tamm … cytokinesbyaffectingglomerularfiltrationrateandacting asaurinarycytokinetrap*...
TRANSCRIPT

Tamm-Horsfall Protein Regulates Circulating and RenalCytokines by Affecting Glomerular Filtration Rate and Actingas a Urinary Cytokine Trap*
Received for publication, January 30, 2012, and in revised form, March 22, 2012 Published, JBC Papers in Press, March 26, 2012, DOI 10.1074/jbc.M112.348243
Yan Liu‡, Tarek M. El-Achkar§, and Xue-Ru Wu‡¶�1
From the Departments of ‡Urology and ¶Pathology, New York University School of Medicine, New York, New York 10016, the�Veterans Affairs New York Harbor Healthcare System, Manhattan Campus, New York 10010, and the §Division of Nephrology,St. Louis University and St. Louis Veterans Affairs Medical Center, St. Louis, Missouri 63108
Background: Kidneys play a key role in cytokine catabolism, but the mechanism remains unclear.Results: Deficiency of Tamm-Horsfall protein (THP), a kidney-specific protein, leads to marked increase of circulating cyto-kines, via decreased glomerular clearance and loss of urinary cytokine trapping.Conclusion: THP influences systemic cytokine clearance.Significance: Defects of THP may underlie disease pathogenesis of kidneys and other vital organs.
Although few organ systems play a more important role thanthekidneys in cytokine catabolism, themechanism(s) regulatingthis pivotal physiological function and how its deficiency affectssystemic cytokine homeostasis remain unclear. Here we showthat elimination of Tamm-Horsfall protein (THP) expressionfrom mouse kidneys caused a marked elevation of circulatingIFN-�, IL1�, TNF-�, IL6, CXCL1, and IL13. Accompanying thiswere enlarged spleens with prominent white-pulp macrophageinfiltration. Lipopolysaccharide (LPS) exacerbated the increaseof serum cytokines without a corresponding increase in theirurinary excretion inTHPknock-out (KO)mice. This, alongwiththe rise of serum cystatin C and the reduced inulin and creati-nine clearance from the circulation, suggested that diminishedglomerular filtration may contribute to reduced cytokine clear-ance in THP KO mice both at the baseline and under stress.Unlike wild-type mice where renal and urinary cytokinesformed specific in vivo complexes with THP, this “trapping”effectwas absent inTHPKOmice, thus explainingwhy cytokinesignaling pathways were activated in renal epithelial cells insuch mice. Our study provides new evidence implicating animportant role of THP in influencing cytokine clearance andacting as a decoy receptor for urinary cytokines. Based on theseand other data, we present a unifying model that underscoresthe role of THP as a major regulator of renal and systemicimmunity.
Cytokines are a structurally and functionally diverse group ofsecreted proteins that play essential roles in immunity, inflam-mation, organ development, cell differentiation, and survivaland tissue repair (1, 2). Most soluble cytokines are pleiotropic,
capable of interacting with cognate membrane receptors pres-ent on multiple cell types and, in doing so, eliciting divergentbiological responses (3). Although a physiological level of cyto-kines is an engine to drive the requisite biological activities,their overproduction can lead to detrimental consequences (4,5). In fact, emerging clinical and experimental evidence sug-gests that cytokines are the principal mediators of the complexinter-organ cross-talk and their imbalance underlies multior-gan failure (6). It is therefore no great surprise that undernormal conditions cytokines in the circulation are kept atextremely low levels (2, 7).Of all organ systems, the kidneys play a leading role in the
clearance of circulating cytokines (8–11). With molecularmasses of the mature peptides ranging from 8 to 25 kDa,most cytokines can pass freely through the molecular sievesof the ultrafiltration barrier of the glomeruli in the kidney(7). Indeed, radiolabeled cytokines administered intrave-nously in experimental animals are eliminated within min-utes to hours from the bloodstream and are traced primarilyto the urine (12, 13). For some cytokines, a significant por-tion of the glomerulus-filtered load is endocytosed by theepithelial cells comprising the proximal tubules and destinedfor lysosomal degradation, with the rest excreted in the urine(14, 15). Alternatively, urinary cytokines could be reab-sorbed via the paracellular route along the renal tubules (16),but this possibility has yet to be experimentally validated.Despite the demonstrated importance of the kidneys in cyto-kine catabolism, many key questions remain. For instance,how is cytokine filtration through the glomeruli regulated?Would a reduced glomerular filtration affect renal cytokineclearance and hence systemic cytokine retention, and if so,would that have a systemic pathological effect? In addition,free urinary cytokines that have escaped the epithelialuptake are known to be harmful for renal tubular andurothelial cells (17–19). Does a decoy receptor exist in theurine that can serve as a scavenger to prevent the cytokinesfrom overactivating the renal epithelial and urothelial cellsand causing urinary tract pathology?
* This work was supported, in whole or in part, by National Institutes of HealthGrant R01DK56903 and Merit Review Awards from the Veterans Adminis-tration’s Research Program (to T. E.-A. and X.-R. W.).
1 To whom correspondence should be addressed: 423 E23 St., Rm. 18064South, New York, NY 10010. Fax: 212-951-5424; E-mail: [email protected].
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 287, NO. 20, pp. 16365–16378, May 11, 2012Published in the U.S.A.
MAY 11, 2012 • VOLUME 287 • NUMBER 20 JOURNAL OF BIOLOGICAL CHEMISTRY 16365
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Tamm-Horsfall protein (THP)2 is a 90-kDa, heavily glycosy-lated protein made by the functionally specialized epithelialcells of the kidney comprising the thick ascending limb of loopof Henle (TAL) (20–25). Mature THP is anchored onto theluminal leaflet of the apical membrane of TAL until its con-trolled release into the urine by protease and/or lipase activities(26, 27). Although reactive substances to polyclonal THP anti-bodies were found in non-kidney sources including serum,brain, liver, and amniotic fluid (23, 28, 29), recent RT-PCR andtransgenicmice harboring a THP promoter/reporter transgenedemonstrated that THP is expressed only in the kidneys (30,31). The relationship of THP with immunity was not appreci-ated until it was “rediscovered” as a potent immune suppressantin the urine of pregnant women and renamed uromodulin (32,33). However, the primary sequences of these two proteins areidentical, although less is certain about their exact glycosylationcontents (34–36). In vitro, uromodulin binds avidly with IL1,IL2, and TNF, and inhibits antigen-mediated T cell prolifera-tion or the IL1-mediated T-cell colony response by phytohem-agglutinin (37–40). Uromodulin also suppresses the cytotoxic-ity of monocytes. Interestingly, the urinary level of uromodulinincreases during the second and third trimester of normal preg-nancy, a physiological response attributed to an increased needfor immune suppression during this period. In contrast, uro-modulin ismarkedly reduced or even undetectable in hyperten-sive preeclampsia patients (41, 42). Notably, most if not all ofthe in vitro immune suppressive properties of uromodulinwerelater found to be shared by THP isolated from healthy maleurine, suggesting a functional commonality between THP anduromodulin (20, 35). Despite these intriguing observations,controversies persisted regarding the role of THP in immunity.First and foremost, unlike the recombinant cytokines, nativecytokines did not appear to interact with THP in vitro (43).Second, because normally THP is not significantly present inthe interstitium or circulationwhere there are immune cells, itsproposed immunosuppressive role would have to be local, e.g.to be confined to the urinary tract. Third, THP is an effectiveimmune stimulator when directly exposed to neutrophils,monocytes, dendritic cells, via up-regulation of target cell cyto-kines and enhancement of their chemotaxis/phagocytosis (44–46). The effects of THP on dendritic cell maturation andautoantibody production after intravenous THP injection areboth dependent on TLR-4 signaling (46). These results seem tobe in direct conflict with those suggesting a role for THP inimmune suppression. Efforts in elucidating an in vivo role ofTHP in immune regulation and in reconciling the seeminglydiametrically opposing functions of the THP have been ham-pered by the lack of experimentalmodels, althoughmore recentdata support an immunosuppressive role for THP in the ische-mia-reperfusion model of acute kidney injury (47, 48).In the present study, we explored the in vivo net effects of the
loss of THP on circulating and renal cytokine homeostasisunder both steady-state and stressed conditions. We studiedthe renal mechanisms whereby the absence of THP in knock-
out mice alters cytokine clearance and hence systemic cytokinelevels. Additionally, we examined whether, in the wild-typemice, renal and urinary THP forms natural complexes withcytokines and whether such a binding/trapping effect is absentin the THP KO mice using several independent but comple-mentary approaches. We also determined the pathophysiolog-ical consequences of un-trapped urinary cytokines on renal epi-thelial activation.On the basis of our data, we present a unifyingmodel that reconciles the current controversies and explainswhyTHPplays an important role in renal and systemic cytokinehomeostasis. Finally, because gene mutations, polymorphisms,and reduced expression of THP occur frequently in pathologi-cal conditions in humans, our findings suggest that local andsystemic cytokine imbalances caused by THP deficiency mayplay a key role in the disease pathogenesis of the kidneys andother vital organs.
EXPERIMENTAL PROCEDURES
Genetically EngineeredMice Lacking Tamm-Horsfall Protein(Uromodulin)—Mice defective for the THP gene were gener-ated using homologous recombination, resulting in the deletionof a region of the THP gene from 650 bp upstream of the tran-scriptional initiation site to the middle of intron 4 (49).Homozygous THP KO mice lacked THP mRNA or protein inthe kidney, as evidenced by Northern blotting, RT-PCR, in situhybridization, Western blotting, and immunohistochemistry.BothTHPKOmice and their wild-type littermates used for thisstudy were maintained in a specific pathogen-free facility andbred in parallel in a 129/SvEv background for 6 generations.Themicewere tested vigorously to be free of any viral, bacterial,or parasitic infection. Mice, 2–3 months old, of both genderswere used for this study. The genotype of the KO mice wasverified using PCR of tail genomic DNA with specific primers(forward, 5�-gaagggactggctgctattg-3�; reverse, 5�-aatatcacggg-tagccaacg-3�). The genotype of the wild-type mice was verifiedsimilarly with specific primers (forward, 5�-agggctttacaggggat-ggttg-3�; reverse, 5�-gattgcactcagggggctctgt-3�). All animal-re-lated procedures were carried out in accordance with federaland local regulations and under an active protocol approved bythe Institutional Animal Care and Use Committee (IACUC).Quantitation of Serum, Urine, and Kidney Cytokines by
ELISA—Whole bloodwas obtained from the retro-orbital sinusroute of 2–3-month-old mice under anesthesia with ketamineand xylazine, and the serumwas isolated routinely and stored at�80 °C until use. The same cohorts (WT (n� 23) and KOmice(n � 22)) were used for the ELISAs to test serum cytokines,although, due to finite volume recovery, not all mice yieldedsufficient amounts for all the assays, hence the slightly variednumbers for different cytokines. The 24-h urine samples werecollected using autoclaved single-mouse metabolic cages(VWR; NALGENE*) into the collecting tubes containing 20 �lof 10% thymol and 10 �l of a mixture of protease inhibitors (80�M aprotinin, 5 mM bestatin, 1.5 mM E-64, 5 mM EDTA, 2 mM
leupeptin, and 1mMpepstatinA; ThermoScientific). After cen-trifugation at 500 � g, the urine supernatants were kept at�80 °C until use. Total kidney proteins were extracted byhomogenizing freshly dissected kidney tissue in lysis buffercontaining a final concentration of 20 mM Hepes (pH 7.5), 150
2 The abbreviations used are: THP, Tamm-Horsfall protein; TAL, thick ascend-ing limb of loop of Henle; GFR, Glomerular filtration rate; KO, knock-out; IP,immunoprecipitation.
THP Deficiency and Cytokine Imbalance
16366 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 20 • MAY 11, 2012
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

mMNaCl, 1 mM EGTA, 1.5 mMMgCl2, 10% glycerol, 1% TritonX-100, and a mixture of protease inhibitors (same as above).After centrifugation at 17,000 � g at 4 °C for 20 min, the super-natant was collected and stored at �80 °C until use. Proteinconcentrations were determined using a BCA reagent (Pierce)with bovine serum albumin as a standard.Measurement of cytokine levels in serum, urine, and kidney
was carried out using commercial ELISA kits consisting of anti-bodies and protein standards for mouse cytokines includingIFN-�, IL1�, TNF-�, and IL6 (Thermo Fisher Scientific), andCXCL1 (mouse equivalent of human IL8) and IL13 (R&D Sys-tems). In the case of ELISA involving kidney proteins, cytokinestandards were used as positive controls; and sample dilutionbuffer, bovine serum albumin, and purified mouse THP wereused as negative controls.Lipopolysaccharide Challenge—Lipopolysaccharide purified
from Escherichia coli (0111:B4) using ion-exchange chroma-tography (500,000 endotoxin units/mg; Sigma) was reconsti-tuted in autoclaved phosphate-buffered saline to a final concen-tration of 1 mg/ml. Four milligrams of LPS per kilogram ofmouse body weight was injected intraperitoneally into ran-domly selected groups of THP KO and wild-type mice. At var-ious time points post-injection (1.5, 3, 6, 12, and 24 h), the bloodsamples were obtained via the retro-orbital sinus route fromanestheticmice, whereas kidney specimenswere procured afterthe mice were euthanized. The sera and total kidney proteinextracts were then subject to ELISA as described above.Determination of Serum Cystatin C and Renal Inulin and
Creatinine Clearance—Serum cystatin C, a marker for renalglomerular filtration function (50, 51), was measured by ELISAusing a kit specific for themouse protein (BioVendor), based onthe manufacturer’s instructions.The glomerular filtration rate (GFR) was determined using
intravenous delivery of FITC-conjugated inulin following a pre-viously established protocol (52). Briefly, FITC-conjugated inu-lin (Sigma) was reconstituted in 0.9%NaCl to a final concentra-tion of 5%. After dialysis in the dark against 0.9% NaCl (dialysismembrane cut-off: 1,000-Da; Spectrum Laboratories) toremove unbound FITC, the solution was filtered through a0.22-�m pore filter (Millipore), and the filtrate was theninjected intravenously via the retro-orbital route under isoflu-rane-induced anesthesia. After injection, 20 �l of blood sam-ples were collected via the saphenous vein at 3, 7, 10, 15, 35, 55,and 75 min. The plasma was employed for determining theconcentrations of FITC-inulin at 485-nm excitation and538-nm emission wavelengths. The fluorescence readings wereplotted into a 2-phase exponential decay curve using nonlinearregression with Internet-based SPSS software. GFR was deter-mined with the formula: GFR � I/(A/� � B/�) (I, the quantityof administered FITC-inulin; A and B, the y intercept values ofthe two decay rates; � and �, decay constants for the distribu-tion and elimination phases) (53). GFR was expressed asml/min (with or without being referenced to the body weight).Fractional creatinine clearance rate was determined by
measuring the concentrations of the serumand24-h urine sam-ples using a commercial kit (Biovision, Mountain View, CA).After dilution in the assay buffer, serumand urine sampleswereincubated in a 96-well microplate with a solution containing
creatinase, creatininase, enzyme mix, and color probe at 37 °Cfor 1 h. The creatinine concentration was calculated by plottingthe reading at 570 nm against a standard curve. Creatinineclearance was expressed as: urinary creatinine concentration�24-h urine volume/serum creatinine concentration.Serum cystatin C level and inulin and creatinine clearance
were determined at the baseline and after LPS challenge. In thelatter situation, blood samples were obtained 12 h after the LPSchallenge.In Vivo Interaction of Renal Cytokines with Tamm-Horsfall
Protein—THP KO mice and their wild-type controls (withoutLPS challenge) were sacrificed and their kidneys dissected outand homogenized in a lysis buffer designed for immunoprecipi-tation (50 mM Tris/HCl (pH 8.0), 150 mM NaCl, 1% NonidetP-40, 0.5% sodium deoxycholate, 0.1% SDS, and a mixture ofprotease inhibitors (same as above)). After centrifugation at17,000� g at 4 °C for 20min, the supernatant was collected andits total protein concentration was determined using the BCAreagent. The supernatant containing 1 mg of the total kidneyprotein extract was supplemented with protein A-conjugatedagarose (Bio-Rad). After centrifugation, the supernatant wasreconstituted with the addition of a rabbit anti-THP antibody(54) to a final dilution of 1:250, and the mixture was incubatedat 4 °C overnight. As negative controls, isotype, nonimmune,normal rabbit sera were similarly diluted and subjected to thesame experimental procedures as with the rabbit anti-THPantibody. Fresh protein A-agarose was then added to the mix-ture and again incubated at 4 °C overnight. After centrifuga-tion, the agarose beadswerewashed 4 times in immunoprecipi-tation buffer and the bound protein complexes were eluted byboiling the agarose beads in 200�l of the aforementioned bufferfor 10 min. Fifty microliters of the elutes were then added intothe ELISA plates that were pre-coated with antibodies againstmouse cytokines (Thermo Fisher Scientific), and ELISA wasthen carried out as described above. In parallel experiments,equal amounts (100 �g) of the total kidney protein extractswere added directly into ELISA plates followed by ELISA toquantify the amount of renal cytokine input (THP-bound andunbound cytokines).Native Polyacrylamide Gel Electrophoresis of THP-Cytokine
Complex—One hundred micrograms of total kidney proteinsextracted with the immunoprecipitation buffer describedabovewere resolved on a 6% polyacrylamide gel in nondenatur-ing and nonreducing conditions. The proteins were then elec-trotransferred onto an Immobilon-PVDF membrane andimmunoblotted with goat anti-mouse IL1� antibody. Follow-ing the incubation with a secondary donkey anti-goat antibodyconjugated with horseradish peroxidase, the IL1� band wasvisualized with enhanced chemiluminescent (ECL) reagents(Pierce). The membrane was stripped and re-blotted with rab-bit anti-THP antibody followed by a secondary, goat anti-rabbitantibody conjugated with peroxidase. To rule out nonspecificbinding, the samemembranes were stripped again and re-blot-ted with isotype antibodies (nonimmune normal goat serum inplace of goat anti-mouse IL1� antibody; nonimmune normalrabbit serum in place of rabbit anti-THP antibody).
THP Deficiency and Cytokine Imbalance
MAY 11, 2012 • VOLUME 287 • NUMBER 20 JOURNAL OF BIOLOGICAL CHEMISTRY 16367
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Co-sedimentation of Cytokine-THP Complex by Centrifu-gation—Freshly collected 24-h urine samples from wild-typeand THP KOmice were centrifuged at 17,000 � g at 4 °C for 15min. The pellets were re-suspended in distilled water; and thesuspension and the corresponding supernatantswere subjectedto ELISA. The status of THP was determined by SDS-PAGEfollowed by Coomassie Blue staining as well as Western blot-ting using a rabbit anti-THP antibody (54).Double Immunofluorescent Staining—Coronal sections (5
�m thick) of paraffin-embedded kidneys from the wild-typemice (without LPS challenge) were de-paraffinized and under-went antigen unmasking in citrate buffer (pH 6.0) using micro-
wave treatment (9/10poweroutput for20min).The sectionsweredouble-stained with (i) goat anti-IL1� and rabbit anti-THP or (ii)goat anti-IFN-� and rabbit anti-THP, followed by two secondaryantibodies (Alexa Fluor 594-conjuated donkey anti-goat IgG (redfluorescence) and Alexa Fluor 488-conjuated donkey anti-rabbitIgG (green fluorescence)). Isotype, nonimmune antibodies (nor-mal goat serum in place of anti-IL1� and goat anti-IFN-�; normalrabbit seruminplaceofanti-THP)wereused inparallel asnegativecontrols with identical staining procedures.Real-time Quantitative PCR Quantification of Renal Cyto-
kine Production—Total RNAs were extracted from WT andTHP KO mouse kidneys using the TRIzol RNA extraction kit
FIGURE 1. Splenic enlargement and macrophage infiltration in THP-KO mice. A, gross anatomy of spleens from a representative wild-type (WT) mouse anda THP KO mouse. Note the increased size and uneven surface of the spleen from the KO mouse. B, comparison of the spleen weight and spleen/body weightratio of randomly selected wild-type (n � 32) and THP KO (n � 32) mice. Note that THP KO mice had significantly increased spleen weight without (left panel)or with (right panel) body weight as a reference. Error bars represent S.D. C, histopathology and immunohistochemistry of the spleens. H&E staining showed agreatly increased number of macrophages in the white pulp of the spleens of THP KO mice (upper and middle panels on right) that were strongly labeled withan antibody against mouse F4/80 (anti-Mø), a macrophage-specific marker (lower right panel). Magnification of upper panels, �100; middle and lower panels,�200. D, quantification of anti-Mø staining. Stained sections from WT and THP KO mice (3 sections per mouse from 3 WT and 3 THP KO mice) were scanned forMø-stained area/intensity. Average reading for WT mice was set at 1 and that for THP KO mice expressed as fold-changes (�S.D.) in reference to the WT mice.
THP Deficiency and Cytokine Imbalance
16368 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 20 • MAY 11, 2012
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from
(Invitrogen). Double-stranded cDNAs were then synthesizedand used as templates for real-time PCR using a QuantiTectSYBR Green PCR Kit (Qiagen). The primers used were: 1)IFN-�, sense, 5�-AAC GCT ACA CAC TGC ATC TT GG-3�;antisense, 5�-GAC TTC AAA GAG TCT GAG G-3�; 2) IL1�,sense, 5�-TCG GGA GGA GAC GAC TCT AA-3�; antisense,5�-GTG CAC CCG ACT TTG TTC TT-3�; 3) TNF-�, sense,5�-ATG AGC ACA GAA AGC ATG ATC-3�; antisense,5�-TAC AGGCTTGTCACT CGAATT-3�; and 4) IL6, sense,5�-AACGATGATGCACTTGCAGA-3�; antisense, 5�-GGAAATTGGGGTAGGAAGGA-3�. PCR conditionswere: 95 °Cfor 10 min for the first cycle, 95 °C for 20 s, 58 °C for 30 s, and72 °C for 30 s for 45 cycles. The products of the PCRs werequantified by measuring the extent of incorporation of SYBRGreen I into newly synthesized DNA and expressed as ratiosversus the amount of�-actin thatwas amplified in parallel usingprimers: sense, 5�-TGT TAC CAACTGGGACGACA-3� andantisense, 5�-TCT CAG CTG TGG TGG TGA AG-3�.Histopathology, Immunohistochemistry, and Western Blot-
ting—Spleens fromwild-type and THPKOmice were paraffin-embedded, sectioned, and stained routinely with hemotoxylinand eosin. Alternatively, paraffin sections underwent antigenunmasking as described above and then immunohistochemicalstaining using a rat antibody against the mouse macrophagemarker (F4/80(CI:A3)) (Novus). After incubation with a sec-ondary goat anti-rat antibody conjugated with horseradish per-oxidase, the reaction was developed in 50 mM Tris/HCl buffer(pH 7.4) containing 3,3�-diaminobenzidine tetrahydrochlorideand H2O2. For quantification of the staining area and intensity,three antibody-stained sections per mouse of 3WT and 3 THPKO mice were scanned and analyzed using NIH Image J soft-ware (rsb.info.nih.gov/ij/index.html). The average reading fromWT mice was set at 1 and that from THP KO mice wasexpressed as fold-changes in reference to the WT mice.For the assessment of renal neutrophil infiltration, paraffin-
embedded kidney sections from wild-type and THP KO miceunderwent antigen retrieval as described, followed by incuba-tion with a primary rat antibody against mLy-6G, a cell surfacemarker of mouse peripheral neutrophils (BioXCell; 1:10,000dilution), and then with a secondary, donkey anti-rat antibodyconjugated with Alexa Fluor 594 (Invitrogen; 1:500 dilution).The sections were double-stained with a rabbit anti-mouse col-lagen type IV polyclonal antibody (Abcam Inc.; 1:500 dilution)visualized by anAlexa Fluor 488-conjugated donkey anti-rabbitantibody (Invitrogen Corp.). The sections were subsequentlycounterstained with DAPI to reveal the nuclei. Positivelystained neutrophils were counted at �400 magnification from20 nonoverlapping fields from each section, three sections eachof kidney, and 8 mice per genotype. The neutrophil count wasexpressed as the number of neutrophils per 10 high-powerfields. Cross-sections of amouse bladder inoculated with a uro-pathogenic E. coli strain (UTI89) were used as positive controlsfor the neutrophil antibody.Key signaling molecules in the cytokine activation pathway
were determined by Western blotting using total kidney pro-tein extracts. After SDS-PAGE, the proteins were transferredonto an Immobilon-PVDFmembrane and blottedwith primaryantibodies against I�B,NF-�B (p65), phosphorylated Stat3, and
MAPK (loading control), with subsequent steps carried outroutinely.Statistical Analysis—Mann-Whitney U test (two-sided) was
used to assess the differences of spleen weights between wild-type and THP KO mice using SPSS software. Student’s t testswere performed to evaluate the differences between the wild-type and THP KOmice for all other applicable assays using thesame software. A p value of less than 0.05 was considered sta-tistically significant.
RESULTS
Splenic Enlargement andMacrophage Infiltration inTHPKOMice—An unexpected, yet frequent finding during gross anat-omy of our THP KO mice was the markedly increased sizes ofthe spleens. Additionally, whereas spleens from the wild-typelittermates always had smooth, shinning surfaces, those of theTHP KO mice were often uneven, dull-colored and nodular(Fig. 1A). Two groups of randomly selected THP KO (n � 32)and wild-type (n � 32) mice, both aged 3 months and in thesame (129/SvEv) background, were therefore sacrificed andtheir spleen and bodyweightsmeasured. It became evident thatthe average spleen weight of THP KO mice was significantlygreater than that of wild-type mice, either without (Fig. 1B, leftpanel) or with (Fig. 1B, right panel) adjustment to the bodyweight. Severely enlarged spleens were also occasionally noted,with the spleen weights three times as heavy as those of thewild-type controls. H&E staining of the enlarged spleensshowed disorganized white-pulp and prominent infiltration bythemacrophages (Fig. 1C), the identity of which was confirmed
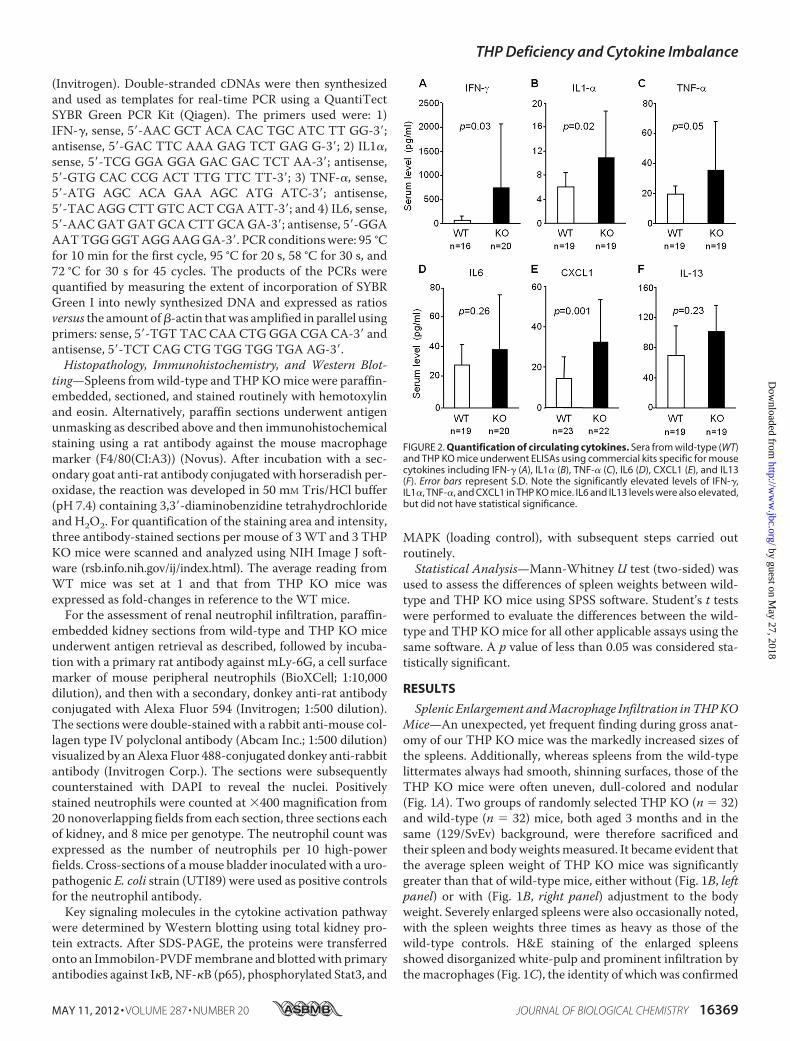
FIGURE 2. Quantification of circulating cytokines. Sera from wild-type (WT)and THP KO mice underwent ELISAs using commercial kits specific for mousecytokines including IFN-� (A), IL1� (B), TNF-� (C), IL6 (D), CXCL1 (E), and IL13(F). Error bars represent S.D. Note the significantly elevated levels of IFN-�,IL1�, TNF-�, and CXCL1 in THP KO mice. IL6 and IL13 levels were also elevated,but did not have statistical significance.
THP Deficiency and Cytokine Imbalance
MAY 11, 2012 • VOLUME 287 • NUMBER 20 JOURNAL OF BIOLOGICAL CHEMISTRY 16369
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

by immunohistochemical staining using an antibody againstmouse F4/80, a marker specific for the macrophages. Spleensfrom the THP KOmice contained a considerably greater num-ber of macrophages than those from the wild-type mice (Fig.1D). No other organ exhibited inflammatory changes in theTHPKOmice (data not shown). These findings revealed a here-tofore unsuspected, specific extra renal pathological conse-quence of the loss of THP expression in the kidney.Marked Increase of Circulating Cytokines in THP KO Mice—
Our finding that loss of THP, a protein made exclusively in thekidneys, would lead to splenomegaly prompted us to explorewhether this was due to a rise in circulating cytokines. UsingELISA, we quantified the serum levels of inflammatory cyto-kines IFN-�, IL1�, TNF-�, IL6, and CXCL1 (a mouse homo-logue of IL8) and a noninflammatory cytokine IL13. All but IL6and IL13 were significantly higher in THP KOmice than in thewild-type controls (Fig. 2, A–F). IL6 and IL13 levels were alsoelevated, although they did not reach statistical significance(Fig. 2,D and F). The serum level of IFN-� (averaged 750 pg/mlin THP KOmice; Fig. 2A) was considerably higher than that ofthe other cytokines (averaged 10–35 pg/ml; Fig. 2, B–F), sug-gesting IFN-�, a key activator of macrophages (55), as a keyreason for the splenomegaly observed in our THP KO mice.The relatively large standard deviation in Fig. 2 (also see below)might reflect the fact that, despite 6 generations of back-cross-ing, the mice we used had not reached an in-bred status,although other possibilities cannot be completely ruled out.When challenged with lipopolysaccharide, the increase of
the circulating cytokines over the steady-state level was alsomore pronounced in THP KO mice than in wild-type controls(Fig. 3, A–F). The time course of induction varied among thecytokines. TNF-�peaked around 1.5 h after LPS administration
and leveled off quickly after 6 h (Fig. 3C), in accordance with itsbeing an early responder. IL1� lagged in its peak time (e.g. at3 h) and subsided also less quickly (after 12 h) (Fig. 3B). In
FIGURE 3. Circulating cytokines in response to LPS treatment. Wild-type (WT, filled circle; n � 8) and THP knock-out (KO, filled square) mice (n � 8) wereinjected intraperitoneally with LPS, and their blood was obtained at 1.5, 3, 6, 12, and 24 h after injection and processed for ELISA. Note that although serumcytokine levels increased in both WT and KO mice in response to LPS, KO mice had significantly higher levels of cytokines, except IL13, at various time pointsthan WT mice (asterisks denote the time points where statistical significance was present; see text for details). IL13 levels were also higher in KO mice than inWT mice at all time points, but did not reach statistical significance.
FIGURE 4. Urinary excretion of cytokines. Twenty-hour urine samples col-lected with metabolic cages were subject to ELISA for IFN-� and IL1�. Theconcentrations were normalized against simultaneously measured creati-nine. Total excretion in a 24-h period was also calculated based on the con-centrations and urine volumes. Note that urinary excretion of IFN-� and IL1�was not higher in THP KO mice than in WT controls. Other cytokines shown inFigs. 2 and 3 were also measured with the same set of urine samples but wereundetectable, thus not shown here.
THP Deficiency and Cytokine Imbalance
16370 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 20 • MAY 11, 2012
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

comparison, IFN-� and IL6 did not reach the highest levelsuntil 6 h and remained high until after 24 h (Fig. 3, A and D).CXCL1 and IL13, on the other hand, peaked early (e.g. at 1.5 h)and remained high throughout the 24-h observation period.Taken together, although LPS induced cytokines in both wild-type and THP KO mice, it had more profound effects on THPKO mice.Effect of THPDeficiency onUrinary Cytokine Excretion—The
kidney plays an important role in the clearance of cytokines (9,14, 15), but the exact mechanisms involved are not well under-stood. For example, clearance depends on at least three princi-pal factors: glomerular filtration rate, cytokine reabsorption,and tubular degradation. Because of the elevated levels of serumcytokines in THP KO mice, we examined their urinary excre-tion rate. The urinary concentrations of IFN-� and IL1� inTHPKO mice were comparable with those in WT mice (with orwithout adjustment to creatinine), and the total amounts oftheir urinary excretion in 24 h were not significantly differentbetween THP KO and wild-type mice (Fig. 4). This suggestedthat the clearance of IFN-� and IL1� was reduced in THP KOmice. Despite their elevated levels in the serum, urinary TNF-�,
IL6, CXCL1, and IL13 were undetectable in the THP KO miceas with the wild-type controls (data not shown). This may bebecause these latter cytokines undergo significant post-filtra-tion handling (such as reabsorption) by the kidney without anyresidual urinary excretion. Our repeated attempts to determinethe urinary levels of the various ILs in LPS-treated mice weremetwith limited success, because thesemice became extremelyill andwere not amenable to urine collection inmetabolic cages.Reduction in Glomerular Filtration as a Possible Contributor
to Elevated Serum Cytokines—Because cytokine clearancedepends in part on GFR (9, 14, 15), we used several methods toassess the differences in GFR between WT and THP KO mice.First, we measured the serum level of cystatin C, a low molec-ular mass (13.3 kDa) and positively charged protein removedfrom circulation almost exclusively by glomerular filtration (50,51, 56, 57). We found the steady-state level of serum cystatin Cto be significantly higher in THPKOmice than in the wild-typemice (Fig. 5A, left two columns). We also determined the clear-ance of FITC-conjugated inulin from the circulation and foundit to be significantly lower in THP KO mice without (Fig. 5B,upper panels) or with (Fig. 5B, lower panels) it being referenced
FIGURE 5. Serum concentration of cystatin C and inulin and creatinine clearance before and after LPS challenge. A, serum cystatin C. The concentrationof serum cystatin C of WT mice (n � 8) and THP KO (n � 8) was determined using a commercial ELISA kit for mouse cystatin C. Note that at the baseline THP KOmice had significantly elevated levels of cystatin C (left two columns). LPS treatment increased the serum cystatin C level (24 h post-treatment) significantlymore in THP KO mice than in WT controls (right two columns). B, inulin clearance. Subsequent to inulin-FITC administration (at 12 h post-treatment), theconcentrations of serum inulin-FITC were quantified at different time points. Note that inulin clearance was significantly lower in THP KO mice than in WTcontrols at baseline (left two columns) and after LPS challenge (right two columns), without (upper panel) or with (lower panel) being referenced to the bodyweight. Error bars represent S.D. C, creatinine clearance. The concentrations of urine and serum creatinine were determined using a commercial kit, and urinevolume was measured with metabolic cages. Note that creatinine clearance was significantly lower in THP KO mice than in WT controls, without (upper panel)or with (lower panel) being referenced to the body weight.
THP Deficiency and Cytokine Imbalance
MAY 11, 2012 • VOLUME 287 • NUMBER 20 JOURNAL OF BIOLOGICAL CHEMISTRY 16371
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

to body weight. The extent of serum cystatin C elevation andinulin clearance reduction in THP KO mice worsened whenthey were challenged with LPS (Fig. 5, A and B, right two col-umns). Specifically, inulin clearance under the untreatedcondition in THP KOmice was 22.5% lower than that of WTmice (Fig. 5B). After LPS treatment it was 46.9% lower thanin WT mice. Last, we determined the fractional excretionrate of creatinine both at steady-state and after LPS chal-lenge (Fig. 5C), yielding data highly consistent with those ofthe inulin clearance. These data point to a glomerular filtra-tion defect in THP KO mice, both at the baseline and uponLPS challenge, as a possible contributor of a reduced renalcytokine clearance. The reduction in inulin and creatinineclearance in THP KOmice was somewhat disproportional tothe increase in serum cytokines, especially for IFN-� andIL1�, suggesting that additional post-filtration tubular han-dling may be involved in regulating the level of circulatingcytokines (see later).Renal and Urinary Cytokines Bound to THP in Wild-type
Mice, a “Trapping” Effect Lost in THPKOMice—Although pre-vious studies showed in vitro binding of several recombinantcytokines to THP, it remained highly controversial as towhether such a binding occurs in vivowith native cytokines (20,21, 23, 24). To clarify this issue, we tested whether cytokinescould co-purify with THP from total kidney protein extracts ofwild-type mice, but not from THP KO mice. Kidney proteinextracts from both types of mice (without LPS challenge) weresubject to immunoprecipitation using an anti-THP antibody,followed by ELISA of cytokines on the immunoprecipitatedproducts. Compared with the THP KO mice from which littleof the four tested cytokines (IFN-�, IL1�, TNF-�, and IL13)could be co-precipitated by the THP antibody, these cytokineswere readily precipitated in wild-type mice (Fig. 6). Approxi-mately 1% IFN-�, 25% IL1�, and 10% IL13 of their respectivetotal inputs were co-purified (Fig. 6, A, B, and D, compare theupper panels with the corresponding lower panels), whereasonly 0.1% TNF-� was co-purified with THP (Fig. 6C, comparethe upper panel with the lower panel). The relatively low pro-portion of IFN-� and TNF-� that were THP-bound could be areflection of the relatively high total cytokine pool in the renalepithelial cells of which most was cytoplasmic and not in directcontact with the apical surface located or urinary THP (see Fig.7C). It is also possible that different cytokines bound to THPwith different affinities and/or capacities (see “Discussion”).Despite the overall higher cytokine concentrations in the start-ing materials of the THP KOmice (filled bars in the lower pan-els of Fig. 6,A–D), little if any cytokine was precipitated by THPantibody in THP KO mice, again supporting the specific cyto-kine-THP interaction occurring only in the WT mice. In addi-tional negative control experiments done in parallel, normalnonimmune rabbit serum replacing the anti-THP failed toimmunoprecipitate any appreciable level of cytokines (data notshown), further demonstrating the specificity of the THP-cyto-kine interaction in the WT mice. These experiments, per-formed three times with three independent cohorts, yieldedhighly consistent results.To verify the co-immunoprecipitation data, we performed
native gel electrophoresis by resolving total kidney proteins
under nondenaturing and nonreducing conditions. UponWestern blotting, we found that IL1� co-migrates with THP inwild-type mice, but not in THP KO mice (Fig. 7A). Isotypecontrol antibodies from the same animal species where the pri-mary antibodies were generated did not produce any band, thusestablishing the specificity of the antibody-antigen reaction(Fig. 7A, lower panels). Taking advantage of the property ofurinary THP to form high-molecular mass polymers (58), wealso tested whether urinary cytokines could be co-sedimentedwith THP by high-speed centrifugation.We found that, in con-trast to the THPKOmice in which IL1� was almost exclusivelypresent in the urine supernatant, an appreciable amount of thiscytokine was found in urine pellets of wild-type mice whereTHP was present (Fig. 7B), thus further supporting the THP-cytokine interaction. Finally, in wild-type mouse kidneys, IL1�and IFN-� were co-localized immunohistochemically withTHP at the apical surface of the thick ascending limb of loop ofHenle, where THP is normally located (Fig. 7C). Both IL1� andIFN-� had a wider tubular distribution (in proximal and distaltubular cells in addition to TAL), and was more cytoplasmicthan THP, thus explaining why only some of the interleukinswere THP-bound in the co-immunoprecipitation (IP) experi-ment (Fig. 6).
FIGURE 6. In vivo binding of renal cytokines to THP. Top panels, co-immu-noprecipitation of THP-bound cytokines followed by cytokine ELISA. Totalkidney protein extracts from wild-type (WT) and THP KO mice (2 representa-tive mice per genotype shown) were subject to IP using a rabbit anti-THPantibody. The IP products were loaded into microtiter wells pre-coated withantibodies against IFN-� (A), IL1� (B), TNF-� (C), or IL13 (D), to capture THP-bound cytokines. This was followed by conventional ELISA (see “Experimen-tal Procedures” for details). Starting materials for IP contained the sameamount of total kidney proteins. Lower panels, ELISA quantification of renalcytokine input (e.g. THP-bound and -unbound cytokines), expressed as permilligram of total kidney proteins. Note that, whereas THP KO mice hadhigher renal cytokines (lower panels, filled bars), little could be precipitatedwith anti-THP antibody (upper panels, filled bars). In contrast, in wild-typemice, each of the four tested cytokines could be precipitated by the anti-THPantibody (upper panels, open bars) strongly suggesting an in vivo THP-cyto-kine interaction (see text for details).
THP Deficiency and Cytokine Imbalance
16372 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 20 • MAY 11, 2012
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Effects of THP Deficiency on Renal Cytokine Content andProduction and Renal Inflammation—To test the potentialeffects of the unbound urinary cytokines in the absence ofTHP on renal epithelial cells, we compared the renal cyto-kine levels of THP KO mice versus those of wild-type mice(Fig. 8). The steady-state levels of IFN-�, IL1�, IL6, and IL13were all significantly higher in THP KO mice than in wild-
type mice (Fig. 9, A, B, D, and F). TNF-� and CXCL1 werealso higher although the difference was not statistically sig-nificant (Fig. 8, C and E). Western blotting analyses showedthat there was a down-regulation of I�B and up-regulation ofNF�B and phosphorylated Stat3, supporting the cytokine-mediated activation of signaling pathways in THP KOmousekidneys (Fig. 9A).
FIGURE 7. Native gel electrophoresis, centrifugal co-sedimentation, and immunohistochemical co-localization of the IL-THP complex. A, total kidneyproteins were extracted with a nondenaturing/nonreducing buffer, resolved by a native polyacrylamide gel, electrotransferred onto an Immobilon PVDFmembrane, and immunoblotted with anti-IL1-�. The membrane was then stripped and re-immunoblotted with anti-THP antibody. To demonstrate strictspecificity, the same membranes were stripped again and blotted with nonimmune, isotype antibodies from the same animal species as with anti-IL1� andanti-THP. Short lines on the left edge of the left panels denote the molecular mass standards (from top to bottom: 130, 95, 72, 56, 43, 34, 26, 17, and 11 kDa). Notethat IL1� co-migrated with THP in wild-type (WT) mice, but not in THP KO mice, indicating in vivo IL1�-THP complex formation. Also note the lack of anynonspecific reaction with isotype antibodies. B, left panel, 24-h urine samples from wild-type and THP KO mice were centrifuged at high speed and their pelletswere re-suspended and subject to ELISA of IL1�. Note that urinary IL1� could be precipitated by centrifugation in the WT mice, which contained THP, but notin the THP KO mice, which lacked THP. B, right panel, Coomassie Blue-stained SDS-PAGE (left strip) and Western blotting (right strip; using rabbit anti-THP) of thepellets from the high-speed centrifugation (left panel), showing the presence of THP (arrow) in WT mice but not in THP KO mice. Short lines on the right edge ofthe right strip denote the molecular mass standards (from top to bottom: 130, 95, 72, 56, 43, 34, 26, and 17 kDa). C, paraffin-embedded kidney sections fromwild-type mice were double-stained with (i) anti-IL1� and anti-THP antibodies (upper panels); (ii) anti-IFN-� and anti-THP (middle panels); and (iii) isotype,nonimmune antibodies as negative controls for anti-IL1�/anti-IFN-� and anti-THP (lower panels). Note the significant co-localization of IL1� or IFN-� with THP(yellow color, arrows). IL1� and IFN-� were present in more renal tubules (arrowheads) than THP. All panels in C were �200 magnification.
THP Deficiency and Cytokine Imbalance
MAY 11, 2012 • VOLUME 287 • NUMBER 20 JOURNAL OF BIOLOGICAL CHEMISTRY 16373
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

To discern whether the increased activation of cytokine sig-naling in THP KO mice was due to increased renal cytokinesynthesis or increased cytokine reabsorption by or increasedinteractions with renal epithelial cells, we performed real-timePCRs of cytokine mRNA expression (Fig. 9B). The levels ofcytokine production were not significantly different betweenWT and KO mice. In addition, a mouse neutrophil-specificantibody, which strongly labeled neutrophils in a uropatho-genic E. coli-infected mouse bladder, did not detect apparentneutrophil infiltration in eitherWT or THP KOmice (Fig. 9C).These data are in favor of the idea that increased renal cytokinecontent (Figs. 6 and 8) results from increased reabsorption ofurinary cytokines and/or increased interaction of urinary cyto-kines with renal epithelial cells in the absence of THP.
DISCUSSION
Functional THP Regulates Serum Cytokine Levels—One ofmajor findings we made in this study was that the loss of THP,a urinary proteinmade exclusively by kidney epithelial cells, ledto a marked elevation of circulating cytokines in THP KOmice(Figs. 2 and 3). This occurred reproducibly in multiple cohortsand both at steady-state (Fig. 2) and after LPS challenge (Fig. 3);it affected most cytokines examined (Figs. 2 and 3) and coin-cided with a prominent, nonrenal organ anomaly, e.g. spleno-megaly (Fig. 1). The increase in serum cytokines was notaccompanied by a corresponding increase in their urinaryexcretion (Fig. 4), suggesting decreased renal clearance, espe-cially for IFN-� and IL1�. Furthermore, our data indicate asignificantly reduced glomerular filtration rate in THP KOmice, as evidenced by the increased serum concentration of
cystatin C, a marker for the glomerular filtration function, aswell as decreased inulin and creatinine clearance (Fig. 5).Together, our data suggest that THP deficiency can cause sys-temic cytokine imbalance at least partially by affecting glomer-ular filtration. It should be noted that reduced GFR was alsoobserved in an independent THP KO model (59), althoughchanges involving circulating cytokines were not investigated.THP as Decoy Receptor for Urinary Cytokines—Our present
study also provided the first experimental evidence that THPformed complexes with cytokines in vivo. Whether THP iscapable of binding cytokines has been fiercely debated, mainlybecause virtually all previous binding studies were done in vitrousing recombinant cytokines (37, 60). Although recombinanthuman cytokines consistently bound to purified THP, thisresult could not be reproduced with native cytokines (43). Inthe present study, we employed four independent methods, e.g.co-IP, native gel electrophoresis, co-sedimentation, and doubleimmunofluoresent staining, using in vivo isogenic materials,e.g.mouse native THP and mouse native cytokines (Figs. 6 and7). We demonstrated that THP-cytokine complexes existed insitu in wild-type mice. In stark contrast, such complexes werelost during THP deficiency. In addition, THP appears to bindboth inflammatory and noninflammatory cytokines, suggestingthat it is the chemical properties of THP that determine itspromiscuousness in cytokine interaction (see later). Further-more, by comparing the renal cytokine levels of wild-type miceversus those of THP KO mice and determining the cytokinesignaling pathways, we showed that the unbound urinary cyto-kines inTHPKOmicewere capable of binding to and activatingrenal epithelial cells (Figs. 8 and 9). Based on these results, wepropose a new role for THP as a urinary trap that serves tointeract with and inactivating glomerulus-filtered cytokines,thus preventing them from activating renal epithelial andurothelial cells. The trapping role played by urinary THP maynot be limited to cytokines. Previous in vitro studies suggestedthat THP also binds to complement C1 and C1q (61–63). It ispossible that THP is a promiscuous urinary trap not only forcytokines, but also for other immunological effectors. The pre-viously noted IgG-THP interaction that has thus far beenshown only in vitro (64) could have presented technical prob-lems in ELISA, Western blotting, and immunohistochemistyinvolving THP-containing materials from wild-type mousekidneys.However, itmust be emphasized thatwe includedmul-tiple controls in our experiments, in particular the isotypenonimmune antibodies to replace THP-specific antibodieswherever applicable.We found no evidence of any nonimmuneIgG-THP binding using in vivomaterials.It appears that cytokines bind with THP via distinct struc-
tural determinants. In vitro studies showed that the high-man-nose moieties of THP are required for its interaction with IL1and TNF, because de-glycosylation or inclusion of free manno-ses effectively blocks the THP-cytokine interaction (65, 66).Interestingly, high-mannose glycosylation of THP is extremelyconserved, being present in THP isolated from all mammalsstudied to date (67, 68). This suggests a strong structural/func-tional relationship in the THP-cytokine interaction and adds tothe functional importance of THP-cytokine binding as an evo-lutionarily conserved mechanism in renal protection. It cannot
FIGURE 8. Renal cytokine content. A–F, total protein extracts from groups ofwild-type (WT) and THP KO mice were used to assess cytokine productionusing ELISA. The cytokine levels were expressed as a ratio to the total kidneyprotein input (e.g. pg of cytokine/mg of total proteins). Note that all the sixcytokines assayed without LPS challenge were higher in KO mice than in WTcontrols, although the increases for TNF-� and CXCL1 were not statisticallysignificant.
THP Deficiency and Cytokine Imbalance
16374 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 20 • MAY 11, 2012
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

FIGURE 9. Status of renal cytokine signaling pathway (A), renal cytokine production (B), and status of renal neutrophil infiltration (C) in WT and THPKO mice (all 3-months of age; n � 10 per genotype). A, Western blotting showing steady-state down-regulation of I�B and up-regulation of NF-�B (p65) andphosphorylated Stat3 in renal protein extracts from THP KO mice, as compared with those from WT mice. B, real-time PCR quantification of renal mRNA levelsof IFN-�, IL1�, TNF-�, and IL6, showing that production of these cytokines was not significantly different between KO mice and WT mice. Values wereaverages � S.D. C, immunofluorescent staining of neutrophils in the kidneys of WT and THP KO mice using an antibody against a cell surface marker of mouseperipheral neutrophils. Top left panel, staining of a cross-section of a mouse urinary bladder experimentally inoculated with a uropathogenic E. coli strain (strainUTI89; 12-h post-infection), as a positive control showing intense labeling of multiple neutrophils in the bladder mucosa. Middle and right panels, representativeimages of renal cortex (top) and outer medullary regions (bottom) from WT and THP KO mice. Arrows denote neutrophils. Lower left panel, neutrophil count per10 high power fields (HPF; at �400 magnification). Note that neutrophils were rarely detected in the kidneys of both WT and THP KO mice.
THP Deficiency and Cytokine Imbalance
MAY 11, 2012 • VOLUME 287 • NUMBER 20 JOURNAL OF BIOLOGICAL CHEMISTRY 16375
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

be ruled out that binding affinities between THP and differentcytokines may vary depending on different primary structuresand secondary modifications of the cytokines.Unifying Model of THP in Regulating Local and Systemic
Immunity—On the basis of our current data and those pub-lished previously, we propose a model that depicts from amechanistic standpoint how the kidney acts as a major cata-bolic organ for cytokines. Normally, small sized cytokines passfreely through the glomerular filtration barrier (7). This filtra-tion process is regulated by a functional THP, which ensureshigh glomerular clearance, hence low circulating cytokines.Once filtered through the glomeruli, the bulk of urinary cyto-kines is retrieved by the proximal tubules and degraded via thelysosomal pathway (14, 15). Those that have escaped the uptakeand/or exceeded the absorption limit move downstream withthe urine flow but are soon captured by urinary and/or mem-brane-bound THP at the TAL. Although free cytokines arecapable of binding to and overactivating the renal epithelial andurothelial cells (18, 19), THP binding traps the free cytokines,thus blocking their biological activities. The trapping effectmayalso prevent the uptake of cytokines by the distal tubular cells asrecently suggested (69) or by para-cellular re-absorption (16),although the existence or lack thereof of the latter route needsexperimental validation. Our model underscores the criticalimportance of THP, a urinary protein normally insulated fromthe immune system, in ensuring both renal protection and sys-temic immune homeostasis. In emphasizing the physiologicalroles of THP in immune suppression, we are by nomeans rulingout its immune stimulatory effect under certain pathologicalconditions (44, 70, 71). Such immune stimulatory responses aredistinguishable from the suppressive functions of THP in con-text, biological end points, and clinical correlates.THP Deficiency-mediated Cytokine Imbalance as Key Medi-
ator of Chronic Kidney Diseases and Multiorgan Failure—Evi-dence is accumulating that THP deficiency, both qualitativeand quantitative, exists in a wide spectrum of pathophysiolog-ical conditions in humans. Germline mutations in the THPgene, which result in THP misfolding and failure to exit fromthe endoplasmic reticulum, cause juvenile-onset hyperurice-mic nephropathy, type II medullary kidney cystic disease, andglomerulocystic disease (25, 72, 73). Single nucleotide polymor-phisms in the promoter and coding regions of the human THPgene, which may affect THP gene expression and function, arestrongly associatedwith chronic kidney disease (51). Decreasedurinary THP excretion has also been found in type I diabetes,acute tubular necrosis, and hyperprostaglandin E syndrome(74–79). The exact cause and pathogenesis of many of thesediseases remain unclear. Based on our data, it can be speculatedthat the local and systemic cytokine imbalance caused by THPdeficiency could be amediator in the genesis and progression ofsome of these conditions. Given the fact that cytokines play akey role inmultiorgan failure caused by renal insufficiency (80),THP deficiency could also contribute to this complicatedpathophysiological process for which little is currently known.Elucidating how renal cytokine homeostasis can be maintainedduring the various pathophysiological conditions will likelyprovide insights into improved therapeutic options.
Acknowledgment—We thank Dr. Duane Hickling, Department ofUrology, New York University School of Medicine, for providing amouse urinary bladder inoculated with a uropathogenic E. coli strainUTI89 as a positive control.
REFERENCES1. O’Shea, J. J., Ma, A., and Lipsky, P. (2002) Cytokines and autoimmunity.
Nat. Rev. Immunol. 2, 37–452. Thomson,A.W., and Lotze,M.T. (2003)TheCytokineHandbook, 4th Ed.,
Academic Press, Boston, MA3. Ozaki, K., and Leonard, W. J. (2002) Cytokine and cytokine receptor plei-
otropy and redundancy. J. Biol. Chem. 277, 29355–293584. Rittirsch, D., Flierl, M. A., and Ward, P. A. (2008) Harmful molecular
mechanisms in sepsis. Nat. Rev. Immunol. 8, 776–7875. Ratsimandresy, R. A., Rappaport, J., and Zagury, J. F. (2009) Anti-cytokine
therapeutics. History and update. Curr. Pharm. Des. 15, 1998–20256. Li, X., Hassoun, H. T., Santora, R., and Rabb, H. (2009) Organ cross-talk,
The role of the kidney. Curr. Opin. Crit. Care 15, 481–4877. Dworkin, L. D., and Brenner, B. M. (1992) in The Kidney: Physiology and
Pathophysiology (Seldin, D.W., andGiebish, G., eds) pp. 979–1016, RavenPress, New York
8. Kudo, S., Mizuno, K., Hirai, Y., and Shimizu, T. (1990) Clearance andtissue distribution of recombinant human interleukin 1� in rats. CancerRes. 50, 5751–5755
9. Bocci, V., Pacini, A., Muscettola, M., Pessina, G. P., Paulesu, L., andBandinelli, L. (1982) The kidney is the main site of interferon catabolism.J. Interferon Res. 2, 309–314
10. Greig, N. H., Soncrant, T. T., Wozniak, K. M., and Rapoport, S. I. (1988)Plasma and tissue pharmacokinetics of human interferon-� in the rat afterits intravenous administration. J. Pharmacol. Exp. Ther. 245, 574–580
11. Bemelmans, M. H., van Tits, L. J., and Buurman, W. A. (1996) Tumornecrosis factor, function, release, and clearance. Crit. Rev. Immunol. 16,1–11
12. Bocci, V., Mogensen, K. E., Muscettola,M., Pacini, A., Paulesu, L., Pessina,G. P., and Skiftas, S. (1983) Degradation of human 125I-interferon � byisolated perfused rabbit kidney and liver. J. Lab. Clin. Med. 101, 857–863
13. Newton, R. C., Uhl, J., Covington,M., and Back,O. (1988) The distributionand clearance of radiolabeled human interleukin-1� in mice. LymphokineRes. 7, 207–216
14. Kudo, S., and Goto, H. (1999) Intrarenal handling of recombinant humaninterleukin-1� in rats. Mechanism for proximal tubular protein reabsorp-tion. J. Interferon Cytokine Res. 19, 1161–1168
15. Nakamura, A., Suzuki, T., and Kohsaka, T. (1995) Renal tubular functionmodulates urinary levels of interleukin-6. Nephron 70, 416–420
16. Katz, A. I., and Emmanouel, D. S. (1978) Metabolism of polypeptide hor-mones by the normal kidney and in uremia. Nephron 22, 69–80
17. Wang, S. N., Lapage, J., and Hirschberg, R. (1999) Glomerular ultrafiltra-tion and apical tubular action of IGF-I, TGF-�, and HGF in nephroticsyndrome. Kidney Int. 56, 1247–1251
18. Benigni, A., and Remuzzi, G. (2001) How renal cytokines and growthfactors contribute to renal disease progression. Am. J. Kidney Dis. 37,S21–24
19. Benigni, A., Zoja, C., and Remuzzi, G. (1995) The renal toxicity of sus-tained glomerular protein traffic. Lab. Invest. 73, 461–468
20. Serafini-Cessi, F.,Malagolini, N., andCavallone, D. (2003) Tamm-Horsfallglycoprotein. Biology and clinical relevance. Am. J. Kidney Dis. 42,658–676
21. Devuyst, O., Dahan, K., and Pirson, Y. (2005) Tamm-Horsfall protein oruromodulin. New ideas about an old molecule.Nephrol. Dial Transplant.20, 1290–1294
22. Säemann, M. D., Weichhart, T., Hörl, W. H., and Zlabinger, G. J. (2005)Tamm-Horsfall protein. A multilayered defence molecule against urinarytract infection. Eur. J. Clin. Invest. 35, 227–235
23. Kokot, F., andDuława, J. (2000)Tamm-Horsfall protein updated.Nephron85, 97–102
24. Kumar, S., and Muchmore, A. (1990) Tamm-Horsfall protein, uromodu-
THP Deficiency and Cytokine Imbalance
16376 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 20 • MAY 11, 2012
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

lin (1950–1990). Kidney Int. 37, 1395–140125. Rampoldi, L., Scolari, F., Amoroso, A., Ghiggeri, G., and Devuyst, O.
(2011) The rediscovery of uromodulin (Tamm-Horsfall protein): Fromtubulointerstitial nephropathy to chronic kidney disease. Kidney Int. 80,338–347
26. Rindler, M. J., Naik, S. S., Li, N., Hoops, T. C., and Peraldi, M. N. (1990)Uromodulin (Tamm-Horsfall glycoprotein/uromucoid) is a phosphatidyl-inositol-linked membrane protein. J. Biol. Chem. 265, 20784–20789
27. Fukuoka, S., andKobayashi, K. (2001) Analysis of the C-terminal structureof urinary Tamm-Horsfall protein reveals that the release of the glycosylphosphatidylinositol-anchored counterpart from the kidney occurs byphenylalanine-specific proteolysis. Biochem. Biophys. Res. Commun. 289,1044–1048
28. Dawnay, A. B., and Cattell, W. R. (1981) Serum Tamm-Horsfall glycopro-tein levels in health and renal disease. Clin. Nephrol. 15, 5–8
29. Phimister, G. M., andMarshall, R. D. (1983) Tamm-Horsfall glycoproteinin human amniotic fluid. Clin. Chim. Acta 128, 261–269
30. Zhu, X., Cheng, J., Gao, J., Lepor, H., Zhang, Z. T., Pak, J., and Wu, X. R.(2002) Isolation of mouse THP gene promoter and demonstration of itskidney-specific activity in transgenic mice. Am. J. Physiol. Renal Physiol.282, F608–617
31. Zhu, X., Cheng, J., Huang, L., Gao, J., Zhang, Z. T., Pak, J., and Wu, X. R.(2003) Renal tubule-specific expression and urinary secretion of humangrowth hormone. A kidney-based transgenic bioreactor growth. Trans-genic Res. 12, 155–162
32. Pennica, D., Kohr, W. J., Kuang, W. J., Glaister, D., Aggarwal, B. B., Chen,E. Y., andGoeddel, D.V. (1987) Identification of humanuromodulin as theTamm-Horsfall urinary glycoprotein. Science 236, 83–88
33. Hession, C., Decker, J. M., Sherblom, A. P., Kumar, S., Yue, C. C., Mattal-iano, R. J., Tizard, R., Kawashima, E., Schmeissner, U., and Heletky, S.(1987) Uromodulin (Tamm-Horsfall glycoprotein), a renal ligand for lym-phokines. Science 237, 1479–1484
34. Kumar, S. (1998) Are Tamm-Horsfall protein and uromodulin identical?Eur. J. Clin. Invest. 28, 483–484
35. Olczak, T., Kubicz, A., Kokot, F., and Duława, J. (1998) Tamm-Horsfallprotein isolated from urine of pregnant and nonpregnant women hassimilar oligosaccharides. Eur. J. Clin. Invest. 28, 475–482
36. Smagula, R.M., VanHalbeek, H., Decker, J. M.,Muchmore, A. V.,Moody,C. E., and Sherblom, A. P. (1990) Pregnancy-associated changes in oligo-mannose oligosaccharides of human and bovine uromodulin (Tamm-Horsfall glycoprotein). Glycoconj. J. 7, 609–624
37. Sherblom,A. P., Decker, J.M., andMuchmore, A. V. (1988) The lectin-likeinteraction between recombinant tumor necrosis factor and uromodulin.J. Biol. Chem. 263, 5418–5424
38. Rosenstreich, D. L., Yost, S. L., and Brown, K. M. (1987) Human urine-derived inhibitors of interleukin 1. Rev. Infect. Dis. 9, S594–598
39. Muchmore, A. V., Shifrin, S., and Decker, J. M. (1987) In vitro evidencethat carbohydrate moieties derived from uromodulin, an 85,000-daltonimmunosuppressive glycoprotein isolated from human pregnancy urine,are immunosuppressive in the absence of intact protein. J. Immunol. 138,2547–2553
40. Muchmore, A. V., and Decker, J. M. (1986) Uromodulin. An immunosup-pressive 85-kilodalton glycoprotein isolated from human pregnancy urineis a high affinity ligand for recombinant interleukin 1�. J. Biol. Chem. 261,13404–13407
41. Nesselhut, T., Rath, W., Grospietsch, G., Weber, M. H., and Kuhn, W.(1989) Urinary protein electrophoresis profile in normal and hypertensivepregnancies. Arch. Gynecol. Obstet. 246, 97–105
42. Nesselhut, T., Rath, W., Grunow, E., Kaufholz, G., Ostermai, U., Cillien,N., and Kuhn, W. (1993) The relationship between urinary Tamm-Hors-fall glycoprotein excretion and urinary activity of glycosidases in normalpregnancy and pre-eclampsia. Eur. J. Obstet. Gynecol. Reprod. Biol. 48,23–31
43. Moonen, P., Gaffner, R., andWingfield, P. (1988) Native cytokines do notbind to uromodulin (Tamm-Horsfall glycoprotein). FEBS Lett. 226,314–318
44. Su, S. J., Chang, K. L., Lin, T. M., Huang, Y. H., and Yeh, T. M. (1997)Uromodulin and Tamm-Horsfall protein induce human monocytes to
secrete TNF and express tissue factor. J. Immunol. 158, 3449–345645. Su, S. J., and Yeh, T. M. (1999) The dynamic responses of pro-inflamma-
tory and anti-inflammatory cytokines of human mononuclear cells in-duced by uromodulin. Life Sci. 65, 2581–2590
46. Säemann, M. D., Weichhart, T., Zeyda, M., Staffler, G., Schunn, M.,Stuhlmeier, K. M., Sobanov, Y., Stulnig, T. M., Akira, S., von Gabain, A.,von Ahsen, U., Hörl, W. H., and Zlabinger, G. J. (2005) Tamm-Horsfallglycoprotein links innate immune cell activation with adaptive immunityvia a Toll-like receptor-4-dependent mechanism. J. Clin. Invest. 115,468–475
47. El-Achkar, T.M.,Wu, X. R., Rauchman,M.,McCracken, R., Kiefer, S., andDagher, P. C. (2008) Tamm-Horsfall protein protects the kidney fromischemic injury by decreasing inflammation and altering TLR4 expres-sion. Am. J. Physiol. Renal Physiol. 295, F534–544
48. El-Achkar, T. M., McCracken, R., Rauchman, M., Heitmeier, M. R., Al-Aly, Z., Dagher, P. C., and Wu, X. R. (2011) Tamm-Horsfall protein-defi-cient thick ascending limbs promote injury to neighboring S3 segments inan MIP-2-dependent mechanism. Am. J. Physiol. Renal Physiol. 300,F999–1007
49. Mo, L., Zhu, X. H., Huang, H. Y., Shapiro, E., Hasty, D. L., and Wu, X. R.(2004) Ablation of the Tamm-Horsfall protein gene increases susceptibil-ity of mice to bladder colonization by type 1-fimbriated Escherichia coli.Am. J. Physiol. Renal Physiol. 286, F795–802
50. Séronie-Vivien, S., Delanaye, P., Piéroni, L., Mariat, C., Froissart, M., Cris-tol, J. P., and SFBC “Biology of Renal Function and Renal Failure”WorkingGroup (2008) Cystatin C, current position and future prospects. Clin.Chem. Lab. Med. 46, 1664–1686
51. Kottgen, A., Glazer, N. L., Dehghan, A., Hwang, S. J., Katz, R., Li, M., Yang,Q., Gudnason, V., Launer, L. J., Harris, T. B., Smith, A. V., Arking, D. E.,Astor, B. C., Boerwinkle, E., Ehret, G. B., Ruczinski, I., Scharpf, R. B., IdaChen, Y. D., de Boer, I. H., Haritunians, T., Lumley, T., Sarnak, M., Sisco-vick, D., Benjamin, E. J., Levy, D., Upadhyay, A., Aulchenko, Y. S., Hofman,A., Rivadeneira, F., Uitterlinden, A. G., van Duijn, C. M., Chasman, D. I.,Pare, G., Ridker, P. M., Kao, W. H., Witteman, J. C., Coresh, J., Shlipak,M. G., and Fox, C. S. (2009) Multiple loci associated with indices of renalfunction and chronic kidney disease. Nat. Genet. 41, 712–717
52. Qi, Z., Whitt, I., Mehta, A., Jin, J., Zhao, M., Harris, R. C., Fogo, A. B., andBreyer, M. D. (2004) Serial determination of glomerular filtration rate inconsciousmice using FITC-inulin clearance.Am. J. Physiol. Renal Physiol.286, F590–596
53. Sturgeon, C., Sam, A. D., 2nd, and Law,W. R. (1998) Rapid determinationof glomerular filtration rate by single-bolus inulin. A comparison of esti-mation analyses. J. Appl. Physiol. 84, 2154–2162
54. Cavallone, D., Malagolini, N., Monti, A., Wu, X. R., and Serafini-Cessi, F.(2004) Variation of high mannose chains of Tamm-Horsfall glycoproteinconfers differential binding to type 1-fimbriated Escherichia coli. J. Biol.Chem. 279, 216–222
55. Mosser, D. M., and Edwards, J. P. (2008) Exploring the full spectrum ofmacrophage activation. Nat. Rev. Immunol. 8, 958–969
56. Gilbert, R. E., Game, D. S., and Advani, A. (2011) in Brenner and Rector’sTheKidney (Taal,M.W., Chertow,G.M.,Marsden, P. A., Skorecki, K., Yu,A. S. L., and Brenner, B. M., eds) pp. 384–420, Elsevier, Philadelphia, PA
57. Ichikawa, I., and Brenner, B. M. (1984) Glomerular actions of angiotensinII. Am. J. Med. 76, 43–49
58. Jovine, L., Qi, H., Williams, Z., Litscher, E., and Wassarman, P. M. (2002)The ZP domain is a conserved module for polymerization of extracellularproteins. Nat. Cell Biol. 4, 457–461
59. Bachmann, S., Mutig, K., Bates, J., Welker, P., Geist, B., Gross, V., Luft,F. C., Alenina, N., Bader, M., Thiele, B. J., Prasadan, K., Raffi, H. S., andKumar, S. (2005) Renal effects of Tamm-Horsfall protein (uromodulin)deficiency in mice. Am. J. Physiol. Renal Physiol. 288, F559–567
60. Muchmore, A. V. (1986) Uromodulin, an immunoregulatory glycoproteinisolated from pregnancy urine that binds to and regulates the activity ofinterleukin 1. Am. J. Reprod. Immunol. Microbiol. 11, 89–93
61. Rhodes, D. C. (2000) Binding of Tamm-Horsfall protein to complement1q measured by ELISA and resonant mirror biosensor techniques undervarious ionic strength conditions. Immunol. Cell Biol. 78, 474–482
62. Rhodes, D. C. (2002) Binding of Tamm-Horsfall protein to complement
THP Deficiency and Cytokine Imbalance
MAY 11, 2012 • VOLUME 287 • NUMBER 20 JOURNAL OF BIOLOGICAL CHEMISTRY 16377
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

1q and complement 1, including influence of hydrogen-ion concentration.Immunol. Cell Biol. 80, 558–566
63. Rhodes, D. C. (2006) Importance of carbohydrate in the interaction ofTamm-Horsfall protein with complement 1q and inhibition of classicalcomplement activation. Immunol. Cell Biol. 84, 357–365
64. Rhodes, D. C., Hinsman, E. J., and Rhodes, J. A. (1999) Errors in reportedassociation between Tamm-Horsfall protein and IgG. Kidney Int. 56, 759
65. Muchmore, A. V., Sathyamoorthy, N., Decker, J., and Sherblom, A. P.(1990) Evidence that specific high-mannose oligosaccharides can directlyinhibit antigen-driven T-cell responses. J. Leukoc. Biol. 48, 457–464
66. Dall’Olio, F., Chiricolo, M., Malagolini, N., Franceschi, C., and Serafini-Cessi, F. (1991) Immunosuppressive activity of Tamm-Horsfall glycopro-tein oligosaccharides. Effect of removal of outer sugars and conjugationwith a protein carrier. Cell. Immunol. 137, 303–315
67. Pak, J., Pu, Y., Zhang, Z. T., Hasty, D. L., and Wu, X. R. (2001) Tamm-Horsfall protein binds to type 1 fimbriated Escherichia coli and prevents E.coli from binding to uroplakin Ia and Ib receptors. J. Biol. Chem. 276,9924–9930
68. Serafini-Cessi, F., Monti, A., and Cavallone, D. (2005) N-Glycans carriedby Tamm-Horsfall glycoprotein have a crucial role in the defense againsturinary tract diseases. Glycoconj. J. 22, 383–394
69. Langelueddecke, C., Roussa, E., Fenton, R. A.,Wolff, N. A., Lee,W. K., andThévenod, F. (2012) Lipocalin-2 (24p3/neutrophil gelatinase-associatedlipocalin (NGAL)) receptor is expressed in distal nephron and mediatesprotein endocytosis. J. Biol. Chem. 287, 159–169
70. Kreft, B., Jabs, W. J., Laskay, T., Klinger, M., Solbach, W., Kumar, S., andvan Zandbergen, G. (2002) Polarized expression of Tamm-Horsfall pro-tein by renal tubular epithelial cells activates human granulocytes. Infect.Immun. 70, 2650–2656
71. Dong, X., Swaminathan, S., Bachman, L. A., Croatt, A. J., Nath, K. A., andGriffin, M. D. (2005) Antigen presentation by dendritic cells in renal
lymph nodes is linked to systemic and local injury to the kidney. KidneyInt. 68, 1096–1108
72. Scolari, F., Caridi, G., Rampoldi, L., Tardanico, R., Izzi, C., Pirulli, D.,Amoroso, A., Casari, G., and Ghiggeri, G. M. (2004) Uromodulin storagediseases. Clinical aspects andmechanisms.Am. J. Kidney Dis. 44, 987–999
73. Bleyer, A. J., Trachtman, H., Sandhu, J., Gorry, M. C., and Hart, T. C.(2003) Renal manifestations of a mutation in the uromodulin (Tamm-Horsfall protein) gene. Am. J. Kidney Dis. 42, E20–26
74. Schröter, J., Timmermans, G., Seyberth, H.W., Greven, J., and Bachmann,S. (1993)Marked reduction of Tamm-Horsfall protein synthesis in hyper-prostaglandin E-syndrome. Kidney Int. 44, 401–410
75. Torffvit, O., Agardh, C. D., and Thulin, T. (1999) A study of Tamm-Horsfall protein excretion in hypertensive patients and type 1 diabeticpatients. Scand. J. Urol. Nephrol. 33, 187–191
76. Torffvit, O., and Agardh, C. D. (1994) Urinary excretion rate of NC1 andTamm-Horsfall protein in the microalbuminuric type I diabetic patient. J.Diabetes Complications 8, 77–83
77. Tsai, C. Y., Wu, T. H., Yu, C. L., Lu, J. Y., and Tsai, Y. Y. (2000) Increasedexcretions of �2-microglobulin, IL6, and IL8 and decreased excretion ofTamm-Horsfall glycoprotein in urine of patients with active lupus nephri-tis. Nephron. 85, 207–214
78. Sejdiu, I., and Torffvit, O. (2008) Decreased urinary concentration ofTamm-Horsfall protein is associated with development of renal failureand cardiovascular death within 20 years in type 1 but not in type 2 dia-betic patients. Scand. J. Urol. Nephrol. 42, 168–174
79. Bernard, A. M., Ouled, A. A., Lauwerys, R. R., Lambert, A., and Van-deleene, B. (1987) Pronounced decrease of Tamm-Horsfall proteinuria indiabetics. Clin. Chem. 33, 1264
80. Carrero, J. J., Yilmaz, M. I., Lindholm, B., and Stenvinkel, P. (2008) Cyto-kine dysregulation in chronic kidney disease. How can we treat it? BloodPurif. 26, 291–299
THP Deficiency and Cytokine Imbalance
16378 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 20 • MAY 11, 2012
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Yan Liu, Tarek M. El-Achkar and Xue-Ru WuGlomerular Filtration Rate and Acting as a Urinary Cytokine Trap
Tamm-Horsfall Protein Regulates Circulating and Renal Cytokines by Affecting
doi: 10.1074/jbc.M112.348243 originally published online March 26, 20122012, 287:16365-16378.J. Biol. Chem.
10.1074/jbc.M112.348243Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/287/20/16365.full.html#ref-list-1
This article cites 77 references, 16 of which can be accessed free at
by guest on May 27, 2018
http://ww
w.jbc.org/
Dow
nloaded from