the effect of ph and temperature on the kinetic parameters of
TRANSCRIPT

THE JOURNAL OF EIOLOGICAL CHEMIGTRY Vol. 243, No. 7, Issue of Aprii 10,pl.r. 1401-1414, lSF8
Printed in U.S.A.
The Effect of pH and Temperature on the Kinetic Parameters of Phosphoglucose I somerase
PARTICIPATION OF HISTIDINE AND LYSINE IN A PROPOSED DUAL FUNCTIOn’ MECHANISM*
(Received for publication, June 8, 1967)
JOHN E. D. DYSOX AND ERNST A. KOLTMANN
From the Department oj Biochemistry, University of California, RiveTside, Calijornia Q&iOi?
SUMMARY
Phosphoglucose isomerase (D-glucose-6-phosphate ketol- isomerase, EC 5.3.1.9) from rabbit skeletal muscle has been subjected to a detailed kinetic study of the effect of pH and temperature on the Michaelis constants, the maximal velocities, the dissociation constant for the competitive inhibitor 6-phosphogluconate, and the equilibrium constant of the reaction.
A coupled pH-stat assay was established as being suitable for measurement of initial velocities in the forward direction between pH 6.0 and 10.0; analogously, the validity of a coupled spectrophotometric assay of the reverse reaction was demonstrated for the range of pH 6.0 to 9.5.
Graphical and computer analysis of K, and Vm,, data, obtained with these assays as a function of pH, suggest the participation in isomerase catalysis of two ionizable groups in the enzyme with average pK values of 6.75 and 9.3, at 30”. From the change in pK with temperature, values of 7,700 and 16,000 cal per mole, respectively, have been cal- culated for the heats of ionization for these two groups. These data are considered to be consistent with the involve- ment of both an imidazole group of histidine and an e-amino group of lysine.
Analysis of Ki data for 6-phosphogluconate, obtained as a function of pH for the forward and the reverse reaction, showed no pK at alkaline pH; this has been interpreted as suggesting that the e-amino group of lysine is not involved in the binding of 6-phosphogluconate to the enzyme.
On the basis of this evidence, a dual function mechanism is proposed in which the protonated c-amino nitrogen first
* This is the seventh paper in the series dealing with studies on phosphohexose isomer&es. The work was supported in part by rrants from the United States Public Health Service IAM 07203) and the Xation2tl Science Foundation (GB 2236) and by Cancer Research Funds of the University of California. Part of the work described in this paper is from a dissertation submit,ted by John E. I). Dyson to the Graduate Division of the University of California, Riverside, in partial fulfillment of the requirements for the degree of Doctor of Philosophy.
catalyzes the opening of the hexose ring structure to yield the straight chain form of hexose B-phosphate, which then undergoes isomerization via formation of an enediol inter- mediate, with the nonprotonated nitrogen of the imidazole group participating as a base catalyst in this second step.
One of the factors fundamental to the understanding of an enzyme mechanism is a knowledge of the amino acid residues involved in the catalytic process, since this is a prerequisite for the construction of reaction mechanisms involving the enzyme molecule. Essentially, two main routes are available to obtain such information. First, a residue can be chemically identified, if its modification can be directly correlated to a corresponding loss in enzyme activity, or if the formation of a covalent link between the substrate and t’he amino acid residue is possible, permitting the isolation of a stable complex. Second, a careful kinetic study of the catalyzed reaction may be made under con- ditions which affect the ionization of one or more participating amino acid residues. From such studies, the pK values for chemical groups involved in the catalytic process can be ob- tained, and the corresponding amino acid residues which partic- ipate in this process may thus be identified.
Some effort has been expended in this laboratory in applying both of these approaches to phosphoglucose isomerase (D-ghCOSe-
6-phosphate ketol-isomerase, EC 5.3.1.9), an enzyme that is strategically located in the glycolytic pathway a.nd, in addition, is of considerable interest with respect to its mechanism of action. Deuterium exchange experiments conducted with partially purified phosphoglucose isomerase preparations from rabbit muscle, human erythrocytes, and bakers’ yeast led Rose and O’Connell to suggest that a base with no exchangeable hydrogens, i.e. a carboxylate group or the nonprotonated nitrogen of an imidazole group, is involved in catalyzing the interconversion of glucose B-phosphate and fructose B-phosphate (1). Subse- quently, Hines and Wolfe reported on studies of the pH-depend-
1401
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1402 Kinetics and Mechanism of Phosphoglucose Isomeruse Vol. 243, No. 7
ent kinetics of phosphoglucose isomerase highly purified from bovine mammary gland (2). These authors derived pK values of 5.96 and 9.11 from rate measurements at 30” and assigned the lower value of the two to an imidazole group, thus supporting the earlier proposal of Rose and O’Connell (I).
Protein modification studies undertaken after the enzyme became available in crystalline form from rabbit muscle (3) are in agreement with the concept of histidine as a critical amino acid residue (4) in phosphoghxose isomerase. The present study was initiat,ed as a collateral to the chemical approach; it utilizes, in addition to the pH dependence of the kinetic parameters, t’he effect of temperature on the pK values and the t,hus derived heats of ionization as a means to identify chemical groups partici- pating in cat,alysis. Preliminary reports on part of this work have been presented (5, 6).
The validity of identifying “catalytic” amino acid residues on the basis of kinetically derived pK values has been questioned. It has been pointed out that some uncertainty may be int’roduced by the influence of equilibrium steps preceding or following the rate-limiting step (e.g. References 7-9). In such instances, the measured pK values may differ somewhat from the true pK values. It seems unlikely, however, that the differences intro- duced would be sufficienbly large in the present instance t,o affect an interpretation with respect to the nature of the groups involved. Moreover, in the present case a considerable amount of evidence other than kinetic supports the participation of the proposed amino acid residues. Criticism has also been directed toward systems in which two pK values are close together, thereby facilitating alternative ionization pathways (IO), a situation which does not prevail for phosphoglucose isomerase.
It must furthermore be considered (1 l-13) that an ionization which is observed to alt,er either V,,,, or K, or both may do so by altering the conformation of the active sit’e and may not involve an amino acid side chain directly concerned wit,h the catalytic process. If t,his were the case, any chemical modifica- tion of such an ionizing group might lead to a change in conforma- tion wit#h ensuing loss of activity, and doubt may be cast on the interpretation of almost any method being used to identify catalytically essential amino acid residues.
In discussions related to the problem of interpreting a kinet- ically observed ionization as an effect on either conformation or catalysis, one argument, however, appears never to have been raised, and yet seems obvious from most studies on pa-dependent kinetics. In profiles of V,,,,, and of V,,,,,/R, with respect to pH, there are normally only two inflection points attributable to ionizations that affect, enzyme activity or affinity for substrat,c. I f one were to account for t’hese in terms of conformation changes, it would seem necessary to assume, for t,he groups t,hat are responsible for binding of substrate to enzyme, either that (a) they are all nonionizing, (b) their ionization has no effect on t’he affinity of enzyme for substrate, or (c) their ionizat,ion is outside of t,he pH range in which kinetic studies are being made. If these premises do not apply, it, is reasonable to assume that distinct pK values should be observed corresponding to both conformation changes and to groups directly participating in substrate binding. Since more than two pK values are rarely found within the stability range of most enzymes, the smallest number of assumptions appears to be associated with the inter- pret,ation identifying the observed pK values with the chemical groups involved in the catalytic process.
Moreover, as will be reported in a future communicat~ion, con-
formational changes in the phosphoglucose isomerase molecule have been studied as a function of pH, with the aid of difference spectrol?hotometry. No spectral changes have been found wit.hin the pH range of 6.5 to 10, and therefore no indication is given of alterations either in the environment of aromatic resi- dues or in the peptide backbone structure. Thlls, the failure to observe such spectral changes in the l)H range of the kineti- cally operable pK values also argues against their correlation with conformational cha,ngea.
EXPERIJIESTAL PROCCUURI;B
Enzymes-Phosplroglucose isomerase was prepared from rabbit skeletal muscle according t,o the method previously described (3, 14). Preparat,ions were stored at O-2” as crystalline sus- pensions in 2.2 M ammonium sulfate, pH 7.0. Under thesr con-
ditions the enzyme may be kept several mont,hs without de- tectable loss of activity. Glucose 6-phosphate dehydrogenase (n-glucose-6-l)hosl)hate:~ADP oxidorcductasr, EC 1.1.1 .49)
from yeast (specific activity, 240 units per mg at pH 8.5 and 30”) and 6-phosphogluconate dehydrogenase (6.l)hosl’ho-n-glucoiiatc: NXDP oxidoreductase (decarboxylating), EC 1.1. I .44) from sheep liver or yeast (specific activity, 1.5 and 15 units per mg, respect’ively, at pH 7.0 and 30”) were purchased from Boehringcr- Mannheim. Phosphofructokinase (ATP : n-fructose-B-phosphate I-phosphot,ransferase, EC 2.7.1.11) was prepared as described elsewhere (15). The preparations used had specific activities of 140 to 160 units per mg at pH 8.5 and 30”, and were free of de- tectable phosphoglucose isomerase and ATPase activity.
Substrates and Reagents-The disodium salt of glucose B- phosphate (95 to lOO(i;, pure), the t~ric~yclohesylammorriuln salt or the trisodium salt of 6-phosl,hogluconate (98 to lOO(,;; pure), t,he trisodiurn salt (90 c< pure) or the tetracyclohexylammonium salt (98 to lOOc/ pure) of fructose 1,6-diphosphatc, the di- sodium salt of adenosine 5’-triphosphate (98 t,o 100’5; pure), and the sodium salt of nicotinamide adenine dinucleotidc phos- phate (97 to 100% pure) were purchased from lioehringer- Mannheim or Sigma. All solutions of substrates and in- hibitors were st’andardized by enzymatic assay, assuming a molecular absorptivity coefficient of 6.22 x 10” 31-l cnl-’ for NADPH (16). The values obtained were in agreement with t,he stated purities.
Commercially obtainable preparatiollu of l’ruc?osc G-phosphate were found t,o contain from 2 to 4~; of glucose &phosphat~r as cont,amination. This xas removed as follows. l’hc barium salt (Uoehringcr-$lannheim) of fruct,ost 6-phoxphat,e (5 g) was dissolved in glass-distilled water, and the bariurii was removed with an excess of Dowex 50-H+ (50 g, wet \&ght,). The rrsulting solution was dccolorized n-ith acid-washed Norit charcoal and adjusted t,o pH 9.2 \T-ith 4 s ?U’H&H. The colorless solution was applied to a colu~nn (40 x 2 cm) of Dower l-SOL”- (200 to 400 mesh), which was then washed with 2 litrrq of glass-distilled water. The column was thereafter eluted with a solution con. taining 2.5 m&f sodium borate, 2.5 mhf NH&XI, and 5 mx am- monium sulfatr, unt,il no further glucose (i-phosphate was found in the effluent. The fruct,ose 6-phosphate was finally elutcd with 500 ml of 0.1 x ammonium sulfate adjusted to pH 3.0 wit,h H2S04. The solution containing t,he fructose G-phosphate was treated with t,hc calculated amount of barium acct,atc to pre- cipitate the sulfate present, this being followed by lyophilization
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of April 10, 1968 J. E. D. Dyson and E. A. Noltynann 1403
to remove water and ammonium acetate. The dry product was dissolved in 200 ml of 0.01 N acetic acid and applied to 100 g (wet weight) of Bio-Rad AGI-X8 resin (100 to 200 mesh, acetate form). At this acetate concentration, the borate passes through the resin but the fructose 6-phosphate is retained. The fructose 6-phosphate was then eluted with 250 ml of 0.5 M ammonium acetate, adjusted to pH 3.0 with acet’ic acid. The solution was lyophilized to dryness to remove mater and ammonium acetate. The dry powder was dissolved in glass-distilled water and treated with a small quantity of Dowex 50 to ensure that all cations had been removed; the- solution was decolorized with acid-washed Norit charcoal and finally adjusted to pH 6.8 with 1 r\’ KOH. The yield was about SO%, based on the calculated original weight of free fructose B-phosphate with a glucose B-phosphate contami- nation of 0.3 to 0.6% remaining.
Tris (hydroxymethyl)aminomethane, A grade (heavy metals
less than 1 ppm), and imidazole, A grade, m.p. 88.9”, chromato- graphically homogeneous, were purchased from Calbiochem; 2- aminoethanol (ethanolamine), m.p. g-10”, was obtained from Matheson, Coleman, and Bell. All other chemicals were of analytical reagent grade and met American Chemical Society specifications. Solutions were prepared in deionized, doubly distilled water.
Methods
Forward Reaction (Glucose 6-Phosphate 3 Fructose 6-Phosphate) -Reaction velocities in the forward direction were determined with the recording pH-stat assay previously described (17). The standard reaction mixture contained the following final concen- trations in a total volume of 3.0 ml: ATP, 5 mnf; MgSO4, 5 m&I; and KCl, approximately 80 mM, depending on the concentration required to bring ‘the ionic strength to a final value of 0.12. Phosphoglucose isomerase aliquots of 25 to 50 ~1 were used, equivalent to 0.1 to 0.4 unit of enzyme, the larger amounts of en- zyme being required at the extremes of the pH range examined. All phosphoglucose isomcrase dilut,ions were made in glass- distilled water, since sufficient buffer was transferred with the enzyme aliquot to ensure its stability. Both the concentration of glucose B-phosphate and the amount of phosphofructokinase added were varied according to the desired conditions of pH and temperature. Phosphofructokinase aliquots were diluted with 0.05 M cysteine, pH 8.5, and preincubated at room temperature for 5 min in order to ensure maximal activity (17). The pH meter of the Radiometer Titrator-Titrigraph combination was standardized against, pH 7 Beckman standard buffer at’ the be- ginning of each experiment, and periodically checked for linear- ity between pH 4 and 10. Temperatures above ambient were controlled with a constant temperature circulating bath (Haake, Berlin, Germany), and those below ambient with a Magni-Whirl constant temperature refrigerated bath (Tslue &I Electric Com- pany, Blue Island, Illinois).
At pH values below 8.2, the stoichiometry ITith respect to H+ of the reaction
Fructose-6-P + ATP phosphofructokinase
fructose-l,G-di-P + ADP + H+
is less than 1. In order, therefore, to establish the validity of the assay over the pH range in which it was used, the stoichiometry was determined under the standard assay conditions as a function of pH at 30”. This determination was made by measuring the
NaOH consumed during complete conversion of known aliquots of glucose B-phosphate to fructose 1,6-diphosphate at. a given pH value. The results of a series of experiments carried out over the pH range 6.0 to 10.0 are shown in Fig. 1. The values indi- cated by the solid line drawn in this figure have been used for the
correction of all velocity measurements made at pH values below 8.2. The curve drawn in Fig. I agrees essentially wit’h that ob- tained by hlberty, Smith, and Bock (18) on direct titration of of ATP. Furthermore, these authors had found that for a tem- perature difference of 13” the pKZ of XTP changed by only 0.02. The values for the stoichiometry indicated in Fig. 1 have there- fore also been utilized for the correction of velocity measurements made at 10” and 40”.
In order to ensure that the coupling enzyme phosphofructo- kinase did not become rate-limiting at any pH, and also to es- tablish its stability over t.he pH range examined, the minimum amount of phosphofructokinase required to obtain true initial velocities was det)ermined as a function of pH. In these experi-
0.0’ ’ I I I I I I I I
6.0 6.5 7.0 7.5 8.0 8.5 9.0 9.5 10.0
DH
FIG. 1. Stoichiometry as a function of plI for the reaction sequence catalyzed by phosphoglucose isomerase and phospho- fructokinase in the PI-I-stat assay of the forward reaction. The titration vessel contained, in a volume of 3 ml, final concentrations of 5 mM ATP, 5 mu MgSO4, KC1 to bring t’he ionic strength to 0.12, and sufficient phosphoglucosc isomerase and phosphofructo- kinase to ensure a rapid completion of the conversion of glucose B-phosphate (G-6-P) to fructose 1,Cdiphosphate at each given pH value. The total amount of glucose G-phosphate added was varied from 2.6 to 0.3 pmolee, end the concentration of the NaO1-I titrating agent, from 4 to 30 rnM. The temperature was 30”. The solid line was drawn as the best visual fit to the exuerirnentsl points. The values indicated by this line were utilizid for the correction of all velocity measurements in the forward direction at pH values below 8.2.
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
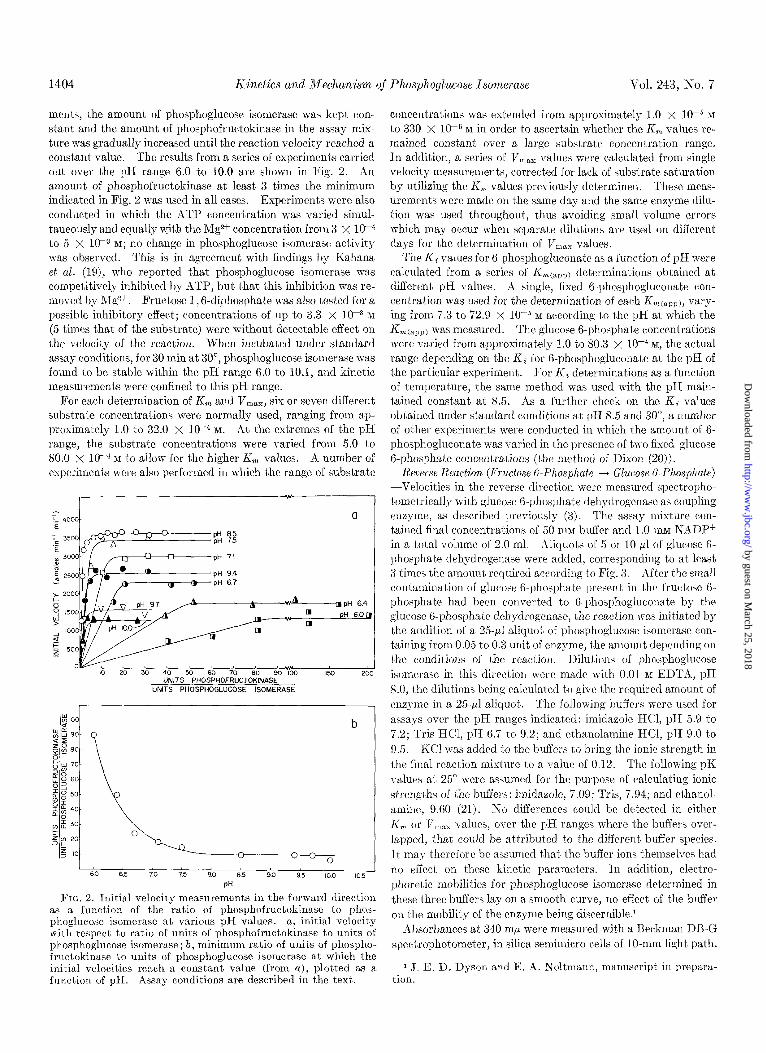
1404 Kinetics and Mechanism of Phosphoglucose Isomerase Vol. 243, No. 7
men&, the amount of phosphoglucose isomerase was kept con- stant and the amount of phosphofructokinase in the assay mix- ture was gradually increased until the reaction velocity reached a con&ant value. The result’s from a series of experiments carried out over the pH range 6.0 to 10.0 are shown in Fig. 2. An amount of phosphofructokinase at least 3 times the minimum indicated in Fig. 2 was used in all cases. Experiments were also conducted in which t,he ATP concentration was varied simul- taneously and equally u:it,h the lVlg* concentration from 3 x low4 to 5 x 10m3 RI; no change in phosphoglucose isomerase activity was observed. This is in agreement with findings by Kahana et al. (19), who report’ed that phosphoglucose isomerase was competitively inhibited by ATP, but that this inhibition was re- moved by Mgzt. Fructose 1 , 6-diphosphate was also tested for a possible inhibitory effect; concentrations of up to 3.3 x 10h3 M (5 times that of t)he substrate) were without detectable effect on the velocity of the reaction. When incubated under standard assay conditions, for 30 min at 30”, phosphoglucose isomerase was found to be stable within the pH range 6.0 to 10.1, and kinetic measurements were confined to this pH range.
For each determination of K, and V,,,, six or seven different substrate concentrations were normally used, ranging from ap- proximately 1.0 to 32.0 x 10M4 M. At the extremes of the pH range, the substrate concentrations were varied from 5.0 t’o 80.0 x 10U4 M to allow for the higher Xi, values. A number of experiment,s were also performed in which the range of substrate
IO 20 30 40 UNlTS50PHO&OF~CT08K9NASE 90 1x3 200
UNITS PHOSPHOGLUCOSE ISOMERASE
! 60 65
I 70 75 80 8.5 9.0 95 100 10.5
PH
FIG. 2. Initial velocity measurements in the forward direction as a function of the ratio of phosphofructokinase to phos- phoglucose isomerase at various pH vdues. a, initial velocity with respect to ratio of units of phosphofructokinase to units of phosphoglucose isomerase; b, minimum ratio of units of phospho- fructokinase to units of phosphoglucose isomerase at which the initial velocities reach a constant value (from a), plotted as a function of pH. Assay conditions are described in the text.
concentrations was extended from approximately 1.0 x lop5 M
to 330 X lop5 M in order to ascertain whether the K, values re- mained constant over a large substrate concentration range. In addition, a series of V,,,,, values were calculated from single velocity measurements, corrected for lack of substrate saturation by utilizing the K, values previously determined. These meas- urements were made on the same day and the same enzyme dilu- tion was used throughout, thus avoiding small volume errors which may occur when separate dilutions are used on different days for the determination of V,,, values.
The Ki values for 6-phosphogluconate as a function of pH were calculabed from a series of Z<m(app) determinations obtained at different pH values. A single, fixed 6-phosphogluconate con- centration was used for the determination of each KnLCapp), vary- ing from 7.3 to 72.9 x lo-” M according to t,he pH at which the K m(aDp) was measured. The glucose A-phosphate concentrations were varied from approximately 1 .O to 80.3 x 10m4 M, the actual range depending on the Ki for 6-phosphogluconate at the pH of the particular experiment. For Ki determinations as a function of temperature, the same method was used with the pH main- tained constant at 8.5. As a further check on the Ki values obtained under standard conditions at pH 8.5 and 30”, a number of other experiments were conducted in which the amount of 6- phosphogluconate was varied in t,he presence of two fixed glucose 6-phosphate concentrations (the method of Dixon (20)).
Reverse Reaction (Fructose 6-Phosphate --f Glucose 6-Phosphate) -Velocities in the reverse direction were measured spectropho- tometrically with glurosr 6-phosphate dehydrogenase as coupling enzyme, as described previously (3). The assay mixture con- tained final concentrations of 50 mM buffer and 1 .O mM XiADP+ in a total volume of 2.0 ml. Xliquot,s of 5 or 10 ,uI of glucose 6- phosphate dehydrogenase were added, corresponding to at least 3 times the amount rcquircd according to Fig. 3. Aft,er the small contaminat,ion of glucose 6.phosphate preselrt in the fruct,ose 6- phosphat,e had been converted to 6-phosphogluconate by the glucose 6-phosphate dehydrogenase, t,he reaction was initiated by the addition of a 25.~1 aliquot of phosphoglucose isomerase con- taining from 0.05 to 0.3 unit of enzyme, the amount depending on the conditions of the reaction. Dilutions of I’hosphoglucose
isomerase in this direction were made with 0.01 nr EDTX, pH 8.0, the dilutions being calculat,ed to give t,he required amount of enzyme in a 25,~l aliquot. The following buffers were used for assays over t,hc pH ranges indicated: imidazolc HCl, pH 5.9 to 7.2; Tris HCl, pH 6.7 t,o 9.2; and et,hanolamine HCI, pH 9.0 to 9.5. KC1 was added to the buffers to bring the ionic st’rengt,h in the final reaction mixture to a value of 0.12. The following pK values at 25” were assumed for the purpose of calculat’ing ionic strengths of the buffers: imidazole, 7.09; Tris, 7.94; and et,ha.nol- amine, 9.60 (21). No differences could be detect,ed in either K, or Ir,,,,, values, over the pH ranges where t,he buffers over- lapped, that could be atbributed to the different buffer species. It may therefore be assumed that the buffer ions themselves had no effect on these kinetic parameters. In addit,ion, electro- phoretic mobilities for phosphoglucose isomerase determined in these three buffers lay on a smooth curve, no effect of the buffer on the mobility of the enzyme being discernible?
ilbsorbances at 340 rnp were measured with a Beckman DB-G spectrophotometer, in silica semimicro cells of lo-mm light path.
1 J. E. D. Dyson and E. A. Noltmann, manuscript in preparn- tion.
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
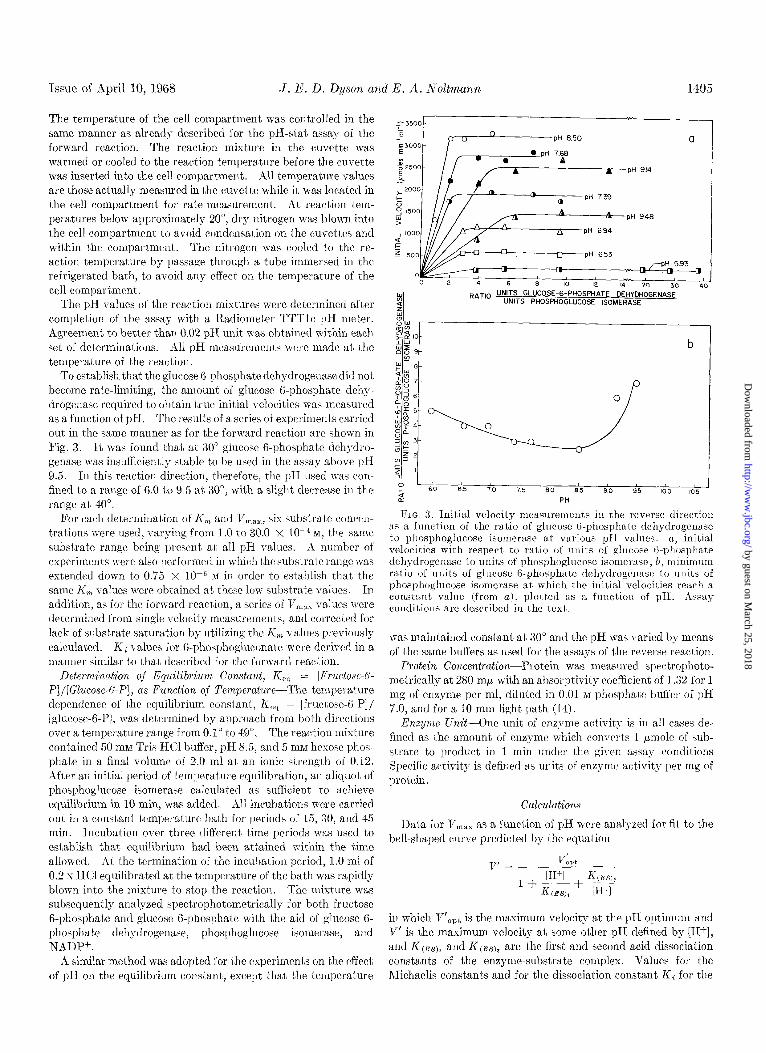
Issue of April 10, 1968 J. 13. D. Dyson and E. A. Noltmann 1405
The temperature of the cell compartment was controlled in the same manner as already described for the pH-stat assay of the forward reaction. The reaction mixture in the cuvette was warmed or cooled to the reaction temperature before the cuvette was inserted into the cell compartment. All temperature values are those actually measured in the cuvette while it was located in the cell compartment for rate measurement. At reaction tem- peratures below approximately 20”, dry nitrogen was blown into the cell compartment to avoid condensation on the cuvettes and within the compartment. The nitrogen was cooled to the re- action temperature by passage through a tube immersed in the refrigerated bath, to avoid any effect on the temperakre of the cell compartment.
The pH values of the reaction mixtures were determined aft,er completion of the assay wit#h a Radiometer TTTlc pH meter. Agreement to better than 0.02 pH unit was obtained within each set of determinations. All pH measurements were made at t,he temperature of the reaction.
To establish that the glucose B-phosphate dehydrogenase did not become rate-limiting, the amount of glucose 6-phosphate dehy- drogenase required to obtain true initial velocities was measured as a function of pH. The results of a series of experiments carried out in the same manner as for the forward reaction are shown in Fig. 3. It was found that at 30” glucose B-phosphate dehydro- gcnase was insufficiently stable to be used in the assay above pH 9.5. In this reaction direction, therefore, the pH used was COW fined to a range of 6.0 to 9.5 at 30”, wit,h a slight decrease in the range at 40”.
For each determination of K, and 1/1,18X, six substrate concen- trations were used, varying from 1 .O to 30.0 x lo-” M, the same substrate range being present at all pH values. A number of experiments were also performed in which the substrat,e range was extended down to 0.75 x IO-” RI in order to establish that the same K, values were obtained at these low substrate values. In
addition, as for the forward react,ion, a series of Ti,,, values were determined from single velocity measurements, and corrected for lack of substrate saturation by utilizing the K, values previously calculated. Ki values for 6-phosphogluconate were derived in a manner similar to that described for the forward reaction.
Determination oj” Equilibrium Constant, K,, = [Fructose-6-
P]/[Glucose-6-P], as Function oj Teynperature-The temperature dependence of the equilibrium constant, K,, = [fructose-B-P]/ [glucose-B-P], was determined by approach from both directions over a temperature range from 0.1” to 49”. The reaction mixture contained 50 ml1 Tris HCl buffer, pH 8.5, and 5 mM hexose phos- phate in a final volume of 2.0 ml at an ionic strength of 0.12. After an initial period of temperature equilibration, an aliquot of phosphoglucose isomerase calculat,ed as sufficient to achieve equilibrium in 10 min, was added. All incubations were carried out in a constant temperature bath for periods of 15, 30, and 45 min. Incubation over three different time periods was used to establish that equilibrium had been att,ained within the t’ime allowed. At the termination of the incubation period, 1 .O ml of 0.2 N HCI equilibrated at the temperature of the bath was rapidly blown into the mixture to stop the reaction. The mixt,ure was subsequently analyzed spectrophotometrically for both fructose 6-phosphate and glucose 6-phosphate with the aid of glucose 6- phosphate dehydrogenase, phosphoglucose isomerase, and NADP+.
A similar method was adopted for the experiments on t’he effect of pH on t,be equilibrium constant, except that the temperature
n PH 739
A J PH 948
L1 PH 694
-pH 653 pH 593
Q =,b I; ,c Y- I RAT,O UNITS GLUCOSE-6-PHOSPHATE DEHYDROGENASE
UNITS PHOSPWOGLUCOSE ISOMERASE
-0 1
40
P I
t 60 6.5 70 75 80 85 90 9.5 - 105
n PH
FIG. 3. Initial velocity measurement,s in the reverse direction its a function of the ralio of glucose (i-phosphate dchydrogetlnse to phosphoglncose isomerase at various pIr valises. (2, initial velocities with respect to ratio of units of glllcose &phosphate dehgdrogenase to units of phosphoglucose isornerssc; b, minimum ratio of units of glucose 6-phosphate dehydrogerlase to units of phosphoglucose isomerase at which the initial velocities reach a constant valrle (from a), plotted as a fuuctiolr of pH. Assay conditions are described in the text.
was maintained constant at 30” and the pH was varied by means of the same buffers as used for the assays of the reverse reaction.
Protein Co?lcentration-Protein was measured spectrophoto- metrically at 280 rnh with an absorptivity coefficient of 1.32 for 1 mg of enzyme per ml, diluted in 0.01 M phosphate buffer of pH 7.0, and for a IO-mm light path (14).
Enzyme Unit--One unit of enzyme activity is in all cases de- fined as the amount of enzyme which converts 1 pmole of sub- &rate to product in 1 mill under the given assay conditions Specific act’ivity is defined as units of enzyme activity per mg of protein.
Calculations
Data for V,x,,, as a function of pH were analyzed for fit to the bell-shaped curve predicted by t’hc equation
in which V’,,t is the maximum velocity at the pH optimum and 8’ is t.he maximum velocity at some other pH defined by [Hf], and KcESJ1 and KcES)2 are the first and second acid dissociation constants of the enzyme-substrate complex. Values for the Michaelis constants and for the dissociation constant Ki for the
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1406 Kinetics and Mechanism of Phosphoglucose Isomerase Vol. 243, No. 7
enzyme&phosphogluconate complex, determined as a function of pH, were plotted in logarithmic manner and interpreted as originally suggested by Dixon (22).
Michaelis constants (K,), maximum initial velocities (V,,,), and their ratio (V,,,/K,), together with standard errors and weight’ing factors, were calculated with the aid of the Riverside Camps I13M 7040 digital comput’er; for this purpose a program developed by Cleland (23) was used, in which the format st,at,e- mcnt? had been modSed to suit t,he IUM 7040. This program uses weighted velocity measurement’s and substrate concentra- t’ions in a manner similar to that described by Wilkinson (24). The beat fit of the V,,,, data to a bell-shaped curve, together with the ionization constants and their standard errors, was also ob- tained from computer analysis by using the V,,,, values and t,heir weighting factors previously determined. Cleland’s pro- gram for a least squares fit to a bell-shaped curve was adapted for this purpose (23). The V,,,, values obtained from single veloc- ity measurements were included in the data, their weighting being set equal to the highest weighting for the Vr,,, values obtained from velocity measurements as a function of the substrate con- centration. This approximation was considered justified, since the relative accuracy of the single velocity measurements, per- formed with the same enzyme dilution, is probably greater than that, of i;he VI,,, values extrapolated from velocity measurements as a function of substrate concentration, which were obtained with different enzyme dilutions and might therefore have in- cluded inherent dilution errors.
RESULTS 4ND DISCUSSION
Kinetic Parameters at Optimum pII-The standard kinetic con- stants obtained for rabbit muscle phosphoglucose isomerase at pH 8.5 and 30” are summarized in Table I. The values for K, are comparable with the majority of previously reported deter- minations made under similar conditions with phosphoglucose isomerases from various sources. A significant difference exists only with respect to the values of Kahana et al. (19) obtained with
TABLE 1
Kinetic parameters for rabbit muscle phosphoglucose isomerase at pH 8.5, SO”, cd 0.12 ionic strength
Parameter Forward reaction: glucose- Reverse reaction: fmctose- 6-P + fructose-h-P 6-P + &mm-6-P
K, (moles liter-l)“. 3.10 i 0.56 x 10-d 1.74 f 0.13 x 10-d V max (fimoles min-1
mg-l)b. 586 * 14 1037 f 13 Molecular activity”. 7.62 z!z 0.19 X lo4 13.48 f 0.17 x 104 Hnldane equilibrium
constant calculated from above values. 0.32
Equilibriumconstant obtained experi- mentally. / 0.30
-
a Forward direction: mean of six determinations made with dif- ferent enzyme preparations over a period of 2 years. Reverse direction: mean of five determinations.
b Determined from six velocity measurements in either direc- tion made on the same day with the same enzyme dilut,ion and cor- rected for lack of substrate saturation with the I& values given above.
~Moles of substrate converted per min per mole of enzyme, hnsed on a molecular weight of 130,000.
crude preparations of phosphoglucose isomerase from rabbit muscle and rabbit brain. Their experimental data resulted in uncorrected K, values approximately one order of magnitude smaller than were obtained in the present study and most other reports. Considerable effort has been expended to find an ex- planation for this divergence.
rZ variation of K, with substrate concentration cannot be the cause for the difference, since measurements in the present study and in that of Kahana et al. were made over similar substrate concentration ranges. A number of experiments conducted in both reaction directions, in which the substrate concentrations were varied over a 300.fold range from 0.01 1x1~ to 3.2 mM, re- sulted in data that did not indicate deviat,ion from straight lines when plotted in a reciprocal manner. Individual Ii, values were furthermore determined for each fraction of the enzyme isolation procedure (3), and no variation could be observed in data obt,ained from the crude homogenate through the five times crystallized phosphoglucose isomerase.2 Finally, two independ- ent methods were used in the present work for the assay of rcac- tion velocities in the forward and the reverse directions, and good agreement was found between the measured equilibrium constant and the one calculated from the Haldane relationship (Table I), thus testifying to the validity of the assay methods (for other criteria concerning the rate measurements, see “Methods”). It appears, therefore, that thus far no explanation for the dis- crepancy between the I& values other than a difference in methodology3 is forthcoming.
EJect 0~” pIl on Kinetic Parameters
Dependence of K, on pH
The pH dependence of K, is shown in Figs. 4 and 5, plotted as pK, with respect to pH for the forward reaction, and as pK, and log (V,,,,/K,) with respect to pH for the reverse reaction. The parameter log (V,,,/K,) was used in the plot for the reverse reaction in order to indicate more clearly the inflection points, since, owing to the small differences between the pK values for
the free enzyme and those for the enzyme-substrate complex, a plot of pKm with respect t,o pH is almost linear.
The values for the ionization constant,s of the free enzyme are
2 The average Ii,, determined over all the fractions of the en- zyme isolation procedure, u-as 1.56 + 0.07 X 10e4 M, under stand- ard conditions in the reverse direction.
3Kahann et al. (19) used coupled fluorimetric assays for both directions. It was pointed out by these authors that the coupling enzymes used for assaying the forward reaction were not free from ” . interfering contaminations, but that it was possible to reduce assav difficulties “to the uoint at which DPNH dissaoearance over the time interval concerned did not exceed fructose-di-P formation by more than lOo/,.” The substrate used by Kahana el al. for most of the assays in the reverse direction contained 14.8ojb as much glucose B-phosphate as fructose B-phosphate (page 2179 of Eeference 19) compared with a glucose O-phosphate con- tamination of 0.3 to 0.6% in the fructose G-phosphate samples utilized in the present work. The high glucose 6-phosphate con- tamination of the fructose 6-phosphate used by Kahsna et al. necessitated subst,antial corrections of all their rnle measuremeni,s for the initially produced f-phosphogluconat,e, which were made by these authors assuming a KC of 0.005 mnx; the latter was also calculated by correcting measured velocities for unavoidably present contributions from contamination of the substrate. In the present work, a Kc of 0.15 IIIM was found at pH 8.0 with enzy- matically standardized G-phosphogluconate in assays of both the forward and the reverse reaction and with substrates and coupling enzymes essentially free from interfering contaminations.
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
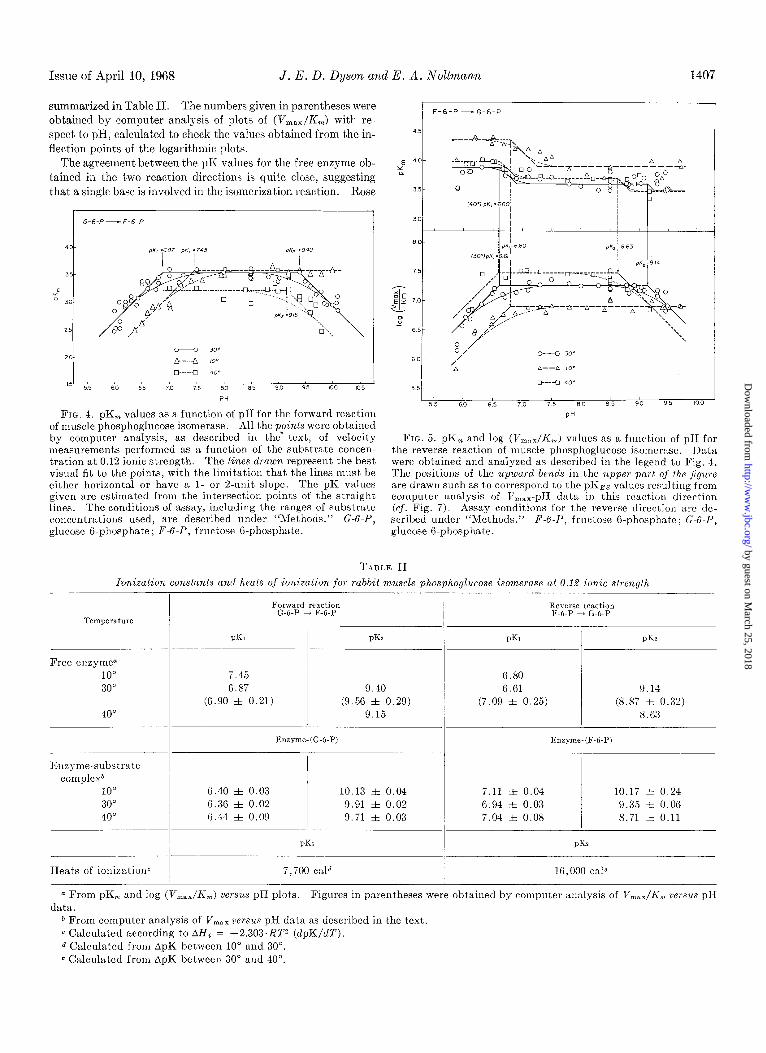
Issue of April 10, 1968 J. E. D. Dyson and E. A. Noltmann 1407
summarized in Table II. The numbers given in parentheses were
obtained by computer analysis of plots of (V,,,/R,) with re-
spect to pH, calculated to check the values obtained from the in- flection points of the logarithmic plots.
The agreement between the pK values for the free enzyme ob- tained in the two reaction directions is quite close, suggesting
that a single base is involved in the isomerization reaction. Rose
1 ! I F-6-p -G-6-P
4%
-
d a
x z: t H
FIG. 4. pK,, values as a function of pH for the forward reaction of muscle phosphoglucose isomerase. All the points were obtained by computer analysis, as described in the‘ text, of velocity measurements performed as a function of the substrate concen- tration at 0.12 ionic strength. The lines drawn represent the best visual fit to the points, with the limitation that the lines must be either horizontal or have a I- or 2.unit slope. The pK values given are estimated from the intersection points of the straight lines. The conditions of assay, including the ranges of substrate concentrations used, are described under “Methods.” G-G-P, glucose 6.phosphate; F-6-P, fructose G-phosphate.
FIG. 5. pK,, and log (V,,:,,/I<,,,) values as a function of pII for the reverse reaction of muscle phosphoglucose isomerase. I>ata were obtained and analyzed as described in the legend to Fig. 4. The positions of the upward bends in the upper pnrt of the ,/igure are drawn such as to correspond to the EKES values resulting from computer analysis of V,,,,- pI1 data in this reaction direction (cf. Fig. 7). Assay conditions for the reverse directiou are de- scribed under “Methods.” F-6-1’, fructose G-phosphate; G-6-1’) glucose B-phosphate.
TABLE II
ionization constunts and heats of ionization for rabbit muscle phosphoglucose isomeruse at 0.12 ionic strength
Forward reaction G-6-P + F-6-P
Reverse reaction F-6-P i G-6-P
Temperature
PKI PKZ PK1 PK~
Free enzyme” 10” 30”
40”
7.45 6.87 9.40
(6.90 zt 0.21) (9.56 + 0.29) 9.15
G.80 6.61 9.14
(7.09 * 0.25) (8.87 zt 0.32) S.G3
Enzyme-(G-6-P) Enzyme-(F-6-P) -.
-.
Enzyme-substrate complexb
10” 30” 40”
G.40 i 0.03 10.13 f 0.04 G.36 + 0.02 9.91 + 0.02 G.44 3~ 0.09 9.71 + 0.03
i.11 f 0.04 10.17 f 0.24 6.94 A 0.03 9.35 i 0.06 7.04 It 0.08 8.71 f 0.11
pK? PKS
Heats of ionizationc 7,iO0 cald 16,000 tale
Q From pK, and log (V,,,/K,) versus pH plots. Figures in parentheses were obtained by computer analysis of V,,,,/K,,, versus p1-T data.
b From computer analysis of V max versus pH data as described in the text. c Calculated according to AHi = -2.303~RP (dpK/dT). d Calculated from ApK between 10” and 30”. c Calculated from ApK between 30” and 40”.
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1408 Kinetics and Mechanism of Phosphoglucose Isomerase Vol. 243, No. 7
(13) pointed out that for phosphoglucose isomerase the ratio of the initial rates of tritium labeling of substrate and product is the same, whether fructose B-phosphate or glucose 6-phosphate is the starting compound. He concluded that a single base must be involved in the catalyt.ic reaction; otherwise, different ratios would be obtained (13). The interpretation of the present data on the basis of the participation of a single base is thus supported by previous experiments.
pK1 Value for Free Enzyme-The pK1 values obtained agree closely with those found in protein molecules for imidazole groups (25), and suggest the participation of a histidine residue in the catalytic interconversion of glucose B-phosphate and fructose B-phosphate. Although side chain carboxyl groups have been found with pKi,t values as high as 7.3, these have all been anom- alous groups unavailable to the solvent, which, in the denatured protein, could be titrated in the normal range with pK values of approximately 4.5 (25). Thus, although caution is necessary in implicating a histidine residue on the basis of its pK alone, a carboxyl group seems very unlikely. The only other group with a pK approaching that found is a terminal amino group; little information is available concerning its pKr,t values, although pK values of approximately 7.8 might be anticipated from data on small molecules (25). Participation of a terminal amino group seems improbable, however, in view of the data of Rose and O’Connell (1). These authors postulated, on the basis of the fraction of 14C associated with deuterium when fructose 6-phos- phate I-D, PC was converted to glucose 6-phosphate, that the base involved in the isomerization reaction is a base with no ex- changeable hydrogens, i.e. a carboxylate group or the nonpro- tonated nitrogen of an imidazole ring (1).
The apparent heat of ionization calculated with the average ApK values obtained from the forward and reverse reactions at 10” and 30” is 7,700 cal. This value would appear to eliminat’e a phosphate ionization of the substrate as responsible for the pKi, especially since direct titration of both glucose B-phosphate and fructose 6-phosphate under the conditions of activity meas- urement yielded a pK value of 6.15 f 0.02 for the free substrates. A Alli of 7,700 cal is similar to that found for histidylglycine (26)) and, although the dat,a on apparent heats of ionization for imid- azole in protein molecules are rather limited, values of 6,500 to 7,500 cal are usually found (25, 27). As Lindley has pointed out (2x), proximity of two ionizing groups in the protein molecule will lead to apparent heat,s of ionization, determined for the two groups, approaching a value intermediate between the two true values, the discrepancy depending on how closely the pK values of the two groups concerned approach each other. A carboxyl group has a heat of ionization in the range of -2,000 to 2,000 Cal; thus, in order to obtain for it an apparent heat of ionization of 7,700 cal, there must, be a group adjacent to the carboxyl group with a heat of ionization of the order of 13,000 cal. The only groups in protein molecules with heats of ionization of this mag- nitude arc terminal and e-amino groups and guanidinium groups.
As discussed later, it seems probable that the group with a pK? of approximately 9.3 is an e-amino group of lysine. How- ever, as pointed out by Tanford (29), the presence of a positively charged amino group adjacent to a carboxyl group in a protein molecule will lead to a lowering of the pKi,,t of the carbosyl group. A carboxyl group may thus be discounted in the present case, and the evidence is most strongly in favor of an imidazolc being the residue involved. Experiments carried out in this laboratory or1 the rcactioli of phosl)hoglucose isomerase with
iodoacetamide4 and on methylene blue-sensitized photooxidation (30) are also in agreement with the involvement of histidine. Thus, three independent methods are all indicative of histidine as the amino acid residue participating in the catalytic process.
pKz Value for Free Enzyme-An assignment for the pKZ of about 9.3 cannot be made as readily as for the pK1. The e- amino group of lysinc, the phenolic hydroxyl group of tyrosine, and the sulfhydryl group of cysteine have pK values in this re- gion. It seems possible, however, to eliminate tyrosine and cysteine from consideration on the basis of the following evidence.
First, the apparent heat of ionization obtained from the average ApK values of the forward and reverse reactions at 30” and 40” is 16,000 cal; this value, although rather high, is of the right order of magnitude for an e-amino group. It is much too high however for a phenolic hydroxyl or a sulfhydryl group, which have heats of ionization of approximately 7,000 cal (25, 27, 29) and 6,900 cal (31), respectively. As pointed out above, the presence of an adjacent group with a lower heat of ionization tends to decrease the true value measured for a particular chemical group (28). In the present case, the value obtained is somewhat higher than the values usually assigned (13,000 cal) to terminal and e-amino groups and guanidinium groups, which have the highest heats of ionization found in proteins. It is thus unlikely that the heat of ionization measured here has been affected by the presence of an adjacent group.
Second, on spectrophotometric titration’ of the enzyme, no ionization of the phenolic hydroxyl groups can be detected until a pH of approximately 10.2 is reached. In 8 M urea, however, all the phenolic hydroxyl groups are available to the solvent and titrate normally. When incubated for 30 min at 30”, the enzyme begins to lose activity at about pH 10, the loss of activity in- creasing with pH up to approximately pH 11.3, where it is virtu- ally complete.’ The time dependence of the titration of the phenolic hydroxyl groups in phosphoglucose isomerase indicates that a time-dependent conformation change must take place in the enzyme molecule before any phenolic hydroxyl groups become available to the solvent. It seems reasonable to equate the loss of catalytic act,ivity, beginning at pH values above 10, with this time-dependent change in conformation, which also commences at about pH 10. Thus, full activity on the one hand, and avail- ability of any of the phenolic hydroxyl groups to the solvent on the other hand, appear to be two factors normally not found in the same phosphoglucose isomerase molecule.
Availability to the solvent, however, may be considered a necessary criterion for an amino acid residue to be involved in a catalytic process with such hydrophilic molecules as glucose 6-phosphate and fructose B-phosphate. Although the possibility cannot be entirely eliminated, it seems unlikely that the hexose B-phosphates could enter a hydrophobic environment, such as that in which the phenolic hydroxyl groups must be located in order not to react with the solvent, even if steric considerations would allow it. Thus, it appears justifiable to eliminate tgrosine as a participant in the catalytic process.
With regard to the sulfhydryl groups of cyeteine, experiments in this laboratory have shown that 8 out of the 12 sulfhydryl groups present in phosphoglucose isomerase can react. with organic mercurial compounds bcforc any loss of enzyme activity occurs (32). Since by its nature, a group involved in the cata- lytic l~rocess would bc assumed to be among the more reactive of
4 K. Schnackerz :~rrd I<. A. Noltrnann, linpnblishctl experiments.
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
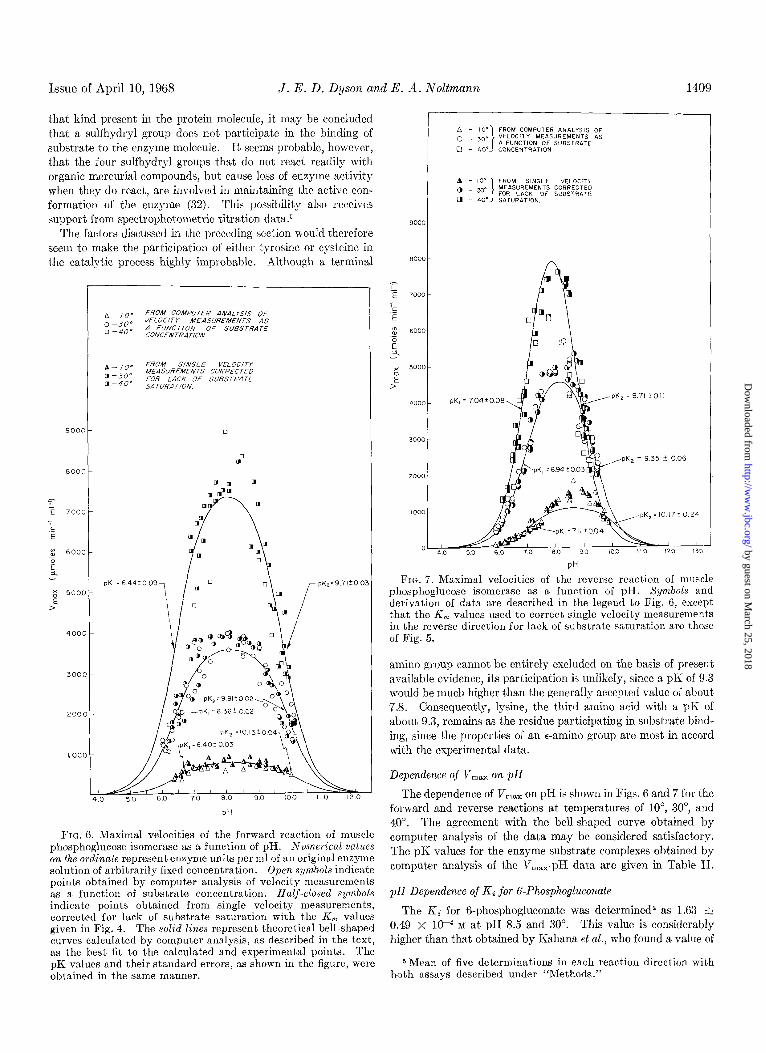
Issue of April 10, 1968 J. E. D. Dyson and E. A. Noltmann 1409
that kind present in the protein molecule, it may be concluded that a sulfhydryl group does not participate in the binding of substrate to the enzyme molecule. It seems probable, however, that the four sulfhydryl groups that do not react readily with organic mercurial compounds, but cause loss of enzyme activity when they do react, are involved in maintaining the active con- formation of the enzyme (32). This possibility also receives support from spectrophotometric titration data.l
The factors discussed in the preceding section would therefore seem to make the participation of either tyrosine or cysteine in the catalytic process highly improbable. Although a terminal
900
800
_I L E 700
;
E
5 600 0
5
g 500
Z
400
300
200
LOO
b--/O” FROM COMPUTER ANALYSIS OF
0 -30” VELOCITY MEASUREMENTS AS
0 -40” A F”NCNON OF SUBSTRLITE CONCEN TRA TION
A--/O” FROM SINGLE VELOCITY
O-30 MEA.wREhfENTS CORRECTED
0 -40” FOR LACK OF SUBSTRATE SA TURA TION.
pK,=6,44tO09, / q 0
FIG. 6. Maximal velocities of the forward reaction of muscle phosphoglucose isomerase as a function of pH. AJumerical values on the ordinate represent enzyme units per ml of an original enzyme solution of arbitrarily fixed concentration. Open symbols indicate points obtained by computer analysis of velocity measurements as a function of substrate concentration. Half-closed symbols indicate p0int.s obtained from single velocity measurements, corrected for lack of substrate saturation with the K, values given in Fig. 4. The solid lines represent theoretical bell-shaped curves calculated by computer analysis, as described in the text, as the best fit to the calculated and experimental points. The pK values and their standard errors, as shown in the figure, were obtained in the same manner.
pK, = 704f008 3i.l ,pK, = 871 foil
“43 ,pK,=935+006
FIG. 7. Maximal velocities of the reverse reaction of muscle phosphoglucose isomerase as a function of pH. Symbols and derivation of data are described in the legend to Fig. 6, except that the K, values used to correct single velocity measurements in the reverse direction for lack of substrate saturation are those of Fig. 5.
amino group cannot be entirely excluded on the basis of present available evidence, its participation is unlikely, since a pK of 9.3 would be much higher than the generally accepted value of about 7.8. Consequently, lysine, the third amino acid with a pK of about 9.3, remains as the residue participating in substrate bind- ing, since the properties of an e-amino group are most in accord with the experimental data.
Dependence of V,,, on pH
The dependence of V,,, on pH is shown in Figs. 6 and 7 for the forward and reverse reactions at temperatures of lo”, 30”, and 40”. The agreement with the bell-shaped curve obtained by computer analysis of the data may be considered satisfactory. The pK values for the enzyme-substrate complexes obtained by computer analysis of the V,,,-pH data are given in Table II.
pH Dependence of Ki for 6-Phosphogluconate
The K; for 6-phosphogluconate was determined5 as 1.63 f 0.49 x 10-J M at pH 8.5 and 30”. This value is considerably higher than that obtained by Kahana et al., who found a value of
5 Mean of five determinations in each reaction direction with both assays described under “Methods.”
PH
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
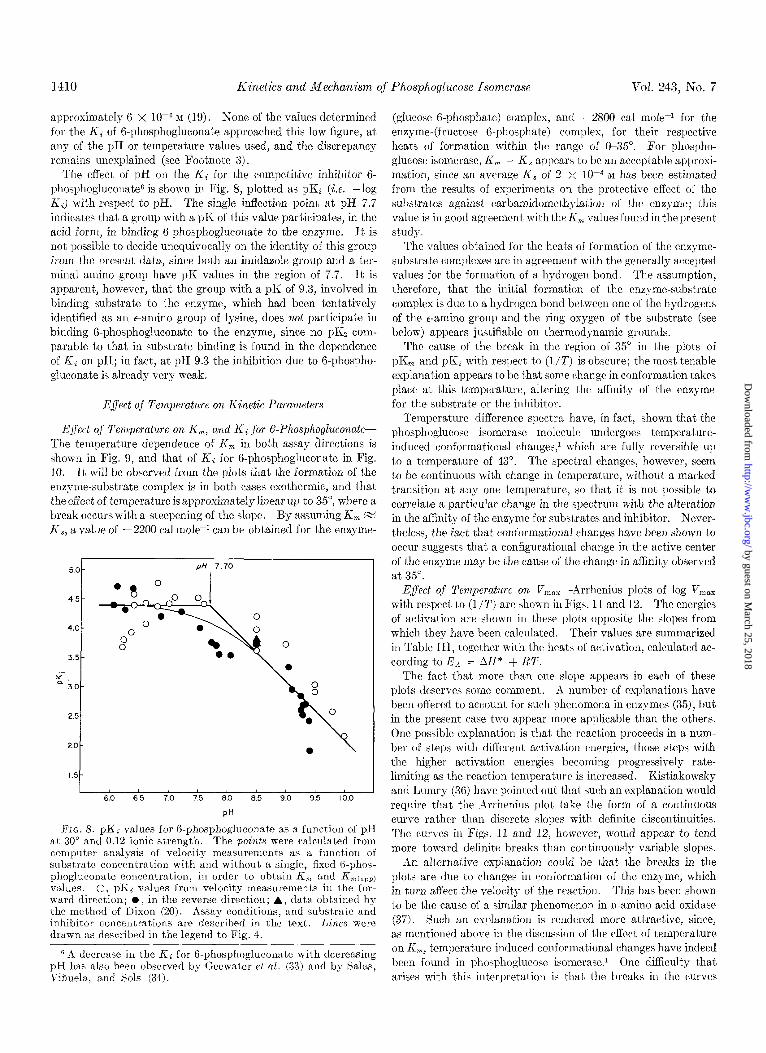
l-1-10 Kinetics and Mechanism of Phosphoglucose Isomeruse Vol. 243, No. 7
approximately 6 x lo+ M (19). None of the values determined for the Ki of 6-phosphogluconate approached this low figure, at any of the pH or temperature values used, and the discrepancy remains unexplained (see Footnote 3).
The effect of pH on the K; for the competitive inhibitor 6- phosphogluconate6 is shown in Fig. 8, plotted as pKi (i.e. -log KJ with respect to pH. The single inflection point at pH 7.7 indicates that a group with a pK of this value participat,es, in the acid form, in binding A-phosphogluconate to the enzyme. It is not’ possible to decide unequivocally on the identity of this group from the present data, since both an imidazole group and a ter- minal amino group have pK values in the region of 7.7. It is apparent, however, that the group with a pK of 9.3, involved in binding substrate to the enzyme, which had been tentatively identified as an e-amino group of lysine, does not participate in binding 6-phosphogluconate to the enzyme, since no pKz com- parable to that in substrat,e binding is found in the dependence of Ki on pH; in fact, at pH 9.3 the inhibition due to B-phospho- gluconate is already very weak.
ISfleet of Temperature on Kinetic Parameters
Eflect of Temperature on K,, and K; for &Phosphogluconate- The t,emperature dependence of K, in both assay directions is shown in Fig. 9, and that of Ki for 6-phosphogluconate in Fig. 10. It will be observed from the plots that the formation of the enzyme-substrate complex is in both cases exothermic, and that the effect of temperature is approximately linear up to 35”, where a break occurs with a steepening of the slope. By assuming K, C K,, a value of -2200 cal mole-l can be obtained for t#he enzyme-
PH 7.70
4.0 -
3.5-
.- ‘:
3.0 -
2.5 -
2.0 -
1.5-
6.0 6 5 7.0 75 60 8.5 9.0 9.5 IO.0
PH
Fro. 8. pKi values for 6.phosphogluconate as a function of pH at 30” and 0.12 ionic strength. The points were calculated from computer analysis of velocity measurements as a function of subst,rate concentration with and without a singIe, fixed 6-phos- phoglurorrate concentration, in order to obtain K,, and Krncapp) values. 0, pKi values from velocity measurements in the for- ward direction; O, in the reverse direction; A, data obtained by the method of Dixon (20). Assay conditions, and substrate and inhibitor concentrations are described in the text. Lines were drawn as described in the legend to Fig. 4.
6 A decrease in the Kc for 6-phosphogIuconate with decreasing pH has also been observed by Geewater et al. (33) and by Salas, Wiuela, and Sols (34).
(glucose B-phosphate) complex, and -2800 cal mole@ for the enzyme-(fructose 6-phosphat’e) complex, for their respective heats of formaCon within the range of O-35”. For phospho- glucose isomerase, Ii, = K, appears to bc an acceptable approxi- mation, since an average K, of 2 X 10-h M has been estimated from the results of experiments on the protective effect of the substrates against carbamidomethylation of the enzyme; this value is in good agreement with the K, values found in the present study.
The values obtained for the heats of formation of the enzyme- substrate complexes are in agreement with the generally accepted values for the formation of a hydrogen bond. The assumption, therefore, that the initial formation of the enzyme-substrate complex is due to a hydrogen bond between one of the hydrogens of the e-amino group and the ring oxygen of the substrate (see below) appears justifiable on thermodynamic grounds.
The cause of the break in the region of 35” in the plots of pK, and pKi with respect to (l/T) is obscure; the most tenable explanation appears to be that some change in conformation takes place at this temperature, altering the affinity of the enzyme for the substrate or the inhibit,or.
Temperature difference spectra have, in fact, shown that the phosphoglucose isomerase molecule undergoes temperature- induced conformational changes,l which are fully reversible up to a temperature of 43”. The spectral changes, however, seem to be continuous with change in temperature, without a marked transition at any one temperature, so that it is not possible to correlate a particular change in the spectrum with the alteration in the affinity of the enzyme for substrates and inhibitor. Never- theless, the fact that conformational changes have been shown to occur suggests that a configurational change in the active center of the enzyme may be the cause of the change in affinity observed at 35”.
EJect of Temperature on Vmar-Arrhenius plots of log Vm, with respect to (l/T) arc shown ill Figs. 11 and 12. The energies of activation are shown in these plots opposite the slopes from which they have been calculated. Their values are summarized in Table III, together with the heats of activation, calculated ac- cording to Ea = All* + RT.
The fact that more t,han one slope appears in each of these plots deserves some comment. A number of explanations have been offered to account for such phenomena in enzymes (35), but in the present case t,wo appear more applicable than the others. One possible explanation is that the reaction proceeds in a num- ber of steps with different activat,ion energies, those steps with the higher activation energies becoming progressively rate- limiting as the reaction temperature is increased. Kist’iakowsky and Lumry (36) have pointed out that such an explanation would require that the Arrhenius plot, take the form of a continuous curve rather than discrete slopes with definit,e discontinuities. The curves in Figs. 11 and 12, however, would appear to tend more toward definite breaks than continuously variable slopes.
dn alternative explanation could be that the breaks in the plots are due to changes in conformation of the enzyme, which in turn affect the velocity of the reaction. This has been shown to be the cause of a similar phenomenon in n-amino acid oxidase (37). Such an explanation is rendered more attractive, since, as mentioned above in hhe discussion of the effect of temperature on K,, temperature-induced conformational changes have indeed been found in phosphoglucose isomerase.’ One difficulty that arises with this interpretation is that the breaks in the curves
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
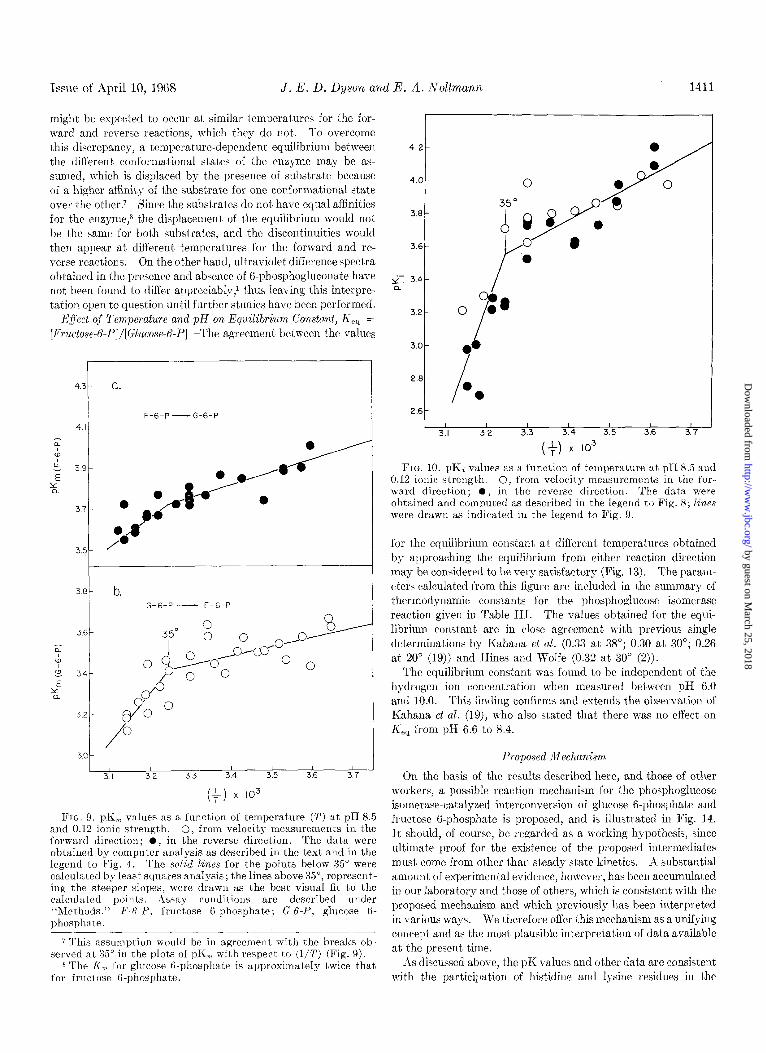
Issue of April 10, 1968 J. E. D. Dyson and E. A. Noltmann 1411
might be expected to occur at similar temperatures for the for- ward and reverse reactions, which they do not. To overcome this discrepancy, a temperature-dependent equilibrium between the different conformational states of the enzyme may be as- sumed, which is displaced by the presence of substrate because of a higher affinity of the substrate for one conformational state over the other.7 Since the substrates do not have equal affinities for the enzyme,8 the displacement of the equilibrium would not be the same for both substrates, and the discontinuities would then appear at different temperatures for the forward and re- verse reactions. On the other hand, ultraviolet difference spectra obtained in the presence and absence of 6-phosphogluconate have not been found to differ appreciably,1 thus leaving t,his interpre- t,ation open to question until further studies have been performed.
IQj’ect of Temperature and pH cm Equilibrium Constant, K,, = [li’ructose-6-P]/[GIcose-6-P]-The agreement, between the values
,E a
33
I
38 I b. G-6-P - F-6-P
36-
34-
32
/ 0
3om I I I I / /
3.1 32 3.3 ,,;: 35 3.6 37
103
FIG. 9. pK,, values as a function of temperature (2’) at pH 8.5 and 0.12 ionic strength. 0, from velocity measurements in the forward direction; l , in the reverse direction. The data were obtained by computer analysis as described in the text and in the legend to Fig. 4. The solid lines for the points below 35” were calculated by least squares analysis; the lines above 35”, represent- ing the st,eeper slopes, were drawn as the best visual fit to the calculated points. Assay conditions are described under “Methods.” F-&P, fructose B-phosphate; G-t-P, glucose 6- phosphate.
7 This assumptioll would be in agreement with the breaks ob- served at 35” in the plots of pK, with respect to (l/T) (Fig. 9).
8 The I<,, for glucose G-phosphate is approximately twice that for fructose G-phosphate.
0 $8
I 00 I
2.8 0
0 2.6 i,/
I I I I I , I 3.1 3.2 3.3 3.4 35 3.6 3.7
if) x IO3
FIG. 10. pK< values as a function of temperature at pH 8.5 and 0.12 ionic strength. 0, from velocity measurements in the for- ward direction; O, in the reverse direction. The data were obtained and computed as described in the legend to Fig. 8; lines were drawn as indicated in the legend to Fig. 9.
for the equilibrium constant at different temperatures obtained by approaching the equilibrium from cibher reaction direction may be considered to be very satisfactory (Fig. 13). The param- eters calculated from this figure are included in the summary of thermodynamic constants for the phosphoglucose isomerase reaction given in Table III. The values obtained for the equi- librium constant are in close agreement with previous single determinations by Kahana et al. (0.33 at 38”; 0.30 at 30”; 0.26 at 20” (19)) and Hines and Wolfe (0.32 at 30” (2)).
The equilibrium constant was found to be independent of the hydrogen ion concentration when measured between pH 6.0 and 10.0. This finding confirms and extends the observation of Kahana et al. (19), who also stated that there was no effect on K,, from pH 6.6 to 8.4.
Pr0po.d !lIechanism
On the basis of the results described here, and those of other workers, a possible reaction mechanism for the phosphoglucose isomerase-catalyzed interconversion of glucose B-phosphate and fructose 6-phosphate is proposed, and is illustrated in Fig. 14. It should, of course, be regarded as a working hypothesis, since ultimate proof for the existence of the proposed intermediates must come from other than steady state kinetics. A substantial amount of experimental evidence, however, has been accumulated in our laboratory and those of others, which is consistent with the proposed mechanism and which previously has been interpret)ed in various ways. We therefore offer this mechanism as a unifying concept and as the most plausible interpretation of data available
at the present time. As discussed above, the pK values and other dat’a are cnnsistent
with the participation of histidinc and lysjne residues in the
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from
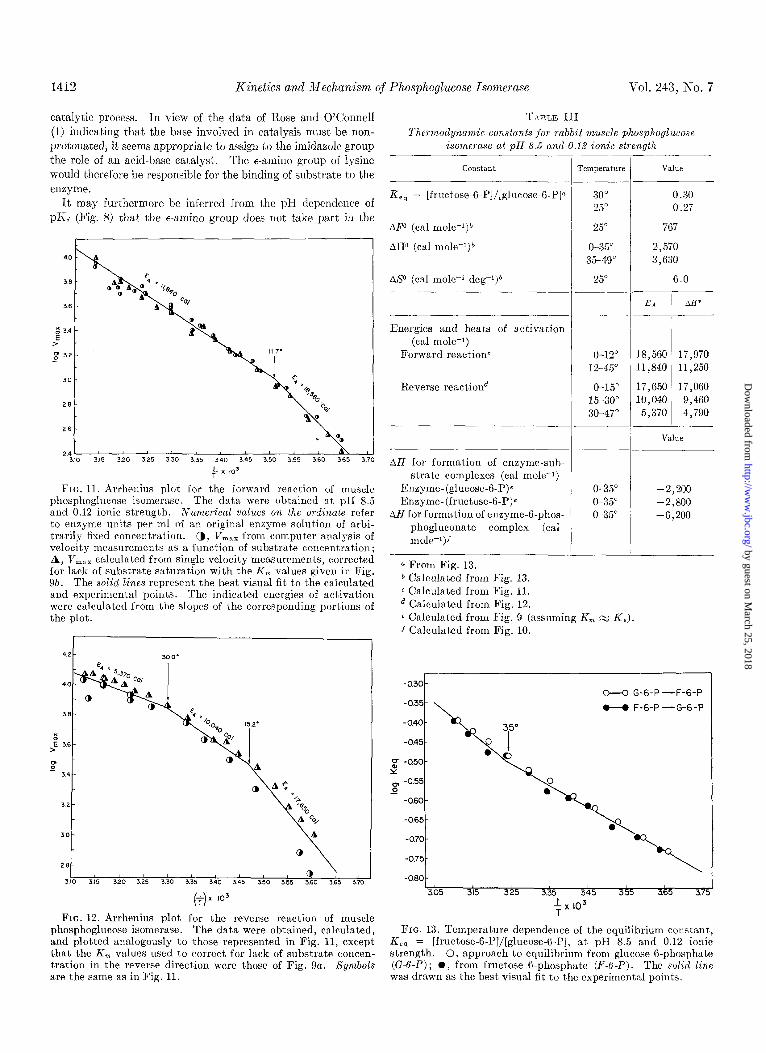
1412 Kinetics and Mechanism of Phosphoglucose Isomeyase Vol. 243, No. 7
catalytic process. In view of the data of Rose and O’Connell
(1) indicating that the base involved in catalysis must be non- protonated, it seems appropriate to assign to the imidazole group
the role of an acid-base catalyst. The e-amino group of lysine
would therefore be responsible for the binding of substrate to the enzyme.
It may furthermore be inferred from the pH dependence of
pKi (Fig. 8) that the e-amino group does not take part in the
FIG. 11. Arrhenius plot for the forward reaction of muscle phosphoglucose isomerase. The data were obtained at pH 8.5 and 0.12 ionic strength. Numerical values on the ordinate refer to enzyme units per ml of an original enzyme solution of arbi- trarily fixed concentration. a, V,,, from computer analysis of velocity measurements as a function of substrate concentration; A, V,,, calculated from single velocity measurements, corrected for lack of substrate saturation with the K, values given in Fig. 95. The solid lines represent the best visual fit to the calculated and experimental points. The indicated energies of activation were calculated from the slopes of the corresponding portions of the plot.
(3 x IO3
FIG. 12. Arrhenius plot for the reverse reaction of muscle phosphoglucose isomerase. The data were obtained, calculated, and plotted analogously to those represented in Fig. 11, except that, the K, values used to correct for lack of substrate concen- tration in the reverse direction were those of Fig. 9a. Symbols are the same as in Fig. 11.
T>LBLE III
Thermodynamic constants .,for rabbit mz~scle phosphoglzccose isome?ase at pH 8.5 and 0.16 ionic strength
constant
K,, = [fructose-6-P]/[glucose-6-P]a
AP (cal mole-l)”
AH0 (cal mole-l)b
AS0 (cal mole-' deg-l)b
Energies and heats of activation (cal mole-l)
Forward reactionc
Reverse reactiond
AH for formation of enzyme-sub- strate complexes (cal moleW1)
Enzyme-(glucose-6-P)e Enzyme- (fructose-6-P)e
AH for formation of enzyme-B-phos- phogluconate complex (cat mole-r)f
I-
-l-
Temperature
30" 25"
25"
O-35" 3549O
25"
O-12” 12-45"
O-15" 15-30" 30-47"
O-35" -2,200 O-35" -2,800 G35" -6.200
-
-.
-
-/-
a From Fig. 13. b Calculated from Fig. 13. c Calculated from Fig. 11. d Calculated from Fig. 12. e Calculated from Fig. 9 (assuming K, M K,). f Calculated from Fig. 10.
VELlW
0.30
0.27
767
2,570 3,630
G.0
EA La*
18,560 17,970 11,840 11,250
17,650 17,060 10,040 9,460
5,370 4,790
g -0.50. Y
s -0.55-
-060-
-065
-070
-0.75
-060 305 315 325 3.35 345 3.55 3.65 3.7
$x 103
FIG. 13. Temperature dependence of the equilibrium constant, Ke, = [fructose-K-P]/[glucose-B-P], at ~1% 8.5 and 0.12 ionic strength. 0, approach to equilibrium from glucose 6-phosphate (G-6-P) ; 0, from fructose B-phosphate (F-6-P). The solid line was drawn as the best visual fit to the experimenta points.
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Issue of April 10, 1968 J. E. D. Dyson and E. A. Noltmann
binding of 6-phosphogluconate. Also, 6-phosphogluconate exists in the straight chain form, whereas the hexose 6-phosphates are almost entirely in the ring form when in solution. Thus, it is reasonable t,o assume that the e-amino nitrogen of lysine binds by protonation and subsequent hydrogen bonding to the ring oxygen of the hexose B-phosphates, thereby causing the ring to open prior to the isomerization reaction per se.
The participation of phosphoglucose isomerase in opening the hexose ring structure has also been postulated by Salas et al. (34) on the basis of their observation that the enzyme from yeast catalyzed the conversion of oc-glucose B-phosphate to &glucose 6-phosphate, but not the reverse. Their finding indicates that a particular positioning of the C-l hydroxyl group is necessary, possibly attained through binding to a particular group on the enzyme. Such a requirement would be in agreement with the highly selective stereospecificity of phosphoglucose isomerase, exemplified by the fact that it only labilizes a particular one of the two hydrogens on C-l of fructose 6-phosphate (38).
In attempting to formulate a mechanism for phosphoglucose isomerase, it is necessary to consider the possible binding of the phosphate group of the hexose B-phosphates. It is notable that all the competitive inhibitors so far found for phosphoglucose isomerase bear a terminal phosphate group. Tests have been carried out in this laboratory on possible inhibitory effects of free glucose and fructose, but no inhibition wm detectable. No in- hibition could be detected with fructose 1,6-diphosphate or glucose 1 ,6-diphosphate, although mannose B-phosphate, ri- bulose 5-phosphate, galactose B-phosphate, sorbitol6-phosphate, glucosamine 6-phosphate, 6-phosphogluconate, and erythrose 4-phosphate have been shown by various workers to be inhibitors (34, 39, 40). It is understandable that the diphosphate com- pounds do not inhibit, since substitution of a phosphate group on C-l would induce considerable steric hindrance. However, the fact that free glucose and fructose do not inhibit would appear to indicate some function in binding for the phospha$e group, since it is difficult to visualize steric requirements which would be met by the hexose 6-phosphat,es but not by the free sugars. Furthermore, phosphate has been reported to be a weak competi- tive inhibitor of phosphoglucose isomerase (34) ; this has been confirmed in this laboratory, and a K; of 5 x 1O-2 M has been determined. The exact manner in which the phosphate group affects binding is unknown, however, and this is indicated by the arrows in Fig. 14.
The proposed reaction sequence is as follows.
where ES denotes enzyme-substrate complex; G, glucose 6- phosphate; F, fructose B-phosphate; C, the straight chain form of the substrate; R, the ring form of the substrate; and X, the assumed enediol intermediate.
The proposed reaction mechanism of Fig. 14 is drawn for the conversion of glucose B-phosphate to fructose 6-phosphate and will be discussed for that direction. It will be understood, how- ever, that the reaction is fully reversible. In Complexes 2,3, and 4 of Fig. 14, the bond angles around C-2 of the substrate have been distorted in order to maintain the same relationship between the substrate molecule and the chemical groups participating in the catalytic reaction.
Complex 1: ??8,c,,-The enzyme-substrate complex is pro-
0 ESGFd
PHOSPHOGLUCOSE ISOMERASE
G-6-P
F-6-P
PHOSPHOGLUCOSE ISOMERASE
Q ES F(R)
@ ESocc,
FIG. 14. Proposed mechanism for the phosphoglucose isom- erase-catalyzed reaction. G-6-P, glucose F-phosphate; P-6-P, fructose 6-phosphate; other abbreviations are defined in the text.
duced initially by formation of a hydrogen bond between one of the hydrogens of the protonated c-amino group of lysine and the ring oxygen of the substrate. Following formation of the hydro- gen bond, the proton is transferred from the nitrogen to the ring oxygen; at the same time the hydrogen bond, which was orig- inally between t)he hydrogen and the oxygen, is transferred to between t,he hydrogen and the nitrogen. Protonation of the ring oxygen results in an electron shift, which opens the ring, with the formation of the straight chain form of the substrate.
Complex 2: EX o(c)-After ring opening, the hydrogen at- tached to C-2 of the substrate is abst’ract’ed by nucleophilic at- tack of the basic imidazole nitrogen? The pair of electrons which formed the bond between C-2 and the hydrogen is shifted, with the formation of a double bond between C-l and C-2, to give the assumed enediol anion intermediate.
Complex 8: EXcx,-Conversion of the enediol intermediate into the straight chain form of fructose 6-phosphate occurs by transfer of the hydrogen on the imidazole nitrogen to C-l of the substrate, the transition state perhaps having the form of a rr complex between the double bond and the hydrogen. Formation of the bond between C-l and the hydrogen is accompanied by electron shift,s which result in the loss of a prot’on from the oxygen on C-2 and the gain of a proton by the oxygen on C-l of the sub- strate.
9 Nucleophilic attack by a base to give an intermediate enediol was first suggested by Rose and O’Connell (1).
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1414 Kinetics and Mechanism o.f Phosphoglucose Isomerase Vol. 243, No. 7
Complex 4: BXFcc)-E’ollowing the formation of fructose 6- phosphate from the enediol intermediate, a ring structure is re- sumed by transfer of a pair of electrons from the oxygen-hydrogen bond to form a bond between the oxygen and C-2. On rupture of the oxygen-hydrogen bond, the hydrogen bond between the hydrogen and the nitrogen is transferred t,o between the hydro- gen and what, is now the ring oxygen. On formation of the bond between the oxygen and C-2, the ketosc oxygen picks up a prot,on from the solvent to complete the ring form of fructose 6-phos- phate, ESR(R).
A significant feature of this proposed dual function mech- anism for phosphoglucose isomerase is that it rationalizes the isomerization as occurring via an enediol intermediate, as gen- erally accepted, and supported by other experimental evidence (1, 38, 41). In addition, it takes into consideration the fact that the concentration of the straight chain forms of the hexose K-phosphates in solution is too low (34, 42) for them to be the true substrates and yet to account for Michaelis constants of lop4 hf with respect to total hexose G-phosphate (2). The pro- posed mechanism avoids the dilemma (34) of having to postulate that either only the open (1, 13, 43) or the ring forms (2) of the hexose B-phosphates act as substrates, and it appears to be in agreement with all the experimental facts available at the present time.
Acknowledgments-We wish t,o t#hank Dr. W. W. Cleland of the University of Wisconsin for making available his computer programs for processing enzyme kinetic data. We also grate- fully acknowledge discussions with Dr. V. Massey of the Uni- versit,y of Michigan on the interpretation of temperature-induced conformation changes and their effect on Arrhenius plots.
1.
2. 3. 4.
5.
G.
7.
8.
9.
10. 11.
REFERENCES
ROSE, I. A., AND O'CONNELL,E. L., J. Bid. Chem.,236, 3086 (19Gl).
HINES,M. C., AND WOLFE, R. G., Biochemistry, 2, 770 (1963). NOLTMANN, E. A., J. Biol. Chem., 239, 1545 (1964). NOLT~LZANN, E. A., AND CHATTERJEE, G. C., Fed. Proc., 25,
754 (1966). DYSON, J. E.. AND NOLTMANN, E. A., Abstracts ef the Pac<fic
Slope Biochemical Conference, Eugene, Oregon, -1966, p. i2. DYSON. J. E. D.. AND NOLTPIIANN. E,. A.. Ped. PTOC.. 26. 390
(196i). ' PELLER, L., AND AI,BERTY, R. A., J. Amer. Chem. Sot., 81,
5907 (1959). ALBER'~~, R. A., AND BLOOMFIELD, V., J. Biol. Chem., 233,
2804 (1963). BRUICE, T. C., AND SCHMIR, G. L., J. Amer. Chem. Sot., 81,
4552 (1959). LINDLEY, H., B&hem. J., 82, 418 (1962). KOSHI~AND, 2). E., JR., Advance. Enzymol., 22, 59 (1960).
12.
13. 14.
15.
16.
17.
18.
19.
20. 21.
22. 23. 24. 25.
26.
27.
28. 29. 30.
31.
32.
33.
34.
35.
36.
37.
38.
39. 40.
41. 42.
43.
KOSOJYER, E. M., diolecular biochemislry, McGraw-IIill Book Company, Inc., New York, 1962, p. 47.
ROSE, 1. A., Annu. 12ev. Biochem., 35, 23 (1966). NOLTMANN, E. A., iI1 S. P. COLO\\-ICK AND N. 0. KA~LAK
(Editors). Methods in enzumoloou. vol. IS. Academic Press. i&w Y&l;, 1966, p. 557. " ""' '
DYSON, J. E. U., Ph.1). dissertation, University of California, Itiverside, 1966.
HORECKER, l3. L., AND KORXBERG, A., J. Biol. Chem., 175, 385 (1948).
DYSON, J. E., AILD NOLI'MANN, E. A., Ad. Biochem., 11, 362 (1965).
ALLIERTY, K. A., SMITH, It. M., AAD BOCK, R. X4., J. Bid. Chem., 193,425 (1951).
KAHANA, S. E., LOWRY, 0. H., SCHULZ, 2). W., P~ssoxx~au, J. V., AND CRA\V~ORD,E. J.,J. Zol. Chem., 235,2178 (1960).
IhXON, i\l., Biochem. J., 55, 170 (1953). RAUEI\T, II. M. (Editor), Biochemisches Taschenbuch, T’ol. 11,
Springer-Verlag, ISerlin, 1964, p. 69. I)IXON, M., Biochem. J., 55, 161 (1953). CLELAND, W. W., Xafure, 198, 463 (1X13). WILKINSON, G. N., Biochem. J., 80, 324 (1961). STEJNHARDT, J., AND BEY~HOK, S., in H. NEURA~'H (Editor),
The proteins, Vol. II, Ed. 2, Academic Press, New York, 1964, p. 139.
COHN, E. J., AKD EDS~LL, J. T., Proieins, amino acids, and
peplirles as ions and &polar ions, Reinhold Publishing Company, New York, 1943.
EDHALL, J. T., AND WYMAN, J., Biophqsical chemislry, I’d. I, Academic Press, New York, 1958, p. 464.
LINDLEY, II., Biochim. Biophp. Ada, 59, 235 (1962). TANFORD, C., ildvance. Protean Chem., 17, 69 (1962). CHATTERJEE, (:. C., AND NOLTMANN, E. A., Ezcrope. J. Bio-
them., 2, 9 (1967). BENESCH, It. E., AND HE~ES('H, It., J. Amer. Chem. Sot., 77,
5877 (1955). CHAT'L'ERJEE. G. C., AND NuI,,~M.~NN, E. A.. J. Hiol. Chem.,
242, 3440 (1967). GEX;\I*ATER. n. M. J.. E~ANSHA\Z.. E. 11.. MIIRTIN. It. b:.. AND
PARR, C:W., Biochem. J., 97: 121, (1665). ' ' SALAS, &I., VII%ELA, E., AND SOLS, A.! J. Biol. Chem., 240,
561 (1965). Dixon-, M., ANU W~un, E. C., Enqmes, Ed. 2, Academic
Press, New York, 1964, p. 158. KISTIAKOTVSKY, G. B., AND LUMRY, le., J. Amer. Chem. Sot.,
71, 2006 (1949). MASSEY, V., CVRTI, u., AND GANTRER, H., J. Bid. Chem.,
241, 2347 (1966). TOPPER, Y. J., J. Biol. Chem., 225,419 (1957) ; in P. D. BOYER,
H. LARDY, AND K. MYRB~~CK (Editors), The enzymes, Vol. 5, Acadetnic Press, New York, 1961, p. 429.
PARR, C. W., Xalure, 178, 1401 (1956). GR.?zI, E., DE FLORA, A., AND PONTREMOLI, S., Biochem.
Biophys. Res. Commltn., 2, 121 (IUGO). ROSE. I. A.. Brookhaven Sump. Biol.. 15. 293 11962) Los, J. M., SIMPSON, L. B.,“A&I WI&&, K., J. A&r. Chem.
Sot., 78, 1564 (1956). PARR, C. W., AND WHI~'TAKER,M., Niochem.J., 74,31~ (1960).
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from

John E. D. Dyson and Ernst A. NoltmannIN A PROPOSED DUAL FUNCTION MECHANISM
Phosphoglucose Isomerase: PARTICIPATION OF HISTIDINE AND LYSINE The Effect of pH and Temperature on the Kinetic Parameters of
1968, 243:1401-1414.J. Biol. Chem.
http://www.jbc.org/content/243/7/1401Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/243/7/1401.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on March 25, 2018
http://ww
w.jbc.org/
Dow
nloaded from