th three
cases o f "c ross mat ings" (0,05 %), where the male achieved
cloaca! contact and was not aggressively jabbed o f f . For example,
nest no. 13 female was thus involved (26 June 1981) some s ix days
be fore la y in g an egg, and on 23 June, nest no. 17 male was observed to copulate w i th an u n id en t i f ie d adu l t female at a roost
point whi le h is partner incubated the egg.
Table 8. C op u la t io n a t tem p ts observed per p e n o . of breeding
c y c le . The f i g u r e s f o r nest no. 61 are in b racke ts as the n e s t l i n g d i s a p p e a r e d b e fo r e f l e d g i n g , and tne t o v 1 f o r
successful s i te s excludes th is s i te .
' le s t
617
1618
29
4
26
61
30
13
57
55
45
3
63
7
•lean
p r e - l a y . A /E A
31
47
42
37
38
44
47
16
38
27
30
46
2145
49
11 2
76
63
39
73
(59)
69
56
69
In c .B
88215
06 0 3 2 0 0 0 0 0 0
B/E N e s t ! C/E prOP 0/E " o t a ;t C 5 0 %
20 0 3 2 5 41
11 20 27 0 3 75
2 15 14 49 45 108
_ - 38
• - 43
0 13 22 5 60
• ■ » . 53
0 ( ID (4 1 ) 27
- 38
* 29
30
. ■ * . 46
* - 21
. - 45
0 3 20 23 71
0 3 3 1 1 116
5S I V , 155 62,
T o ta l f o r s u c c e s s fu l s i t e s : 471

51
Nest-bu i ld in t ,No sign o f nest bu i ld ing at any s i te was observed before the
end o f March 1982 and a t on ly two s i t e s , success fu l i n the
previous cycle, were there any remain of the nest s t ruc ture . In
fac t , by the end o f December 1981, nest materia l on the s i te s of
a l l f a i l e d breeding attempts had disappeared, presumably removed
by wind and ra in .C o l lec t ion o f nest material in earnest was f i r s t observed on
2 5 A p r i l 1982, some ten weeks b e fo re the average la y in g date. Th is i s s i m i l a r to u a p p e t - f te e d V u l tu r e behav iour (Pennycuick
1976). Branches comprising the nest s t ruc ture were co l lec ted , in
most cases, from two separa te areas on the s lopes o f the k lo o f
and ca r r ied back to the nest in the b i l l (contra Houston 19/6:20
fo r Ruppell 's G r i f fo n , where a l l the branches used fo r bu i ld ing . e r e seen to be s to le n from o th e r s i t e s . ) . F i l c h i n g was seen on
two occas ions in the p r e - l a y in g pe r iod in 1981 and on severa l
occasions in the same period in 1982. In t o t a l , nest materia l was seen to be taken from o th e r s i t e s on 19 occas ions , by occupants
o f e i g h t d i f f e r e n t s i t e s . Only tw ic e was the s i t e occupy
p re s e n t when the m a te r ia l was taken, and on 13 occas ions tne
materia l was taken from an adjacent s i te .At s ix nests where male and female cou ld be d i s t i n g u is h e d
(sexes de term ined by ob se rva t io n of c o p u la t io n , * t l e a s t one
partner ringed), the frequency o f c o l le c t in g forays by e i th e r sex
could be determined. Col lect ing forays were mainly determined for
th ree nests o n ly - those s i t u a te d on NW2 c l i f f where the
c o l le c to r 's sex could be determined with greater c e r ta in ty . Of 98
collections, males co l lec ted 75 t imes, s i g n i f i c a n t l y more Cb «)
than d id females ( \ 2,,=" expected values t e s t , p<0,01) al though
at one nest, the difference was not significant (p=0,0b, n » 31). The female g e n e r a l l y packed the materia l into the s t r u c t u r e , although both sexes were often seen bending over and rearranging
the contents (Figure '<■'?).

52
/

53
4.2.2 The incubat ion period
In 1981, 356 nest-days of observation of 16 nests w i th eggs
were made, and I accumulated a fu r the r 99 nest-days of 17 nests
during an e igh t day consecutive period in 1982.No double c lu tc h e s (Mundy & Ledger 1975, Mundy 1982) or
rep l acemen c c lu tc h e s were recorded dur ing the ac tua l study
p e r io d , a l though the egg at nest no. 55 was la i d s i g n i f i c a n t l y
la te r (on 14 July 1980, which was exact ly two standard deviat ions
l a t e r ) than the es t im a te d mean o f 5 June f o r t h a t year and was
thus l i k e l y to have been a replacement clutch.For 16 in c u b a t io n per iods in 1981, which inc ludes one egg
lo s t a f te r 41 days, one adul t was a t the s i te at 9 \ 7 p e r cent of f i e counts (range 91,2%-100%) and s i tes were unoccupied only 0,09
per cen t o f t h i s obse rva t ion p e r io d , an average of 359 t h i r t y -
minute counts per nest. T h s behaviour i s ind icated in Figure 2).
A t a l l nes ts , the frequency o f a s in g le a d u l t a t the nest was s i g n i f i c a n t l y h ig he r than f o r any o th e r stage o f the breeding
cycle (Sheffe's te s t , p<0,01). A partner remained at the s i te 2,0
per cen t o f tne t im e (range 0%-8,8%). B i rds g e n e ra l l y Incubated the eggs very t i g h t l y and on only one occasion d id one a d u l t
leave an egg unoccupied , fo r 182 minutes, fo r no apparent reason
(bu t see 4.2.3.).O f 3n average o f 78,5 c o p u la t io n s ooserved a t s i x nests
throughout the breeding cycle, 5,3 per cent occurred during the
incubation period (Table 8).The resu l ts of analysis of eggs for pestic ide contamination
a r e p r e s e n t e d i n c h a p t e r t h r e e ( T a b l e 6 ) , and t h e t h i c k n e s s or
s h e l l f r a g m e n t s measu r ed a r e g i v e n i n a p p e n d i x d.
A W h i t e - n e c k e d Raven was o b s e r v e d t o c a u s e an i n c u b a t i n g
a d u l t t o s t a n d , b u t r e m a i n on i t s n e s t , and j a b a g g r e s s i v e l y as
t h e r a v e n l a n d e d on a n e s t i n g l e d g e a t A a s v o g e l v l e i . Two r a v e n s
t h e n l anded 1,5 m above a n o t h e r i n c u b a t i n g a d u l t , w h i c h r e m a i n e d
t i g h t on i t s n e s t . One s u b s e q u e n t l y l a n d e d a t a t h i r d n e s t and
pecked a t t h e o u t l y i n g n e s t m a t r i a l : t he a t t e n d a n t a d u l t j a b b e d
a g g r e s s i v e l y b u t r e m a i n e d i n an i n c u b a t i n g p o s i t i o n . A t l e a s t
f o u r r a v e n s w e r e s ee n o v e r the Potberg k l o o f , b u t no p r e d a t o r y
a t t e m p t s we r e o b s e r v e d .

54
Laying datesLaying dates fo r 1981 and 1982 were determined by checking
the colony re gu la r ly , although precise dates fo r three nests were
no t ob ta ined in 1982. Dates f o r 1977, 1979 and 1980, as w e l l as
the dates o f la y in g in both 1981 and 1982 a t Aasvogel v1ei were
e s t im a te d us ing an average in c u b a t io n pe r iod o f 56 days and a
known-age / wing leng th curve (Mundy 1982). Wing leng ths o f
nes t l ings measured before 1981 were obtained from CDNEC records.
Thus on ly l a y in g dates o f s i t e s t h a t produced la rge n e s t l i n g s
were ob ta ined f o r years o th e r than 1981 and 1982. These lay ing
dates are de p ic ted ir. F igure 23, and s i g n i f i c a n t d i f f e re n c e s
(Student's t - t e s t ) between the means of any years are indicated;
both the Potberg and Aasvogelvlei means are compared here.
PorecnG (re!
ISSi
121!
i)4 =
* " PB y e i r s 1 * , A / V ' i . IZ *
1220
122
1227if
net .tgez"
1 9 8 1 "
M S V V i f V £t (*VV)
tssz
1981
MAY 18 Z«I j In Jimc m Mr iz 19
F igure 23. Dates o f egg lay ing a t Potberg and Aasvogel v l e i . The mean (±s.d.) and range are shown, as w e l l as s i g n i f i c a n t d i f f e r e n c e s (1: leve l : y e a r x < ; 5 . leve l : y e a r x ).

55
In 1577 on ly , did the l in k o f higher r a in f a l l and la te r mean
lay ing date not occur, although the di f ferences are very s l ig h t .
C on s ide r in g t h i s r e l a t i o n s h i p however, in 1980 alone was the
preceding s ix months' p rec ip i ta t ion s ig n i f i c a n t l y less than tha t
o f 1981 ( t - t e s t , p=0,05). The extreme example w is the 1981 mean
date , which was s i g n i f i c a n t l y l a t e r than any o the r year a t
Potberg (F igu re 23), and s i m i l a r on ly to the mean date for the same year a t Aasvogelv 1 ei ( t , - t e s t , 20 t i . f ,, p=0,Q9). 1 he overai i
average la y in g date f o r the f i v e years is 17 June (no. eggs*58,
s.d.*17,5), and th is is c le a r ly la te r than the early-May average
fo r the Sl erpoort , Magaliesberg colony (Mundy 1982:171) and the
s im i l a r average fo r the Colleywobbles, Transkei colony (Vernon et
ajL 1982).The fo l low in g considers the evidence as to whether ra is ing
an o f fso r ing successful ly might delay egg-laying in the fo l low ing year . A l l 1981 eggs were l a i d s i g n i f i c a n t l y l a t e r tnan in 1980,
except the l i k e l y replacement c lu tch mentioned. Of the three long
p o s t - f l e d g i n g dependence per iods in 1980/1, an egg was l a id at
' h° same s i t e be fo re the mean 1981 la y in g date on one occasion and a f t e r i t on another , n e i th e r date being s i g n i f i c a n t l y
d i f f e r e n t ; the t h i r d s i t e was not a c t i v e in the subsequent season . A f t e r the p o s t - f l e d g in g dependence periods observed in
1981/2, eggs were l a i d a t the same s i t e s be fore the mean 1982 date on fo u r o f the f i v e occassions and s i g n i f i c a n t l y l a t e r by
the confirmed same adul ts (Robertson 1983b) in one instance. At
the s i t e w i th the longest 1981 PFOP, the fo l low ing year's egg was
la id oefore the mean date.The most accura te t imes o f la y in g were ob ta ined f o r three
eggs in 1981, where both were l a id between 16H30 and 07h30 the
nex t day. At one o f tnese s i t e s (nes t no. 63), the same co lou r
r inged female l a i d an egg in 1982 between 17h30 and 15h00, on
exact ly the same date as the previous year.
Changeovers, and incubation of the ggChangeovers were general ly accomplished w i th in about three
minutes o f the mate's a r r i v a l a t the nest, a l though c e r ta in
p a r tn e rs gave the impress ion of being "keen" to r e l i e v e an
i n c u b a t o r w h ic h w a s " r e l u c t a n t " to lea ve . In such c a s e s ,

56
nanycvey " '1 ' wau- ,ad oeen yewily nudyed, f f r,h( nest ; t h 1 s behaviour wa: more f r e q u e n . du r ing the
nestr ing period. During what was most l i k e l y the f i r s t changeover
i t nest no.16, the incoming adul t al lowed the egg to r o l l cut of
the cup by s tand ing on the r im. A f t e r a few feeb le a t tem p ts at
re p la c in g the egg w U h i t s b i l l , i t con t inued to " incuba te " the
empty nest. Dur ing the next 90 m inu tes , the b i r d a t tempted on
hree occassions to replace the egg, which lay some 8 cm from the
r 1m. I rep laced the egg a t ii>h40, a f t e r i t had been exposed fo r
4 ,0 m in u te s ( c l e a r w e a th e r , 9 -13 k n o t b re e z e ) . T h is egg
subsequently produced the begging juven i le in Figure 42 A pair
of C a l i fo rn ia Condors Gymnogyps cal i fo rn ianus was also observed
to al low the egg to r o l 1 ou t o f the nes t , a l though in th a t case
the egg shattered (Anon 198?). This was possibly the fate o f the
egg from nest no. 25 in 1982 i see 4.2.5.).A t o t a l o* 186 changeovers o f b i r d s incuba t in g eggs were
,oted in 198), averag ing 11,5 per s i t e ,o r 0,5 per nest-day of
observa t ion . However, a number o f changeovers a t the o u t l y i n g
s i tes were probably missed: i f the NW2 ledge (d i r e c t l y opposite
the observation point) is considered in i s o la t io n , 61 changeovers
were documented for four incubation periods during 38 nest-days,
i .e . 0,7 changeovers r ' nest per day. As an extreme example, a t me s i te , three changeovers on the same day were observed on two
d i f f e r e n t days, Changeovers occurred in the morning, be io re a
jay's foraging or, in the afternoon, a f te r foraging. The e a r l ie s t nstance noted where the incoming oartner had fed that morning
„ as : in30 : i f the changeovers are then d iv ided i n to p r e - 1 ln30
61, 33%) and pos t -1 lh30 groups (125, or 67%), then the
incoming a d u l t had ob ta ined food on 64 per cent o f the l a t t e r
(8,6% cases unknown). The overa l l average changeover time was 12 hrs 54 mins, 50 mins e a r l i e r than t h a t documented f o r the
■Jkeerooort, Magal iesberg colony (Mundy 1982:16:). 1 he two average
changeover t imes o f LOhlO and H h l 2 probably r e f l e c t tne
s i tua t ion more accurately, and the fo u r -h o u r separa t ion agree:-
v i th the average o f the feed in orays (3 hrs 42 mins) ob ta ined
-or the colony (3.2.2.).incuba t ing b i r d s o c c a s io n a l ly stood up to s t r e t c h , f la p

i t s long a x is , Mundy 1982: 161), and tne longest pen,: 1 net
cases tha t the egg was exposed was 4 minutes :1 istanc meo,
average 1,2 mins).Measured incubation s t in ts , i.e. the length of time betweer
two changeovers a t a s i t e , were obtained whenever possible when changeovers were observed during consecutive days of observation.
As w i th changeovers, these measured s t i n t s are divided in to those where two changeovers occurred w i t h i n one obse rva t ion jay
( "w i th in -day") , and longer s t in ts . In 1981, the mean "w ith in-day
s t i n t was 3 hrs 15 mins (n=44) and t h i s was not s i g n i f i c a n t l y d i f f e r e n t to the 1982 f i g u r e o f 3 hrs 31 mins ( t - t e s t , 60 d . f . ,
p=0,30). The mean o f 1981 "long" s t in t s was 26 hrs 21 mins (n=72i and th is was s ig n i f i c a n t l y longer than the 1982 mean of 22 nrs 35
mins ( t - t e s t , 98 d . f . , p=0,01), due to the i n c lu s io n o f 11 ex tra long" s t i n t s (mean = 44 hrs 3 mins, range 40 hrs 45 mins- 48 hrs
8 mins). I f these 11 values are excluded, the 1981 mean of 2^ nrs 48 mins is not s ig n i f i c a n t l y longer than those obtained in 1982
( t - t e s t , 86 d . f . , p=0,37). In f i v e o f these e x t r a - lo n g cases, the ind iv idua l concerned was id e n t i f i a b le ( r in g e d ) , c o n f i rm in g t h a t
no changeover had been missed.Sexes cou ld be d i s t i n g u is h e d a t f i v e nests due to tne
presence o f e i t h e r a metal r inged ( two nests) or c o lo u r r inged p a r tn e r . I on ly those ins tances where the r i n g / s w a s / w e r e
c l e a r l y r e s ig h te d a t both changeovers, are the correspond ing
s t i n t s used in "a lys is of any sex-related di f ferences in length.
The mean f o r males (1981 and 1982 f i g u r e s combined) was not s i g n i f i c a n t l y longer than the me i fo r females ( t - t e s t , 30 d.t..
p=0,32), and the observed f requenc ies o f e i t h e r sex was no”
s i g n i f i c a n t l y d i f f e r e n t (X2"*" expected values tes t , p=0,72).Five addled eggs ocurred in two years , three dur ing -ne
o b s e rv a t io n year. In two o f these three cases no change ’ n
incubation behaviour a f te r the expected hatching date was noted
For example, adults a t nest no. 29 continued to i -ubate normally f o r 18 days a f t e r the expected ha tch ing date, un t i 1 I col lec ten the egg f o r a n a ly s is . At one n< , t however, the incu ba t in g adui t
tended to stand over the egg, ra ther than s i t on i t , from some i J
days a f te r the expected hatching date. Twenty four days a f te r tne
expected hatching date, the Incumbent adul t (male) deserted th is

58
qyy f o r 152 w ins , re tu rn e d to stand Mud,1 i t to r .ibb w ins , to
leave i t unattended th a t n ig h t (I c o l l e c t e d the egg the next
day).
Length of the periodA t s ix s i t e s in 1981 where both lay ing and ha tch ing dates
were accurately obtained, an average incubation period of 5/ days
(954 C.L.s 55,6-58,4) was documented.
4 .2 .3.The nestl ing period
A t o t a l o f 377 nest-days o f obse rva t ion o f ten s i t e s wi th
n e s t l i n g s were made in 1981 and 15 nest-days, a t fou r nests , in
1982. In 1981, seven nest l ings l ived longer than 11 days and six
fledged, and of ten nestl ings colour ringed (average age=76 days)
in 1982, seven f ledged (see 4.2.5.).E leven per c e n t o f c o p u l a t i o n a t t e m p t s observed a t
successfu l s i t e s through the 1981 cyc le occurred dur ing the
n e s t l i n g pe r iod (Table 8). The mean ha tch ing date f o r the years1977 and 1979 - 1982 was then 14 August, c a lc u la te d using the
mean laying date of 17 June and a 57-day incubation period.
Parental care of the nestl ingDur ing obse rva t ions o f the seven nests in 1981, n e s t l in g s
were l e f t unattended fo r 1,2 per cent o f the observed n e s t l i n g
pe r iod (range 0 ,1 4 -4 ,5 * ) , and one n e s t l i n g was unattended for
s ig n i f i c a n t l y more time than three other nests (Scheffe s test ,
p<0,05). Six nestl ings were seen unattended at least once during
the pe r iod ( i .e . one never) , and the average age at the f i r s t
such i n s t m c e was 71,7 days (range 41-93 days). N e s t l in g s were
accompanied by one adul t 76,4 per cent of the counts. Both adults
were at the nest 22,4 per cent of the time: in a l l cases th is was s ig n i f i c a n t l y greater than dun g the incubation period and, in
three of the f ive cases, s ig n i f ic a n t ly less than during the PFDP
(Scheffe 's t e s t , p<0,05) (F igure 21).During t h i s pe r io d in 1981, sexes were d i s t i n g u is h a b le at

59
onlv one active nest and both parents attended the nes t l ing for
s i m i l a r leng ths o f t im e I t - t e s t , Z l d . f . , p=0,43). Both male arid female attendance s t i n t s were s i g n i ' l c a n t l y shorter than those
obtained during the incubation period ( t - t e s t s , p<0,01>.The mean " long" attendance s t i n t o f 21 nrs 56 mins (n-71)
documented was not s ig n i f i c a n t l y shorter than the s im i la r mean
o b ta in e d du r ing in c u b a t io n , i f the 11 " e x t r a - lo n g " s t i n t s are
excluded ( t - t e s t , 129d.f„ p=0,09). I f these values are included,
the d i f fe rence becomes s ig n i f ica n t . The mean "w i th in day s t m t
o f 3 hrs 39 mins (n-ST) was not s i g n i f i c a n t l y longer than the
f i g u r e f o r tne in cu b a t io n pe r iod ( t - t e s t , 95d. f . , p=U,V). ■ "e average changeover t ime documented fo r t h i s period is then 12h29,
which i s s i g n i f i c a n t l y e a r l i e r than t h a t documented f o r the incubation per iod ( t - t e s t , 3 7 5 d . f „ p-0,491. Of 193 changeovers,
65 8 per cen t occurred a f t e r Uh30 and on 86 per cen t o f these,
the incoming adu l t had obtained food (4.6% of cases unknown).For i t s f i r s t few weeks, the nes t l ing is brooded closely by
the a d u l t , making the p rec ise t im e o f ha tch ing d i f f i c u l t to
d is c e rn . There i s subsequently a g rada t ion in attendance
behav iour o f the paren ts , p a r t i c u l a r l y a f t e r the n e s t l i n g i s
do rsa l ly feathered. This behaviour varied between nests, e.g. at
one s i t e a d u l t s o f te n " incuba ted" the empty nest w h i le the n e s t l ing stood nearby. A four month o ld nest l ing wandered 2m to
an ad jacen t s i t e c o n ta in in g a f i v e month o ld n e s t l i n g on th ree
occasions: both pecked gently at each other's necks before being aggressively jabbed at by an attendant adult . S im i la r behaviour
was never observed f o r a d u l t s o f ad jacen t s i t e s , b a n n e rs
con t inued to b r in g f resh green sprays to the nest du r ing t m s
period.Most feed ing bouts were mouth to mouth, w i th tne n e s t l i n g
a c t ive ly c a l l i n g and tapping the parent's b i l l whi le drawing food
from i t s mouth. On a t leas t seven occasions, parents tha t had nut fed were observed to go through the motions of r e g u r g i t a t i n g w i t h o u t any food appearing, a l though in o bse rva t ion i t is
d i f f i c u l t to be accura te about t h i s (chapter th re e , 3.3.,. he
"ex-attendant" partner consumed food tha t was regurgi ta ted onto
the nest by the forager on six occassions.Nestl ing development is described in Mundy (196^), and tne

60
= = = = = = = = - “ ' “ i -
#1 #iiiSi
1n 1979 and, f i n a l l y , absent In 198
CONEC records).
mm

Figure 24. Simuli id in fe s ta t io n o f a 47-day old nest l ing.
400 -
e 300
200 -
— | ------- - I ----------- T ! ......... i ' • I
?6 77 78 79 80 81 82
Year
' igure 25. Simul i id presence re I ' ted to the amount o f ra in in the ; i x months preceding September i#). The degree of in fe s ta t io n is
:1assed as heavy (H), medium (M) o r absent (A), and the mean
$ mount of r a in f a l l fo r these months is shown (x).

Length of the periodWhere no days of observation preceded the day the f ledg l ing
was f i r s t seen away from the nes t , the f le d g in g date was not accurately known ( four cases known to w i th in f i ve days). Where at l e a s t two o b s e rva t ion days preceded the date o f f i r s t o b s e r v e d
f l i g h t , t h a t day was taken as the f le d g in g date. Some f le d g in g
dates were on ly known to w i t h i n an accuracy o f th ree days. In 1981 mean nest l ing periods were 148,0-150,3 days (range 139-171)
days). Both these means were s ig n i f i c a n t l y longer than the 1982
mean of 137 days (range 125 - 147 days) ( t - t e s t fo r mean of 148, l l d . f . , p=0,04).The in d iv i d u a l w i t h the 125-day n e s t l i n g pe r iod
died an est imated three weeks a f t e r f ledging. I t was found, under
f o l i a g e in the k l o o f r i v e r , an es t im a ted two weeks a f t e r i t s
death (P.v.d. Westhuizen pers. comm.). The next shortest nestl ing
period I documented was 136 days, and th is b i rd survived at least
f i v e months.The mean fo r a l l ava i lab le period lengths is then 142 or 143
days ; th i s compares well w i th Mundy's (1982:188) est imate o f 140 days. The mean 1981 f le d g in g date was 21-23 January 1982, and
th is contrasts w i th 28 December 1982 fo r 1332 f ledgl ings.

63
4 . 2.4.The p o s t - f l e d g in g dependence period (Pf 'DP)
A f t e r f l e d g in g , young ra p to rs may v i s i t the nest s i t e and
remain dependent on th e i r parents for what is known to be a very
var iab le length of time (Newton 19/9). In th is study, termination
o f the period was taken as the date o f the las t recorded v i s i t of
the juven i le to i t s natal nest s i te . Parent- juveni le in te rac t ions were observed at the breeding area only ; i t was not attempted to
observe any such in te rac t ions , i f these occur, at feeding s i tes ,
as i s the case e.g., in the Black V u l tu re Coragyps a t ra tu s
(Cathartidae) (Jackson 1975).From 8 May 1981, 118 PFDP-days o f observation were made of
three 1980 juven i les (i .e. nestl ings in 1980), jnd 137 PFDP-days o f s i x j u v e n i l e s in the f o l l o w in g season. A minimum o f tnree
hours of observation every day between 21 December and 13 January
1983 (23 consecutive days) were made o f 1982 juven i les around tne
t ime of f l e d g in g . J u ve n i les are c l e a r l y d i s t i n g u i s h a b le in the
f i e l d by various plumage characters (e.g. black-brown eye colour
and a long, lanceo la te d and streaked r u f f ) , as documented in Mundy (1982:43). Colour r inged as n e s t l i n g s , each j u v e n i l e was
then in d iv id u a l ly recognisable. The observed PFDPs are depicted
1n F igure 26.
Length of the periodIn 1981, th ree j u v e n i l e s con t inued to v i s i t the nest s i t e s
for 221, 194 and 175 days a f te r f ledging, although they were la s t
seen r e c e iv in g food a t 210, 189 and 159 days re s p e c t i v e l y . The
PFDPs of three other juven i les did not exceed 11 and 133 days in
length (i .e. two of these juven i les were resighted in tne k loo f ,
but never seen at th e i r nests).In 1981, f ive juven i les (of six that fledged) held PFDPs of
longer than 13 days: 15-19 days, 22-25 days, 83 days, 12U-124
days and 139-142 days. The s i x t h j u v e n i l e (nes t no. 7) was la s t resignted in the k lo o f s ix days a f te r f ledging, and one day after-
f ledging at i t s s i te .Three 1981 j u v e n i l e s from Aasvoge!v le i held pe r iods o f no
longer than 82, 84 and 85 days (estimated ages from wing lengths
a t r i n g i n g , us ing a 143-day n e s t l i n g pe r iod ) , as they were

55
63
13
s.■
v s/ l
tmm ^ s s 'B le i f f o f *c to « lin e r t# -
n J J A S 0 " N D J F M AM J J
1981 1982— «— :— i— :— r— i— i— ; ! : i : f
i
AS ON D J F M A M J J
F igu re 26. Dlanram o f the breeding cyc les o f I n d iv id u a l nests
(numbered on le f t ) . The Incubation period is represented by dark
b lo c k s , the n e s t l i n g per iod by empty b locks and the PFDP by
hatched bloc s.

1980-i— i— i— r
30
50
55
1981
63
13
’H e ieu * aessie?* *»•«:*> #ciw4iamofif
"'l'-
M J J A S 0 NO J F M AM J ' J
1981 1982-i r
16
—
17
63
- I I L . > ■ 1 :__;---L _LJ A S ON D J F M A M J J
F igu re 26. Diagram o f the breeding cyc les o f i n d iv i d u a l nests
(numbered on le f t ) . The incubation period is represented by dark
b lo c k s , the n e s t l i n g pe r iod by empty b locks and the PFDP by
hatched blocks.

resighted at Fotoerg from 6 Apr i l 1982.Although observations were made or 1982 juveni les, no PFDP
lengths were obtained, except fo r those tha t were recovered dead.
Only one j u v e n i l e was re s ig h te d in the k l o o f f i v e months a f t e r
f l e d g in g (A.F. Boshoff p-j ts. comm.).At nest no. 30, the 1980 PFDP over lapped w i t h the next
season's incubation period sy two days; the juven i le v i s i te d tne
s i t e eleven days l a t e r but was ag g re ss ive ly chased o f f by the
incubating adul t. At nest no. 55, there was a s im i la r overlap of
fo u r days, as i 1 l u s t r a t e d in F igure 2 . No feeding was observed
a t any nest du r ing t h i s pe r iod o f ove r lap . For the th ree 1981
PFDPs o f longer than 82 days, the next season's egg was , a id 41
days a f te r the periods had ended (range 6-79 days).
Behaviour o f juven i lesThe presence o f j u v e n i l e s a t t h e i r nests i s d isp layed fo r
th ree nests in F igures 27 - 29. On average, the f i v e 1981 juven i les were at t h e i r natal s i tes at 36 per cent of the counts
made on days dur ing t h e i r re spe c t ive PFOPs. I f the th ree long periods are considered alone, th is f igure is 26 per cent. For the
th ree long PFDPs, j u v e n i l e s were alone a t t h e i r s i t e s 1,8, 1,9
and 2,5 per cen t of the counts.I n d i v i d u a l s were o f te n seen to land ( c lu m s i l y ) a t var ious
p laces on the slopes o f the k l o o f , where they would spend t ime
perched. During the course o f the s tudy, two j u v e n i l e s were
e x t r i c a t e d from undergrowth t h a t had trapped them, and i t is
l i k e l y that a t least one other had died as a re s u l t o f landing in
vegetat ion. One of those captured had landed on top of the parked
Combi at the mouth o f the k lo o f ! Thus i f a j u v e n i l e was not at
i t s s U e or v i s i b l e in the k l o o f , i t cou ld not n e c e s s a r i ly be concluded as foraging. A rad io- t rack ing study s i n i t i a t e d with
an aim o p d o c u m e n t in g the t r a n s i t i o n to f u l l f o r a g i n g
c a p a b i l i t i e s , but lapsed a f t e r two marked indi <duals died and
the t r a n s m i t t e r on the t h i r d stopped fu n c t io n in g (Boshof f ,
Roberts t . i & Norton in prep.), -i ven t h i s c o n s id e r a t io n , the
number o f days dur ing which j u v e n i l e s were considered to have
foraged, as a p ro p o r t io n o f the number o f days observed, are
presented in Table 9. As i f to emphasise the v a r i a b i l i t y in

I
i lDAYS DURING PFOP
■iqure 71. Diaqram depict ing the periods when <■ juven i le (nest no.vas at Hs nest during i t s PFDP (shaded areas). Each empty block representsm observation day, and days of no observation are eq l l y spaced between
these; Figures 29 and 30 have the same format. Aggressive interactions)etween the juven i le and i t s parents were observed at the points indicated
bv arrows.

I t
t i m e >f day
hflfi
i 't
13
in
hays miRiNG p rn r
L
I• inure 28. Diaqrar' depicting the periods when a juvenile (nest no. 16) was a, , nest durinn its PFDP (see Figure 28). An aggressive interaction
ween un juvenile and its oarents was observed at the point indicated.
'm

DAYS CUR INC PFDP
Figure 29. Diaoram depict ing the period, when a juven i le (nest no. 63) was at i t s nest during i t s PFDP (see Figure 28). Observations ceased
before termination nt th is PFDP.

behaviour during th is period, nest no.7 juven i le returned to the
colony w i th a f u l l crop the day a f te r i t fledged! This ind iv idua l
was not observed a f te r th is day.
Table 9. The number o f days th a t j u v e n i l e s , du r ing t h e i r PFDP, were considered not to have foraged. Column A gives the number o f
days between f ledging and the f i r s t observed foraging attempt ;
in brackets is the number o f observation days between these two
events.
Nest No. days ju v . observed
No. days j u v . d id n o t fo rage
D ro p . * COl . A
6 26 16 62 43 (16)
16 49 14 29 17 (6)
4 9 8 89 25 (8 )
17 8 6 75 1 (1)
63 43 15 35 2 M )
7 2 1 (50) 1 (1 )
At the nest s i t e , j u v e n i l e s s o l i c i t f o r food by means o f a
range o f s ig n a ls , in c lu d in g neck -s tabb ing , hoarse-ca M in g ,
crouching and wing-shaking (Mundy 1982:189)(see Figure 22). Only
once was a perched juven i le heard to ca l l hoarsely away from the s i t e (no a d u l t nearby); no neck-s tabb ing or w ing-shak ing was
observed away from a s i t e (a l though see "Parents a t the nes t " ) .
On s ix occasions w ith two ind iv idua ls , xhe juven i le ca l led in the
a i r some 15 secs before landi j . Juveni les indicated recognit ion of a parent by commencing s o l i c i t i n g behaviour before the parent
landed. I f the a d u l t f lew o f f w h i le the j u v e n i l e was begging,
t h a t behav iour would cease. The occurrence o f f o o d - s o l i c i t i n g
behaviour i s g iven in Table 10, as is the p ro p o r t io n o f days

A b s o lic it in g behaviour C DNest No. Jays No. days Juv. B/A I am only im only am ♦ pm No. day* No. days 0 / C I
observed v is ite d nest s o !le tt in g ad 'seen ’motions
I Z 85 3 10 5 IB I I 61
63
26
16 49 32 65 2 12 2 16 10 83
44 33
5017 8 6 75 I 3 - 4
4 j 79 67 4 20 2 26 15 58
i » 67 t £««12 f , 46 J * *6 7 *"53%
TVole 10. The occurrence of fo o d -s o l ic i t in g behaviour during the
PFDP o f f i v e j u v e n i l e s in 1981/2. Column D g ives the number o f days t h a t a v j I t s were observed to go through the motions o f
regurg i ta t ing food.
C/8
82
50
75
67
90
T -7 3 i

NestA
No. days oBserved
BNo. d i / S Juv. v is ite d nest
8/A *.s o lic it in g behaviour
am only pm only am » pmc
No. dayss o lic it in gseen
0No. days ad H 'motions'
0/C X C/8
6 26 22 85 3 10 5 18
16 49 32 65 2 12 2 16 10 63 50
4 9 4 44 2 \ - 3 1 33 75
17 a 6 75 1 3 - : 4 2 50 67
63 43 29 67 4 20 Z 26 15 58 90
2 • 6 /1 $*•12 $ **46 $ *•6 7 *•531 7 .7 3
Table 1C. The occurrence of fo o d -s o l ic i t in g behaviour during tne
PFDP o f f i v e j u v e n i l e s in 1 9 8 1 / 2 . Column D gives tne n u m b e r of
days t h a t a d u l t s were observed to go through the mot ions of
regurg i ta t ing food.

du r ing each 1981 PFOP th a t the j u v e n i l e v i s i t e d i t s s i t e (co l .
B/A).A du l ts w i th an empty crop were seen to go through the
motions or r e g u r g i t a t i n g , and j u v e n i l e s o l i c i t i n g behaviour
would continue a f t e r th is had ceased (although up to some 5U0g of
food is no* v i s i b l e as a d is te n s io n - see 3.3). As both b i r d s
often fc nto the c l i f f , actual t rans fe r of food was d i f f i c u l t to ascertain f, om the observation point. Even when the juven i le
had appa ren t ly rece ived a s u b s ta n t ia l amount o f food, tne s o l i c i t i n g behaviour would con t inue s p o ra d ic a l l y through the
afternoon. The number of days where an adul t was observed to 'go
through the motions" are indicated in Table 10 (col. D/C): these
are 1 ike 1 > to be minimum f ig u r e s , as such instances are e a s i l y
missed by an observer.A l l j u v e n i l e s th a t had PFDPs v i s i t e d t h e i r s i t e s w i t h i n a
maximum o f fou r ob se rva t ion days o f t h e i r re sp ec t ive f le d g in g
dates: a 1982 j u v e n i l e , equipped w i t h i r a d i o - t r a n s m i t t e r , did
not v i s i t i t s s i t e once du r ing the 17 days th a t i t remained in
the v i c i n i t y o f the k l o o f ( i .e . i t d id not forage). Each day i t made a number o f e x p lo ra to ry f l i g h t s in the k l o o f and o f te n
soared past i t s s i t e where an a d u l t (which on a t le a s t e ig h t
occasions had a bulging crop) was present, but landed most t imes
a t a po in t ca 15 m below i t s si te. On i t s second to f i f t h f l i g h t s i t landed in v e g e ta t io n , and was caught and weighed two days
a f t e r f l e d g in g (w e ig h t 6,9 kg). A f t e r 19 days of no t o b ta in in g
food, i t was found dead 4 kms from the k loo f (weight 5,2 kg).An i n t e r s p e c i f i c i n t e r a c t i o n between a j u v e n i l e and two
Secretary Birds Sagi t ta r iu s serpentarius is described in 4.2.6.
Parents at the nestDuring the PFDP of each 1981 juven i le , adults l e f t the nest
vacant f o r 14 per cen t o f th observa t ion per iod (F igure 21) ;
there was no s ig n i f i c a n t d i f ference between any of the f ive nests
(Schef fe 's t e s t , p > 0 ,0 5 ) . At three o f the f i v e nests , each s i t e was unoccupied fo r s i g n i f i c a n t l y more t ime than dur ing the
in cu ba t ion or n e s t l i n g per iods (Schef fe 's t e s t , p<0 ,01 ) , w h i le no t s i g n i f i c a n t l y d i f f e r e n t from the p r e - la y in g per iod or

behaviour shown during re tent ion of the site.On average, 15,0 per cent o f c o p u la t io ns observed through
the breeding cycle occurred during th is period (s ix nests, Table
8). At nest no. 16, an egg was l a id s ix days a f t e r the j u v e n i l e ' s l a s t recorded v i s i t ; 45.4 per cent o f cop u la t io ns observed at
th is s i te occurred during tha t PFDP (the s i te with the copulat ing
a d u l t s in F igure 22). A male paren t mounted a 225 day-o ld
ju ve n i le in a copulat ion attempt, and emitted a loud groan before the s h r ie k in g j u v e n i l e jabbed i t o f f . The parent then pecked
gently at the juven i le 's back. This was the only inc ident of th is k in d observed. S im i l a r behaviour has been observed in Gannets
Sul a bassana (Nelson 1978).Two o bse rva t ions in d ic a te d th a t adu l ts d i f f e r e n t i a t e d
between j u v e n i l e s and recognised t h e i r own o f f s p r i n g . In one
case, nest no. 16 j u v e n i l e (no. 16 j u v . ) , 12 days a f t e r f l e d g in g ,
landed near nest no. 57 and walked onto the vacant nest. A
juve n i le had fledged from th is s i te seven days before, and was in
f u l l view o f the s i t e a t the t ime. At 09h30 an a d u l t landed and nc. 16 juv. i n i t i a t e d normal fo o d -s o l ic i t in g behaviour. A f te r the
a d u l t gave i t two aggress ive jabs on i t s wing, the s o l i c i t i n g stopped and the a d u l t stood in an aggress ive posture over i t ,
neck extended f o r fou r minutes, be fore moving o f f the nest .
S ix teen minutes l a t e r , no. 57 juv . landed on the s i t e ledge,
looked at the in t rude r and stood facing out. Twelve minutes a f te r
t h i s , the adul t jabbed aggressively at the intruder and repulsed
i t from the s i t e . In the second case, no. 63 juv . (8 days a f t e r
f ledging) landed between two nest s i tes , one o f which (nest no.6)
held a dependant juven i le 30 days older than the in truder; no. 6
j u v . was not a t the s i t e a t the t ime. Both adu l ts ag g re ss ive ly jabbed at no. 63 ju v . 's heal and back and stood over i t , w h i l s t the ju ve n i ie crouched and shrieked submissively, with th e i r necks
extended in c lass ic th reat posture (Mundy 1982:188). Four minutes a f t e r landing, the juven i le was aggressively pushed o f f the ledge
by nest no. 6 a d u l t . On three ihe r occasions, a j u v e n i l e du r ing
i t s PFDP was aggressively repulsed a f t e r landing at a s i te other
than i t s own, a l though these ins tances d i f f e r e d from the two c i te d above in tha t the s i tes concerned had produced no s im i la r ly
aged o f fsp r ing that cycle.

Terminat ion o f the period
Aggressive i nteract ions were noted in f i ve of e ight observed
PFDPs dur i ng t h i s study: observa t i ons ceased before the end o fone long 1961 PFDP, thus t h i s r a t i o i s more c o r r e c t l y f i v e ofseven cases. One 1981 PFDP is excluded from this as the j uven i le
was a t the s i t e f o r one day only. In t h a t case i t appeared to
have fore ted s u c c e s s fu l l y the day a f t e r f l ed g in g and i t i s not
known whether i t d i spersed or d ied a f t e r t ha t day, but i t was
never r es i gh ted in the k l o o f again. The aggress ive encounters
ranged from a parent jabbing sharply at. the s o l i c i t i n g o f f spr i ng
wh i l e a l l o w i n g i t to remain a t the s i t e (7 ins tances, 2 nes ts ) ,
to the parent chasing the juveni le o f f the s i te (11 instances, 4nests), or prevent ing i t from landing at the s i te (4 instances, 1
nest). In three of the seven instances, the juveni le jabbed back
at the adul t but ceased to s o l i c i t . During the 11 instances that
the j uven i l e was chased o f f the s i t e a f t e r landing, i t was not atthe s i t e f o r longer than three minutes. Parents prevented the
o f f s p r i n g from landing by extending t h e i r necks (3 i nstances
wi th i n 15 minutes at one s i te , both adul ts present), or by moving
quick ly to i t s projected landing spot and ca l l i ng loudly, causing i t to veer o f f .
At. the f i r s t recorded aggressive encounter in each case, the average age o f the j u v e n i l e was 290 days (range 175-361 days).
This t ranslates into some 147 days, or f i ve months, of PFDP. The f i r s t recorded case of aggtessive jabbing at nest no. 6 occurreu 36 days before the period ended, and the j uven i le was also chased
o f f the s i te f i ve t imes that day and prevented from landing once
(F igure 27). Six days l a t e r i t was fed by a parent a t the s i t e ,
a f t e r two bouts o f aggress ive j abb ing ( l a s t i n g a few seconds each). ; n three o f the o ther cases, the j u v e n i l e d id not spend
t ime a t the s i t e a f t e r aggression was recorded (one i n d i v i d u a l v i s i t ed the s i t e lu and 11 days a f t e r i t had been chased o f f , but i t ne i ther so l i c i t e d nor was o f f ed any food).
Retent ion of the s i te
Of the nine si tes that fa i led in breeding in 1981, six were ac t i v e in 1982. At two o f these s i t e s , one pa r t ne r had a metal
r i ng and both bi rds were act ive in both cycles. Af ter the pa i r at

nes t no. 63 f a i l e d dur i ng the i ncuba t i on per iod in 19132, both
(colour r inged) partners were seen to spend t ime at, and repulse
i n t r u d e r s f rom, t h a t s i t e . These ins tances served to con f i rm behaviour tha t was i nd ica t i ve of re ta in ing a speci f ic nest s i t e
f o r the purposes o f breeding in the fo l lowing season.
Si tes t h a t f a i l e d i n 1981 and were ac t i v e in 1982 were
unoccupied f o r 43 per cent o f the counts ( range 12«-5Q%, 537
si te-days o f observa t i on , mean o f 91 s i te-days per s i t e ) , and
only one s i t e was attended s i g n i f i c a n t l y more than any other si te (Schef fe ' s t e s t , p <0 , 01 ) . On average f o r s i x s i t e s , a s i n g le
a d u l t a t tended tne si te f o r 24 per cent o f tne counts, one si te being s i g n i f i c a n t l y ess attended than the other f ive. Both s i tes
were attended by adul ts f o r 33 per cent of the counts on average, and there were no s i g n i f i c a n t di f ferences between s i tes in t h i s
respect.For 11 per cen t oT the counts at s i t e s t ha t produced
juveni les in 1981, adul ts were not at the s i t e a f t e r the PFDP had
ended.

75
4.2.5.Breedi ' ig success
As a l l breeding s i t e s and roo s t i ng areas at Potberg were
c l e a r l y v i s i b l e , the d i f f i c u l t i e s in measuring the var ious
parameters associated wi th the product i on o f young (see Newton
1979:128-130) were amel iorated to a considerable extent. Thus i t
i s c e r t a i n , a t l e a s t a t Potberg, t ha t dur ing the study per iod
the re l ev an t i n f o r m a t i o n on a c t i v e s i t e s , eggs l a i d , n e s t l i n g s hatched and the numbers of non-breeding b i r d s was ob ta i ned; i t was assumed t h a t b r e e d i n g o c c u r r e d o n l y a t P o t o e r g and
Aasvoge l v l e i ( chapte r f i v e ) . Success i s then expressed as the mean number o f young produced (nest l ings f ledged) per t e r r i t o r i a l
p a i r . This cor responds to Howard's (1979) quaternary level
est imate of reproduct ive success. The survival j uveni les unt i l
t e r m i n a t i o n o f pa ren ta l care (Howard's Ojx c i t. f i f t h l eve l
es t im a te o f r e p r o du c t i v e success), and the p ro po r t i on o f b i rds breeding, i s cons idered in chapter f i v e . In both years a t nest
no. 8 where a complete nest was b u i l t and the (non- r i nged,
presumed same) adul ts copulated f requent ly (Table 7), no egg was
l a i d but the s i t e was i nc l uded as ac t i ve . The p a r t i a l l y ac t i ve
s i te discussed in 4.2.1. was not taken as act ive in 1981.
Table 11. Breeding data f o r 1981 and 1982 at Potoerg (PB) and Aasvogelvlei (AW).
Colony PB
C l i f f NWI nw? KF £ « |
(A)3rf>pfilno pairs in *> 2 17 '
Eiqs l aid 9 6 2 U j
Egqs hatched 5 5 - to J
N o s t l . r inged 4 1 - 7 1
( B ) N p ' t l . f ledged
(B/A)Success
3 3 fi
0, 35'
NWI
PB
NW? Nf 1901 1 1
I I 6 1 1H in 9
10 fi 1 17 10 8
10 4 14 1 7
7 3 10 6 5 II
5 2 7 4 5 9
0,39 0 .40 0.55

76
Breeding data for 1981 and 1982 f or PotDerg and Aasvogelvlei are presented in Table 11, and separated according to the
locat ion o f each s i te for the Potberg colony (see Figures 18, 19
and 20). The Potberg success f i gu re s f o r the two years were not
s i g n i f i c a n t l y d i f f e r e n t ( \ 2=0,043, I d . f . , p>0,25) . Cons ider ing
both co lon ies f o r the two years o f the study per i od (54 p a i r -
years) , 22 n e s t l i n g s f ledged, an average of 0,41 n e s t l i n g s per p a i r (or 0,39 as I ' improved' t h i s f i g u r e by rep lac i ng an egg in
1981 (?), descr ibed in 4.2.2.) ; f o r the purposes o f s t a t i s t i c a l
c a l c u l a t i o n s , t h i s s i t e i s taken as successfu l . Due to the low expected f requenc ies i nvo l ved , the f i gu re s f o r Potberg and
Aasvogelvlei were not compared s t a t i s t i c a l l y ; in a l l chi-square tests , Yates' cor rect ion fac tor is used (Schef ler 1969:95).
The Potberg 1981 success f igure is s i g n i f i c a n t l y lower than
the Kranzberg f igure for the same year ( \ 2 =4,67, Id. f . , p <0,05)
as we l l as the Col 1 eywobbl es f i g u r e (X 2 =5,81, I d . f . , p<0,025) ;
f i g u r e s were taken from J.C. Dobbs and P.C. Benson (pers comm.) and Vernon et a_l_. (1982) r e s p e c t i v e l y . I t was not s i g n i f i c a n t l y
lower thai , the p rev ious two years combined (X2 = l , 32 , I d . f . , p=0,25). The 1982 Potberg f i g u r e was s i g n i f i c a n t l y lower than
t h a t o f 1981 at Col ley wobbles (X2 =4,85, I d . f . , p<0 ,05 ) , but not s i g n i f i c a n t l y lower than the i982 Kranzberg f igure ( \ 2=2,66,
Id. f . , p >0,05). Between 1975 and 1982, nest l ings were documented
at 98 of 165 act ive s i tes (1975-1980 data from CDNEC records), a f i g u r e o f 0,59 n e s t l i n g s per s i t e . As 13 eggs were documented,
and t he i r outcome unknown, th is f igure is more accurately given
as 0,51-Q,C7 n e s t l i n g s ra ised per a c t i v e s i t e , or pa i r . This
f igure is not s i g n i f i c a n t l y higher than a mean success f igure for the Maga 1 iesberg co lon ies (Mundy 1982:193) ( \ 2= 0,19, Id. f . ,p > 0 , 2 ).
Of s i tes that were act ive in both study years a t Potberg, 2 out o f 13 (0,15) produced f l e d g l i n g s in both years , 7 out o 13 (0,54) produced one f l e d g l i n g , out o f 13 (0,38) produced large
n e s t l i n g s , and 3 out o f 13 (0,23) never produced one. Obv ious ly ,
s i t e s t h a t produced f l e d g l i n g s a lso produced large n e s t l i n g s :
the ca tego r i es are not separated here. The d i f f e r e n c e in the success o f a t t e m p t s between NW1 and NW2 c l i f f s was not
s i gn i f i can t (X2 *3,34, Id. f . , p >0,05). The frequency of breeding

w
77
a t p a r t i c u l a r s i t e s i s i nc l uded in chapter s i x (see a l so Table
Causes of breeding failure
Possible causes of breeding f a i l u re are l i s ted in Table 12,
1 inked to observed causes o f f a i l u r e f j r the two years and both co l o n i es , and are discussed in t h i s sect ion. Many o f the
ca tego r i es are d i r e c t l y l i n k ed , and an a t t emp t was made to
d i f f e re n t i a te between proximate and u l t imate causes (Newton 19 9:
129), and include both.
Table 12. Possible causes of breeding f a i l u re an.' t h e i r est imated occurrence a t Potberg (PB) and Aasvoge l v l e i (AW) dur i ng the
study period.
Category Incidence in 1981 Inci ence in 1982
? 'j AVV PB AVV
1. Dead egg 3 2 2 i
2 . Depredation 2 :■ ■*. ‘
j . 'lest content
Knocked o f f nest
1 1 ?
4. rieather 3 *
5. Parent death * * -
6. Human disturbance - 3 -
7, unknown 1 1 2
Total : 9 4 9
Category one. This grouping Includes eggs that are i n f e r t i l e and those that died as a re su l t of pest ic ide conta nation or changes
in the m i c r o - c l i m a t e o f the egg, the l a t t e r a r e s u l t of I ncuba t i ng b i r d s vaca t i ng the neat (see ca tego r i es f ou r and

78
eight). The e f fec ts of pest ic ide contamination are discussed in chapter three (3.3.). A p ro po r t i on of a l l eggs l a i d must be
i n f e r t i le (Brown & Amador 1968). No nest s i t e a t e i t h e r colony
produced an addled egg in both years.The thickness of eggshel l fragments, as wel l as sect ions of
e g g s that were co l lec ted fo r contaminant analysis, were measured
d i r e c t l y and are presented in appendix d.Category two. Ravens and Baboons Papio u r s inus are known to prey
on v u l t u r e eggs (P.C. Benson pers. comm., Green 1977, Houston
1976, pers. obs.) and Black Eagles Aqui la verreauxi i on nest l ings
(Bvwen 1970, Mundy et aj_. in prep., Steyn 1973). No successful
predatory at tempt was observed during th i s study : the breeding f a i l u res comprising t h i s category are those that were most l i k e l y
a r e s u l t o f d e p r e d a t i o n . Examples i n c l u d e the sudden disappearance of a young n e s t l i n g which could not be located
a f t e r searching the base o f the c l i f f , or the d i scovery o f egg
and content remains in the nest whose measured shel l thickness
was concluded un l i ke l y to have been ext ra- th in . The at tempts that
I observei are documented in appendix b.Category three. Incidents included here may occur as a r e su l t of
disturbance (category six) causing the adul t rapid ly to vacate
the nest , or dur i ng changeovers (e.g. i n c i d e n t descr ibed i n4.2.2.), or as a r e s u l t o f i n t r u d e r r epu l s i on behaviour . E i t he r
o f these was l i k e l y the f a te o f nest no. 25 in 1982, as remains of the shel l and contents were found wi th i n 0.5 m of the nest.
At one nest at Aasvogelvlei in 19°l , a downy whi te nest l i ng
was round dead a t the base of a r e l a t i v e l y high nest s t r u c t u r e (some 0,7 m o f f the s l op ing ledge) , p a r t i a l l y in a h o r i z o n ta l
crack. I t i s probable t h a t i t had f a l l e n out o f the nest ( adu l t s disturbed ? -see category six) and consequently starved to death.4 Mart ial Eagle Polemaetus be 11icosus nest l ing that f e l l out of
i t s nest in the same year at Potberg was attended and fed on the
ground (pers. obs.). I t is p s i b l e t h a t the v u l t u r e n e s t l i n g
died as a r e s u l t o f the ac tua l f a l l ; in e i t h e r case, the
u l t imate cause is unknown.Ca te go r y four . D i r e c t e f f e c t s of i nc l ement weather i nc lude
phys ica l d e s t r u c t i o n o f the nest and consequent m o r t a l i t y , as wr 11 as s tarvat ion o f nest l ings at c r i t i c a l stages of growth, or

/
79
a t ha tch ing, through i n h i b i t i o n of fo rag ing oy the adu l t s , ' r 1981, Potberg experienced the highest incidence of ra in fa l m / b
years (2.1.2.): l ocated w i t h i n the w i n t e r r a i n f a l I reg ion , t h i
p rec ip i ta t i on i s thus highest during the egg and young ne s t ! ”
periods.Category f i v e No evidence of parent death was notrd cer ta in y
not one o f the e ight ringed bi rds died during a breeding cycle
e i t he r year.Category s i x Three large nest l ings vacated t he i r nests between 18
and 21 November 1982 ard a h e l i c o p t e r was observed to pass
through the k l o o f on 24 November. Al though not d e f i n i t e , i t is
l i k e l y tha t hel icopters had entered the k loof during the previous
week. Subsequent post-mortems of two of the nest l ings revealed
substant ial f a t reserves (chapter three), thus three out of 18 of the 1982 breeding at tempts were concluded as having been lost due
to d i r e c t human d i s tu rbance . With the except ion o f di s t . rbance
dur i ng r i n g i n g ope ra t i ons (4.1.) and f i v e searches of the c l i f f
bases (3.2.1.), th i s was the only d is tu rbance i n c i d e n t observed, i n marked c o n t r a s t to some other co lon ies (J.C. Doobs & P.C.
Benson pers. comm., Komen 1983). I emphasise t h a t one nersor
wa l k i ng q u i e t l y along the bases o f NW1 and NW2 breeding c l i f f s dur i ng the cyc le r e s u l t s in every a t t endan t adu l t vaca t i ng 1 ts
s i te . Although Aasvogelvlei is located on pr ivate property, the
disturbance there i s considered minimal.
4.2.6.General behaviour
Intraspecific behaviour
Repulsion of intruders from the nest s i te
The behaviour o f the co lour r inged p a i r in p a r t i c u l a . and
a t o ther nests where one par r was r inged, served to con f i rm
t h a t I n t r u d e r s l a n d i n g near an occup ied nes t s i t e are aggressively chased o f f . Consistent acts of repulsion may then be
used to indicate an occupied si te. Between July 1981 and 31 May
1982 (139 obse rva t i on days), 437 such cases were noted at 2/
d i f f e r e n t s i t e s . A f u r t h e r 74 ins tances were noted < H p t e

dur i ng the nine day observa t i on s t i n t from 26 Ju l y to 3 August
1982.The agonist i c behaviour was c l ea r l y graded : as a f i r s t sign
o f aggress ive i n t e n t , the s i t e occupant would stand, and peer
i n t o the nest . A few aggress ive jabs in the d i r e c t i o n o f the i n t r u d e r would lead to the occupant jumping o f f t-he s t r u c t u r e , g i v i n g a hoarse c a l l and engaging con t a c t by j abb ing , u n t i l the
i n t r u d e r l e ' t . In f i v e i ns tances where j u v e n i l e s landed at f o r e i gn s i t e s and remained f o r longer than a few seconds, the
occupants jabbed a t and stood over them w i t h necks extended in the threat posture : two such instances are described in 4.2.4.
S i g n i f i c a n t l y more i ns tances occurred a f t e r 12h00 than
before t h a t t ime ( \ 2 " = " expected values t e s t , p<0,0Ql ) . In a l l
but 67 cases (15%), the i n t r u d e r l e f t immediate l y : o f these 67 Ins tances, the i n t r u d e r was age-es t imated as an immature 54
times. Thus an adul t in t ruder landing at a s i te is more l i k e l y to be repulsed immediately than i f i t were in immature plumage. In
at least 11 instances, immatures were "al lowed" to remain wi th in
3 m of the s i t e . Not a l l I n t r u d e rs were age-est imated however.
T h i r t y per cent o f the 131 i ns tances occurred a t s i t e s w i t h no egg, n e s t l i n g or dependent j u v e n i l e ( i .e . pa i r s t ha t had f a i l e d
in breed ing, or where no egg was produced). A f t e r f a i l u r e in breed ing, r e pu l s i ons were documented a+ a l l s i t e s except one,
which was abandoned a f t e r f a i l u r e , and was not ac t i v e in the fo l lowing season. Members o f adjacent s i tes recognised each other
and no aggression was recorded save for when a member wandered
too near to i t s adjacent si te.Int ruders landing on a communal nest ing ledge were general ly
chased o f f oy the occupants of more than one s i te , r esu l t i ng in a summated repulsion ef fec t . This e f f ec t was pa r t i cu l a r l y evident
when i n t e r s p e c i f i c p r e d a t o r y a t t e m p t s were observed ( " I n t e r s p e c i f i c behaviour " , and appendix b). I n t r ud e r s were not
to lerated at any point on the 4Z breeding ledge (Figure 22) by a
member of any nest on that ledge.An a d u l t from the s i t e t h a t was ac t i v e in both years but
never produced an egg, was repulsed from various si tes w i th eggs on 19 observed occasions in 1981 and 15 in 1982 (observed during
the nine day s t i n t ) . On two o f the occasions in 1981, both

81
members v i s i t ed foreign s i tes together. An extreme case occurred
21 days a f t e r the 1981 average l ay i ng date, when one nest no.8
occupant was aggressively repulsed ten t imes from four d i f f e ren t
s i t e s w i t h i n 200 m o f i t s own. Each s i t e i t v i s i t e d had an egg, and i r each case the i nc uba t i ng ad u l t j u m p e d o f f the nest
s t r u c t u r e to repu l se i t . Ten days before the (es t imated) 1932
average laying date, an incubat ing adul t was o f f i t s egg 11 times
in 14 minutes to repul se the p a i r t h a t kept on l anding a t the s i t e : th i s pai r f i n a l l y landed at nest no.8. This behaviour was
not observed by the occupants of any other s i te.The only t ime that a colour ringed immature was observed to
v i s i t i t s natal s i t e (one breeding a t t emp t had passed s ince i t f l e d g e d ) , i t c a r r i e d n e s t m a t e r i a l t o the s i t e b u t was
aggressively chased o f f twice by the at tendant adul t.
Interspecific behaviour
P r e d a t o r y a t t e m p t s on n e s t l i n g s by Black Eagles are
described in deta i l in appendix b.
Both eagles o f t en soared past the v u l t u re breeding 1 edges ( w i t h i n some 20 m), and t h i s e l i c i t e d c l e a r l y aggress ive
responses on occasions observed between November 1981 and
February 1982, i .e. when there were l arge v u l t u r e n e s t l i n g s
present. On seven other occasions when the eagles soared past the
breeding ledges, I recorded no discernib le aggressive response :
t h e s e occasions occur red when there were e i t h e r eggs ( f ou r
instances) or empty nests.The eagles' nest was si tuated between NE and Nw2 c l i f f s and
vu l t u re s consequent ly passed i t f r e qu e n t l y . On eleven o f these
ocass ions, on* o f the eagles dived at the v u l t u re causing i t to
f lex i t s wings. These aggressive i nteract ions, i n i t i a t e d by the
eagles, were on ly observed dur i ng the eagles' breeding cyc le . A f t e r the eyas disappeared in , l y 1981, I observed no s i m i l a r
i n c i d e n t s u n t i l May 1982, the onset o f t ha t season's breeding
attempt.On 23 February 1982, I observed an aggressive I n t e r a c t i o n
between a s i x month o ld j u v e n i l e and two Secretary Bi rds in an
open f i e l d 5 km from t h t k l o o f . I di sturbed the j u v e n i l e , which

landed 40 m f rom the two b i r d s : both ran towards i t andattempted to stamp on i t in charac te r i s t i c fashion. The vul ture
responded wi th loud staccato c a l l s and a f t e r some 40 secs the two
wandered of f .I n t e r a c t i o n s betweer v u l t u re s and White-necked Ravens are
documented in 4.2.3.Although Peregrine "aIcons Falco peregrinus nested w i th i n 5
m o f one v u l t u r e nes t , and o f t en perched c lose to v u l t u r e s , no
aggressive in teract ions were observed.Chacma Baboons were observed per i od ica l l y in the k loof and
when near (e.g. below) the breeding ledges, vul tures would watch
them closely. No predatory at tempts were observed.

4 . 3 . Discussion
4.3.1.The p r e - l a y i n g per i od
On the evidence a v a i l a b l e , s i t e s are ac t i v a te d some s ix
months or more before egg-laying. I can only conclude that pai r
format ion preceded occupancy of a s i t f : single vul tures were not seen c o n s i s t e n t l y a t the few s i t e s i n i t i a t e d dur i ng the study per iod . A l l b i p a re n ta l species possess mechanisms f or wringing
mates together and achieving c o p u la t i o n , e s s e n t i a l l y a s i g n a l - response r e a c t i o n c h a i n (P a te r s o n 1982). I d i scerned few
c o u r t sh ip procedures except , perhaps, mutual preening and scapular act ion (Ber rut i 1981). Obviously, many signals perceived
by e i ther sex may not be perceived eas i ly by human observers. The cha rac te rs o f the s p e c i f i c - m a t e re c o g n i t i o n system func t i on
e f f e c t i v e l y i n the normal or recognised h a b i t a t o f the species (Paterson op. c i t . ), and f o r r a p to r s i n genera l , p a i r f o rmat ion
tor instance i s associated wi th potent ia l breeding places (Newton
1979). In the Cape V u l t u r e t h i s i s the breeding co lony , and
s p e c i f i c a l l y the ne s t i ng 1 edges, where 95,7 per cent o f a l l c op u l a t i o n a t t emp ts occurred. In c o n t r a s t to the feeding area
which, ce r t a i n l y In t h i s colony's case, i s t o t a l l y man-inf luenced
(chapter three), the breeding area is unaffected by humans. Thus
aspects of breeding behaviour t h a t I observed t ha t are not d i r e c t l y l inked to feeding, e.g. nest l ing attendance and foraging
t imes , should be r e p re s e n ta t i v e o f the species throughout i t s
range.As nest mater ial disappeared from some si tes before the next
season's cyc l e , the s i t e r a the r than the nest s t r u c t u r e (Newton
1979) wou ld appear t he more v a l u a b l e resource. Th is was emphasised by in t ruder repulsion behaviour throughout the year.
Houston's (1976) observat ion of a sex-related di f ference in
behaviour du r i ng t h i s per i od con f i rmed, and as a number o f
ind iv iduals were recognisable, males w 're documented to co l l ec t
nest material more f requent ly than females. I emphasise that the
behaviour i s not d i s t i n c t i v e f o r Cape Vu l t u re males ; very s im i l a r behaviour occurs in Gannett (Nelson 19/8). Such behaviour
would, in e f f ec t , reduce the female's energy expenditure ; is i t

84
then related to piuJuct ion of the egg ? A range of raptor females increase i n we igh t p r i o r to egg produc t i on (Newton 1979:Fi gure
15). However, in g r i f f on vul tures that share parental attendance du t i e s , the increase occurs in both sexes (Houston 1976). Thus
energy is accumulated not merely to enable the female to produce
eggs, but to supplement the energy budget o f both dur i ng the subsequent cycle periods (Newton 19/9). Also, the g r i f f on vul ture
d u t r h o f one egg weighs between three and f i v e per cent o f the female's body we igh t (Mundy 1982) and, in c o n t r a s t to sma l l e r
raptors, places a r e l a t i v e l y small demand on her reserves. These
con s i de r a t i on s i n d i c a t e t h a t the bias in c o l l e c t i n g forays is r e l a te d to some o ther f a c t o r , such as cou r t sh ip (ak in to Gannet
behaviour ?).Far more c o p u l a t i o n a t t emp ts occur than may be considered
adequate for inseminat ion, and t h i s has been noticed in a number
o f v ec i es (reviewed in Newton 1979 and Steyn 1982). I t ce r t a i n l y
occurs a t Potberg in the m a j o r i t y o f pa i r s , a l b e i t at * reduced
frequency, throughout the year (also Mundy 1982) ; th i s contrasts
wi th Houston's (1976) observat ion of none a f t e r a month past the
mean l ay i ng date. As I spent more t ime in observat i on , and was
c o n s i s te n t through the l ength o f one cyc le , I would gain a more
accurate re f l ec t i on of the bi rds' bt. iaviour.The act i t s e l f i s considered to serve bio logical funct ions
o the r than immediate f e r t i l i s a t i o n (Power 1980), i nc l ud ing
i nduc t i on o f ovar i an a c t i v i t y (Er ickson A Zenone 1976) and
inducement of endocrine changes f a c i l i t a t i n g successive stages of
nes t i ng (Hinde 1970). I t has been imp l i c a te d in pa i r - bond ing (Newton 1979), but anomal ies such as high f requenc ies of
a t t emp ts , in species w i t h weak p a i r bonds, are documented (e.g. Black-shouldered Ki te FI anus caeruleus, reviewed in Steyn 1982).
Noting the strong t a c t i l e s t imu l i in Gannet copulat ions, Nelson (1978) suggested them to be a prolonged source o f sexual s t i m u l a t i o n ; s i m i l a r s t i m u l i both t a c t i l e and a u d i t o r y , are
apparent in v u l t u r e mat ings. The e f f e c t , i f any, o f the male's
loud ca l l s on adjacent pai rs would be Incidental .Er ickson & Zenone (1976) cons ider t h a t many copu la t i ons
spread over a long t ime r e f l e c t an a t t empt by the male to
i ncrease his con f i dence o f p a t e r n i t y . This i s based on Tr i vers'

(1974) suggest ion t h a t in species where the male co n t r i b u te s ex t e ns i v e l y to pa ren ta l care, i t i s im po r tan t , in ev o lu t i o n a ry
terms, to the male tha t the eggs are f e r t i l i s e d by his own sperm. L im i t ing copulat ions to the female's t ime of ovulat ion opens the p o s s i b i l i t y o f her being inseminated p r io r to his e f f o r t s . Across
a range o f species then, the g rea t e r the male's parenta l i nves tment , the s t r ong e r the behaviou** t h a t would ensure his
genet ic p a t e r n i t y . I f t h i s holds, the occurrence o f c o l o n i a l breeding would again emphasise the importance of t h i s beh viour,
as c o lo n ia l i t y e f f e c t i v e l y raises the female's chances of contact w i t h w i t h o ther males (e.g. a t the roos t po in t s ) . Copulat ions c e r t a i n l y occur a t areas o ther than nest s i t e s ( t h i s study,
Houston 1976).
4.3.2.The Incubat ion pe r i od
Given the o f t en markedly c i rcannual rhythm o f l ay i ng (e.g.
t h i s study, tunr 'y 1982), v u l t u r e s c l e a r l y respond to c e r t a i n
prox imate f a c t o - s t h a t prov ide a genet ic-based s t i mu lus to i n i t i a t e breeding behaviour (Lack 1954). Food a v a i l a b i l i t y at the
t ime may then resu l t in minor modi f i cat ions in the laying date.
For raptors in general , mean laying dates are ea r l i e r in good as
opposed to poor food c o n d i t i o n s ; food t he re fo re acts as both a prox imate and u l t i m a t e c o n t r o l l i n g f a c t o r (Newton 1979). For
example, Yom-Tov (1974) advanced the l ay i ng date o f the Crow Co.-vus cojrone by o f f e r i ng supplementary food in the p r e - l a y i n g
per i od and compar ing the r e s u l t s w i t h c o n t r o l s , a 1 though the
e f f e c t was sub t l e (Brent & Daan 1980). Also, under good food condi t ions one would expect an increase in the number of breeding
a t t emp ts , o ther t h i ngs being equal (chapter f i v e , 5.3.2.). However, long per i ods of p r e c i p i t a t i o n or heavy c loud, through
i n h i b i t i n g v u l t u r e s ' f o rag ing , e f f e c t i v e l y reduces the ra te of accumula t ion o f the necessar (4.3.4.) body reserves in the
per iod preceding l ay i ng . This would act to delay the date o f l a y i n g , and i f t h i s co n s i de ra t i o n i n f luences the date to a
considerable extent (i .e. i f i t alone were largely responsible),
the late 1981 laying date then indicates a period of reduced food
a v a i l a b i l i t y . This exp lana t i on seems the most f e a s i b l e f o r the

86
ra in-cor re la ted var ia t i ons in mean laying date that I documented.
The markedly c i rcannua1 l ay i ng date o f p a r t i c u l a r pa i r s ,
c e r t a i n l y both i d e n t i f i a b l e par tne rs a t nest no. 63 where the date was e x a c t l y the same in both study years would, however,
quest ion the degree o f inf luence of these proximate controls.I t must constant ly be borne in mind that the food source of
t h i s colony i s an " a r t e f a c t " o f s t oc k - f a r m ing p r ac t i ces . As the
food source has changed markedly w i t h i n l i v i n g memory, i t is
u n r e a l i s t i c to r e l a t e o v e ra l l t i m i n g o f the colony 's breeding
cycle to i t s cur rent food source. I r t h i s regard, Houston (197o) concluded that the breeding season of the congener Gu rueppe im
in East Af r i ca was so t imed that the f ledging date coincided wi J i a r e l a t i v e l y abundant per i od o f food. This l i n k c e r t a i n l y doe^
not hold at Potberg, where f l e d g l i n g s appear dur ing a t ime of
r e l a t i ve decrease in food abundance.I t i s of i n t e res t to note that throughout the Cape Vul ture's
range there is a gradat ion in laying dates, from ea r l i es t in the
Transvaal (ear ly May) through to l a tes t in the southwestern Cape
(mid June). Th is appears to be a r e f l e c t i o n o f env i ronment -
i nduced m o d i f i c a t i o n o f what i s l a r g e l y the i n n a t e l y predetermined t ime o f i n i t i a t i o n o f breeding, but the f a c to r s
involved are unknown.The accurately determined period length of 57 days compares
favourably wi th the 56-day est imate of Mundy (1982:165) and the
57,6 day per i od o f Boshof f & Cur r ie (1981). I t i s c l e a r l y longer than the f i g u re s given in Brown A Amadon (1968) and McLa^hlan &
L 1 vers 1 dge (1978).
4.3.3.The nestl ing period
Immedia te ly a f t e r ha tch ing , the tendency was f o r both
parents to stay at the nest t oge ther f o r longer s t r e tches V.an
during the Incubation period. Nest l ings were essent ia l l y guarded
a l l the t ime by at least ve parent ( th i s stud), Mundy 1982), and t h i s contrasts wi th attendance behaviour o f other vul ture species
(Mundy 1982, Pennycuick 1976). Predat ion may account f o r a s ub s ta n t i a l po r t i o n o f egg and n e s t l i n g loss in co l on ia l
s i tuat ions, e.g. gu l l s , reviewed in Hunt & Hunt (1976). The high

parenta l at tendance by Gu_ cop ro theres , f o r the stage o f the per i od a f t e r the n e s t l i n g i s d o r s a l l y fea thered, could then
r e f l e c t an ant i -predator response ; more l i k e l y i t is an e f f ec t
o f c o l o n i a l i t y and the t r a f f i c o f s t rangers to the nest (Mundy
1982). For example, the a b i l i t y of parents to repulse Black Eagle predatory a t t emp ts i s enhanced as a r e s u l t o f t h e i r co l o n i a l
nes t i ng ha b i t (4.2.6., appendix b).That at tendance s t i n t s o f both sexes were sh o r te r than
during the incubat ion period re f l ec t s the increased attendance of
both parents , as we l l as not such c lose at tendance by any one
parent . Th's made changeovers more d i f f i c u l t to d i scern and
re s u l t e d in a g rea te r p r op o r t i o n o f w i t h i n - d a y s t i n t s being documented. The e a r l i e r changeover t ime r e f l e c t s the weather
c o n d i t i on s o f approaching summer, as compared to the previous period (see chapter three), as wel l as the st imulat ion provided
by the nest l ing to return to the nest ea r l i e r .Nes t l i ng growth and c o n d i t i o n , as we l l as aspects o f the
q u a l i t y o f food as i t a f f e c t s n e s t l i n g s , are considered in
chapter three (3.2.4. and 3.3.).Nes t l i ngs o f the Lappet - faced Vu l t u re are known to be
parasi t ised by Hippoboscid f l i e s Icosta meda (Anthony 1976), and in 1981 Potberg nest l ings were heavi ly fed on by s imu l i i d f l i e s .
As parents remove f l i e s , the number of f l i e s that I observed was probably greater than i t would have been had the parents not been
disturbed (by me). S imu l i i d f l i e s require water in which to breed (Ledger 1979), and the stream in the k l o o f prov ides a su i t a b l e
ha b i t a t . The Potberg c r l o n y i s the only one I know of w i t h a
sui table breeding ground fo r these f l i e s w i t h i n some 400 m of the
breeding ledges (see also Mundy 1982, Tarbotor * Al lan in press). That the degree of nest l i ng i n fes ta t i on is related to preceding p rec ip i ta t i on , i s indicated in Figure 25, although other factors
may inf luence t he i r presence.S i m u l i i d s are known to vectors o f protozoan pa ras i t es
(Fal l is et al. 1973), and are l i k e l y vectors of avian haematozoa i n t o v u l t u r e n e s t l i n g s (Boshof f & Cur r ie 1981, t ioshof f 1981).
Blooa smears o f 1979 n e s t l i n g s (a year o f heavy i n f e s t a t i o n )
contained one or more species of Leucocytozoon, wi th the degree of i nfec t ion varying from low-grade in some samples to a higher

parasl taemiu in others (Boshoff & Curr ie The f ac t that thee a r l y blood f i l m taken from a 22-day o l d n e s t l i n g revealed no
Leucocytozoon presence probably r e f l ec ts an aspect of the f l i e s
l i f e cycle. For example, gametocytes of L. neavei were f i r s t seen in peripheral blood of Guinea Fowl Numida meleagris nest l ings 14
days af ter i n fec t i on from sporozoi tes from S. adersi (Fal l is et a 1. 1973). The subsequent f l e d g in g success of those Potberg
n e s t l i n g s w i t h high pa ras i t aemia i s unknown, thus whether
Leucocytozoon cont r ibutes to Cape Vul ture mor ta l i t y or morbidi ty remains undetermined. The n e s t l i n g t h a t ex h ib i t e d a low-grade
Leucocytozoon i n fec t i on (and which was fed on by a large number
o f s i m u l i i d s ) in t h i s study was l a s t seen f l y i n g s i x days a f t e r
i t f ledged, and i t did not have a PFDP. Given the necessi ty of at least some supplementary aid from the parents during th is stage
o f i t s l i f e (4.3.4.), even i f i t d i spersed, the chances are t ha t
i t d id not l i v e long. However, no l i n k between i t s probable demise and the parasi taemia i s suggested ; the nest l ing cer ta i n l y
l i v e d f o r some th ree months w i t h th' is i n f e c t i o n . Thus more
observat ions of the fate of f l edgl ings known to be infected wi th b lood pa ras i t e s are needed before any conc lus ion regard ing the
e f f ec ts of e i ther parasi te can be reached.In a prev ious study o f haematozoa from southern A f r i c a n
v u l t u r e s , the Cape Vu l tu re was the cn i y species t h a t was not
i n f e c te d w i t h blood pa ra s i t e s (Greiner & Mundy 1979). This was ascr ibed to i t s unique nes t i ng h a b i t , as we l l as the t i m in g of
i t s breeding season. In t h e i r f i r s t weeks of l i f e , nest l ings have a re l a t i v e l y sof t skin and are e i ther naked or sparsely covered
w i t h f ea the rs ; consequent ly i t i s a t t h i s stage t h a t they are p a r t i c u l a r l y vu lne rab le to the a t t e n t i o n s of b i t i n g i nsec ts
(Markus 1974). My observat ions of the d i f f e ren t extent of feeding
by f l i e s a t two sequen t i a l v i s i t s to the nest would tend to suppor t t h i s . Gre iner & Mundy (1979) sampled ne s t l i n g s f -om
s u n r e r r a i n f a l l co l o n i es , - ing which t ime "appropr ia te o r n 1 t h o p h l-l1c v e c t o r s a re e v i d e n t l y sca rce o r absen t ..
precluding transmission of potent ial pathogens These resul ts
then i n d i c a t e another unique e f f e c t u f t h i s co lony 's l oca t i o n ,
and consequent impl i ca t ions for the members. The adul ts display a
behav ioura l t r a i t r e l a te d to the presence o f the f l i e s in t h a t

they remove f l i e s w i t h t h e i r b i l l s , and i t has oeen r e f l e c t e d upon nreviously that many hosts may be evolut ionar i1y adapted to
t he i r blood pare .es (Markus 1974).
4.3.4.The post - f ledging dependence pe r i od
Parental care extends beyond the pre- f lodging periods in i
wide var ie ty o f bi rds (Ashmole & Tovar 1968, Burger 1981, Diamond ,975, Feare 1975, Nelson 1976, Skut .h 19,6), i n c l u d i n g rap tor s
(reviewed in Brown & Amador 1968, Newton 1979, see also Sherrod
1983). I t i s , however , l a r g e l y u n s t u d i e d due to obv ious
d ' f f i c u l t i e s i n o b s e r v i n g the j u v e n i l e s a f t e r they f l y . Pa r t i cu l a r l y lengthy indiv idual PFDP lengths have been observed
in Crowned Eagles Stephanoaetus coronatus (11,5 months)(Brown
1966), Harpy Eagles Harpi a harpy ja (10 months) (Fowler & Cope 1954), and the C a l i f o r n i a Condor (7 months) (Koford 1953). In
most members o f the A cc i p i t r i da t , much shorter periods have been
documented (e.g. Johnson 1973, Newton ] 979 :Tab 1e 18, Sherrod
1983, Snyder & Wi ley 1976). New World Black Vu l t u res Coragyps
a t r a t u s have been observed to con t i nue to feed j u v e n i l e s away from the nest s i t e some six months a f t e r f ledging (Jackson 1975);
s im i l a r behaviour (away from the s i te) has not been observed in
any a c c i p i t r i d s p e c i e s . I n d i v i d u a l PFDPs have a l s o been
documented f o r Old World v u l t u r e s (Anthony 1976, Mundy 1982, Mundy & Cook 1975, Pennycuick 1976), a l though observa t i ons are
minimal. As a general ru le in raptors, the larger species remain
dependent longer (Newton 1979).Tne occurrence of prolonged parental care ce r t a i n l y seems to
be c o r r e l a t e d w i t h t he use o f s k i l l e d f e e d i n g methods,
exp lo i ta t i on of scarce foods and prof ic iency in accurate f l y ing
(Ashmole A Tovar 1968, Koford 1953, Stonehouse A Stonehouse
1963), and the improvement of these wi th age and experience (Dunn 1972, Lack 1968, Orians 1969). ' surplus of fa t , laid down during
the nest l ing and post - f ledging periods, would serve to t ide the j u v e n i l e over du r i ng per i ods o f ea r i y s e l f - f e e d i n g when i t s
s k i l l s were not mastered to the po in t wnere hunt ing was more l i k e l y to be successfu l (Cade 1982, Drent A Daan 1980, She-rod
1983). Using these reserves, a juveni le may survive approximately

90
r,ree weeks w i t h o u t i ;■ ■ latdnce I documented, a f t e r
t h i s engtn o f t ime the ' r e n i l t wa-, some t w o - t h i r d s c c i t s
normal" oody weight.The lack of proficiency in f l y i ng is probably accentuated in
young g r i f fons , whose wing loadings approach the possible maximum
Pennycuick 1972). This was i l l u s t r a t e d by the numbr,- o f days
that lapsed before the f i r s t ooserved foraging at tempt in three ) f the 1981 PFDPs (Table 9). Lack o f p r o f i c i e n c y in the f ac t o r s l i s ted above would act to increase the p robab i l i t y of mor ta l i t y
) f th is age-c l ass , and th is would account , p a r t l y , f o r the
v a r i a t i o n in l ength o f PTDP th a t i s o f t en observed ( t h i s study, Newton 1979). The lengths would also be i n f l uenced by the par t i cu l ar , local , food condi t ions : i t is c l ear l y advantageous
to the juven i le to a t ta in e f f i c i e n c y in feeding as soon as
possible.in pract ice, i t is very d i f f i c u l t to document the proport ion
if food supp l ied by the parents compared to the p ropo r t i on
obtained by the ju ve n i le through the d i f f e r e n t stages o f the
JPDP. A re l a t i v e l y higher concentrat ion of avai lable food would f a c i l i t a t e th is t rans i t i on (see also chapter six). This was borne
j t in observat i ons o 1 Oys te r ca t che rs Haematopus o s t r a i e g u s , -nere the age at which paren ta l feeding ceased va r ied w i t h the
ib itat , and consequently wi th food (Mor ton-Gr i f f i ths 1969).uveni le Cape Vul tures i n i t i a t e contact wi th t he i r parents,
j ' i not v i ce versa. In the breeding k l o o f t h i s occurs a t the s i t e only. The behaviour o f the j u v e n i l e t h a t never
- e v l s i t e d i t s s i t e , and consequent ly s tarved, emphasised t h i s i spec t : i t s parents made no a t t emp t to v i s i t i t a t i t s o f ten jsed perch po in t 15 in below them. Also, j u v e n i l e s spent very
l i t t l e t ime alone at th e i r nest s i t e s , and were c l e a r l y s t im u la te d to v i s i t them by the presence o f the parents.
Juveniles may so l ic i t food from adul ts other than t he i r parents, nowever my c i r c u m s t a n t i a l e\ lence i nd i ca tes t h a t parents i i f ferent ia te between s imi lar ly -aged of f spr ing and supply food to
their own o f f spr i ng only. Given what I consider the necessity of larental feeding aid during th is c r i t i c a l period, th is t r a i t is
• idapt ive value in increasing the probab i l i t y of the parents'
r ' product i ve , uccess. I t i s surel y s i g n i f i c a n t t h a t the

/
Aasvogelvlei Immatures ar r ived at Potberg some f ,5 nonth. i f - ’ f ledging : t he i r "dispersal " pr sumably fo l low-d -fDP,
than taking the place of i t .Al though Newton (1979) c i t e s two re fe ren t ‘ *
authent icated" cases of adul ts terminat ing the period by drivinc, t h e i r young away, these do not in f a c t r e f e r to such behaviour.
The over t aggressive terminat ions that I observed, together * V
Rowe's (1947) observat ions of a j uveni le Black Eagle, would V
f near to be the only recorded Instances in the Accipi t r idae.The t r ad i t iona l view of the re la t ionship between parent
o f fspr ing assumes un i la te ra l parental investment (Wi lson 19 however, the phenomenon o f p a r e n t - o f f s p r i n g c o n f l i c t *
apparent ly c l a r i f i e d by Tr ivers (1974), who interpreted i t as t!
outcome of natural select ion operat ing in opposite d i rect ions or
the two generat i ons . Tne t i m i n g o f the per iod o f c o n f l i c t is explained in terms of the inclusive f i tness of the j uveni le , and
o f the reproduc t i ve f i t n e s s o f the parent ; both f i t ne ss es ar
de f i ned in Wi lson (1975). In vu l t u re s w i t h such an extend* I
breeding cycle, the cost to the <emale in terms Oi her a b i l i t y *
produce an egg in the next season becomes c l ea r l y apparent as the
PFDP approaches that season's laying date. This was I l l u s t r a te d
by the Increase in frequency of emulat i ons during the PFDP as i t overlapped wi th the pre- lay ing period.
In overal l terms, a "choice" must be made between deferr ing
the next season's breeding at tempt and cont inuing to supply the j u v e n i l e w i t h food, 0 " t e r m i n a t i n g the PFDP. I f the former , the
j u v e n i l e ' s i n c l u s i v e f i t n e s s d im in i shes , but i t s r ep roduc t i ve f i t n e s s i s enhanced on ly i f i t could not have surv ived w i t hou t
the a d d i t i o n a l p a r e n t a l h e l p , c e t e r i s pa r i bus . F a c t o r s
I n f l u e n c in g t h i s " sw i t c h " are those t ha t necess i ta ted the
( l e n g t h y ) p e r i o d i n the f i r s t p l a ce : the j u v e n i l e ' srequ i rements , and i t s p r o f i c i e n c y in f l i g h t in r e l a t i o n to i t s
a b i l i t y to ob ta i n i t s p a r t i c u l food- type. My observa t i ons
i n d i c a t e t h a t Cape Vu l t u r es , a t l ea s t a t Potberg, at tempt breeding in the season f o l l o w i n g a successful c yc l e , thereby opt ing for the l a t te r .
The aggress ive i n t e r a c t i o n s between parents and t h e i r o f f s p r i n g t r ecogn i sab le to me) formed a subset o f a l l the

i n t r aspec i f i c in teract ions that I observed. The observat ions of
overt aggressive terminat ion of the per i od are tnen cons i ^ ten w i t h p a r e n t - o f f sp r ing c o n f l i c t theory d r i v e r s 1974), and are
not neccessar i ly "anthropomorphic and erroneous" (Brown & Amadon
1968). The case of the juveni le that was chased o f f the s i t e and subsequently fed at the s i t e , i s i nd ica t i ve of the extended t ime-
per i od o f c o n f l i c t o f f i t n e s s es of e i t h e r ‘ p a r t i c i p a n t " (as
i l l u s t r a t e d in hi Ison 1975: Figure 16-2).

93
CHAPTER FIVE : POPULATION DYNAMICS
5 . 1 . Int roduct ion
This chapter i s concerned w i t h breeding ra tes , surv iva l
rates and the e f f ec ts of th : consequent reproduct ive rate on the southwestern Cape (sub) popu la t i on . D i r e c t l y concerned w i th
numbers of ind iv iduals , i t documents the state ( including i n f l ux
and e f f lux ) of the populat ion as i t was during the study period,
and provides a framework for populat ion trend predict ion.G r i f f o n v u l t u r e s , together w i t h sea -b i rds and o ther large
rap to r s , have a very low rep ro duc t i v e ra te (Amadon . 964, Wynne
Edwards 1955). I n d i v i d u a l s can be d i f f e r e n t i a t e d according to
breeding s ta tus and c a p a b i l i t y ; the non-breeding segment then i n c l u d e s n o n - b r e e d i n g " b r e e d a b l e " adu l t s , sub-adu l ts and
immatures, e.g. f ou r year o l ds , and o ther immatures, e.g.
j u v e n i l e s . The term "breedable" r e f e r s to i n d i v i d u a l s t h a t are
capable o f breeding, but for various reasons are not act ive in a p a r t i c u l a r year , and i s used because some immature plumaged
ind iv iduals are f u l l y capable o f breeding ( t h i s study, see Newton ' 9 / 9 :T a b le 21). The demographic parameters associated wi th the
dynamics o f t h i s popu la t i on ( i n c l u d i n g those given by Brown & Cade 1972) are then cons idered separa te l y . Breeding success
dur ing the study per i od and f o r the prev ious s i x years i s
documented in chapter four (4.2.5.).
5.2.Informat ion concerning the demographic parameters
5.2.1.Popu1 a t i on size and st ructure
Informat ion per ta in ing to colony size p r i o r to t h i s study is
co l la ted in Boshoff & Curr ie (1 U). The number of act ive Potberg
nest s i t e s f o r the 1975 - 1963 per i od , as wel l as Aasvogelv le i 1981-1983, are dep i c ted in F igure 30. The t o t a l s o f a l l b i r d s present in the colony were very cons tan t (F igure 17), save for
the drop of some 17 per cent o f the t o t a l f o l l o w i n g the
h e l i c o p t e r i n c i d e n t (see 4 .2 .6 . ) . S i m u l t a n e o u s coun ts a t

94
I. :
Aasvogelvlei produced 82 (Potoei j 57) in 1981 and 74 (Potberg 59)
i n 1982 ( A . F . B o s h o f f , N.G. Pa lmer pers . comm.). F i g u r e 17 documents counts made j u s t before dark at Aasvogelvlei during the
study per iod.
30 -
37
,
n o . o f
a c t i v e
s i t e s 20 _
10
21
22
19
t7 17
11 15
14
1—1075 197ft
" r ""
'977 '973 1979 •380 1931 1982 1)83
Figure 30. The number o f ac t i v e nests a t Potberg (numbered) and
Aasvogelvlei for che l as t nine years.

Three immatures, r inged as n e s t l i n g s a t Aasvoge l v l e i in 1981, were res i g h te d a t Potberg (4.2.4.), and t h i s conf i rmed
inter -colony movement. As the colonies are 120 km apart ( l ine of
s i g h t ) , t h i s i nvo l ves some 2,5 hours f l y i n g t ime , using an average soeed o f 50 kph (Boshof f , Robertson & Norton in prep.,
Pennycuick 1972). These two co lon ies were t he re fo re assumed to c o n s t i t u t e a s i n g le popu la t i on . This popu la t ion was, in turn ,
assumed to be d i s c r e t e in terms o f i n f l u x o f b i r ds from o ther co lon ies : no r i nged b i r d from any o ther colony has ever been
found in the area, nor has any co l o u r r inged " f o re i g n e r " been res i gh ted ( t h i s study, Boshof f & Cu r r i e 1981). However one
immature, sighted in 1982 at Potberg and age-estimated as having
fledged in 1981, was not r inged and therefore not from Potberg or
Aasvoge l v l e i . This i n c i d e n t was the on l y suggest ion of i n f l u x f rom other, unknown, areas. A f t e r a newspaper r ep o r t which
documented vul tures at Perdeberg (Cape Times 15 Apr i l 1983), an
o l d s i t e in the Langeberg range (33 57S ; 21 32E), I checked the area, which is 18 <m SW of Aasvogel v l e i , in Apr i l 1983. I counted
28 vul tures and saw one Aasvogel v le i 1982 immature. The amount
o f "whi tewash" suggested t h a t t h i s l o c a t i o n was used f a i r l y f r e q u e n t l y . Cons ider i ng e f f l u x , a lumber o f b i r d s r inged at
Potberg have been resighted or recovered in various areas of the Cape Province (but not near any other colonies) before i n i t i a t i o n
of th is study iBoshoff 1981,Boshoff & Curr ie 1981:5). No s im i l a r
i n c i d e n t s were documented dur ing t h i s study. These instances c n f i rm t ha t immatures may d i sperse f rom t h e i r natal colony (Mundy 1982, Piper et 1981), and therefore cannot be assumed
as dead merely b e c . s e they are no longer r es i gh ted at the
colony.The f o rag ing range o f the Potberg vu l t u re s was recorded
i n d i r e c t l y (pos ta l survey) and d i r e c t l y ( r a d i o - t r a c k i n g ) : the
fo l lowing instances document sight ings made away from the defined
foraging range (see 3.2.1.). Se- -rate sight ings were made of one v u l t u r e (some i n d i v i d u a l ?) in February and A p r i l ly82, 105 km
WSW of the k loo f and ca 40 km SE of Aasvoelkop, a defunct vul ture roos t (Boshof f & Vernon 1980, I. B e l l , _ex P. Steyn j_n 11 t t .,
M a r t i n 1983). A s i n g l e a d u l t was observed 45 km SSE o f
Aasvogelvlei (118 km ENE of Potberg) in August 1982 (A.F. Boshoff

96
I n m t . ) . The three other sight ings given by Mart in (1983) were w i th i n a 60 km radius o f the k loof , some 1,5 hours vu l t u re - f l y i ng
t ime d i s t a n t . Records f o r the decade 1970 - 1979 are given in Boshoff a_L (1983:F,gure 17).
Age est imat ions of a l l b i rds in the colony (divided into two classes) were made at the s ta r t of each breeding cycle . Thus at the s t a r t o f the 1981 cy c le , 43 (75 %) were es t imated as f i v e
years o ld and o l de r ( "breedable" age). Seventeen breeding pa i r s
comprise 34 breeders, which const i tuted 61 per cent of the then cur rent to ta l o f 57 bi rds. The proport ion documented at the s ta r t o f the 1982 cyc le was o f 40 b i r d s f i v e years and o lde r (70 % of
the then c u r r e n t t o t a l ) . Thus 34/43 (79 %) o f "breedable" b i r d s were ac t i v e in 1981, and 36 / 40 (90 ») in 1982.
At Aas voge 1 v 1 e i , a maximun, o f 19 vu l t u r es was counted i n 1981, of which at least two were c l ea r l y immatures. Probably one
o f these was a c t i v e (5.2.2.) and in t h a t year , ten s tes were
a c t i v e , i .e. 20 breeders a t the colony. In 1982, 16 was the highest count, and of nine breeding pai rs, eight eggs were la id.
5.2.2.Deferred matur i ty
Vu l t u res are Inc luded in the group of b i r d s which show a deferred matur i ty by .,ot breeding in t he i r f i r s t few years, and
t h i s obviously a f fec ts the breeding dynamics o f the populat ion in question. In 1981 then, o f 34 act ive breeders 2 were est imated by
plumage characters as immatures and 2 o f 36 in 198?. Cer tainly at two s i t e s t h a t were a c t i v a te d f o r the 1982 cyc le , both females
had brown eyes and were es t imated as no o l de r than f ou r years (and into t h e i r f i f t h ) . Of seven ringed breeders in 1981, and six in 1982, only two were of known age (Robertson 1983b), as others had e i t h e r a metal r i n g ( f o u r ) , or had miss i ng co lou r r i ngs
( two) . Of the known age p a i r in 1981, the male was s i x years old (and i n to i t s seventh) and the female four years old ; i t i s
poss ib l e , but unknown, whethe t h i s same p a i r us tne same (ac t i ve ) s i t e the prev ious cyc le . A female, w i t h a miss ing r i ng and only observed in 1981, was e i t h e r r inged in 1974 or 1975 i f
r i nged at Potberg : more l i k e l y the l a t t e r , i t was then f i v e years old when i t produced an egg in 1981. And the age of another

(male) wi th missing r ings could not be determined wi th adequate
accuracy. Al l three breeding adul ts wi th metal r ings could have been r inged in any year between 1951 and 1973 (Boshof f & Cur r ie
1981). A female ringed in 1977 at Potberg commenced breeding in
the 1983 cycle (possible mate replacement, see 4.2.1.), then six years o l d and i n t o i t s seventh year . I t was observed as not
breeding in the previous two years.No »*i nged b i r d s breo a t Aasvogel v I ei in e i t h e r year . In
1981, A.F". Rr>shoff and I c lassed only one breeder (subsequent
dead egg, as an immature, and est imated i t at four years old.
5.2.3.Frequency of active sites, and of breeding
As wi th other raptors, ind iv iduals show breeding per i od ic i ty by net breeding in p a r t i c u l a r years : t h i s i s p a r t i c u l a r l y
ev i den t in a colony where a l l "breedabIe" b i r d s are v i s i b l e . I f
i t i s assumed that pai rs (or a t least one of the pai r ) occupy the
same s i t e when ac t i v e in two consecut ive years , breeding
frequency can be determined by examining act ive s i tes as wel l as observ ing recogn isab le i n d i v i d u a l s . Cons ider ing 64 s i t e s over
eight years (average annual increment of 1,6 new s i tes documented per year from 1975), the same s i te was act ive in two consecutive
years 15 t imes (2 years : 15) ; in 3 years : 8 t imes ; in 4 years : 4 t imes ; in 5 years : 4 t imes and in 6,7 and 8 years : tw i ce
(data from CDNEC records). I f two adjacent years are considered over the seven cyc les , 67 per cent or two t h i r d s o f the s i t e s
that were act ive in one year were act ive in the next (range 45% - 8 3%). Fourteen o f seventeen (82v ) ac t i v e 1980 s i t e s were ac t i ve in 1981 (15/18 or 83., in 1982). Thus during the study period, one f i f t h of the s i tes " l a i d o f f " between the two years, as compared
t o a f a c t o r o f one t h i r d f o r the y e a r s 1975-1982. Th i s corresponds to a pai r breeding at a spec i f i c s i te at a frequency of about two in every three years.
Of seven recogn i sab le in , / i dua l s ac t i ve a t s i x s i t e s in 1981, f i v e were a c t i v e at the same s i t e ( f ou r s i t es ) the fo l lowing year, a decrement o f less than one th i rd . Both bi rds of
the co lour r inged p a i r were a c t i v e a t the same s i t e in both years.

5.2.4.Age-c lass survival rate ,
The f o l l o w i n g e v i d en c e conce rns s u r v i v a l of marked i n d i v i d u a l s , f i r s t - y e a r im ma tu res and o t h e r age groups separately, by documenting how long each was seen in the colony
(depic ted in F igure 31). The a l t e r n a t i v e reason ( to death) o f non-sight ing, of dispersal and r ing loss, must be borne In mind.
One opened metal r ing, of a b i rd colour ringed in 1975, was found
a t the base o f NWl c l i f f in 1982. I t i s poss ib l e t ha t t h i s individual was act ive in 1981 (Figure 31, 'YW').
1981 1982-i—4— 1— 4— I— l— I-
met.1.295:met.1.7
gyr
gyw =m e t. r .26-1-
ma rk i ng
on b i r d
1983
- + — t -
zz--.e t . 1 .1 ; — :
■
A M J ' J A ) N J J I H A " j J . S O N 0
Figure 31. diagram depict ing the length of t ime ringed vul tures
were res i gh ted in the k l o o f . The dark shading i nd i ca tes b i r d s
r inged in 1980 ( r sy , bsw, bsg, bry) and dur ing the study per iod.
Ihree immatures ringed at Aasvogelvlci were resighted from Apr i l 1 W . For met.1.29, see t ex t "Adult survival " .

99
Immature survivalOnly two of e i g h t 1980 f l e d g l i n g s were d e f i n i t e l y seen in
the kloof up to one year a f t e r f ledging ; however two others were no t r inged. Thus no more than f ou r remained a l i v e and in the
k loo f throughout t h e i r f i r s t year, al though th i s f igure is most
l i k e l y 2/8 (Table 13.). As observat i ons began some three months a f t e r f l ed g i ng , the average of 44,7 per cent i s probably lower
than the "t rue" value.
Table 13. The number of months, in the year a f t e r f ledging, that
juveni les were resighted in the kloof . As bi rds from nests 55 and 30 were not co lour r i nged, t h e i r presence in the k l o o f was
uncertain.
Bird no. months a f t e r » of max.
f ledging that b ird posslb.e
was resiqhted
RSY 12 100
BSW 3 25
ese 12 100
BRY 4 33
n.65 m m .6 50 (? )
n.30 min.6 50 (? )
8RG 0 0
WYB 0 0
Average 5.4 44.7
,>,R 10 83
8SR 4 33
WYR 2 17
WBfi 0 .5 4
RSG 0.3 2.5
A ">Y 12 100
Average 4 .8 40.0
O m b tued average 5 .1 42 ?

In 1981, a l l n e s t l i n g s were r inged and cnly one was
res igh ted in tne k l o o f one year a f t e r f l edg ing . On average, immatures rema ned for 40 per cent of the maximum possible periodin t h e i r f i r s t year . The mean f o r ihe two years i s one o f 43 per
cent "survival " in the f i r s t year (Table 13).
Of seven t h a t f 1 edged in 1982, qnly one was present i n thek l o o f s i x months l a t e r . Three were recovered dead w i t h i n o e
month of f ledging.Thus, fo r these three years combined, f o u r (or a maximum of
six) of 21 immatures were resighted a l i ve one year a f t er they had
f ledged, a "surv ival " rate of some 19 - 29 p*r cent for t h i s age
class.
Adul t survivalOf the seven recognisable ind iv i dua l s act ive at six s i tes in
1981, f i ve were present one year later. One "missing" b i rd had a
metal r ing only, thus could have easi ly been missed as i t was no longer seen a t a nest s i t e in 1982 and cou ld have been confused
wi th other, s i m i l a r l y r inged, bi rds (Figure 31: met.1.29). Adult su r v i v a l i s also i nd i c a te d in the t o t a l counts (F igure 17).
Al though the speed of nate replacement i s unknown, no breeder was
considered to have died wh i l s t engaged in a cycle ; cer ta i n l y no r inged pa r t ne r went miss ing. One (non- r ingod) ad u l t t h a t died
during the study period was found (an est imated six weeks a f t er deatn) 4 n f rom the top of the w a t e r f a l l a t the head o f the
kloof .
General survivalTable 14 documents the b i r d s tha were res igh ted in 1981,
according to when they were r inged. I spent a large amount o f
t ime in observat ion and, as a l l perch areas were v i s i b l e from the observa t i on p o in t , am c o n f i d e n t tha t I observed every r inged
b i r d . Assuming t h a t the co lour r inged v u l t u re s had not l o s t t he i r r ings, of 123 bi rds that id been ringed up unt i l 1980, 14
(11,4%) were sighted M 1981. I f "breedable" colour r inged birds alone are cons idered ( i .e . 1974 - 1976), f ou r of 48 (8,3%)
surv ived to breed in tne k l oo f . Of course, an unknown number of
non ringed bi rds from those years may or may not be breeding.

101
Table 14. The number o f ne s t l i n g s t h a t were metal r inged and
colour ringed at Potberg (data from Boshoff & Curr ie 1981, CDNEC records), a;,J the number tnat were subsequently resighted during
the study per iod. Two i n d i v i d u a l s ( xa , xb ) w i th miss ing r ings
could not be accurately placed.
Year no. nestlings
metal ringed
no. nestlings
colour ringed
no. n est l ings
not ringed
no. resighted no. resighted no. breeding
in 1981 in 1962
p re - '7 2
1972
19/3
1974
1975
1976
1377
1978
1980
12
14
14
22
15
11
13
7
9
6
??
min. 6
2
8
5
2
2
1 2
},ab
ab
,ab
ab

102
b. j .DIscussion
5.3.1.Pnjulat ion s t r u c t u r e
The has cer ta i n l y decl ined in numbers wi th in l i v ing
memory, but i t appears to have stabi l i sed. The drop in the number
of act ive s i tes between 1975 and 19/b (Figure 30) would suggest a sudden dec l i ne in colony s t r eng th , a no i t i s not u, l i k e l y t ha t
th i s was due to an i nd i r ec t poisoning event. Breeding a c t i v i t y in
the subsequent y e a r s sugges t an absence o f any form o f persecution and, as a resul t of discussions wi th farmers in thearea (chapter th ree) , I would cons ider such an event to be
u n l i k e l y in the f u t u re . Nonetheless, the co lo n ia l nes t i ng and
feeding hab i t s o f t h i s species do render a colony o f t h i s s ize very vulnerable to sudden exterminat ion by as l i t t l e as a single,
a l b e i t u n i n t e n t i o n a l , po isoning event . That the a v a i l a b l e
evidence suggests that i t is a discrete populat ion increases the
p o s s i b i l i t y o f such a "c lean sweep", and also ra i ses quest ions concerning the v i a b i l i t y , i n genet i c terms, o f the populat i on,
hese aspects are discussed fur ther in chapter six.A f i g u r e of some 30 per cent o f i mmatures in 1982 r e f l e c t s
e i t h e r a high ad u l t su r v i v a l or a r e l a t i v e l y low j u v e n i l e
su r v i v a l r a te : tne (suggested) high m o r t a l i t y o f r inged b i rds would i n d i c a t e the l a t t e r . However, the numbers o f b i r d s at Aasvogelvl r i serves to emphasise that colonies may d i f f e r in the
y oport ions of cer ta in age-classes counted, pa r t i cu l a r l y i i the
colony is checked on l y s p o ra d i c a l l y . That even less b i r d s than
double the number o f ac t i v e nests were counted in both years save for one count) indicates a d i f f e ren t foraging behaviour to
that at Potberg (or a very d i f f e re n t breeding strategy I). Also,
i t i s known th a t the popu la t ion o f Cape Vul tu res i s s p a t i a l l y Heterogeneous wi th ? . least, four components (Mundy 1982, Piper et
ij_. 1981). However a t Potberg, where colony numbers were very . t ab le , the p ro po r t i on of "bn- dab 1 e" b i r ds breeding in e i t h e r
year cannot be considered low, i f i t is remembered that factors such -is an (unknown) shew in the male : female ra t i o in a colony
t h i s size would inf luence partner a v a i l a b i l i t y considerably For -xar.ple, in the C a l i f o r n i a Condor popu la t i on o f some 60

103
I n d i v i d u a l s in 1946, \ fo rd \ 19; , r r ent
proport ion of male-.Both the lo c a t io n and th^ p t • ,enc - Aasvogei v l e i
immature suggested that the vul tures at ^erdeberg "ormed part of the 1982 A usvoge lv le i popu la t ion . ' t - . poss ib le th a t at least one breeding a t te m p t occurred in the .angeberg in 1981 (e.g.
Perdeberg ?), and th is may explain the presence of the non-ringed
immature I saw at Potberg. As only three vul tures were counted at
Aasvogel v l e i in January 1983 and the two co lo n ies are so close together, I assumed that the birds counted at Perdeberg were the
same as those that were at Aasvogelvlei in 1982.
5.3.2.Deferred m a tu r i t y
The (successful ) four year old female oreeding record is the
youngest age of a documented g r i f f o n vu l tu re breeding attempt , a European G r i f f o n Vu1tu re Gyps fu lvus was found breeding in the
w i l d a t f i v e years o f age (Terrasse 1977) and a c ap t ive G.
c o p ro th eres bred u n success fu l ly in i t s s ix th year (Mundy 1982).
Mundy (o£. c i t .) then assumed the age o f f i r s t breeding f o r the species to be s i x years . But why do b i rd s not breed in th e i^ immature years ? According to Lack (1968:297) ".. they (b i rd s )
have evolved physiological regula t ing factors to prevent ea r l ie r
m a tu r i t y because t h i s .. enables them to leave more o f f s p r i n g than i f they mature e a r l i e r . This view im p l ie s , f i r s t , t h a t any Ind iv idua ls which attempt to breed when younger are un l ike ly to succeed and, secondly tha t the attempt to breed exposes the birds
to a higher m o r ta l i t y than i f they do not attempt to breed." This v iew ( c o r r e c t l y ) s t r e s s e s c e r t a i n c o n s t ra in t s o f natura l s e le c t io n , and i t a lso de f ines the phenomenon o f de ferred
matur i ty as an adaptation (sensu Wil l iams 1966).I t i s known th a t young b i r d s have a high m o r t a l i t y ra te
( e.g. P iper e_t aj_. 1981, 5.2 J . , presumably because they are less e f f i c i e n t in f i n d in g food than o ld e r b i rd s (Lack 1968, c ' i p t e r fou r ) . A t r a n s i t i o n to e f f i c i e n t feeding as a r e s u l t of
e; r .r ience gained could then be viewed as t id in g the indiv idual over a se r ies o f le v e ls (Newton 1979:136) or energy thresho lds
(Drent & Daan 198U, Vernon & Robertson 1983). The end r e s u l t i

an Indiv idual capable o f accumulating s u f f i c ie n t reserves so that i t is able to accommodate each l e v e l , and thereby exceed the (summated) breeding th resho ld . The balance o f an i n d iv i d u a l ' s
energy budget i s then expected to vary seasona l ly (e.g., w i th re la t i v e food abundance in re la t ion to requirements, q.v. Houston 1976), annually and w i th age, where age is l inked w i tn increasing exper ience (Vernon & Robertson 1983). Viewed in i t s c o r re c t
pe rspe c t ive then, de fe r red m a tu r i t y i s a consequence ( sensu
Wi l l iams 1966) of Inexperience.In d i v i d u a l v a r ia t i o n s in the age o f f i r s t breeding would
then re su l t from environmental v a r ia t i o n s , and consequent food a v a i l a b i l i t y (Lack 1968, Newton 1979), as well as in t raspec i f ic
r e la t i o n s (e.g., a v a i l a b i l i t y o f mates and nest s i t e s ) th a t i n f lu e n c e a t ta in m e n t o f the breeding thresholds. These factors
can be expected to vary between the d i f f e re n t colonies throughout southern Afr ica.
5 ,3 .3 .Survival
Given the assumption that ringed vultures have not los t a l l t h e i r r in g s , whether the "m iss ing " v u l tu re s are dead, or a l i v e
elsewhere, the fac t remains that they do not const i tu te part of
th is populat ion's breeding segment.Survival est imates for th is species have been made, based on
r ing recoveries (Houston 1974b, Piper et ah 1981), although the
authors have c r i t i c i s e d the r e s u l t s (o£. c i t . ) , e.g., " c l e a r l y
nonsense" (Mundy 1982:267). Indeed, i f based sole ly on recoveries
o f dead b i r d s , a l l es t im a tes o f a g e -s p e c i f i c s u r v i v a l , tha t assume a constant survival rate for pa r t icu la r age-classes, are
l i a b le to be untrustworthy (l.akhani & Newton 1983). Also, in the
absence of re l ia b le estimates, pred ic t ion of population numbers cons t i tu tes l i t t l e more than a numerical exercise. For example,
the expected annual increment "breedable" v u l tu re s from the Potberg colony was c a lc u la te d using past breeding data and
es t im ated a g e -s p e c i f i c s u rv iv a l ra tes o f 40 per cent p.a. f o r f i r s t years, 35 per cent p.a. over the next four years and 92 per cent p.a. fo r a d u l ts . From December 1982 to December 1987, the
p op u la t ion "dropped" by f i v e b i rd s : i f the a d u l t su rv iva l

105
est imate was increased to 95 per cent p.a., the population would
show an increase o f two ! These c o n s id e ra t io n s do, however, emphasise the overwhelming importance o f high a d u l t su rv iva l rates, for animals with such a low reproductive rate. As w i th the Andean Condor V u l tu r gryphus and the C a l i f o r n i a Condor, these
v u l t u re s d is p la y such low re p ro d u c t iv e ra tes th a t t h e i r p o pu la t io ns cou ld not p e r s i s t except f o r the e x t ra o rd in a ry
longevity of the adults" (Mertz 1971).

/
1C*
CHAPTER SIX : GENERAL DISCUSSION
The e v o lu t io n o f e x c lu s ive scavengers I s proposed' to have occurred under c e r t a i n c o n d i t io n s , as descr ibed by Houston
(1979). The proposal is based on studies of g r i f f o n vultures and other scavengers in the re la t i v e ly undisturbed (by man) s i tua t ion
in the Serengeti in East A fr ica (Houston 1972, 1974a, 1976, Kruuk 1967, P e t r id e s 1959, Pennycuick 19 72). In a prev ious paper,
Houston (1974a) remarked that the s i tua t ion in the Serengeti ".. may not be typ ica l o f the a c t i v i t i e s o f g r i f f o n vultures in other reg ions o f A f r i c a today". As one stands on top o f the Potberg
today (F igu re 3), i t is indeed d i f f i c u l t to re - c re a te , in one's mind, the scenario o f Coastal Rhenosterbosveld supporting numbers o f Bontebok and other bovids (chapter two). And i f the degree of
man's in f lu e n c e on l i f e communit ies in general is viewed as a continuum (Myers 1979), the remnant v u l t u r e po pu la t ion in the
sou the rn -sou thwes te rn Cape is su re ly p o s i t io n e d towards an e x t r e m i t y . This s ta tement needs, however, to be q u a l i f i e d . In
terms o f t h e i r feed ing , the b i rd s are t o t a l l y dependent on the stock farmer, whereas in the breeding area the s i tua t ion in the
k l o o f remains remarkably und is turbed . Throughout t h e i r range then, Cape Vultures have undergone a marked t ra n s i t io n in feeding
•co logy in the l a s t two to three c e n tu r ie s and the aim o f t h i s study was to examine the nature and e f fec ts o f th is a l te red food
supply on the reproductive performance o f a p a r t i c u la r colony.Food is considered l im i t i n g i f i t prevents a population from
increasing and i f s u f f i c ie n t , i t must be so in quant i ty , qua l i ty
and a v a i l a b i l i t y to the animal concerned (Newton 1980). I
attempted to determine how much food would be avai lable to the vul tu res i r the area surrounding the colony by incorporat ing tne
various factors that inf luence '.s a v a i la b i l i t y . I then attempted to determine how much food i n d i v i d u a l s , and the whole colony,
a c tu a l l y ob ta ined. The former was determined by ( i n fo rm a l ) personal in terv iews w ith farmers, as well as a postal survey ; I
cons ider th a t both o f these methods f u l f i l l e d a va luab le
e x t e n s io n r o l e . The l i m i t e d n a tu r e o f the fe e d in g range
determined by these methods, and subsequently supported by the

107
r a d io - t r a c k i n g study was expected, given th a t i t s s ize is dependant on the ( summated) requ irements o f i n d iv i d u a l colony
members. In passing, i t is o f in te res t to note that the existence o f a feed ing range is not n e c e ssa r i ly an e f f e c t o f man's
in f lu e n c e s . Although g r i f f o n v u l tu re s o r ig i n a t e d under and are most s u i te d to c e r t a i n c o n d i t io n s (Houston 1979), pe r iphe ra l
p o p u l a t i o n s , in a reas o f low o r no m i g r a t o r y u n g u la te c o n c e n t ra t io n s , would have fed on the carcasses o f re s id e n t
species. The sou them-southwestern Cape would have beer such an area (chapter two).
Practica l d i f f i c u l t i e s with the est imat ion of crop content
o f i n d iv i d u a l b i rd s re s u l te d in t h i s sec t ion being not as accurate as I had o r ig in a l l y hoped. In re trospect , I should have concen t ra ted , du r ing the ob se rva t ions , on a more r igo rous
documentation of a smaller number o f nests (e.g.those on NW2).
Thus in c o n t r a s t to the S e re n g e t i , where food va r ie s seasonally in d is t r ib u t io n and abundance (Houston 1974a), food at Potberg v a r ie s in abundance on ly . The I eve, f v a r i a t i o n in the
q u a n t i t y a v a i l a b le w i t h i n an area o f some 60 km rad ius o f the colony (excluding south !) is considered highly un l ike ly to f a l l
below the colony's requirements : in general terms there is thus a s p a t ia l s u f f i c i e n c y . A s i m i l a r co e lus ion was reached w i th
Gymnogyps ca1i fo rn ianus , another large-carcass scavenger (M i l le r e t a_L 1965). In terms o f temporal a v a i l a b i l i t y o f food, the
evidence I ob ta inea in d ic a te d th a t both a d u l ts and n e s t l in g s b ta ined s u f f i c i e n t q u a n t i t i e s dur ing the n e s t l i n g per iod.
Although th is period coincides wi th inclement weather , and 1961
was exceptional ly wet in th is respect, the condit ion of nestl ings as in d ic a te d by growth in both wing length and body mass
suggested th a t the n e s t l i n g s nonetheless rec ieved s u f f i c i e n t
quant i t ies of food.But is t h i s food a v a i la b le to a l l age groups ? Young
v u l tu re s are less e f f i c i e n t a ‘ o b ta in in g food than o ld e r b i rds (Houston 1976, Mundy 1982), and a t Potberg young b i r d s have a
very low s u r v iv a l ra te (chapter f i v e ) . Ju ve n i les may a lso have very lengthy PFDPs, and i f the length o f the Pf'DP is i n d i c a t i v e
o f the a v a i l a b i l i t y o f food to th a t age c lass (chapter fou r ) ,
then my ob serva t ions r e f l e c t a d i f f i c u l t y tha t those juveni les
experience in o b ta in in g food. Are the chances o f an immature

108
ind iv idua l being behavioural ly excluded by adults from ingesting food g rea te r a t a sma I 1 ( e.g. sheep) carcass than a t a la rge (e.g
h e r t e b e e s t ) c a rc a s s ? Two f a c t o r s c o u ld r e i n f o r c e t n i s disadvantage. F i r s t l y , the s m a l le r carcass would presumably be eaten up f a s t e r , and by fewer b i rd s and secondly , a d u l t s , which
outnumber j u v e n i l e s by a r a t i o o f some three to one (chapter f i v e ) , g e n e ra l l y i n i t i a t e feeding at the carcass (Mundy 1982).
Th is proposal i s e s s e n t i a l l y a c o r o l l a r y o f t h a t o f F ishe r 's (1952), which accounted fc the surge in numbers o f the Fulmar
Fulmarus g lac ia l is . the i n i t i a t i o n and development of whaling prov ided a r i c h and e a s i l y a v a i la b le food source (discarded
o f f a l ) t h a t acted to increase the s u r v iv a l ra tes o f the younger age classes. The very small numbers of ringed b irds surviv ing to breed at e i th e r colony (Table 14) h ighl ighted the current lack of
r e c r u i tm e n t (and, consequent ly , an im p o r ta n t advantage o f the r in g in g programme !). A f law in my study was then a lack o f
carcass-watches th a t would have provided relevant information. Comparative PFDP lengths from o the r c o lo n ie s , e.g. those in the
Transvaal where the food source is d i f f e r e n t , would also I l l u m i n a t e t h i s aspect . This proposal i s , however, te s ta b le . A l i m i t e d supplementary feeding scheme, soeci f i c a l ly designed to
ass is t juven i les through th is period of t ra n s i t io n (chapter four) has been i n i t i a t e d a t Potberg, and is o u t l i n e d in appendix a.
N e s t l ing s w i l l con t inue to be co lo u r r inged fo r a t leas t f i v e yea rs , and du r ing the months a f t e r f le d g in g , the j u v e n i l e s ' use
o f t h i s source as w e l l as t h e i r s u r v i v a l , w i l l be monitored (CDNEC-A.F. Boshoff pers. comm.). Other th ings being equal,
documentation o f subsequent j u v e n i l e s u r v iv a l ra te s w i l l then
al low fo r a conclusion regarding the proposal's v a l i d i t y . These v u l tu re s have thus achieved a t r a n s i t i o n in t h e i r food source,
bu t I suggest t h a t t n i s t r a n s i t i o n has not been achieved w i th
equal sucess by a l l age groups.
The evidence indicates that th is jub-populat ion const i tu tes a d is c re te e n t i t y (chapte r t ve ) : d i s t r i b u t i o n , maps should
in c o rp o ra te t h i s (e.g. Newman 1983, Steyn 1982), as the nearest known breeding conspeci f ics are i . the no r theas te rn Cape, some
500 km from t h i s area. In the Cape Prov ince, t h i s species has undergone f l u c t u a t i o n s in numbers in the past, * 1 though i t is
l i k e l y t h a t i t s range was continuous u n t i l approx im ate ly one

cen tu ry ago (Boshof f & Vernon 1980). I f one assumes I s o la t i o n since then, and th is is most l i k e l y an over-est imate o f the t ime- span, some f ive iso lated generations are involved. What then are
the consequent imp l ica t ions for the remnant group ? Obviously a
smaller size increases the chances o f e l im ina t ion in a pa r t icu la r area, and the CDNEC has attempted to reduce the l i k e l i h o o d o f
such an event, e.g. in d i re c t poisoning, by means of considerable extension work w i th farmers. Less apparent are b io log ica l e f fec ts
o f such s ize and i s o l a t i o n , and as these two a t t r i b u t e s are unique to th is population i t is re levant to consider them in some
d e t a i l here. For example, in small popu la t ions he te rozygos i ty decreases through the a c t io n of genet ic d r i f t , a l though in
populat ion sizes o f 100 to 1000 the loss of v a r i a b i l i t y is s l igh t in each generation 'Roughgarden 1979). Also, only one indiv idual
migrat ing between "iso la ted" groups per generation is enough to counter the e f f e c t o f d r i f t (o£. c i t .) : i t i s th e re fo re h ig h ly
un l ike ly that these vul tures display any phenotypic disadv cage resu l t ing from th is e f fe c t alone. Secondly, a recent analysis of
the e f f e c t s o f inb reed ing accounted fo r the h igher m o r t a l i t y
ra tes o f inbred ca p t iv e young ungula tes (Ba l lou & RaM< 1982).
The frequency o f inb reed ing in the c o lo n ia l (e.g. g r i f f o n v u l t u re ) s i t u a t i o n , where b i rd s appear to breed in t h e i r nata l
areas (chapter four) , is presumably greater than tha t in so l i ta ry
breeders, other things being equal. Consequently, on the basis of
t h i s a lone, i .e . exc lud ing any poss ib le behavioura l t r a i t s tha t might prevent inbreeding, oie would expect a r e la t i v e l y low level o f h e te ro z y g o s i t y in t h i s sub-populat ion. Bal lou & Ralls (1982)
concluded by emphasis ing th a t an awareness o f the e f f e c t s o f
inb ree d ing , such as decreased f e r t i l i t y , i s v i t a l to the sound conservation management of small ( isolated) populations. Genetic
d r i f t and inbreeding then re su l t in decreased heterozygosity, and
a recent e lec trophoret ic study of allozymes and soluble proteins
o f Cheetahs Ac inonyx jubatus vealed dramat ica l ly low levels of var ia t ion (O'Brien et aj_. 1983). These levels could not, however,
be l inked to any (deleterious) phenotypic effects . Nevertheless, i t should be borne in mind tha t d e le te r io u s e f f e c t s are not o f
necessity a d i re c t consequence of reduced v a r i a b i l i t y : the l ink c e r t a i n l y has been demonstrated (Ba l lou & R a l ls 1982 & re fs ,
therein) , but not (as yet) to be causal . Furthermore, any study

o f t h i s i n t e r e s t i n g aspect (e.g. between southern Cape and northern Transvaal vultures ?) should c r i t i c a l l y question which p r o te in s the study is to be based on : O'Brien et ̂ aJL (1983) did
not consider th is .Co lon ia l n e s t ing provides opportun it ies fo r ind iv iduals to
observe where o the rs go in search o f food (Loman <4 Tamm 1980, Ward & Zahavi 1973) and, given the proposed 1 ink between a migratory food source and the evolut ion of these scavengers, The
advantages acc ru ing to an in d iv id u a l nesting near conspecifics
are apparent . Furthermore, where the food source is evenly d i s t r i b u t e d th roughout a p a r t i c u l a r area (e.g. feeding range), the network o f i n d i v i d u a l s spreading over the area (Houston
1974a) together w i th local enhancement (Hinde 1970/ at a carcass c l e a r l y f a c i l i t a t e the d iscovery o f food. Where numbers in the
colony d e c l in e , i s the re a c r i t i c a l leve l below which these advantages e f f e c t i v e l y break down ? The r a d io - t r a c k in g study v i v i d l y demonstrated the c a p a b i l i t y o f an in d iv i d u a l to cover
la rge t r a c t s o f ground r a p id l y and a persona l , v is c e ra l , understanding i s t h a t under t h i s a l t e re d food regime the advantages of co lon ia l breeding, in terms of ac tua l ly f ind ing the
carcass, are not as g rea t as they were under the c o n d i t io n s of the "Serengeti scenar io " . The Potberg colony i s c u r r e n t l y a t a reduced leve l, i.e. there are vacant nest s i tes in the k loo f that have been successful in the past, and in an area of apparent food abundance. Is the tendency o f t h i s sou the rn-sou thwes tern Cape
population to sp l in er in to smaller bre- ng groups a re f lec t ion o f th is decreased advantage, in combination w ith other, unknown,
social factors?T h is s tu d y has d e m o n s t ra ted t h a t these v u l tu re s are
essen t ia l ly se l f -sus ta in ing under the condi t ions of .he "farmland
s c e n a r i o " , g iv e n a lack o f p e r s e c u t i o n and a c o n t in u e d
wi l l ingness on the part of the farming community to ( in d i rec t ly ) support them. The resu l ts of ’<* l im i te d feeding programme should provide for a conclusion as to whether manipulative management procedures are neccessary in order to maintain the i r presence in
the area. Of equal importance, I t r u s t t h ^ t the study has i l l u s t r a t e d aspects o f t h e i r l i f e h i s t o r y , and in d ica ted those
that warrant fu r the r investigation.

APPENDIX A
An a c t i v e management plan f o r the co lony , o f which the
1 im i te d feeding programme forms a major p a r t , was accepted in
p r i n c i p l e by the CDNEC a t a meeting in A p r i l 1983. The lo c a t io n o f the feeding s i tes on pr iva te property was accepted by the farm owner in October 1983. Most o f the p o in ts l i s t e d in the sec t ion " P u b l i c i t y campaign" have been at tended to. The management
proposals are out l ined in th is appendix.
Aims
a. To improve the survival rate o f f i r s t yeat birds.
b. To increase the breeding success.c. To remove, or lessen, the threat of poisoned carcasses in the
fo rag ing area, as we l l as reduce the p e s t i c id e le v e ls th a t are
cur ren t ly found in addled eggs.
Manipulation of _che food supply
Although research in d ic a te s temporal v a r ia t i o n s in the
quanti ty of food avai lable to the vu l tu res, a spatial suf f ic iency i s concluded to e x i s t in the area surrounding the colony. The
time of year during which newly fledged birds are in part icular
need o f an adequate and e a s i l y a v a i la b le food supply (due to nexperience in f l y in g and foraging a b i l i t i e s ) coincides w i th one
such p e r io d . T h is s tud y h i g h l i g h t e d the c r i t i c a l l y low r e c ru i tm e n t to breeding d isp layed by the Potberg co lony, m o r ta l i t y being highest during the months fo l lowing fledging. Thera tionale for a l im i ted feeding programme is discussed fu r the r in
chapter s ix . I t is th e re fo re considered th a t a supplementary
feeding programme be implemented dur ing the January to June
period.[ he supply o f uncontaminated food w i l l a lso ac t to reduce
the le v e ls of contaminants c u r r e n t l y found in eggs as we l l as
ensure an adequate supply during the months preceding laying.

112
Deta i ls of the feeding programme
A supplement o f no less than 1U0 kg o f carcass per week
should be provided. A supply o f bone fragments in the size range 1-5 cm obtained from previous carcasses should be made avai lable
the s i t e . This supplement must be in the form o f whole
carcasses to a t t r a c t the b i r d s , p rov ide a balanced d ie t and simulate natural feeding behaviour. Carcasses must be s l i t along the stomach to al low the vul tures entry in to the body cavi ty , and
must be removed f o r burn ing or b u r ia l not l a t e r than two days a f te r placement so as to prevent the accumulation of b lowf ly eggs
(and consequent farmer antagonism).To ensure that the vultures re ta in the ’ r foraging behaviour,
th ree a l t e r n a te feed ing s i t e s should be used. The approximate
location of these is shown in Figure 1.
irteae 3.
JP
Figure 1. The approximate location of the feeding s i tes (marked
as stars) in re la t ion to th' olony (arrow).
Although i t would be ideal to have three s iu c / in the D
Hoop Nature Reserve, t h i s is not poss ib le as the colony 's fo rag ing area, determined ( to an unknown ex ten t ) by prevai 1 mg
winds and the orographic e f fec ts of the Potberg, f a l l s mainly to

113
""r."":.".-..... .Duration of the feeding programme
considered v i t a l :_ f i r s t year survival rate - number of eggs la id per year . number of b i rds fledged per year
fo r a minimum of f ive years.
D i s t u r b a n c e at the co1on^
W m

, w r? i j 'iu ■ « 'i".
uu l ie i ly nflp't; .r
■ nv .1 i in here i s to each e v e , ) -.tock f . r m e r w i t h in the fo ray ing range or the Potbery and Aasvoge1v 1ei co lon ies . The
f l i g h t o f the v u l t u r e s should be made aware to the farmers as wel as the advantages which vul tu res have fo r them. The colonies emain extremely vulnerable t the threat of poisoned carcasses n the fo rag ing area to combat in se c t plagues, e.g. b lo w f ly ;
here, one carcass has the p o te n t ia l to k i l l h a l f the breeding iegment o f the Potberg colony. Thus an appeal must be made to p r e v e n t i a lbe i t accidental poisoning event.
,onciusio,
M o r t a u t y lu r in g e r t a m stages o f the breeding cyc le
.u r ren t iy l im i t s recru itment to the reproductive segment o f the
.o iony. Even in the absence o f o ther m o r t a l i t y fa c to rs (e.g. a
ooisoning event, c human d is tu rb a nce ) , such c r i t i c a l l y low re c ru i tm e n t le v e ls w e v e n tu a l ly re su l t in ex t inc t ion of the
: o 1ony. the programme o u t l i n e d above i s designed to a l l e v i a t e s t re ss a t the p a r t i c u l a r stages th a t e x h i b i t high m o r t a l i t y
ra tes. M o n i to r in g the e s s e n t ia l demographic aspects o f the population w i l l enable an evaluation of the programme a f te r f ive
md a decision on i t s future.

s lopes), w ithout d is tu rb ing the birds.
P ub l ic ; ty campaign
The aim here i s to reach every stock farmer w i t h in the fo rag ing range o f the Potberg and Aasvoge1v le i co lon ies . The
p l i g h t o f the v u l tu re s should be made aware to the farmers as well as the advantages which vu l tu res have fo r them. The colonies remain extreme 1y vulnerable to t he threat of poisoned carcasses in the fo ra g in g area to combat in s e c t plagues, e.g. b lo w f ly ;
here, one carcass has the p o te n t i a l to k i l l h a l f the breeding segment o f the Potberg colony. Thus an appeal must be made to
prevent a (a lb e i t accidental) poisoning event.
Conclusion
M o r t a l i t y dur ing c e r ta in stages of the breeding cycle
curren t ly l im i t s recruitment to the reproductive segment of the
co lony. Even in the absence o f o the r m o r t a l i t y fa c to rs te.g. o
po ison ing event , or human d is tu rb a nce ) , sue1’ c r i t i c a l l y ow re c ru i tm e n t le v e ls w i l l e v e n tu a l l y re su i t in ex t inc t ion of the colony. T ? programme o u t l in e d above is designed to a l l e v i a t e
s t re ss a t the p a r t i c u l a r stages tha i. e x h i b i t high m o r ta l i t y
ra tes . Monito ng the e s s e n t ia l demographic aspects of the population w i l l enable an evaluation of the programme a f te r f ive
years and a decision on i t s futv e.

115
A P P E N D I X B
The accounts o f the f o l l o w in g observa t ions th a t I made a t
Potberg are taken from "Predatory a t tempts on Cape Vu l tu res by Black Eagles" (Mundy, Robertson, Komen & O'Connor in orep.).
At 15h45 on 24 J u ly 1981 a subadu l t eagle swooped down
towards a c l i f f and caused 25 ro o s t in g v u l tu re s to f l y o f f , but
(the) two incubating birds remained on t h e i r eggs. At 15h3Q on 14 January 1982 a j u v e n i l e entered the gorge and dived down to
w i t h i n 10 m o f a breeding ledge, e l i c i t i n g hoarse c a l l s and
extended necks from the vul tures and causing 9 of 12 birds to f l y o f f , including a f led g l ing which had fledged a week ea r l ie r . Tne three tha t remained were attending the only act ive nests on that
c l i f f , although a nest l ing of 114 oays was the only nest occupant
at the time. The eagle then dived past another breeding area and e l i c i t e d f u r t h e r hoarse c a l l s , causing fo u r more b i rd s to f l y
o f f .At 08h4Q on 13 December 1981 loud c a l l s anounced the ar r iva l
o f a female eagle on a vu l ture s i te where the vul tures had fa i le d
in t h e i r breeding a t tem p t in J u ly . The s i t e was 4 m above an a d u l t a t tend ing a 116-day o ld n e s t l i n g . W i th in 30 seconds, fu r the r hoarse c a l l s accompanied the landing of the male eagle on
a ledge 3 m higher . At Q8h50 a j u v e n i l e v u l tu r e also landed at
th is las t s i te and aggressively chased the eagle o f f by jabbing a t i t v ig o ro u s ly w i th i t s b i l l and e m i t t i n g a h ig h -p i tche d s tacca to c a l l . The female eagle l e f t three mir ' tes l a t e r and
between 09h02 and l lh55, and together with the male, the eagles e l i c i t e d aggressive responses 11 more times from four attendant parent vultures by soaring past th e i r section c l i f f repeatedly.
At 10hQ3 on 24 December 1981 an eagle dived at f ive vultures perched on c c l i f f , causing a j u v e n i l e to f l y o f f ; no c a l l was
heard. The eagle repeated the behaviour and caused three o f the remain ing fou r a d u l ts to jump o f f . ‘ s i t soared in the v i c i n i t y
o f the c l i f f once again, the l a s t v u l t u re jumped o f f and tne eagle dived a f t e r i t , extend ing i t s legs as i t passed over the
v u l t u r e , but no c o n tac t was made. The v u l tu re w1 need s l i g h t l y , f lexe d i t s wings, thereby inc reas ing i t s speed, and f lew on. At

115
A P P E N D I X B
The accounts o f the f o l l o w in g observa t ions th a t I made a t
Potberg are taken from "Predatory a t tem pts on Cape Vu l tu res by Black Eagles" (Mundy, Robertson, Komen & O'Connor in prep.).
At 15h45 on 24 J u ly 1981 a subadu l t eagle swooped down
towards a c l i f f and caused 25 ro o s t in g v u l tu re s to f l y o f f , bu t (the) two incubating birds remained on t ^ e i r eggs. At 15h30 on 14 January 1982 a j u v e n i l e entered the fo rge and dived down to
w i t h i n 10 m o f a breeding 1 edge, e l i c i t i n g hoarse c a l l s and
extended necks from the vul tures and causing 9 of 12 birds to f l y o f f , including a f ledg l ing which had fledged a week ea r l ie r . The three that remained were attending the only act ive nests on that c l i f f , although a nest l ing o f 114 days was the only nest occupant
at the time. The eagle then dived past another breeding area and e l i c i t e d f u r t h e r hoarse c a l l s , causing fou r mere b i rd s to f l y
o f f .At 08h40 on 13 December 1981 loud c a l l s anounced the arr iva l
o f a female eagle on a vu l tu re s i te where the vul tures had fa i led in t h e i r breeding a t tem p t in Ju ly . The s i t e was 4 m above an a d u l t a t tend ing a 116-day o ld n e s t l i n g . W i th in 30 seconds, fu r the r hoarse c a l l s accompanied the landing o f the male eagle on
a ledge 3 m higher . At 08h50 a j u v e n i l e v u l tu r e a lso landed a t th is las t s i te and aggressively chased the eagle o f f by jabbing a t i t v ig o ro u s ly w i th i t s b i l l and e m i t t i n g a h ig h -p i t ch ed
s tacca to c a l l . The female eagle l e f t three minutes la te r and between 09h02 and l lh55, and together w i th the male, the eagles
e l i c i t e d aggressive responses 11 more times from four attendant parent vul tures by soaring past th e i r section c l i f f repeatedly.
At 10h03 on 24 December 1981 an eagle dived at f ive vultures perched on a c l i f f , causing a j u v e n i l e to f l y o f f ; no c a l l was heard. The eagle repeated the behaviour and caused three of the remain ing fou r a d u l ts to jump o f f ; i t soared in the v i c i n i t y
o f the c l i f f once again, the l a s t v u l t u r e jumped o f f and tne eagle dived a f t e r i t , extending i t s legs as i t passed over the
v u l t u r e , but no con tac t was made. The v u l t u r e winced s l i g h t l y ,
f le xe d i t s wings, thereby inc reas ing i t s speed, and f lew on. At

lO h l l the male eagle landed 1,5 m from a 133-day old nestl ing and 2 5 m from s i m i l a r aged n e s t l i n g on e i t h e r side. A l l three
attendant adults rushed towards the eagle and no is i ly jabbed at
i t , and the eagle was f i n a l l y forced o f f the ledge e ight seconds
a f t e r i t had landed.

1 1 7
APPENDIX C
The fo l low ing is a copy o f the le t t e r , reduced in size, that
was used in the postal survey (chapter three) :
PROVINCIAL AOMINISTRATION 0? THB CAPS 0? 1003 HOPS
DEPARTMENT ON M TVRS a S V I’ O W tS iT A i CONSERVATION
T u lephone : N ap kfl 5«30
Dear S ir
Fat be ( 3e Hoop N ature Reserve)P .u . 3 re iaad o rp3RS3AS3CRPt :? c
I aa employed by the above Departm ent to c a rry out research on -he Cape V u ltu re . As you s ty *r .o * , ape V u ltu re s ;eundergone i d rs a a tis d e c lin e in r.e pest fs» i t c a : • » , e s p e c ia lly in the Cape Province * r.e re there i r e r no i than 6 - breeding p a irs . The is or » ia or ay re o e iro l : totry and understand the fa c to rs ca .s in g t h i : d e c lin e ;V u ltu re s occur only in Southern A fr ic a and are there '> • urc o n s erva tio n p r io r i t y .
: ecu ld l ik e to es tim ate the fo rag in g e re * » v i i t u »sfrom tne Potberg ao u r.ta in . em ch is the or i -a m g v u .tu r ecolony in the a o u tn -*e s te rr . Tape P ro v in ce . ' - 1- 1 th e re fo rebe extrem ely g ra te fu l i f you :ou.d an see: • •.ion - * - o * ,detach and re •- urn the s l ip i n tne p re -id -, e , - d enve lope .»nu.d e l jo ap p rec ia te hear.no from yo. a c o .t any v u . lu res found in d is tre s s on your f a n .
In o rd er fo r he to aane f u l l uoe e l tne in fo rm atio n aup pu ie i by "ou ‘ ~ - 1 . - ■ . .mu « n r - i i n - i r address as re iu eb e1c*.
fo u rs f a i t h f u l ly
In o rd e r f o r he to aaite f u l l uoe ot t .e in .o ra a t.o n e u p p n e u by "on i t la e s s e n t ia l th a t fou supp-y your address <\a r e q u e s te d
- 1 - ; tr. • you n a n t ic ip a t io n of yo -r c o -o p e ra tio n .
A .S . RC3ERT50N
TEA.-I HERS
PLEASE TICK APPROPRIATE AN3*ER NIKS
Hoe o ften : i von :*« v u ltu re s m your firm '' ADDRESS
About once a dayAbout once a week
About once a monthAbout once a yearAbout once per f iv e years
About once per ten yearn
Never
3 . 1 . ASS. V IR AfRIXAAN"

117
APPENDIX C
The fo l low ing 1s a copy of the l e t t e r , reduced in size, that
was used in the postal survey (chapter three) :
PBOVINCUL OMINISTRATlO.'t < - <3 Ca? . 0 ? V S 0 2 HOPS
3S?A R m :iT o? XAT-? s m s e w ta w A i OWSEHVATZOS
' e le e b o n e i 7 sp k e i S9 *.b«r? ( 3a Hoop H; -u re 3 «9t r v » )P.O. 3r»" .aadorp 3RS3AS30SP72iC
3e a r S i r
I «o tao loysd by ‘.he above 3« p e rb a « n t to c a r r y o u t re s e a rc h on the rape V u l t u r e . A3 you aay k r .o* . Cape V u l tu re s have undergone a d ra e a t lC d e c l i n e i n "he pes t fe» i e c s ie a . e e o e c i a l ’. y in the Cape P ro v in c e "h e re th e re are - :« no c re than o- breedir . .- c a . r s . Che h e ; o r a ia o f ay re s e a rc h . to t r y end unders tand tr.e f a c t o r s c a u s in g th u s d e c . m e ; -a - V u l t u r e s occu r o n ly i n S o u th e rn A f r i c a and a re t h e r e : o r e our c o r .a e rv e t ic r . p r i o r i t y .
I wou ld l i k e to e s t im a te the f o r a g in g a rea o f the v u l t u r e s f r o e t r s P o tbe rg m ou n ta in , w h ich i s the o n ly r e a a .a m g v u . t u r e c o lo n y the i c i t n - w e e t e r r . Cape P r o v in c e . - * " u . i the re : o re be e x t re m e ly g r a t e f u l i f yo . cou ld a is w e r the q u e s t ion be.ow, d e ta c n and r e t u r n the s l i p i n tne p r e - e id r e s s e l e n ve lo pe , wou ld a ls o a p p re c ia te h e a r in g from : o a c o . t any v u . t u r e s found in d i s t r e s s on yo u r f a r n .
In o rd e r f o r sa to aaite f u l l use of the in f o r m a t i o n s u p p l ie d by you i t i s » i s e n t i a l t h a t you su pp ly , * r address as req u e s te d be lo w . ' Ah', jc you i n a n t i c i p a t i o n o f 1 ; r c o - o p e r a t i o n .
f o u r s f a i t h f u . - y
A . S . RCBSRTSOM
___________________ TSAR HERS --------------------
P L E A S E T IC K A P P R O P R IA T E A N S *E S .
How o f t e n d o y o u s e e v u l t u r e s o n y o u r f a r m " A P 3 S S 3 3
About once a day .......About once a week .......About once a ronth About once a year About once per f i v e years About once per ten y e a - '
lever
\ . ASS. v t> AfRIKAAN-

118
A P P E N D I X D
The fo l low ing thicknesses of shel l fragments were measured
d i r e c t l y , by the Western Foundation o f Ver teb ra te Zoo.ogy,
C a l i fo rn ia , using a Federal Bench comparator thickness guage (L.
K i f f in 1 i t t . ). Ten measurements were made o f each sample and, where known, the fa te o f the sample is i n d ic a te d (hatched^ ;
n e s t l i n g subsequently f 1 edged). The o r i g i n a l p o s i t i o n o f the fragment, where known, is also indicated (xe : equatorial ; xP ;
p o la r ) .
Nest Date T h ick n e ss
membrane p re s e n t
(mm)
membrane a bsen t
Fate C o l l e c t o r
10 1974 0,671 0 ,626 add led CDNEC
40 1974 0,564 • add led CDNEC
3 1975 0,668® . 0 ,6 1 0 p 0,626 a dd led CDNEC
4 1975 0,708® add led CDNEC
42 1975 0 ,641® . 0 ,661p 0,562 embryo dead CDNEC
55 1975 0 ,760 0 ,627p ; 0 ,6 4 9 P add led CDNEC
13 1981 - 0,649 add led ASR
16 1981 - 0 ,622 h a tched ' ASR
29 1981 0 ,759 0,661 a d d e d ASR
63 1981 - 0 ,680 h a tc h e d 1 ASR
17 1982 0,852®-,0,360p add led ASR
13 AW 1982 0 ,6 4 6 ® ;0 ,5 8 5 p add led ASR

119*
A P P E N D I X E
The f o l l o w in g is a l i s t o f poisons used in the farmlands
s u r r o u n d in g the c o lo n y , in o rd e r1 o f amounts so l d a t the
Bredasdorp-Napier Co-operative (S. Lambrecht. pers. comm.). From d iscuss ions w i t h farmers in the area, i t would appear th a t
"Dazzel" is , or ce r ta in ly was, tne most frequently used poison to
-ombat Blow f l i e s .
Brard Main component Manufacturer
1. Sheep dips
Super Golden Fleece
Sumifleece
Dazzel NF
Sama dip
Topclip purple
( Pr im ate*, Parecto)
Oiazinon
Zip dip
Y-8HC Dip
Oiazinon, A n itraz Coopers
Shell
Oiazinon Agricura
Phoxim 3ayer SA
Oiazinon, Rotenone Cifca Geigy
Oiazinon Coopers
Triazophos Hoechst
Lindane, Trichlorophenol Agricura
2. Sheep b lo w -f ly s tr ide
Vetraz in SP 50
Dazzel
PAB
In je c t
Karbadip
Nexa je t
Bromex OFF
Vetrazin
Oiazinon
Oichlofenthion
Fenthion ethyl
Carbaryo1
3romophos-ethy1
Oichlofenthion
Ciba Geigy
Agricura
Mil borrow
Bayer SA
Agricura
Hoechst
Coopers

120
REFERENCES
ACOCKS, J.P.H. 1975. Veld types of South A f r i c a . Mem. Bot. Surv.
S. A fr . No. 40, 128 pp.AMADON, D. 1964. The evo lut ion o f low reproductive rates in birds
Evolution 18: 105-110.ANON. 1982. Condor rescuers get egg on t h e i r faces. New Sc ient is t
18 March: 699.ANTHONY, A.J. 1976. The Lappet-facec Vultures of the Gonarezhou.
Bokmakierie 28: 54-57.ASHMOLE, N.P. & TOVAR, H.S. 1968. Prolonged paren ta l care in
Royal Terns and other binds. Auk 85: 90-100.BALLOU, J. & RALLS, K. 1982. Inbreeding and juven i le m or ta l i ty in
small po pu la t ions o f ungulates : a d e ta i le d ana lys is . B io l .
Conserv. 24: 239-272.BERRUTI, A. 1981. D isp lays o f the sooty a lba t rosses Phoebefr ia
fusca and P. nalpebrata. Ostr ich 52: 98-103.BOSHOFF, A.F. 1981. Notes on two Cape V u l tu re co lon ies in t-'e
southwestern Cape Province, South Afr ica. Vulture News 5: 3-
10.BOSHOFF, A.F. & CURRIE, M.H. 1981. Notes on the Cape V u l tu re
colony at Potberg, Bredasdorp. Ostr ich 52: 1-8.BOSHOFF, A.F. & VERNON, C.J. 1979. Supplementary notes on the
Cape Vulture in the Cape Province. Cape Town: Cape
Dept. Nature Env. Cons.BOSHOFF, A.F. & VERNON, C.J. 1980. The pa s t and p re s e n t
d i • t r ib u t io n and status o f the Cape Vulture in the Cape
Province. Ostr ich 51: 230-250.BOSHOFF, A.F., VERNON. C.J. & BROOKE, R.K. 1983. H i s t o r i c a l a t la s
o f the d i u r n a l r a p t o r s o f the Cape P ro v in c e iA v e s :
Fa 1 con i formes). Ann. Cape Prov. Mus. ina t . i s t . ) 14: 173-
297.BOWEN, P. 1970. Some observations of the Cape Vulture. Afr. Wild
L i fe 24: 125-128.BROOKE, R.K. ( in press). The ra re id vu lne rab le b i rd s of South
A f r i c a . S. A f r . Natn. Sci . Programmes. Rep.BROWN, L.H. 1966. Observat ions on some Kenya eagles. I b i s 108:
531-572.

121
BROWN, L.H. & AilADON, D. 19b8. Eagles, hawks and fa lcons o f the
world. Felthan : Country L i fe .BROWN, L.H & CADE, I .J . 1972. Age c lasses and popu la t ion
dynamics o f the Bateleu^ and Afr ican Fish Eagle. Ostr ich 43:
1-16.BRYANT, D.M. & GARDINER, A. 1979. Energe t ics o f growth in House
Mart ins Delichon urbica. J. Zool. Lond. 189: 275-304.
BURGER, J. 19bl. On becoming independant in Herr ing G u l ls :parent-young c o n f l i c t . Amer. Nat. 117: 444-456.
CADE, T.J. 198?. The Falcons o f the wor ld . I thaca , N.Y. : Cornel l
Univers i ty Press.DAY, J., SIEGFRIED, W.R., LOUW, G.N. & JARMAN, M.L. (eds). 1979.
Fynbos ecology : a p r e l im in a r y syn thes is . S. Afr . Nat. S c i .
Progr. Reo. 40.DIAMOND, A.W. 1975. Biology and behaviour of fr igatebirds Fregata
spp. on Aldabra A to l l . Ib is 117: 302-323.DRENT, R.H. & DAAN, S. 1980. The prudent parent : energe t ic
adjustments in avian breeding. Ardea 68: 225-252.DUNN, E.K. 1972. Age ar 3 f ish ing a b i l i t y of Sandwich terns. Ibis
114: 360-366.du PLESSIS, S.F. 1969. The past and present geographical
d i s t r i b u t i o n o f the Peri ssodac ty1 a and A r t i od a c ty1 a in
southern Afr ica. M.Sc. thesis, Univ. of Pretoria.
ERICKSON, C.J. 4 ZtNONE, P.G. 1976. Courtship dif ferences in male Ring Doves : avoidance oi cuckoldry ? Science 192: 1353-1354.
ESTEP, D.Q. & BRUCE, K.E.M. 1981. The concept o f rape in non- humans : a c r i t ique . Anim. Beh. 29: 1272-1273.
FALL IS, A.M., JACOBSON, R.L. & RAYBOLD, J.N. 1973. Haematozoa in d o m e s t i c c h ic k e n s and Guinea Fowl in Tanzan ia and t ra nsm iss io n o f Leucocytozoon i eavei and Leucocytozoon
schoutedeni. J. Protczool. 20: 438-442.FANNIN, A S WEBB, D. 1975. Notes on the breeding of the Crowned
Eagle. Honeyguide 82: 36.FEARE, C.J. 1972. The seasonal p a t te rn of feeding in the Rook
( Corvis f r u g i 1egus) in norths Scot land. Proc. 15 th In t .
orn. Congr. 643.FEARE, C.J. 1975. P o s t - f le d g in g care in Crested and Sooty Terns.
Condor 77: 368-370.

122
FISHER, J. 1952. The Fulmar. London: Co l l ins .FITCH, H.S., SWENSON, F. & TILLOTSOf D.F. 1946. Behaviour and
food habits of the Red-tai led Hawk. Condor 48: 205-237.FOWLER, C.W. & COPE, J.B. 1964. Notes on the Harpy Eagle in
B r i t i s h Guiana, Auk 81: 257-273.GESSAMAN, J.A. 1973. Ecological energetics o f homeotherms. Logan,
Utah: Utah State Universi ty Press Monogr. Series 20.GREEN, A.A. 1977. New nesting records fo r Puppell 's Grif fon Gyps
rueppel 1 i i in East Afr ica. Bul l . B r i t . Orn ithol . Club 97: 9.
GREEN, R. 1976. Breeding behaviour o f Ospreys Pandion hal iaetus
in Scotland. Ib is 118: 475-490.GREINER, E.C. & MUNDY, P.J. 1979. Haematozoa from southern
Afr ican vultures, wi th a descript ion of Haemoproteus janovyi
sp. n. J. P a r a s i to l . 65: 147-153.HEYDORN, A.E.F. & T1NLEY, K.L. 1980. E s tu a r ies o f the Cape. Part
1. Synopsis of the Cape coast-natural features, dynamics and
u t i l i s a t i o n . CSIR Res. Rep. 380.HINDE, R.A. 1970. Animal behaviour. 2nd ed. New York: McGraw-
H i l l .HOUSTON, D.C. 1972. The ecology of Serengeti v u l tu re s . 0. P h i l ,
thesis, University of Oxford.HOUSTON, D.C. 1974a. Food searching in g r i f f o n v u l tu re s . E. A f r .
W i ld l . J. 12: 63-77.HOUSTON, D.C. 1974b. M o r t a l i t y o f the Cape Vu l tu re . O s t r ich 45:
57-62.HOUSTON, D.C. 1974c. The role of g r i f fon vul tures Gyps ofr i-anus
and Gyps ruppe 1111 ( s i c ) as scavengers. J. "oo i . Lond. 172:
35-46.HOUSTON, D.C.1975. Eco log ica l i s o l a t i o n o f A f r i c a n scavenging
b i rd s , Ardea 63: 55-64.HOUSTON, D.C. 1976. Breeding o f the White-backed and Ruppell 's
G r i f f o n V u l tu re s , Gyps a f r i c a n us and Gyps rueppel 1 i i . Ib i s
118: 14-40.HOUSTON, D.C. 1978. The e f f e c t food q u a l i t y on breeding
s t ra teg y in g r i f f o n v u l tu re s (Gyps spp.). J. Zool. Lond. 186:
175-184.

123
■ ,Ni ). , N . n, .dd j /Ld tv ■ I f scavengers, pp. 263-ZWo
In: a .R.E. nc i a i r & M. Norton-Gr i f f i ths teds). Serenget i . lynam ‘ ,n o system, hicago: U n iv e rs i t y o f Chicago
Pres .OWARO. I '>. » e s t im a t in g re p ro du c t ive >- .cess in natura l
populations. Am. Nat. 114: 221-231.N T _ g.L. & HUNT, M.W. 1976. G u l l c n i c k s u r v i v a l : the
s ig n i f i c a n c e o f growth ra tes , t im in g o f breeding and
t e r r i t o r y size. Ecology 57: 62- ;5.ACKSON, .A. 1975. Regurgitat ive feeding of young Black Vultures
in December. Auk 92: 802-B03.ARVIS, SIEGFRIED, W.R. & CURRIE, M.H. 1974. Conservation
o f the Cape Vu l tu re in the Cape Province. J. sth. A f r .
^ i I d ! . Mgmt Ass. 4: 29-34. uHNSON, S.J. 1973. P o s t - f le d g in g a c t i v i t y o f the Red-ta i leu
Hawk. Raptor Res. 43-48.• OFORD, C.B. 1953. The C a l i f o r n i a Condor. Nat. Audubon Res. Rep.
4: 154.. o.MEN, J. 1983. The energy demands on parent Cape Vultures at the
Skeerpoort breeding co lony (a b s t ra c t ) . Second Symposium on
A f r i c a n predatory b i rd s : Programme and Abs t rac ts . Durban.
Natal Bird Club.- RUUK, H. 196.. Compet i t ion f o r food between v u l tu re s in East
Afr ica. Ardea 55: 171-193. cACK, D. 1954. The natural regula t ion o f animal numbers. Oxford :
in ive rs i ty Press.. ACk, I/, I9b8. Eco log ica l adap ta t ions fo r breeding in b i rds .
_ondon: Methuen.AKHANI, K.H. & NEWTON, I. 1983. E s t im a t in g a g e -s p e c i f i c b i rd
s u rv iv a l ra tes - can i t oe done ? J. An i m. Ecol. ^2. 83-91. .EUGER, J.A. 1974. co lour-r ings for vultures. Safring News 3: 23-
28.EDGE h , .A. 19 79. A f r ica n in se c t l i f e , rev. edn. Cape Town:
3tru1k.rDGER, .A. '8( . -u lu res poisoned in Capri v i . V u l tu re News
3:15.

L0CK1E, J.D., RATCL1FFE, D . A . & B A L H A R R Y , R. 19b< Breeding success and o rg a n o -c h lo r i ne residues in Golden
West Scotland. J. appl. E co l . 6: 361-389.LOMAN, J. & TAMM, S. 1980. Do roos ts serve a, in fo rm a t ion
centres" fo r crows and ravens ? A m. Nat. 115: 385-305.
LOU W, D.F.J. 1970. Prevent ion o f lamb m o r t a l i t y . Supp I . t The
Wool Grower 24: i - i v .MARKUS, M.B. 1974. Arth ropod-borne disease as a poss ib le factor
l i m i t i n g the d i s t r i b u t i o n o f b i rd s . J. P a ras i t . 4: 609-
612.MARTIN, R.J. & MARTIN, E. 1983. Atlassing in the eastern squares.
Promerops 157: 1.McLACHLAN, G.R. & LIVERSIDGE, R. 1978. Roberts Dirds o f >outr
A fr ica , 4th ed i t ion . Cape Town: John Voelcker B,ro Book irun<
MERTZ, D.B. 1971. The mathematical demography of the ' . a l i f o r
Condor population. Am. Nat. 105: 437-453.MILLER, A.H., McMILLAN, 1.1. & McMlLLAN, E. 1965. The . ’ ent
s ta tus and w e l fa re o f the C a l i f o r n i a Condor. Mat. 1 idup-
Soc., Res. Rep. 6.MONNIG, H.O. & YELDMAN, F.J. 1976. Handbook n ,tock useases.
2nd edn. Cape Town: Tafelberg.MORRIS, A. & MUNDY, P. 1981. In c id e n ts o f po isoning v u l tu re s in
Zimbabwe. Zimb. Sci. News 15: 127-129.MUNDY, P.O. 1982. The comparative b io lo gy o f southern A f n c a r
vul tures. Johannesburg: VulCure Study Group.MUNDY, P.J. & COOK, A.W. 1975. Hatching and rearing of two M i -
by the Hooded Vulture. Ostr ich 46: 45-50.MUNDY, P.J. & LEDGER, J.A. 1975. Notes on the ape , "
Honeyguide 83: 22-28.MUNDY, P.J. & LEDGER, J.A. 1976. Gr i f fon vul tures, carnivor u
bones. S. A f r . J. Sci. 72: 106-110.MUNDY, P.J., GRANT, K.I. , TANNOCK, J. S WESSEi b, . . 198; .
Pesticide residues and eggshell thickness of g r i f fo n vu i - e
eggs in southern Afr ica. J. Wiid l. Manage. 4b: 69- 3.MUNDY, P.J., MORRIS, A. & HAXEN, C.M. 1983. The p rop o r t ion of a"
impala e d ib le to v u l tu re s . A f r . J. Ecol. 21: 75 - 'b.

125
MUNOY, P.J. & KOMEN, J. 1983. Why do we weigh baby v u l tu re s ? (a b s t ra c t ) . Second Symposium on A f r i c a n Predatory B i rds :
Programme and Abstracts. Durban: Natal Bird Club.MYERS, N. 1979. The s ink in g ark. A new look a t the problem o f
disappearing species. Oxford: Pergamon Press.NELSON, J . B . 1976. The breeding b io lo g y o f F r ig a te b i rd s . L iv in g
Bird 14: 113-155.NELSON, J.B. 1978. The Sul idae. Aberdeen: Oxford U n iv e rs i t y
Press.NEWMAN, K. 1983. B i rds o f sui _rn A f r i c a . Johannesburg:
MacMi11 an.NEWTON, I. 1979. Popu la t ion ecology o f ra p to rs . Berk.hamsted:
Poyser.NEWTON. I. 1980. The ro le of food in l im i t i n g b i rd numbers. Ardea
68: 11-30.NORTON-GRIFFITHS, M. 1969. The o rg a n is a t io n , c o n t ro l and
development o f pa ren ta l f e e d in g in the O y s te r c a t c h e r
(Haematopus ostralegus). Benaviour 34: 55-114.
O'BRIEN, S.J., WILDT, D.E., GOLDMAN, 0., MERRIL, C.R. & BUSH, M. 1983. The Cheetah is depauperate in genet ic v a r ia t i o n .
Science 221: 459-462.ORIANS, G.H. 1969. Age and hunt ing success in the Brown Pel ican,
Pelcanus occidental i s . Anim. Behav. 17: 316-319.
PATERSON, H.E.H. 1982. Darwin and the o r i g i n o f species. S. A fr .
J. Sci. 78: 272-275.PEAKALL, D.B. & KEMP, A.C. 1976. Organoch 1 o r in e residues in
herons and raptors in the Transvaal. Ostr ich 47: 139-141.
PENNYCUICK, C.J. 1972. Soaring behaviour and performance of some East Afr ican b i rd : , observed from a motor-gl ider . Ib is 114:
178-218.PENNYCUICK, C.J. 1976. Breeding o f the Lappet- faced and White-
headed Vultures (Torgos t rachel io tus Forster and Trigonoceps
o c c ip i ta l is Burche 11) on the Serengeti Plains. E. Afr. Wildl.
J. 15: 67-84.PETRIDES, G.A. 1959. Competition fo r food between f ive species of
East Afr ican vultures. Auk 76: 104-106.

126
PIPER, S.E., MUNDY, P.J. & LEDGER, J.A. 1981. Est imates of survival in the Cape Vulture Gyps coprotheres. J. Anim. Ecol. 50: 815-825.
POWER, H.W. 1980. On bi rd cucko 1 dry and numan a d u l te ry . Am. Mat.
116: 705-709.RICKLEFS, R.E. 1977. On the evolut ion of reproductive strategies
in birds : reproductive e f f o r t . Am. Nat. I l l : 453-478. ROBERTSON, A. 1983a. The fo rag ing area o f the Potberg Cape
V u l t u r e c o lo n y ( a b s t r a c t ) . B i r d s and Man Symposium: Programme 8> Abs t rac ts . Johannesburg : Wi twa te rs rand Bird
Club.ROBERTSON, A. 1983b. Known-age Cape Vu l tu res breeding in the
w i 1d. Ostr ich 54: 179.
ROUGHGARDEN, J. 1979. Theory o f p o p u l a t i o n g e n e t i c s and e v o lu t io n a ry ecology : an in t r o d u c t io n . London: C o l l i e r
MacMi 11an Publ ishers.ROWE, E.G. 1947. The breeding biology of Aguila verreauxi Lesson.
Ib is 89: 576-606.
SCHEFLER, W.C. 1969. S t a t i s t i c s f o r the B io lo g ic a l Sciences.London: Addison-Wesley Publishing Company.
SCHOENER, T.W. 1969. Sizes o f feeding t e r r i t o r i e s amoung b i rds .Ecology 49: 123-141.
SCHULZE, B.R. 1965. C l im ate o f South A f r i c a . P a r t 8. General Survey. Weather Bureau Publ. 28.
SHERROD, S.K. 1983. Behavior o f f l e d g l i n g Peregr ines. New York:
The Peregrine Fund, Inc.SIEGFRIED, W.R., FROST, P.G.H., COOPER, J. & KEMP, A.C. 1976.
South A f r i c a n Red Data Book - Aves. S. A f r . Natn. Sci.
Programmes Rep. No. 7.SKEAD, C.J. 1980. H i s t o r i c a l mammal Inc idence in the Cape
Province. Vol. 1: The western and no r the rn Cape. Cape Town: Cape Department o f Nature and Environmental Conservation.
SKUTCH, A.F. 1976. P a re n t b i r d s and t h e i r young. A u s t in :
Univers ity of Texas Press.SNYDER, N.F. & WILEY, J.W. 1976. Sc I s ize dimorphism in hawks
and owls of North America. Ornithological Monographs 20.

/
STEYN, P. 1973. Eagle Days. London: Purnel l .
STEYN, P. 1982. B i rds o f prey o f southern A f r i c a Cape Town: David Phi 1 ip.
STONEHOUSE, B. 5 STUNEHOUSE, S. 1963. The F r i g a t - B i rd Fregata aquila of Ascension Island. Ib is 103: 409-422.
AR130TON, W.R. & ALLAN, D.G. in p re ss . The s t a t u s and
conservation o f birds o f prey in the Transvaal. Transv. Mus. monogr.
■ ERRASSE, J.-F. 1977. Matur i te s e x u :1le du Vautour fauve,
premieres donnees obtenues dans la nature. Oiseau 47: 214- 218.
RIVERS, R.L. 1974. Parent-offspr ing .onfl i c t . Am. Zool. 14: 249- 264.
VERNON, C.J., PIPER, S.E. & SCHULTZ, U.M. 1982. The breeding
success of the Cape Vultures at Col lywobbles, Transkei, 1981. Vulture News 8: 26-29.
VERNON, v.J. & ROBERTSON, A. 1983. Factors regulat ing breeding in the Cape Vulture. Vulture News 7: 10-13.
WARD, P. & ZAHAVI, A. 1973. The importance o f cer ta in assemblages
of b i rd s as " i n fo r m a t io n - c e n t r e s " f o r fo o d - f in d in g . Ib is 115: 517-534.
WILLIAMS, G.C. 1966. Adaptation and Natural Selection: a c r i t ique
o f some c u r r e n t e v o lu t io n a ry thought. P r ince ton : Pr inceton U n iv e rs i t y Press.
WILSON, E.O. 1 9 7 5. Sociob io 1 ogy. The new syn thes is , ambridge (Mass.): Harvard University Press.
WYNNE-EDWARDS, V.C. 1955. Low re p ro d uc t ive ra tes in b i rd s , especia l ly sea-birds. Acta Int. Ornithol. Congr. 11: 540-547.
YQM-TOV, f . 19-4. The e f f e c t o f food and p reda t ion on breeding
density and success, c lutch size and laying date of the Crow Corvus corone L. J. Anim. Ecol. 43: 479-498.


Author Robertson A SName of thesis The Feeding Ecology and breeding biology of a Cape Vulture colony in the Southwestern Cape Province 1983
PUBLISHER:University of the Witwatersrand, Johannesburg ©2013
LEGAL NOTICES:
Copyright Notice: All materials on the Un i ve r s i t y of t he W i t w a t e r s r an d , Johannesbu r g L i b r a r y website are protected by South African copyright law and may not be distributed, transmitted, displayed, or otherwise published in any format, without the prior written permission of the copyright owner.
Disclaimer and Terms of Use: Provided that you maintain all copyright and other notices contained therein, you may download material (one machine readable copy and one print copy per page) for your personal and/or educational non-commercial use only.
The University o f the W itwatersrand, Johannesburg, is not responsible for any errors or omissions and excludes any and all liability for any errors in or omissions from the information on the Library website.