selection of reference genes for quantitative real-time pcr during flower bud development in cms7311...
TRANSCRIPT

SHORT COMMUNICATION
Selection of reference genes for quantitative real-time PCRduring flower bud development in CMS7311 of heading Chinesecabbage (Brassica rapa L. ssp. pekinensis)
Xiaoyong Xu • Zeping Yang • Xilu Sun •
Lugang Zhang • Zhiyuan Fang
Received: 4 May 2013 / Revised: 31 October 2013 / Accepted: 31 October 2013 / Published online: 20 November 2013
� Franciszek Gorski Institute of Plant Physiology, Polish Academy of Sciences, Krakow 2013
Abstract The accuracy of quantitative real-time PCR
(qRT-PCR) depends on the stability of the reference gene
used for normalization. In heading Chinese cabbage (Bras-
sica rapa L. ssp. pekinensis), the most stable reference genes
for qRT-PCR during flower bud development have not been
elucidated. In this study, the statistical software geNorm was
used to test eight candidate reference genes during flower
bud development in male sterile (Ms) and fertile (Mf) plants.
The result revealed that the stability order was Tub/GAP-
DH [ Cyp [ EF1a [ U34559 [ BrTip41 [ Apr [ 18S
rRNA, Tub and GAPDH were the most stable genes [average
expression stability (M) 0.614], and the combined use of six
reference genes [pairwise variation (V) 0.15] was suggested
to be the optimal reference gene for qRT-PCR during flower
bud development. Furthermore, the expressions of
BcPME31 during flower bud development normalized with
the combined use of six reference genes and with GAPDH or
Tub alone were compared; the various results also suggested
that selection of the optimal reference gene was necessary for
gene expression analysis.
Keywords Flower bud development � Heading
Chinese cabbage � Male sterility � qRT-PCR �Reference gene selection
Abbreviations
CMS Cytoplasmic male sterility
GAPDH Glyceraldehyde-3-phosphate dehydrogenase
qRT-PCR Quantitative real-time polymerase chain
reaction
Ms Male sterile
Mf Male fertile
Rf Restorer gene
Introduction
The use of gene expression analysis is very important in
functional research, and this has led to the development of
various methods to quantify gene expression. Compared
with traditional methodologies such as Northern hybrid-
ization and quantitative competitive polymerase chain
reaction (PCR), the use of quantitative real-time PCR
Communicated by P. Sowinski.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11738-013-1437-0) contains supplementarymaterial, which is available to authorized users.
X. Xu � Z. Yang � X. Sun � L. Zhang (&) � Z. Fang
College of Horticulture, State Key Laboratory of Crop Stress
Biology in Arid Areas, Northwest A&F University, Yangling
712100, Shaanxi, People’s Republic of China
e-mail: [email protected]
X. Xu
e-mail: [email protected]
Z. Yang
e-mail: [email protected]
X. Sun
e-mail: [email protected]
X. Xu
College of Horticulture, Shanxi Agriculture University, Taigu
030801, Shanxi, People’s Republic of China
Z. Fang (&)
Institute of Vegetables and Flowers, Chinese Academy of
Agricultural Sciences, Beijing 100081, People’s Republic of
China
e-mail: [email protected]
123
Acta Physiol Plant (2014) 36:809–814
DOI 10.1007/s11738-013-1437-0

(qRT-PCR) is more responsive, rapid, and provides high-
throughput results. This has leaded to its being widely used
to identify gene expression in various fields of biological
research. However, the accuracy of qRT-PCR is dependent
on the choice and stability of reference genes used for
normalization. Presently, a number of reference genes have
been widely used for qRT-PCR, including b-actin, glyc-
eraldehyde-3-phosphate dehydrogenase (GAPDH), 18S
rRNA, EF-1-a, ubiquitin, and a or b-tubulin (Gu et al.
2011; Qi et al. 2010). However, more and more studies
have suggested that no single reference gene is universal
for all experiments (Gutierrez et al. 2008; Tong et al. 2009;
Dong et al. 2012). This makes it important to choose the
optimal reference genes in each biological system before
using qRT-PCR to investigate gene expression.
Heading Chinese cabbage (Brassica rapa L. ssp. pekin-
ensis) is one of the most important leafy vegetables in Asia.
With the completion of the Chinese cabbage Chiifu-401-42
genome sequencing project, it follows that functional
genomics will become more significant in this species,
making transcript analysis both necessary and important.
Currently, several candidate reference genes have been
compared under drought stress and downy mildew infection
(Qi et al. 2010), but the stability of these genes in different
flower bud stages has not been verified. Furthermore, male
sterility is an unusual phenomenon in higher plants, and there
is no difference in phenotype from normal plants except for
the presence of dysfunctional or degenerate stamens. It was
reported that the expression of thousands of genes was
changed during flower stamen development (Alves-Ferreira
et al. 2007). Therefore, it is necessary to define the reference
gene for qRT-PCR before assessing the expression of genes
of interest during flower bud development in the male sterile
plant of heading Chinese cabbage.
In this study, the statistical software geNorm (Vande-
sompele et al. 2002) was used to evaluate the expression
stability of eight candidate reference genes during flower
bud development in male sterile (Ms) and male fertile (Mf)
plants of heading Chinese cabbage. Based on the statistical
result, the expression level of BcPME31 during flower bud
development was also quantized. This is the first report
about the selection of reference genes during flower bud
development and will be of special interest in the field of
male sterility relative research.
Materials and methods
Plant material preparation
Ms and Mf plants used in this study have been described
previously (Xu et al. 2013). The flower buds of Ms and Mf
plants were collected and sorted into four groups based on
the bud size length (\1.0, 1.0–2.0, 2.0–3.0, and
4.0–5.0 mm in length). Buds of analogous size were mixed
together and stored at -80 �C until required.
Total RNA isolation and cDNA synthesis
Total RNA was isolated from the floral bud samples uti-
lizing Bizol (BIOER, China) according to the manufac-
turer’s instructions. Recombinant DNase I (Takara, Japan)
was used at 37 �C for 30 min to remove trace DNA. The
quality of total RNA was determined by agarose gel elec-
trophoresis and an ultraviolet spectrophotometer (Nano-
Drop 2000C, Thermo Scientific, Wilmington, DE).
Equal amounts of total RNA (about 2 lg) in all samples
were used for first-strand cDNA synthesis by Prime-
ScriptTM RT reagent kit (TaKaRa, Japan), according to the
manufacturer’s instructions. Purified cDNA was dissolved
in ddH2O and stored at -20 �C for use. RNA extraction
and cDNA synthesis from all of the different samples were
performed for three biological replicates.
Primer design
The optimal reference gene was determined from nine
candidate genes in all samples. The designs of primers for
six candidate genes including EF1a, GAPDH, Ubc, 18 s
rRNA, Apr, and Cyp were excerpted from previous studies
(Qi et al. 2010), with the exception of Tub where primers
were redesigned to amplify a fragment about 110 bp. Two
candidate genes, BrTIP41 and U34559, were accorded to
the gene TIP41 in Brassica napus and AT4G33380 in
Arabidopsis, respectively (Chen et al. 2010; Czechowski
et al. 2005), and their sequence annotations were referred
to as Bra011516 and Bra034559 in the Brassica Databases
(BRAD http://brassicadb.org/brad/). The primers were
designed with a Tm value of approximately 58–62 �C
using the software Primer Premier 5 (www.PremierBiosoft.
com). The specificity of each primer was confirmed by
BLAST searching against the Brassica Database. The pri-
mer sequences are listed in Table S1.
QRT-PCR and data analysis
QRT-PCR was carried out in an optical 8-well plate (Ax-
ygen, USA) using the Bio-Rad IQ5 System (Bio-Rad,
America). SYBR Green II (Takara, Japan) was used to
monitor dsDNA synthesis. The reactions were performed
in triplicate in a final volume of 20 ll (SYBR� Premix Ex
TaqTM (29) 10 ll; forward and reverse primers (10 lM)
0.4 ll each; tenfold diluted cDNA 2 ll; sterile ddH2O
7.2 ll) using the following program: initial denaturation at
95 �C for 3 min, followed by 40 cycles (95 �C for 20 s,
60 �C for 30 s, 2 �C for 30 s). Melting curve analysis was
810 Acta Physiol Plant (2014) 36:809–814
123
performed to check for the occurrence of primer dimers
and non-specific PCR products by using cycles of 95 �C for
15 s followed by a constant increase of temperature
between 60 and 95 �C. The amplification specificity was
also determined by electrophoresis and sequence analysis
of their PCR products.
A standard curve for each gene was generated using ten-
fold serial dilutions of pooled cDNAs with equal templates of
every sample. The efficiency of these primers was calculated
using LinRegPCR (Ramakers et al. 2003). The statistical
software program geNorm (Vandesompele et al. 2002) was
employed to compare the expression stability of these can-
didate genes in eight samples from Ms and Mf plants.
Results and discussion
Primer specificity and efficiency
To evaluate the amplification specificity, agarose gel
electrophoresis was performed for amplicons of the can-
didate reference genes derived from pooled cDNA and
genomic DNA templates (Fig. S1). All primer pairs gen-
erated only single amplicon band in Ms and Mf cDNA
pools. Except BrTip41 whose forward primer was com-
posed of parts of two exons and could not amplify any
band, the rest of the primer pairs could amplify the specific
band with DNA templates; especially four genes including
GAPDH, Ubc, Apr, and U34559 showed amplification
differences in amplicon size between DNA and cDNA
templates, as their primers span an intron in the genome.
As a result, these four genes could be used for RNA
extraction quality assay. All these interest bands were
cloned and sequenced; except non-specific amplification of
genomic DNA by Ubc primers, the sequences of the
remaining bands showed high identities with expected
sequences (data not shown).
Before carrying out qRT-PCR, all of the RNA samples
of high quality were verified by PCR results of GAPDH
and Apr. There was no genomic DNA contamination in
cDNA templates. Neither primer dimers nor unexpected
products were found (data not shown). Amplification effi-
ciency ranged from 93.9 % in U34559 to 124.3 % in Ubc,
and coefficient of determination (R2) varied from 0.994 to
0.998 (Table S1). Except Ubc, which was abolished for the
amplification efficiency over 120 %, the remaining genes
were demonstrated to be efficient and specific for the target
gene amplification.
Expression dynamics of the candidate reference genes
RNA transcription levels were measured to evaluate the
expression stability of the remaining eight candidate genes
(Fig. S2). A relatively wide range of Ct values was
observed from 7.06 to 34.03 in eight tested samples, and
most Ct values were between 20 and 30. The least abun-
dant transcripts were of BrTip41 with Ct value 34.03. The
transcript level of 18S rRNA was expressed highly with the
minimum Ct value of 7.06.
Statistical analysis of gene expression stability
The Ct datasets of eight candidate genes were transformed
into relative quantification data using Microsoft Excel with
the 2-DCT method and then imported into geNorm for
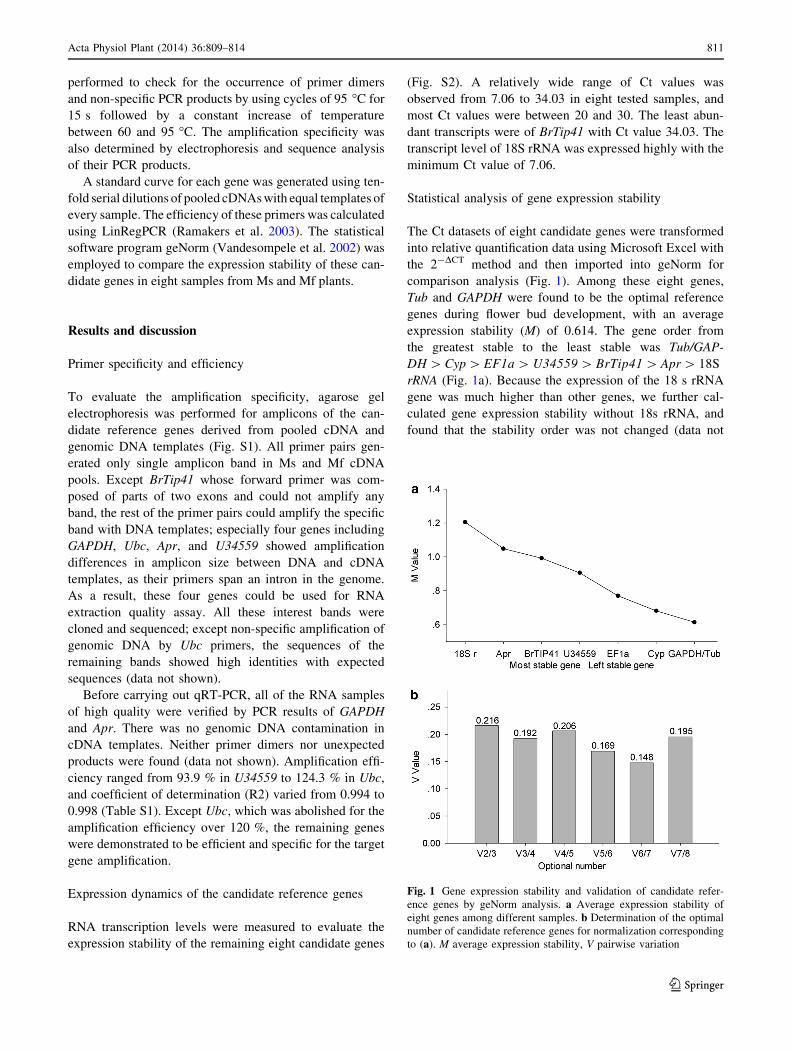
comparison analysis (Fig. 1). Among these eight genes,
Tub and GAPDH were found to be the optimal reference
genes during flower bud development, with an average
expression stability (M) of 0.614. The gene order from
the greatest stable to the least stable was Tub/GAP-
DH [ Cyp [ EF1a [ U34559 [ BrTip41 [ Apr [ 18S
rRNA (Fig. 1a). Because the expression of the 18 s rRNA
gene was much higher than other genes, we further cal-
culated gene expression stability without 18s rRNA, and
found that the stability order was not changed (data not
Fig. 1 Gene expression stability and validation of candidate refer-
ence genes by geNorm analysis. a Average expression stability of
eight genes among different samples. b Determination of the optimal
number of candidate reference genes for normalization corresponding
to (a). M average expression stability, V pairwise variation
Acta Physiol Plant (2014) 36:809–814 811
123
shown). The Ct values of eight candidate genes were also
analyzed by BestKeeper (Pfaffl et al. 2004) (Table S2).
After the geNorm analysis, Tub showed the best value
among all candidate genes, with GAPDH next and 18S
rRNA the least stable gene. The main difference was that
the stability order of the other five genes was not consistent
with that of geNorm analysis.
Although Tub was proved to be one of the most variable
genes in considering male and female reproductive tissues,
spikelets, roots, and leaves of apomictic and sexual Bra-
chiaria brizantha (Silveira et al. 2009) and in leaves, roots,
and stems of two soybean cultivars exposed to cadmium
(Wang et al. 2012), it was also regarded as one of the most
stable genes in all developmental stages and under all stress
conditions of Platycladus orientalis (Chang et al. 2012)
and in our experiment. All these results illustrate that ref-
erence gene Tub was not suited for all experiments.
GAPDH was regarded as one of the most stable genes in
various stress environments and in different tissues during
plant development (Qi et al. 2010), and has been widely
used as reference gene in Chinese cabbage (Zhang et al.
2010, 2009). This gene was also identified as the second
stable gene in our experiment. These results suggest that
GAPDH had a wide range of adaptability.
Additionally, geNorm also recommended the minimum
number of reference genes required for determining a
reliable normalization factor. Vandesompele et al. (2002)
have proposed 0.15 as the threshold value for the pairwise
variation (V), below which, the additional reference gene is
not required. The least pairwise variation in our samples
was 0.148 (Fig. 1b), which suggested that the combined
use of six reference genes may be necessary for normal-
izing the expression of the interest gene.
Expression analysis of BcPME31 in eight samples
Pectin methylesterase (PME/PE) plays important roles in
pollen development and pollen tube growth (Tian et al.
2006). Reducing PME expression led to abnormal or
shrunken pollen, and lower pollen vigor in Chinese cab-
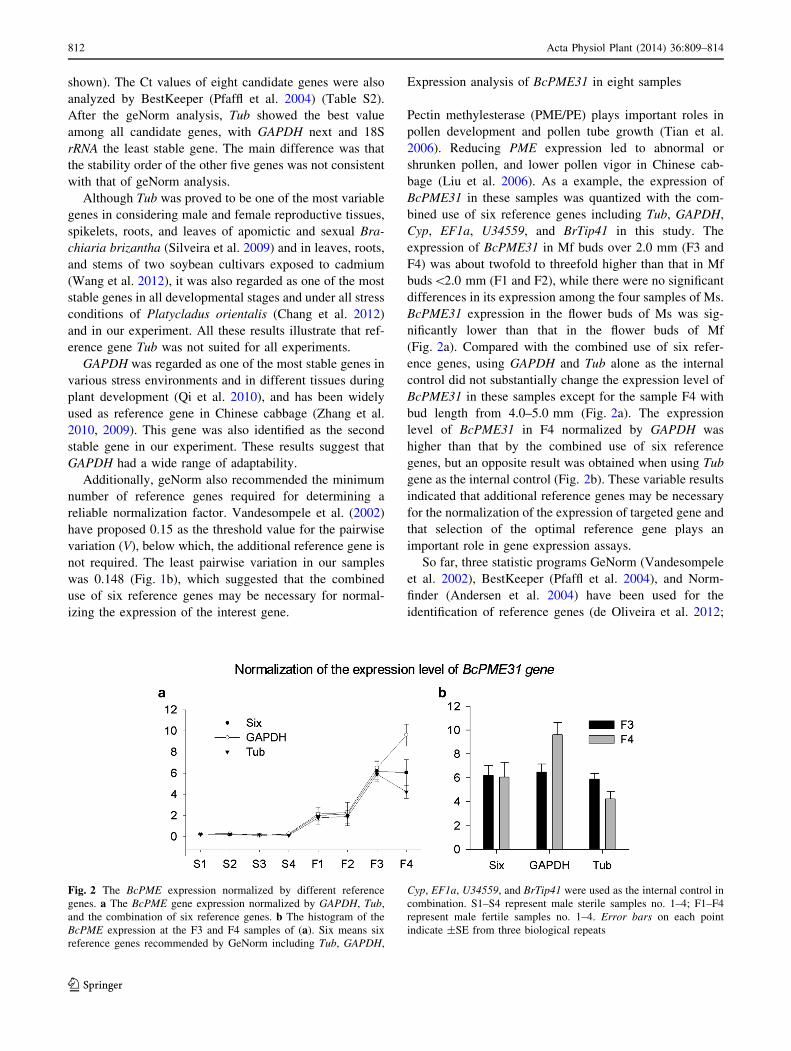
bage (Liu et al. 2006). As a example, the expression of
BcPME31 in these samples was quantized with the com-
bined use of six reference genes including Tub, GAPDH,
Cyp, EF1a, U34559, and BrTip41 in this study. The
expression of BcPME31 in Mf buds over 2.0 mm (F3 and
F4) was about twofold to threefold higher than that in Mf
buds\2.0 mm (F1 and F2), while there were no significant
differences in its expression among the four samples of Ms.
BcPME31 expression in the flower buds of Ms was sig-
nificantly lower than that in the flower buds of Mf
(Fig. 2a). Compared with the combined use of six refer-
ence genes, using GAPDH and Tub alone as the internal
control did not substantially change the expression level of
BcPME31 in these samples except for the sample F4 with
bud length from 4.0–5.0 mm (Fig. 2a). The expression
level of BcPME31 in F4 normalized by GAPDH was
higher than that by the combined use of six reference
genes, but an opposite result was obtained when using Tub
gene as the internal control (Fig. 2b). These variable results
indicated that additional reference genes may be necessary
for the normalization of the expression of targeted gene and
that selection of the optimal reference gene plays an
important role in gene expression assays.
So far, three statistic programs GeNorm (Vandesompele
et al. 2002), BestKeeper (Pfaffl et al. 2004), and Norm-
finder (Andersen et al. 2004) have been used for the
identification of reference genes (de Oliveira et al. 2012;
Fig. 2 The BcPME expression normalized by different reference
genes. a The BcPME gene expression normalized by GAPDH, Tub,
and the combination of six reference genes. b The histogram of the
BcPME expression at the F3 and F4 samples of (a). Six means six
reference genes recommended by GeNorm including Tub, GAPDH,
Cyp, EF1a, U34559, and BrTip41 were used as the internal control in
combination. S1–S4 represent male sterile samples no. 1–4; F1–F4
represent male fertile samples no. 1–4. Error bars on each point
indicate ±SE from three biological repeats
812 Acta Physiol Plant (2014) 36:809–814
123

Han et al. 2010; Mallona et al. 2010; Wan et al. 2010). Of
these, GeNorm was widely used for its additional function
regarding the minimum number of reference genes
required for determining a reliable normalization factor. In
this study, GeNorm was employed to compare eight can-
didate reference genes in Ms and Mf plants of heading
Chinese cabbage during flower bud development. The
statistical result revealed that the stability order was Tub/
GAP-
DH [ Cyp [ EF1a [ U34559 [ BrTip41 [ Apr [ 18S
rRNA, and the combined use of six reference genes was
optimal for qRT-PCR. Furthermore, the expressions of
BcPME31 normalized with the combined use of six refer-
ence genes and with GAPDH or Tub alone were compared;
the results also suggested that the selection of the optimal
reference genes was necessary in gene expression-related
research.
Author contribution For this paper, Zhiyuan Fang and
Lugang Zhang designed the study, gave direction, and
provided research facilities. Xiaoyong Xu performed most
of the experiments and wrote the manuscript. Zeping Yang
helped with statistics analysis. Xilu Su helped with qRT-
PCR test. All authors reviewed and commented on the final
manuscript. The authors declare that no conflict of interest
exists.
Acknowledgments This work was supported by grants from the
National Natural Science Foundation of China (No. 30871717) and
the National Science & Technology Pillar Program
(2012BAD02B01).
References
Alves-Ferreira M, Wellmer F, Banhara A, Kumar V, Riechmann JL,
Meyerowitz EM (2007) Global expression profiling applied to
the analysis of arabidopsis stamen development. Plant Physiol
145(3):747–762. doi:10.1104/pp.107.104422
Andersen CL, Jensen JL, Orntoft TF (2004) Normalization of real-
time quantitative reverse transcription-PCR data: a model-based
variance estimation approach to identify genes suited for
normalization, applied to bladder and colon cancer data sets.
Cancer Res 64(15):5245–5250. doi:10.1158/0008-5472.can-04-
0496
Chang E, Shi S, Liu J, Cheng T, Xue L, Yang X, Yang W, Lan Q,
Jiang Z (2012) Selection of reference genes for quantitative gene
expression studies in Platycladus orientalis (Cupressaceae)
using real-time PCR. PLoS One 7(3):e33278. doi:10.1371/
journal.pone.0033278
Chen X, Truksa M, Shah S, Weselake RJ (2010) A survey of
quantitative real-time polymerase chain reaction internal refer-
ence genes for expression studies in Brassica napus. Anal
Biochem 405(1):138–140. doi:10.1016/j.ab.2010.05.032
Czechowski T, Stitt M, Altmann T, Udvardi MK, Scheible W-R
(2005) Genome-wide identification and testing of superior
reference genes for transcript normalization in Arabidopsis.
Plant Physiol 139(1):5–17. doi:10.1104/pp.105.063743
de Oliveira LA, Breton MC, Bastolla FM, Camargo Sda S, Margis R,
Frazzon J, Pasquali G (2012) Reference genes for the normal-
ization of gene expression in eucalyptus species. Plant Cell
Physiol 53(2):405–422. doi:10.1093/pcp/pcr187
Dong M, Zhang X, Chi X, Mou S, Xu J, Xu D, Wang W, Ye N (2012)
The validity of a reference gene is highly dependent on the
experimental conditions in green alga Ulva linza. Curr Genet
58(1):13–20. doi:10.1007/s00294-011-0361-3
Gu C, Chen S, Liu Z, Shan H, Luo H, Guan Z, Chen F (2011)
Reference gene selection for quantitative real-time PCR in
Chrysanthemum subjected to biotic and abiotic stress. Mol
Biotechnol 49(2):192–197
Gutierrez L, Mauriat M, Guenin S, Pelloux J, Lefebvre JF, Louvet R,
Rusterucci C, Moritz T, Guerineau F, Bellini C, Van Wuyt-
swinkel O (2008) The lack of a systematic validation of
reference genes: a serious pitfall undervalued in reverse
transcription-polymerase chain reaction (RT-PCR) analysis in
plants. Plant Biotechnol J 6(6):609–618. doi:10.1111/j.1467-
7652.2008.00346.x
Han LQ, Yang GY, Zhu HS, Wang YY, Wang LF, Guo YJ, Lu WF,
Li HJ, Wang YL (2010) Selection and use of reference genes in
mouse mammary glands. Genet Mol Res 9(1):449–456
Liu LC, Yu XL, Ye WZ, Xiang X, Cao JS (2006) Functional
confirmation of BcMF3 gene related to male sterility of Chinese
cabbage by antisense RNA. J Zhejiang Univ (Agric Life Sci)
32(05):473–478 (in Chinese)
Mallona I, Lischewski S, Weiss J, Hause B, Egea-Cortines M (2010)
Validation of reference genes for quantitative real-time PCR
during leaf and flower development in Petunia hybrida. BMC
Plant Biol 10:4. doi:10.1186/1471-2229-10-4
Pfaffl MW, Tichopad A, Prgomet C, Neuvians TP (2004) Determi-
nation of stable housekeeping genes, differentially regulated
target genes and sample integrity: BestKeeper—Excel-based tool
using pair-wise correlations. Biotechnol Lett 26(6):509–515.
doi:10.1023/b:bile.0000019559.84305.47
Qi JN, Yu SC, Zhang FL, Shen XQ, Zhao XY, Yu YJ, Zhang DS
(2010) Reference gene selection for real-time quantitative
polymerase chain reaction of mRNA transcript levels in Chinese
Cabbage (Brassica rapa L. ssp pekinensis). Plant Mol Biol
Report 28(4):597–604. doi:10.1007/s11105-010-0185-1
Ramakers C, Ruijter JM, Deprez RH, Moorman AF (2003) Assump-
tion-free analysis of quantitative real-time polymerase chain
reaction (PCR) data. Neurosci Lett 339(1):62–66
Silveira ED, Alves-Ferreira M, Guimaraes LA, da Silva FR, Carneiro
VT (2009) Selection of reference genes for quantitative real-time
PCR expression studies in the apomictic and sexual grass
Brachiaria brizantha. BMC Plant Biol 9(84):1471–2229
Tian GW, Chen MH, Zaltsman A, Citovsky V (2006) Pollen-specific
pectin methylesterase involved in pollen tube growth. Dev Biol
294(1):83–91. doi:10.1016/j.ydbio.2006.02.026
Tong Z, Gao Z, Wang F, Zhou J, Zhang Z (2009) Selection of reliable
reference genes for gene expression studies in peach using real-
time PCR. BMC Mol Biol 10(71):1471–2199
Vandesompele J, De Preter K, Pattyn F, Poppe B, Van Roy N, De
Paepe A, Speleman F (2002) Accurate normalization of real-time
quantitative RT-PCR data by geometric averaging of multiple
internal control genes. Genome Biol 3(7):research0034.1–
research0034.11. doi:10.1186/gb-2002-3-7-research0034
Wan H, Zhao Z, Qian C, Sui Y, Malik AA, Chen J (2010) Selection of
appropriate reference genes for gene expression studies by
quantitative real-time polymerase chain reaction in cucumber.
Anal Biochem 399(2):257–261. doi:10.1016/j.ab.2009.12.008
Wang Y, Yu K, Poysa V, Shi C, Zhou Y (2012) Selection of reference
genes for normalization of qRT-PCR analysis of differentially
expressed genes in soybean exposed to cadmium. Mol Biol Rep
39(2):1585–1594
Acta Physiol Plant (2014) 36:809–814 813
123

Xu X, Sun X, Zhang J, Huang W, Zhang L, Fang Z (2013) Identification
of candidate genes associated with male sterility in CMS7311 of
heading Chinese cabbage (Brassica campestris L. ssp. pekinensis).
Acta Physiologiae Plantarum 35(11):3265–3270. doi:10.1007/
s11738-013-1346-2
Zhang JY, Li Y, Shi GJ, Chen XF, Wang JJ, Hou XL (2009)
Characterization of alpha-tubulin gene distinctively presented in
a cytoplasmic male sterile and its maintainer line of non-heading
Chinese cabbage. J Sci Food Agric 89(2):274–280. doi:10.1002/
Jsfa.3438
Zhang CW, Qi L, Hou XL, Shi GJ, Zhang JY (2010) Differential gene
expression analysis of a new Ogura CMS line and its maintainer
in non-heading Chinese cabbage by cDNA-AFLP. Acta Phys-
iologiae Plantarum 32(4):781–787. doi:10.1007/s11738-010-
0463-4
814 Acta Physiol Plant (2014) 36:809–814
123