studies of atrazine effects on dna in mice...
TRANSCRIPT

STUDIES OF ATRAZINE EFFECTS ON DNA IN MICE BY CHROMOSOMAL ABERRATION TEST AND AGAROSE GEL ELECTROPHORESIS ASSAY
Iman Abd ElMoneim Darwish Mahmoud
J. Egypt. Soc. Toxicol. (Vol. 39: 25-38 July 2008)
WWW.jest.eg.net/ Zoology Department, Women's College for Arts, Science and Education, Ain-Shams University, Cairo, Egypt
ABSTRACT The herbicide atrazine was tested for its ability to induce chromosomal
aberrations in mouse bone marrow. Also, the alkaline single cell gel electrophoresis was used to measure DNA strand breaks in both kidney and liver tissues. Mice were given orally a single dose of 28.125, 56.25, 112.5 and 225 mg/kg body weight (1/64, 1/32, 1/16 and 1/8 of LD50) and repeated daily doses of atrazine. Mice were sacrificed 24 hrs after the last dose.A slight increase chromosomal aberration was observed for the vehicle group (corn oil) compared with the negative control (untreated mice). A significant increase in the percentage of chromosome aberrations in bone marrow was observed after oral administration of atrazine at 56.25, 112.5, and 225 mg/kg body weight for 24 hours. The mean of mitotic index induced by this herbicide did not differe significantly from that observed in the negative control. Atrazine – induced numerical chromosomal aberrations in 24h treatment with a dose-response relationship. Moreover, the present data clearly indicate that atrazine possesses the potential, at least to a limited extent to cause alterations in the cellular DNA in both kidney and liver tissues. In conclusion, the induction of chromosome aberrations and DNA damage following exposure to atrazine indicates a potential for clastogenicity.
INTRODUCTION Atrazine (2-chloro 4 (ethylamino) 6-
(isopropylamino) 1,3,5 triazine) is one of the most commonly used herbicides in North America is frequently detected in ground and surface water (Dooley et al., 2006). Atrazine is the most heavily applied agricultural pesticide for crop production in the United States. Both animal and human studies have suggested that atrazine is possibly carcinogenic, but results have been inconclusive (Rusiecki et al., 2004).
The three herbicides atrazine, cyanazine or simazine significantly increased the rate of apparent dominant lethals, x-linked recessive lethals and x or y loss after treatment by larval feeding in Drosophila melanogaster (Murnik and Nash, 1977). Nishi et al. (1979) suggested that maleic hydrazide and its salts had weak inducibility of cytotoxicity and positive cytogenetic effects on Chinese hamster V79 cells in vitro. However, trifluralin and simazine, but not atrazine, significantly increased sister chromatid exchanges per cell with the highest concentrations in human lymphocyte (Ghiazza et al., 1984). Meschini et al. (1988) showed that maleic hydrazide was genotoxic in mammalian cells in vitro but apparently
not in vivo. In 1992, Meisner et al. demonstrated dose- related cytogenetic damage not associated with mitotic inhibition or cell death due to the alachlor-atrazine combination suggesting an additive model in vitro.
Roloff et al. (1992) showed little chromosome damage in human lymphocytes exposed in vitro to either linuron or atrazine, whereas significant chromosome damage was observed in lymphocytes simultaneously exposed to linuron and atrazine. In 1993, Kligerman et al. found a concentration related increase in sister-chromatid exchange in splenocytes of rat and mouse exposed through their drinking water to a mixture of pesticides (aldicarb, atrazine, dibromochloropropane, 1,2-dichloropropane, ethylene dibromide and simazine) that simulated contaminated groundwater in California. Exposing human lymphocyte cultures to concentrations of N-nitrosoatrazine (NNAT) as low as 0.0001 microgram/ml results in significant elevations in chromosome breakage as well as an increased mitotic index (Meisner et al., 1993). Moreover, Lioi et al. (1998) observed a dose- related increase in the percent of aberrant cells, an increase of sister chromatid exchanges/cells, a significant reduction of the mitotic index, and increase of glucose-6-

Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
26
phosphate dehydrogenase (G6PD) enzyme activity in human peripheral lymphocytes exposed in vitro to glyphosate, vinclozolin, atrazine and DPX-E9636. In contrast, Ribas et al. (1998) concluded that atrazine was able to exert a weak cytotoxic effect using sister- chromatid exchanges, chromosome aberrations and micronuclei in cultured human peripheral blood lymphocytes.
Taets et al. (1998) observed karyotype damage in Chinese hamster ovary (CHO) cells with a combination of atrazine and simazine or atrazine and cyanazine as measured by flow cytometry. Kligerman et al.(2000)showed that atrazine, simazine, and cyanazine were failed to produce any significant increases in sister chromatid exchanges or chromosomal aberrations in human lymphocyte cultures at the limits of solubility and toxicity. Garaj-Vrhovac and Zeljezic (2001) showed an increased number of chromosomal aberrations, sister chromatid exchange (SCE) frequency, micronucleus (MN) frequency and values of Comet assay parameters in Croatian workers occupationally exposed to a complex mixture of pesticides containing atrazine, alachlor, cyanazine, 2,4- dichlorophenoxy-acetic acid and malathion. In addition, at extremely low levels, atrazine was found to damage animals chromosomes in cell culture as mentioned by Rayburn et al. (2001).
The mean value of sister chromatid exchange, chromatid and chromosome breaks, micronuclei and DNA migration in workers occupationally exposed to a mixture of pesticides (atrazine, alachlor, cyanazine, 2,4- dichlorophenoxyacetic acid and malathion) were significantly higher in the comparison with the control group (Garaj-Vrhovac and Zeljezic, 2002 and Zeljezic and Garaj-Vrhovac, 2002). Moreover, Malik et al. (2004) demonstrate a does'nt dependent increase of SCE in cultured lymphocytes after exposure to atrazine. In 2006, Bouilly et al. showed that the chromosome pairs (1,5,9 and 10) were affected by the loss of 1 chromosome (61%, 15%, 42% and 42% respectively) in the progeny of oysters contaminated by atrazine using the restriction enzyme (HaeIII) digestion chromosome banding.
On the other hand, atrazine affected the circadian rhythms of RNA; DNA and total protein differently, in relation to the specific endocrine tissue of male rats (Nicolau and Socoliuc, 1980). Pino et al. (1988) detected DNA breaks in cell suspensions obtained from stomach, kidney and liver, but not in those from lung by the DNA alkaline elution technique in rats given orally a single high dose or repeated daily doses of atrazine. In 1997, Clements et al. indicated that the herbicide atrazine currently used in Southern Ontario (Canada) is capable of inducing DNA damage in tadpoles using the alkaline single cell gel DNA electrophoresis (SCGE) or Comet assay. In contrast, no significant increase in maleic hydrazide- induced DNA damage, as measured by the Comet assay, could be demonstrated in either leaves
of tobacco (Nicotiana tabacum var.xanthi) or roots of field beans, Vicia faba (Gichner et al., 2000).
Moreover, the Comet assay on nuclei from Tradescantia staminal hairs is a useful tool to monitor genotoxic agents such as maleic hydrazide as concluded by Alvarez-Moya el al. (2001). Tennant et al. (2001) indicated that atrazine was induced a small dose- related increase in DNA damage in mice using the alkaline single cell gel electrophoresis (SCGE) assay. Zeljezic and Garaj-Vrhovac (2001) found statistically increased levels of DNA damage in the Comet assay in terms of tail length and tail moment after the period of high exposure to a mixture of pesticides. Undeger and Basaran (2002) observed the DNA damage in lymphocytes of pesticide- exposed workers was significantly higher than that in the controls (P< 0.001).
Zeljezic and Garaj-Vrhovac (2004) found statistically significant increase of tail length for the blood samples, kidney, liver, bone marrow and spleen examined in mice treated with two pesticide formulations (Bravo and Gesaprim) containing alachlor and atrazine as active ingredients compared to the control. For both pesticides DNA of kidney and liver showed largest increase in migration. Chang et al. (2005) stated that both alachlor and atrazine were showed dose related increases in DNA damage in blood cells of common carp (Cyprinus carpio) using the single – cell gel electrophoresis method at environmentally relevant concentrations, (<100ppb). In addition, Monroy et al. (2005) showed DNA damage in normal human cells (GM38) and human fibrosarcoma (HT1080) cells at glyphosate concentrations of 4.0-6.5 and 4.75-5.75, mM respectively using the Comet assay. Recently, Bhalli et al. (2006) indicated that occupational exposure to pesticides causes DNA damage with the single cell gel electrophoresis (SCGE) assay or Comet assay.
In the present study, the genotoxic potential of atrazine, as revealed by bone marrow chromosome aberration and DNA amount using the agarose gel electrophoresis assay in both liver and kidney tissues of mice is reported.
MATERIALS AND METHODS Animals:
Male mice, aged 10-12 weeks and weighing 25-30g were used. All animals were kept under the same laboratory conditions of temperature (20±2oC) under 12 hrs light/dark cycle and 50% humidity. The mice were received standard laboratory chow and water ad libitum. Six animals were used for each treatment and control group.
Test agent: Atrazine (CAS No 1912-24-9, purity 97.7% with
the empirical formula C8 H 14 CIN5) was obtained from Sigma Chemical Company.

Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
27
Doses: The herbicide was tested in male mice in four
doses approximately 1/64, 1/32, 1/16, and 1/8 of published LD50 values (Gebel et al., 1997). Atrazine was dissolved in corn oil and administered orally into mice at dose levels of 28.125, 56.25, 112.5 and 225 mg/kg body weight. Gavage volumes were 0.1 ml per 10 g body weight. For negative control, corn oil only was used.
Chromosomal aberrations (CAs) assay: Mice were administered with a single dose of
28.125, 56.25, 112.5 and 225 mg/kg body weight via the oral gavage route for 24 hours and chromosome preparations were made as described earlier (Agarwal et al., 1994). Mice were injected (i.p.) with colchicine (4 mg/kg) 2 hrs prior to sacrifice. The bone marrow cells were aspirated in buffer solution and centrifuged at 1000 rpm for 10 minutes. The pellets obtained were mixed in aqueous solution of KCl and left for 20 min at 37ºC. Cells were recentrifuged, fixed in cold Carnoy's fluid (methanol – glacial acetic acid, 3:1) and dropped on clean chilled slides. Finally, slides were air dried and stained with 4% Giemsa.
Mitotic index (M.I.) was calculated from 1000 cells/animal and expressed in percentage (Rios et al., 1995).
Mitotic index = number of metaphases x 100 1000 cells
Statistical analysis: Data from control and treated animals for
mitotic preparation were evaluated with Student's t-test at p<0.05 (Snedecor and Cochran, 1980).
Preparation of genomic DNA from mouse tissue:
Mice injected with atrazine were sacrificed 24 hrs after the last injection and both liver and kidney tissues were removed for the determination of DNA content.
Prepare the tissues: 1- Excision and immediately mince of the tissue and freeze in liquid nitrogen.
2- Grind 250 mg tissue with hemogenizer. Suspended in 2.5ml digestion buffer (1MTris-HCl,1mM EDTA, 1% sodium dodecyl sulphate (SDS), 10 mg/ml pancreatic RNase or nucleic acid purification grade lysis).
DNA extract: 1-Samples were incubated, shaking in tightly capped tubes, 1 h at 37oC.
2-Then proteinase K was added followed by incubation at 37oC/12h.
3-Extract samples with an equal volume of phenol.
4-Add one volume of phenol/chlorophorm/isoamyl alchohol.Centrifuge at 10.000 rpm for 10 minutes (If phases do not resolve well, add another volume digestion buffer and repeat centrifugation. If thick white material appears at interface, repeat organic extraction). Transfer top layer (aqueous) to a new tube.
5- Add 1/10 vol of 5 M NaCl and 1vol of 100% cold ethanol. Centrifuge at 10.000 rpm for 2 minutes.
6- Wash with 70% ethanol, air dry and resuspended in Tris EDTA (TE, 10 mM Tris – HCl, 1 mM EDTA, pH 8.0) buffer at 1mg/ml (Gross-Bellard et al., 1972 and Sambrook et al., 1989).
Gel preparation: Gel was prepared using 1.8% electrophoretic
grade agarose (BRL). The agarose was boiled with tris borate EDTA buffer (1X TBE buffer, 89 mM tris, 89 mM boric acid, 2 mM EDTA, pH 8.3), and then 0.5 microgram / ml ethidium bromide was added to agarose mixture at 40oC. Gel was poured and allowed to solidify at room temperature for 1 h before samples were loaded. The gel was stained with ethidium bromide and detected by UV transilluminator. The gel is photographed with plaroid camera to obtain a permanent record of the band pattern in the gel (Aquadro et al., 1992).
RESULTS
In control mice, the mean frequency of chromosome aberrations per 50 cells was 3.83±1.47. Most of the cells (85.33%) have a diploid chromosome number 2N= 40. Treatment of mice with corn oil alone does not induce any significant effect on the bone marrow chromosomal aberrations as shown in Table (I).
The percentage of aberrant cells increased in a concentration- related manner after atrazine treatment, Fig. (1).
Atrazine-induced both chromosome type aberrations and chromatid-type aberrations at all concentrations tested. Even at lower concentrations (1/64 LD50) abnormal metaphases were observed at 1.39 times higher rate than those in untreated control level. About 17.33% of the examined cells showed one or another type of structural chromosomal anomalies following treatment with 1/32 LD50 (56.25 mg of atrazine). This value was increased to 18.33% at 112.5 mg/kg concentration. Chromosomal aberrations were observed in 18.67% of the metaphase of mice treated with 225 mg/kg body weight (1/8 LD50).

Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
28

Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
29
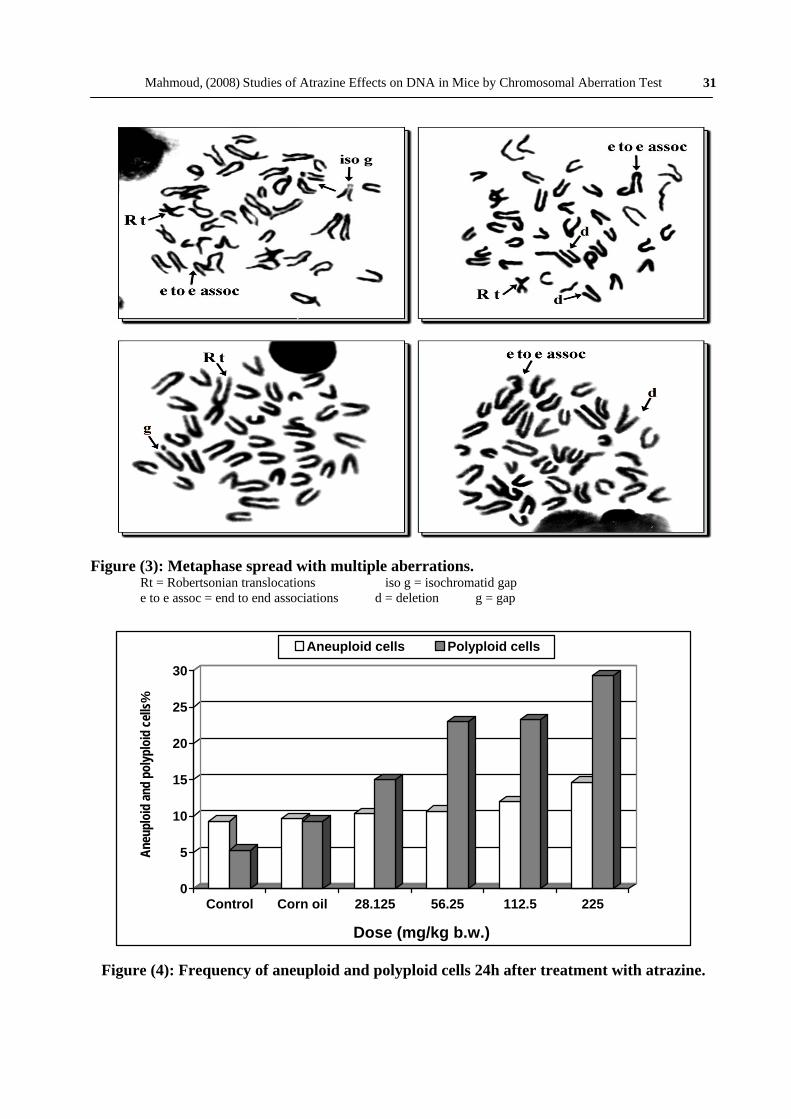
The centromeric attenuation, Robertsonian translocation and acentric fragments were the main aberrations noticed with atrazine. Secondary constrictions, acentric fragment, double minutes and gaps were rarely observed, Fig. (2). Attentive assessment of the distribution of gaps revealed that the distal regions of the long chromosomes were more vulnerable to the herbicide. Cells containing more than one aberrations were less frequent in occurrence, Fig. (3).
A slight increase in the mitotic index values in atrazine treated animals was noted at the lowest tested dose, then this value was found to be decreased gradually with increase the dose of the herbicide, though it still remained higher compared to untreated animals.
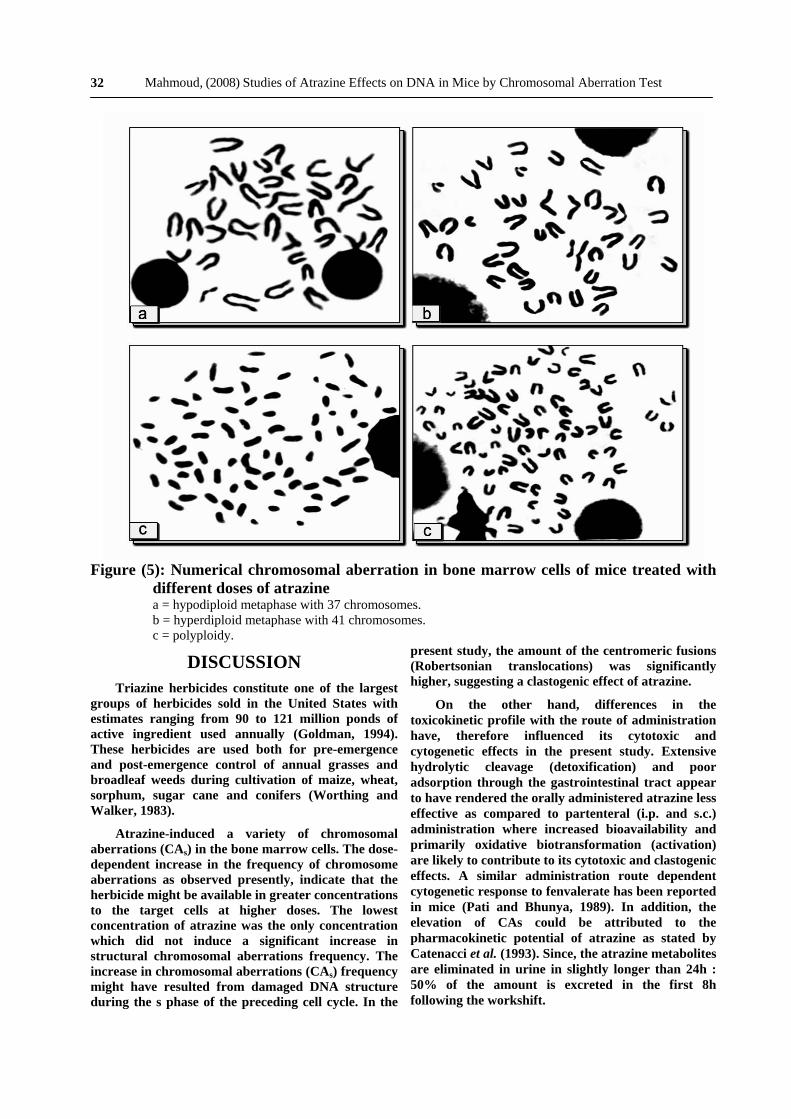
The frequency of hypoploidy also increased with increasing concentrations of atrazine but statistically significant differences from the controls were observed only at the dose of 225 mg/kg body weight. On the other hand, there was a clear dose dependent increase in the induction of polyploidy, Fig. (4). The frequency of polyploid metaphases in the bone marrow cells of mice was significantly increased at all dose levels compared with controls. In polyploid metaphases, the
chromosomes presented as a minute and highly shrunken units which were not easy to identify and even impossible to karyotype, Fig. (5). This type of aberration showed the highest score among other types.
DNA concentration: The amount of DNA in the control group was
38.5µg/ml and 30.8µg/ml in liver and kidney tissues, respectively (Table II). After 24hr of treatment with corn oil, the DNA amount in the both tissues were reduced to 36.8µg/ml and 24µg/ml, respectively. The intensity of DNA in mouse liver increased at low concentration of atrazine, while the high concentration of the herbicide reduced such intensity. The daily administration of atrazine for 5 consecutive days at 1/16 LD50, induced marked decrease of DNA concentration in the liver tissue to 30.9µg/ml; Furthermore, the DNA. amount in kidney tissue treated with atrazine decreased gradually till it reached 26.2µg/ml in the high dose. While, DNA level in this tissue was 28.4µg/ml and 32.3µg/ml due to treatment with 112.5 mg/kg body weight (1/16 LD50) for 5 and 10 consecutive days, respectively.
0
2
4
6
8
10
12
14
16
18
20
Control Corn oil 28.125 56.25 112.5 225
Dose (mg/kg b.w.)
Fig. (I): Percentage of structural chromosomal aberrations in bone marrow cells induced by atrazine via the oral gavage route.
Table (II): The concentration of DNA in both liver and kidney tissues of mice treated with atrazine.
Treatment
tissue Control Corn oil
28.125 mg/kg b.w.
56.25 mg/kg b.w.
112.5 mg/kg b.w.
225 mg/kg b.w.
112.5 mg/kg b.w. for 5 days
112.5 mg/kg b.w.for 10 days
Liver 38.5 36.8 46.5 37.2 41.7 29.3 30.9 34.2
Kidney 30.8 24 30.9 26.9 26.6 26.2 28.4 32.3
Stru
ctur
al ch
rom
osom
al ab
erra
tions
%

Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
30
Figure (2): Mitotic chromosomes from bone marrow cells of mice treated with atrazine for 24h. Rt = Robertsonian translocation ac f = acentric fragment b= break R = ring s c= secondary constractions d= deletion
Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
31
Figure (3): Metaphase spread with multiple aberrations. Rt = Robertsonian translocations iso g = isochromatid gap e to e assoc = end to end associations d = deletion g = gap
0
5
10
15
20
25
30
Control Corn oil 28.125 56.25 112.5 225
Dose (mg/kg b.w.)
Aneuploid cells Polyploid cells
Figure (4): Frequency of aneuploid and polyploid cells 24h after treatment with atrazine.
Aneu
ploi
d an
d po
lyplo
id ce
lls%
Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
32
Figure (5): Numerical chromosomal aberration in bone marrow cells of mice treated with different doses of atrazine a = hypodiploid metaphase with 37 chromosomes. b = hyperdiploid metaphase with 41 chromosomes. c = polyploidy.
DISCUSSION Triazine herbicides constitute one of the largest
groups of herbicides sold in the United States with estimates ranging from 90 to 121 million ponds of active ingredient used annually (Goldman, 1994). These herbicides are used both for pre-emergence and post-emergence control of annual grasses and broadleaf weeds during cultivation of maize, wheat, sorphum, sugar cane and conifers (Worthing and Walker, 1983).
Atrazine-induced a variety of chromosomal aberrations (CAs) in the bone marrow cells. The dose-dependent increase in the frequency of chromosome aberrations as observed presently, indicate that the herbicide might be available in greater concentrations to the target cells at higher doses. The lowest concentration of atrazine was the only concentration which did not induce a significant increase in structural chromosomal aberrations frequency. The increase in chromosomal aberrations (CAs) frequency might have resulted from damaged DNA structure during the s phase of the preceding cell cycle. In the
present study, the amount of the centromeric fusions (Robertsonian translocations) was significantly higher, suggesting a clastogenic effect of atrazine.
On the other hand, differences in the toxicokinetic profile with the route of administration have, therefore influenced its cytotoxic and cytogenetic effects in the present study. Extensive hydrolytic cleavage (detoxification) and poor adsorption through the gastrointestinal tract appear to have rendered the orally administered atrazine less effective as compared to partenteral (i.p. and s.c.) administration where increased bioavailability and primarily oxidative biotransformation (activation) are likely to contribute to its cytotoxic and clastogenic effects. A similar administration route dependent cytogenetic response to fenvalerate has been reported in mice (Pati and Bhunya, 1989). In addition, the elevation of CAs could be attributed to the pharmacokinetic potential of atrazine as stated by Catenacci et al. (1993). Since, the atrazine metabolites are eliminated in urine in slightly longer than 24h : 50% of the amount is excreted in the first 8h following the workshift.

Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
33
There is clear evidence that atrazine produces similar effects on plant and mammalian cells. It induces sister chromatid exchanges (SCEs) in root meristem cells of Allium cepa during the s phase, suggesting that the plant cell system shows similarities and may replace mammalian cells in screening and testing mutagens in vitro. Therefore, long treatment with maleic hydrazide(MH) will certainly cover the s period, while pulse treatment with MH will presumably damage few cells which are not in the s phase (Gonzalez-Gil and Navarrete, 1986).
Moreover, MH increased dominant lethals, X linked recessive lethals and X or y loss in Drosophila melanogaster (Murnik and Nash, 1977).
As it has been shown by Natarajan et al. (1981) that false-positive results in the sister chromatid exchange (SCE) assay may be caused by inhibitors of poly (ADP-ribose) - polymerase.
The possibility that the mutagenic activity of MH was due to the presence of impurities such as hydrazine or other compounds was considered. Therefore, Chinese hamster ovary (CHO) cells were treated with three different samples of MH of varying grades of purity. The magnitude of SCEs induction was similar for the three different samples, suggesting that the effect was not due to the presence of impurities. In addition, maleic hydrazide does not seem to induce SCEs in bone marrow of mice treated in vivo (Meschini et al., 1988). These data obtained in vivo can not be considered conclusive, as it is well known that cells other than bone marrow may be sensitive cell population.
Similarly, kligerman et al. (1993) found a concentration related increase in SCE in the splenocytes of rat and mouse subjected to the mixture of pesticides and ammonium nitrate that stimulated groundwater in California.
In 2004, Malik et al. suggested that an important part in the variation of SCE frequency reported by different laboratories when conventional SCE analysis is applied after exposure to a certain concentration of atrazine, is due to differences in cell cycle kinetics of cultured lymphocytes, rather than to a true biological variation in the cytogenetic end point used.
Additionally, in chromosome studies, MH itself induced chromosomal aberrations in Chinese hamster V79 cells when directly added to the medium, as previously demonstrated by other workers (Nishi et al., 1979). Similar studies reported by Taets et al. (1998) and Rayburn et al. (2001) using CHO cells.
In a series of in vivo and in vitro studies, Meisner et al. (1992) report problematic findings, concerning the clastogenicity of atrazine. This study's methodology is open to question because bone
marrow, a rapid cycling tissue, should not be used for analyses following chronic treatments as damaged cells are lost during subsequent cell divisions. Nevertheless, the authors report no increase in CAs in mice subjected to atrazine through their drinking water. In the in vitro part of this study, the authors treated human lymphocytes with from 0.01 to 1.0 µg/ml atrazine and reported a small increase in CAs at 0.1 and 100 µg/ml.
Other investigators reported that maleic hydrazide caused various chromosomal aberrations in cultured human cells (Ghiazza et al., 1984; Meisner et al., 1992 and 1993; Roloff et al., 1992; Lioi et al., 1998; Garaj-Vrhovac and Zeljezic, 2001 and Zeljezic and Garaj-Vrhovac, 2002).
However, several authors indicated that the herbicide atrazine was not effective using sister-chromatid exchange (SCE), chromosome aberrations and/or micronuclei (MN) in cultured human peripheral blood lymphocytes, irrespective of the presence of metabolic activation, which would mean a general lack of effectiveness of atrazine to induce clastogenic and aneugenic damage in cultured human lymphocytes (Ribas et al., 1998; Kligerman et al., 2000 and Malik et al., 2004). The lower sensitivity of MH on mammals and mammalian cells, as well as bacteria and fungi, may depend on the higher capacity of detoxication or lower levels to break down MH into products acting on cells cytogenetically, rather than that MH can not penetrate and reach the target nuclei better in mammalian cells than in plant cells (Nishi et al., 1979).
Recently, Bouilly et al. (2006) described aneuploidy in the pacific oyster, Crassostrea gigas, contaminated by atrazine using the G-banding technique and the restriction enzyme HaeIII.
On the other hand, there was a non significant difference (P>0.05) in mitotic index of treated mice when compared to control group. The present results are in agreement with the findings of Ugulava and Khubutiya (1975) who reported that the herbicides Dacthal (Chlorthal-dimethyl), diphenamid, atrazine, monuron and simazine caused insignificant changes in mitotic activity and chromosome changes were non-specific and not different from spontaneous mutations, nuclear proteins and DNA in onion root cells were affected hardly at all.
In 1992, Meisner et al. demonstrated dose related cytogenetic damage not associated with mitotic inhibition or cell death, with damage due to the alachlor-atrazine combination suggesting an additive model in human lymphocytes. Similary, Zeljezic and Garaj-Vrhovac (2002) mentioned that there were no differences in the proliferative rate index (PRI) between the control and treated group regardless of the sampling period in workers occupationally

Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
34
exposed to a mixture of pesticides (atrazine, alachlor, cyanazine, 2,4-dichlorophenoxyacetic acid and malathion).
The increase in polyploid cells in treated animals was significant at all dose levels but the increase in hypoploidy was found to be significant only at the higher dose. Polyploidy observed after treatment with atrazine caused an effect on the mitotic spindle. The relative damage of the spindle and the chromosomes may be distributed to the extent that the spindle is aborted at some extent or is absent, which results in the duplication of chromosome complement or polyploidy. This type of polyploidy is known as endopolyploidy and may be caused by different processes (Geitter, 1953 and Temtamy et al., 1982).
Endomitotic reduplication is a specialized process of doubling the chromosome material with the tendency of cells to pass from G2 to the next s phase without undergoing cell division. Microtubular / spindle disturbances are considered important to this process with implications of possible aneuploidy (Dean and Danford, 1984). Moreover, the induction of aneuploidy and polyploidy might be determined by quite different mechanisms, the former occurred at lower doses, involving missegregation of one or a few chromosomes, the latter occurred at higher doses, affecting the whole mitotic apparatus (Athwal and Sandhu, 1985).
On the other hand, induction of endomitotic reduplication has also been observed with a number of chemicals that influence DNA replication (Bean et al., 1992 and Gurr et al., 1993). A variety of chemicals including spindle disturbances also produce endomitotic reduplication and/or aneuploidy (Warr et al., 1993).
From the obtained results, it is clear that the decrease in the DNA content was depending on the tissue examined. The DNA amount was gradually decreased in the kidney tissue following 28.125, 56.25, 112.5 and 225 mg/kg of atrazine exposure for one day. However, no dose response could be found for DNA amount in the liver tissue. Moreover, the results showed that, there was a slight decrease in the concentration of DNA in kidney cells after oral administration of atrazine compared to liver cells of mice. The present results suggest that atrazine may be associated with genetic toxicity and that liver DNA was the target for atrazine.
In 1985, Gluth et al. mentioned that the amount of radioactivity which derived from 14C – labeled pollutants was 3-4 in liver, kidney and intestine, but just 1 in blood, muscle and gills of carp exposed for 6,24 and 72hrs to high external concentrations of atrazine.
Neskovic et al. (1993) found that atrazine led to changes of varying intensity, depending on the
parameter tested, the organs and tissues examined, as well as the atrazine concentration in carp. Furthermore, Scutaru et al. (1998) showed that atrazine concentrations measured in the liver of rats were higher than those found in the kidney, but both can be ranked as low compared with the amount of the administered doses.
In addition, the effects of a multiple dose indicated that the DNA amount in mouse liver and kidney were restored to normal levels, presumably due to a rapid degradation and elimination of atrazine. The present results is in agreement with the data obtained by Gordana et al. (1982) who indicated that ceruloplasmin and acid phosphatase activities were returned to normal levels in liver, kidney and spleen of rats administered a second dose of ATR after the recovery from the first treatment.
As the concentration of atrazine increased in the present work, the chromosomal aberrations values gradually increased as well, suggesting a high frequency of lesions in the DNA molecule. Similarly, Timson (1968) expected, theoretically, from the chemical structure of maleic hydrazide (MH) that it can interfere with DNA synthesis by substitution for the pyrimidines, and thus it would lead to faulty replication leading to a reduction in the number of successful mitoses in phytohaemagglutinin-stimulated human hymphocytes.
Some more recent studies have also been contradictory. Apparently, maleic hydrazide does not damage DNA per se, but only after transformation by the plant metabolism to some damaging derivatives (Plewa and Gentile, 1982). Pino et al. (1988) found atrazine to cause a very marginal increase in alkaline labile sites using the DNA alkaline elution assay on DNA from the stomach, kidney, and liver of rats treated orally with atrazine.
Studies by Taets et al. (1998) made use of flow cytometry attempt to measure clastogenicity induced by three triazines alone and in various combinations. These authors report that atrazine can induce clastogenicity as measured by changes in the coefficient of various (CV) of the DNA content of the G1 peak at concentrations down to 0.003 µg/ml.
The absence of DNA migration induced by maleic hydrazide was not affected in tobacco by either pH of the MH solution, the sampling time after MH treatment or continous MH treatment for 14 days (Gichner et al., 2000). Futhermore, using Comet assay, Tennant et al. (2001) showed that atrazine in concentrations of 250 and 500 mg/kg body weight significantly increased DNA migration in leukocytes of mice treated in vivo.
Marc et al. (2004) stated that the extent of the inhibition of DNA synthesis by formulated glyphosate was correlated with the effect on the cell cycle of sea

Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
35
urchin development. They concluded that formulated glyphosate's effect on the cell cycle is exerted at the level of the DNA response checkpoint of s phase. In 2005, Chang et al. showed dose-related increases in DNA damage in blood cells of common caip (Cyprinus carpio) treated with alachlor or atrazine at environmentally relevant concentrations (<100 ppb) using the single cell gel electrophoresis method. Atrazine at concentrations as low as 7 ppb induced liver cytochrome p450 IAI gene expression.
In 2006, Liu et al. found that atrazine was able to cause apoptosis in grass carp (Ctenopharyngodon idellus) cells from cell line ZC7901. Moreover, DNA fragmentation was detected by the TUNEL reaction and agarose gel electrophoresis.
Unlike atrazine, the commercial formulation Gesaprim and the adjuvant mixture increased DNA damage and the number of apoptotic cells in human lymphocytes. Metabolic activation additionally enhanced the DNA damaging potential of Gesaprim and the adjuvant mixture but did not affect atrazine genotoxicity (Zeljezic et al., 2006).
In conclusion, by using the gel agarose DNA electrophoresis on boh liver and kidney, genotoxic effects of atrazine in vivo can be detected and more knowledge on the specificity of its mutagenic action for both organs could be gained. Also, used on different organs in in vivo genotoxicity studies, the gel DNA electrophoresis assay could provide a good assessment of potential pesticide carcinogenicity.
Finally, the results of the present study indicate that atrazine have definite interactions with DNA metabolism in mice, resulting in chromosomal aberrations, indicating potential mutagenic effects. Human exposure to this agent should be restricted.
REFERENCES Agarwal, D.K.; Chauhan L.K.S.; Gupta S.K. and
Sundararaman V. (1994): Cytogenetic effects of deltamethrin on rat bone marrow. Mut. Res.; 311: 133-138.
Alvarez-Moya, C., Santerre Lucas, A.; Zuniga Gonzalez, G., Torres Bugarin, O., Padilla Camberos, E. and Feria Velasco, A. (2001): Evaluation of genotoxic activity of maleic hydrazide, ethylmethane sulfonate, and N-nitrosodiethylamine in Tradescantia. Salud Publica Mex.; 43 (6): 563-569.
Aquadro, C., Noon, W. and Began, D. (1992): RELP Analysis Using Heterogous Probs. In Hholzel, A.R., ed. Molecular Genetic Analysis of Populations, a Practical Approach. P. 115-157, IRL Press, Oxford.
Athwal, R.S. and Sandhu, S.S. (1985): Use of a human x mouse hybrid cell line to detect aneuploidy induced by environmental chemicals. Mut. Res.; 149: 73-81.
Bean, C., Armstrong, M.J. and Galloway, S.M. (1992): Effect of sampling time on chromosomal aberration yield for 7 chemicals in Chinese hamster ovary cells. Mut. Res.; 265: 31-44.
Bhalli, J.A., Khan, O.M. and Nasim, A. (2006): DNA damage in Pakistani pesticide manufacturing workers assayed using the Comet assay. Environ. Mol. Mutagen.; 47 (8): 587-593.
Bouilly, K., Leitao, A.; Chaves, R.; Guedes-Pinto, H.; Boudry, P. and Lapegue, S. (2006): Endonuclease banding reveals that atrazine-induced aneuploidy resembles spontaneous chromosome loss in Crassostrea gigas. Cell Biochem. Biophys.; 45 (2): 203-213.
Catenacci, G., Barbieri, F., Bersani, M.; Ferioli, A.; Cottica, D. and Maroni, M. (1993): Biological monitoring of human exposure to atrazine. Toxicol. Lett.; 69 (2): 217-222.
Chang, L.W., Toth, G.P., Gordon, D.A., Graham D.W., Meier, J.R., Knapp, C.W., Jr. deNovelles, F.J., Campbell, S. and Lattier, D.L. (2005): Responses of molecular indicators of exposure in mesocosms: common carp (Cyprinus carpio) exposed to the herbicides alachlor and atrazine. Environ. Toxicol. Chem.; 24 (1): 190-197.
Clements, C., Ralph, S. and Petras, M. (1997): Genotoxicity of select herbicides in Rana catesbeiana tadpoles using the alkaline single-cell gel DNA electrophoresis (Comet) assay. Environ. Mol. Mutagen.; 29: 277-288.
Dean, B.J. and Danford, N. (1984): Assays for the detection of chemically induced chromosomal damage in cultured mammalian cells, in : S.Venitt and Parry J.M. (Eds.), Mutagenicity Testing, A Practical Approach, IRL Press, Oxford, pp. 187-232.
Dooley, G.P., Prenni, J.E., Prentiss, P.L., Cranmer, B.K., Andersen, M.E. and Tessari, J.D. (2006): Identification of a novel hemoglobin adduct in Sprague Dawley rats exposed to atrazine. Cell Biochem. Biophys.; 45 (2): 203-213.
Garaj-Vrhovac, V. and Zeljezic, D. (2001): Cytogenetic monitoring of Croatian population occupationally exposed to a complex mixture of pesticides. Toxicol.; 165 (2-3): 153-162.
Garaj-Vrhovac, V. and Zeljezic, D. (2002): Assessment of genome damage in a population of

Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
36
Croatian workers employed in pesticide production by chromosomal aberration analysis, micronucleus assay and Comet assay. J. Appl. Toxicol.; 22 (4): 249-255.
Gebel, T., Kevekordes, S., Pav, K., Edenharder, R. and Dunkelberg H. (1997): In vivo genotoxicity of selected herbicides in the mouse bone-marrow micronucleus test. Arch. Toxicol.; 71: 193-197.
Geitter, L. (1953): Endomitose und endomitoti sche polyploidisierung. Protoplasmatologia; 60: 1-89.
Ghiazza, G., Zavarise, G., Lanero, M. and Ferraro, G. (1984): Sister chromatid exchanges induced in human lymphocyte chromosomes by trifluralin, atrazine and simazine. Boll Soc. Ital Biol. Sper.; 60 (11): 2149-2153.
Gichner, T., Menke, M.; Stavreva, D.A. and Schubert, I. (2000): Maleic hydrazide induces genotoxic effects but no DNA damage detectable by the Comet assay in tobacco and field beans. Mutagenesis; 15 (5): 385-389.
Gluth, G.; Freitag, D.; Hanke, W. and F. Korte, (1985): Accumulation of pollutants in fish. Comp. Biochem. Physiol.; 81 (2): 273-277.
Goldman, L.R. (1994): Atrazine, simazine and cyanazine: notice of initiation of Special Review. Federal Register; 60412-60443.
Gonzalez-Gil, G. and Navarrete, M.H. (1986): Induction of SCE by maleic hydrazide throughout the cell cycle. Mut. Res.; 163: 57-61.
Gordana, W., Straus, B. and Stankovic, V. (1982): Effects of atrazine on ceruloplasmin and acid phosphatase in rat liver, kidney and spleen. Acta. Pharm. Jugosl; 32 (1): 53-58.
Gross-Bellard, M.; Oudet, P. and Chambon, P. (1972): Isolation of high molecular weight DNA from mammalian cell. Euro. J. Biochem.; 36: 32-37.
Gurr, J.R., Lin, Y.C., Ho, I.C., Jan, K.Y. and Lee, T.C. (1993): Induction of chromatid breaks and tetraploidy in Chinese hamster ovary cells by treatment with sodium arsenite during G2 phase. Mut. Res.; 319: 135-142.
Kligerman, A.D., Chapin, R.E., Erexson, G.L., Germolec, D.R., Kwanyuen, P. and Yang, R.S.H. (1993): Analyses of cytogenetic damage in rodents following exposure to simulated groundwater contaminated with pesticides and a fertilizer. Mut. Res.; 300: 125-134.
Kligerman, A.D., Doerr, C.L., Tennant, A.H. and Zucker R.M. (2000): Cytogenetic studies of three
triazine herbicides. I. In vitro studies. Mut. Res.; 465: 53-59.
Lioi, M.B., Scarfi, M.R., Santoro, A., Barbieri, R., Zeni, O., Salvemini, F., Di Berardino, D. and Ursini, M.V. (1998): Cytogenetic damage and induction of pro-oxidant state in human lymphocytes exposed in vitro to glyphosate, vinclozolin, atrazine, and DPX-E9636. Environ. Mol. Mutagen.; 32 (1): 39-46.
Liu, X.M., Shao, J.Z., Xiang, L.X. and Chen, X.Y. (2006): Cytotoxic effects and apoptosis induction of atrazine in a grass carp (Ctenopharyngodon idellus) cell line. Environ. Toxicol.; 21 (1): 80-89.
Malik, S.I., Terzoudi, G.I. and Pantelias, G.E. (2004): SCE analysis in G2 lymphocyte prematurely condensed chromosomes after exposure to atrazine: the non-dose-dependent increase in homologous recombinational events does not support its genotoxic mode of action. Cytogenet. Genome Res.; 104 (1-4): 315-319.
Marc, J., Belle, R.; Morales, J.; Cormier, P. and Mulner-Lorillon, O. (2004): Formulated glyphosate activates the DNA- response checkpoint of the cell cycle leading to the prevention of G2M transition. Toxicol. Sci.; 82 (2): 436-442.
Meisner, L.F., Belluck, D.A. and Roloff, B.D. (1992): Cytogenetic effects of alachlor and/or atrazine in vivo and in vitro. Environ. Mol. Mutagen.; 19 (1): 77-82.
Meisner, L.F.; Roloff, B.D. and Belluck, D.A. (1993): In vitro effects of N-nitrosoatrazine on chromosome breakage. Arch. Environ. Contam. toxicol.; 24 (1): 108-112.
Meschini, R., Quaranta, M.T., Fiore, M., Polcaro, C., Possagno, E. and Palitti, F. (1988): Chromosomal damage induced by maleic hydrazide in mammalian cells in vitro and in vivo. Mut. Res.; 204(4): 645-648.
Monroy, C.M., Cortes, A.C., Sicard, D.M. and de Rostrepo, H.G. (2005): Cytotoxicity and genotoxicity of human cells exposed in vitro to glyphosate. Biomedica; 25 (3): 335-345.
Murnik, M.R. and Nash, C.L. (1977): Mutagenicity of the triazine herbicides atrazine, cyanazine, and simazine in Drosophila melanogaster. J. Toxicol. Environ. Health; 3 (4): 691-697.
Natarajan, A.T.; Csukas, I. and Van Zeeland, A.A. (1981): Contribution of incorporated 5- bromodeoxy uridine in DNA to the frequencies of sister chromatid exchanges induced by inhibitors

Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
37
of poly (ADP-ribose) polymerase. Mut. Res.; 84:125-132.
Neskovic, N.K., Elezovic, I., Karan, V., Poleksic, V. and Budimir, M. (1993): Acute and subacute toxicity of atrazine to carp (Cyprinus carpio L.). Ecotoxicol. Environ. Saf.; 25(2): 173-182.
Nicolau, G.Y. and Socoliuc, E. (1980): Effects of atrazine on circadian RNA, DNA and total protein rhythms in the thyroid and adrenal. Endocrinologie; 18 (3): 161-166.
Nishi, Y., Mori M. and Inui, N. (1979): Chromosomal aberrations induced by maleic hydrazide and related compounds in Chinese hamster cells in vitro. Mut. Res.; 67: 249-257.
Pati, P.C. and Bhunya, S.P. (1989): Cytogenetic effects of fenvalerate in mammalian in vivo test system. Mut. Res.; 222:149-154.
Pino, A. Maura, A. and Grillo, P. (1988): DNA damage in stomach, kidney, liver and lung of rats treated with atrazine. Mut. Res.; 209: (3-4): 145-147.
Plewa, M.J. and Gentile, J.M. (1982): The activation of chemicals into mutagens by green plants, in : de Serres F.J. and Hollaender A. (Eds.), Chemical Mutagens, Principles and Methods for their Detection, vol. 7, Plenum. New York pp. 401.
Rayburn, A.L., Bouma, J. and Northcott, C.A. (2001): Comparing the clastogenic potential of atrazine with caffeine using Chinese hamster ovary (CHO) cells. Toxicol. Lett.; 121 (1): 69-78.
Ribas, G., Surrallés, J., Carbonell, E., Creus, A., Xamena, N. and Marcos R. (1998): Lack of genotoxicity of the herbicide atrazine in cultured human lymphocytes. Mut. Res.; 416: 93-99.
Rios, A.C.C., Salvadori, D.M.F.; Oliveira, S.V. and Ribeiro L.R. (1995): The action of the herbicide paraquat on somatic and germ cells of mice. Mut. Res.; 328 (1): 113-118.
Roloff, B.D., Belluck, D.A. and Meisner, L.F. (1992): Cytogenetic studies of herbicides interactions in vitro and in vivo using atrazine and linuron. Arch. Environ. Contam. Toxicol.; 22 (3): 267-271.
Rusiecki, J.A., De Roos, A., Lee, W.J., Dosemeci, M., Lubin, J.H., Hoppin, J.A., Blair, A. and Alavanja, M.C. (2004): Cancer incidence among pesticide applicators exposed to atrazine in the agricultural health study. J. Natl. Cancer Inst.; 96 (18): 1375-1382.
Sambrook, J., Fritsch, E. and Maniatis, T. (1989): Molecular Cloning: A Laboratory Manual Cold Spring Harbour Laboratory, Press, New York.
Scutaru, B., Giersch, T., Cozmei, C. and Hock, B. (1998): Immunoenzymatic determination of atrazine in rat tissue samples. Toxicol.; 127 (1-3): 11-16.
Snedecor, G.M. and Cochran, W.G. (1980): "Statistical methods "7th ed. Oxford and J.B.H. publishing com.
Taets, C., Aref, S. and Rayburn, A.L. (1998): The clastogenic potential of triazine herbicide combinations found in potable water supplies. Environ. Health Perspect.; 106 (4): 197-201.
Temtamy, S.A., de Hondt, H.A. and El–Ghor, M.A. (1982): Effect of novalgin on chromosomes of Rattus norvegicus. Egypt. J. Genet. Cytol.; 11: 105-111.
Tennant, A.H., Peng, B. and Kligerman, A.D. (2001): Genotoxicity studies of three triazine herbicides : in vivo studies using the alkaline single cell gel (SCG) assay. Mut. Res.; 493 (1-2): 1-10.
Timson, J. (1968): The effect of maleic hydrazide on the mitosis of phytohaemagglutinin stimulated human lymphocytes cultured in vitro, Caryologia; 21: 157-159.
Ugulava, N.A. and Khubutiya, R.A. (1975): Chromosome aberrations and weight changes in cell nuclei under the influence of herbicides. Materialy logo Mezhdunarodnogo Simpoziuma Stran Chlenov SEV, Pushchino, SSSR; 1: 57-59.
Undeger, U. and Basaran, N. (2002): Assessment of DNA damage in workers occupationally exposed to pesticide mixtures by the alkaline Comet assay. Arch. Toxicol.; 76 (7): 430-436.
Warr, T.J., Parry, E.M. and Parry, J.M. (1993): A comparison of two in vitro mammalian cell cytogenetic assays for the detection of mitotic aneuploidy using 10 known or suspected aneugens. Mut. Res.; 287: 29-46.
Worthing, C.R. and Walker, S.B. (Eds.) (1983): The pesticide Manual, 7th ed. The Lavenham Press, Lavenham.
Zeljezic, D. and Garaj–Vrhovac, V. (2001): Chromosomal aberration and single cell gel electrophoresis (Comet) assay in the longitudinal risk assessment of occupational exposure to pesticides. Mutagenesis; 16 (4): 359-363.

Mahmoud, (2008) Studies of Atrazine Effects on DNA in Mice by Chromosomal Aberration Test
38
Zeljezic, D. and Garaj–Vrhovac, V. (2002): Sister chromatid exchange and proliferative rate index in the longitudinal risk assessment of occupational exposure to pesticides. Chemosphere; 46(2): 295-303.
Zeljezic, D. and Garaj–Vrhovac, V. (2004): Genotoxicity evaluation of pesticide formulations containing alachlor and atrazine in multiple
mouse tissues (blood, kidney, liver, bone marrow, spleen) by Comet assay. Neoplasma; 51 (3): 198-203.
Zeljezic, D., Garaj–Vrhovac, V. and Perkovic, P. (2006): Evaluation of DNA damage induced by atrazine and atrazine- based herbicide in human lymphocytes in vitro using a Comet and DNA diffusion assay. Toxicol. In Vitro.; 20 (6): 923-93