soil moisture variability in a temperate deciduous forest ...rvd/pub/ees_2014.pdf · insights from...
TRANSCRIPT

THEMATIC ISSUE
Soil moisture variability in a temperate deciduous forest:insights from electrical resistivity and throughfall data
Yuteng Ma • Remke L. Van Dam •
Dushmantha H. Jayawickreme
Received: 1 December 2013 / Accepted: 15 May 2014 / Published online: 4 June 2014
� Springer-Verlag Berlin Heidelberg 2014
Abstract In deciduous forests, soil moisture is an important
driver of numerous physical, microbial, and biogeochemical
processes. Therefore, characterizing the interactions between
vegetation and soil moisture is critical to forecast long-term
water resources and ecosystem health. However, these inter-
actions are difficult to measure, both in time and space.
Recent studies have shown the ability of electrical resistivity
imaging (ERI) to characterize the spatial and temporal
dynamics of soil moisture at a range of scales. However, no
study has yet attempted to use ERI to describe spatiotemporal
variability of soil water in relation to vegetation structure and
throughfall. In this study, at a mature forest site in Michigan,
USA, we captured spatial and temporal dynamics of soil
moisture using weekly ERI measurements augmented with
throughfall and soil temperature measurements, and a detailed
vegetation survey for five adjacent quadrats. Our results show
that throughout the growing season, the soil moisture gradu-
ally declined despite strong variations in cumulative monthly
rainfall. This decline was occasionally halted, but not
reversed, during weeks with high precipitation. Spatial vari-
ability of electrical resistivity and soil moisture was not
related to soil temperature differences but showed a strong
correlation with canopy variables.
Keywords Electrical resistivity � Deciduous forests �Forest ecology � Soil moisture � Throughfall
Introduction
The temperate-climate deciduous forests with four distinct
seasons are a common vegetation type in the northern
hemisphere (Hansen et al. 2000; Wen 1999; Keddy 1996).
With world’s population relying on forests for a range of
natural products and ecosystem services, it is critical that
these systems are properly understood and managed
(McElrone et al. 2013). Previous work has shown how
changes in precipitation, temperature, and wind may have
important consequences for the type and density of trees
(e.g., Reed and Desanker 1992; McKenney-Easterling
2000). Moreover, changes in CO2 concentration have been
shown to affect plant water demand and the soil water
balance of temperate forests (Schafer et al. 2002). In this
complex system, soil moisture is both an indicator and a
driver of change. Therefore, understanding the spatial and
temporal dynamics of near-surface soil moisture is a crit-
ical endeavor (Asbjornsen et al. 2011; Peng et al. 2013).
However, monitoring and quantifying soil water fluxes at
relevant spatial scales remains difficult because of the
above- and belowground heterogeneities inherent to forest
ecosystems.
Soil moisture content varies temporally as a result of
soil water inflow (precipitation and snow melt) and outflow
(root water uptake, evaporation, and deep drainage) dif-
ferences, driven by seasonal and stochastic weather events.
Soil moisture content also varies spatially because of soil
texture, vegetation canopy, litter density, root distribution,
and other landscape heterogeneities (e.g., Wilson et al.
2000; Griffiths et al. 2009; Xu et al. 2013; He et al. 2014).
Y. Ma � R. L. Van Dam (&)
Department of Geological Sciences, Michigan State University,
288 Farm Lane, East Lansing, MI 48824, USA
e-mail: [email protected]
R. L. Van Dam
Institute for Future Environments, Queensland University of
Technology, Gardens Point Campus, 2 George Street, Brisbane,
QLD 4000, Australia
D. H. Jayawickreme
Department of Geography, Bridgewater State University,
Bridgewater, MA 02325, USA
123
Environ Earth Sci (2014) 72:1367–1381
DOI 10.1007/s12665-014-3362-y

Over the years, different methods have been proposed to
assess the influence of these heterogeneities on soil water
(e.g., Breda et al. 2006; Wang et al. 2013). However,
integrating the data from these different methods and
quantifying the collective influence of multiple variables
on spatiotemporal soil water dynamics remains difficult.
In forests, a substantial portion of the precipitation may be
lost due to canopy interception and subsequent canopy
evaporation. The fraction of precipitation that reaches the
ground as stemflow or throughfall is important for soil water
balance and catchment hydrology (Bryant et al. 2005),
nutrient cycling (Michalzik et al. 2001), soil respiration
(Borken et al. 2006), and a range of microbial and biogeo-
chemical processes (Schliemann and Bockheim 2014). The
throughfall fraction, which is a critical component of the soil
water balance, depends on among others the canopy storage
capacity, precipitation intensity and duration, and atmo-
spheric variables such as wind and relative humidity that
influence evaporation rates (e.g., Klaassen 2001; Bryant
et al. 2005). Prior studies of throughfall variability have
shown that canopy interception decreases with distance from
tree stems (Johnson 1990). However, in a detailed study
using randomly spaced rain gauges in a deciduous forest,
Carlyle-Moses et al. (2004) found that the effect of stand
variability on throughfall was insignificant for larger pre-
cipitation events ([5 mm). Similarly, other studies have
looked at canopy attributes of different species and conse-
quent influence on throughfall fractions (e.g., Crockford and
Richardson 2000; Bryant et al. 2005). However, these studies
have rarely explored the influence of throughfall on soil
moisture development in the near surface, which is perhaps
the most important for understanding the aspects of forest
hydrology and biogeochemistry.
Soil moisture measurements have traditionally been
taken using gravimetric measurements of physical soil
samples. Such measurements provide accurate data, but are
time-consuming, invasive, and offer limited temporal and
spatial resolution. To overcome these limitations, time-
domain reflectometry (TDR) equipment, neutron probes,
and tensiometers are now frequently used (Breda et al.
1995; Wullschleger et al. 1998). These are useful methods
to obtain time series of soil water content, although
drawbacks include their invasive nature, limited spatial
sensitivity, susceptibility to errors resulting from poor
sensor-soil contact, and the need for multiple sensors when
soil moisture patterns over even modest field scales need to
be described.
In recent years, electrical resistivity imaging (ERI) has
become a popular method to study spatial and temporal
variability of soil properties (Jayawickreme et al. 2014). It
has been used for soil moisture assessment (e.g., Zhou et al.
2001; Michot et al. 2003), soil temperature studies (Pepin
and Livingston 1995; Musgrave and Binley 2011), and soil
structure and texture exploration (Omonode and Vyn 2006;
Lusch et al. 2009; Chaplot et al. 2010; Basso et al. 2012).
Furthermore, ERI is increasingly used to study the long-
term impacts of land-use conversions (Jayawickreme et al.
2011; Ahmed et al. 2012; Nosetto et al. 2013), agricultural
water management (Islami et al. 2011; Garre et al. 2013),
and to monitor water content in shallow bedrock (Nijland
et al. 2010; Yamakawa et al. 2012). A few recent studies
have also successfully used ERI to assess the relationships
between vegetation variables and soil moisture (Garcia-
Montiel et al. 2008; Robinson et al. 2012). However, no
study has yet attempted to utilize ERI to describe spatio-
temporal variability of soil water in relation to vegetation
structure and throughfall.
The primary objective of this work is to improve our
understanding of soil plant dynamics in temperate-climate
deciduous forests, with a focus on both seasonal change
and spatial variability in soil water content. In particular,
we explore whether the spatial pattern of soil water remains
the same during growing season and whether spatial vari-
ability of soil water is correlated to vegetation structure and
throughfall; we do not explicitly consider stemflow and the
role of plant litter and associated evaporation. To answer
these questions, we studied the growing season (May–
November) soil moisture patterns along with several key
vegetation and water balance attributes including stand
distribution and density, tree basal area, the leaf area index
(LAI), precipitation, and throughfall in a mature forest plot
in Michigan, USA.
Study site and methods
The study site is a patch of deciduous trees in central
Lower Michigan on a glacial till plane with nearly flat
topography (Fig. 1). The forest patch is laterally extensive
in an east–west direction but bordered to the south by a
recent clear-cut and to the north by a grassland. A north-
east- to southwest-oriented transect, 64.5 m in length and
with minimal topographic variations, was used for soil
water measurements with ERI (Fig. 2); the transect
extended another 60 m (40 electrodes) in the northeast
direction in the grassland (Jayawickreme et al. 2010), but
was not used for this study. Soil texture observations from
several boreholes along the transect show that the top
40–60 cm consists of clay loam, and is underlain primarily
by medium- to fine-grained sand, with porosities of 0.47
and 0.39, respectively (Jayawickreme et al. 2008). The
water table fluctuates between 3 and 4 m below the surface,
with highest water levels occurring before the onset of
active photosynthesis in early spring, and lowest water
levels in late fall around leaf senescence (Jayawickreme
et al. 2010).
1368 Environ Earth Sci (2014) 72:1367–1381
123

Vegetation structure
The forest consists of randomly distributed mostly mature
trees ([30 years old) dominated by sugar maple (Acer
saccharum). To characterize the vegetation structure at the
study site, a detailed vegetation survey was conducted in
June 2013 following standard protocols (e.g., Montgomery
and Chazdon 2001). Eight quadrats measuring 9 9 9 m
were laid out sequentially as shown in Fig. 2; each quadrat
centered on one of the TIF/iButton locations described
below. In this study, we only use the results from 5
quadrats (quadrat 2–6; quadrat 1 was only partly covered
by resistivity image points and quadrats 7 and 8 were
affected by the forest–grassland boundary). We mapped the
location of each tree in the quadrats and measured the
diameter at breast height (DBH). Basal area (BA) was
derived from the DBH measurements and scaled to 1
hectare size for easy comparison with values typically
given in the literature (e.g., Wardle et al. 2004). Crown
area (CA) of each tree was estimated using the maximum
canopy extent in four radial directions. The exact heights of
trees at the site were not measured, but they were estimated
to range between 25 and 35 m. Similarly, Crown depth
(CD) was also characterized only qualitatively, with a high
CD corresponding to trees where the first leafed branch
occurs at a height lower than half the tree height. The
results from the survey are summarized in Table 1.
Leaf area index (LAI) is a measure of canopy foliage
density that is broadly defined as the amount of leaf area in
a canopy per unit ground area (Chen et al. 1997). LAI has
been recognized as a key descriptive variable of forest
ecosystems because of the significant role green leaves
play in many biophysical processes. It may also correlate
with light transmittance and with some aspects of inter-
ception and throughfall (Lovett et al. 1996; Crockford and
Richardson 2000; Montgomery and Chazdon 2001). For a
database of measurements (n = 187) for temperate decid-
uous broad-leaved forests from around the world, Asner
et al. (2003) reported LAI values ranging from 1.1 to
8.8 m2/m2, with a mean of 5.1 m2/m2. For this study, a
complete LAI survey was conducted using a Decagon LP-
80 instrument at the peak of the growing season in 2012,
with LAI values (1.8–5.2 m2/m2; average 4.4 m2/m2) that
fall within the range suggested by Asner et al. (2003).
Aboveground monitoring equipment
The climate data obtained from a Michigan Automated
Weather Network (MAWN) station, located approximately
Fig. 1 Location of field site in Michigan
Fig. 2 Detailed layout of the field site showing the location of the
resistivity electrodes, 9 9 9 m vegetation survey quadrats, soil and
environmental sensors, and trees. One tipping-bucket rain gauge was
placed in the forest (survey quadrat 4); the location of the identical
rain gauge in the adjacent grassland plot is not shown in this figure
Table 1 Results from the vegetation survey for each survey quadrat,
giving the number of trees, average diameter at breast height (DBH),
basal area (BA), and average crown area (CA); non-living trees were
included in the calculations of DBH and BA
Quadrat # Total
trees
# Dead
trees
High
CD
DBH (m),
average
BA
(m2/
ha)
CA (m2),
average
2 3 0 2 0.48 102.1 105.7
3 7 1 2 0.28 77.9 36.4
4 1 0 1 0.37 13.0 81.3
5 4 1 1 0.27 32.6 39.0
6 6 0 3 0.25 43.5 24.0
See Fig. 2 for locations of the trees
Environ Earth Sci (2014) 72:1367–1381 1369
123

1.5 km from the site, were augmented with additional
equipment installed on-site. Precipitation information was
obtained from a tipping-bucket rain gauge that was placed
in the grassland. Data loggers recorded the time of every
0.2 mm of rainfall, but suffered from a few outages so that
the data cover only part of the study period. However,
sufficient volume of data was available for the comparison
of data from the on-site tipping-bucket and MAWN
weather station and to analyze the effect of different types
of storms on interception and throughfall.
Canopy throughfall in forests is considered quite vari-
able; in previous research, this has been measured in var-
ious ways, including using troughs, funnels, and standard
rain gauges (e.g., Crockford and Richardson 2000). Here,
we used two approaches for measuring throughfall. To
study the effects of rainfall duration and intensity on
throughfall, we used a tipping-bucket rain gauge in the
forest (Fig. 2). This complements the tipping-bucket rain
gauge that measured un-intercepted rainfall in the adjacent
grassland. To study the effects of canopy interception on
the spatial variability of forest throughfall, we used a novel,
low-budget method (throughfall integrating funnel, TIF)
that measures cumulative rainfall (Dunkerley 2010). In this
method, precipitation is collected in a funnel and guided
over a calcium sulfate hemihydrate tablet (‘‘Plaster of
Paris’’). As water flows over the tablet, a small amount of
calcium sulfate dissolves and is removed from the tablet.
Because the weight loss associated with a single precipi-
tation event is small, this method cannot be used to
quantify individual events. In this study, we used the
method to quantify the cumulative rain and throughfall
during approximately 4-week-long intervals (‘‘tablet
periods’’).
A total of 24 TIFs were installed at the site, including
18 in the forest (Fig. 2). All funnels were located
*65 cm above the ground surface. The TIFs in the
grassland are above the vegetation canopy and therefore
measured cumulative precipitation. Immediately before
field installation, the tablets were oven-dried at 120 �C
for 24 h and weighed. After approximately 4-week
intervals, the tablets were replaced with new ones. The
removed tablets were again oven-dried for 24 h and
weighed. The weight loss recorded during the tablet
period is proportional to the volume of water that has
passed over the tablet during that time interval (Dun-
kerley 2010). To facilitate comparisons between differ-
ent tablets and periods of different lengths, the weight
losses were normalized by starting weights (fractional
weight loss) and scaled to 28-day periods. The
throughfall fraction is then obtained from these data by
comparing the weight losses of TIFs placed under the
forest canopy with the average of those in the grassland.
Belowground monitoring equipment
The study site was instrumented with a suite of equipment
to monitor soil properties (Fig. 2). For soil temperature
measurements, a multidepth array (5, 10, 20, 60, and
100 cm below the surface) of Thermochron iButton sensors
(model DS1922L with a resolution of 0.0625 �C and an
accuracy of 0.5 �C) was installed. Additionally, iButton
sensors were installed 5 cm below the surface along the
forest transect, at 4.5 m lateral spacing. The sensors were
programmed to log data every 2 h and were downloaded
once every few months. A groundwater observation well at
the site was used for water-level and groundwater tem-
perature measurements. Soil moisture capacitance probes
at 20 and 80 cm depths in one location in the forest were
used for comparisons with ERI moisture estimates (Fig. 2).
Soil electrical resistivity data for monitoring soil water
dynamics were collected using a permanent transect. One
of the primary reasons for permanent electrode arrays is the
better repeatability of measurements, which improves the
signal-to-noise ratio for time-lapse resistivity surveys. The
transect consisted of 44 graphite electrodes spaced 1.5 m
apart (Fig. 2). The graphite rods, with 12 mm diameter and
30 cm length, were installed in 2006; they were buried
flush with the surface but made no contact with the soil for
the top 10 cm. To maximize data collection efficiency and
limit electrode-to-cable connection problems, the elec-
trodes were permanently wired to a central take-out loca-
tion using insulated 24-gauge wire. This setup eliminated
electrode position errors and incorrect wiring or connec-
tivity problems. Subsequent to electrode installation, the
area surrounding the transect was only minimally disturbed
by field activity at the site.
Approach
The study period spanned the 2012 growing season from
May through November. Based on the climate data
obtained from the nearby weather station, the temperature
range during the study period was comparable to the long-
term average (Fig. 3). According to long-term precipitation
data for the region, mean monthly precipitation from May
through September varies between 75 and 85 mm, and is
slightly lower (*60 mm) in October and November. The
average annual rainfall for the region is approximately
760 mm/year. Compared to the long-term mean, 2012
weather data showed that the study period was not typical,
with drier conditions in June and July, close to normal in
August and September, and a wetter November (Fig. 3).
During the study period, 24 individual electrical resis-
tivity datasets were collected using an AGI SuperSting
1370 Environ Earth Sci (2014) 72:1367–1381
123

(R8/IP) resistivity system with external switchbox and a
Wenner array configuration at approximately weekly
intervals (Fig. 3). Each dataset along the 44-electrode
transect in the forest had 301 image points distributed over
14 image depths (a-spacings ranged from 1.5 to 22 m). An
analysis of contact resistance data from six datasets (iden-
tified with green lines in Fig. 3) throughout the growing
season (n = 1,806) shows limited correlation with electrode
location or a-spacing, indicating consistent data quality. As
expected, the average contact resistance did increase during
the growing season as the soil dried. In this study, no repeat
measurements or reciprocal data were collected to limit the
overall survey time to less than 90 min. However, previous
work at this site has shown that data quality is very high with
less than 0.2 % of data failing a tight repeat error criterion of
1 % (Jayawickreme et al. 2010). These repeat errors did not
correlate with geometric factors or environmental variables
(e.g., precipitation events and air temperature). Reciprocal
data from a nearby site with similar soil type, comparable
setup, and data collection procedures had an average reci-
procal error of 0.33 % (n = 289 with a-spacings that ranged
from 0.75 to 13.5 m).
To obtain insight into the temporal dynamics at the site,
we analyzed the difference between individual datasets.
Different procedures for inverting time-lapse geophysical
data have been proposed (Miller et al. 2008) including
(a) using one inverted resistivity model as a base model for
subsequent datasets, (b) inverting the difference between
two apparent resistivity datasets, and (c) subtracting
resistivity models after inverting them separately. The first
approach with a base inversion and monitoring datasets is a
common method to obtain differences between two data-
sets. However, potential disadvantages include the large
impact of the choice of base dataset on the resulting dif-
ferences and the possibility of error propagation in datasets
with non-systematic data errors (Jayawickreme et al.
2010). Here, we used the third approach by inverting the
datasets independently and subtracting the resulting models
to obtain the difference. Potential drawbacks of this
approach include the non-uniqueness of individual inver-
sions and the possibility that small errors in the data mask
actual resistivity changes (Daily et al. 2005).
Prior to inversion, clear outliers were removed from the
resistivity datasets. This applied to less than 1 % per
dataset with an average of 0.3 % per dataset. The inver-
sions were performed using AGI 2D Earthimager software.
The initial inversion step involved calculating a forward
model based on the pseudosection resistivity distribution.
We used a finite-difference mesh with a width and height
of 0.5 m. The total mesh consisted of 129 by 22 rectangular
cells plus eight padding cells around the domain, except the
surface. We then used an iterative Occam l2-norm smooth
inversion (Constable et al. 1987). All inversions were
halted at the same iteration step, at RMS data misfit levels
ranging between 2.8 and 4.3 %. This procedure not only
provides a comparable heterogeneity in each resistivity
model but also improves the ability to compare the spatial
distribution of resistivity and water content over time. No
topographic correction was applied as the elevation dif-
ferences along the line were very small and had no sig-
nificant effect on the inversion results.
Temperature correction
The influence of temperature on measured apparent resis-
tivity can be removed using empirical models. A linear
Fig. 3 Climate variables for the field site, including mean daily air
temperature (red line) and daily precipitation during the experiment
(blue vertical bars); data were obtained from the MAWN weather
station at Hancock Turfgrass Research Center, located 1.5 km from
the field site (42.7110, -84.4760). Also shown is the mean daily
temperature based on a record from 1980 to 2009 for East Lansing
(black line). Green hatched lines indicate dates of tablet installation
and replacement, with gray horizontal lines indicating the total
rainfall for each of the tablet periods. The black vertical lines at the
top of the graph indicate ERI data collection days
Environ Earth Sci (2014) 72:1367–1381 1371
123

model was used in this study to correct for the temperature
effect, similar to that in Jayawickreme et al. (2010).
According to Sen and Goode (1992), the resistivity at a
base (reference) temperature can be calculated using
qref
qt
¼ c T � Trefð Þ þ 1; ð1Þ
where qref is the resistivity at a reference temperature
(T = 25 �C) and qt is the measured resistivity at temper-
ature T. The fractional change in resistivity per unit change
in temperature, c, is constant over the temperature range of
interest. Based on the data for glacial till materials, we used
a value of 0.018 (Hayley et al. 2007; Jayawickreme et al.
2008).
Temperature corrections were performed after inverting
the resistivity data. For these corrections, we used tem-
perature readings obtained from the temperature iButton
loggers at depths of 20, 40, 60, and 100 cm (Fig. 4). One
reading, closest to the time of resistivity data collection,
was used for each depth. Temperature was assumed to be
constant at 10.25 �C at 10 m depth after Jayawickreme
et al. (2010). No spatial variability in soil temperature was
incorporated in this correction because average soil tem-
perature data from the iButtons at 5 cm depth (Fig. 2) show
only very minor variations between sensor locations.
Water content conversion
In order to understand soil moisture distribution in space
and its dynamics in time, the resistivity models obtained
through inversion and after correction for the temperature
effect need to be converted to water content. Although the
relationship between electrical resistivity and water content
is well known to be soil-specific, there have been a few
efforts in recent years to establish general pedotransfer
functions (e.g., Hadzick et al. 2011). In this study, we used
material-specific relationships between resistivity (q) and
water content (h) developed in laboratory experiments
(Jayawickreme et al. 2010). Based on the laboratory
results, most of the soil materials tested follow a power
function that can be expressed as S ¼ qs
q
� �m�1, where S is
the saturation (water content/porosity) and qs is the bulk
resistivity of soil at 100 % saturation. The m value was
estimated as 1.16 for sand and 0.67 for clay loam, and qs is
71.53 and 68.15 Xm, respectively (Jayawickreme et al.
2008). After inverted resistivities were corrected for the
temperature effect, the soil water content was calculated.
Soil moisture contents obtained using this procedure cor-
relate well with soil moisture values obtained using the
capacitance probes, but are slightly underestimated:
h(ERI) = 0.93 9 h(cp) ? 0.02 (Jayawickreme et al. 2010).
Results
Resistivity data
The ERI data collection interval allowed for the analysis of
long-term changes corresponding to the periods between
tablet swaps as well as shorter-duration soil water
dynamics. For the analysis of shorter-duration dynamics,
we categorized weeks as having high ([15 mm) or low (0–
5 mm) cumulative precipitation. Plots of resistivity change
between datasets were generated by calculating the percent
difference between (temperature-corrected) resistivity
models usingqnþ1�qn
qn
� �� 100, where qn?1 and qn are the
resistivity values from ending and starting models,
respectively.
During the 2 weeks with low cumulative precipitation at
the beginning of the growing season (Table 2), soil resis-
tivity shows a general increase, as would be expected
(Fig. 5). The week in early June (June 6–13) shows a lower
increase than the week in late June (June 27–July 3), even
though both weeks had B1 mm of water input. For weeks
with high cumulative precipitation (Table 2), the resistivity
response depended strongly on the time of the growing
season (early, mid, or late). During the week with 19.6 mm
of rainfall in early June (May 30–June 6), soil resistivity
shows a very limited decrease, except for a few small areas
near the surface (Fig. 6). The absence of a widespread
decline in resistivity in response to throughfall shows that
Fig. 4 Soil temperature measurements at different depths (0.2, 0.4,
0.6, and 1 m) during ERI data collection. Second-order polynomials
fitted to these data, and extended via a linear function to a constant
temperature at depth (see text), were used for temperature correction
of inverted resistivity models. The first three depths used for
temperature corrections are indicated with dotted horizontal lines
1372 Environ Earth Sci (2014) 72:1367–1381
123

during the early growing season, the vegetative demand for
soil water outweighed the input from throughfall and
stemflow. In contrast, a similar cumulative precipitation
(17.5 mm) later in the growing season (August 30–Sep-
tember 6) caused the shallow soil resistivity to decrease
noticeably, although no change was evident below *0.5 m
depth (Fig. 6). The week in October with 39.6 mm of
cumulative precipitation triggered an even stronger nega-
tive response from the surface to about 0.5 m depth
(Fig. 6). This image also shows the initiation of lateral
resistivity heterogeneities where the negative resistivity
changes span the entire 2 m depth. The locations of most of
these heterogeneities also correlate with areas of minor
resistivity reductions following rainfall in early June
(Fig. 6 top panel).
Inverted resistivity models for the six dates during
which the throughfall tablets were installed or replaced
show a general increase in resistivity from early growing
season in late May until late September (Fig. 7). The
inverted models show some distinct lateral variability with
relatively high resistivity patches at *18.75 and *50 m
distance. There are also some areas with smaller-scale
lateral variability in resistivity, the dimensions of which
appear to change during the growing season. The resistivity
model for the last measurement on October 24 (Fig. 7)
shows a considerable resistivity reduction compared to the
earlier dates. This reduction can be attributed to the sig-
nificant rainfall (81 mm) between September 26 and
October 24 (Table 2). The influence of this rainfall was
accentuated by lower evaporative demand (lower temper-
atures), and fall senescence, which substantially reduced
the interception losses. In the October model, much of the
spatial variability that developed during the growing sea-
son has disappeared.
The first two-and-a-half months of the study period
(mid-May through July) were characterized by rainfall
quantities more than 50 % below average and relatively
high air temperatures. During this period, the resistivity
increased considerably, which corresponds to a distinct
decrease in soil moisture (Fig. 8). During the second half
of the growing season in July and August, rainfall amounts
were significantly higher with totals close to the mean
monthly precipitation for the region (Fig. 3). Thus, near-
surface resistivity occasionally dropped during weeks with
high cumulative rainfall (Fig. 6, center panel). Overall, and
despite the high rainfall, however, the soils continued to
lose moisture during the second half of the growing season
Fig. 5 Percent change in
resistivity for approximately
1-week time periods with low
cumulative precipitation from
June 6 to June 13 and from June
27 to July 3. For this and the
following figures, tick marks on
the horizontal axes correspond
to the boundaries of vegetation
quadrats 2–6 (Fig. 2). All data
shown in this and following
figures (Figs. 6, 7, 8, and 9)
have been temperature-
corrected
Table 2 Details of cumulative precipitation for selected low-
(0–5 mm) and high- ([15 mm) precipitation weeks discussed in the
text, and for the tablet swap periods
Start date End date Cumulative
precipitation
(mm)
Number
of days
Normalized
rainfall
(mm/day)
May 30 June 6 19.55 7 2.8
June 6 June 13 1.01 7 0.14
June 27 July 3 0 6 0
August 30 September 6 17.53 7 2.5
October 10 October 17 39.56 7 5.7
May 30 June 27 31.24 28 1.1
June 27 July 25 24.38 28 0.87
July 25 August 30 65.52 36 1.8
August 30 September 26 55.37 27 2.1
September 26 October 24 81.03 28 2.9
Environ Earth Sci (2014) 72:1367–1381 1373
123

Fig. 6 Percent change in
resistivity for approximately
1-week time periods with high
cumulative precipitation from
May 30 to June 6, August 30 to
September 6, and October 10 to
October 17
Fig. 7 Inverted and
temperature-corrected
resistivity models of six ERI
datasets collected on days of
tablet installation and
replacement (see Fig. 3)
1374 Environ Earth Sci (2014) 72:1367–1381
123

(Fig. 8). The soil water deficit recovered only after Sep-
tember, when plant water use started to decrease and major
rain events took place. The percent change in soil moisture
between two successive tablet periods is shown in Fig. 9. A
decrease in soil moisture indicates that the evapotranspi-
ration and possible soil water drainage exceeded the water
input from throughfall and stemflow for that period. This
series of images clearly shows that the largest changes in
soil moisture occurred early in the growing season during
the first two tablet periods (May 30 to June 27 and June 27
to July 25; Fig. 9). Despite a significant increase in
cumulative rainfall (Fig. 3), soil drying continued for the
next two tablet periods (July 25 to August 30 and August
30 to September 26; Fig. 9). These latter two periods had a
comparable amount of rainfall (65 and 55 mm, respec-
tively; Table 2), but the behavior was quite different with a
small but distinct near-surface soil moisture increase for
the third period from July 25 to August 30. The reduced
drying observed for the third and fourth periods compared
with the first two is likely a direct result of the increased
precipitation. It may be augmented, however, by a lower
plant water demand related to the lower daily mean tem-
perature. Another possibility is that at this time of the year
when the upper soil layer is already dry, the trees use more
water from near of below the water table. This was sug-
gested by Jayawickreme et al. (2008), who showed that the
effective rooting depth of these trees potentially extends to
at least 4 m below the surface during particularly dry
growing seasons. The situation changed for the last tablet
period when water content strongly increased (Fig. 9).
Vegetation structure
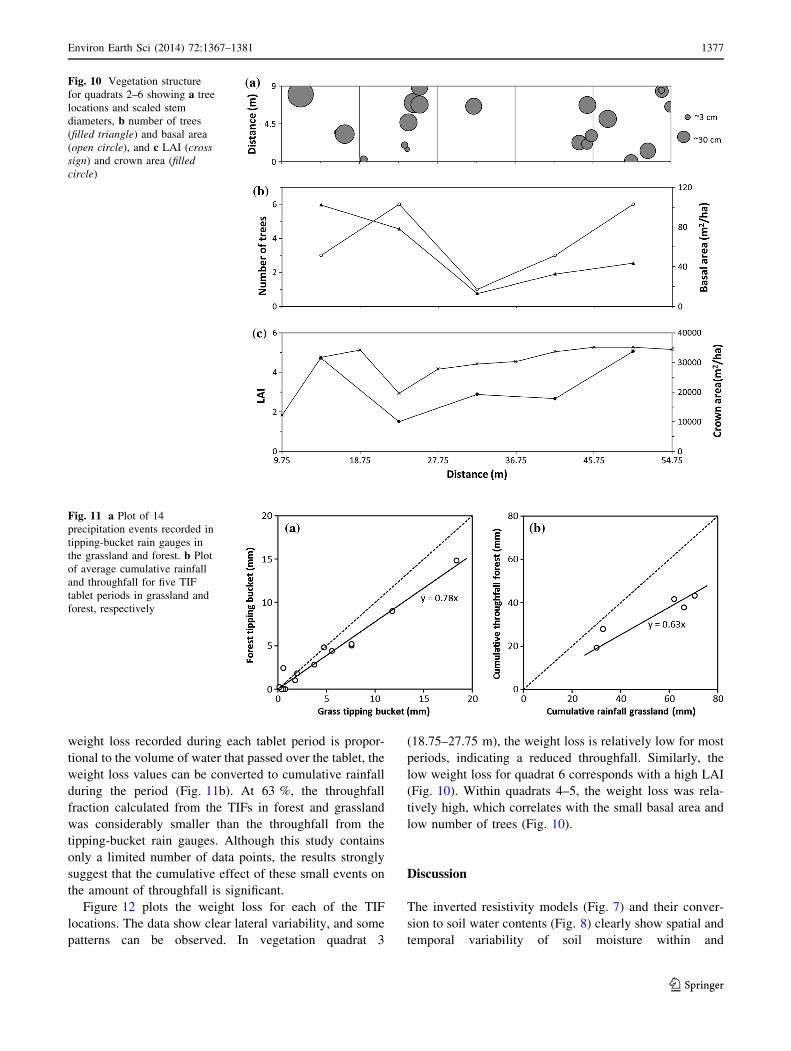
Results of the vegetation survey are given in Fig. 10 with
additional details in Table 1. The survey shows that along
the measurement line there are two distinct areas with a
clustering of trees (Fig. 10a). These clusters are separated
by an approximately 15-m-wide area with low tree density
(27–42 m). This zone encompasses all of quadrat 4 and
part of 5. The number of live trees in each quadrat varies
between 1 (quadrat 4) and 6 (quadrats 3 and 6). The basal
area depends on stem diameter (DBH) and is not neces-
sarily correlated with the number of trees (e.g., quadrat 2 in
Fig. 10b). Quadrat 6, close to the forest edge, has a high
number of trees but the basal area is relatively small. LAI
shows a clear high in quadrats 5 and 6, as does crown area
for quadrat 6. The low values for the LAI and crown area in
vegetation quadrat 3 (Fig. 10c) are surprising as this
Fig. 8 Volumetric water
content for the six dates of
tablet installation and
replacement
Environ Earth Sci (2014) 72:1367–1381 1375
123

quadrat has a relatively large number of trees (Table 1).
However, it must be noted that there is not necessarily a
perfect correlation between stem and canopy measure-
ments because of, among other factors, encroachment of
tree canopies from outside the quadrats. Also, with the LAI
measurements in mid-morning, there may have been some
excess light that entered the forest from the southern
boundary.
Throughfall
As rainfall interception by forest canopies depends on a
large range of factors related to climatic conditions (e.g.,
rainfall duration and intensity, drop size, and wind speed)
and vegetation (e.g., leaf shape and inclination, and LAI), it
is difficult to make generalizations about interception los-
ses and throughfall rates, even for specific forests (Crock-
ford and Richardson 2000). However, it is well known that
for small events, a larger percentage of rainfall is inter-
cepted than for large events (e.g., Link et al. 2004). Once
the canopy storage capacity has been exceeded, for tree
species with downward pointing leaves, excess water will
drip off and fall to the ground. It is therefore understood
that throughfall rates are not linear with event size.
The tipping-bucket rain gauge in the grassland recorded
14 precipitation events during the study period, ranging
from 0.2 to 18 mm. Comparison of these data and with the
tipping-bucket rain gauge in the forest below the forest
canopy gave a forest throughfall fraction of 78 %
(Fig. 11a). There was little difference in this fraction
between events of varying amounts, with a linear rela-
tionship between event size and throughfall fraction
(R2 = 0.97). All events were of relatively short duration,
low intensity, and small size (amount). This likely kept all
events below the canopy storage capacity, resulting in the
linear relationship between event size and throughfall
fraction.
The smallest events in the dataset are not well repre-
sented in the tipping-bucket throughfall analysis. At very
low rainfall amounts, the signal-to-noise ratio becomes
very small and their overall effect cannot be established.
The analysis performed using the throughfall integrating
funnels has the potential to estimate the effect of these
small events, albeit not on an individual event basis. As the
Fig. 9 Percent change in water
content for the five tablet
periods. The anomaly around
29 m in some of the plots is due
to a bad data point in the
September 26 dataset
1376 Environ Earth Sci (2014) 72:1367–1381
123
weight loss recorded during each tablet period is propor-
tional to the volume of water that passed over the tablet, the
weight loss values can be converted to cumulative rainfall
during the period (Fig. 11b). At 63 %, the throughfall
fraction calculated from the TIFs in forest and grassland
was considerably smaller than the throughfall from the
tipping-bucket rain gauges. Although this study contains
only a limited number of data points, the results strongly
suggest that the cumulative effect of these small events on
the amount of throughfall is significant.
Figure 12 plots the weight loss for each of the TIF
locations. The data show clear lateral variability, and some
patterns can be observed. In vegetation quadrat 3
(18.75–27.75 m), the weight loss is relatively low for most
periods, indicating a reduced throughfall. Similarly, the
low weight loss for quadrat 6 corresponds with a high LAI
(Fig. 10). Within quadrats 4–5, the weight loss was rela-
tively high, which correlates with the small basal area and
low number of trees (Fig. 10).
Discussion
The inverted resistivity models (Fig. 7) and their conver-
sion to soil water contents (Fig. 8) clearly show spatial and
temporal variability of soil moisture within and
Fig. 10 Vegetation structure
for quadrats 2–6 showing a tree
locations and scaled stem
diameters, b number of trees
(filled triangle) and basal area
(open circle), and c LAI (cross
sign) and crown area (filled
circle)
Fig. 11 a Plot of 14
precipitation events recorded in
tipping-bucket rain gauges in
the grassland and forest. b Plot
of average cumulative rainfall
and throughfall for five TIF
tablet periods in grassland and
forest, respectively
Environ Earth Sci (2014) 72:1367–1381 1377
123

between the study quadrats. Similarly, spatial variability is
observed in the data on vegetation characteristics and
throughfall (Figs. 10, 12). In this section, we will address
the persistence of these spatial patterns throughout the
growing season, as well as the correlation between soil
moisture and the other measured variables.
Figure 13 presents the average water content below
each vegetation quadrat, for depth ranges of 0–50,
50–100, and 100–200 cm to emphasize different parts of
the root zone. The results show that at the beginning of
the growing season, the soil moisture contents were
highest, irrespective of the depth range. Soil moisture
distribution appears fairly constant across the array with
relatively small differences between vegetation quadrats.
At most times, quadrat 6 had the lowest water content
values along the array. The changes in water content
during tablet periods (Fig. 14) show a clear drying pattern
(negative change), except for the last period from Sep-
tember 26 to October 24. The changes throughout the
growing season are most significant for the 0- to 50-cm-
depth range (Fig. 14a). The smallest change along the
survey transect nearly always occurred in vegetation
quadrat 4 (between 27.75 and 36.75 m), which had the
lowest number of trees, the smallest basal area, and a
slightly higher throughfall volume (high TIF fractional
weight loss; Fig. 12). This suggests that the small and
slow resistivity response in this quadrat is driven by a
smaller soil water extraction by plants and the greater
throughfall volume, which likely kept the soil replenished
with moisture.
To compare soil moisture at different depths along the
measurement transect with data on throughfall and soil
temperature we used only the sensors located in the centers
of the five vegetation quadrats. Sensors located on the
quadrat edges (Fig. 2) were not included in these com-
parisons. The correlation between soil temperature (both
absolute and differenced between data collection periods)
and vegetation parameters and soil moisture did not pro-
duce statistically significant correlations. This was an
expected result based on the earlier observation of limited
spatial variability in soil temperature along the transect.
During the first 2 months of the growing season, the soil
dried relatively fast, especially in the shallow soil layers
(Fig. 14a). A comparison of soil moisture change and
normalized tablet weight loss for these months shows that
in quadrats with more throughfall, the drying is less pro-
nounced (Fig. 15a). This result is as expected, but the
correlations are relatively weak. This may be the result of
the previously discussed strong heterogeneity in through-
fall that is difficult to capture using the TIF method and
limited measurement locations. During the third and fourth
tablet periods, when soil moisture content changed rela-
tively little (Fig. 14), there is no correlation between tablet
weight loss and the soil moisture change (Fig. 15a). During
the final tablet period, strongest wetting is concentrated in
quadrats with less throughfall. This result may indicate that
in these quadrats the growing season moisture deficit was
most significant.
A quantitative comparison between soil moisture and
vegetation characteristics shows a strong negative corre-
lation with crown area (Fig. 15b) and LAI during the
growing season. This correlation between soil moisture and
canopy indicators was strongest at the start of growing
season, when quadrats with more canopy (high LAI or
large crown area) had distinctly lower soil moisture con-
tents. The slope of this correlation gradually dropped
throughout the growing season (although the correlation
coefficients remain strong), which suggests that the root
water uptake was uniform along the transect and uncorre-
lated with canopy structure. The final period from Sep-
tember 26 to October 24 at the end of the growing season,
which was characterized by high rainfall amounts (Fig. 2),
coincided with leaf fall-off. Higher throughfall quantities
along the transect resulted in disappearance of the corre-
lation between soil moisture and canopy indicators
(Fig. 15b), as would be expected. The lower depth intervals
showed comparable behavior, although the effect of can-
opy became less significant with depth. Analysis of the data
shows no strong correlation between DBH and the number
of trees with soil moisture distribution; this is no surprise as
these vegetation variables do not significantly impact
interception and throughfall.
Fig. 12 Tablet weight loss for
each of the TIF locations along
the survey transect. To enable
direct comparisons, the weights
have been normalized to correct
for different starting weights of
the tablets and scaled to 28-day
periods
1378 Environ Earth Sci (2014) 72:1367–1381
123

0
0.1
0.2
0.3
0.4
9.75 27.75 45.75
So
il m
ois
ture
Distance (m)9.75 27.75 45.75
Distance (m)
9.75 27.75 45.75
Distance (m)
5/30 6/277/25 8/309/26 10/24
(a) (b) (c)
Fig. 13 Average ERI-derived soil moisture for vegetation survey quadrats 2–6 from a 0–50 cm, b 50–100 cm, and c 100–200 cm depth
-100
0
9.75 27.75 45.75
So
il m
ois
ture
ch
ang
e (%
)
Distance (m)
9.75 27.75 45.75
Distance (m)
9.75 27.75 45.75
Distance (m)
5/30-6/276/27-7/257/25-8/308/30-9/269/26-10/24
(a) (b) (c)
Fig. 14 Percent change in soil moisture between tablet periods for vegetation survey quadrats 2–6 from a 0–50 cm, b 50–100 cm, and
c 100–200 cm depth
Fig. 15 Comparison of soil moisture (averaged per quadrat), vege-
tation properties, and throughfall data. a Change in soil moisture in 5
vegetation quadrats versus tablet weight loss. To enable direct
comparisons, the weights were normalized to correct for different
starting weights of the tablets and scaled to 28-day periods. b Average
soil moisture in 5 vegetation quadrats versus crown area. Linear
regression lines (dashed) are given for datasets on May 30 (first and
wettest), September 26 (driest), and October 24 (last), and the average
(solid line). In both a and b, soil moisture from 0–50 cm was
averaged for each quadrat
Environ Earth Sci (2014) 72:1367–1381 1379
123

Conclusions
This work presented a study of the effects of vegetation
structure on near-surface soil moisture, and its relation
with throughfall, in a temperate deciduous woodland. The
temporal dynamics of soil moisture monitored during the
growing season showed near-continuous drying. This was
especially the case early in the growing season when
precipitation was significantly below normal at the site.
However, this soil drying continued well into the second
half of the growing season when precipitation amounts
were substantially higher. During weeks with significant
precipitation amounts, only minor interruptions to this
drying trend were observed. In the early parts of the
growing season, even with substantial throughfall
amounts, no significant interruption to the drying occur-
red, likely as a result of high plant water consumption.
Later in the growing season, similar throughfall produced
more distinct wetting signatures. Based on the resistivity
data gathered, we also observed that with influxes of
precipitation during the growing season, significant spa-
tial heterogeneities in resistivity or soil moisture devel-
oped. These patterns in resistivity and soil moisture
correlated with some vegetation characteristics, in par-
ticular with the crown area of the trees and LAI. The
number of trees and DBH had no significant effect on the
distribution of soil moisture at the site. The spatial vari-
ability in soil moisture was also weakly correlated with
throughfall. The data presented in this paper provide
insights into the important relationships between vegeta-
tion characteristics of forests, including stand properties
and interception, and soil moisture that are still noticeably
absent in the literature.
Acknowledgments This research was funded by the US National
Science Foundation (NSF Grant EAR-0911642). Any opinions,
findings, and conclusions or recommendations expressed in this
publication are those of the authors and do not necessarily reflect the
views of the NSF. We acknowledge Agustin Brena, David Hyndman,
Anthony Kendall, Alex Kuhl, James Loop, and Ryan Nagelkirk for
field assistance and discussion of results. The manuscript benefited
from constructive comments by three anonymous reviewers.
References
Ahmed YAR, Pichler V, Homolak M, Gomoryova E, Nagy D,
Pichlerova M, Gregor J (2012) High organic carbon stock in a
karstic soil of the Middle-European Forest Province persists after
centuries-long agroforestry management. Eur J For Res
131:1669–1680
Asbjornsen H, Goldsmith GR, Alvarado-Barrientos MS, Rebel K,
Van Osch FP, Rietkerk M, Chen J, Gotsch S, Tobon C, Geissert
DR, Gomez-Tagle A, Vache K, Dawson TE (2011) Ecohydro-
logical advances and applications in plant–water relations
research: a review. J Plant Ecol 4:3–22
Asner GP, Scurlock JMO, Hicke JA (2003) Global synthesis of leaf
area index observations: implications for ecological and remote
sensing studies. Glob Ecol Biogeogr 12:191–205
Basso B, Fiorentino C, Cammarano D, Cafiero G, Dardanelli J (2012)
Analysis of rainfall distribution on spatial and temporal patterns
of wheat yield in Mediterranean environment. Eur J Agron
41:52–65
Borken W, Savage K, Davidson EA, Trumbore SE (2006) Effects of
experimental drought on soil respiration and radiocarbon efflux
from a temperate forest soil. Glob Change Biol 12:177–193
Breda N, Granier A, Barataud F, Moyne C (1995) Soil water
dynamics in an oak stand I. Soil moisture, water potentials and
water uptake by roots. Plant Soil 172:17–27
Breda N, Huc R, Granier A, Dreyer E (2006) Temperate forest trees
and stands under severe drought: a review of ecophysiological
responses, adaptation processes and long-term consequences.
Ann Forest Sci 63:625–644
Bryant ML, Bhat S, Jacobs JM (2005) Measurements and modeling of
throughfall variability for five forest communities in the
southeastern US. J Hydrol 312:95–108
Carlyle-Moses DE, Flores Laureano JS, Price AG (2004) Throughfall
and throughfall spatial variability in Madrean oak forest
communities of northeastern Mexico. J Hydrol 297:124–135
Chaplot V, Lorentz S, Podwojewskia P, Jewitt G (2010) Digital
mapping of A-horizon thickness using the correlation between
various soil properties and soil apparent electrical resistivity.
Geoderma 157:154–164
Chen JM, Rich PM, Gower ST, Norman JM, Plummer S (1997) Leaf
area index of boreal forests: theory, techniques, and measure-
ments. J Geophys Res 102:29429–29443
Constable SC, Parker RL, Constable CG (1987) Occams inversion: a
practical algorithm for generating smooth models from electro-
magnetic sounding data. Geophysics 52:289–300
Crockford RH, Richardson DP (2000) Partitioning of rainfall into
throughfall, stemflow and interception: effect of forest type,
ground cover and climate. Hydrol Process 14:2903–2920
Daily W, Ramirez A, Binley A, LaBrecque D (2005) Electrical
resistance tomography-theory and practice. In: Butler DK (ed)
Investigations in geophysics. Society of exploration geophysics,
vol 13. p 525
Dunkerley DA (2010) A new method for determining the throughfall
fraction and throughfall depth in vegetation canopies. J Hydrol
385:65–75
Garcia-Montiel DC, Coe MT, Cruz MP, Ferreira JN, da Silva EM,
Davidson EA (2008) Estimating seasonal changes in volumetric
soil water content at landscape scales in a savanna ecosystem
using two-dimensional resistivity profiling. Earth Interact
12:1–25
Garre S, Coteur I, Wongleecharoen C, Kongkaew T, Diels J,
Vanderborght J (2013) Noninvasive monitoring of soil water
dynamics in mixed cropping systems: a case study in Ratchaburi
Province, Thailand. Vadose Zone Journal 12
Griffiths RP, Madritch MD, Swanson AK (2009) The effects of
topography on forest soil characteristics in the Oregon Cascade
Mountains (USA): implications for the effects of climate change
on soil properties. For Ecol Manag 257:1–7
Hadzick ZZ, Guber AK, Pachepsky YA, Hill RL (2011) Pedotransfer
functions in soil electrical resistivity estimation. Geoderma
164:195–202
Hansen MC, Defries RS, Townshend JRG, Sohlberg R (2000) Global
land cover classification at 1 km spatial resolution using a
classification tree approach. Int J Remote Sens 21:1331–1364
Hayley K, Bentley LR, Gharibi M, Nightingale M (2007) Low
temperature dependence of electrical resistivity: implications for
near surface geophysical monitoring. Geophys Res Lett
34:L18402
1380 Environ Earth Sci (2014) 72:1367–1381
123

He L, Ivanov VY, Bohrer G, Maurer KD, Vogel CS, Moghaddam M
(2014) Effects of fine-scale soil moisture and canopy heteroge-
neity on energy and water fluxes in a northern temperate mixed
forest. Agric For Meteorol 184:243–256
Islami N, Taib S, Yusoff I, Ghani AA (2011) Time lapse chemical
fertilizer monitoring in agriculture sandy soil. Int J Environ Sci
Technol 8:765–780
Jayawickreme DH, Van Dam RL, Hyndman DW (2008) Subsurface
imaging of vegetation, climate and root-zone moisture interac-
tions. Geophys Res Lett 35:L18404
Jayawickreme DH, Van Dam RL, Hyndman DW (2010) Hydrological
consequences of land-cover change: quantifying the influence of
plants on soil moisture with time-lapse electrical resistivity.
Geophysics 75:43–50
Jayawickreme DH, Santoni CS, Kim JH, Jobbagy EG, Jackson RB
(2011) Changes in hydrology and salinity accompanying a
century of agricultural conversion in Argentina. Ecol Appl
21:2367–2379
Jayawickreme DH, Jobbagy EG, Jackson RB (2014) Geophysical
subsurface imaging for ecological applications. New Phytol
201:1170–1175
Johnson RC (1990) The interception, throughfall and stemflow in a
forest in highland Scotland and the comparison with other
upland forests in the UK. J Hydrol 118:281–287
Keddy PA (1996) Ecological properties for the evaluation, manage-
ment, and restoration of temperate deciduous forest ecosystems.
Ecol Appl 6:748–762
Klaassen W (2001) Evaporation from rain-wetted forest in relation to
canopy wetness, canopy cover, and net radiation. Water Resour
Res 37:3227–3236
Link TE, Unsworth M, Marks D (2004) The dynamics of rainfall
interception by a seasonal temperate rainforest. Agric For
Meteorol 124:171–191
Lovett GM, Nolan SS, Driscoll CT, Fahey TJ (1996) Factors
regulating throughfall flux in a new New-Hampshire forested
landscape. Can J For Res 26:2134–2144
Lusch DP, Stanley KE, Schaetzl RJ, Kendall AD, Van Dam RL,
Nielsen A, Blumer BE, Hobbs TC, Archer JK, Holmstadt JLF,
May CL (2009) Characterization and mapping of patterned
ground in the Saginaw Lowlands, Michigan: possible evidence
for Late-Wisconsin permafrost. Ann Assoc Am Geogr 99:1–22
McElrone AJ, Choat B, Gambetta GA, Brodersen CR (2013) Water
uptake and transport in vascular plants. Nat Educ Knowl 4:6
McKenney-Easterling M (2000) The potential impacts of climate
change and variability on forests and forestry in the Mid-Atlantic
Region. Clim Res 14:195–206
Michalzik B, Kalbitz K, Park JH, Solinger S, Matzner E (2001)
Fluxes and concentrations of dissolved organic carbon and
nitrogen: a synthesis for temperate forests. Biogeochemistry
52:173–205
Michot D, Benderitter Y, Dorigny A, Nicoullaud B, King D, Tabbagh
A (2003) Spatial and temporal monitoring of soil water content
with an irrigated corn crop cover using surface electrical
resistivity tomography. Water Resour Res 39:1138
Miller CR, Routh PS, Brosten TR, McNamara JP (2008) Application
of time-lapse ERT imaging to watershed characterization.
Geophysics 73:G7–G17
Montgomery RA, Chazdon RL (2001) Forest structure, canopy
architecture, and light transmittance in tropical wet forests.
Ecology 82:2707–2718
Musgrave H, Binley A (2011) Revealing the temporal dynamics of
subsurface temperature in a wetland using time-lapse geophys-
ics. J Hydrol 396:258–266
Nijland W, van der Meijde M, Addink E, de Jong SM (2010)
Detection of soil moisture and vegetation water abstraction in a
Mediterranean natural area using electrical resistivity tomogra-
phy. Catena 81:209–216
Nosetto MD, Acosta AM, Jayawickreme DH, Ballesteros SI, Jackson
RB, Jobbagy EG (2013) Land-use and topography shape soil and
groundwater salinity in central Argentina. Agric Water Manag
129:120–129
Omonode RA, Vyn TJ (2006) Spatial dependence and relationships of
electrical conductivity to soil organic matter, phosphorus, and
potassium. Soil Sci 171:223–238
Peng W, Song T, Zeng F, Wang K, Du H, Lu S (2013) Spatial
distribution of surface soil water content under different
vegetation types in northwest Guangxi, China. Environ Earth
Sci 69:2699–2708
Pepin S, Livingston NJ (1995) Temperature-dependent measurement
errors in time domain reflectometry determinations of soil water.
Soil Sci Soc Am J 59:38–43
Reed DD, Desanker PV (1992) Ecological implications of projected
climate change scenarios in forest ecosystems in northern
Michigan, USA. Int J Biometeorol 36:99–107
Robinson JL, Slater LD, Schafer KVR (2012) Evidence for spatial
variability in hydraulic redistribution within an oak-pine forest
from resistivity imaging. J Hydrol 430:69–79
Schafer KVR, Oren R, Lai C-T, Katul GG (2002) Hydrologic balance
in an intact temperate forest ecosystem under ambient and
elevated atmospheric CO2 concentration. Glob Change Biol
8:895–911
Schliemann SA, Bockheim JG (2014) Influence of gap size on carbon
and nitrogen biogeochemical cycling in Northern hardwood
forests of the Upper Peninsula, Michigan. Plant Soil
377:323–335
Sen PN, Goode PA (1992) Influence of temperature on electrical
conductivity of shaly sands. Geophysics 57:89–96
Wang Y, Zhu H, Li Y (2013) Spatial heterogeneity of soil moisture,
microbial biomass carbon and soil respiration at stand scale of an
arid scrubland. Environ Earth Sci 70:3217–3224
Wardle DA, Walker LR, Bardgett RD (2004) Ecosystem properties
and forest decline in contrasting long-term chronosequences.
Science 305:509–513
Wen J (1999) Evolution of eastern Asian and eastern North American
disjunct distributions in flowering plants. Annu Rev Ecol Syst
30:421–455
Wilson KB, Hanson PJ, Baldocchi DD (2000) Factors controlling
evaporation and energy partitioning beneath a deciduous forest
over an annual cycle. Agric For Meteorol 102:83–103
Wullschleger SD, Meinzer FC, Vertessy RA (1998) A review of
whole-plant water use studies in trees. Tree Physiol 18:499–512
Xu S, Liu LL, Sayer EJ (2013) Variability of above-ground litter
inputs alters soil physicochemical and biological processes: a
meta-analysis of litterfall-manipulation experiments. Biogeo-
sciences 10:7423–7433
Yamakawa Y, Kosugi K, Katsura S, Masaoka N, Mizuyama T (2012)
Spatial and temporal monitoring of water content in weathered
granitic bedrock using electrical resistivity imaging. Vadose
Zone J 11. doi:10.2136/vzj2011.0029
Zhou QY, Shimada J, Sato A (2001) Three-dimensional spatial and
temporal monitoring of soil water content using electrical
resistivity tomography. Water Resour Res 37:273–285
Environ Earth Sci (2014) 72:1367–1381 1381
123