stbroid metabolism in tbb pu-incubation...
TRANSCRIPT

STBROID METABOLISM IN TBB
PU-INCUBATION MEDIUM
by
Charles Pak Wai Tsang

MBTABOLISM OF STEROIDS IN THE PRE-INCUBATION MEDIUM FROM ADR.BNAL GLANDS
by Charles Pak Wai Tsang
A thesis submitted to the Faculty of Graduate
Studies and Research in partial fulfilment of
the requirements for the degree of
Master of Science
Department of Investigative Medicine
McGill University, Montreal. August, 1965

ACKNOWLBDGEMENTS
I wish to express my deepest appreciation to Professor
J. s. L. Browne for his interest, advice and encouragement in
this work, and for supplying the laboratory facilities in the
Department of Investigative Medicine.
I wish to express my profound gratitude to Dr. A. Carballeira
for his continued interest, criticism and guidance throughout
this study.
I would like to thank Dr. c. J. P. Giroud and Dr. J. Stachenko
for their valuable advice, suggestions, and gift of llp-oH-proges•
terone.
I am also grateful to Dr. W. Klyne from whom I obtained llp
OH-progesterone for crystallization etudies.
I aœ thankful for the very kind help which I have received
from my fellow students and staff members in this laboratory. In
particular I wish to thank Mrs. R. Lucis, Mr. A. Kehdi, and Mr.
D. MeKillan for technical assistance. I also wiah to thank Mr.
H. Triller, Mr. M. Leong, and Mr. A. Mehdi for reading this manus
eript.
My gratitude is due to my wife who has helped typing the
initial manuscript.
I would like to express my appreciation to Miss B. Wagmann
who bas skillfully typed this thesis.
Lastly, but not leest, I am much indebted to the late Mr.
v. Kazmin for his guidance in analytical procedures.

To My
SIS TER,
COUSINS,
and
MI WIP'E
for their
Love and Kindness

TABLE OF COBTBNTS
I. UVIIW OF LITERATUU
PART I - INTRODUctiON
1. Important Steroids iao1ated from the Mammalien Adrena1 Cortex
2. J'unctional Zonation
3. Lack of 17a-hydroxylated Steroids in the &at Adrenals
4. llp-OH-Progesterone as a potentia1ly important Intermediate Metabolite in Steroid Biosynthesis
PAI.T II
Some Reactions involved in the Biosynthesis of Adrenocortical Steroids
1. Isomerization of ~5-3p-ol to ~4-3-keto
2. Steroid Hyd.roxylation
i.
1
2
2
4
4
6
8
9
a) Location of Enzymes 9
b) Oxygen Requirement 10
c) MDPH &equirement 11
d) NADPH-generating Systems 12
e) Mechanism of Steroid Hydroxylation 13
f) Effects of Freezing on Tissue Response to NADPH 13
Origin of the present Study
II. MATUIALS AND MBTBODS
1. Materials
2. Incubation Procedure
3. Extraction of Steroids
14
15
15
16
18

ii.
4. Paper Chroœatography for the Separation of Steroids 19
S. Detection of Steroids on Chroœatograms 21
6. Blution of Steroids from the Paper Strips 22
7. Quantitative Determination of Steroids 22
8. Systemetic Separation of Steroids 23
9. Characterization of Corticosterone, Cortisol, and llp-oH-Progesterone 24
Acetylation 25
Oxidation 26
Crystallization of llp-oH-Progesterone 26
Test for a.ketol group 27
Test for the dihydroxy-acetone aide-chain 27
III. DSULTS 29
IXPIISSION OF IBSULTS 29
SBCTION A 31
1. Incubation with Labe11ed Progesterone
2. Co-factor aequirementa
3. Incubation with Labelled Pregnenolone
4. Incubation with Labelled Cholesterol
5. Conclusion drawn from Section A
SBCTION B
1. Bffect of Time on Pre-incubation
2. Bffect of Temperature on Pre-incubation
31
38
41
48
48
49
49
52

iii.
TA:BLB OF CONTBNTS (Cont' d.)
SlcriON C SS
Tissue versus PIM SS
SBCTION D 60
Characterization of the aajor Radioactive Conversion Products from Labelled Substrates by the Bovine or &at Adrenal PIM 60
(I) llP-OB-Progesterone (froa rat adrenal PI!O
(II) Cortisol and Corticosterone (froœ bovine adrenal PI!O
(III) Tentative Corticosterone isolated frou the rat adrenal PIM, and progesterone froa the bovine adrenal PIM
IV. GBNBIAL DISCUSSION
v. s UMM&.R.Y
60
64
65
68
73

e
Trivial or Abbreviated Naœe
18-0H-B
Cortisol (Compound F)
Aldosterone
Cortisone (Compound E)
19-0H•DOC
18-0B-DOC
11-deoxy-cortiaol (Compound S)
Corticosterone (Compound B)
llP·OH-progesterone
17a-OB-progeaterone
11-deoxycorticosterone (DOC)
11-keto-progesterone
Pregneno1one
Progesterone
Systematic N-.e
llp,l8,21-tribydroxy•pregn-4-ena-3,20-dione
llp,l7a,21•tribydroxy-pregn-4-ene-3,20-dione
llp,21-dibydroxy-pregn-4•ene-3,20-dione-18-al
17a,21-dibydroxy-pregn-4-ene-3,11,20-trione
19,21-dibydroxy-pregn-4-ene-3,20-dione
18,21-dibydroxy-pregn-4-ene-3,20·dione
17a,21-dibydroxy-pregn·4-ene-3,20-dione
11J,21-dibydroxy-pregn-4-ene-3,20-dione
llp-bydroxy-pregn-4-ene-3,20-dione
17a-bydroxy-pregn-4-ene-3,20-dione
21-bydroxy-pregn-4-ene-3,20-dione
Pregn-4-ene-3,11,20-trione
Jp-bydroxy-pregn-5-ene-20-one
Pregn-4-ene-3,20-dione
e
~ •

e e
Other Abbreviationa
A cm Adrenocorticotropic hormone
G·6-P Glucose 6-phosphate
NAD Nicotinamide adenine dinucleotide (oxidized form)
NADP Nicotinaœide adenine dinucleotide phosphate (oxidized form)
NAD PH Nicotinaaide adenine dinucleotide phosphate (reduced form)
PIM Pre-incubation medium or media
c.p.m. counta per minute
<Cl •

I. RBVIBW or Ll'J.UATUU
INTRODUCTION
The bioayntheais of adrenocortical ateroids has been intensively
studied in the last decade or so. Comprehensive reviewa have appeared
covering aultiple aspects of the proceasea involved in the elaboration
of the cortical aecretory products under varioua expertœental condi
tions (1, 2, 3, 3a). ln this theais, attention vas focuaed in review
ing those aspects of the !2 vitro ateroid biosynthesis more connected
to the present experimental work. ln the first part of the review, a
brief survey of the more important corticoids of the C-21 series pro
duced by the mammalian adrenal cortex will be given. The site of syn
thesia of these steroids within the cortex will also be discussed.
More emphasia will be made on the 0·21 steroids synthesized and se
creted by the beef and rat adrenals, becauae most of the present studies
have been done on the former species, with two exper~ents also carried
out on the latter. Within tais part of the review, a short paragraph
is also devoted to llP-OH·progesterone, which bas been of some interest
to us.
The second part of the review will deal mainly vith the enzymatic
reactions involved in the transformation, and certain hydroxylations
of the steroid molecule. ln connection vith this, the aasociated
enzyme systems, co-factor requireaents, and affect of temperature will
be discussed. A brief description of the origin of this atudy will be
given at the end of the review.

-2·
PAU I
1. Imeortant Steroids ieolated from the Mammalien Adrenel Cortex
Cortisol and corticosterone are quantitatively and qualita
tively the most important glucocorticoids synthesized and released
by the mammalien adrenal cortex. In .an (4, 5), the monkey (6, 7),
the guinee pig (6, 8), and the cattle (9), cortisol ia produced in
much higher concentration then corticosterone. In the rat (10, 11,
12, 13), the rabbit (14, 15), and the mouse (16, 17), corticosterone
is, however, the major secretory product. &ecently, Birmingham and
Ward (18), and Peron (19, 20) have, independently, identified 18-
hydroxy-11-deoxycorticoaterone (18-oB-DOC) as the second most abun
dant steroid of the rat adrenal. Peron (19, 20) bas also isolated
18-hydroxycorticosterone (18~-B) in the seme species. Sandor and
Lanthier (21) bave reported the isolation of 18-0B-B from the zona
g1omeru1oaa of the beef adrenal.
Aldosterone is the chief mineralcorticoid of probebly all the
mammalien apecies. Bowever, it is aynthesized in very amall con·
centrations in the adrenel cortex. This hormone waa first iaolated
by Simpson and Tait in 1952 (22).
2. Functionel Zonation
Tbus far we have been diacussing the isolation of a few important
corticoids from the adrenal cortex without particularly pointing out
(with the exception of 18-oB-B) the sites of syntbesis of these compounds
in the cortex. In general, in the few species investigated, cortisol

-3-
is produced pri .. rily in the inner zones or zona fasciculata-reticu
larie (Z.F.), aldosterone in the outer zone or zona glomeruloaa (Z.G.),
and corticosterone in all zones. This vas first demonstrated by Ayres
et al. (25) who incubated the separated Z.G. and Z.F. of the bovine
adrenal cortex. St.ultaneously, Giroud et al. (26) showed that aldo
sterone waa produced mainly in the Z.G. of the rat adrenals. These
findings have been well confirmed (27-31), including evidence obtained
fraa the huœan adrenal cortex (32).
Bovever, the original concept of functional zonation, i.e. dif
ferent zones may produce different steroids with different biological
actions, must be credited to Dean and Greep (23, 24) who obaerved that
following hypophysectomy, the inner zones of the adrenal cortex atrophied
with an accompanying disorder in carbohydrate metabolism, whereas in
sodium deficient rats hypertrophy of the Z.G. occured.
We have thus far reviewed briefly some of the important steroids
of interest produced in the different zones of the adrenal cortex.
Now a summary of the principal C-21 steroids isolated from the dif
ferent zones in the beef or rat adrenal cortex is tabulated below:
!&· !:!.· Aldosterone
Corticosterone Cor ti cos ter one
18-oB-B
18-oB-DOC
BOVINE
Z,G. hl•
Aldosterone
Corticosterone Corticosterone
18-oH-B
Cortisol

The adrenal cortex also synthesizes and secretes the C-19 steroids.
However, these compound& are not the concern of this thesis.
3. Lack of 1~-hydrolylated Steroids in the Rat Adrenals
Since there have been some conflicting resulta as to whether the
17a-bydroxylated steroids are synthesized in the rat adrenal or not,
it seeme appropriate to insert a few lines to discuss this point.
In the earlier works of Beard et al. (35), Eisenstein (36), Bof
mann (37), and recently Brownell et al. (38), these authors bave re
ported to bave detected 17a-hydroxylated compound& in the rat adrenals.
The criteria for identification are invariably based on a positive
Porter-Silber reaction, and similar mobility with the authentic com
pound& in some chromatographie systems. However, Birmingham et al.
(39) and Ward and Birmingham (40), and lately Laplante (41) have pre
sented no evidence for the presence of 1~-hydroxylated steroid in
this species. Further, Birmingham and Ward (18), and Ward and Birming
ham (42), and independently Peron (19) have isolated 2o-18 hemiketal
of 18-0H-DOC from the rat adrenal. This compound reacts with Porter
Silber reagent, but does not reduce tetrazoliuœ chloride (42).
It is now generally accepted that the rat adrenal does not
produce 17a-hydroxylated steroids.
4. 118-oH-P as a potentia11y important Intermediate Metabolite in
Steroid Biosznthesis
We bave been particu1arly interested in this steroid compound not
only because of its association with our work, as has been explained

•
•
-5-
earlier, but also because there has been increasing evidence to suggest
that this may be an important interœeàiate metabolite in steroid bio
synthesis.
The earlier work of Levy et al. (43) showed that the beef adrenal
did not œetabolize 11~-0H-progesterone, even though small amounts of
this steroid were obtained by perfusing the gland with progesterone.
Eichhorn and Hechter (44) found that homogenates of beef adrenals
hydroxylated llp-oH-progesterone at a slower rate than progesterone
or DOC. On the basis of these findings, the above mentioned authors
concluded that 11~-oB-progesterone was not an important intermediate
metabolite in the beef adrenals. On the other band, Brownie et al.
(45) demonstrated that beef aitochondria readily converted proges
terone to ll~·OH-progesterone. Giroud and Stachenko et al. (46) showed
that beef gloaerulosa slices metabolized ll~·OH-progesterone as easily
as corticosterone or DOC to aldosterone. More recently, Stachenko and
Giroud (31) showed that 11,-oB-progesterone was readily coaverteè to
cortisol by the beef fasciculata sliees. (Ro comparison was made witb
17a-oH-progesterone). Roberts et al. (47) indicated that rat adrensl
bomogenate bydroxylated significant amounts of labelled progesterone
to 11~-0H-progesterone in the presence of cyclic 3',5'-adenosine mono
phosphate (eyclic AMP).
Kraulis and Birmingham (48) reported that the rat and the frog ad
renal quartera ~ vitro converted stable llp-oH-progesterone to corti
costerone more efficiently than added deoxycorticosterone. Oertel and
Eik-Res have isolated and identified llp-oR-progesterone, and 11-keto
progesterone in the canine adrenal vein blood (49). Sandor et al.(50) have

-6-
tentatively identified labelled llp-OB·progeaterone froœ the duck
and pullet adrenal slices incubated with labelled pregnenolone or
progesterone. Very recently, Kittinger (51) with the use of gas
chromatography has been able to identify 11~-0B•progesterone froœ
the rat adrenal tissue. Thus, there is accumulating evidence that
11~-0B-progesterone may be an important interaediate metabolite in
steroid bioayntheais of the adrenal cortex.
P.A.ItT II
Some Reactions involved in the Bioaratheaia of Adrenocortical Steroids
The biosynthesia of adrenocortical steroids starts with acetate.
The sequences of reactions involved are not fully known. Those wbich
are most generally accepted today are based on the work of Bechter
et al. (52) with the perfused ox adrenal gland. The work of others
bas resulted in modification of the original scheme, and additions
to it, but without any drastic alteration. Theae changes were re
viewed by Grant (2). There is reason to believe that the asme or
very similar sequences are followed. in man (53, 54, 55).
A schema of the more important stepa in the synthesis is given
on page 7.
Bovever, in this review, our interest is in the enzyme systems
involved in the later part of the syntheaia, atamting from pregnen
olone, i.e. the isomerization and hydroxylation of the steroid mole
cule.

-7-
A Scheae of the Bios79thesis of Adreaocortical Steroids
Acetate---~ Mevalonic acid--·r Squalene '
A ~ 2oa-hydroxycholesterol ~---·Cholesterol
1 B ~ 'il 'lj
2oa,22B·dihydroxycholesterol Cholesterol esters ! ,c y
Pregne•olone
v
1)
B
Progesterone·-·t Corticoids
A: "2oa-hydroxylase11 (56)
1: 11 22-hydroxylase11 (57)
C: 1120,22-desaolase" (58)
D: A5-3p-bydroxysteroid. debydrogenase (59)
B: Isomerase-·transfers double bond froa carbone S and. 6 to carbone 4 and. S (60)
F: llfS, 17a, 21, ••• etc. steroid hydroxylases (Reference given in text).

.a.
1. Isomerization of~S.Jp-ol to~4-3-keto
The coaversiou of pregneaoloae to progesterone consista of at
least two operatioaa as is indicated in the scheme of page 7. These
reactions are: dehydrogenation of the 3p-hydroxyl group, and shifting
of the double bond froœ carbone 5 and 6 to carbone 4 and 5. At least
two enzymes or enzyme systems are involved, the ~5-3p-bydroxysteroid
dehydrogenase (59) and isomerase (60). In practice, the two enzymes
or enzyme systems are difficult to separate and are studied together
under the name of the former, wbich we shall be discussing.
The ~5-Jp-bydroxysteroid dehydrogenase is now known to be present
in all steroid producing tissues, for exemple, the corpus luteua and
the testis (59), and lately it bas also been demonatrated to be present
also in the liver and kidney (61). Its presence in the adrenal cortex
waa first demonstrated by Saœuels et al. (59), who at the seme t~
showed that RAD st~lated this reaction by acting as a hydrogen acceptor.
Tbese findings were confirmed by Byer and Samuels (62), and also Balkerston
et al. (63) froa aicrosoœal etudies of the beef adrenals. The latter
authors fouad that there was a losa of activity of this enzyme after
mild dialysis, and that the activity could be restored by the addition
of RAD. Lately, Kowal et al. {64, 65) from etudies of the acetone
powder preparations of the corpus luteu. and the adrenal cortex, found
that NAD was the preferred co-factor of this reaction and that NADP
could replace NAD with less tban 501 of the activity. NADH, however,
inhibits this reaction which can be overcome by adding excess NAD. This
reaction is also inhibited by the C-19-~4·3-ketoateroids and progesterone

-9-
according to the seme authora. Of particular interest ia their
observation that their preparations could not utilize cholesterol
or 20-deoxypregnenolone suggesting that an oxygen function at the
C-20 position is necessary for the activity of the dehydrogenase.
Kistochemical etudies by Baille et al. (66) seem to indicate
that the enzyme is present mainly in the Z.G. Bowever, Levy and
aubin (67) have evidence that the enzyme is present in all zones
of the rat adrenal cortex. ln the human adrenal cortex, the enzyme
is also found in all zones {68). Sub-cellularly, this enzyme is
found in the microsomal fraction {62).
2. Steroid Htdroxylation
Kydroxylation of the steroid molecule, at least at the 11, 17
and 21 positions, require the specifie hydroxylating enzymes (bydrox
ylases), molecular oxygen, and NADPB as co-factor. We sball make
a brief survey of the locations of the different enzymes, the oxygen
and co-factor requirements, generation of the co-factor NADPH, and
finally a brief mentioning of the mecbanism of bydroxylation which
is, at present, virtually speculative.
a) Location of Bnzyœes
llp-hxdro!Ylase: This enzyme is present in all zones of the
cortex (see page 102 of reference 1) and bas been most intensive1y
studied. lt is located in the mitochondrie (69, 70). A soluble
fraction of it bas been prepared by Grant (71), Tomkins et al. (72)
and Sharma et al. (73). Tomkins et al. (72, 74) claimed tbat they

-10-
have obtained, from the calf adrenal acetone powder extracts» tbree
distinct enzymes, and one beat-stable co-enzyme, all of wbicb were
required to catalyse llp-hydroxylation, besides o2 and NADPB. The
beat-stable co-factor was obtained fra. an aqueous extract of rabbie
liver acetone powder. Sweat (75) bas partially purlfied certain
fractions iDvolved in llp-hydroxylation by .aans of ammoniua-sulphate
precipitation. Tbere ia evidence for the exiatence of at least two
llp-hydroxylases: one ia specifie to progesterone and is present ln
Z.G.; the otber to DOC and 1a present throughout the cortex. (See
page 102 of reference 1).
21-hydroxylase:
The enzyme is found throughout the cortex, Plager and Saœuels
(76) designated this enzyme as the 'soluble fraction', on the ground
that after centrifuging the hoaogenate at 20,000 g for half an hour,
the enzyme reaained in the aupernatant. Later, Ryan and Bngel (77)
showed that the 20»000 g supernatant fraction was sedimented at
105,000 g for balf an hour, and deaignated the 105,000 g sediments
as the 'aicrosoaal fraction'.
l~hzdroxylase:
17a-hydroxylase is present pr~rily ln the zona fasciculata, and
sub-cellularly, in the aicrosoaal fraction (78). For this reason, it
is difficult to separate 17a-hydroxylase from 21-hydroxylase by centri
fugation alone, since both enzyaes are in the microsomal fraction.
b) O!Y&en Requirement
Hayano and Dorfman (70, 79) noted that o2 waa utillsed by the

-11-
beef homogenates. and Saffran and Bayliss (80) observed that the
rat adrenals failed to produce corticoids when o2 was replaced by
N2• Direct evidence that molecular oxygen is involved in llp-hy
droxylation was obtained from the work of Hayano et al. (81. 82, 83),
18 . 18 using o2, D2o and H2 o. This was later confirœed by Sweat et
al. (84). Involvement of molecular oxygen in 1~ and 21 hydroxyla-
tion was similarly elucidated by Hayano et al. {85) from etudies
on micro-organism. Ryan and Bngel (86) also obtained s~ilar find-
inga with 17a-bydroxylation from the beef adrenal microsomal studies.
c) NADPB &eguirement
Saffran and Bayliss (80) observed no corticoid production in
rat homogenates incubated alone. Hayano and Dorfman {87) noted
that the 11-·hydroxylation of DOC with washed adrenal homogenate
required the addition of fuaaarate and magnesium ions (Mg*). ATP
and NAD were also needed in the case of aged or acetone powder pre-
parations. NADP alone could replace all these additives (87. as.
89) • Wben NADP was used, the requirement for Hg++ vas no longer
present, indicating the transformation of NAD in the presence of
ATP and Ma++ to NADP.
However, it was Sweat and Lipcomb (90) who demonstrated that
NADPH was the actual co-factor required for the hydroxylation.
These authors also pointed out that the di- and tri-carboxylic acide
of the Krebs cycle, which stimulated the hydroxylation, did so by
acting as substrates for the generation of NADPB, for instance, iso-
citrate + NADP' )oxalosuccinate + NADPH. These findings were

-12-
confirmed by Grant (71). &yan and Engel (86) have shawn that 21-hy
droxylation is also HADPH-dependent.
The specificity of 1~-hydroxylase for RADPH bas alao been demon
strated in the testis (91, 92), and this ia implicated in the adrenal
(34, 93). It ia also well establisbed tbat RADPB or RADPH-generating
systems atimulate steroid production 1! vitro (94, 95, 96). It ia
now generally accepted that ateroid hydroxylation is RADPH-dependent.
Lately, Halkerston et al. (63) and Constantopoulos et al. (97) have
demonatrated tbat NADPH is alao required for the cleavage of the cho
lesterol aide chain. Whether this is associated with the bydroxylation
at the 20 and 22 positions is not yet clear.
d) R&DPH•senerating Systems
We just discussed the importance of RADPH in ateroid hydroxylation.
It will be informative to know the generation of this co-factor in the
adrenal.
Glock and McLean (98), and Kelly et al. (99) have found that the
adrenal cortex containa a higb levet of G-6-P debydrogenaae, and 6-P
gluconic debydrogenase. aecently, McKerna (lOO) bas made a syatematic
study of the dehydrogenaaes in the beef adrenal cortex, and the rat ad
renale. He bas found that in botb species G-6-P debydrogenaae bas the
bigbest activity. Next come 6-phospbogluconic debydrogenase, iaocitric
dehydrogenase and matie dehydrogenase in decreasing order of activity.
It will be clear that the source of hydrogen atoas must be froœ the
aubatrates of the specifie dehydrogenaaes. Ultimately, these substrates
aust be derived froœ glucose. Gordon (101) bas shawn that the adrenal
cortex readily metabolizea glucose which stbaalatea glycolysia ander

-13-
both aerobic and anaerobie conditions. However, the oxidative pathway
is stiœulated to a much greater extent. Respiration without glucose
declines over the 3-hour period, but with glucose it remaina constant
during that period. Aeeording to Haynes' hypothesis (94), the stimu
lating effeet of ACTH on cortieoid synthesis is, in essence, to increase
the breakdown of glycogen to G-6-P, which is then oxidized, produeing
more NADPH as a result. However, Gordon (101) bas evidence that the
adrenal gland does not store much glycogen, and would depend in large
measure on the rapid rate of oxidation of the eirculating glucose.
e) Mechanism of Steroid Hydroxylation
The mechanism for steroid hydroxylation is not clear. It is
postulated (102) that the process involved activation of moleeular
oxygen and the substrate before transfer of the oxygen atom can take
place. The nature of the role of NADPH in the process is not known.
It is speculated that some metal ions, and certain catalytic proteine
may play a part in the activation mechanism. Lately, a new eytochrome
(P-450) found in the adrenal mitochondrie seems also to be involved
in the activation (102a).
f) Bffects of Freezin& on Tissue Response to NADPH
Most interesting and relevant to the present etudies is the ob
servation first made by Raines (120) that freezing increases the corti
steroid yield in hog adrenals. Koritz and Peron (95) found that freez
ing of the adrenal tissue before incubation inereases the response to
G-6-P and NADP stimulation. The latter authors (96) have also noted
that freezing bas the same effeet on the rat adrenal homogenates in the

-14-
presence of NADP and G-6-P. More recently, Hall and Koritz (103)
have provided evidence that freezing of the adrenal homogenates
causes a greater increase in conversion of la-labelled cholesterol
than of endogenous precursors to corticosterone in the presence of
the above additives. The mechanism of stimulation by freezing is
not known.
Origin of the present Study
In the beginning of his study, the author was doing adrensl
incubations in connection with the affects of Angiotensin on aldo·
sterone secretion. In the course of these experimenta, he was
attracted by one observation, i.e., the incubation medium alwsys
turns turbid after incubation with the tissue. He was interested
to know if this turbidity could be caused by the presence of enzymes
or particles containing the enzymes, sinee these partieles are
sufficiently big to obstruct light.
He has sinee been involved in this study, without realizing
for some time that Schtinbaum et al. (104), in 1959, has already
detected NADP-dependent dehydrogenase activity in the incubation or
pre-incubation medium from the rat adrenal quartera.
The main findings are contained in this thesis.

-15-
II. HATI1UALS ABD MI'1'BODS
1. Materials
a) Baffer Solution (KIBG)
Xrebs-Ringer-bicarbouate solution (pB 7.4) containing 200 -a 1
glucose, saturated with 951 o2-S% co2 (lOS).
b) Additives (per al incubation aediuœ)
i) NAD
ii) NADP
. •
:
0.39 p.1110le
0.83 IJ.DlOle
iii) G-6-P : 2.53 ~ole
T'beee ch811lical8 were purchaaed frœ the Sipua Cheaical Company
in their sodium salta (except NAD) and were readily soluble in the
buffer solution at pB 7.4. T'bey were prepared and added t.mediately
prior to incubations.
c) Labelled Steroid Substrates
14c-labelled progesterone, and/or 14c- or la-tabelled pregneuolone
have been uaed as steroid precursora. T'beee compound& were obtained
from New England Nuclear Corp. The purity of the compouuda was eatab-
lished either before or sûaultaneoualy with the experiaeats. The radio-
active .aterials vere diluted with the noa-radioactive standards so that
the specifie activitiea were uaually within the range of 4-8 x 106 c.p.a.
/p.mole. The aaounts of suhstratea uaed varied slightly la the neighbor-
hood of 0.17 p.mole per l.S g tissue per 15 ml incubation medium. The
aaounts of tissue and the incubation volumes were always kept constant
unlesa otherwise atated.
d) Vials and Glassware
All glaasware and vials were clean and free of radioactivity before

-16-
use. Vials for radioaetivity eounts were used ouly ouce.
Glassware after use was thoroaghly rinaed vith methanol and vater,
kept in ehroaiaa aulfurie eleauiug solution for two daya, and rinsed
again for several hours vith tap vater. The final rinsing was doue
vith distilled vater.
2. Incubation Procedure
a) &eaaon for Choiee of Tissue
For the purpose of preparing the pre-incubation aedi• (PIM),
bovine adrenal cortical allees vere used moat of the t~e. Rat adrenal
tissue waa used only twiee, the first t~ being the pilot experiœent.
The reaaon for this is that theae expert-enta, as it haa been explained
before, branehed out froa the original exper~ents vith Angiotenain,
whieh requtred good separation of the zona gloaerulosa, the bovine
adrenal being a suitable gland for this purpoee.
b) Bovine Adrenal Slieea
Bovine adrenals vere obtained froa a local alaaghter-bouse. The
glands vere kept ehilled until pre-incubation whieb vas started about
two hours after receiving the glands. Pat was removed frOID the glands.
The method of Stachenko and Giroud (30) vas followed cloaely to separate
the zona glomerulosa from the zona fascieulata; sliees vere made by
cutting the gland parallel to its surface vith the Stadie-Riggs micro
tome (106). The first alice was taken as the gloœeruloaa tissue. The
second was diaearded. The third was taken as the fascieulata. The
glomerulosa and the faaciculata slicea can be usually differentiated
by the naked eye, the former being light yellowish, and the latter

-17-
brownish. When slices of the whole cortex were needed. the gland vas
eut at right angles to ita surface; the aedullary tissue vas removed.
In all cases. slices vere about 0.3 mm thick. and 15 mm long.
c) Jlat Adrenal Tissue
llat adrenals vere obtained frOil lUlle Sprague-Dawley rats, veigh·
ing 150-180 g. These aniiiUlla vere anesthetized vith Neahutal (0.3 mlfanimal
intraperitoneally). Bach adreual vas cleaned free of fat. Quartered
or whole aclrenals vere ueed. The adreual tissue was kept cold in
IIBG until ueed.
d) Preparation of Pre-incubation Media. (PI!Q
Bovine cortical alices or rat adrenals (vbole or quartered) were
vashed three timea vith 10 ml KIBG just before blottiug and veigbing.
1.5 g of bovine cortical slicea or 400-750 ag of whole or quartered
rat adrenals vere weighed out on an electrical or aicrotoraion balance
and placed in a lOO al beaker containing 15 al IBIG. In any one ex-
periment, there were uaually 2·8 auch pre-incubations depending on the
design of the experiment. The beakers were incubated in a Dubnoff
Metabolic lncubator, shaking at 65 cycles per ainute under 951 o2-51
oo2 at 38° C (unlesa otberwise atated) for 1 hour. At the end of the
pre-incubation period, after allowing the tissue to settle, the pre
incubation aedia (the term being uaed after Saffran and Baylias (80),
who vere the firat to introduce pre-incubation) fraa all the beakers
were pooled into a 250 al flask and gently swirled to euaure unifora
mixing.
c) Incubation of PIM with Steroid Precursors
Tbrougbout the exper1ments, attention vas focused on the couver•

-18-
sion producta froa the pre-incubation media. However, in one expert-
ment, coapariaons vere aade between the PIM, and the tissue in con
verting added labelled progesterone. ln this case, the tissue vas
alao used after pre-incubation. Steroid precursors dissolved in
ethanol were introduced into lOO ml incubation beakers. The ethanol
was evaporated by gentle waraing. Co-factors vere added as desired.
15 ml (except otherwise stated) of PIM were finally pipetted into the
beakers, which vere then incubated in the Dubnoff Metabolic Incubator
for 3 hours under the saœe conditions as described for the pre-ineu-
bation.
3. Extraction of Steroids
At the end of the three-hour incubation period, the media were
extracted three timea with two volumes (30 ml) of freahly distilled
ehloroform-ethyl acetate (1:1). The extracts were dehydrated over
anhydrous sodium sulfate, filtered, and taken to drynesa under redueed
pressure at 50° c. The reaidue vas redisaolved in 10 ml distilled
methanol, from whieh a 0.1 ml aliquot (i.e., 11) was transferred to a
vial for radioactivity meaaurement. (The method will be described
later). Prom the values obtained in c.p.m., the total radioaetivity
recovered at this stage wae ealeulated.
The extraction of steroids in the tissue was as follows:
The tissue was homogenized vith S ml KRBG and extracted witb 3 volumes
0 of diatilled acetone over-nigbt at 4 C, during which period the protein
was alao preeipitated. The precipitate was filtered off and washed 3
timea with 10 ml of fresh acetone. The acetone vas evaporated under

-19-
0 reduced pressure at 35 C leaving behind only the aqueous residue.
Bereafter, the extraction of the steroids from the aqueous solution
vas the saœe as described for the media. before, with one addition:
the extract was vashed once each vith 1/2 volume of IN NaOB, IN HCl,
and finally distilled water in order to remove soae of the fat froœ
the extract.
4. Paper Chromatol!aPh7 for the Separation of Steroids
a) Preparation of Paper
Whatman No. 1 chromatography papers, 56 ca long and 17 ca vide,
vere washed in a Soxhlet apparatua with methanol-benzene (1:1) for 3
days. The papers vere huns in the dark to dry and then stored in a
folder to protect them from dust. The washed paper vas eut along
the le~gth to form strips, 1.5- 2 cœwide, 43 ca long, attached to
a coœmon head 13 ca long. The line of application (starting line)
vas 2 ca from the common head.
b) Application of Steroids
Concentrated steroid extracts were quantitatively transferred
to a 15 ml centrifuge tube and evaporated to dryness under a stream
0 of nitrogen in a vater-bath, kept under 50 c. The residue was dis-
solved vith 2 drops of methanol-benzene (1:1) and applied to the paper
along the starting line by means of a capillary pipette. The solvant
mixture vas evaporated vith a dry stream of air. The wbole operation
was repeated twice to ensure complete transfer.
c) Solvent Szstems uaed
The solvant systems of Bush (107) and of Zaffaroni and Burton

-20-
(108) vere useô. The for.er systems require an equilibration perioô of
froa 3/4 to 12 hours before the addition of the mobile phase. In the
latter systems impregnation of the paper before application of ateroiôa
is necessary: the paper was wetteô by passing through the stationary
phase ôiluteô with an equal volume of methanol. The paper was blotted
between clean sheets of Whataan No. 2 filter paper and hung up for 3 ain.
before application. After application the paper waa run right away in
a chromatography jar without equilibration. All chromatogr ... vere de
velopeô by the descending aetbod in a constant te111Perature room at 2f c.
The systeme used are listed below:
1) The Bush Solvent Szstell8
Names and Ref. Mobile Pbaae/Stationarz Phase
(10 7) Benzene/aetbanol :water 100 50 : 50
Bush C (107) Toluene:ethylacetate/methanol:water 90 : 10 50 : 50
Bush A (107) Petroleum ether/methanol:water lOO 80 : 20
(109) Iao-octane/n-butanol:water lOO 50 : 90
ii) Zaffaroni SiatellS
Equilibration Time
45 ain.
45 min.
45 min.
12 hra ..
The mobile ana atationary phases are mutually saturated with respect
to each other (exception: T.B.G.)
Na.ea and &ef. Mobile Phase/Stationarz Phase
T.E.G. (llO)
L.P .G. (111)
T.P .G. (112)
Toluene/ethylene glycol
Ligroin/propylene glycol
Toluene/propylene glycol
1

-21-
s. Detection of Steroids on Chromatosr ...
After development of the chromatogra., the paper vas dried (3 ain.
- 12 hrs.) at room ta.perature. Areas containing ateroida on the strips
were located by~ a) ultraviolet absorption and/or, b) radioactivity
detection.
a) Ultraviolet Ligbt Absorption
Steroids poasessing an a-p unsaturatei ketone group absorb ultra
violet light at 238-240 mp.
The paper waa exposed to ultraviolet light source using a Corning
filter no. 9863. The areas of absorption would appaar as dark spots
(sensitivity: 2 ~g steroid(s)jca2). The positions of the spots were
either aarked with a pencil or photographei on the Kodak standarè
contact photographie paper.
b) ladioactivity Detection
Radioactive conversion products derived froa 14c. or la-labelled
precursors were detected by, (i) radioautography and/or, (ii) radio
chr0118togra scanner.
1) Bx &adioautographx
The chromatograa was covered with a no sereen Kodak sensitivity
X-ray fila in a light-tight standard X-ray cassette for 3 days. The
fila was then developed by standard X-ray fila developing techni4ues.
&adioactivity of 2,000-3,000 disintegration per ainute of 14c could
be detected on the film after a 3-day exposure. The filawas not
sensitive to la-labelled compound& under these conditions.
ii) By bdiochromatogra Scanner (Packard Model 7200)
Tbe paper was scanned on the apparatus at the desired sensitivity

•
-22-
and speed. the scanner can detect both 14c. and Ja-tabelled materiel.
6. Blution of Steroida froœ the Paper Strips
Areas on the etripa containing the steroids were eut out and
eluted with 30 al of ketone-free, 3-times distilled ethanol fra. a
30-al syringe uaing a 22-gauge needle, especially prepared (bent at
right angles) for this purpose. At the end of the elution, the
ethanol solution vas evaporated in a flash avaporator under 5~ c.
7. guantitative Deteraination of Steroids
The ~4-3-keto steroids were deterained quantitatively by (a)
the ultraviolet light absorption method and (b) radioactivity measure
ment. The foraer measured both the endogenous production as well as
conversion products froa added radioactive steroid precursors, whereas
the latter aeasured only the conversion products.
a) Ultraviolet Lis!t Absorption Method
The steroid was dissolved in a known voluae of ethanol (3-5 al),
and transferred to a silica cell (3 ml capacity). The solution vas
measured at 240 ~ (teated aax~l absorption for these compound&) in
Beckman DU Spectrophotometer against the paper blank (derived from
the blank incubation which contained only the buffer solution). The
paper blank vas also aeasured against the reagent blank (ethanol) to
check its cleanliness. The light source was B2 lamp. A standard
curve relating the optical density (O.D.), and the concentration (in
~g/al) of the corresponding authentic compound waa made. This is a
linear function within limita of concentrations. Once the O.D. vas
known, the amounts of steroid in solution could be deterained froa

-
•
-23-
the standard curve.
Individuel ateroids were measured at least after two paper chro-
matographic purifications. Biah paper blank absorption at 240 ~ was
seldom encountered (O.D. range: 0.05-0.15). It was therefore con
sidered unaeceaaary to apply the Allen (113) correction. aince in
theae experimenta, relative. rather than absolute values vere of
interest.
b) ladioactivitX Measurement
Radioactivity of 14c- or la-labelled compound& vas measured in
a Packard liquid scintillation spectrometer. Materiels were counted
twice. A blank and radioactive standard vere included in a set. Ali
quota representing 1/100 to 1/30 of the total aample were transferred
in methanol to the counting vials. The methanol vas evaporated in air
or under a strea. of nitrogen and redissolved in 5 ml of scintillation
phosphor (toluene pbosphor containing 0.31 of 2,5-diphenyloxazole, and
0.01% of p-bis 1,2(5 phenyloxazolyl) 1-benzene). The efficiency for
14c and 3a eounts was 701 and 371 respectively, with an aceuracy of
± 21.
8. Sxstematic Separation of Steroida
Tbe paper chromatographie systems used bave been described in
detail. A systematic separation of the steroids of interest will now
be given.
The initial separation of the steroids froa the incubation extracta
was carried out in the Bush a5 system for 3 hours. In order of de
creasing polarity, the fractions located parallel to authentic 18-0B-

•
-24-
corticosterone, cortisol, aldosterone, cortisone and progesterone
(the solvent-front fraction) were eluted. The fractions in the
regions of cortisol, aldosterone and cortisone were re-chromato
graphed in the Bush C system for 3 hours. (In this system, aldo
sterone separates vell from cortisone). The fraction in the region
of corticosterone was re-chromatographed in the B2B system for 5
hours. (In this system, 11-deoxycortisol will be separated from
corticosterone). The solvent-front fraction was re-cbromatographed
in the LPG system for either 8 or 50 hours. In the 8-hour run,
pregnenolone, progesterone, or cholesterol would be well separated
from the more polar steroids, e.g., tl--OB-progesterone, 17a-OB
progesterone, 11-deoxycorticosterone, and 11-keto-progesterone.
These compound& would still be poorly or not separated at all. In
the 50-hour run, pregnenolone, progesterone, or cholesterol would
have run off the paper, being collected over this period in a flask.
The more polar steroids, as mentioned above, would be well separated.
All quantitative measurements of the coœpounds (unless otherwise
stated) were measured after purification by two systems of paper
chromatography described above.
9. Characterization of Corticosterone1 Cortisol, and ll@-Bydroxy-
2rosesterone
A number of conversion products were obtained when the PIM from
the beef or rat adrenal tissue vas incubated with labelled pregneno
lone or progesterone. However, only the ones of interest to us vere
partially characterized. They vere: Corticosterone and/or Cortisol

•
-25-
fro. the beef or rat adrenal PIK and 11~-0H-progeaterone from the rat
PŒ.
ln general~ proof of identity of the respective compounds waa
baaed on the fo11owing criteria:
1) ldentical aobility of the radioactive tentative compound with that
of the non-radioactive authentic carrier in at leaat ~o paper chro
matographie systems before acetylation or oxidation~ and in one
paper systea after acetylation or oxidation.
2) Maintenance of constant specifie activity in the ayateas run througb
as deacribed in (1) above~ and after successive cryatallization.
(This waa perforaed only on 11~-oH-progeaterone).
3) Max~um u.v. absorption in the region between 238-242 ~·
4) Tests of functional groups with,
(a) Tetrazolium derivative M and B 1767, and
(b) Porter-Silber reagent.
The paper chromatographie systems uaed for the partial character
ization of the individual compound& will be indicated in "aeaults."
Present1y, the methoda for acetylation, oxidation, cryatallization,
and tests on functional groupa of the steroida will be given.
Acetzlation
The method described by Bush (114) was followed. The steroid
(30-80 ~g) waa diaaolved in a aixture of pyridine (0.1 ml) and acetic
anhydride (0.4 ml), and heated at 60P C for 15 min. Under these con
ditions, the 21-hydroxyl group of ateroid a-ketols is completely es
sterified, and the 18-hydroxyl group of aldosterone (hemiacetal fora)

•
-26-
is only esterified to 15X. The reagenta were tben reaoved by a stream
0 of aitrogen at 50 c. The ateroid (acetylated or not) could be directly
applied to paper.
Oxidation
The ateroid (30-80 ~8) waa disaolved in 1 al of glacial acetic
acid, and an aqueous solution of 0.5 ml chromic acid (2X chroatua
trioxide) was added. The tube waa shaken gently, atoppered and left
in the dark for 10 min. Under these conditions, the 11~- and 11a-
hydroxyl groups of the typical C-19 and C-21 steroida are oxidized
coapletely, while the side chain of the C-21 steroids remains intact.
At the end of 10 min., the solution was diluted with 5 volumes of
distilled water and extracted 4 tiRes with an equal voluae of athyl-
acetate-chloroforœ. The organic phase was washed with 1 ml saturated
sodium bicarbonate and water, dried over Na2so4, filtered and evap
orated.
Crystallization of 11@-oH-Proaesterone
A saall fraction (5-7 ~8) of the isolated radioactive product
was mixed with approximately 10 mg of non-radioactive authentic coa-
pound (kind1y given by Dr. W. Klyne), and disaovled in sufficient
quantity of acetone. The solution was filtered through a sintered
glass funnel to remove contaminants (mainly fibre) froa the paper
chromatograms. The filtrate was evaporated to dryness. Crystalli-
zation waa carried out in a small tube using three different solvant
mixtures: (a) acetone-methanol, (b) acetone-benzene, and (c)
acetone-ethanol. The steroid or steroid mixutre was diasolved in a

•
-27-
minimal volume of wara acetone, the 'good' solvent in wbich the steroid
dissolves well. About half volume of a second solvent (methanol, benzene
or ethanol) was added, in which the steroid is only sparingly soluble.
Upon gentle heating over a steam-water bath, acetone being more volatile,
would be evaporated first. When acetone was almost completely evaporated,
crystal& would appear. Occasional cooling and standing was sometiœes
necessary. After crystallizatian was completed, the mother liquor was
withdrawn vith an ultra-fine, self-..de transfer pipette. Samples of
crystals and mother liquor were dried, weighed and taken for radioactivity
counts.
Test for a-ketol sroup
c-21 steroids having a a-ketol group in the side chain reduce
tetrazolium salts (116).
2 mg of the tetrazolium derivative M and B 1767 (2:5-diphenyl-3·
(4-styrylpbenol)-tetrazolium chloride) was dissolved in 2 ml of 95~
ethanol and 5 ml of 2.5 N NaOH (123).
1/2-cm strips containing about 5 ~g of the isolated steroids were
passed through the solution, washed once with distilled water and
blotted in between two sheets of filter paper. A reference strip con
taining 2 ~g corticosterone gave a purple color tœœediately, while 6 ~g
of authentic progesterone did not.
Test for the dihydrO!X•acetone aide chain
The Porter-Silber reagent (117, 118) reacts with the 17, 21-di·
hydroxy-20-keto-steroids, as we11 as 21-hydroxysteroids with a 20-18
hemiketal linkage (except those with a hydroxyl group at c-11 (119) •
Paper strips containing the steroids were passed through the

•
-28-
Porter-Silber reagent. A reference strip containing 3 ~~ of cortisol
would give a yellow color in 1/2 hour at 60P c •

•
-29-
Ill. USULTS
BXPRISSIONS Of DSULTS
Resulta are expreased qualitatively and quantitatively. The
quantitative data are expreased in ~oles, specifie activities,
and/or per cent conversion of the labelled precursors. The per
cent conversion is based on the total radioactivity recovered froœ
the incubation medium with ethylacetate-chloroforœ, and not based
on the radioactivity initially added. This method of expression is
warranted from the following considerations: (a) Extractions of
known amounts of progesterone, corticosterone, and cortisol with
ethylacetate-chloroform were reaaonably high and close to each other
(85~-96~), with progesterone having the highest percent recovery.
Resulta froœ a control experiment are ahown below:
CO!!Pouncla Adele cl
(c.p ••• x 105)
Progesterone-4-14c (52 J18) 4.37
.. 4.36
CO!IJ?ounds Acldecl
Corticosterone
Il 30
Cortisol 30
n 30
aecovered
(c.p ••• x 105)
4.15
4.22
B.ecoverecl
26.0
26.6
25.3
25.8
~ &ecovery
94.7
96.8
~ aecovery
85.3
86.7
84.3
86.0

(b) From the actuel experimenta it had been observed that the re•
coveries of radioactivity from a set of incubations remained remark-
ably constant whether or not there had been conversion of the added
precursor. This can only be true if the recoveriea of the conver-
sion products and the added precursor are st.ilar. The fo11owing
figures taken from two experimenta will illustrate the point. lach
figure representa a single incubation;
Total ladioactivitX &ecovered
Experimenta Precursor No Conversion Good Conversion
X•7 Progesterone 6 1.48 X 10 c.p.m. 1.43 x 106 c.p.m.
" 1.47 x 106 c.p.m.
" 1.37 x 106 c.p.a.
X-12 Progesterone 6.95 X 106 c.p.a. 6.55 x 106 c.p.a.
" 7.15 x 106 c.p.m.
" 6.96 x 106 c.p.m.
!hua, on the grounds that the recoveriea of the compound& are
high and similar to each other. it is conaidered justified to base
the per cent conversion on the total radioactivity recovered from
the extraction of the incubation mediuœ, aince the relative values
obtained under this condition will remain unaltered.
Th~ alternative way of expression, baaed on the radioactivity
added, auffers from the following inconveniences or disadvantages:
i) It necessitates the counting of an extra saœple to estab1iah
• the actuel activity added •

-31-
ii} The percentage calculated under this condition will be lover
than the other, since the tosses occurring during extractions
are not compenaated.
SBCliON A
Under this section, it will be sbovn tbat the pre-incubation
media contain a number of enzymes involved in steroid metabolism,
and the conditions under which this can be demonstrated. It should
be emphasized that the resulta obtained here do not indicate whether
the enzymes exist in the free state or are contained in the sub
cellular particles.
1. Incubation with Labelled Progesterone
Pigure 1, (Page 32} shows the resulta of a pilot axperiment in
which labelled progesterone was incubated with the PIM prepared from
the rat ~uartered adrenals in the absence or presence of NADP and
G-6-P (collectively called 'co-factors' in Pig. 1). The initial
separation of the conversion products in the Bush Bs system are shovn
on the left. The re-chromatography of the solvent-front fraction
from the Bush B5 system to the LPG system ia indicated on the right.
To the left of each coluœn are the ultraviolet photocopies, and to
the right, the corresponding radioautograms. The ultraviolet photo
copies show, indiscriminately, the products released to the PIM from
the tissue during the pre-incubation period, as well as the conversion
products derived froœ the labelled precursor during the period of
incubation. The radioautograu show exclusively the conversion

•
•
FIGUBB I
CoDversion of Progesterone-4-14c by PIK
(llat Adrenal Quartera)

-CONVERSION of PROGESTERONE - 4 - 14c by P 1 M
( Rat Adrenal Quarters)
no PIM PIM alone PIM +
Co-factors
Bush 8 5 3 hrs.
STD
Cortisol
Aldosterone
Corticosterone
x
PIM alone PIM +
Co-factors
L PG 30hrs.
•
STD
DOC

Compound a
up .. oa-P
Corticosterone
-33-
TABLE I
Conversion of Progesterone-4-14c by PIM
(Rat Adrenal Quartera)
~ Conversionl3 hra. Yieldl3 hra.~Qmo1el
c.p.a. /incubation /incubation
4.5 x 105 23.1 0.040
3.6 x 105 18.9 0.033
Incubation aixture contained: Progesterone·4-14c (0.189 ~le or 59.4
~g) with a total radioactivity of 1.95 x 106 c.p.a.;
NADP (0.38 J.llllOle/DÜ); G-6-P (2.53 J.liDOle/al); PIM (15 DÜ), prepared
froa an equivalent of 450 œg rat adrenal quartera pre-incuhated for 1
hour.

-34-
products from the labelled precursor. Fig. 1 is intended to show
the following:
i) The purity of the labelled precuraor used, by chromatographie
standard (first column, Bush Bs)•
ii) The inability of the PIM to convert labelled progesterone with
out the addition of co-factors. (Two small radioactive banda
can be aeen on the firat column of the LPG system. However,
quantitatively, these are too amall to be of any significance).
iii) The capacity of the PIM to couvert labelled progesterone in the
presence of the added co-factors (third column, Bush Bs, and
second column, LPG). More than two radioactive fractions were
detected, with the conspicuous absence of aldosterone. However,
interest was focused only on two main fractions: (a) the frac
tion in the region of authentic corticosterone, and, (b) frac
tion X in the region of authentic lltJ-OH-progesterone, which is,
however, not on the standard strip in Fig. 1 because this com
pound was not available at the time of this experiment. Both
fractions in the respective regions mentioned bave been partially
characterized as corticosterone and 11~-0H-progesterone respec
tively (see Section D). Table 1, (Page 33) gives the quantitative
analysis of the two main fractions after each bas been run through
two systems of paper chromatography as described under 1Methods'.
lt will be aeen that under these conditions, !l-B-OH-progesterone
was produced in slightly larger amounts than corticosterone.
From here on, except in one case, all the experimental resulta are from
the bovine adrenal PIM incubations. As will be clear from Pig. 2,

-35-
PIGUU 2
Conversion Products from Progesterone-4-14c by PIM
(Bovine Adrenals)

e
CONVERSION PRODUCTS from PROGESTERONE- 4- 14C by PIM
Glomerulosa
G-6-P NADP G-6-P
(Bovine Ad renais)
STD
Cortisol
Cortisone
Corticosterone
Progesterone
Bush B5
Fasciculata
G-6-P NADP G-6-P
STD
Cortisol
Aldosterone Cortisone
Corticosterone
Progesterone
e

Ceounds
Corticosteron.e
Corticosterone
Cortisol
Cortisol
Ccapound S
-36-
TABLE Il
Conversion of Progeaterone-4•14c by PIM
{Bovine Adrenala)
Yieldl3 hrs.~~aole) ~ Converaionl3 hrs.
/incubation c.e.m. /incubation
(Z .. P.) 0.033 2.4 x 105 17.52
{Z.G.) 0.034 2. 7 x 105 18.53
(Z.P .) 0.041 1.9 x 105 14.10
(Z.G.) 1.1 x 104 0.76
(Z.P .) 0.019 1.2 x to5 8.75
Bach incubation mixture contained: Progeaterone-4-14c (0.181 ~ole or
56.8 ~g) with a total radioactivity of 1.42 x 106 e.p.m.; RADP (0.83
~le/ml); G-6-P {2.53 ~le/ml); PIK (15 ml) prepared froa 1.5 g tis
sue pre-in.cubated for one hour.
Z.G. • zona glo.eruloaa
z.r. - zona fasciculata

-37-
(Page 35) no less than six or seven radioactive conversion products
vere always obtained, when labelled progesterone was used as the
precursor. However, it is beyond the scope of this study, nor are
there enough authentic materiels to analyse each of these fractions.
Rather, relatively detailed qualitative and quantitative studies
vere concentrated on the two main fractions in the regions of authentic
cortisol and corticosterone initially isolated in the Bush Bs systea.
lt is thought that this approach will throw soae ligbt on soae of the
facts about the PIM.
Fig. 2 shows the distribution of the various radioactive frac
tions in the Bush B5 systea from a comparative study in which the PIM
prepared froa the isolated glomerulosa and fasciculata vere incubated
with labelled progesterone. More detailed quantitative resulta will be
shown in Table Il, (Page 36) after re-chroaatography of the main frac
tions in the appropriate systema as bas been indicated under '~thods."
However, it will suffice to point out that no radioactive materiel in
the region of authentic aldosterone was detected under various incu
bation conditions froa the PIM, including the one prepared froa the
gloaerulosa tissue. It will also be noted froa Fig. 2 that G-6-P
added by itself did not activate the enzyae activities. Table Il
shows clearly that althougb the radioactive fraction in the region of
standard corticosterone was produced in about equal quantity by the
PIM from both zones, the radioactive products in the regions of cor
tisol and compound S vere obtained almost exclusively froa the fas
ciculata•PIM incubate. Tbese findings are in agreement vith the known
fact that the l~hydroxylating enzyme is located mainly in the

-38-
Z.F., as it has been discuased in the'aeview of Literature'.
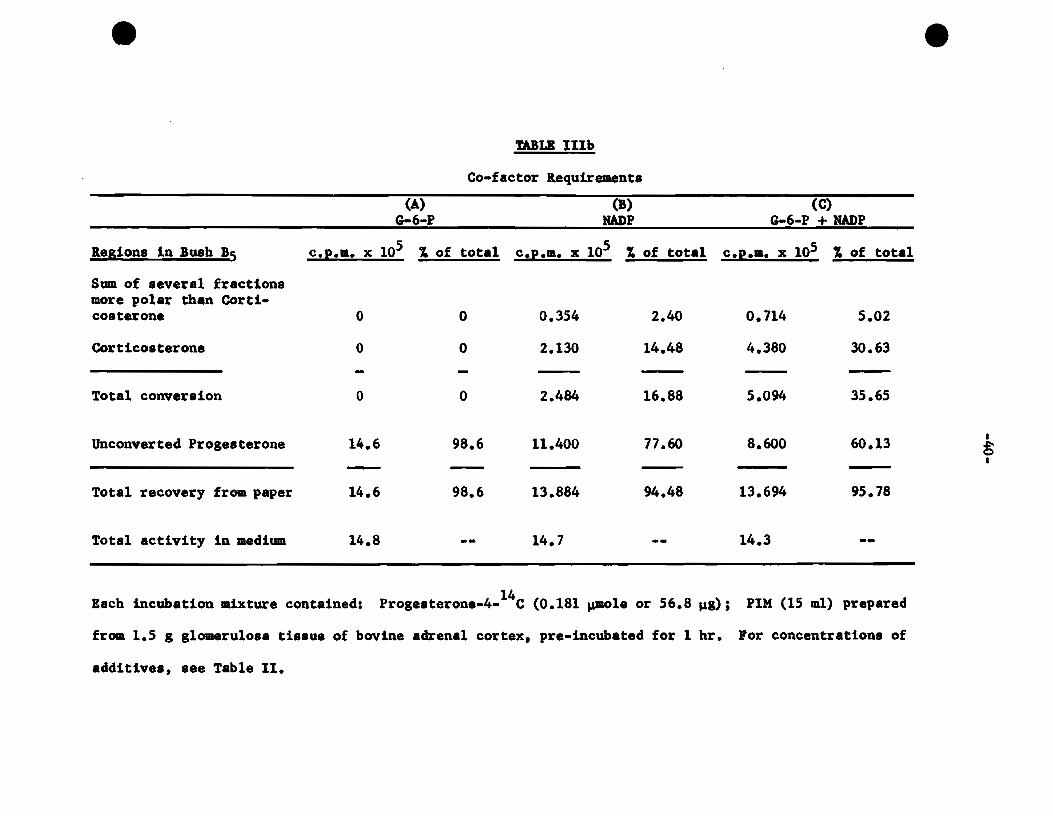
2. Co-factor Reguirements
It bas been indicated in Fig. 1 and Fig. 2 on Page 32 and
Page 35 respectively, that the demonstration of enzyme activities
in the PIM requires the supplementation of both NADP and G-6-P,
and that the latter by itaelf is ineffective. The object of the
next two experimenta was to find out whether NADP was the actuel
co-factor required, or that both additives were needed. The PIM
from the Z.G. were incubated with labelled progesterone in the pre
sence of either or both additives. The resulte of the two experi
menta vere tabulated separately in Tables Ilia, (Page 39) and IIIb,
(Page 40), because the specifie activitiea of progesterone used in
the two cases were different, and in one experiment (Table Ilia)
double the uaual amounts of PIM were uaed (30 ml). The resulta in
both Tables give the counts and percentage of all the radioactive
fractions aeparated in the Bush B5 system in order to give a better
account of the over-all conversions. On the other band, the sub
fractions further isolated from the main fraction in the region of
corticosterone, and from the aolvent-front fraction were not listed
separately, because individually, theae are quantitatively insignifi
cant. However, it should be stated that the fraction in the region
of corticosterone in the Bush 15 system contains more than 601 of the
compound, and the solvant-front fraction contains well over 95~ of
unconverted progesterone. With this as the background, the resulta
sbown in Tables Ilia and IIIb can be interpreted witb more accuracy.

e e
TABLE Ilia
Co-factor Requirements
(A) (B) (C) G-6-P HADP G-6-P + NADP
Restons in Bush B5 ~·.P~ m. x 10 6 l Qf_total c.p.~ 10 6 ~ of total C_tl)elll• X 10 6 t. of total
18-0B•B * 0 0 0.01 0.83 0.02 1.82
Cortisol 0 0 o.o1 0.83 o.o4 3.63
19-0R•DOC ** 0 0 0.03 2.50 o.o3 2.73
Corticosterone 0 0 0.36 30.00 0.58 52.72
- - - - -Total conversion 0 0 0.41 34.16 0.67 60.90
Unconverted Progesterone 1.10 91.66 0.50 41.60 0.20 18.18
- - -Total recovery from paper 1.10 91.66 0.91 75.76 0.87 79.08
Total activity in mediua 1.2 --- 1.2 --- 1.1
Bach incubated mixture contained: Progesterone-4-14c (0.198 pmole or 62 ~g); PIM (30 ml) prepared from an equivalent of 3 g glomeruloaa tisaue of bovine adrena1 cortex pre-incubated for 1 hr. Por concentrations of additives, see Table 11.
* Based only on chromatographie mobility.
** This position corresponds with that of a band characterized in this Departœent as 19-0R•DOC by Lucia, a. et al. (121).
• w \0 1
e e
TABLB Illb
Co-factor aequirementa
(A) (B) ·(c) G-6-P NADP G-6-P + NADP
Regions in Bush Bs ç~p.a., li: l.Q 5 l of total c.p.a. x 10 5 %. of total c.J!.aa. x 105 1 of total
Sum of severa! fractions more polar than Corti-coster one 0 0 0.354 2.40 0.714 5.02
Corticosterone 0 0 2.130 14.48 4.380 30.63
- -Total conversion 0 0 2.484 16.88 5.o94 35.65
Unconverted Progesterone 14.6 98.6 11.400 77.60 8.600 60.13 1 g 1 -
Total recovery from paper 14.6 98.6 13.884 94.48 13.694 95.78
Total activity in medium 14.8 -- 14.7 -- 14.3 --14 Bach incubation mixture contained: Progesterone-4- C (0.181 1J.180le or 56.8 Jlg); PIM (15 ml) prepared
froa 1.5 g glomerulosa tissue of bovine adrenal cortext pre-incubated for 1 hr. For concentrations of
additives, see Table II.

-41-
It will be noted that in both experimenta, no conversion was
observed when only G-6-P was added. The recoveries of unconverted
progesterone under this condition were 91.66~ and 98.6X reapectively
in the two experimente. The average recovery was within the l~it
of wbat is expected from any elution of standard progesterone. It
can therefore be concluded that there was no conversion in the pre
sence of G-6-P.
Wben NADP was added, the conversion observed, both totally,
and in the main fraction were about half of those in which both NADP
and G-6-P were present (Tables Ilia and IIIb). The corresponding
unconverted progesterone recovered was also given. Total recovery
froœ the papergram was given in each case. It will be seen that the
recovery was lover when there was conversion than when there was no
conversion. This can be explained by the fact that some of the con
version products may be more difficult to elute, and also, the more
the number of elutions, the greater will be the chances of losses.
From the findings of these two experimenta, it can be concluded
that NADP is the only exogenous co-factor required to activate the
enzymes present in the PIM. G-6-P by itself is totally ineffective,
but its addition to NADP will further enhance the enzyme activities.
3. Incubation with Labelled Preggenolone
In the foregoing experimenta we have seen that the pre-incubation
media contain enzymes capable of transforming progesterone to a number
of its metabolites. The purpose of the next two experimente is to
find out whether the PIM also have the capacity to transform pregneno-

-42-
lone~ a C-21-45·38-ol steroid~ to other ateroida possesaing a
~4-3-keto structure~ or, more specifically, to see, if ~5-JB.
hydroxysteroid dehydrogenase is also present in the PIH. At the
saœe time, the effect of HADP, NAD, or a combination of the two
were alao investigated. Sufficient amounts (more than 0.16 ~le
or 50 ~g) of labelled pregnenolone were added, so that should there
be any reaaonable transformation of the precursor to some of its
~4-3-keto metabolites, the latter could be detected on the paper
graœ by ultraviolet light. ( ~5-38-ol steroids do not absorb ultra
violet ligbt with maximum absorption at 240 ~~ whereaa ~4-3-keto
steroids do.)
In the first experiment, 15 ml of the PIM from the bovine
adrenal cortex (glomerulosa and fasciculata) were incuhated with
pregnenolone-7a-3a in the absence or presence of NAD. The resulta
are shawn in Table IVa, (Page 43). It will be seen that, without
NAD, the only product detected was in the region of progesterone
(6.61) in the LPG system. With NAD present, by far the largest
conversion product was located in the region of progesterone (63.91).
This region was clearly u.v. absorption positive. A small radio•
active fraction was also detected in the region of 17a-oH-progester
one and DOC (not separated) in the LPG system (5 hrs.), but the con
version to this fraction was only 3.71.
In the second experiœent, the PIM from the glomerulosa of the
bovine adrenal cortex was incubated with 14c-labelled pregnenolone
in the presence of NAD, NADP, or the combination of the two. In
all cases, G-6-P was added. The conversion products can be seen

-43-
TABLE IVa
Conversion of Pregneno1one-~3H by PIM
(Bovine Adreual Cortex)
a.esious
Progesterone
17a.OB•P and DOC
Unconverted Pregnenolone
Total recavery frOID two papers
No Additive
umple 1 of total ymole % of total
o.oot 6.6 o.to5 63.9
o.o o.o 3.7
11.9 8.1
78.5 15.7
3 Bach incubation mixture contained: Pregnenoloue-7a- H (0.161 ~mole)
witb a total radioactivity of 75.7 x 105 c.p.a.; PIM (15 al). NAD
(if added) : 0.39 ~le.

-44-
TABLB IVb
Conversion of Pregnenolone-4-14c by PIM
(Bovine Adrenal Glomerulosa)
NADP NAD+ NADP
Res ions ~le '1 total y.mole '%. total y.mo1e '1 total
Corticosterone o.os 0.019 11.40 0.019 11.3
Progesterone .os 51.50 0.016 9.90 .025 15.0
Cortisol o.oo 0.59 0.54
17a-oH-P +DOC 7.90 o.o o.oo
Sum 59.48 21.89 26.84
14 Bach incubation mixture contained: Pregnenolone-4- C (0.167 ~ole or
53 ~g) vith a total radioactivity of 5.98 x 105 c.p.m.;
PIM (15 ml.) For concentrations of additives. see Page 29 in 'Methode'.

-45-
FIGURB 3
Conversion of Pregaenolone-4-14c by PIH
(Glomerulosa: Bovine Adrenala}

e
CONVERSION of PREGNENOLONE - 4 - 14C by P 1 M ( Glomerulosa : Bovine Adrenals)
A B C
Corticosterone
Pregnenolone Progesterone
.. DOC ...... .
NADP NAD NADP NAD
Bush B5 3 hrs.
A' B' C' STD
NADP NAD NADP NAD
LPG 4hrs.
DOC
( Pregnenolone)
Progesterone
e

-46-
in Fig. 3, Page 45, and the quantitative resQlta in Table IVb, Page
44. The first part of the experiment (Table IVb) confirmed the re-
sulta obtained in the previous expert-eut, that is, in the presence
of NAD alone, tbe transformation of pregnenolone vas liaited aostly
to tentative progesterone. A fraction in the region of 17a-ou-P and
DOC was also obtained (7.91) but was not investigated. RADP vith
G-6-P waa also effective in activating the ~5-JB-ol steroid dehydro-
genese. The conversion products were tentative progesterone (9.9%)
and corticoaterone (11.4%) again indicating the importance of exogenoQ&
NADP in certain steroid hydroxylation reactions. The addition of RAD
to NADP st~ulate the transformation of pregnenolone to tentative
progesterone (15%), bQt not to corticosterone (11.3%). This would
&Qggest tbat NAD has little effect on the hydroxylation reactiona.
Resulta in Table IVb also indicate that the over-all conversiona in
the presence of NAD, RADP, and G-6-P were not as great as in the pre-
sence of NAD and G-6-P, suggesting inhibition by the addition of NADP.
Tbese resulta agree well with the recent findings of laval,
Forchielli and Dorfman (64) that NADP can replace NAD as the co-factor
of ~5-3p-hydroxysteroid dehydrogenaae with about lesa than 50% of the
activity. These aQthors have also found that NADH inhibits the isomeri-
zation reaction, which can be partially overcome by excessive NAD. If
this is the case, then this aay explain our resulta why addition of
NADP to NAD seeœed to inhibit the reaction, since it is obvious that
at least parts of the NADP added vith G-6-P were reduced to NADPH which
in turn might reduce parts of the NAD to NADB.

-47-
TABLE V
Pailure of the PIM to convert Cboleatero1·4-14c
(A) (1) No Additive NADP + G-6-P
Total radioactivity s s froaa aedia 8.04 x 10 9.1S x 10
Activity of labelled Cholesterol after
6.93 x lOS 8.12 x lOS two paper systems
'1 recovery of label led Cholesterol s6.r' 88.7'1
14 Bach incubation contained: Cholesterol-'+• C (0.11 ~le or 43 ~g)
PIM (30 ml). For the concentration of additives, see Table I.

-48-
4. Incubation with Labelled Cholesterol
Carballeira et al. (122) have obaerved that the bovine adrenal
cortical tissue alices aeem to utilize choleaterol-4-14c only when
aupplemented by the NADPH-generating system. It would therefore be
of intereat to know if the PIM from the bovine adrenal cortex could
metaboliae labelled cholesterol. Double the usuel amounts (30 ml
inatead of 15 ml) of PIM from the glomeruloaa were incubated with
cholesterol-4-14c (0.11 ~mole or 43 ~g) under the following conditions:
(a) witbout additives, and, (b) with NADP and G-6-P. The resulta are
shown in Table Y, (Page 47). No radioactive conversion product was
obaerved under both incubation conditions. The recoveries of un
converted labelled cholesterol after the two paper chromatographie
systems Bush 15 and LPG were 86.11 in (a) and 88. 71. in (b). Tbus,
from the negative finding of radioactive conversion products, the
high and aimilar recoveries of uncooverted cholesterol (after two
systems of paper chromatography) under both incubation conditions as
stated above, it waa concluded that the PIM did not utiliae the added
labelled cholesterol to any appreciable extent even in the presence
of HADP and G-6-P.
S. Conclusion drawn from Section A
In the light of all the experimental evidence obtained in this
section, the following main pointa ean be stated:
1) The PIM from the bovine adrenal cortex, when supplemented with
the proper co-enzymes, bas the capacity to tranaform added labelled
preguenolone or progesterone to a number of metabolites. Of theae

-49-
tentative progesterone froa pregnenolone, as well as cortico
sterone and cortisol froa added progesterone have been partially
characterized, as will be seen in Section D. On the basis of
these findings, it can be establiahed that the bovine adrenal
PIM contains the following enzymes: (1) the ~5-3p-hydroxy
steroid dehydrogenase, (ii) the llp, 17a and 21 steroid hydrox
ylases, and (iii) the NADP·dependent dehydrogenase, wbich reduce
NADP to NADPB. AIDOng these, G-6-phosphate dehydrogenase muet be
present since the addition of G-6-P greatly sttmulatea the
hydroxylation reactions.
2) Froœ the pilot expertœents, llP and 21 steroid hydroxylase acti
vities were also detected, and there is good resson to believe
that all the other enzyme activities {with the exception of 17a
hydroxylase) detected in the bovine adrenal PIK are also present
ln the rat adrenal PIM.
Sch~nbaum et al. (104) have already reported the detection of
NADP-dependent dehydrogenase activities in the PIM froa the rat
quartered adrenals.
SECTION B
1. Bffect of Tiœe on Pre-incubation
In Section A, evidence has been presented for the presence of a
number of enzymes in the pre-incubation media. lt was aasuœed that
the enzymes, in wbatever for.a, were released into the aedia froœ the
tissue during the period of pre-incubation. !be next expertment re
presents an attempt to test the above asaumption. Tissue allees from

-so-
the adrenal cortex were prepared and washed exactly as deacribed be
fore, and pre•incubated for an increasing period of time: 2, 20, 40,
and 60 minutes. At the end of the pre-incubation period, the respec
tive PIM were incubated with labelled progesterone, HADP and Q-6-P for
3 hrs. at 38° C as before. Isolated radioactive corticoaterone and
cortisol were used as the parameters, since these two steroids would
reflect the activities of the three major hydroxylating enzymes (llB,
17a, 21). &esults are indicated in Fig. 4, (Page 51). It is clear
from Fig. 4, that there were little or no enzyme activities in the PIM
in the beginning of pre-incubation, inèicating that the slices had
been thoroughly waahed. As the pre-incubation period waa lengtheneè,
the enzyme activities in the meèia also increased. This ia strong
evidence that the enzymes or particles containing the enzymes were
released into the PIM daring pre-incubation and not before, thua,
ruling out the possibility of contaminations prior to pre-incubation.
It will be noted from Fig. 4, that the curve for cortisol (heavy line)
is roughly linear, suggesting that the rate of inereaae of enzyme
aetivities is proportional to the time of pre-incubation. On the other
hand, the eurve for eorticoaterone (dotted line) tenda to level off,
indicating that the rate of increaae of enzyme aetivities tends to de
cresse slightly with time of pre-incubation.
The author cannot explain this diserepancy in the two carves
except for apeeulating that the relatively high yield of corticoater
one in the mediua might inhibit further formation. Birmingham and
Kurlents (123) have ahown that cortieoids liberated, or cortisol and
corticoaterone added into the medium have an inhibitory effect on
further corticoid production by the rat quartered adrenals.

-51-
FIGURE 4
Effect of Tüne on Pre-incubation
-------Corticosterone
----C"ortisol

-• 15 c: 0 ...... o .. .,. ... • .,..,. .... .,..,. ,.... • _,.till' 0 ,o -0 u ,, • - ,, .... ·-... .. 10
,, 11.0
.. 0 ,, 0 u ,,
c.J - ,o -. , ·~·o.a ., , . .. , • s:. , .. , 0.6 .s:.
rtl ,
-~ ,
' 5 , ' c: , c: 0
, 0.4 ·- , 0 • , ·-.. 0~
• .. • 0.2 • > > c: c 0 • 0 u 0 0.0 u ~ 2 20 40 60
~ • Ti me of pre- incubation .
minutes • an

-52-
The low yield of cortisol in this expert.ent ia not typical of
the PIM froa the bovine adrenal cortex, but bas been encountered oc-
casionally, suggesting the instability of probably the 17a-bydroxylase.
2. Effect of Teœperature on Pre-incubation
After establishing that the enzymes or particles containing the
enzymes vere released into the media froœ the tissue during the period
of pre-incubation, the author thought it vould be of interest to know
if this 1release' or 'leakage' of enzymes could be prevented by low
temperature pre-incubation, since it is a general practice that tissues
are kept cold before the experimente in order to prevent or retard the
process of degeneration. For this purpose, separated glomerulosa (Bxpt.
A) or whole cortical slices (Bxpt. B) of the bovine adrenal, or whole
0 c 0 (uncut) rat adrenals (Expt. C) were pre-incubated at 4, 10, or 38 C.
The pre-incubation period for the bovine adrenal slices was 1 br., and
for the rat adrenals vas 1 1/2 br. At the end of the pre-incubation 0
period, the respective PIMwere incubated under usual conditions (38 C,
3 hrs., 951 o2 - 51 co2 ), vith labelled progesterone, HADP, and G-6-P.
The resulta of the three experimenta are shown in Table VI, (Page 53).
Only the major isolated conversion product or products for each experi-
ment vere cbosen as the parameters, since theae would represent over
801 of the over-all conversion. Froœ Table VI 1 under 11 Conversion'
and 'yield' it can be seen that pre-incubation at 12° C increases the
enzyme activities in the PIM about 1 1/2 times, and at 4° C about 3
times as compared vith pre-incubation at 38° c.

e
TABLB VI
Bffect of Temperature on Pre-incubation
Temperature Experiment Compounds oc
(A) Z.G. Corticosterone 38 (Bovine Adrenal) 4
(B) Bntire Cortex Corticosterone 38 (Bovine Adrena1) 12
(C) Wbole Adrenal Cortieosterone 38 Gland (rat) 4
ll~·OH-P 38 4
Yield/3 hrs. f.lmole
0.034 0.099
0.075 0.101
-
*s.A. c.p.m. c.p.m./JJ'}'AOle
2. 7 x lOS 7.8 x 106 1.0 x 105 7.1 x 106
1.9 x 10; 6 2.5 x 106 2.8 x 10 2.8 x 10
o.o -5.5 x 103 -4.4 x 103 -4.4 x to4 -
1 Conversion /3 brs.
18.5 51.4
27.5 40.7
o.o 0.86
0.69 6.90
Bach incubation mixture contained: Progesterone-4-14c: Bxpt. A - (0.181 f.Lmole; 8.0 x 106 c.p.m./~le
Bxpt. B - (0.170 f.llll01e; 4.1 x 106 c.p.m./J.1Dl0le
Bxpt. C- (0.170 J.1Dl0le; 3.7 x 106 c.p.m./f.lmole
PIM from the rat adrenal waa prepared by pre-incubating 400 mg who1e uncut adrenal with 15 ml buffer for 1 1/2 hours. PIM froœ bovine adrenal: saœe as in Table Il.
*s.A. • specifie activities.
e
1 1,1'1 w •

-54-
Of some interest is the observation that very little enzyme
activities were detected in the PIM from the whole (uncut) rat adrenal
0 gland pre-incubated at 38 C (llp-OB-progesterone 0.691), whereas in
the PIM pre-incubated at 4° C the enzyme activities were increased
more than 10 times,with llp-oB-progesterone being the predominant con-
version product (6.91). Bowever, the yields in these cases vere too
low for an accurate ultraviolet light absorption measurement.
A comparison of the specifie activities of the isolated compound&
shows that the products obtained from the law-temperature pre-incubated
PIM were not more diluted than the corresponding ones from the 3aP c-
preincubated PIM. This indicates that lov-temerpature pre-incubation
does not st~ulate the release of the endogenous steroid precursor which
might contribute to the increased prodution of the isolated compound&.
Tbus it can be concluded that law-temperature pre-incubation in-
creases the enzyme activities in the PIM. These findings are similar
to the observation that freezing the adrenal tissue or homogeaates prior
to incubation enhances the enzyme activities (95, 96, 103, 120). Recently
it has been reported (124) that the hydroxylase and dehydrogenase acti
vities in tissue rapidly frozen to - 160° C are markedly increased,
even though the cytochrome-C-oxidase activity is decreased.
The mechanism by which extremely low temperature apparant1y sti-
mu1ates certain enzyme activities is not clear. It is known that freez-
ing causes damages to cella (124). However, Pincus has mentioned in
'Discussion• in the paper by Haines (120) that freezing is one of the
beat ways to preserve the mitochondrie or tissue. In the case of the

-55-
0 PIM, since pre-incubation of the tissue was never done below 4 C,
destruction of celle was probably not likely. Prevention of enzymes
from destruction may explain in parts the apparent increase in enzyme
activities.
SECTION C
The final set of incubation was designed to compare the capacity
of the adrenal cortical tissue, and the pre-incubation medium to meta-
bolize labelled progesterone under. the stated conditions. Tissue
slices (1.5 g) were pre-incubated as before for one hour with 15 ml
buffer to prepare the PIM. The pre-incubated tissue (1.5 g in 15 ml
fresh buffer) and the PIMwere then separately incubated with labelled
progesterone (0.214 pmole or 67.2 ~g) for three hours in the presence
or absence of NADP and G-6-P. The resulta of a duplicate incubation
of each type will be shown in Tables VIla and VIIb on Pages 58 and 59
respectively. Table VIla shows the conversion of the added labelled
progesterone to the various fractions of interest to us. Table VIIb
deals with the total production of steroids, that is, the conversion
of the added substrate as well as the endogenous production. A few
points should be clarified before discussing the resulta: 1) The
values obtained from the tissue incubation represent the sua of the
steroids released into the medium and those extracted from the tissue
itself after incubation. 2) The total recovery of the radioactivity
from the tissue incubation (55.1 x 104 c.p.œ.) was slightly less than
that from the PIM (66.1 x to4 c.p.m.). Therefore, the per cent con-
version was caleulated on the respective total reeoveries. 3) The

•
•
-56-
specifie activities of the conversion proâucts in the PIMwere cal
culateâ after correction for the secretory products into the medium
âuring pre-incubation.
Results: First let us compare the conversion of aââed labelled
progesterone by the PIM (alvays fortified vith NADP and G-6-P) with
that by the tissue ineubated vithout the additives (Table VIla). The
conversion to corticosterone, compound S, and !~-OB-progesterone
(mostly) by the PIMwaa either equal to or superior to that by the
tissue. Bowever, the accumulation of abnoraally large amounta of
tentative 17a-ou-progesterone is not typical of the PIM. The usuel
major end-products are corttcosterone and cortisol. The reason for the
abnormality is not known, and tnctêentally would explain the low yield
in corticosterone and cortisol as coapared to the previous experimenta.
The conversion to cortisol and cortisone by the PIM was much lower
that that by the tissue. Aaain, this may not be typical. There vas
no detectâble conversion to aldosterone and 18-0B-eorticosterone (18-0B-8),
and this is the outstandina qualitative difference between the tissue
and the PIM. When the tissue vas fortifieâ vith HADP and G-6-P the
couversion of labelleâ progesterone to most of the fractions was in
ereased, but not very considerably. This may be due to exhaustion of
the added precursor. The conversion to tentative aldosterone and
18-0B-B was either not affected or even decreased by NADP and G-6-P.
Generally, the aâdition of NADP and G-6-P to the tissue incubation
seems to affect more (in these experimenta) the endogenous productions
than the utilisation of the adâed labelled progesterone. This can be

-57-
seen (Table VIIb) from the fall in the specifie activities of the com-
pounds listed. Again this may be due to addition of insufficient
labelled precursor~ because the tissue has a great capacity to convert
added progesterone to various fractions including many unidentified
regions. For example, the fraction in the region of corticosterone
in the Bush B5 system from tissue incubates contains at least 401 of
unidentified materiels which are not included in the Tables.
On the whole, the isolated products from the tissue incubates had
lower specifie activities than the corresponding ones from the PIM.
This is due to dilution by the endogenous production. In fact, the
specifie activities of the isolated products in the PIM were either
approaching or equal to that of the added precursor (310 c.p.m. x 104/
~mole) within experimental errors, indicating that the bulk of the
products were derived from the labelled substrate. Regarding total
conversion, the PIM (in these experimenta) had about 40-451 of that
of the tissue. This is significant when one considera that the PIM
contains only a minute fraction of the enzymes in the tissue. The
relatively great capacity of the PIM to utilize the added labelled
substrate is at least partly due to laek of competition from endog-
enous steroid precursors.
- e
TABLE VIla
Conversion of Labelled Progesterone by Bovine Adrenal Cortical Tissue and PIM
Tissue Slices PIM No Additive NA'DP + G•6•P NADP + G-6-P
Coœ.p()undf c.p.m.. x 10 3 1 of total c.p.m.. x 10 3 '%. of total ..Ë!!•m.• x 10 3 1 of total
Compound B 32.0 5.80 43.6 7.91 31.3 4. 73 42.0 7.60 51.5 9.34 32.2 4.89
Cortisol 27.3 4.95 52.3 9.49 8.6 1.3 42.6 7.70 58.1 10.54 9.5 1.4
Cortisone 59.5 10.79 48.7 8.88 0.87 0.13 37.8 6.86 28.9 5.25 1.5 0.3
Compound S s.9 1.07 7.1 1.29 20.9 3.2 ' VI co 9.5 1.73 19.6 3.55 22.7 3.4 1
Aldosterone 4.3 o. 78 10.4 1.88 o.o o.o 4.4 0.80 4.8 0.85 o.o o.o
18-0H·B 30.9 5.60 26.1 4. 73 o.o o.o 43.9 7.79 22.1 4.03 o.o o.o
Mixture of 170MOB-prog. 21.5 3.90 19.8 3.55 104.0 15.74 and DOC 21.4 3.80 23.9 4.32
Unconverted Progesterone 13.1 2.37 11.9 2.15 397.0 60.00 6.0 1.09 o.o o.oo

e e
TABLB Vllb
Conversion of Labelled Progesterone by Bovine Adrenal Cortical Tissue and PIM
Tissue S11cea PIM No Additive NADP + G·6-P NADP + G-6-P
c.p,a, x 104 c,p.m. x 10 4 ~···x 10
4
Co!!pounds l,l!!!Ole /eo1e ymole /tele mte /».mD1e
18-0B.-P 0.018 172 0.018 145 - .. 0.037 119 0.025 88 -
Cortisol 0.0127 210 0.028 187 0.0028 307 0.0745 57 0.108 54 0.0030 316
Corticosterone 0.012 250 0.017 256 o.o1 313 0.023 183 0,028 184 o.o1 322 • VI
\D
17a-OB-Progesterone - 0.-03 287 • - - -0,03 265

-60-
SECTION D
Characterization of the major Radioactive Conversion Products from
Labelled Subatrates by the Bovine or Rat Adrenal PIM
The methode usee for the partial characterization of the radio
active conversion products have been fully described in 'Methode.•
In all cases, the isolated products were first purified by aoing
through two appropriate paper chromatographie syatema before the
tests were begun. Insufficient materia1s prevented the use of in
frared spectroscopie etudies of these products.
(1) 118-0H-Progesterone: A major radioactive product from the rat
PIM. The oriainal fraction was obtained from fraction X in
the LPG systeœ, Fig. 1, Paae 32.
The criteria for characterization vere:
1. ldentical Mobility with Authentic 118-0H-Progesterone
(a) About 5 pg of the isolated radioactive product were œixed vith
approximately 30 ~~ of the non-radioactive authentic 11~-oB
progesterone (kind1y aiven by Dr. C. J. P. Giroud). ldentical
mobi1ity was established with the authentic compound in the
two paper chromatographie systems, Bush A (S hrs.), and LPG
(72 hrs.), shown in Columns A and B respectively of Figure 5,
(Pase 62). Radioautographs are on the right and ultraviolet
photocopies on the left.
(b) The materiel eluted from the last mentioned paper (LPG) vas
subjected to selective oxidation of the llJ-hydroxyl group (115).

-61-
ldentical mobility with the oxidation derivative of the authentic
compound waa again established in the same two systems, Bush A
(3 hrs.), and LPG (31 hrs.), shawn in Columna C and D respectively
of Figure S. The tracinga on the left repreaent the radioactivity
&canning. The correaponding ultraviolet photocopies are in the
middle. The standard strip (StD) of Column c shows authentic 11-
keto-progesterone. The ultraviolet light absorption areas at the
tips of Column C represent the solvent-fronts. The standard atrip
(STD) of Column D shows authentic ll,·OH-progesterone and 11-keto
progesterone in decreasing order of polarity.
2. Constant specifie Activity after Paper Chromatoseaphy
Specifie activity was not tested in the above two systems before
oxidation because of lack of sufficient authentic materiel to set up
the standard curve at that time. However, constant specifie activity
was tested and established in the above mentioned systems after oxi
dation, as will be indicated in Table VIII, (Page 63).
3. Constant specifie Activiti after repeated Crystallization with the
Authentic Compound
Approximately 10 mg of non-radioactive authentic llp-OH-progester
one (kindly given by Dr. W. Klyne) were mixed vith minute amounts of
the isolated radioactive product. The 'mixture' waa aubjected to re
peated crystallization. Resulta are shown in Table IX, (Page 63).
4. Other Properties of the lsolated Product
(i) Absorbed ultraviolet light maximally at 241 ~ indicating

• ~
-62-
~ . • }
. •
•
·"'

... •. . .
!i"1 ~ ~ _,-, ;-_ ~ -------+---H -
<J
-+-Ji -: '" ----' - -,..--+- -+--j -+ 1 j - -~ .- -- - f-+--' 0
~~-\- -1: :- t-_.: 1 ·- x 3
~-1 L - -t ~ ~ ~ 1 --~~~ \ +- Il - -+-J - ~"1 t t
~~ t~==t &
t _j ~ f-+ --4 -- - --.--+--!--+-. . ~ s s 0 0 ,_o -~-r-~ - - · -l-- ... -~+-+- T-
s s . . . ~ , o 0 0 0
-~ r- ,..., " -+--~~ . " --t t- -t · r-L I 1 ~ ~ ~ 0 0 0 1 .
'- . .
<(

-63-
T.ABLI VIII
Characterization of tbe.Cr03 Oxidation Product of llp-oB-Progesterone
System
Bush A
LPG
c.p.m.
29600
22500
TABLE IX
)18·
27.50
20.75
1076
1084
Crysta1lization of llp-oH-Progesterone
c.p.m./mg
Crystallization Sol vents Crysta1s Mother Liquors
Pirst Acetone-methanol 11.98 x 10 3 8.42 x 10 3
Second Acetone-benzene u.ss x 103 11.95 x 103
Third Acetone-ethanol 12.22 x 103 11.95 x 103

-64-
the presence of a 6 4-3-keto struction.
(ii) Could not be acetylated by the method (114) selective
for the 21-hydroxyl group, thus auggesting lack of this
function group.
(iii) Did not reduce tetrazolium chloride, indicating the lack
of an a-ketol structure in the aide-chain.
(iv) Did not react with the Porter-Silber reagent, indicating
the lack of a dihydroxy-acetone aide-chain.
All the above evidence strongly indicates that the steroid in
question is 11~-oB-progesterone, although a co.plete characterization,
including an infrared study, was lacking.
(II) Cortisol and Corticosterone: As major conversion products iso
lated from the bovine adrenal cortical PIM. Both tentative com•
pound& were initially isolated in the Bush B5 system. Chracteri
zation includea (a) establishment of identical mobility, as well
as attainment of constant specifie activity with the respective
authentic non-radioactive carriers in the appropriate paper chro
matographie systems (as will be indicated in Tables X and XI)
before and after acetylation, and (b) tests of functional groups,
including ultraviolet light absorption, reduction of tetrazolium
chloride and Porter-Silber reaction. The resulta are tabulated
in Tables X and XI on Pages 66 and 67 respectively.
(III) Tentative Corticosterone iaolated from the lat Adrenal PIM, and
Froges ter one from the Bovine Adrenal PIM: These compounds were

-65-
charaeterized lesa vigorously, i.e. without the measurement of
specifie aetivity or aeetylation.
Tentative corticosterone from the rat adrenal PIM was found:
(a) to have identieal mobility with the authentic compound in the
systems of Bush B5, TBG, and E2B;
(b) to reduce tetrazolium chloride but to have no reaction with
the Porter-Silber reagent, and
(c) to absorb ultraviolet light maximally at 240 ~·
Tentative progesterone as a conversion produet from labelled
pregnenolone was shown:
(a) to have identieal mobility with the authentic compound in the
LPG system;
(b) to have no reaction with both tetrazolium chloride and Porter
Silber reagent, and
(c) to absorb ultraviolet light maximally at 240 ~·

-66-
TABLE X
Characterization of Cortisol
(a) Paper Chromatographie Systems
Chemical Treatment System c.p.m../v.s
None
None
Acety1ation
* Max. u.v. ab.
240 ~
~~
Bush C
T.P.G.
(b) Other Tests
Porter-Silber Reaction
+
1.01 x 103
1.00 x 103
1.00 x 103
Reduction of Tetrazolium.
+
* The m.aterial in ethanol waa m.easured in the Beckman DU
Spectrophotom.eter against its blank solution. The wave·
length range waa 230•260 op. (See also page 22).

-67-
TABLE XI
Characterization of Corticosterone
(a) Paper Chromatographie Systems
Chemical Treatment System
None
None
Acetylation
Max. u.v. ab.
240 ~
T.B.G.
(b) Other Tests
Porter-s il ber Reaction
2.42 x 103
2.78 x 103
2. 77 x 103
Reduction of Tetrazolium
+

-68-
IV. GENERAL DISCUSSION
A short discussion bas usually been given at the end of each
set of experimenta. ln this section, we intend to diseuse only those
aspects of the expertœents which have some general interest or im-
portance.
lt is not the first t1me that enzyme activities have been de
tected in the pre-incubation media from adrenal tissue incubation.
SchBnbaum, Davidson, and Casselman {104) have earlier reported the
detection of NADP•dependent dehydrogenase activities in the incubation
and the pre-incubation media in which rat quartered adrenals have been
incubated. The present study not only confirma the observation of
the above mentioned authors, but provides further evidence for the
presence of a number of other enzyme activities in the PIM. These
enzyme activities are those of;
(i) The ~?-3p-hydroxysteroid dehydrogenase, and
(ii) the llp, 17a, and 21-hydroxylase.
The capacity of these enzyme activities will be appreciated by mention
ing that as much as 50-601 of the added labelled pregnenolone (50 ~g)
or progesterone (50-60 ~g) may be metabolized within three hours by the
PIM in the presence of the appropriate co-factors, (NAD for isomeri
zation of ~?-3p-ol to b~-3-keto; NADP and G-6-P for hydroxylation
reactions). This loss of considerable enzyme activities to the pre
incubation mediumwould explain at ·least partly the observation first
made by Saffran and Bayliss (80) that the adrenal tissue produces lesa

-69-
corticoids after pre-incubation. At the present time it is not cer
tain whether these enzymes in the PIM are contained in the sub-cellular
particles or exist as the soluble fraction in the buffer medium.
In the beginning of this writing, a preliminary experbaent was
done in which the PIMwas subjected to ultracentrifugation at 105,000
x g for 1 hour, and the sediments and supernatant were tested separately
for enzyme activities. Very little activities were found in the super
natant after this treatment. However, the activities in the sedbaents
were not more than 151 of the control, i.e. PIM with no ultracentrifu
gation. The possibility exista that some parts of the enzyme systems
might have been sedimented, and other parts have reaained in the super
natant and that both fractions are necessary for full enzyme activities.
However, much further work is required to clarify this point.
Of some interest is the consistent failure to observe any conversion
of added labelled progesterone to radioactive aldosterone in the pre
sence of NADP and G-6-P. Concoaitantly, the conversion to the fraction
in the region of 18-oB-B was either detected in very minute amounts
(less than 1%) or not detected at all. These findings may indicate
that 18-hydroxylation, one of the presumed final steps in the bio
synthesis of aldosterone, may not be NADPH-dependent. This view is
supported by the observation tbat the addition of HADP and G-6-P ~
vitro did not increase the conversion of added labelled progesterone
to tentative aldosterone or 18-0B-B by the tissue slices. (See Table
VIIb, Page 59). It also did not st~ulate the endogenous production of
aldosterone, or 18-0B-B. In support of this view is also the general

-70-
observation that aldosterone biosyntheais appeara to be independent
of the action of ACTR. This would aeem logical if one accepta the
theory of Haynes (94), wbich states, in essence, that the action of
ACTH is to increase the availability of HADPH.
Alternatively, it is also possible that the 18-hydroxylase and the
18-hydroxysteroid dehydrogenase (1) are very labile, and that their
destruction in the PIM may be the cause of the failure of aldosterone
foraation froa the added precursor. For instance, an enzyme molecule
containing an SB functional group is very susceptible to oxidation unless
properly protected. All this should provide a good opportunity for
further etudies. Not observed is also the'20,22-desmolase'activity
which splits the cholesterol molecule between carbone 20 and 22 to
form the C-21 steroids. However, evidence is obtained only from one
incubation, and the negative finding may be attributed to the relatively
law specifie activity of the labelled cholesterol used (7.8 x 106 c.p.m.
per ~le), although recovery of the radioactivity of unconverted cho
lesterol as compared to the control, does not support this view. Lately,
Kowal, Forchielli, and Dorfman (64, 65) have observed no evidence that
acetone powder preparations of the adrenal cortex can utilize labelled
cholesterol in the presence of NAD or NADP, although tbe same preparations
can readily metabolize pregnenolone under similar conditions. Bowever,
without further experimental evidence it is not beneficiai to speculate upon
the inability of the PIM to utilize cholesterol. The accuaulation of
llp-oH-progeaterone as a radioactive conversion product from labelled
progesterone incubated with the PIM froœ the rat quartered adrenals

• -71-
was rather unexpected and interesting. Until very recently~ this
intermediate metabolite bas not been found in rat adrenal tissue in
cubations. However, Kittinger (51) bas been able to detect this
compound in the rat adrenal at very small concentration, with the
use of gas chromatography. Roberts et al. (47) bas tentatively
detected this steroid from the rat adrena1 hoœogenates incubated with
label1ed progesterone in the presence of 3',5'-cyc1ic AMP. These
authors observed that 11~-hydroxylase activity vas greatly stt.ulated
by the cyclic AMP. In our case, further investigation as to the cause
of the accumulation of this intermediate metabolite under our incu
bation conditions bas not been taken. However, aensitivity of llp
bydroxylase of the rat adrenal to NADPH stimulation is strongly sus
pected, aince the amounts of NADP and G-6-P added to the PIM were con
siderable (12.5 mg and 12.0 -a respectively). Whatever the cause of
the accumulation, the isolation of llp-oH-progesterone froa the rat
adrenal preparation (PI~ supports the views of Kraulis and Birmingham
(48), and of Stachenko and Giroud (31) that llp-OH-progesterone might
be an fœportant intermediate metabolite in the steroid biosyntbesis of
the normal adrenal cortex, on the ground that the rat and the frog,
as well as the bovine adrenal, respectively, can readily metabolize
this compound. The extremely low concentration of this steroid in the
adrenal tissue (Sl) may be due to its rapid turnover rate as it bas been
suggested by Kraulis and Birmingham (48).
Technically, incubation of the pre-incubation aediua aay be a
convenient vay for the study of corticoid biosynthesis. It bas many

-72-
of the functions of the homogenate incubations, with certain additional
advantagea. For instance, the PIM is remarkably free of fat, which
is always present in homogenates, and interferes with the extraction
(foaming) as well as application to paper.
Witb the use of this œethod of incubation, we have been able to
confirœ the work of Samuels et al. (59), and lately the work of Kowal
et al. (64) that NAD is the preferred co-factor for nS.Jp-hydroxy
ateroid dehydrogenase, and that NADP can replace NAD vith about SOl
of the activity. We have alao confirmed indirectly that NADPB is
esaential for certain ateroid hydroxylationa.

-73-
V. SUMMARY
1. A number of enzyme activities were detected in the pre-incubation
medium (PIM) from the rat quartered adrenals or bovine adrenal
cortical slices.
2. NADP was the only exogenous co-factor required for the hydroxyla
tion at the llp, 17a, and 21 positions of the steroid molecule.
G-6-P by itself bad no effect, but greatly stimulated the hydrox
ylating activities when added with NADP; this strongly suggests
the presence of G-6-phosphate dehydrogenase activity.
3. Labelled progesterone was hydroxylated mainly to corticosterone
and cortisol by the bovine adrenal PIM, or to 11~-0B-progesterone
and corticosterone by the rat quartered adrenal PIM. aadioactive
aldosterone was not observed in either case.
4. The ~5-3~-hydroxysteroid dehydrogenase activity was markedly
stimulated by the addition of NAD, and to a lesser degree by NADP.
Transformation of added labelled pregnenolone in the presence of
NAD alone was lLmited to a fraction having the properties of pro
gesterone.
s. Enzyme activities in the PIM increased with time of pre-incubation.
6. Law-temperature pre-incubation enhanced enzyme activities in the
PIM.

BIBLIOGRAPHY
1. Grant, J.K.: Brit. med. Bull., Vol. 18, Ho. 2, 1962. -2. Grant, J.K.: Symp. biochem. Soc., 18, 24, 1960. -3. Bechter, o., and :Pincus, G.: Fhysiol. l.ev., 2!• 459, 1954.
la. Yates, F.E., Urquhart, J. : Fhysio1. Rev., ~' 359, 1962.
4. Feterson, R.B., and Wyngaarden, J.B.: J. Clin. Invest., 2!• 957, 1955.
S. Ayres, P.J., Garrod, O., Tait, S.A.S., Tait, J.r., Walker, G., and Pearlman, V.B., (1957) In: WolsteDho1me, G.E.W., and Millar, B.C.P., ed. Ciba Foundation Co11oquia on Endocrinology. Vol. 11: Hormones in blood, p. 309, Churchill, London.
6. Boltzbauer, M.: J. Physio1., ]l!, 294, 1957.
7. Aires, M.M., ladekaro, M., and Fajer, A.: International Congres& of Hormonal Steroids, Abstr., 213, 1962.
8. Pajer, A., and Vogt, M.: International Congress of Hormonal Steroids, Abstr., 153, 1962.
9. &ace, G.J., and Wu, B.K.: Gen. and Com. lndocrinol., !, 199, 1964.
10. Bush, I.l.: J. Physiol., l!î• 12P, 1951.
11. Vogt, J.: J. Physiol., 130, 601, 1955.
12. Giroud, C.J.F., Stachenko, J., and Venning, I.H.: Proc. Soc. Bxptl. Biol. Med., ~~ p. 154, and p. 855, 1956.
13. Ward, F.G., and Birminghaa, M.K.: Biochem. J., 76, 269, 1960.
14. Reif, A.l., and Longwe11, B.B.: Bndocrino1ogy, ~~ 573, 1958.
15. Cassner, F.X., Nelson, D.B., Reich, B., Rapala, R.T., and Saœuels, L.T.: Proc. Soc. Bxptl. Biol·- Med., Jl., 829, 1951.
16. Hofmann, :r.G.: Bndocrino1ogy, 12• 712, 1956.
17. Southcott, C.M., Bandy, B.!., Newsoa, S.E., and Darrach, M.: Can. J. Biochem. Physiol., 1!• 913, 1956.

18. Biraingbaœ, M.K., and Ward, P.J.: J. Biol. Cbem., 236, 1661, 1961.
19. Peron, F.G.: Endocrino1ogy, !!• 39, 1961.
20. Peron, P.G.: Prograa of the 43rd Meeting of the Endocrine Society, Abstr., 82, 1961.
21. Sandor, T., Lanthier, A.: Acta Endocrinol., ~~ 355, 1963.
22. Simpson, S.A., Tait, J.F., and Bush, I.E.: Lancet, l• 226, 1952.
23. Deane, H.W., and Greep, &.o.: Am. J. Anat., l!• 117, 1946.
24. Deane, H.W., Shaw, J.H., and Greep, &.o.: 133, 1948.
Endocrinology, 43, .....
25. Ayres, P.J., Gould, &.P., Simpson, s.A.s., and Tait, J.F.: Biochea. J., !!• 19P, 1956.
26. Giroud, C.J.P., Stacheuko, J., and Venning, B.B.: Proc. Soc. Exptl. Biol. Med., !a• 154, 1956.
27. Giroud, C.J.P., Stachenko, J., and Pi1etta, P.: Intern. Symp. A1dos., Geneva, 1957. Little Brown and Co., Boston, 1958, p. 56.
28. Giroud, c.J.P., Stachenko, J., and Piletta, P.: Symposium on Aldosterone, Churchill Ltd., London, 56, 1958.
29. Stacbenko, J., and Giroud, C.J.P.: Bndocrino1ogy, ~~ 743, 1959.
30. Stachenko, J., and Giroud, C.J.P.: Bndocrino1ogy, ~, 730, 1959.
31. Stachenko, J., and Giroud, C.J.P.: Can. J. Biochem., ~. 1777, 1964.
32. Ayres, P.J., Garrod, O., Tait, S.A.S., and Tait, J.F., (1958) In: Muller, A.F., and O'Connor, C.M., ed. An International Symposium on Aldosterone, p. 143, Churchill, London.
33. Plager, J.E., and Samoels, L.T.: Federation Proc., 11, 383, 1952.
34. Plager, J.E., and Samuels, L.T.: Arch. Biocheœ. Biophy., ~. 477, 1953.
35. Beard, &.D.H., 0 1Donnel1, &.J.V., Peron, F.G., Saffran, J.c. So1omon, s.s., Thompson, L.M., Willoughby, H., and Yates, C.H.: &ecent Progr. Hormone Res., ~~ 383, 1954.
36. Eisenstein, A.B.: Proc. Soc. Expt. Biol. Med., ~, 657, 1956.

37. Hofmann, F.G.: Bndocrinology, 60, 382, 1957.
38. Brownell, K.A., Lee, S.L., Beek, R.R., Block, P.K.:
39.
~.
Bndocrinology, 1!• 167, 1963.
Birmingham, M.K., and Kurlents, B.: ll· 510, 1959.
Ward, P.J., and Birminghaœ, M.K.:
Can. J. Biochem. Physiol.,
Biochem. J., 76, 269, 1960 • ..... 41. Laplante, c., Giroud, C.J.P., and Stachenko, J.: Bndocrinology,
]1, 825, 1964.
42. Ward, P.J., and Birmingham, M.K.: Acta Endocr., 12• 110, 1962.
43. Levy, H., Jeanloz, R.W., Jacobsen, R.P., Hechter, o., Schenker, V., and Pincus, G.J.: J. Biol. Chem., 111, 867, 1954.
44. Bichhorn, J., and Bechter, o.: Proc. Soc. lxpt. Biol. (N.Y.), 97, 614, 1958.
45. Brownie, A.c., Grant, J.K., and Davidson, D.W.: Biochem. J., 58, 218, 1954.
46. Giroud, C.J.P., Stachenko, J., and Filetta, P. In: Muller, A. r., and O'Connor, C.M., ed. An International Symposium on Aldosterone. Little Brown and Co., Boston, 1958.
47. Roberts, s., Crearge, J.E., and Fowler, D.D.: Nature, ~' 761, 1964.
48. Kraulis, I., and Birmingham, M.K.: Acta Endocr., il• 79, 1964.
49. Oertel, G.W., and Eik-Nes, K.B.: Bndocrinology, 1, 39, 1962.
50. Sandor, T., Lamoureux, J., and Lanthier, A.: Endocrinology, 73, 269, 1964.
51. Kittinger, G.W.: Steroids, l• 21, 1964.
52. Bechter, o., Zaffaroni, A., Jacobsen, R.P., Levy, H., Jeanloz, R.W., Schenker, v., and Pincus, G.: Recent Progr. Hormone Res., !· 215, 1951.
53. Pincus, G., and Romanoff, E.B., In: Wolstenholme, G.B.W., and Cameron, M.P., ed. Ciba Foundation Colloquia on Bndocrino1ogy. Vol., 8: The human adrenal cortex, p. 97, Churchill, London.
54. Lombardo, M.E., and Hudson, P.B.: Endocrino1ogy, ~' 417, 1959.

55. Lombardo, M.B., R.oitman, B., and Hudson, P.B.: J. Clin. Bndocrin., ,!!. 1283, 1956.
56. Soloœon, S., Levitan, P., and Lieberman, s.: R.ev. Can. Biol., ll· 282, 1956.
57. Dorfman, R..I.: Cancer Res., !Z• 535, 1957.
58. Staple, 1., Lynn, w.s., jr., and Gurin, s.: J. Biol. Cheœ., .2.!· 845, 1956.
59. Samuels, L.T., Helmreich, K.L., Lasater, M.B., and Reich, H.: Science, 113, 490, 1951.
60. Talalay, P., and Wang, v.s.: Bioch~. Biophys. Acta, ]!, 300, 1955.
61. Aoshfaa, Y., Kochakian, C.D., and Jadrijevic, D.: Bndocrinology, J!!, 521, 1964.
62. Byer, K.F., and Saauels, L.T.: J. Biol. Chem., ~. 69, 1956.
63. Halkerston, I.D.K., Bichhorn, J., and Hechter, o.: J. Biol. Cheœ., .ll§., 374, 1961.
64. Kowa1, J., Forchie111, 1., and Dorfman, R.I.: Steroids, 1• 531, 1964.
65. Kowa1, J., Forchie1li, B., and Dorfman, R.I.: Steroids, !, 77, 1964.
66. Baille, A.H., Caaeron, B.H.D., Griffith&, K., and Hart, D. Mck.: J. Bndocrin., ]!, 227, 1965.
67. Levy, H., Deane, H.W., and R.ubin, B.C.: Bndocrinology, ~. 932, 1959.
68. Dawson, I.M.P., Pryse-Daviea, J., and Snape, I.M.: Biochem. J., Eh 16P, 1961.
69. Sweat, M.L.: J. Am. Chem. Soc., 73, 4056, 1951.
70. Hayano, M., and Dorfman, R.I.: J. Biol. Chem., l2l• 175, 1951.
71. Grant, J.K.: Biochem. J., ~' 559, 1956.
72. Tomkins, G.M., Michael, P.J., and Curran, J.F.: Bioch~. Biophys. Acta, l!• 655, 1957.
73. Sharma, D.C., Forchielli, B., and Dorfman, R..I.: J. Biol. Chem., 273, 1495, 1962. -

74. Tomkins, G.K., Curran, J.F., and Michael, P.J.: Bioch~. Biophys. Acta: j!, 449, 1958.
75. Sweat, K.L. (1962) In: Proceedings of the Vth International Congres& of Biochemistry, Koscow, August 10-16, 1961. (Abstracts) Pergamon Press, Oxford.
76. Plager, J.B., and Saœuels, L.T.: Federation Proc., j!, 383, 1952.
77. Ryan, K.J., and Bngel, L.L.: J. Biol. Chem., ~. 103, 1957.
78. Hofmann, F.G.: Bioch~. et Biophys. Acta, lZ• 566, 1960.
79. Bayano, K., and Dorfman, R.I.: J. Biol. Chem., 1Q!, 175, 1953.
80. Saffran, K., and Bayliss, K.J.: Bndocrino1ogy, ~. 140, 1953.
81. Hayano, K., and Dorfman, R.I.: J. Biol. Chem., 211, 227, 1954. -82. Bayano, ~. Lindberg, H.c., Dorfman, R.I., Bancock, J.B.B., and
Doering, W. von B.: Arch. Biochem. Biophys., 12, 529, 1955.
83. Blooa, B.M., Hayano, K., Saito, A., Stone, D., and Dorfman, R.I.: Federation Proc., y, 222, 1956.
84. Sweat, M.L., Aldrich, R.A., Bruin, C.B. de, Fowlks, W.L., Heiselt, L.R., and Mason, H. s.: Federation Proe., ll• 367, 1956.
85. Bayano, M., Saito, A., Stone,D., and Dorfman, R.I.: Bioch~.
86.
87.
Biopbys. Acta, 1!, 381, 1956.
Ryan, K.J., and Bnge1, L.L.: J. Biol. Cbem., 225, 103, 1957. -Hayano, M., Dorfman, R.I.: J. Biol. Chem., lQ!, 175, 1953.
88. Hayano. M •• and Dorfman, R.I.: J. Biol. Cbem •• 211, 227, 1954.
89. Bayano, M., Dorfman, R.I., and Rosenberg, B.: Federation Proc., ll· 290, 1955.
90. Sweat, M.L., and Lipeomb, M.D.: J. Am. Cbea. Soc., ill• 5185, 1955.
91. Lynn, w.s., Jr., and Brown, R.H.: J. Biol. Cbea., ~. 1005, 1958.
92. Lynn, w.s., Jr., and Brown, &.H.: J. Biol. Cbem., ~~ 1015, 1958.
93. Plager, J.B., and Samuels, L.T.: J. Biol. Chea., lll• 21, 1954.

94. Haynes, a.c., Jr., and Berthet, L.: J. Biol. Chem., ~. 115, 1957.
95. Koritz, S.B., Peron, F.G.: J. Biol. Chem., 230, 343, 1958. ........ 96. Koritz, S.B., and Peron, F.G.: J. Biol. Chem., ~~ 1959.
97. Constantopoulos, G., and Tchen, T.T.: J. Biol. Chem., 236, 65. 1961.
98. Glock, G.I., and McLean, P.: Biochem. J., 2!• 171, 1954.
99. Kelly, T.L., Nielson, B.D., Johnson, R.B., and Vesting, c.e.: J. Biol. Chem., ~~ 545, 1955.
100. McKerna, K.W.: Biochim. Biophys. Acta, 1!• 710, 1963.
101. Gordon, J.J.: Biochem. J., il• 701, 1957.
102. Hayano, M. In: Oxygenaaes, edited by Hayaishi, O., Academie Press, New York and London, 1962.
102a. Harding, B., and Nelson, D.: of the Forty-Seventh Meeting.
The Endocrine Society. No. 41, June, 1965.
Program
103. Hall, P.F., and KDritz, S.B.: lndocriuo1ogy, ]l, 135, 1964.
104. Schünbaum, E., Davidson, M., Large, R.E., Casselman, W.G.: Cau. J. Biochem. Physiol., ll• 1209, 1959.
105. Umbreit, w.w., Burris, &.H., and Stauffer, J.F.: Manometric Techniques.
106. Stadie, w.c., and Riggs, B.C.: J. Biol. Chem., 154, 687, 1944. ........ 107. Bush, I.E.: Biochem. J., 22. 370, 1952.
108. Zaffaroni, A., and Burton, R.B.: J. Biol. Chem., ~. 749, 1951.
109. Eber1ein, W.&., and Bongiovanni, A.M.: Arch. liochem., 12• 90, 1955.
110. Nowaczyuaki, W.J., and Koiw, I.J.: 1957.
Lab. Clin. Med., 49, 815, ......
111. Savard, K.: J. Biol. Chem., ~~ 457, 1953.
112. Zaffaroni, A.: Recent Progr. Horm. Res., !• 51, 1953.

113. Allen, W.M.: J. Clin. Endocrino1ogy, 10, 71, 1950.
114. Bush, I.E.: The Chromatography of Steroids, Pergamon Press, p. 358.
115. Bush, I.E.: The Chromatography of Steroids, Pergamon Press, p. 363.
116. Klyne, W.: The Chemistry of the Steroids, p. 166, 1960. (Methuen and Co. Ltd.)
117. Porter, c.e., and Si1ber, R.H.: J. Biol. Chem., 185, 203, 1950.
118. Silber, R.H., and Porter, c.e.: J. Biol. Chem., l!Q, 923, 1954.
119. Birmingham, M.D., and Ward, P.J.: J. Biol. Chem., ~, 1661, 1961.
120. Haines, W.J.: Recent Progress in Hormone Research, 1, 285, 1952.
121. Lucia, R., Carba1leira, A., and Venning, E.H.: Proc. Can. Fed. Biol. Soc., 1964, p. 43.
122. Carballeira, A., Medhi, A., and Venning, E.H.: Proc. Soc. Exptl. Biol. and Med., 117, 751, 1965.
123. Birmingham, M.K., and Kurlents, E.: Endocrinology, 62, 47, 1958.
124. Saunders, J.F.: Science,~~ 181, 1965.