ter signal tran tem - dspace.mit.edu
TRANSCRIPT

I now-,mp-MI1, ip a 70 JJLtot.
wo,
02AT TjQv
M
,00 VOW
TOT&TY.-
.
I A
ter Signal Tran tem
IPA

396 RLE Progress Report Number 136

Chapter 1. Signal Transmission in the Auditory System
Chapter 1. Signal Transmission in the Auditory System
Academic and Research Staff
Professor Lawrence S. Frishkopf, Professor Nelson Y.S. Kiang, Professor William T. Peake, ProfessorWilliam M. Siebert, Professor Thomas F. Weiss, Dr. Bertrand Delgutte, Dr. Donald K. Eddington, Dr. DennisM. Freeman, Dr. John J. Guinan, Jr., Dr. William M. Rabinowitz, Dr. John J. Rosowski, Joseph Tierney, MarcA. Zissman
Visiting Scientists and Research Affiliates
Dr. Jennifer R. Melcher, Dr. Sunil Puria, Dr. Jay T. Rubinstein,' Dr. Devang M. Shah
Graduate Students
C. Cameron Abnet, Henry E. Chung, C. Quentin Davis, Scott B.C. Dynes, Michael P. McCue, Lisa F. Shatz,Konstantina Stankovic, Susan E. Voss
Technical and Support Staff
Janice L. Balzer, Mark R. Nilsen, Michael E. Ravicz,Zhu
1.1 Introduction
Research on the auditory system is carried out incooperation with two laboratories at the Massachu-setts Eye and Ear Infirmary (MEEI). Investigationsof signal transmission in the auditory system involvethe Eaton-Peabody Laboratory for Auditory Physi-ology, whose long-term objective is to determinethe anatomical structures and physiological mech-anisms that underlie vertebrate hearing and toapply this knowledge to clinical problems. Studiesof cochlear implants in humans are carried out atthe MEEI Cochlear Implant Research Laboratory.Cochlear implants electrically stimulateintracochlear electrodes to elicit patterns of auditorynerve fiber activity that the brain can learn to inter-pret. The ultimate goal for these devices is toprovide speech communication for the profoundlydeaf.
1.2and
Signal Transmission in the ExternalMiddle Ear
The goal of our work is to understand the relation-ship between the structures of the external andmiddle ear and their functions.
Frank J. Stefanov-Wagner, David A. Steffens, Meng Y.
1.2.1 Structure-Function Relations inMiddle Ears
SponsorNational Institutes of Health
Grant R01-DC-00194-11
Project StaffDr. John J. Rosowski, Professor William T. Peake,Michael E. Ravicz, David A. Steffens
The middle-ear is a key link in the hearing process.In normal hearing, sound signals are delivered tothe inner ear by the middle ear. Disease, age, andinjury can degrade the performance of the middleear. In part of our work, we investigate the relation-ship between the structures of the middle-ear airspace and tympanic-membrane and the function ofthe middle ear. We are making joint anatomical andphysiological measurements in a series of rodentand cat species with different tympanic membraneand middle-ear air-space morphologies. We willalso use similar methods in cats to investigatewhether the response of the cochlea is completelydetermined by the difference in pressure across thetwo cochlear windows.
In the last year, we have moved towards our goal inthe following ways:
1 Research Affiliate, Massachusetts Eye and Ear Infirmary, Boston, Massachusetts.
397

Chapter 1. Signal Transmission in the Auditory System
1. We have analyzed our measurements onmiddle-ear cavity and tympanic membranefunction and incorporated the results into apaper presenting new measurements of themiddle-ear input impedance in cats2 and into amodel of the mammalian external and middleear.3 These analyses support the theory thatthe primary effect of middle-ear cavities is con-sistent with a "series" model. The cavitiesimpede the volume-velocity of sound at thetympanic membrane but have little effect onhow the membrane and ossicles respond for agiven volume velocity. An additional analysisof anatomical and physiological measurements,made on many mammalian species, points outthat different species have greatly differentmiddle-ear cavity impedances and that the rela-tive magnitudes of the impedance of the cavityand the impedance of the rest of the ear varieswidely among mammals. 4
2. We have taken the "series" model and our dataa step further and analyzed the effect of vari-ations in middle-ear volume on auditory thresh-olds in a desert rodent (the Mongolian gerbil).5
This analysis suggests that the middle-earcavity in a gerbil plays a large role in limitinglow-frequency auditory thresholds even thoughthe cavity has a volume larger than the braincase. This conclusion is contrary to the sug-gestion that the large middle-ear cavities ofthese and other specialized desert rodentshave evolved to permit maximum sensitivity tolow-frequency sound.
3. A series of new measurements tested the"classic" assumption that the inner earresponds to the sound-pressure differencebetween the oval and round window.6 Theexternal and middle ears of cats were preparedso that we could independently control themagnitude and phase of the sound pressure atthe two cochlear windows. The cochlear
2 T.J. Lynch Ill, W.T. Peake, and J.J. Rosowski, "MeasurementsAcoust. Soc. Am., forthcoming.
potential, measured with a round-windowelectrode, was our indicator of cochlearresponse. The potential was determined withsound stimuli presented at both or eithercochlear window. The results indicate that thedifference in sound pressure at the windows isclearly the primary stimulus to the cochlearpotential.
The results of our work have direct significance toour understanding of human disease, animalethology, and the basic hearing process. Chronicand acute otitis media are very common causes ofhearing dysfunction. One significant effect of thesediseases is a reduction in the air volume of themiddle ear. This reduction can be transientlycaused by the accumulation of fluid or more perma-nently produced by the accumulation of debriswithin the middle ear or actual alterations in thebony walls of the cavity. Our work will enable aclearer understanding of the role of the middle-earair spaces in both normal and diseased ears.
The ethological issues we are testing concern therole of enlarged middle-ear air spaces. Our workon a number of rodent species and a wide selectionof cat species will test the popular idea that largemiddle-ear air spaces represent an adaptation toarid environments.
Our work on specification of the effective stimulusto the inner ear also has direct clinical conse-quences in that surgeons routinely manipulate thepressure difference between the cochlear windowswhen they reconstruct severely diseased middleears. Our new measurements support the efficacyof such procedures. Furthermore, the results of ourmeasurements have implications for our funda-mental understanding of inner ear mechanisms.Any indication that the cochlea is sensitive to stimuliother than the difference in sound pressure at thecochlear windows will require basic changes incochlear theories.
of the Acoustic Input Impedance of Cat Ears: 10 Hz to 20 kHz," J.
3 J.J. Rosowski, "Models of External and Middle-Ear Function," in The Springer Handbook on Auditory Research, Volume 6: AuditoryComputation, eds. H.L. Hawkins, T.A. McMullen, A.N. Popper, and R.R. Fay (New York: Springer-Verlag, forthcoming).
4 J.J. Rosowski, "Outer and Inner Ear," in The Springer Handbook on Auditory Research, Volume 4: Comparative Mammalian Hearing,eds. A.N. Popper, and R.R. Fay (New York: Springer-Verlag, forthcoming).
5 M.E. Ravicz and J.J. Rosowski, "The Effect of Middle-Ear Cavity Size on Acoustic Power Flow in the Ear of the Mongolian Gerbil," inAbstracts of the Seventeenth Midwinter Meeting of the Association for Research in Otolaryngology, February 6-10, 1994, p. 145.
6 S.E. Voss, J.J. Rosowski, and W.T. Peake, "Is the Pressure Difference Between the Oval and Round Windows the Stimulus forCochlear Responses?" in Abstracts of the Seventeenth Midwinter Meeting of the Association for Research in Otolaryngology, Feb-ruary 6-10, 1994, p. 87.
398 RLE Progress Report Number 136

1.2.2 Basic and Clinical Studies ofMiddle-Ear Function
SponsorNational Institutes of Health
Grant P01-DC00119 Sub-Project 1Grant F32-DC00073-2
Project Staff
Dr. John J. Rosowski, Professor William T. Peake,Dr. Sunil Puria, Michael E. Ravicz
Our goal of understanding the relationship betweenthe structure and function of the auditory peripheryincludes defining the effect of middle and externalear pathologies on auditory function. To helpachieve this goal, we have continued measure-ments of function in human ears using temporal-bones donated at the time of death, and we haveapplied the models we determined from our animalwork to questions concerning middle-earpathologies and treatments.
Dr. Puria has led a project to measure the pressuretransfer function of the human middle ear.7 Theexcised temporal bones were prepared to allowreproducible placement of a calibrated soundsource and microphone in the ear canal togetherwith a miniature hydrophone in the cochlearvestibule. The preparation procedure included theplacement of "flush" tubes-one near the roundwindow, the other in the vestibular canals-to keepthe cochlea fluid filled. These measurementssuggest that the human middle ear acts as a band-pass pressure amplifier with a maximum gain ofabout 20 dB between 1 and 5 kHz. At lower fre-quencies, the gain is roughly proportional to fre-quency; at higher frequencies the gain decreasesas frequency increases.
With the help of our clinical associates at the Mass-achusetts Eye and Ear Infirmary, we haveexpanded our analysis of previous models of dis-eased and reconstructed middle ears. The resultsof these analyses were presented at two confer-ences8 where we suggested that small modifica-tions in current surgical procedures could lead to
Chapter 1. Signal Transmission in the Auditory System
improved hearing results. These modificationsincluded: stapes prostheses with larger areas,maintaining a volume of at least 0.5 cc for thereconstructed middle-ear cavity, and stiffer grafts forisolating the round window from sound.
1.3 Cochlear Mechanisms
SponsorNational Institutes of Health
Contract P01-DC00119Grant R01-DC00238Gramt R01-DC00473
Project Staff
ProfessorFreeman,Henry E.
Thomas F. Weiss,Dr. Devang M. Shah, C.
Chung, C. Quentin Davis,
Dr. Dennis M.Cameron Abnet,Lisa F. Shatz
Our goal is to study the mechanisms by which themotions of macroscopic structures in the inner earproduce motions of the mechanically sensitive hairbundles of sensory receptor (hair) cells. Becauseof its strategic location, the tectorial membranemust play an important role in the mechanical stim-ulation of hair bundles. However, there have beenfew direct observations of the tectorial membrane,and its critical properties remain obscure.
Previously, we have measured osmotic responsesto isosmotic solutions of different sodium,potassium, and calcium concentration in the iso-lated chick tectorial membrane. That work hasbeen submitted for publication.9 However, the struc-ture and biochemical composition of the tectorialmembrane shows considerable diversity acrossspecies. Therefore, during the past year we haveinvestigated the physicochemical properties oftectorial membranes isolated from the lizard andmouse cochlea using the same techniques. Bas-ically, in this procedure we dissect the tectorialmembrane from the cochlea, place it in a chamberthrough which solutions of known composition areperfused. The tectorial membrane is decorated withmicroscopic latex beads whose three-dimensionalmotions in response to solution changes are meas-
7 S. Puria, J.J. Rosowski, and W.T. Peake, "Middle-Ear Pressure Gain in Humans," in Biophysics of Hair Cell Sensory Systems, eds.H. Duifhuis, J.W. Horst, P. van Dijk, and S. van Netten (River Edge, New Jersey: World Scientific Publishing Co., 1993).
8 S.N. Merchant, J.J. Rosowski, and M.E. Ravicz, "Acoustical Aspects of Type IV and V Tympanoplasty," paper presented at theMeeting of the American Academy of Otolaryngology, Head and Neck Surgery, Minneapolis, Minnesota, October 3-8, 1993; J.J.Rosowski, "Analyses of Middle-Ear Reconstructive Techniques," paper presented at the Meeting of the American Oto-NeurologicSociety, Minneapolis, Minnesota, October 2, 1993.
9 D.M. Freeman, D.A. Cotanche, F. Ehsani, and T.F. Weiss, "The Osmotic Response of the Isolated Tectorial Membrane of the Chickto Isosmotic Solutions: Effect of Na , K+, and Ca2+ Concentration," submitted to Hear. Res.
399

Chapter 1. Signal Transmission in the Auditory System
ured automatically with a system consisting of acompound microscope, CCD camera, videodigitizer, and workstation. Results for all threespecies (chick, lizard, and mouse) show thatincreasing calcium concentration from the lowvalues (20 pmol/L) typical for the solution thatbathes the tectorial membrane in situ (endolymph)to the higher concentrations (2 mmol/L) found innormal extracellular fluids causes the tectorial mem-brane to shrink with a time course of about fiveminutes. The shrinkage is largely reversed whencalcium concentration is lowered but with a slowertime course (tens of minutes).
Tectorial membranes from all three species alsoswell when isosmotic, high-sodium solutions aresubstituted for high-potassium solutions. However,the magnitude of this response is much greater forthe chick (mean of 135 percent) than for either themouse (mean of 14 percent) or the lizard (mean of10 percent). Swelling in high-sodium solutions is atleast partially reversed on return to high-potassiumsolutions. However, irreversible changes resultduring long-duration (60-minutes) exposures tohigh-sodium solutions that give rise to largeswelling responses.
Our results are consistent with the simple hypoth-esis that the tectorial membrane is a polyelectrolytegel with fixed ionizable charge groups that can bindions differentially. This picture of the tectorial mem-brane is similar to that for other connective tissues:the binding of ions modulates the fixed chargedensity which changes the concentration of counterions in the tissue and hence its water content.
The shrinkage and swelling of the tectorial mem-brane in response to changes in ion concentrationmay have implications for hearing. For example, anumber of factors are known to change ion compo-sition of the lymphs including acoustic overstimu-lation, anoxia, systemic administration of ethacrynicacid, or perfusion of the cochlea with ouabain, etc.Our results show that changes in the ionic composi-tion of the bath can lead to changes in the structureof the tectorial membrane. Changes in its structureare likely to cause changes in the delivery of themechanical stimulus to the hair cells and hence toaffect hearing.
1.3.1 Publication
Freeman, D.M., D.A. Cotanche, F. Ehsani, and T.F.Weiss. "The Osmotic Response of the IsolatedTectorial Membrane of the Chick to IsosmoticSolutions: Effect of Na+, K+, and Ca2+ Concen-tration." Submitted to Hear. Res.
1.4 Stimulus Coding in the AuditoryNerve and Cochlear Nucleus
SponsorNational Institutes of Health
Grant P01-DC00119Grant T32-DC00038P
Project Staff
Dr. Bertrand Delgutte, Dr. Peter A. Cariani
We are investigating the neural mechanisms under-lying auditory perception at the level of the auditorynerve and cochlear nucleus. In the past year, wehave focused on temporal patterns of neural dis-charge that convey pitch and speech information.These temporal patterns are analyzed by recordingthe responses of auditory nerve fibers and cochlearnucleus neurons in anesthetized cats. All-orderinterspike interval distributions (autocorrelationhistograms) are computed and the properties ofthese distributions are compared with psychophys-ical data.
Our previous results had shown that the pitch of acomplex tone corresponds to the most frequentinterspike interval in the auditory nerve for a widevariety of stimuli. Responses of cochlear nucleusunits to the same stimuli were recorded to deter-mine whether pitch might also be represented byinterspike interval patterns there. Virtually all singleunits in the cochlear nucleus that responded to thestimuli showed pitch-related intervals, and thesewere usually the most frequent in the distribution.Thus, cochlear nucleus units might convey pitchinformation to more central auditory stationsthrough an interspike interval code.
We have continued our investigation of the neuralmechanisms underlying the perceptual segregationof concurrently-presented vowels. Human listenersidentify pairs of concurrent vowels better when theydiffer in fundamental frequency (FO) than when theyhave the same FO. Previously we had found thatthe constituents of a vowel pair could be identifiedfrom the aggregate interspike interval distributionsof the auditory nerve with an accuracy comparableto human perceptual judgements. During the pastyear, we seeked to explain how vowel identificationmight be improved for pairs with different FOswithout requiring an explicit determination of FO.We found that differences in FO create changingalignments in the onsets of vowel periods, and thatthese changing alignments create transient changesin interspike interval distributions of auditory-nervefibers. One possibility is that these changes allow
400 RLE Progress Report Number 136

the auditory system to perform a spectral analysisover relatively short (5-10 ms) time windows andintegrate these analyses over time to identify thevowel constituents. Along these lines, we devel-oped a new algorithm for identifying the constitu-ents of a double vowel from running interspikeinterval distributions. This algorithm is consistentwith human perceptual data showing effects ofperiod asynchronies on the identification of concur-rent vowels.
1.4.1 Publication
Delgutte B., P. Cariani and M.J. Tramo, "Neuro-physiological Correlates of the Pitch of ComplexTones." J. Acoust. Soc. Am. 93: 2293 (1993).
1.4.2 Binaural Interactions in AuditoryBrainstem Neurons
Sponsor
National Institutes of HealthGrant P01-DC00119
Project Staff
Dr. Bertrand Delgutte, Dr. John J. Guinan, Jr., Dr.John J. Rosowski
Several acoustic cues are important for sound local-ization, including interaural time (ITD), interaurallevel (ILD) differences, and spectral shape. In col-laboration with Dr. T.C.T. Yin and his colleagues atthe University of Wisconsin in Madison, we arestudying the relative contribution of these cues tothe directional sensitivity of neurons in the inferiorcolliculus (IC). The activity of single units wasrecorded in anesthetized cats in response to "virtualspace" (VS) stimuli delivered through closedacoustic systems. These stimuli were generated bypassing broadband noise through digital filters con-structed from head-related transfer functions meas-ured in one cat. We focused on high-CF (> 5 kHz)units, which are likely to be sensitive to spectralfeatures.
The sensitivity of neurons to stimuli whose virtualposition varied along the horizontal and verticalplanes generally resembled that found in previousfree-field studies. For most units, responses tomonaural VS stimuli were less directional thanthose for binaural stimuli, suggesting that binauralinteractions were important for the directionalresponse of these neurons. The type of binauralinteractions (e.g., EE, El, or mixed) determined withVS stimuli was consistent with that found forbroadband noise.
Chapter 1. Signal Transmission in the Auditory System
We also generated modified VS stimuli in whichone or more of the localization cues (ITD, ILD, orspectrum) were set constant for all locations. Forexample, "O-ITD" stimuli were constructed bydelaying the waveform in the leading ear to set theITD to zero for every azimuth. "Delta-ITD" stimuliwere created by taking the waveforms for the posi-tion directly in front, then introducing the appro-priate ITD for each azimuth. Results suggest that,for most of these high-CF units, ILD is the mostpotent cue, followed by spectral shape, then byITD. These results are consistent with human psy-chophysical experiments showing the dominance ofILD cues for high-frequency stimuli. At a moregeneral level, our results demonstrate the useful-ness of virtual-space techniques for bridging thegap between free-field studies of the directionalsensitivity of neurons and dichotic studies of ITDand ILD sensitivity.
1.4.3 Publication
Delgutte, B., P.X. Joris, R.Y. Litovsky, and T.C.T.Yin. "Different Acoustic Cues Contribute to theDirectional Sensitivity of Inferior-ColliculusNeurons as Studied with Virtual-Space Stimuli."Abstracts of the 17th Midwinter ResearchMeeting of the Association for Research inOtolaryngology, St. Petersburg, Florida, Feb-ruary 6-10, 1994.
1.4.4 Electrical Stimulation of the AuditoryNerve
Sponsor
National Institutes of HealthGrant P01-DC00361
Project Staff
Dr. Bertrand Delgutte, Scott B.C. Dynes
We are studying physiological mechanisms of elec-trical stimulation of the cochlea because this infor-mation will help design improved processingschemes for multiple-channel cochlear implants.During the past year, we have studied auditory-nerve fiber correlates of interactions observedpsychophysically when pulsatile electric stimuli areapplied in rapid succession. We measured theresponses of auditory-nerve fibers in anesthetizedcats for pairs of monophasic pulses separated byshort intervals. Results show that neural inter-actions between successive current pulses occur onthree different time scales:
401

Chapter 1. Signal Transmission in the Auditory System
1. The shortest interactions occur when the firstpulse in a pair is subthreshold. For cathodicfirst pulses, the threshold for following cathodicpulses was decreased, while anodic first pulsesincreased the threshold of following cathodicpulses. These results are qualitatively con-sistent with viewing the first pulse as leaving aresidual charge on the neural membrane. Thetime constant of these capacitive effects is100-200 usec. These subthreshold interactionsare likely to play an important role in contin-uous interleaved sampling (CIS) processors forcochlear implants because most pulses areprobably subthreshold in this situation, so thatelicitation of a spike would require interactionover multiple pulses. Particularly interesting isthe result that subthreshold interactions forbiphasic pulses similar to those used in CISprocessors cannot be predicted from simpleideas about charge integration by the neuralmembrane. On the other hand, these inter-actions are consistent with psychophysical datashowing sensitization for biphasic pulses pre-ceded by either anodic/cathodic orcathodic/anodic pulses. Thus, the neural andpsychophysical data are consistent with eachother, but both are at variance with current the-oretical conceptions of neural excitation.
2. When the first pulse was suprathreshold, thethreshold for the second pulse was found todecrease with increasing interpulse delay, asexpected from the refractory properties ofnerve fibers. The decrease was such that at 2msec interpulse delay, the threshold for thesecond pulse averaged about 5 dB greaterthan the threshold for a single pulse. Thus,these interactions due to suprathreshold pulseslast longer than subthreshold interactions.These suprathreshold interactions may notseverely limit the performance of CIS proces-sors providing that there is a sufficient pool ofnerve fibers that discharge at different times(volley principle).
3. In addition to interactions between two succes-sive pulses, further results provide evidence forthe existence of long-term interactions occur-ring cumulatively over multiple pulses.Because these multipulse interactions have notbeen systematically characterized, it is difficultat this point to assess their significance for CIS
10 S.R. Wiener-Vacher, J.J. Guinan, Jr., J.B. Kobler, and B.E. Norris,An Intracellular Labeling Study", submitted to J. Comp. Neurol
processors. We plan to examine these inter-actions in more detail during the next year.
1.4.5 Publications
Dynes, S.B.C., and B. Delgutte. "Neural Responseto Nonsimultaneous Electrical Stimuli: Physiolog-ical Results." Conference on ImplantableAuditory Prostheses, Smithfield, Rhode Island,July, 1993.
Dynes, S.B.C., and B. Delgutte. "Temporal Mech-anisms of Auditory-Nerve Response to MultipleElectric Pulses." Abstracts of the 17th MidwinterResearch Meeting of the Association forResearch in Otolaryngology, St. Petersburg,Florida, February 6-10, 1994.
Rubinstein, J.T., and S.B.C. Dynes. "Latency,Polarity, and Refractory Characteristics of Elec-trical Stimulation: Models and Single-Unit Data."Abstracts of the 16th Midwinter ResearchMeeting of the Association for Research inOtolaryngology, St. Petersburg, Florida, Feb-ruary 6-11, 1993, p. 76.
1.5 Interactions of Middle-Ear Musclesand Olivocochlear Efferents
Sponsor
National Institutes of HealthContract P01 DC00119
Project Staff
Dr. John J. Guinan, Jr.
Our aim is to determine the actions and interactionsof acoustically elicited middle-ear muscle reflexesand olivocochlear efferent reflexes.
During the past year, we have analyzed data andprepared for publication our results on theinnervation of the stapedius muscle. 10 In a previousstudy using intracellular labeling of physiologicallyidentified stapedius motoneurons, we showed thatthere is functional spatial segregation in thestapedius motoneuron pool." In the same animals,we have now traced each sufficiently labeled axon
"Motoneuron Axon Distribution in the Stapedius Muscle of the Cat:
11 S.R. Vacher, J.J. Guinan, Jr., and J.B. Kobler, "Intracellularly Labeled Stapedius-Motoneuron Cell Bodies in the Cat Are SpatiallyOrganized According to Their Physiologic Responses," J. Comp. Neurol. 289: 401-415 (1989).
402 RLE Progress Report Number 136

into the stapedius muscle to determine whether asimilar functional spatial segregation is presentwithin the muscle. Ten labeled axons ofphysiologically identified stapedius motoneuronswere visible in the facial nerve and five could betraced to endplates within the stapedius muscle. Inone case, a stapedius motoneuron innervated onlya single muscle fiber; we think that this is the firstdocumented case of such remarkably fine-grainedmotor innervation. Overall, there were 39 observedbranches from the five axons (we may have missedothers). This indicates an average innervation ratio( 7.8) which is much higher than that obtainedfrom present estimates of the numbers of stapediusmotoneurons and muscle fibers. Muscle fibersinnervated by a single axon were spread over awide area in the muscle, suggesting that spatialsegregation in the stapedius muscle is unlikely.Thus, the reasons for the central functional spatialsegregation in the stapedius motoneuron pool aremore likely related to organizing factors which origi-nate in the brain rather than to organizing factorswhich originate in the muscle.
1.5.1 Publication
Wiener-Vacher, S.R., J.J. Guinan, Jr., J.B. Kobler,and B.E. Norris. "Motoneuron Axon Distributionin the Stapedius Muscle of the Cat: An Intracel-lular Labeling Study." Submitted to J. Comp.Neurol.
1.6 Cochlear Efferent System
Sponsor
National Institutes of HealthGrant 2RO1 DC00235
Project Staff
Michael P. McCue, Konstantina Stankovic
Our aim is to understand the physiological effectsproduced by efferents in the mammalian inner earincluding medial olivocochlear efferents which termi-nate on outer hair cells and lateral efferents whichterminate on auditory-nerve fibers.
Chapter 1. Signal Transmission in the Auditory System
During the past year, we have analyzed data andprepared for publication results on our newly-discovered class of vestibular primary afferentneurons, fibers which respond to sounds at moder-ately high sound levels.12 Like their cochlearhomologues, these vestibular afferent fibers receiveefferent projections from brain-stem neurons. Wehave explored efferent influences on the back-ground activity and tone-burst responses of theacoustically-responsive vestibular afferents. Shock-burst stimulation of efferents excited acoustically-responsive vestibular afferents; no inhibition wasseen. A fast excitatory component built up within100-200 ms of shock-burst onset and decayed witha similar time course at the end of each shockburst. During repeated 400 ms shock bursts at 1.5s intervals, a slow excitatory component grew over20-40 s and then decayed, even though the shockbursts continued. Efferent stimulation excitedacoustically-responsive vestibular afferents withoutappreciably affecting their sound-evoked responses.This provides strong evidence that the excitation isdue to efferent synapses on afferent fibers insteadof efferent synapses on hair cells. Efferent stimu-lation enhanced the tone-induced within-cycle mod-ulation of discharge rate (i.e., increased the a.c.gain) without changing the degree of phase lockingto low frequency tones as measured by the syn-chronization index; i.e., there was little or noimprovement in the bidirectionality (linearity) ofnerve fiber output. The acoustically-responsivevestibular afferents provide a mammalian model forstudying purely excitatory efferent effects in a haircell system. Anatomically, these vestibular efferentsynapses resemble lateral olivocochlear efferentsynapses on cochlear nerve fibers, which suggeststhat lateral efferents may have an excitatory effect.A paper describing these results has been sub-mitted. 13
During the past year, we have begun work on aproject to compare efferent-evoked effects onauditory-nerve fibers with different spontaneousrates. We have implemented and done preliminaryexperiments with a paradigm in which auditory-nerve-fiber rate versus sound level functions withand without efferent excitation are obtained withrandomized presentation of both the sound leveland the presence of efferent stimulation.
12 M.P. McCue, and J.J. Guinan, Jr., "Acoustic Responses from Primary Afferent Neurons of the Mammalian Sacculus,." Assoc. Res.
Otolaryngol. Abstr. 16: 33 (1993).
13 M.P. McCue, and J.J. Guinan, Jr., "Influence Of Efferent Stimulation On Acoustically-Responsive Vestibular Afferents in the Cat,"submitted to J. Neurosci.
403

Chapter 1. Signal Transmission in the Auditory System
J.J. Guinan, Jr. "AcousticPrimary Afferent Neurons ofSacculus." Assoc. Res.16: 33 (1993).
McCue, M.P., and J.J. Guinan, Jr. "Acoustically-Responsive Fibers in the Vestibular Nerve of theCat." Submitted to J. Neurosci.
McCue, M.P., and J.J. Guinan, Jr. "Influence OfEfferent Stimulation On Acoustically-ResponsiveVestibular Afferents in the Cat." Submitted to J.Neurosci.
1.7 Cochlear Implants
Sponsor
National Institutes of HealthGrant PO1-DC00361Contract NO1-DC2-2402
Project Staff
Dr. Donald K. Eddington, Dr. William M. Rabinowitz,Dr. Jay T. Rubinstein, Joseph Tierney, Marc A.Zissman
1.7.1 Project A: Models of Current Spreadand Nerve Excitation During IntracochlearStimulation
Project Staff
Dr. Donald K. Eddington, Dr. Jay T. Rubinstein
The basic function of a cochlear prosthesis is toelicit patterns of activity on the array of survivingauditory nerve fibers by stimulating electrodes thatare placed in and/or around the cochlea. By modu-lating patterns of neural activity, these devicesattempt to present information that the implantedsubject can learn to interpret. The spike activitypatterns elicited by electrical stimulation depend onseveral factors: the complex, electrically heteroge-neous structure of the cochlea, geometry andplacement of the stimulating electrodes, stimuluswaveform, and distribution of excitable auditorynerve fibers. An understanding of how thesefactors interact to determine the activity patterns is
1.6.1 Publications
McCue, M.P., andResponses fromthe MammalianOtolaryngoL Abstr.
404 RLE Progress Report Number 136
fundamental to designing better devices and inter-preting the results of experiments involvingintracochlear stimulation of animal and human sub-jects. As a first step toward this understanding, thegoal of this project is to (1) construct a softwaremodel of the cochlea that predicts the distribution ofpotential produced by the stimulation of arbitrarilyplaced, intracochlear electrodes and (2) use thesepotential distributions as inputs that drive models ofauditory nerve fibers.
This year, we refined our nonlinear, single-unitmodel to include a passive, myelinated cell body(see figures 1 and 2). Varying the membrane capa-citance and resistance to simulate variations in themyelination of the cell body produces profoundeffects on conduction. Without myelination, spikeswill not propagate across the cell body unlessvoltage-sensitive sodium channels are present inthe soma. With increasing myelination, spikesconduct in the orthograde direction but notretrograde (see figures 3 and 4). A model cell bodywith myelin thickness comparable to that of theinternode propagates spikes in both directions.These simulations demonstrate the biophysical fea-sibility of physiologically observed, somatic spikerectification and implicate the anatomic asymmetryof the nodes of Ranvier adjacent to the soma as apossible mechanism.
A multichannel stochastic model of the mammaliannode of Ranvier has also been developed. Thismodel represents N sodium channels in parallelwith a membrane capacitance and leakage resist-ance. Each channel consists of a single-channelconductance obtained from the patch-clamp andfluctuation literature and a gate controlled by threem (activation) particles and one h (inactivation) par-ticle. Particle open and closed times are random(exponential distribution) with mean obtained fromthe reciprocal of the particle's rate constant. Thusthe open/closed time probability density functionsare time-varying. The model has been tested bysubstituting Frankenhauser-Huxley kinetics andmembrane parameters for the mammalian data andcomparing the relative spread of threshold (RS)predicted by the model with that measured forperipheral amphibian nodes. The experimentaldata are very closely approximated by the simu-lations. This is the first direct numerical verificationthat the microscopic sodium channel fluctuationsare sufficient to account for the macroscopic fluctu-ations of threshold to electrical stimulation.
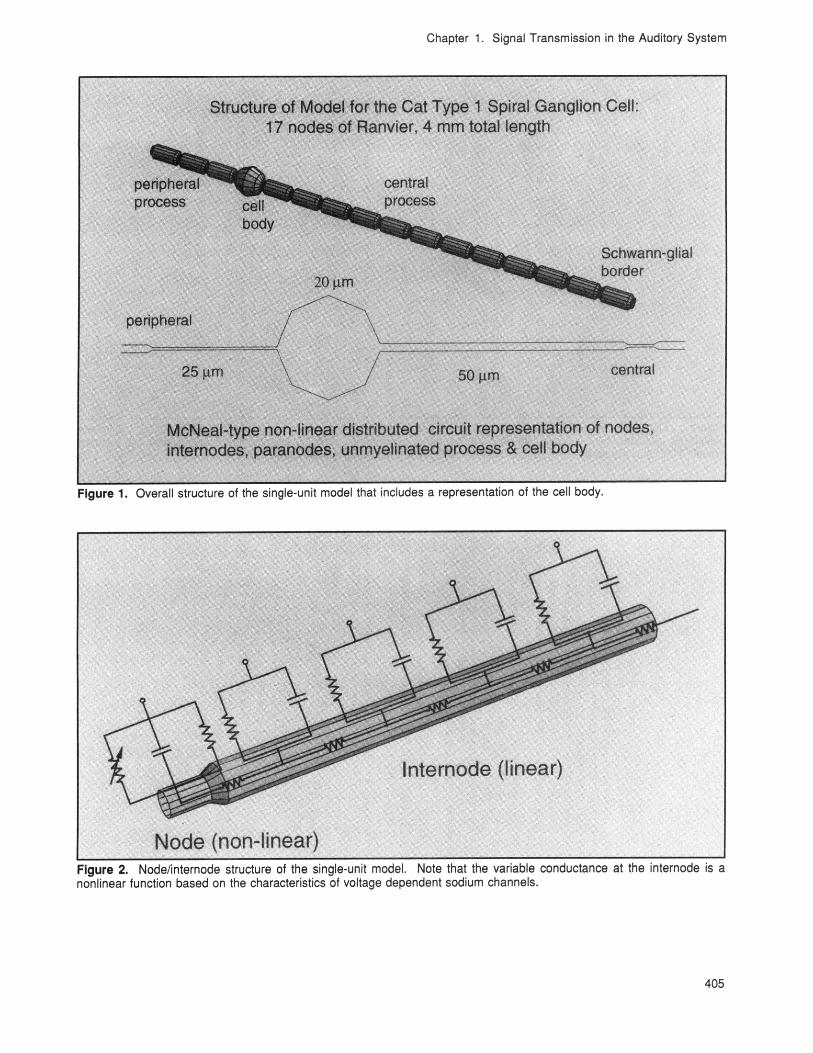
Chapter 1. Signal Transmission in the Auditory System
Figure 1. Overall structure of the single-unit model that includes a representation of the cell body.
Figure 2. Node/internode structure of the single-unit model. Note that the variable conductance at the internode is anonlinear function based on the characteristics of voltage dependent sodium channels.
405

Chapter 1. Signal Transmission in the Auditory System
Figure 3. Computed membrane potential as a function of time for the pictured model nodes. The stimulating electrode'sposition is indicated by the black oval and the cathodic, monophasic stimulus waveform is shown below the picturedmodel. Note that the spike conducts across the cell body in the orthograde direction.
Figure 4. Computed membrane potential as a function of time for the pictured model nodes. The stimulating electrode'sposition is indicated by the black oval and the cathodic, monophasic stimulus waveform is shown below the picturedmodel. Note that even though the model parameters are the same used in the model of figure 3, retrograde conductionis blocked at the cell body.
406 RLE Progress Report Number 136

Chapter 1. Signal Transmission in the Auditory System
1 1
0
I-
ca -2
0
-50 Model (ac-ca)I. -4
-6-6
0 200 400 600 800 1000
Delay (us)
Figure 5. Forward masking results for an anodic-phase-first, biphasic masker followed by a cathodic-phase-first,biphasic probe (ac-ca). Probe threshold is plotted in dB relative to the nonmasked threshold and as a function of thedelay between the masker and the probe. The subject results are represented by the symbols and the single-unit modelpredictions by the solid line.
1.7.2 Project B: Psychophysics ofIntracochlear Electrical Stimulation
Project StaffDr. Donald K. Eddington
One goal of our psychophysical studies is toprovide insight into the mechanisms that limitdevice effectiveness. Using a masking paradigm,we demonstrated in previous years that electricalstimuli delivered simultaneously by twointracochlear electrodes produce strong interactionsand that the strength of these interactions acrosssubjects is negatively correlated (r=-0.8) with theirspeech reception scores. Processing systems thatinterleave stimuli in time reduce these interactionsand produce improved speech reception in manysubjects\ul\d. This year, we focused on measuresof forward masking to investigate the strength ofnonsimultaneous interactions.
The electrical stimuli used in the forward maskingexperiments were single, charge-balanced biphasicpulses (phase duration 50us) presented to thesame intracochlear electrode. The masker ampli-
tude was constant at -2 dB below the unmaskedthreshold with the anodic phase first and cathodicphase second (ac). The following probe stimuluswas presented with cathodic phase first (ca), and itsamplitude was varied using a two-down, one-upadaptive procedure in a three alternative, forcedchoice context to measure threshold. For this ac-camasker-probe condition, the single-unit model pre-dictions are consistent with the subject data (figure5). The subthreshold masker reduces threshold byas much as 6 dB for a 100 us masker/probe delay.This effect diminishes with increased delay and isnegligible for delays longer than 800 us.
The data plotted in figure 6 are from the samesubject using a cathodic-phase-first, anodic-phase-second masker. In this condition (ca-ca), themasker tends to decrease threshold less than theac-ca case and its effect lasts longer than 3 ms.Note that the single-unit model's prediction for theca-ca case is not consistent with the psychophys-ical results. This inconsistency will be explored infuture single-unit studies in a cat. Because thenonsimultaneous interactions are significant, we arebeginning to explore stimulation strategies that willminimize their impact.
407
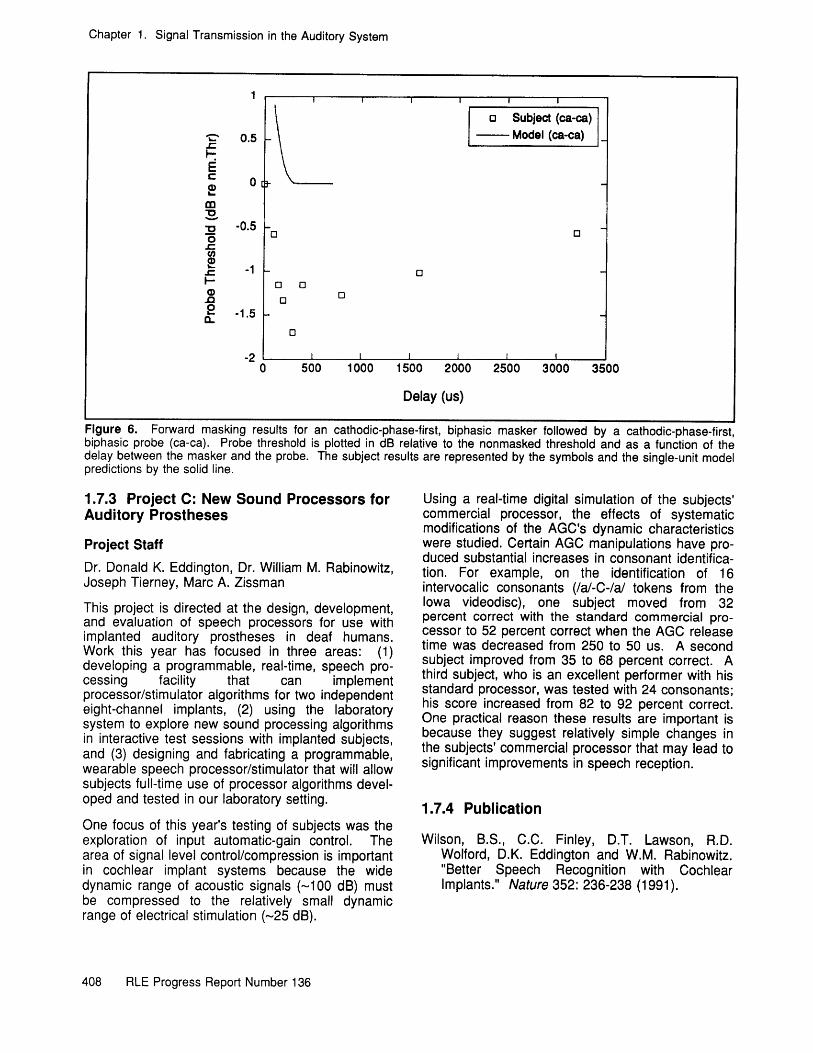
Chapter 1. Signal Transmission in the Auditory System
o Subject (ca-ca)-. 0.5 -- Model (ca-ca)
C 0I-
S -0.5oal
0 0
L -1.5
0
-20 500 1000 1500 2000 2500 3000 3500
Delay (us)
Figure 6. Forward masking results for an cathodic-phase-first, biphasic masker followed by a cathodic-phase-first,biphasic probe (ca-ca). Probe threshold is plotted in dB relative to the nonmasked threshold and as a function of thedelay between the masker and the probe. The subject results are represented by the symbols and the single-unit modelpredictions by the solid line.
1.7.3 Project C: New Sound Processors forAuditory Prostheses
Project StaffDr. Donald K. Eddington, Dr. William M. Rabinowitz,Joseph Tierney, Marc A. Zissman
This project is directed at the design, development,and evaluation of speech processors for use withimplanted auditory prostheses in deaf humans.Work this year has focused in three areas: (1)developing a programmable, real-time, speech pro-cessing facility that can implementprocessor/stimulator algorithms for two independenteight-channel implants, (2) using the laboratorysystem to explore new sound processing algorithmsin interactive test sessions with implanted subjects,and (3) designing and fabricating a programmable,wearable speech processor/stimulator that will allowsubjects full-time use of processor algorithms devel-oped and tested in our laboratory setting.
One focus of this year's testing of subjects was theexploration of input automatic-gain control. Thearea of signal level control/compression is importantin cochlear implant systems because the widedynamic range of acoustic signals (-100 dB) mustbe compressed to the relatively small dynamicrange of electrical stimulation (-25 dB).
Using a real-time digital simulation of the subjects'commercial processor, the effects of systematicmodifications of the AGC's dynamic characteristicswere studied. Certain AGC manipulations have pro-duced substantial increases in consonant identifica-tion. For example, on the identification of 16intervocalic consonants (/a/-C-/a/ tokens from theIowa videodisc), one subject moved from 32percent correct with the standard commercial pro-cessor to 52 percent correct when the AGC releasetime was decreased from 250 to 50 us. A secondsubject improved from 35 to 68 percent correct. Athird subject, who is an excellent performer with hisstandard processor, was tested with 24 consonants;his score increased from 82 to 92 percent correct.One practical reason these results are important isbecause they suggest relatively simple changes inthe subjects' commercial processor that may lead tosignificant improvements in speech reception.
1.7.4 Publication
Wilson, B.S., C.C. Finley, D.T. Lawson, R.D.Wolford, D.K. Eddington and W.M. Rabinowitz."Better Speech Recognition with CochlearImplants." Nature 352: 236-238 (1991).
408 RLE Progress Report Number 136