revisiting the tams1-encoding gene as a species-specific target for the molecular detection of...
TRANSCRIPT

O
Rd
MI
a
ARRA
KTTRTM
I
bhla2zmfptpprioismrec
j
1h
Ticks and Tick-borne Diseases 4 (2013) 72– 77
Contents lists available at SciVerse ScienceDirect
Ticks and Tick-borne Diseases
j ourna l ho me page: www.elsev ier .com/ locate / t tbd is
riginal article
evisiting the Tams1-encoding gene as a species-specific target for the molecularetection of Theileria annulata in bovine blood samples
arcos Santos1, Ricardo Soares1, Pedro Costa, Ana Amaro, João Inácio ∗, Jacinto Gomes ∗
nstituto Nacional de Recursos Biológicos, I.P. – Laboratório Nacional de Investigac ão Veterinária (INRB, I.P. – LNIV), Estrada de Benfica 701, 1549-011 Lisboa, Portugal
r t i c l e i n f o
rticle history:eceived 20 December 2011eceived in revised form 15 June 2012ccepted 27 July 2012
a b s t r a c t
Tropical or Mediterranean theileriosis is a tick-borne hemoprotozoan disease that poses important healthproblems in cattle. The etiological agent is the apicomplexan parasite Theileria annulata that occurs aroundthe Mediterranean basin, Middle East, and southern Asia. PCR-based assays have been developed fordetecting theilerial infections, and the gene encoding an immunodominant major merozoite/piroplasm
eywords:heileria annulataheileriosiseal-time PCRams1
surface antigen of T. annulata (Tams1) is a commonly used genomic target. However, several studiesrevealed that this gene is highly polymorphic raising questions concerning the suitability of Tams1 gene-targeted primers to detect all T. annulata isolates. In this work, we re-evaluated the in silico sensitivityof previously developed primers targeting the Tams1 gene, designed novel and more comprehensiveprimers for its amplification, and established efficient standard and real-time PCR assays for assessing
f T. a
olecular diagnostics the presence or absence ontroduction
Tropical theileriosis (or Mediterranean theileriosis) is a tick-orne hemoprotozoan disease responsible for causing importantealth problems in cattle. The agent of the disease is Theileria annu-
ata, which occurs around the Mediterranean basin, Middle Eastnd Southern Asia (Gubbels et al., 1999; García-Sanmartín et al.,006; Branco et al., 2010; Silva et al., 2010). T. annulata sporo-oites enter the bovine host during tick feeding and rapidly invadeacrophages. Ultimately, merozoites are produced and released
rom the macrophages, invade erythrocytes and develop into piro-lasms (Dobbelaere and Heussler, 1999). The animals that survivehe acute disease become carriers of T. annulata piroplasms andlay an important role as reservoirs for the maintenance of thearasite life cycle (D’Oliveira et al., 1995). Identification of these car-ier animals is of utmost importance in epidemiological studies, fornferring infection risk and for the implementation and monitoringf control programs. Traditionally, detection of Theileria pathogensn infected animals is accomplished by microscopic examination oftained blood smears, which have low sensitivity for the assess-ent of carrier animals, in which low numbers of erythrocytes
emain infected (Altay et al., 2008). Serological methods can bemployed to diagnose subclinical infections, but cross-reactions areommon, current infections and previous exposures are generally
∗ Corresponding authors. Tel.: +351 21 7115302; fax: +351 21 7115385.E-mail addresses: [email protected] (J. Inácio),
[email protected] (J. Gomes).1 RS and MS contributed equally to this work.
877-959X/$ – see front matter © 2012 Elsevier GmbH. All rights reserved.ttp://dx.doi.org/10.1016/j.ttbdis.2012.07.006
nnulata in bovine blood samples.© 2012 Elsevier GmbH. All rights reserved.
not distinguished, and antibodies tend to disappear in long-termcarriers (Altay et al., 2008; García-Sanmartín et al., 2006). A fewmolecular diagnostic-based assays have also been developed, someof which have proven to be effective in detecting theilerial infec-tions in carrier animals. A reverse line blotting (RLB) assay basedon the amplification of the hyper-variable V4 region of the 18SrDNA gene of Theileria (and of closely-related Babesia parasites)and reverse hybridization of the products with species-specificoligonucleotide probes is currently considered to be the most sen-sitive test for detecting T. annulata (Gubbels et al., 1999; Georgeset al., 2001; Bilgic et al., 2010). Nevertheless, since RLB is a rela-tively cumbersome assay, it is not entirely suitable for use in theroutine diagnosis of Theileria infections. Some PCR-based assayshave also been described for detecting T. annulata (e.g. D’Oliveiraet al., 1995; Criado-Fornelio et al., 2009). The gene encoding animmunodominant major merozoite/piroplasm surface antigen ofthis parasite (Tams1) is commonly used as genomic target, andspecific primers for the partial amplification of this gene wereoriginally developed by D’Oliveira et al. (1995). These primers,assigned as N516 (forward) and N517 (reverse), have been fre-quently used by other authors to assess the prevalence and fordiagnosis of T. annulata infections in bovine populations from dif-ferent regions (D’Oliveira et al., 1997; Martín-Sánchez et al., 1999;Almeria et al., 2001; Sparagano et al., 2002; Dumanli et al., 2005;Aktas et al., 2006; Durrani and Kamal, 2008; Bilgic et al., 2010;Mahmmod et al., 2010; Shahnawaz et al., 2011). However, it is
currently known that Tams1-encoding gene is highly polymor-phic (Gubbels et al., 2000), which may constrain the respectivePCR-based detection of all T. annulata genotypes and potentiallycontribute to an underestimation of the real prevalence of this
ick-bo
paptrNpefs
M
S
fPuswbastmaamwwaBatw(tt3tafmafsuBInt
S
TseoPaeto
M. Santos et al. / Ticks and T
athogen. It is noteworthy that many more gene sequences arevailable in public databases at present than when N516 and N517rimers were originally designed and, therefore, it is possible thathe respective complementary targets are located in more variableegions of the gene. In this work, we review the in silico efficiency of516 and N517 primers, designed novel and more comprehensiverimers for amplifying the T. annulata species-specific Tams1-ncoding gene, and we established an efficient real-time PCR assayor detecting this important pathogenic protozoan in bovine bloodamples.
aterials and methods
amples and DNA extraction
For this study, a total of 63 DNA samples, formerly extractedrom bovine (Bos taurus) blood samples of various regions ofortugal, were selected on the basis of a previous characterizationsing an RLB assay (Gomes et al., 2012). DNA samples were clas-ified into 5 groups (Table 1): (I) T. annulata-positive samples, forhich the hyper-variable region V4 of the 18S rDNA was amplified
y PCR, using universal primers for Theileria and Babesia species,nd the presence of the parasite was clearly demonstrated in theame by RLB; (II) T. annulata-weakly-positive samples, for whichhe PCR amplification of the V4 region yielded no detectable frag-
ents in the electrophoresis gel, but the parasite was still detectedfter reverse hybridization with the species-specific probe; (III) T.nnulata mixed infections, corresponding to samples in which aixture of T. annulata and other species of Theileria and/or Babesiaas detected by RLB; (IV) samples with other piroplasms, infectedith Theileria and/or Babesia species, but not with T. annulata;
nd (V) negative samples, for which no infection with Theileria orabesia was detected. Blood samples were originally collected frompparently healthy animals into sterile tubes with EDTA (Vacu-ainer, Becton Dickinson). Total genomic DNA was extracted fromhole-blood samples by a BioSprint®96 automated workstation
Qiagen), using the BioSprint®96 Blood kit (Qiagen) according tohe manufacturer’s instructions, and stored at −20 ◦C until fur-her use. The average total DNA yield in these samples was around5–40 ng/�L (as assessed using a NanoDropTM 1000 Spectropho-ometer from Thermo Scientific). A DNA sample extracted from a T.nnulata-infected macrophage culture (sample THA-1), obtainedrom a calf with clinical diagnosis of theileriosis (confirmed by
icroscopical examination of Giemsa-stained smears and RLBssay), was used as a positive control. A DNA solution extractedrom bovine blood (sample 8182) for which a T. annulata para-itemia of 0.03% was observed using optical microscopy was alsosed as a positive control. DNA samples from Babesia bovis and. bigemina kindly provided by Varda Shkap (Kimron Veterinarynstitute, Israel) were used as negative controls of amplification. Aegative control consisting of sterile distilled water (GIBCO, Invi-rogen) was also used.
equence analysis and primer design
Comparative analysis of Tams1-encoding gene sequences from. annulata, retrieved from NCBI-GenBank, was performed throughequence alignments using the CLUSTAL X v2.0 software (Larkint al., 2007). More than 140 gene sequences of diverse geographicalrigins (e.g. Turkey, Mauritania, Sudan, India, Iraq, Tunisia, Spain,ortugal, and Italy) were analyzed [most of them were obtained
nd published by Shiels et al. (1995), Katzer et al. (1998), Al-Saeedt al. (2010) and, mainly, by Gubbels et al. (2000)]. Alignments con-aining tens of sequences were visually inspected for the presencef highly conserved nucleotide segments within T. annulata (i.e.rne Diseases 4 (2013) 72– 77 73
present in all available Tams1-encoding gene sequences), providingalso enough nucleotide differences when compared to homologousgenes from other closely related species (particularly of the genusTheileria). A novel set of primers was designed, named Tams1 forw(5′-CAA ATT CGA GAC CTA CTA CGA TG-3′) and Tams1 rev (5′-CCACTT RTC GTC CTT AAG CTC G-3′), based on selected conserved seg-ments of the gene (Fig. 1), allowing the amplification of a fragmentwith about 319 bp. Primer properties such as guanine and cytosinecontent and theoretical melting temperature were assessed usingthe OligoAnalyzer software, version 1.0.3 (Teemu Kuulasmaa, Kuo-pio, Finland). The in silico specificities of designed primers wereassessed using the BLAST suite of NCBI-GenBank.
Partial amplification of the Tams1-encoding gene usingstandard and real-time PCR primers Tams1 forw and Tams1 revwere used in standard PCR assays for the detection of T. annulata inDNA samples based on the partial amplification of the respectiveTams1-encoding gene. Reaction mixture in a 25-�L final volumecontained 2.5 mM MgCl2 (Promega), 200 �M of each deoxynucle-oside triphosphate (Promega), 1.0 �M of each primer (STAB VidaLda), 0.5 U of GoTaq®Flexi polymerase (Promega), and 1× of therespective buffer, and 5 �L of the DNA sample solution. The PCRreactions were performed in an automated DNA thermal cycler (MJMini, Biorad) with the following program: (i) one initial denatur-ing step for 10 min at 94 ◦C; 35 cycles of denaturation for 30 s at94 ◦C, annealing for 30 s at 55 ◦C, and extension for 30 s at 72 ◦C;ending with a final extension step for 10 min at 72 ◦C. Amplifiedproducts were visualized under UV light after electrophoresis ina 1.5% (w/v) agarose gel (in 1× TBE buffer) stained with ethidiumbromide. A more sensitive real-time PCR assay was also establishedusing the previous designed primer set. The optimized reactionmixture contained 1× SsoFastTM Evagreen® Supermix (BioRad, CA,USA), 0.3 �M, and 0.15 �M of Tams1 forw and Tams1 rev primer,respectively, for a final volume of 20 �L (including the addition of5 �L of template DNA sample). The real-time PCR was processedin a CFX96TM Real-Time PCR Detection System (BioRad, CA, USA)with the following optimized program: (i) one initial denaturingstep for 2 min at 95 ◦C; and (ii) 45 cycles of denaturation for 15 sat 95 ◦C, annealing for 30 s at 55 ◦C and extension for 30 s at 72 ◦C.An additional step for the determination of the melting curve ofthe amplified fragments was added at the end of the amplifica-tion program, consisting of a 1 ◦C temperature increase every 5 s(beginning at 55 ◦C and ending at 95 ◦C). This additional step allowsconfirmation that the detection of fluorescence is related to theamplification of specific DNA targets and not with the formationof artifacts such as primer dimers. Data obtained in real-time PCRexperiments were analyzed with the CFX ManagerTM Software, ver-sion 1.5 (BioRad). The nucleotide sequence of selected PCR productswas determined using the same above-mentioned Tams1 forw andTams1 rev primers for the sequencing reactions (STAB Vida Lda,Lisbon, Portugal).
Results
Sequence analysis and primer design
The comparative analysis of more than 140 Tams1-encodinggene sequences of T. annulata confirmed the polymorphic feature ofthis gene observed previously by Gubbels et al. (2000) and others.It also allowed assessing that the N516 primer has a complemen-tary target in a highly polymorphic region of the gene (Fig. 1).This was not possible to assess when this primer was designed in
1995 since only a few sequences of the Tams1 gene were avail-able at that time. Moreover, of the more than 140 T. annulataTams1 gene sequences currently available at NCBI-GenBank, a fullycomplementary target of N516 was only found in 44 sequences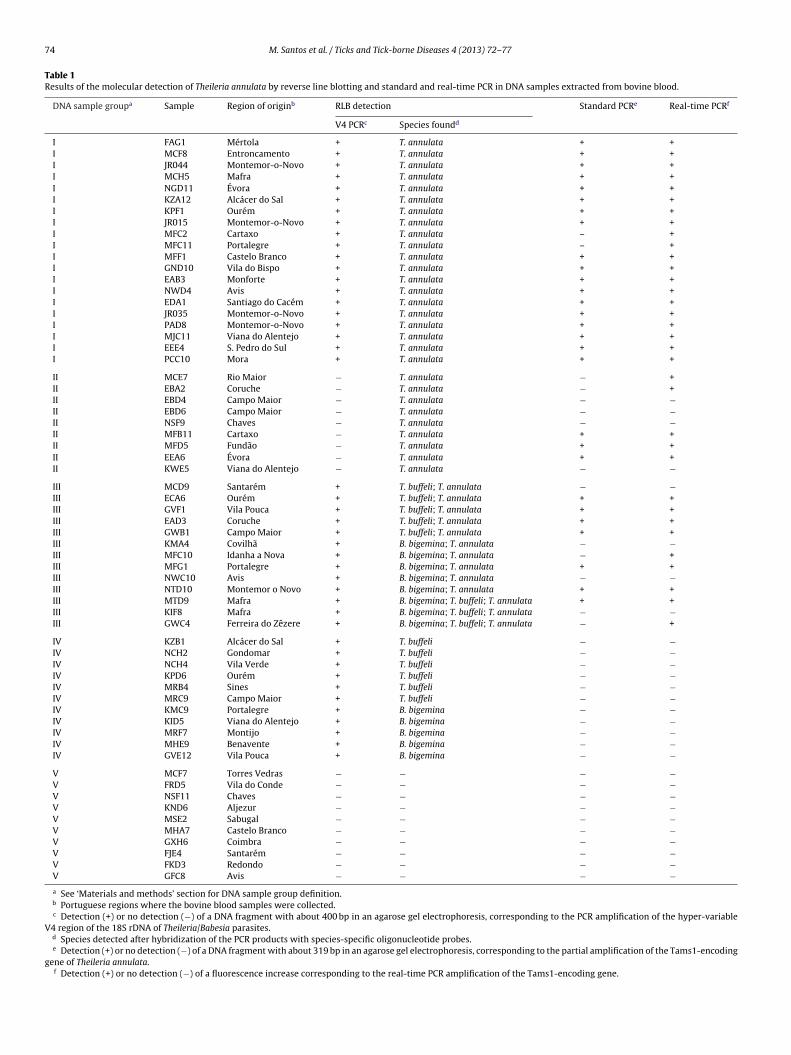
74 M. Santos et al. / Ticks and Tick-borne Diseases 4 (2013) 72– 77
Table 1Results of the molecular detection of Theileria annulata by reverse line blotting and standard and real-time PCR in DNA samples extracted from bovine blood.
DNA sample groupa Sample Region of originb RLB detection Standard PCRe Real-time PCRf
V4 PCRc Species foundd
I FAG1 Mértola + T. annulata + +I MCF8 Entroncamento + T. annulata + +I JR044 Montemor-o-Novo + T. annulata + +I MCH5 Mafra + T. annulata + +I NGD11 Évora + T. annulata + +I KZA12 Alcácer do Sal + T. annulata + +I KPF1 Ourém + T. annulata + +I JR015 Montemor-o-Novo + T. annulata + +I MFC2 Cartaxo + T. annulata – +I MFC11 Portalegre + T. annulata – +I MFF1 Castelo Branco + T. annulata + +I GND10 Vila do Bispo + T. annulata + +I EAB3 Monforte + T. annulata + +I NWD4 Avis + T. annulata + +I EDA1 Santiago do Cacém + T. annulata + +I JR035 Montemor-o-Novo + T. annulata + +I PAD8 Montemor-o-Novo + T. annulata + +I MJC11 Viana do Alentejo + T. annulata + +I EEE4 S. Pedro do Sul + T. annulata + +I PCC10 Mora + T. annulata + +
II MCE7 Rio Maior − T. annulata − +II EBA2 Coruche − T. annulata − +II EBD4 Campo Maior − T. annulata − −II EBD6 Campo Maior − T. annulata − −II NSF9 Chaves − T. annulata − −II MFB11 Cartaxo − T. annulata + +II MFD5 Fundão − T. annulata + +II EEA6 Évora − T. annulata + +II KWE5 Viana do Alentejo − T. annulata − −III MCD9 Santarém + T. buffeli; T. annulata − −III ECA6 Ourém + T. buffeli; T. annulata + +III GVF1 Vila Pouca + T. buffeli; T. annulata + +III EAD3 Coruche + T. buffeli; T. annulata + +III GWB1 Campo Maior + T. buffeli; T. annulata + +III KMA4 Covilhã + B. bigemina; T. annulata − −III MFC10 Idanha a Nova + B. bigemina; T. annulata − +III MFG1 Portalegre + B. bigemina; T. annulata + +III NWC10 Avis + B. bigemina; T. annulata − −III NTD10 Montemor o Novo + B. bigemina; T. annulata + +III MTD9 Mafra + B. bigemina; T. buffeli; T. annulata + +III KIF8 Mafra + B. bigemina; T. buffeli; T. annulata − −III GWC4 Ferreira do Zêzere + B. bigemina; T. buffeli; T. annulata − +
IV KZB1 Alcácer do Sal + T. buffeli − −IV NCH2 Gondomar + T. buffeli − −IV NCH4 Vila Verde + T. buffeli − −IV KPD6 Ourém + T. buffeli − −IV MRB4 Sines + T. buffeli − −IV MRC9 Campo Maior + T. buffeli − −IV KMC9 Portalegre + B. bigemina − −IV KID5 Viana do Alentejo + B. bigemina − −IV MRF7 Montijo + B. bigemina − −IV MHE9 Benavente + B. bigemina − −IV GVE12 Vila Pouca + B. bigemina − −V MCF7 Torres Vedras − − − −V FRD5 Vila do Conde − − − −V NSF11 Chaves − − − −V KND6 Aljezur − − − −V MSE2 Sabugal − − − −V MHA7 Castelo Branco − − − −V GXH6 Coimbra − − − −V FJE4 Santarém − − − −V FKD3 Redondo − − − −V GFC8 Avis − − − −a See ‘Materials and methods’ section for DNA sample group definition.b Portuguese regions where the bovine blood samples were collected.c Detection (+) or no detection (−) of a DNA fragment with about 400 bp in an agarose gel electrophoresis, corresponding to the PCR amplification of the hyper-variable
V4 region of the 18S rDNA of Theileria/Babesia parasites.d Species detected after hybridization of the PCR products with species-specific oligonucleotide probes.e Detection (+) or no detection (−) of a DNA fragment with about 319 bp in an agarose gel electrophoresis, corresponding to the partial amplification of the Tams1-encoding
gene of Theileria annulata.f Detection (+) or no detection (−) of a fluorescence increase corresponding to the real-time PCR amplification of the Tams1-encoding gene.

M. Santos et al. / Ticks and Tick-borne Diseases 4 (2013) 72– 77 75
Fig. 1. Alignment of selected sequences of the Tams1-encoding gene of Theileria annulata (Tann) isolates from distinct locations (Ma, Mauritania; Tu, Turkey; Su, Sudan;I f the
( f nuclp ighlig
(opl(pepcgauIpttfttawgtbAiv
D
fwutrTsdatt(a(siL
n, India; Ba, Bahrain; Pt, Portugal; Sp, Spain; It, Italy) and of homologous genes oNCBI-GenBank accession numbers are indicated for each sequence). Numbering orimers N516 (D’Oliveira et al., 1995), Tams1 forw and Tams1 rev (this study) are h
corresponding to an in silico sensitivity of about 31%). About 56%f the T. annulata sequences harbor 5 or more mismatches com-ared to the fully complementary N516 primer target (mainly
ocated in the region corresponding to the 3′ end of the primer)Fig. 1). It was not possible to assess the variability of the com-lementary target of N517 primer since it is located in a genend for which there are only 4 nucleotide sequences available inublic databases. However, the polymorphisms found in the N516omplementary target of some Tams1-encoding gene alleles sug-ests that the PCR-based detection of T. annulata, using this primernd genomic target, can result in false-negatives and consequentlynderestimate the presence of this parasite in bovine populations.
n fact, we detected recently several cases of bovine blood sam-les clearly positive for the presence of T. annulata, but for whichhe Tams1-encoding gene amplification was negative when usinghe N516/N517 primers (Gomes et al., unpublished results). There-ore, novel forward (Tams1 forw) and reverse (Tams1 rev) primersargeting more conserved regions of the gene were designed andested in this work (Fig. 1). The in silico sensitivities of Tams1 forwnd Tams1 rev primers were 98% (with only 3 Tams1 gene allelesith non-fully-complementary targets, but presenting only sin-
le mismatches) and 99%, respectively. The homologous genes ofhe closely-related species T. lestoquardi, which is unable to infectovines (Leemans et al., 1999), T. parva, only present in somefrican regions, and T. taurotragi all presented several mismatches
n the complementary target region for both primers, rendering itery specific for T. annulata (Fig. 1).
etection of Theileria annulata
DNA samples extracted from bovine blood and previously testedor the presence of T. annulata and other piroplasms by RLB,hich was considered our gold-standard detection method, weresed for the setting of standard and real-time PCR assays usinghe designed Tams1 forw and Tams1 rev primers. Examples ofesults obtained in real-time PCR assays are presented in Fig. 2.he positive control sample 8182, for which a very low para-itemia of 0.03% could be microscopically determined (3 piroplasmsetected in about 10,000 erythrocytes counted), yielded strongmplification signals (Ct values of 25.1). Following the amplifica-ion steps (Fig. 2A), a melting temperature curve analysis confirmedhat the correct Tams1-encoding gene fragments were amplifiedpresenting melting temperatures between 79 ◦C and 83 ◦C) andssessed the eventual formation of artifacts such as primer dimers
with melting temperatures around 73 ◦C) (Fig. 2B). The nucleotideequences of 9 amplified Tams1 gene fragments, with distinct melt-ng temperatures (NWD4 ∼ 79 ◦C; PCC10 ∼ 79.5 ◦C; MFC2 ∼ 80.0 ◦C;PC1 ∼ 80.0 ◦C; JR044 ∼ 80.5 ◦C; THA-1 ∼ 80.5 ◦C; JR015 ∼ 81.0 ◦C;closely-related species T. lestoquardi (Tles), T. parva (Tpar), and T. taurotragi (Ttau)eotide positions uses sequence AF214858 as reference. Complementary targets ofhted inside a box, and mismatches are typed in bold.
JR035 ∼ 81.0 ◦C; and PAD8 ∼ 81.5 ◦C), were obtained and depositedat NCBI-GenBank with accession numbers JX088596–JX088604.
The optimized PCR and real-time PCR detection assays showed100% specificity, since T. annulata was not detected in DNA samplesof Groups IV and V (Table 1). The detection sensitivities were 90%and 100% for the PCR and real-time PCR assays, respectively, takinginto consideration all Group I DNA samples, which harbor singleinfections of T. annulata (Table 1). However, detection sensitivitiesfor DNA samples of Groups II and III were lower for both standardand real-time PCR assays (33% and 56% for Group II and 54% and69% for Group III, respectively) (Table 1).
Discussion
Several nucleic acid testing assays based in the polymerasechain reaction have been developed for detecting T. annulataparasites in biological samples of the bovine hosts and vectorticks (D’Oliveira et al., 1995; Gubbels et al., 1999; Martín-Sánchezet al., 1999; Kirvar et al., 2000; Georges et al., 2001; Sparaganoet al., 2002; Bilgic et al., 2010). The efficiency of these assaysdepends on a variety of factors such as genomic targets and primersequences, reagent manufacturers and concentrations, amplifica-tion conditions and DNA extraction, and respective storage ofsamples (Sparagano et al., 2002). One widely-used genomic targetin PCR-based assays for detecting T. annulata is the gene encod-ing the major merozoite/piroplasm surface protein (Tams1) of thisparasite. Tams1 gene-targeted primer set originally developed byD’Oliveira et al. (1995) has been the most widely used for the PCR-based assessment of T. annulata infections (D’Oliveira et al., 1997;Martín-Sánchez et al., 1999; Almeria et al., 2001; Sparagano et al.,2002; Dumanli et al., 2005; Aktas et al., 2006; Durrani and Kamal,2008; Bilgic et al., 2010; Mahmmod et al., 2010; Shahnawaz et al.,2011). Our in silico analysis revealed that at least one of thoseprimers targets a highly polymorphic region of the Tams1 gene(Fig. 1). In this work, a novel primer set was designed targeting con-served regions of more than 140 Tams1 gene sequences availablefrom public databases (Fig. 1), which amplifies a T. annulata species-specific DNA fragment with about 319 bp. The efficiency of theseprimers was assessed using standard and real-time PCR assays.Several real-time PCR applications were previously described fordetecting Theileria and other closely related Babesia parasites, usinggenomic targets such as the 18S rRNA and cytochrome b genes(Jeong et al., 2003; Buling et al., 2007; Kim et al., 2007, 2008;Sibeko et al., 2008; Criado-Fornelio et al., 2009; Bhoora et al., 2010;
Papli et al., 2011; Ramos et al., 2011), albeit none was specifi-cally developed to target the pathogenic T. annulata. In order toimprove the detection of lower levels of T. annulata in samples,45 cycles were used in real-time PCR assays (Fig. 2A). Following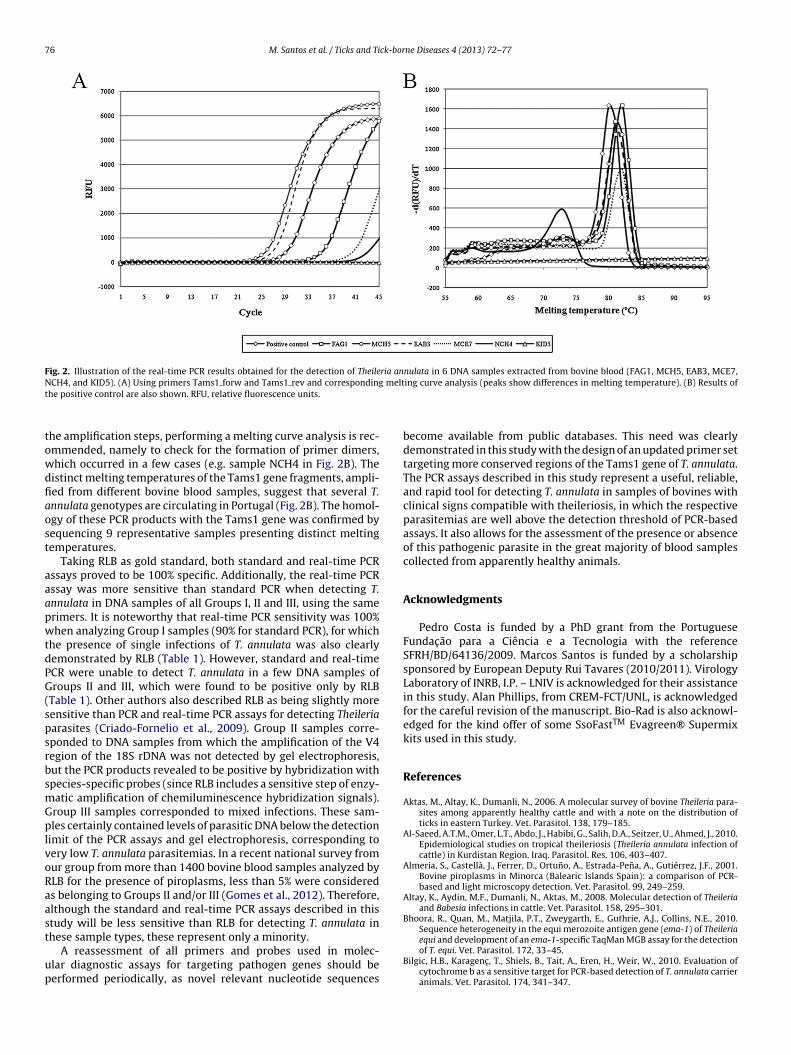
76 M. Santos et al. / Ticks and Tick-borne Diseases 4 (2013) 72– 77
Fig. 2. Illustration of the real-time PCR results obtained for the detection of Theileria annulata in 6 DNA samples extracted from bovine blood (FAG1, MCH5, EAB3, MCE7,N meltt
towdfiaost
aaapwtdPG(spsrbsmGplvoRaast
up
CH4, and KID5). (A) Using primers Tams1 forw and Tams1 rev and correspondinghe positive control are also shown. RFU, relative fluorescence units.
he amplification steps, performing a melting curve analysis is rec-mmended, namely to check for the formation of primer dimers,hich occurred in a few cases (e.g. sample NCH4 in Fig. 2B). Theistinct melting temperatures of the Tams1 gene fragments, ampli-ed from different bovine blood samples, suggest that several T.nnulata genotypes are circulating in Portugal (Fig. 2B). The homol-gy of these PCR products with the Tams1 gene was confirmed byequencing 9 representative samples presenting distinct meltingemperatures.
Taking RLB as gold standard, both standard and real-time PCRssays proved to be 100% specific. Additionally, the real-time PCRssay was more sensitive than standard PCR when detecting T.nnulata in DNA samples of all Groups I, II and III, using the samerimers. It is noteworthy that real-time PCR sensitivity was 100%hen analyzing Group I samples (90% for standard PCR), for which
he presence of single infections of T. annulata was also clearlyemonstrated by RLB (Table 1). However, standard and real-timeCR were unable to detect T. annulata in a few DNA samples ofroups II and III, which were found to be positive only by RLB
Table 1). Other authors also described RLB as being slightly moreensitive than PCR and real-time PCR assays for detecting Theileriaarasites (Criado-Fornelio et al., 2009). Group II samples corre-ponded to DNA samples from which the amplification of the V4egion of the 18S rDNA was not detected by gel electrophoresis,ut the PCR products revealed to be positive by hybridization withpecies-specific probes (since RLB includes a sensitive step of enzy-atic amplification of chemiluminescence hybridization signals).roup III samples corresponded to mixed infections. These sam-les certainly contained levels of parasitic DNA below the detection
imit of the PCR assays and gel electrophoresis, corresponding toery low T. annulata parasitemias. In a recent national survey fromur group from more than 1400 bovine blood samples analyzed byLB for the presence of piroplasms, less than 5% were considereds belonging to Groups II and/or III (Gomes et al., 2012). Therefore,lthough the standard and real-time PCR assays described in thistudy will be less sensitive than RLB for detecting T. annulata in
hese sample types, these represent only a minority.A reassessment of all primers and probes used in molec-lar diagnostic assays for targeting pathogen genes should beerformed periodically, as novel relevant nucleotide sequences
ing curve analysis (peaks show differences in melting temperature). (B) Results of
become available from public databases. This need was clearlydemonstrated in this study with the design of an updated primer settargeting more conserved regions of the Tams1 gene of T. annulata.The PCR assays described in this study represent a useful, reliable,and rapid tool for detecting T. annulata in samples of bovines withclinical signs compatible with theileriosis, in which the respectiveparasitemias are well above the detection threshold of PCR-basedassays. It also allows for the assessment of the presence or absenceof this pathogenic parasite in the great majority of blood samplescollected from apparently healthy animals.
Acknowledgments
Pedro Costa is funded by a PhD grant from the PortugueseFundac ão para a Ciência e a Tecnologia with the referenceSFRH/BD/64136/2009. Marcos Santos is funded by a scholarshipsponsored by European Deputy Rui Tavares (2010/2011). VirologyLaboratory of INRB, I.P. – LNIV is acknowledged for their assistancein this study. Alan Phillips, from CREM-FCT/UNL, is acknowledgedfor the careful revision of the manuscript. Bio-Rad is also acknowl-edged for the kind offer of some SsoFastTM Evagreen® Supermixkits used in this study.
References
Aktas, M., Altay, K., Dumanli, N., 2006. A molecular survey of bovine Theileria para-sites among apparently healthy cattle and with a note on the distribution ofticks in eastern Turkey. Vet. Parasitol. 138, 179–185.
Al-Saeed, A.T.M., Omer, L.T., Abdo, J., Habibi, G., Salih, D.A., Seitzer, U., Ahmed, J., 2010.Epidemiological studies on tropical theileriosis (Theileria annulata infection ofcattle) in Kurdistan Region. Iraq. Parasitol. Res. 106, 403–407.
Almeria, S., Castellà, J., Ferrer, D., Ortuno, A., Estrada-Pena, A., Gutiérrez, J.F., 2001.Bovine piroplasms in Minorca (Balearic Islands Spain): a comparison of PCR-based and light microscopy detection. Vet. Parasitol. 99, 249–259.
Altay, K., Aydin, M.F., Dumanli, N., Aktas, M., 2008. Molecular detection of Theileriaand Babesia infections in cattle. Vet. Parasitol. 158, 295–301.
Bhoora, R., Quan, M., Matjila, P.T., Zweygarth, E., Guthrie, A.J., Collins, N.E., 2010.Sequence heterogeneity in the equi merozoite antigen gene (ema-1) of Theileria
equi and development of an ema-1-specific TaqMan MGB assay for the detectionof T. equi. Vet. Parasitol. 172, 33–45.Bilgic, H.B., Karagenc , T., Shiels, B., Tait, A., Eren, H., Weir, W., 2010. Evaluation ofcytochrome b as a sensitive target for PCR-based detection of T. annulata carrieranimals. Vet. Parasitol. 174, 341–347.

ick-bo
B
B
C
D
D
D
D
D
G
G
G
G
G
J
K
K
sitol. 174, 199–205.
M. Santos et al. / Ticks and T
ranco, S., Orvalho, J., Leitão, A., Pereira, I., Malta, M., Mariano, I., Carvalho, T.,Baptista, R., Shiels, B.R., Peleteiro, M.C., 2010. Fatal cases of Theileria annulatainfection in calves in Portugal associated with neoplastic-like lymphoid cellproliferation. J. Vet. Sci. 11, 27–34.
uling, A., Criado-Fornelio, A., Asenzo, G., Benitez, D., Barba-Carretero, J.C.,Florin-Christensen, M., 2007. A quantitative PCR assay for the detection andquantification of Babesia bovis and B. bigemina. Vet. Parasitol. 147, 16–25.
riado-Fornelio, A., Buling, A., Asenzo, G., Benitez, D., Florin-Christensen, M.,Gonzalez-Oliva, A., Henriques, G., Silva, M., Alongi, A., Agnone, A., Torina, A.,Madruga, C.R., 2009. Development of fluorogenic probe-based PCR assays forthe detection and quantification of bovine piroplasmids. Vet. Parasitol. 162,200–206.
’Oliveira, C., van der Weide, M., Habela, M.A., Jacquiet, P., Jongejan, F., 1995. Detec-tion of Theileria annulata in blood samples of carrier cattle by PCR. J. Clin.Microbiol. 33, 2665–2669.
’Oliveira, C., van der Weide, M., Jacquiet, P., Jongejan, F., 1997. Detection of Theileriaannulata by the PCR in ticks (Acari: Ixodidae) collected from cattle in Mauritania.Exp. Appl. Acarol. 21, 279–291.
obbelaere, D., Heussler, V., 1999. Transformation of leukocytes by Theileria parvaand T. annulata. Annu. Rev. Microbiol. 53, 1–42.
umanli, N., Aktas, M., Cetinkaya, B., Cakmak, A., Koroglu, E., Saki, C.E., Erdogmus,Z., Nalbantoglu, S., Ongor, H., Sims ek, S., Karahan, M., Altay, K., 2005. Prevalenceand distribution of tropical theileriosis in eastern Turkey. Vet. Parasitol. 127,9–15.
urrani, A.Z., Kamal, N., 2008. Identification of ticks and detection of blood protozoain Friesian cattle by polymerase chain reaction test and estimation of bloodparameters in district Kasur, Pakistan. Trop. Anim. Health Prod. 40, 441–447.
arcía-Sanmartín, J., Nagore, D., García-Pérez, A.L., Juste, R.A., Hurtado, A.,2006. Molecular diagnosis of Theileria and Babesia species infecting cattlein northern Spain using reverse line blot macroarrays. BMC Vet. Res. 2 (6),http://dx.doi.org/10.1186/1746-6148-2-16.
eorges, K., Loria, G.R., Riili, S., Greco, A., Caracappa, S., Jongejan, F., Sparagano, O.,2001. Detection of haemoparasites in cattle by reverse line blot hybridisationwith a note on the distribution of ticks in Sicily. Vet. Parasitol. 99, 273–286.
omes, J., Soares, R., Santos, M., Santos-Gomes, G., Botelho, A., Amaro, A., Inácio,J., 2012. Detection of Theileria and Babesia infections amongst asymptomaticcattle in Portugal. Ticks Tick-Borne Dis., http://dx.doi.org/10.1016/j.ttbdis.2012.07.002.
ubbels, J.M., de Vos, A.P., van der Weide, M., Viseras, J., Schouls, L.M., De Vries, E.,Jongejan, F., 1999. Simultaneous detection of bovine Theileria and Babesia speciesby reverse line blot hybridization. J. Clin. Microbiol. 37, 1782–1789.
ubbels, M.J., Katzer, F., Hide, G., Jongejan, F., Shiels, B.R., 2000. Generation of amosaic pattern of diversity in the major merozoite-piroplasm surface antigenof Theileria annulata. Mol. Biochem. Parasitol. 110, 23–32.
eong, W., Kweon, C.H., Kang, S.W., Paik, S.G., 2003. Diagnosis and quantification ofTheileria sergenti using TaqMan PCR. Vet. Parasitol. 111, 287–295.
atzer, F., McKellar, S., Ben Miled, L., D’Oliveira, C., Shiels, B., 1998. Selection for
antigenic diversity of Tams1; the major merozoite antigen of Theileria annulata.Ann. N.Y. Acad. Sci. 849, 96–108.im, C.M., Blanco, L.B., Alhassan, A., Iseki, H., Yokoyama, N., Xuan, X., Igarashi, I.,2008. Diagnostic real-time PCR assay for the quantitative detection of Theileriaequi from equine blood samples. Vet. Parasitol. 151, 158–163.
rne Diseases 4 (2013) 72– 77 77
Kim, C., Iseki, H., Herbas, M.S., Yokoyama, N., Suzuki, H., Xuan, X., Fujisaki, K., Igarashi,I., 2007. Development of Taqman-based real-time PCR assays for diagnosticdetection of Babesia bovis and Babesia bigemina. Am. J. Trop. Med. Hyg. 77,837–841.
Kirvar, E., Ilhan, T., Katzer, F., Hooshmand-Rad, P., Zweygarth, E., Gerstenberg,C., Phipps, P., Brown, C.G., 2000. Detection of Theileria annulata in cattleand vector ticks by PCR using the Tams1 gene sequences. Parasitology 120,245–254.
Larkin, M.A., Blackshields, G., Brown, N.P., Chenna, R., McGettigan, P.A., McWilliam,H., Valentin, F., Wallace, I.M., Wilm, A., Lopez, R., Thompson, J.D., Gibson, T.J.,Higgins, D.G., 2007. Clustal W and Clustal X version 2.0. Bioinformatics 23,2947–2948.
Leemans, I., Brown, D., Hooshmand-Rad, P., Kirvar, E., Uggla, A., 1999. Infectivity andcross-immunity studies of Theileria lestoquardi and Theileria annulata in sheepand cattle. I. In vivo responses. Vet. Parasitol. 82, 179–192.
Mahmmod, Y.S., El-Balkemy, F.A., Yuan, Z.G., El-Mekkawy, M.F., Monazie, A.M.,Zhu, X.Q., 2010. Field evaluation of PCR assays for the diagnosis of tropi-cal theileriosis in cattle and water Buffaloes in Egypt. J. Anim. Vet. Adv. 9,696–699.
Martín-Sánchez, J., Viseras, J., Adroher, F.J., García-Fernández, P., 1999. Nested poly-merase chain reaction for detection of Theileria annulata and comparison withconventional diagnostic techniques: its use in epidemiology studies. Parasitol.Res. 85, 243–245.
Papli, N., Landt, O., Fleischer, C., Koekemoer, J.O., Mans, B.J., Pienaar, R., Josemans, A.,Zweygarth, E., Potgieter, F., Latif, A.A., 2011. Evaluation of a TaqMan real-timePCR for the detection of Theileria parva in buffalo and cattle. Vet. Parasitol. 175,356–359.
Ramos, C.A., Araújo, F.R., Souza, I.I., Bacanelli, G., Luiz, H.L., Russi, L.S., Oliveira, R.H.,Soares, C.O., Rosinha, G.M., Alves, L.C., 2011. Real-time polymerase chain reac-tion based on msa2c gene for detection of Babesia bovis. Vet. Parasitol. 176,79–83.
Shahnawaz, S., Ali, M., Aslam, M.A., Fatima, R., Chaudhry, Z.I., Hassan, M.U., Ali, M.,Iqbal, F., 2011. A study on the prevalence of a tick-transmitted pathogen. Theileriaannulata, and hematological profile of cattle from Southern Punjab (Pakistan).Parasitol. Res. 109, 1155–1160, http://dx.doi.org/10.1007/s00436-011-2360-1.
Shiels, B.R., D’Oliveira, C., McKellar, S., Ben-Miled, L., Kawazu, S., Hide, G., 1995.Selection of diversity at putative glycosylation sites in the immunodominantmerozoite/piroplasm surface antigen of Theileria parasites. Mol. Biochem. Para-sitol. 72, 149–162.
Sibeko, K.P., Oosthuizen, M.C., Collins, N.E., Geysen, D., Rambritch, N.E., Latif, A.A.,Groeneveld, H.T., Potgieter, F.T., Coetzer, J.A., 2008. Development and evalua-tion of a real-time polymerase chain reaction test for the detection of Theileriaparva infections in Cape buffalo (Syncerus caffer) and cattle. Vet. Parasitol. 155,37–48.
Silva, M.G., Marques, P.X., Oliva, A., 2010. Detection of Babesia and Theileria speciesinfection in cattle from Portugal using a reverse line blotting method. Vet. Para-
Sparagano, O.A., Carelli, G., Ceci, L., Shkap, V., Molad, T., Vitale, F., Loria, G.R., Reale,S., Caracappa, S., Bouattour, A., Almeria, S., Castella, J., Corchero, E., Habela, M.,2002. Pan-Mediterranean comparison for the molecular detection of Theileriaannulata. Ann. N.Y. Acad. Sci. 969, 73–77.