russell (2006) tubercle
TRANSCRIPT

7/30/2019 Russell (2006) Tubercle
http://slidepdf.com/reader/full/russell-2006-tubercle 1/9
The bacterium Mycobacterium tuberculosis is the causa-tive agent of one of those human conditions that, despitethe developments of modern medicine, is reluctant torelinquish its grip on mankind. The incidence of tuber-culosis (TB) correlates strongly with reduced socio-eco-nomic status, thereby adding to the burden of the globalpoor1,2. Estimates of host penetrance based on skin-testconversion indicate that up to one-third of the world’spopulation are infected with this bacterium. Over theirlifetime it is projected that an infected individual has a5–10% risk of developing TB3, although this scenarioshows extraordinary regional variation that is impactedto a tragic extent by HIV infection4. Given that such arelatively low number of individuals develop disease andtherefore the capacity to transmit the infection, it fol-lows that the transmission process must be supremely efficient to sustain such levels of infection in its hostpopulation.
The progression of the disease is determined at thelevel of the infection site itself. Once the bacterium hasgained entry into a macrophage and triggered that hostcell to invade the tissue of the lung, the host respondsby remodelling the site of infection into a cellular mass,
the ‘tubercle’ or granuloma that has given the disease itsname. The human tuberculosis granuloma is a beautifuland fascinating structure in which all the acts of the dis-ease are written and performed, as documented in BOX 1.The bulk of the literature on the tuberculosis granulomafocuses on the host component and ignores any influ-ence that the bacterium might have on its progression.However, given that transmission is key to the successof the bacterium, it would be short-sighted to imaginethat M. tuberculosis has not evolved specific strategiesand effectors to maximize progression to a productiveinfection. This Review attempts to redress the balancein favour of M. tuberculosis.
What is now ancient historyTo appreciate the biological significance of the granu-loma it is important to consider tuberculosis in thecontext of human evolution rather than focusing exclu-sively on the current manifestations of the disease. How has TB impacted on mankind through pre-history?Until recently, it was thought that M. tubercu losis evolved from Mycobacterium bovis and was acquiredby humans during the development of agriculture inthe Fertile Crescent around 9,000 years ago5, a hypoth-esis popularized in the book Guns, Germs and Steel by Jared Diamond6. In this scenario, TB would have beenintroduced into a subpopulation of mankind after theiremergence from Africa and one would have predicted,unless other infectious diseases provided comparableselective pressures on fitness and reproduction, thatthe current human populations would exhibit extremetraits of susceptibility However, this is not the case— with the exception of a few extremely rare mutationsin central immune system function7, the populationgenetics of susceptibility to TB reveal relatively mod-est associations of individual genotypes with diseaseprogression7–13.
More recent studies into the evolution of M. tubercu-losis have indicated that M. tuberculosis is actually moreclosely related to a common founder strain than it is to M. bovis. This implies that either humans infected cat-tle with M. tuberculosis, resulting in the divergence of M. bovis , or the two strains evolved in parallel from afounder strain that infected both humans and cattle14–16.The ancestor of M. tuberculosis most likely arose fromthe M. tuberculosis-like species of the M. tuberculosis complex that are found today in central Africa14–17. Thismight indicate that humans were exposed to tuberculo-sis-mediated selection pressure much earlier, and morecomprehensively, than was previously assumed.
Department of Microbiology
and Immunology, College of
Veterinary Medicine, Cornell
University, Ithaca, New York
14853, USA.
e-mail: [email protected]
doi:10.1038/nrmicro1538
Published online
11 December 2006
Founder strain
The ancestral species or strain
that underwent divergent
evolution to produce several
new species or strains.
Who puts the tubercle in tuberculosis?David G. Russell
Abstract | Tuberculosis (TB), an illness that mainly affects the respiratory system, is one of the
world’s most pernicious diseases. TB currently infects one-third of the world’s population and
kills approximately 1.7 million people each year. Most infected individuals fail to progress to
full-blown disease because the TB bacilli are ‘walled off’ by the immune system inside a tissue
nodule known as a granuloma. The granuloma’s primary function is one of containment and
it prevents the dissemination of the mycobacteria. But what is the role of the TB bacillus in
the progression of the granuloma? This Review explores how Mycobacterium tuberculosis
influences granuloma formation and maintenance, and ensures the spread of the disease.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 5 | JANUARY 2007 | 39
7/30/2019 Russell (2006) Tubercle
http://slidepdf.com/reader/full/russell-2006-tubercle 2/9
Airway
Free mycobacteria
Necrotic, caseatinggranuloma centre
Granuloma
Macrophage
LymphocytesLymphocyte
Infected macrophage
Airway
Mycobacterium
Blood vessel
Infected alveolarmacrophages
Mononuclearcells
Alveolarmacrophage
Foamy macrophages
Fibrous cuff Macrophages
Foamy macrophages
Bloodvessels
Although M. tuberculosis can cause primary dis-ease, most notably in neonates, the most commonmanifestation of TB in adults is the reactivation of a pre-existing, chronic infection. Reactivation isfrequently triggered by conditions that compromiseimmune surveillance, including AIDS, poor nutrition,old age and stress, all conditions that would seem toimpact primarily on a post-breeding-age population.But this is a modern interpretation and it is likely that,under the more extreme living conditions to whichprehistoric man was exposed, progression to full-blown, primary disease was more common and would
have impacted on human reproduction. If the humanpopulation ran the gauntlet of such selection pressureen masse as it emerged from Africa, this could havecontributed to the relatively low incidence of markedTB susceptibility traits.
Although reactivation and disease progression areusually thought of as a ‘failure’ of the immune system,this is an extremely humanocentric interpretation thatdenies the key position that transmission occupies in thelife cycle of M. tuberculosis. This article examines theactive role of M. tuberculosis in the biology of the TBgranuloma.
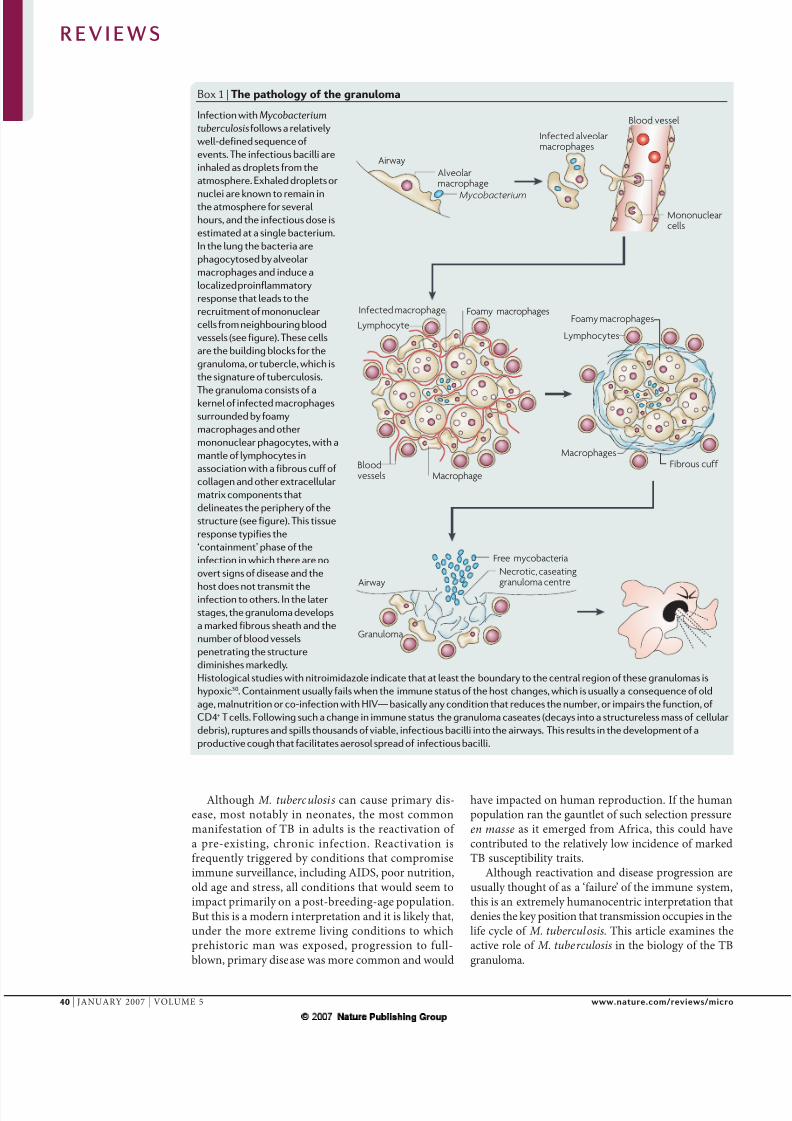
Box 1 | The pathology of the granuloma
Infection with Mycobacterium
tuberculosisfollows a relatively
well-defined sequence of
events. The infectious bacilli are
inhaled as droplets from the
atmosphere. Exhaled droplets or
nuclei are known to remain inthe atmosphere for several
hours, and the infectious dose is
estimated at a single bacterium.
In the lung the bacteria are
phagocytosed by alveolar
macrophages and induce a
localized proinflammatory
response that leads to the
recruitment of mononuclear
cells from neighbouring blood
vessels (see figure). These cells
are the building blocks for the
granuloma, or tubercle, which is
the signature of tuberculosis.
The granuloma consists of akernel of infected macrophages
surrounded by foamy
macrophages and other
mononuclear phagocytes, with a
mantle of lymphocytes in
association with a fibrous cuff of
collagen and other extracellular
matrix components that
delineates the periphery of the
structure (see figure). This tissue
response typifies the
‘containment’ phase of the
infection in which there are no
overt signs of disease and the
host does not transmit theinfection to others. In the later
stages, the granuloma develops
a marked fibrous sheath and the
number of blood vessels
penetrating the structure
diminishes markedly.
Histological studies with nitroimidazole indicate that at least the boundary to the central region of these granulomas is
hypoxic30. Containment usually fails when the immune status of the host changes, which is usually a consequence of old
age, malnutrition or co-infection with HIV— basically any condition that reduces the number, or impairs the function, of
CD4+ T cells. Following such a change in immune status the granuloma caseates (decays into a structureless mass of cellular
debris), ruptures and spills thousands of viable, infectious bacilli into the airways. This results in the development of a
productive cough that facilitates aerosol spread of infectious bacilli.
R E V I E W S
40 | JANUARY 2007 | VOLUME 5 www.nature.com/reviews/micro

7/30/2019 Russell (2006) Tubercle
http://slidepdf.com/reader/full/russell-2006-tubercle 3/9
Infectedmacrophage
Uninfected macrophages
TNF-α, CCL2, CXCL10, IL1-α, IL1-β, ILI8
TNF-α, CCL2, CXCL10, CXCL9,CCL5, CCL3, CCL4
TNF-α(acts as a
positive
feedbackloop)
The cytokine/chemokine storm
ExpandsTNF
production
Downregulatesproinflammatoryresponse
IFN-γ
Lymphocytes(CD4+, CD8+ and T cells andB cells)
Neutrophils
Caseation
The process by which a
tuberculous granuloma decays
into a structureless mass of
cellular debris.
Chemokine
Cytokines involved in specific
inflammatory responses. They
are differentiated into CC or
CXC chemokines on the basis
of their primary sequence.
Natural killer (NK) T cellAn NK T cell is a T cell that
expresses some NK cell
receptors and has some NK-
cell-like functions. They also
express a T-cell receptor that
recognizes CD1b (which binds
glycolipids not peptides).
CD4+ cell
A subpopulation of T cells that
express the CD4 receptor and
respond to antigens presented
on the surface of host cells that
bear major histocompatibility
complex class II molecules. Two
distinct subsets of activated
CD4+ T cells have been
described. T-helper 1 (TH1) cells
produce interferon γ , tumour-
necrosis factor α and interleukin
(IL-)12, and support cell-
mediated immunity. TH2 cells
produce IL-4, IL-5 and IL-13,
support humoral immunity, and
downregulate TH1 responses.
CD8+ cell
A subpopulation of T cells that
express the CD8 receptor.
CD8+ cells recognize antigens
that are presented on the
surface of host cells by major
histocompatibility complexclass I molecules, leading to
their destruction, and are
therefore also known as
cytotoxic T cells.
Cytokine
Member of a large family of
secreted proteins that bind
immune cells through specific
receptors. Cytokine production
results in the activation of an
intracellular-signalling cascade
that commonly regulates
processes such as immune
function and inflammation.
The development of the TB granulomaMost experimental infections conducted with M. tuber-culosis use the murine host. The mouse is a useful toolfor revealing the bacterial factors that are important inthe infection process, and for probing the antigenic andcellular basis of a protective immune response. However,it is seriously compromised as a model for granulomaformation and progression. Mice do not build the highly stratified structures that are observed in humans anddo not normally exhibit necrosis and caseation, whichare significant factors leading to transmission (BOX1).Retrospective histological and molecular studies of TBinfection in humans have yielded invaluable insightsinto the progression of the disease, especially wheninterpreted in the context of in vitro murine macrophagestudies. In vitro analyses of the responses of murine andhuman macrophages to M. tuberculosis infection indicatethat the cells produce a robust proinflammatory responsethrough the activity of Toll-like receptor (TLR) agonists(stimulators of the host’s TLRs) that are abundant on
the surface of the bacteria. It is widely postulated thatalveolar macrophages in the airways, following inter-nalization of inhaled bacteria, are stimulated to invadethe lung epithelium18–20. Production of tumour necrosisfactor (TNF)-α and inflammatory chemokines from theinfected macrophages drives the recruitment of succes-sive waves of neutrophils, natural killer (NK) T cells, CD4+
T cells and CD8+ T cells, each of which produce their owncomplement of chemokines and cytokines that amplify cellular recruitment and remodelling of the infectionsite18,19,21. This inflammatory cascade (BOX 2) is regulatedand superceded by a specific, cellular immune responsethat is linked to the production of interferon (IFN)-γ. At
this stage, the formation of the ‘stable’ granuloma that isresponsible for immune containment during the latent,or subclinical, period of the infection becomes recogniz-able and the stratification of the structure emerges22–26.These studies have been invaluable in defining the overalltrends observed in infected tissue. This, however, is thetraditional view of granuloma progression and its tenetsare being questioned by temporal studies in macaquemonkeys that indicate a more heterogeneous picture inwhich caseation is observed even in early lesions, andthere is marked heterogeneity between the granulomaspresent in a single host27.
More mature-phase granulomas show marked neo-
vascularization and develop an extensive fibrotic capsulethat delineates the margin between the macrophages,granulocytes, foamy macrophages and giant cells, and thelymphocytic infiltrate18,25,26. In the late stage, the centre of the granuloma loses its vascular appearance and becomesnecrotic. In a progressive lesion (BOX 1), the necrosis pre-cedes and probably facilitates caseation, the granuloma
wall breaks down and the bacteria are released into theairways, resulting in transmission. Transmission seemsto coincide with high levels of matrix metalloproteinase9 (MMP9) in the serum, implying that tissue breakdownis an active, destructive process.
Recent histological analyses of human granulomashave been particularly informative in understandingthe biology of these later structures18,22,25. The analysis of active cavitary lesions, compared with ‘non-progressive’tuberculomas, indicated that the non-progressive lesionswere more highly vascularized, which is consistent bothwith the vascular epithelial growth factor (VEGF) pro-duction that is reported in activated macrophages28,29,
Box 2 | The cytokine/chemokine storm
The recruitment of successive waves of cells to the
forming granuloma is driven by a bewildering array
of cytokines and chemokines that is initiated by the
infected macrophage. Tumour necrosis factor (TNF)-α is
undoubtedly the dominant cytokine in this response and
it elevates the production of the chemokines that mediate
recruitment (see figure). Infected macrophages produceTNF-α, and the chemokines CCL2 and CXCL10. In
experimental bead granulomas there is a transient
recruitment of neutrophils before the sustained migration
of macrophages but it is unclear if this occurs in the
natural infection where the insult is less. Analysis of
broncholavage fluid and serum from infected individuals
indicates that there is the production of a barrage of
chemokines, including CCL2, CXCL10, CCL3,4,5 and
CXCL9. Finally, in vivo studies of both infected mice and
macaque monkeys have revealed extensive upregulation
of the CXCR3-binding cytokines CXCL9, CXCL10 and
CXCL11 (REFS 92,93). These chemokines lead to the
recruitment of lymphocytes including first natural
killer (NK) T cells, then CD4+, CD8+ and γδ T cells and
B cells. The quietening down of the response coincideswith the production of interferon (IFN)-γ (see figure). It is
interesting to note that IFN-γ -deficient mice are actually
unable to turn off this proinflammatory cascade and suffer
fatal pathology in their lungs94.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 5 | JANUARY 2007 | 41

7/30/2019 Russell (2006) Tubercle
http://slidepdf.com/reader/full/russell-2006-tubercle 4/9
NeovascularizationThe formation of new blood
vessels in a developing tissue.
This process is stimulated by
the production of vascular
endothelial growth factor. The
term is used most frequently
in cancer biology in which the
tumour develops its own
blood supply through
neovascularization.
Foamy macrophage
A macrophage loaded with
lipid droplets. Such cells are
often observed in tissues with
chronic proinflammatory
stimulus.
Giant cell
A giant, multinucleate
macrophage.
Tuberculoma
The tuberculoma is the
granuloma that is formed
during tuberculosis infection.
This term is most frequently
used by clinicians and has
replaced the more traditional
‘tubercle’.
Phagosome
A membrane-boundcytoplasmic vacuole formed
around a particle ingested by
phagocytosis.
Homotypic fusion
The fusion of identical
compartments or vesicles.
Autophagy
A pathway for the recycling of
cellular contents, in which
materials inside the cell are
packaged into vesicles and are
then targeted to the vacuole or
lysosome for bulk turnover.
and the accepted notion that progression to caseationoccurs in a low-oxygen environment30. IFN-γ -producingT cells were found in both progressive and non-progres-sive granulomas but were located predominantly outsidethe necrotic centre and the fibrous wall.
One of the more intriguing observations in thesestudies was the relative distribution of bacteria andbacterial products. Several studies noted that, althoughbacteria were found in the central, necrotic region of the lesion, a significant portion of bacteria (or bacte-rial antigens) were associated with macrophages in theperipheral leukocytic infiltrate. These macrophageshave been found both bordering the necrotic regionand outside the fibrotic capsule. These regions are thesites of most lymphocyte proliferation. Intriguingly, thebacteria found in these peripheral macrophages werepositive for the expression of ICL1 (isocitrate lyase)23,31,which is known to be upregulated by M. tuberculosis inmacrophages that are exposed to activating cytokines32.These observations indicate that the interactions at theperiphery of the granuloma are dynamic.
The human tuberculosis granuloma is the productof a robust cellular immune response to bacterial com-ponents; it is not by chance that the dead mycobacteriaare used as the active ingredient in Freund’s completeadjuvant. In AIDS patients, the diminished capacity tomount a CD4+ T-cell response correlates with reducedgranuloma-forming capacity and, significantly, areduced ability to prevent metastasis of infection. Mostmonographs detailing granuloma structure and func-tion deal with this immune-mediated containment of infection and tend to treat transmission as a breakdownin immune function, but the bacterium has a vestedinterest in driving transmission and its capacity to divertthe host’s response must be considered in greater depth.In short, how does M. tuberculosis influence this processto maximize its survival and subsequent transmissionunder strong immune pressure?
Survival inside macrophagesPathogenic mycobacterial species survive inside mac-rophages by arresting the normal maturation of theirphagosome, thereby restricting its acidification to pH 6.4and limiting fusion with pre-formed lysosomes33–35. Thecharacteristics of this compartment and its intersectionwith cellular membrane trafficking pathways are illus-trated in FIG. 1. The strategy adopted by M. tuberculosis limits the hostility of the intracellular environment.
However, the Mycobacterium-containing vacuole is notisolated or inert. It communicates with the extracellularenvironment by homotypic fusion with other compart-ments in the early recycling endosomal system36. Theaccessibility of the vacuole was demonstrated by therapidity with which the vacuole acquired GM1 ganglio-side from the macrophage plasmalemma37. These datawere all generated in infected macrophages in culture.Although it still falls short of a true in vivo analysis,it has been shown that M. tuberculosi s-containing
vacuoles in infected macrophages isolated directly frombroncholavage fluid from TB patients fail to acidify anddo not acquire lysosomal cargo38.
Modulation of the phagosome seems to be mediatedby both cell-wall lipids and other bacterial effectors33,35.Mutants that are partially defective in phagosomemodulation reside in vacuoles of pH 5.8, which arrestsbacterial growth39. In addition, immune activation of themacrophage enables it to overcome the influence of thebacterium on phagosome maturation and deliver it toa more acidic (pH 5.2) bactericidal environment. Thefusion of the Mycobacterium-containing phagosomewith lysosomes is achieved through the activity of theIFN-inducible p47 GTPase family 40–42. These data imply that the growth status of the bacteria can be impactedby the immune modulation of their host macrophagesto induce either growth arrest or death.
Given the differential distribution of intracellular andextracellular bacteria observed in human granulomas itis interesting to speculate on which bacteria are capableof replication: those in the necrotic core that are thoughtto be hypoxic, or those at the periphery in macrophagesthat are accessible to the activating cytokines producedby proliferating lymphocytes. Extreme activation of host
macrophages in vitro by cytokines or through the induc-tion of autophagy will result in bacterial death. However,in vivo, it is more likely that the difference betweenactivated and resting macrophages is a continuum thatdetermines bacterial growth rates rather than mediatingclearance. This conclusion is supported by two studiesthat examined the stability of the bacterial populationin infected mice and failed to detect an accumulation of bacterial debris, at both the histological and molecularlevels43,44. Because the relative number of genome equiva-lents was stable and no accumulation of bacterial ‘skel-etons’ was observed, both studies concluded that therewas a relatively stable population of bacteria, probably non-replicative, during chronic infection.
Beyond phagocytesThe development of the granuloma is clearly mediated by different populations of host cells and is absolutely depend-ent on the presence of a robust cellular immune responsedirected against bacterial antigens. However, it remainsin the best interests of the bacterium to manipulate thisresponse locally to ensure persistence and, ultimately, latestage damage and transmission. There is a rich literaturedating back to the 1960s that identified the potency of arange of mycobacterial cell-wall lipids in the manipula-tion of immune response, adjuvanticity and the inductionof granulomas45–54 (and therefore their use in Freund’s
complete and Ribi adjuvant). Despite this fascinating setof early observations, our appreciation of how bacterialcomponents might function as effectors that promote thesuccess of the bacterium are only now emerging. Previousstudies revealed that the M. tuberculosis cell-wall com-ponents lipoarabinomannan and arabinomannan werereleased inside infected cells and trafficked to compart-ments that lacked bacteria55. Further analysis of this proc-ess using M. bovis bacille Calmette–Guérin (BCG) cellsthat had their surface glycoconjugates tagged with TexasRed hydrazide revealed that the mycobacterial constituentswere associated with several intracellular organelles butaccumulated in the internal vesicles in lysosomes56 (FIG. 2).
R E V I E W S
42 | JANUARY 2007 | VOLUME 5 www.nature.com/reviews/micro

7/30/2019 Russell (2006) Tubercle
http://slidepdf.com/reader/full/russell-2006-tubercle 5/9
Transferrin
Early endosome
pH 6.4
pH 4.8
Recycling pathway
Early endosomewith transferrin
Transferrin
Mycobacterium
Lysosome
Time (min)
Mycobacterium
IgG-beads
p H
a
b
04.5
5.0
5.5
6.0
6.5
7.0
7.5
5 10 15 20 25 30
IgG-coated bead
Nucleus
The released bacterial constituents traffic through theinfected host cell and, significantly, were also present in anextracellular microvesicular fraction56,57. Further analysisof the exosomal vesicles revealed the presence of lysosomalmembrane proteins and lysosomal proteases, indicatingthat labelled mycobacterial cell-wall constituents werereleased from a lysosomal compartment. Ultrastructuralanalyses of infected cells showed that the bacterial lipidsaccumulated in multilamellar vesicles that were mor-phologically reminiscent of major histocompatibility
complex (MHC) class II-enriched compartments (MIIC)or multi-vesicular lysosomes (FIG. 2). These mycobacte-rial lipids could also be observed in uninfected bystandercells56. The release of these vesicles seemed to occurthrough a constitutive form of exocytosis57. Clearly, if these lipids possessed biological activities then theirtransfer to bystander cells would expand the bacteria’ssphere of influence beyond the immediate confines of the host cell. Subsequent studies have shown that similar
vesicles containing bacterial components can be gener-ated through the apoptotic death of infected macrophagesand dendritic cells58,59, and that bacterial lipids mightbe released and transferred between cells as complexeswith apolipoprotein60, although it is unclear if all thesepathways are functionally equivalent.
The bioactive lipids of M. tuberculosis
Analysis of the lipid species present in the exosomal frac-tion identified the main peripheral lipids from the bacte-rial cell wall56,61. This implies that release is passive, or atleast non-selective, possibly mediated by the surfactant
environment in the endosomal system. Characterizationof the lipids by thin-layer chromatography, differentialdye labelling electrospray ionization mass spectrometry and collisionally activated tandem mass spectrometry led to the determination that the main released lipidscorresponded to phosphatidylinositol mannoside species2A and 2B, monophosphatidylglycerol, diphosphatidyl-glycerol, phosphatidylethanolamine, trehalose mycolatesand the phenolic glycolipid mycoside B.
Mycobacterial cell-wall-associated proteins belong-ing to the Ag85 family were also detected in the exo-somes from infected macrophages, supporting the view that this is a mass-action effect62. Although it has notbeen documented in the same detail, it is likely that allreleased, hydrophobic components from the bacteriawill follow a similar pathway through, and out of, theinfected macrophage.
Antigens versus effectorsMany of these released bacterial components are knownimmunomodulators, but if they traffic with the releasedbacterial lipids they will aggregate in the multivesicularlysosome, or the MIIC, of the host cell. Whereas the M. tuberculosis-containing vacuole itself is sequesteredoutwith the antigen presenting machinery of the cell63,the bacterial components that are released are fodderfor the antigen-presenting machinery and become
antigens that could be recognized on the surface of theinfected cell.
Protein antigens secreted into the culture filtrate by growing bacteria in culture have been shown to includesome of the more immunogenic bacterial proteins, suchas early secreted antigenic target 6 (ESAT6) and culturefiltrate protein 10 (CFP10) (REFS 64–66), and several of the lipids can be presented by the CD1 family of MHCmolecules67–69. Furthermore, the antigen-loaded vesiclesfrom infected cells are capable of mediating cross-prim-ing58,59. These released antigens could therefore have arole in driving the proliferative response observed at theperiphery of the granuloma.
Figure 1 | Trafficking of Mycobacterium tuberculosis
bacilli in cells. Following phagocytosis, phagosomes
containing IgG-coated beads acidify rapidly to pH 5 (a)
and lower, and the inert particles are subsequently
delivered to the lysosome (b). By contrast, phagosomescontaining M. tuberculosis only acidify to pH 6.4 (a) and
fail to fuse with lysosomes. The vacuoles containing
M. tuberculosis retain many of the characteristic of the
early endosomal system (b). They are accessible to
recycling endosomes, as evidenced by their acquisition
of labelled transferrin when it is added to the external
medium. Components of the peripheral cell wall of the
bacteria that are shed into the bacteria-containing vacuoles
traffic out of the vacuoles and coalesce in dense, lysosomal
compartments. Finally, the bacteria-containing vacuoles,
in common with most endosomal/lysosomal stages, fuse
with delivery vesicles trafficking from the trans-Golgi
network of the host cell. The data imply that the vacuoles
in which M. tuberculosis reside are not particularly hostile
with respect to both pH and hydrolytic activity35
.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 5 | JANUARY 2007 | 43

7/30/2019 Russell (2006) Tubercle
http://slidepdf.com/reader/full/russell-2006-tubercle 6/9
Spread to bystander cells
Lipids released in exosomesfrom cell surface
Lipids are shed
Mycobacteriumtuberculosis
Nucleus
Lipids accumulatein multilamellar bodies Multilamellar bodies coalesce
in multivesicular lysosome
a
b
c
d
Fibrosis
Fibrosis is frequently seen at
sites of chronic inflammatory
stimulation. Cells lay down a
fibrinogen/fibrin skeleton that
is augmented with other
extracellular matrix proteins
like collagen.
Some lipids are more equal than others
The identification of mycobacterial cell-wall lipids thatare involved in the induction or control of the diseasepathology that promotes bacterial survival has pro-ceeded predominantly through direct analysis of the bio-logical activities of individual, purified bacterial cell-wallcomponents. So although the data documenting therelease and trafficking of bacterial lipids inside their hostcell are recent, there is a rich history of publications on thebiomodulatory activities of mycobacterial lipids.
In the 1960s, researchers became interested in thecapacity of defined bacterial cell-wall constituents toinduce granulomatous responses in mice52,53. Since theseseminal studies, several laboratories have explored theeffects of isolated bacterial cell-wall lipids on macrophages
and T cells in culture70,71, and in mice72–77. The relevanceof these data to the tuberculoma is difficult to extrapolatebecause the in vitro experiments were done using selectcell types (usually macrophages) and might lack key play-ers, whereas the in vivo experiments are complex and havelimited accessibility for analysis and manipulation.
Among the in vivo experiments are a few studies thathave localized the tissue response either through theuse of particulate delivery systems for bacterial compo-nents74,78,79 or restriction to a specific site, such as the useof the air-pouch model76; both of these methods facilitateanalysis at specific foci. Trehalose dimycolate (TDM)incorporated into oil droplets45,80 or on particles75,78,79 induces strong granulomatous reactions at sites of accu-mulation. More recently, lipid-coated particles that wereincorporated into an extracellular matrix gel were inocu-lated into mice to study the waves of cellular recruitmentand tissue remodelling induced by the bacterial lipids79.Fluorescently tagged lipids were internalized by mac-rophages and newly recruited inflammatory cells, andthese cells generated a proinflammatory response that
involved TNF-α, interleukin (IL)-1α, IL-6, IL-10, CCL2and IFN-γ -inducible protein-10 (CXCL10). Bacterial-lipid-bearing matrices recruited greater numbers of inflammatory cells than control matrices that containedphosphatidylglycerol or no lipid.
Histological characterization (FIG. 3) indicated thatleukocytes arrived by two routes, either adherence tothe surface and migration into freely floating matricesor extravasation directly from the circulation into newly
vascularized matrices. Leukocytes arrived in succes-sive waves: neutrophils, followed by macrophages andmature dendritic cells, and finally T cells, NK cells andB cells. This model aptly demonstrates the inflammatory properties of peripheral BCG cell-wall lipids, reproduc-ing cellular and cytokine responses that are typical of granulomas in Mycobacterium-infected tissues. In addi-tion the structure also reveals that many of the structuralcharacteristics of TB granulomas (a capsular structure,extensive fibrosis and neovascularization) were recapitu-lated in this artificial model79. The similarities imply thatthe bacterial lipids have a significant role in shaping thetissue response at the infection site.
The in vivo granuloma model was used to assay the bioactivity of the individual lipid species withrespect to cell recruitment and c ytokine induction.BCG-derived cell-wall lipids were f ractionated andthe isolated fractions, including phosphatidylinositol
dimannosides, cardiolipin, phosphatidylglycerol, phos-phatidylethanolamine, trehalose monomycolate, TDMand mycocide B were assayed for activity 61,78. Trehalosemono- and dimycolates were found to induce profoundrecruitment of a range of immune effector cells in vivo,indicating that the trehalose mycolates, particularly TDM, are the most bioactive lipids in the mycobacte-rial cell-wall fraction78. The potency of the responseto TDM in comparison to other mycobacterial lipidsis clear even at the histological level. These data areconsistent with the rich body of literature document-ing the bioactivity of trehalose mycolates both in vitro and in vivo51,61,71,72,81–83.
Figure 2 | Release and trafficking of bacterial lipids. Mycobacterium tuberculosis
sheds its peripheral cell-wall components into its vacuole. These lipids accumulate
in multilamellar bodies (a) in mixed micellar structures that contain both host- and
pathogen-derived components. The multilamellar bodies coalesce in the
multivesicular lysosome, known as the major histocompatibility complex class
II-enriched compartment in antigen-presenting cells (b). The vesicles gain egress
from the infected macrophage by exocytosis and are released as exosomes into the
external milieu (c). The exosomes, carrying pathogen-derived lipids and proteins,
are internalized by neighbouring cells (d)56,57,62.
R E V I E W S
44 | JANUARY 2007 | VOLUME 5 www.nature.com/reviews/micro
7/30/2019 Russell (2006) Tubercle
http://slidepdf.com/reader/full/russell-2006-tubercle 7/9
Matrigel matrix
Lymphocyte cuffing Cells around the beads
Bloodvessels
Fibrouslayer
Polystyrenebead coatedwith TDM
Fat cells at base of peritoneal cavity
Neovascularization
Signature-tagged
mutagenesis
A technique to screen large
numbers of distinct mutants for
those that fail to survive an
animal infection. Each mutant
is tagged with a unique DNA
sequence (called a signature
tag), which allows a specific
mutant to be tracked within a
large pool of bacteria.
Genetic screens for cell-wall lipid functionsThe use of genetics to probe the biological activity of the cell-wall lipids is problematic, owing to the restrictednumber of mutants available because the major classes of lipids seem to be essential. However, three lipid modu-lators have been analysed by mutational approaches,including phthiocerol dimycocerosate (PDIM), TDM
and a phenolic glycolipid. Although the infection studieshave been carried out predominantly in mice, these dataare probably significant to the regulation of pathology atthe infection site in humans.
The activity of TDM in modulating the host inflam-matory response has been confirmed and extended by mutational studies that revealed that TDM is modifiedby an unusual cyclopropane addition. Mutants defectivein either the production of cyclopropane or the additionof cyclopropane to TDM have a markedly enhancedinflammatory response at a comparable bacterial load inmurine infection experiments84–86. This in vivo pathology translated into a five-fold increase in the level of TNF-α
released by infected macrophages in vitro. These resultsare intriguing because they imply that the pathologicalproperties of the lipid have been ‘designed’ to mediate alevel of damage appropriate to the survival and persist-ence strategies of the bacteria.
Signature-tagged mutagenesis screens were used toidentify loci involved in the synthesis and export of PDIM, which resulted in a growth defect in macrophagesand in mice87–89. Interestingly, the growth defect of themutated M. tuberculosis strains in mice seemed to betissue specific, and was restricted to the lung whereasbacterial loads in the liver and spleen were unaffected.The authors hypothesized that the lipid had a role inregulating the immune response within the infected tis-sue site89, although more work is needed to elucidate thetissue-dependency of the phenotype.
Finally, it has long been known that the W-Beijingfamily of human TB isolates are particularly virulent inhumans, and induce extreme pathology at a relatively low bacterial load in mice. Analysis of bacterial gly-colipids in mice and rabbit models identified a phenolic
glycolipid, similar to PDIM, that was unique to theseW-Beijing strains90,91. Deletion of genes in a locus thatencodes the machinery for polyketide and PDIM syn-thesis led to loss of this lipid and a marked reduction inthe hypervirulence phenotype. Analysis of the responseof macrophages to purified phenolic glycolipid indi-cated that the lipid suppressed their proinflammatory response, resulting in depressed production of TNF-α,IL-6, IL-12 and CCL2. How this translates into virulencein vivo remains to be determined but the data recipro-cate the theme that virulence — and pathology — mustbe regulated to yield a pathogen capable of chronicity,reactivation and transmission.
Clearly, a genetic route in generating bacterialmutants with structurally defined defects in theircell-wall constituents is attractive. The data emergingfrom such studies are tantalizing but they underlinethe complexity of the interaction and emphasize thedivide between macrophage experiments in vitro and the results of in vivo challenge in a multicellularenvironment.
TransmissionThis Review has attempted to generate a picture of a strong host immune response that provides thebuilding blocks that form the granulomatous tis-sue at the infection site while emphasizing that the
arrangement of these blocks is clearly influenced by effectors released by M. tuberculosis . It is relatively easy to envisage how the inflammatory cell-wallcomponents of M. tuberculosis can aggravate its hostinto a strong proinflammatory response that causesthe tissue to compartmentalize this persistent irritantbehind a fibrous capsule. Artificial models with iso-lated lipids arrayed on particles reciprocate many of the characteristics of true tubercles during their early stages of development. The real challenge, however, isto understand what the bacterium can do to drive thelate-stage damage that leads to transmission. TDM isan interesting candidate for such a mediator. It seems
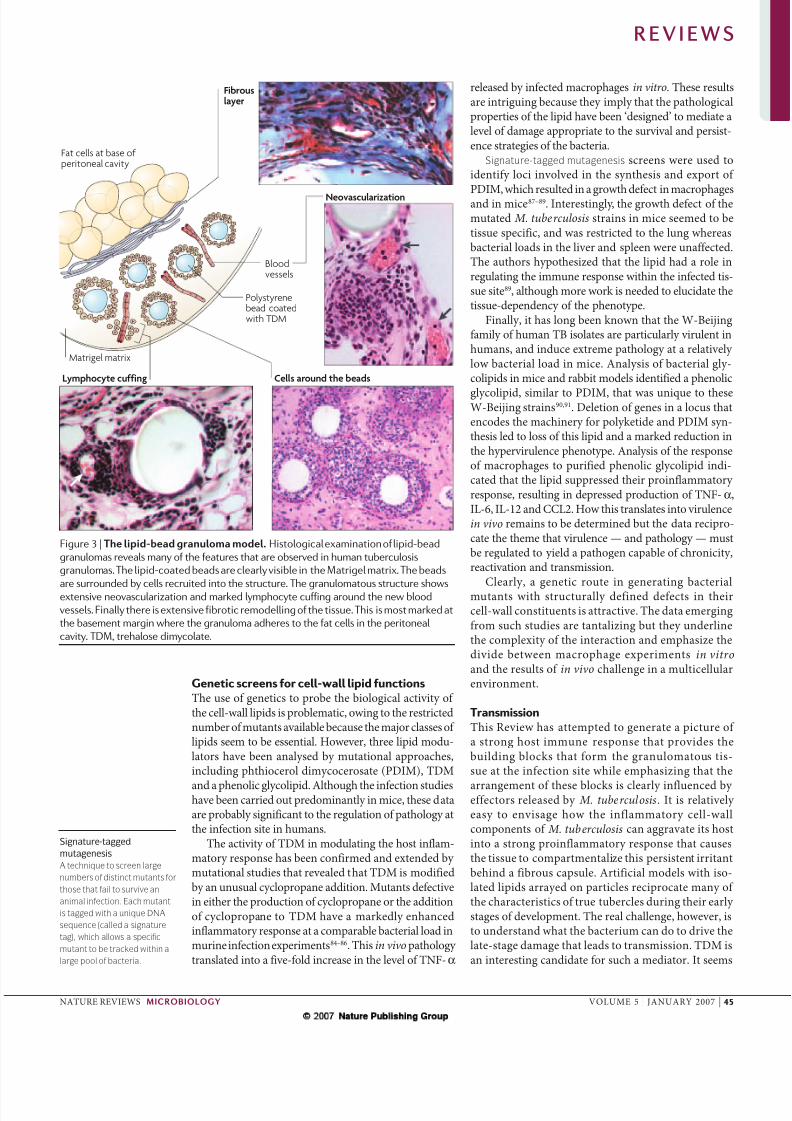
Figure 3 | The lipid-bead granuloma model. Histological examination of lipid-bead
granulomas reveals many of the features that are observed in human tuberculosis
granulomas. The lipid-coated beads are clearly visible in the Matrigel matrix. The beads
are surrounded by cells recruited into the structure. The granulomatous structure showsextensive neovascularization and marked lymphocyte cuffing around the new blood
vessels. Finally there is extensive fibrotic remodelling of the tissue. This is most marked at
the basement margin where the granuloma adheres to the fat cells in the peritoneal
cavity. TDM, trehalose dimycolate.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 5 | JANUARY 2007 | 45

7/30/2019 Russell (2006) Tubercle
http://slidepdf.com/reader/full/russell-2006-tubercle 8/9
1. Corbett, E. L. et al. The growing burden of tuberculosis:global trends and interactions with the HIV epidemic.
Arch. Intern. Med. 163, 1009–1021 (2003).2. Frieden, T. R., Sterling, T. R., Munsiff, S. S., Watt, C. J.
& Dye, C. Tuberculosis. Lancet 362, 887–899 (2003).3. Harries, A. D. & Dye, C. Tuberculosis. Ann. Trop. Med.
Parasitol. 100, 415–431 (2006).4. Dye, C. Global epidemiology of tuberculosis. Lancet
367, 938–940 (2006).5. Sreevatsan, S. et al. Restricted structural gene
polymorphism in the Mycobacterium tuberculosis complex indicates evolutionarily recent globaldissemination. Proc. Natl Acad. Sci. USA 94,9869–9874 (1997).
6. Diamond, J. Guns, Germs and Steel , (W. W. Norton & Company, New York, 1997).
7. Casanova, J. L. & Abel, L. Genetic dissection of immunity to mycobacteria: the human model. Annu.Rev. Immunol. 20, 581–620 (2002).
8. Cooke, G. S. & Hill, A. V. Genetics of susceptibility tohuman infectious disease. Nature Rev. Genet. 2,967–977 (2001).
9. Fulton, S. A. et al. Inhibition of majorhistocompatibility complex II expression and antigenprocessing in murine alveolar macrophages byMycobacterium bovis BCG and the 19-kilodaltonmycobacterial lipoprotein. Infect. Immun. 72,2101–2110 (2004).
10. Baghdadi, J. E. et al. An autosomal dominant majorgene confers predisposition to pulmonary tuberculosisin adults. J. Exp. Med. 203, 1679–1684 (2006).
11. Malik, S. et al. Variants of the SFTPA1 and SFTPA2genes and susceptibility to tuberculosis in Ethiopia.
Hum. Genet. 118, 752–759 (2006).12. Bornman, L. et al. Vitamin D receptor polymorphisms
and susceptibility to tuberculosis in West Africa: acase-control and family study. J. Infect. Dis. 190,1631–1641 (2004).
13. Cervino, A. C. et al. Fine mapping of a putativetuberculosis-susceptibility locus on chromosome15q11–13 in African families. Hum. Mol. Genet. 11,1599–1603 (2002).
14. Brosch, R. et al. A new evolutionary scenario for theMycobacterium tuberculosis complex. Proc. Natl
Acad. Sci. USA 99, 3684–3689 (2002).15. Gutierrez, M. C. et al. Ancient origin and gene
mosaicism of the progenitor of Mycobacteriumtuberculosis. PLoS Pathog. 1, e5 (2005).
Although not the original paper proposing that the‘founder strain’ for tuberculosis came from central
Africa and predated the speciation of M. bovis, thispaper contains a thorough and extremely
accessible discussion of the key issues.16. Mostowy, S., Cousins, D., Brinkman, J., Aranaz, A. &
Behr, M. A. Genomic deletions suggest a phylogeny forthe Mycobacterium tuberculosis complex. J. Infect.Dis. 186, 74–80 (2002).
17. Mostowy, S. & Behr, M. A. The origin and evolution of
Mycobacterium tuberculosis. Clin. Chest. Med. 26,
207–216 (2005).
18. Ulrichs, T. & Kaufmann, S. H. New insights into the
function of granulomas in human tuberculosis.
J. Pathol. 208, 261–269 (2006).
19. Flynn, J. L. & Chan, J. What’s good for the host is
good for the bug. Trends Microbiol. 13, 98–102
(2005).20. Algood, H. M., Lin, P. L. & Flynn, J. L. Tumor
necrosis factor and chemokine interactions in the
formation and maintenance of granulomas in
tuberculosis. Clin. Infect. Dis. 41 (Suppl. 3), 189–193
(2005).
21. Algood, H. M., Chan, J. & Flynn, J. L. Chemokines and
tuberculosis. Cytokine Growth Factor Rev. 14,
467–477 (2003).
22. Ulrichs, T. et al. Differential organization of the local
immune response in patients with active cavitary
tuberculosis or with nonprogressive tuberculoma.
J. Infect. Dis. 192, 89–97 (2005).
23. Fenhalls, G. et al. Distribution of IFN-γ , IL-4 and TNF-α
protein and CD8 T cells producing IL-12p40 mRNA in
human lung tuberculous granulomas. Immunology
105, 325–335 (2002).24. Tully, G. et al. Highly focused T cell responses in
latent human pulmonary Mycobacterium
tuberculosis infection. J. Immunol. 174, 2174–2184
(2005).
25. Kaplan, G. et al. Mycobacterium tuberculosis
growth at the cavity surface: a microenvironment with
failed immunity. Infect. Immun. 71, 7099–7108
(2003).
26. Dheda, K. et al. Lung remodeling in pulmonary
tuberculosis. J. Infect. Dis. 192, 1201–1209
(2005).
27. Lin, P. L. et al. Early events in Mycobacterium
tuberculosis infection in cynomolgus macaques. Infect.
Immun. 74, 3790–3803 (2006).
Temporal studies on granuloma development
are extremely challenging, particularly in
higher primates. Therefore this study by Lin
and colleagues on infections in macaques is
extremely valuable and questions some of
the accepted dogma of granuloma progression.28. Alatas, F. et al. Vascular endothelial growth factor
levels in active pulmonary tuberculosis. Chest 125,
2156–2159 (2004).
29. Ragno, S. et al. Changes in gene expression in
macrophages infected with Mycobacterium
tuberculosis: a combined transcriptomic and
proteomic approach. Immunology 104, 99–108
(2001).
30. Tsai, M. C. et al. Characterization of the tuberculous
granuloma in murine and human lungs: cellular
composition and relative tissue oxygen tension. Cell.
Microbiol. 8, 218–232 (2006).
31. Fenhalls, G. et al. In situ detection of
Mycobacterium tuberculosis transcripts in human
lung granulomas reveals differential gene
expression in necrotic lesions. Infect. Immun. 70,
6330–6338 (2002).
32. McKinney, J. D. et al. Persistence of Mycobacterium
tuberculosis in macrophages and mice requires the
glyoxylate shunt enzyme isocitrate lyase. Nature 406,735–738 (2000).
33. Deretic, V. et al. Mycobacterium tuberculosis
inhibition of phagolysosome biogenesis and
autophagy as a host defence mechanism. Cell.
Microbiol. 8, 719–727 (2006).
34. Russell, D. G. Mycobacterium tuberculosis: here
today, and here tomorrow.Nature Rev. Mol. Cell Biol.
2, 569–577 (2001).
35. Russell, D. G., Purdy, G. E., Owens, R. M., Rohde, K. &
Yates, R. M.Mycobacterium tuberculosis and the
concept of the ‘4 minute’ phagosome. ASM News 71,
459–463 (2005).
36. Sturgill-Koszycki, S., Schaible, U. E. & Russell, D. G.
Mycobacterium-containing phagosomes are
accessible to early endosomes and reflect a
transitional state in normal phagosome
biogenesis. EMBO J. 15, 6960–6968
(1996).
37. Russell, D. G., Dant, J. & Sturgill-Koszycki, S.
Mycobacterium avium- and Mycobacterium
tuberculosis-containing vacuoles are dynamic,
fusion-competent vesicles that are accessible to
glycosphingolipids from the host cell plasmalemma.
J. Immunol. 156, 4764–4773 (1996).
38. Mwandumba, H. C. et al. Mycobacterium tuberculosis
resides in nonacidified vacuoles in endocytically
competent alveolar macrophages from patients with
tuberculosis and HIV infection. J. Immunol. 172,
4592–4598 (2004).
This study demonstrates that the non-acidification
of M. tuberculosis vacuoles in macrophages in
culture is reciprocated in infected macrophages
isolated by broncholavage from tuberculosis
patients.
39. Pethe, K. et al. Isolation of Mycobacterium
tuberculosis mutants defective in the arrest of
phagosome maturation. Proc. Natl Acad. Sci. USA
101, 13642–13647 (2004).
40. MacMicking, J. D., Taylor, G. A. & McKinney, J. D.
Immune control of tuberculosis by IFN-γ -inducible
LRG-47. Science 302, 654–659 (2003).
41. Schaible, U. E., Sturgill-Koszycki, S., Schlesinger, P. H. &
Russell, D. G. Cytokine activation leads to acidification
and increases maturation of Mycobacterium avium-
containing phagosomes in murine macrophages.
J. Immunol. 160, 1290–1296 (1998).
42. Via, L. E. et al. Effects of cytokines on mycobacterial
phagosome maturation. J. Cell Sci. 111, 897–905
(1998).
43. Munoz-Elias, E. J. et al. Replication dynamics of
Mycobacterium tuberculosis in chronically infected
mice. Infect. Immun. 73, 546–551 (2005).
44. Rees, R. J. & Hart, P. D. Analysis of the host-parasite
equilibrium in chronic murine tuberculosis by total and
viable bacillary counts. Br. J. Exp. Pathol. 42, 83–88
(1961).
45. Ribi, E. et al. Induction of resistance to tuberculosis in
mice with defined components of mycobacteria and
with some unrelated materials. Immunology 46,
297–305 (1982).
46. Hamamoto, Y., Kobara, Y., Kojima, A., Kumazawa, Y. &
Yasuhira, K. Experimental production of pulmonary
granulomas. I. Immune granulomas induced by
chemically modified cell walls and their constituents.
Br. J. Exp. Pathol. 62, 259–269 (1981).
47. McLaughlin, C. A., Parker, R., Hadlow, W. J.,
Toubiana, R. & Ribi, E. Moieties of mycobacterial
mycolates required for inducing granulomatousreactions. Cell. Immunol. 38, 14–24 (1978).
48. Yamamoto, K. & Karinuma, M. Genetic control of
granuloma response to oil-associated BCG cell wall
vaccine in mice. Microbiol. Immunol. 22, 335–348
(1978).
49. Granger, D. L., Yamamoto, K. I. & Ribi, E. Delayed
hypersensitivity and granulomatous response after
immunization with protein antigens associated with a
mycobacterial glycolipid and oil droplets. J. Immunol.
116, 482–488 (1976).
50. Meyer, T. J., Ribi, E. & Azuma, I. Biologically active
components from mycobacterial cell walls. V.
Granuloma formation in mouse lungs and guinea pig
skin. Cell. Immunol. 16, 11–24 (1975).
51. Moore, V. L., Myrvik, Q. N. & Kato, M. Role of cord
factor (trehalose-6, 6′-dimycolate) in allergic
granuloma formation in rabbits. Infect. Immun. 6, 5–8
(1972).
to have evolved structurally to induce a measureddegree of pathology 85,86, and its biological activity isregulated by its mode of presentation73,78,80. The recentobservation that TDM associates with lipid droplets innecrotic regions of murine granulomas80 is intriguingbecause it provides a potential means of enhancingthe activity of TDM in the late-stage granulomas,allowing it to be at its most destructive when dam-age is needed to drive transmission. In earlier in vivo experiments, the toxicity of TDM was enhancedmarkedly by its incorporation into oil droplets orlarge particles45,51,73,78.
Profiling the transcriptional response in defined areasof human TB granulomas through the use of microarray analysis of laser-capture micro-dissected regions fromhuman tissue might provide some insights into the path-ways activated in the host during this crucial transition.The application of this information to more appropriateanimal models such as guinea pigs, rabbits or monkeysmight even allow these pathways to be probed experimen-tally. Although it might be a pipedream to imagine wecould ever block the transmission of M. tuberculosis, weshould at least aim to understand how the propagation of this successful pathogen is achieved.
R E V I E W S
46 | JANUARY 2007 | VOLUME 5 www.nature.com/reviews/micro

7/30/2019 Russell (2006) Tubercle
http://slidepdf.com/reader/full/russell-2006-tubercle 9/9
52. Bekierkunst, A. et al. Granuloma formation induced in
mice by chemically defined mycobacterial fractions.
J. Bacteriol. 100, 95–102 (1969).
53. Bekierkunst, A. Acute granulomatous response
produced in mice by trehalose-6, 6-dimycolate.
J. Bacteriol. 96, 958–961 (1968).
54. White, R. G., Jolles, P., Samour, D. & Lederer, E.
Correlation of adjuvant activity and chemical structure
of wax D fractions of Mycobacteria. Immunology 7,
158–171 (1964).
This paper marks the beginning of our appreciation
of the biological activities that reside within theisolated components of the mycobacterial cell wall.
55. Xu, S. et al. Intracellular trafficking in
Mycobacterium tuberculosis and Mycobacterium
avium-infected macrophages. J. Immunol. 153,
2568–2578 (1994).
56. Beatty, W. L. et al. Trafficking and release of
mycobacterial lipids from infected macrophages.
Traffic 1, 235–247 (2000).
This paper documented the intracellular release
of cell-wall lipids by live bacteria inside their host
macrophage. It demonstrated that these lipids
coalesced in the multi-vesicular l ysosomes, were
exocytosed as vesicular bodies and were
internalized by neighbouring cells.57. Beatty, W. L., Ullrich, H. J. & Russell, D. G.
Mycobacterial surface moieties are released from
infected macrophages by a constitutive exocytic event.
Eur. J. Cell Biol. 80, 31–40 (2001).
58. Schaible, U. E. et al. Apoptosis facilitates antigen
presentation to T lymphocytes through MHC-I andCD1 in tuberculosis. Nature Med. 9, 1039–1046
(2003).59. Winau, F., Kaufmann, S. H. & Schaible, U. E. Apoptosis
paves the detour path for CD8 T cell activation against
intracellular bacteria. Cell. Microbiol. 6, 599–607
(2004).
60. van den Elzen, P. et al. Apolipoprotein-mediated
pathways of lipid antigen presentation. Nature 437,
906–910 (2005).61. Rhoades, E. et al. Identification and macrophage-
activating activity of glycolipids released from
intracellular Mycobacterium bovis BCG. Mol.
Microbiol. 48, 875–888 (2003).62. Beatty, W. L. & Russell, D. G. Identification of
mycobacterial surface proteins released into
subcellular compartments of infected macrophages.
Infect. Immun. 68, 6997–7002 (2000).
63. Ullrich, H. J., Beatty, W. L. & Russell, D. G. Interaction
of Mycobacterium avium-containing phagosomes with
the antigen presentation pathway. J. Immunol. 165,
6073–6080 (2000).64. Arend, S. M. et al. Detection of active tuberculosis
infection by T cell responses to early-secreted
antigenic target 6-kDa protein and culture
filtrate protein 10. J. Infect. Dis. 181, 1850–1854
(2000).
65. Fortune, S. M. et al. Mutually dependent secretion of
proteins required for mycobacterial virulence. Proc.
Natl Acad. Sci. USA 102, 10676–10681 (2005).66. Weldingh, K. et al. Two-dimensional electrophoresis for
analysis of Mycobacterium tuberculosis culture filtrate
and purification and characterization of six novel
proteins. Infect. Immun. 66, 3492–3500 (1998).67. Moody, D. B. et al. T-cell activation by lipopeptide
antigens. Science 303, 527–531 (2004).
68. Ulrichs, T., Moody, D. B., Grant, E., Kaufmann, S. H. &
Porcelli, S. A. T-cell responses to CD1-presented
lipid antigens in humans with Mycobacterium
tuberculosis infection. Infect. Immun. 71, 3076–3087
(2003).
69. Dascher, C. C. & Brenner, M. B. CD1 antigenpresentation and infectious disease. Contrib.
Microbiol. 10, 164–182 (2003).
70. Puissegur, M. P. et al. An in vitro dual model of
mycobacterial granulomas to investigate the
molecular interactions between mycobacteria and
human host cells. Cell Microbiol. 6, 423–33 (2004).
71. Syed, S. S. & Hunter, R. L. Jr. Studies on the toxic
effects of quartz and a mycobacterial glycolipid,
trehalose 6, 6′-dimycolate. Ann. Clin. Lab. Sci. 27,
375–383 (1997).72. Actor, J. K., Olsen, M., Hunter, R. L. Jr & Geng, Y. J.
Dysregulated response to mycobacterial cord factor
trehalose-6, 6′-dimycolate in CD1D-/- mice.
J. Interferon. Cytokine Res. 21, 1089–1096 (2001).73. Behling, C. A., Perez, R. L., Kidd, M. R., Staton, G. W.,
Jr & Hunter, R. L. Induction of pulmonary granulomas,
macrophage procoagulant activity, and tumor necrosis
factor-α by trehalose glycolipids. Ann. Clin. Lab. Sci.
23, 256–266 (1993).74. Bentley, A. G. et al. In vitro delayed hypersensitivity
granuloma formation: development of an antigen-
coated bead model. J. Immunol. 134, 4163–4169
(1985).75. Lima, V. M. et al. Role of trehalose dimycolate in
recruitment of cells and modulation of production of
cytokines and NO in tuberculosis. Infect. Immun. 69,
5305–5312 (2001).
76. Sakaguchi, I. et al. Trehalose 6, 6′-dimycolate (Cord
factor) enhances neovascularization through vascular
endothelial growth factor production by neutrophils
and macrophages. Infect. Immun. 68, 2043–2052
(2000).77. Yamagami, H. et al. Trehalose 6, 6′-dimycolate (cord
factor) of Mycobacterium tuberculosis induces foreign-
body- and hypersensitivity-type granulomas in mice.Infect. Immun. 69, 810–815 (2001).
78. Geisel, R. E., Sakamoto, K., Russell, D. G. & Rhoades,
E. R. In vivo activity of released cell wall lipids of
Mycobacterium bovis bacillus Calmette-Guerin is due
principally to trehalose mycolates. J. Immunol. 174,
5007–5015 (2005).
This study detailed the systematic analysis of the
biological activities of released mycobacterial lipids
and concluded that trehalose dimycolate was the
most biologically active in a reconstituted
granuloma model that facilitated temporal
dissection of the cells, cytokines and chemokines
responsible for the response.
79. Rhoades, E. R., Geisel , R. E., Butcher, B. A.,
McDonough, S. & Russell, D. G. Cell wall lipids from
Mycobacterium bovis BCG are inflammatory when
inoculated within a gel matrix: characterization of a
new model of the granulomatous response to
mycobacterial components. Tuberculosis (Edinb) 85,
159–176 (2005).80. Hunter, R. L., Olsen, M., Jagannath, C. & Actor, J. K.
Trehalose 6, 6’-dimycolate and lipid in the
pathogenesis of caseating granulomas of tuberculosis
in mice. Am. J. Pathol. 168, 1249–1261 (2006).
This study built on the previous work from this
group and noticed that trehalose dimycolate (TDM)
associated with large lipid deposits within the
granuloma. The formation of macromolecular arrays
of TDM enhances its biological activity markedly.
81. Ryll, R., Kumazawa, Y. & Yano, I. Immunological
properties of trehalose dimycolate (cord factor) and
other mycolic acid-containing glycolipids-a review.
Microbiol. Immunol. 45, 801–811 (2001).
82. Hamasaki, N. et al. In vivo administration of
mycobacterial cord factor (Trehalose 6, 6′-dimycolate)
can induce lung and liver granulomas and thymic
atrophy in rabbits. Infect. Immun. 68, 3704–3709
(2000).
83. Bekierkunst, A. & Yarkoni, E. Granulomatous
hypersensitivity to trehalose-6, 6′-dimycolate (cordfactor) in mice infected with BCG. Infect. Immun. 7,
631–638 (1973).
84. Glickman, M. S., Cox, J. S. & Jacobs, W. R. Jr. A
novel mycolic acid cyclopropane synthetase is
required for cording, persistence, and virulence of
Mycobacterium tuberculosis. Mol. Cell 5, 717–727
(2000).
85. Rao, V., Fujiwara, N., Porcelli, S. A. & Glickman, M. S.
Mycobacterium tuberculosis controls host innate
immune activation through cyclopropane modification
of a glycolipid effector molecule. J. Exp. Med. 201,
535–543 (2005).
86. Rao, V., Gao, F., Chen, B., Jacobs, W. R. Jr &
Glickman, M. S. Trans-cyclopropanation of mycolicacids on trehalose dimycolate suppresses
Mycobacterium tuberculosis-induced inflammation
and virulence. J. Clin. Invest. 116, 1660–1667
(2006).
87. Camacho, L. R. et al. Analysis of the phthiocerol
dimycocerosate locus of Mycobacterium tuberculosis.
Evidence that this lipid is involved in the cell wall
permeability barrier. J. Biol. Chem. 276,
19845–19854 (2001).
88. Camacho, L. R., Ensergueix, D., Perez, E., Gicquel, B.
& Guilhot, C. Identification of a virulence gene cluster
of Mycobacterium tuberculosis by signature-tagged
transposon mutagenesis. Mol. Microbiol. 34,
257–267 (1999).
89. Cox, J. S., Chen, B., McNeil, M. & Jacobs, W. R. Jr.
Complex lipid determines tissue-specific replication of
Mycobacterium tuberculosis in mice. Nature 402,
79–83 (1999).
90. Reed, M. B. et al. A glycolipid of hypervirulent
tuberculosis strains that inhibits the innateimmune response. Nature 431, 84–87
(2004).
91. Tsenova, L. et al. Virulence of selected Mycobacterium
tuberculosis clinical isolates in the rabbit model of
meningitis is dependent on phenolic glycolipid
produced by the bacilli. J. Infect. Dis. 192, 98–106
(2005).92. Fuller, C. L., Flynn, J. L. & Reinhart, T. A. In situ
study of abundant expression of proinflammatorychemokines and cytokines in pulmonary
granulomas that develop in cynomolgus macaques
experimentally infected with Mycobacterium
tuberculosis. Infect. Immun. 71, 7023–7034
(2003).93. Rhoades, E. R., Cooper, A. M. & Orme, I. M. Chemokine
response in mice infected with Mycobacteriumtuberculosis. Infect. Immun. 63, 3871–3877 (1995).
94. Cooper, A. M. et al. Disseminated tuberculosis ininterferon γ gene-disrupted mice. J. Exp. Med. 178,
2243–2247 (1993).
AcknowledgementsThis work was supported by grants from the National Institute
of Allergy and Infectious Diseases and the National Heart,Lung and Blood Institute of the National Institutes of Health,
USA. The author would like to acknowledge the work of past
and present members of the laboratory, most notablyE. Rhoades, R. Geisel and K. Sakamoto.
Competing interests statementThe author declares no competing financial interests.
DATABASESThe following terms in this article are linked online to:
Entrez Genome Project: http://www.ncbi.nlm.nih.gov/
entrez/query.fcgi?db=genomeprj
Mycobacterium bovis | Mycobacterium tuberculosis
FURTHER INFORMATIONDavid G. Russell’s homepage:http://www.vet.cornell.edu/
public/microbiology/russellnew2.htmAccess to this links box is available online.
R E V I E W S
NATURE REVIEWS | MICROBIOLOGY VOLUME 5 | JANUARY 2007 | 47