some characteristics of a phenol-oxidizing pseudomonas' · 5hydroxy...
TRANSCRIPT

Some Characteristics of a Phenol-Oxidizing Pseudomonas'M. K. HAMDY, E. L. SHERRER, C. I. RANDLES, H. H. WEISER AND W. D. SHEETS
Department of Bacteriology and Waste Treatment Laboratory, Engineering Experiment Station, Ohio State University, Columbus, Ohio
Received for publication September 16, 1955
In a previous communication, Sheets et al. (1954)have shown that three phenol-resistant organisms, asporeforming rod, a coccus and a short rod, belongingto the genera Bacillus, Micrococcus and Pseudomonasrespectively, were isolated from a trickling filter, op-erated continuously at 54 C for the treatment of phe-nolic wastes. The Pseudomonas-like rod predominatedover the other two types. A pure culture of this organ-ism was capable of oxidizing phenol at a relatively highrate (Hamdy et al., 1954).
Microorganisms that oxidize such aromatic com-pounds as phenol, m-cresol, and naphthalene have .beenisolated by Kramer and Doetsch (1950), Czekalowskiand Skarzynski (1948), and Gray and Thornton (1928).
Physiological data on phenol-oxidizing bacteria havebeen reported by Egorova (1946), Evans (1947), Stanier(1948), and Sleeper and Stanier (1950). Kilby (1948)postulated the breakdown of phenol as: phenol -÷ cate-chol -- o-benzoquinone -> 1,2,5, trihydroxybenzene ->5 hydroxy o-benzoquinone -- 3-ketoadipic acid -* -suc-cinate + acetate.
Sleeper and Stanier (1950) reported that catechol isan intermediate in the oxidation of phenol by cells ofPseudomonas fluorescens. Stanier et al. (1950) confirmedthat g-ketoadipic acid is an intermediate in the oxida-tion of catechol by P. fluorescens while hydroxyhydro-quinone is highly improbable.An investigation was undertaken in this laboratory
with the aim of securing more information concerningthe characteristics of and the mechanism of phenolbreakdown by an unidentified strain of Pseudomonasisolated from the trickling filter exposed to phenolliquor.
MATERIAL AND METHODS
The culture media, the bacteriological and chemicalmethods employed in this study were the same as thosepreviously described (Hamdy et al., 1954). The organ-ism used in this work was the pseudomonad isolatedfrom a trickling filter by Sheets et al. (1954). Oxygenuptake was measured using conventional manometricmethods. Fernbach culture bottles were used to growthe organism in salt-solution medium2 with phenol while
1 Engineering Experiment Station Bulletin No. 157.2 Salt solution, made of 1.0 g ammonium nitrate, 0.5 g
dipotassium hydrogen phosphate, 0.01 g ferric chloride, 0.1 gsodium chloride, 0.2 g magnesium sulfate per liter of distilledwater.
Florence flasks were used to grow the organism on nu-trient broth with or without phenol. Aeration was ac-complished by bubbling sterile air through the culturesto increase the number of cells. Centrifugation was usedto separate the cells, which were.then washed with dis-tilled water. In order to compare the results of themanomnetric experiments,, approximately equal cell con-centrations, as measured turbidimetrically with a Beck-man model B spectrophotometer at a wave length of500 m,u, were prepared. All manometric data were ob-tained at 46.5 C, as no activity was observed within areasonable length of time at 37 C.
EXPERIMENTAL RESULTS
Morphological and Biochemical Characteristics ofthe Pseudomonad
Cells 0.5 to 0.8 by 1.3 to 3.0 microns, straight, motileby polar flagellum (figure 1), gram negative. These rodsfrequently showed unstained areas, probably vacuola-tions, especially in old cultures. The organism grewwell on ordinary culture media, single, or in pairs whenthe culture was incubated at 37 C or at room tempera-ture, and in very long chains when the culture was in-cubated at 54 C (figures 2 and 3).
The agar colonies. Circular, 3 to 5 mm diameter, yel-low, smooth, glistening, entire, opaque.Agar slant. Filiform, raised, smooth, glistening, un-
dulate; the growth on the slant tends to become darkgray with age.
Nutrient broth. Turbid with heavy sediment.Optimum temperature. It grows well at both 37 and
54 C with the latter appearing to be more favorable. Itgrows slowly at room temperature and very poorly at10 C.
Fermentation. No acid or gas was formed from thefollowing carbohydrates and alcohols: arabinose, glu-cose, fructose, galactose, sucrose, maltose, lactose,starch, inulin, dextrin, glycerol, mannitol and sorbitol.Slight acidity was observed in xylose.
Action on inorganic nitrogen compounds. The organ-ism utilizes ammonium salts and urea when furnishedwith no other source of nitrogen.
Nitrate is not reduced, starch is not hydrolyzed andgelatin is not liquefied. No action was seen on bloodagar and milk was not changed. Catalase, indol andMRVP tests were netative. The organism does not pro-duce hydrogen sulfide. It grows well on citrate medium.The organism is aerobic, attacks phenol, and resists
71
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
M. K. HAMDY ET AL.
*i.X.~ ~ ~ ~ ~ ~~~~. ....:t: ....
FIG. 1. Photomicrograph of Pseudomonas X54 demon- FIG. 3. Photomicrograph of Pseudomonas X54 grown forstrating the polar flagella (X 1800). 24 hours on tryptone glucose extract agar at 54 C (X 1100).
FIG. 2. Photomicrograph of Pseudomonas X54 grown for24 hours on tryptone glucose extract agar at 37 C (X 1100).
high concentrations of it (Hamdy et al., 1954; Sheetset al., 1954).Based on the characteristics listed above, the organ-
ism was identified as a member of the genus Pseudo-monas and was designated Pseudomonas X54. Pseudo-monas X54 shows an unusual temperature optimum forthe genus Pseudomonas. It has a yellow to grayish colorin agar colonies, fails to produce acid or gas from glu-cose and fails to reduce nitrates to nitrites. Theseproperties are not characteristic of Pseudomonas. How-ever, the culture was originally isolated from sewage,where the average temperature was around 22 C, thenafter isolation, the temperature of the trickling filter
was gradually brought to 54 C in the presence of phenol.The temperature optimum of the culture may havebeen influenced by this treatment. In other properties,the morphology, gram stain, motility, aerobiosis, andorigin from water, the culture is typical of the genusPseudomonas.The culture does not correspond to the species of
Pseudomonas listed in Bergey's Manual of DeterminativeBacteriology (Breed et al., 1948). The binomial Pseudo-monas phenolis is suggested for the culture.
The growth of Pseudomonas X54 on other compounds.Fourteen compounds including phenol were tested assole sources of carbon. Washed cells of the organismwere placed in Erlenmeyer flasks containing 200 ml ofsalt solution medium and 2 X 10-3 moles of the testedcompound and the flasks were incubated at 48 C. Table1 represents the results of this experiment indicatingthe vast potentialities of this organism to metabolizecompounds other than phenol as a sole source of carbon.As indicated in table 1, some compounds formed dark-colored complexes which did.not appear until the tem-perature of the flasks was raised to 48 C. Evidentlythis may be due to the highly reactive nature of thehydroxyl group in the ortho and the para positions onthe benzene ring. Kramer and Doetsch (1950) used hy-droquinone and quinone while Sleeper et al. (1950) usedcatechol in their studies on the bacterial oxidation ofaromatic compounds. Yet neither of the aforementionedauthors reported encountering this difficulty, probablydue to the fact that their investigations were done at30 C incubation.
Effect of pH on growth and phenol oxidation. Theeffect of pH on phenol oxidation and on the growth ofthe Pseudomonas X54 in both salt solution medium
72 [VOL.,4
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

PHENOL-OXIDIZING PSEUDOMONAS
Carbon Source
Phenol .. ...........
Catechol ............
Resorcinol ......
Hydroquinone .....
Quinone .............
Pyrogallic acid.....Phloroglucinol ......
m-cresol ............
Cyclohexanol ........
Benzoic acid ......
Phenylacetic acidSalicylic acid........Aniline ..............
Glucose .............
Days Allowing Visible Growth
3 daysA dark color was formed4 daysA dark color was formedA dark color was formed
4 days
Growth after
5 Days
++++
++
+++++
1 day
* ++++ = growth equal to or better than cells grown in2 X 10-3 moles of phenol per liter of salt solution medium;+ = slight growth; - = no growth.
TABLE 2. The effect of pH on the growth and phenoloxidation of Pseudomonas X54
Growth in SaltMedium Solution Medium Growth in Phenol Conc.pHm ofand 250 ppm Nutrient Broth in ppm.t
Phenol
4.6 _* - 1905.0 _ _ 1845.5 ± + 1406.0 + ++ 1116.5 ++ +++ 377.0 +++ ++++ 07.5 ++ ++++ 228.0 + ++ 1608.5 _ i 1809.0 - - 189
*-_ = no growth; i = very slight; ++ = moderate;++++ = heavy growth, after 3 days' incubation.
t Phenol concentration in ppm after 3 days' incubation at54 C in salt solution medium containing 200 ppm phenol.
with phenol and in nutrient broth is shown in table 2.Flasks containing buffered nutrient broth or salt solu-tion medium with 200 ppm phenol were prepared hav-ing initial pH values ranging from 4.5 to 9.0. The flaskswere inoculated with an active culture using 2 per centinoculum of the organism and incubated at 54 C for 3days. At the end of the incubation period phenol analy-ses using Hill and Herndon method (1952) were per-formed on the salt solution medium containing phenol.The results indicated that the optimum pH for boththe growth of the organism and the phenol oxidationranged from 6.5 to 7.5.
Tyrosine as a possible source of carbon and nitrogen.This experiment was conducted to investigate the abil-ity of the Pseudomonas organism to grow on an aromaticamino acid as a sole source of carbon and nitrogen underboth aerobic and anaerobic conditions. Salt solution
TABLE 3. The utilization of tyrosine by PseudomonasX54 in salt solution medium incubated at 54 C
Time of Incubaion Phenol*Oxygen Tension Analysis
1 day 5 days
Aerobic ............... ++++t +++++ NegativeAnaerobic ............ ++ + + Negative
* Phenol analysis was made at the end of every 24 hours'incubation.
t Growth equal to or better than cells grown aerobically in2 X 10- moles of phenol in a liter of salt solution medium.
medium containing 2 X 10-3 moles of tyrosine per literof solution was used. The results shown in table 3 indi-cated that the organism can grow actively under aerobicconditions and very slowly under anaerobic conditions.It was also found that this aromatic amino acid canreplace the ammonium phosphate in the salt solutionmedium as a source of nitrogen. Daily analyses ofphenol in tyrosine salt solution medium failed to detectits presence under either aerobic or anaerobic condi-tions, indicating that phenol is not an end product inthe metabolism of tyrosine by the Pseudomonas strain.A dark compound was observed during the aerobic
oxydation of tyrosine probably due to melanin (Green-berg, 1951).Manometric studies. Warburg manometric techniques
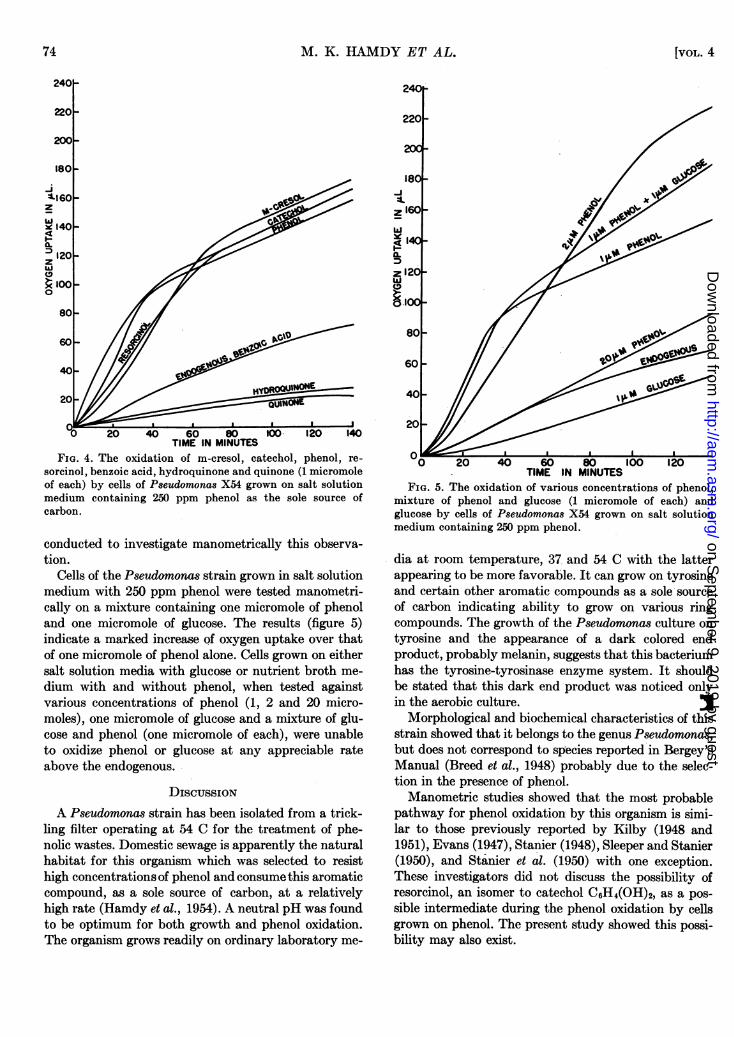
were used at 46.5 C to secure information on the mecha-nism of phenol oxidation by the Pseudomonas culture.Cells grown in 250 ppm phenol in salt solution mediumwere tested against the following compounds using onemicromole of each: m-cresol, catechol, phenol, resorci-nol, benzoic acid hydroquinone and quinone. The results(figure 4) show three different types of activity againstthese various substrates; those having the same rate ofoxygen uptake as phenol, those with a similar rate tothe endogenous, and those with a lower rate of oxygenuptake than the endogenous. The slightly lower activityin resorcinol, by phenol-grown cells, may be due to in-hibition. Catechol showed a more rapid rate of oxygenuptake than phenol which might be explained on thebasis of the toxic effect of phenol. Fogg and Lodge(1945) reported that catechol had a phenol coefficientof 0.5. This toxic effect was more pronounced at a highconcentration of phenol while 2 micromoles of phenolshowed some inhibitions; 20 micromoles of phenol de-creased oxygen uptake to that of the endogenous (fig-ure 5).
Results in a previous report (Hamdy et al., 1954)have established that the presence of lactose inhibitedphenol oxidation by the Pseudomonas culture, probablydue to competition in adaptative enzyme production.Accordingly, it was advisable in a biological treatmentof the phenolic waste, to use media free from carbonsources other than phenol to accomplish phenol oxida-tion more effectively. The following experiments were
TABLE 1. The ability of Pseudomonas X54 to grow onvarious compounds in salt solution medium at 48 C
19561 73
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
M. K. HAMDY ET AL.
-i
zw
;q-z(9
0
20 40 60 80 100 120 140TIME IN MINUTES
FIG. 4. The oxidation of m-cresol, catechol, phenol, re-sorcinol, benzoic acid, hydroquinone and quinone (1 micromoleof each) by cells of Pseudomonas X54 grown on salt solutionmedium containing 250 ppm phenol as the sole source ofcarbon.
conducted to investigate manometrically this observa-tion.
Cells of the Pseudomonas strain grown in salt solutionmedium with 250 ppm phenol were tested manometri-cally on a mixture containing one micromole of phenoland one micromole of glucose. The results (figure 5)indicate a marked increase of oxygen uptake over thatof one micromole of phenol alone. Cells grown on eithersalt solution media with glucose or nutrient broth me-dium with and without phenol, when tested againstvarious concentrations of phenol (1, 2 and 20 micro-moles), one micromole of glucose and a mixture of glu-cose and phenol (one micromole of each), were unableto oxidize phenol or glucose at any appreciable rateabove the endogenous.
DISCUSSIONA Pseudomonas strain has been isolated from a trick-
ling filter operating at 54 C for the treatment of phe-nolic wastes. Domestic sewage is apparently the naturalhabitat for this organism which was selected to resisthigh concentrations of phenol and consume this aromaticcompound, as a sole source of carbon, at a relativelyhigh rate (Hamdy et al., 1954). A neutral pH was foundto be optimum for both growth and phenol oxidation.The organism grows readily on ordinary laboratory me-
TIME IN MINUTESFIG. 5. The oxidation of various concentrations of phenol,
mixture of phenol and glucose (1 micromole of each) andglucose by cells of Pseudomonas X54 grown on salt solutionmedium containing 250 ppm phenol.
dia at room temperature, 37 and 54 C with the latterappearing to be more favorable. It can grow on tyrosineand certain other aromatic compounds as a sole sourceof carbon indicating ability to grow on various ringcompounds. The growth of the Pseudomonas culture ontyrosine and the appearance of a dark colored endproduct, probably melanin, suggests that this bacteriumhas the tyrosine-tyrosinase enzyme system. It shouldbe stated that this dark end product was noticed onlyin the aerobic culture. 3
Morphological and biochemical characteristics of thisstrain showed that it belongs to the genus Pseudomonas,but does not correspond to species reported in Bergey'sManual (Breed et al., 1948) probably due to the selec-tion in the presence of phenol.Manometric studies showed that the most probable
pathway for phenol oxidation by this organism is simi-lar to those previously reported by Kilby (1948 and1951), Evans (1947), Stanier (1948), Sleeper and Stanier(1950), and Stanier et al. (1950) with one exception.These investigators did not discuss the possibility ofresorcinol, an isomer to catechol C6H4(OH)2, as a pos-sible intermediate during the phenol oxidation by cellsgrown on phenol. The present study showed this possi-bility may also exist.
74 [VOL. 4
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

PHENOL-OXIDIZING PSEUDOMONAS
The cells of this organism when grown on glucosesalt solution medium, with and without phenol, failedto take up oxygen at a significant rate when phenol was
added as a substrate, indicating that adaptive enzymes
are responsible for phenol oxidation (Hamdy et al.,1954). Cells grown on salt solution medium containingphenol showed no activity when tested on glucose; a
mixture of phenol and glucose was oxidized at the same
rate as phenol alone. After 35 to 40 minutes, when thephenol had presumably been oxidized, some oxygen
uptake was still occurring.The minimum theoretical amount of oxygen required
to completely oxidize one micromole of phenol is 157microliters. When Warburg experiments were observedfor a long time, the phenol grown cells were able toconsume almost the required amount of oxygen pre-
viously mentioned, for complete phenol oxidation,after subtracting the endogenous, indicating the fissionof the benzene ring of the phenol during the oxidationprocess.
It should be stated that a yellow color was observedin the trickling filter during the phenol oxidation. Thiscolor intermediate compound disappeared shortly aftercomplete oxidation of phenol. No attempts were madeto identify this yellow-colored compound. Kilby (1951)has reported that slightly yellow medium was observedafter inoculation and incubation of the vibrio culturein mineral salt medium containing phenol.
SUMMARY
A phenol-oxidizing bacterium has been isolated andidentified as a strain of Pseudomonas. Morphologicaland biochemical characteristics have been reported. Theability of this culture to grow on and oxidize other aro-
matic compounds was investigated; manometric stud-ies revealed that phenol oxidation by this strain is ac-
complished by adaptive enzymes. We suggest the namePseudomonas phenolis.
REFERENCES
BREED, R. S., MURRAY, E. G. D., AND HITCHENS, A. P. 1948Bergey's Manual of Determinative Bacteriology, 6th ed.The Williams & Wilkins Company, Baltimore.
CZEKALOWSKI, J. W., AND SKARZYNSKI, B. 1948 The break-down of phenols and related compounds by bacteria. J.Gen. Microbiol., 2, 231-38.
EGOROVA, A. A. 1946 Physiological data on phenol-oxidizingthermophilic bacteria. Mikrobiologiya, 15, 467-477.
EVANS, W. C. 1947 Oxidation of phenol and benzoic acid bysome soil bacteria. Biochem. J., 41, 373-382.
FOGG, A. H., AND LODGE, R. M. 1945 The mode of anti-bacterial action of phenols in relation to drug-fastness.Trans. Faraday Soc., 41, 359-365.
GRAY, P. H., AND THORNTON, H. G. 1928 Soil bacteria thatdecompose certain aromatic compounds. Zentr. Bakt.,Parasitenk, Ab., II 73, 74-96.
GREENBERG, D. M. 1951 Amino Acids and Proteins, p. 20.Charles C Thomas, Publisher, Springfield, Illinois.
HAMDY, M. K., SHERRER, E. L., WEISER, H. H., AND SHEETS,W. D. 1954 Microbiological factors in the treatment ofphenol wastes. Appl. Microbiol., 2, 143-148.
HILL, R. H., AND HERNDON, L. K. 1952 Determination ofphenols by the p-nitrosodimethylaniline method. Sewageand Ind. Wastes, 24, 1389-1396.
KILBY, B. A. 1948 The bacterial oxidation of phenol to,8-ketoadipic acid. Biochem. J., 43, V-VI.
KILBY, B. A. 1951 The formation of ,3-ketoadipic acid bybacterial fission of aromatic rings. Biochem. J. 49, 671-674.
KRAMER, N., AND DOETSCH, R. H. 1950 The growth ofphenol-utilizing bacteria on aromatic carbon sources.Arch. Biochem., 26, 401-405.
SHEETS, W. D., HAMDY, M. K., AND WEISER, H. H. 1954Microbiological studies on the treatment of petroleumphenolic wastes. Sewage and Ind. Waste, 26, 862-868.
SLEEPER, B. P., AND STANIER, R. Y. 1950 The bacterialoxidation of aromatic compounds. I. Adaptive patternswith respect to polyphenolic compounds. J. Bacteriol.,59, 117-127.
STANIER, R. Y. 1948 The oxidation of aromatic compoundsby fluorescent Pseudomonads. J. Bacteriol., 55, 447-494.
STANIER, R. Y., SLEEPER, B. P., TSUCHIDA, M., AND MAC-DONALD, D. L. 1950 The bacterial oxidation of aromaticcompounds. III. The enzymatic oxidation of catechol andprotocatechuic acid to ,B-ketoadipic acid. J. Bacteriol.,59, 137-151.
75
on Septem
ber 9, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from