respuesta electrofisiologica de la planta de … · savia, concluyendo que la señal se propaga por...
TRANSCRIPT

RESPUESTA ELECTROFISIOLOGICA DE LA PLANTA DE
AGUACATE (Persea americana Mill.) AL ESTRÉS HÍDRICO
ELECTROPHYSIOLOGICAL RESPONSE OF THE AVOCADO PLANT (Persea
americana Mill.) TO WATER STRESS.
L Rios-Rojas 1 L Gurovich2
1Corporación Colombiana de Investigación Agropecuaria – Agrosavia C.I. Palmira. Palmira,
Valle del cauca (Colombia).
2. Pontificia Universidad Católica de Chile. Santiago (Chile).
Correo electrónico: [email protected]
Resumen
Las plantas responden diferencial y rápidamente a estímulos ambientales mediante señales
eléctricas, optimizando así sus procesos fisiológicos de adaptación. Este comportamiento fue
medido y usado como indicador del estado hídrico de las plantas, al someterlas a déficit hídrico
durante periodos prolongados de tiempo. El experimento se desarrolló entre octubre 10 y
noviembre 15 de 2013, en la Pontificia Universidad Católica de Chile, en cuatro especies leñosas
frutales: Citrus limon, Persea americana, Prunus domestica y Prunus persica. Para las
mediciones se utilizó un multivoltímetro Keithley y un sensor electrofisiológico. Se midieron
simultáneamente el contenido de humedad del suelo y variables climáticas; parámetros que fueron
relacionados con el potencial eléctrico, para identificar el efecto del estrés hídrico inducido. Se
encontró que el potencial eléctrico presentó un ciclo Circadiano en las plantas experimentales,
con un máximo y un mínimo que se corresponden en magnitud en el tiempo y que este ciclo se
altera en su amplitud y frecuencia cuando la planta se somete a déficit hídrico y sufre estrés. El
ciclo recupera su comportamiento “normal” cuando la planta vuelve a ser hidratada. Así, el
comportamiento eléctrico de la planta puede ser usado como una herramienta para el manejo de
precisión del riego, indicando cuándo regar, más no indica el volumen a regar, así que es necesario
medir simultáneamente la humedad del suelo. Finalmente, los resultados aquí obtenidos no
pueden ser generalizados a otras especies leñosas, debido a sus diferencias fisiológicas y
morfológicas y por tanto investigación en otras especies sería necesaria.
Abstract
Plants react differently and quickly to environmental stimuli through electrical signals in order to
optimize their physiological adaptation processes. This behavior was measured and used as an
indicator of plant water status, when subjected to water stress for prolonged periods of time. The
experiment was conducted between October 10th and November 15th, 2013, at the Pontifical

Catholic University of Chile, on four woody plant species: Citrus lemon, Persea americana,
Prunus domestica and Prunus persica. Measurements were carried out using a Keithley
multivoltimeter and an electrophysiological sensor. Soil moisture content and climatic variables
were measured simultaneously; the latter parameters which are related to the electric potential
used to identify the effect of water stress. It was found that the electric potential showed a
circadian cycle in the experimental plants with a maximum and a minimum corresponding in
magnitude over time; this cycle was altered in its amplitude and frequency when the plant was
subjected to stress by water deficit. The cycle returned to its "normal" behavior when the plant
was hydrated again. Thus, the electrical behavior of the plant can be used as a tool for handling
precision irrigation, indicating when to water, but does not indicate the volume to water that is
needed, thus it is necessary to measure soil moisture simultaneously. Finally, the results obtained
in this study can’t be generalized to other woody species, due to physiological and morphological
differences amongst species and therefore it is necessary to conduct further studies on other
species.
Palabras Claves: Potencial eléctrico, estrés hídrico, riego, Persea americana.
Introducción
Las plantas responden eléctricamente a estímulos del medio ambiente (Burdon-Sanderson, 1873;
Darwin, 1896), como un rápido ajuste fisiológico para adaptarse (Fromm, 2006). Respuestas
eléctricas (PE, potencial eléctrico) frente a diferentes estímulos bióticos y abióticos han sido
reportados: respiración, fotosíntesis y fototropismo (Volkov y Ranatunga 2006; Pavlovic y
Macuso, 2011), inducción floral (Wagner et al., 2012), presión osmótica (Schroeder y Hedrich,
1989), mecanismos de defensa a daño por herbívoros (Fromm y Lautner, 2007; Volkov y
Ranatunga, 2006, Heil y Ton, 2008), daño mecánico y por insectos (Brenner et al., 2006),
intensidad de la luz (Cabral, 2011, Volkov, et al., 2012), temperatura (Kai, et al., 2011, Cabral,
2011), estimulación mecánica (Volkov y Ranatunga, 2006), disponibilidad hídrica (Brenner, et
al., 2006, Gurovich y Hermosilla, 2009; Oyarce y Gurovich, 2010; Ríos-Rojas et al 2014, 2015).
Estos y otros estímulos han sido reportados en Fromm y Lautner (2007), Baluška y Mankuso
(2009), y Gurovich (2012). Las señales se expresan como potenciales de acción (PA) y
potenciales de variación (PV) y su magnitud será tan alta como intenso haya sido el estímulo
(Volkov et al, 2012; Wang et al, 2009). La señal generada puede viajar largas distancias y se da
principalmente por las conexiones simplásticas del floema (Bose, 1926; Van Bel y Ehlers, 2005).
El floema se convierte en un sistema de comunicación electroquímica, operando como una red
neuronal simple (Mancuso, 1999; Zimmermann et al, 2009; Fromm y Lautner, 2012; Volkov,
2012). Pruebas de transmisión de señales a altas velocidades indican que existen señales
diferentes a señales hidráulicas (Mullendore et al, 2010) y hormonales (Rodríguez et al., 2011;
Fromm, 2006; Gibert, 2006) que generan ajustes fisiológicos instantáneos. Gibert (2006) en sus
experimentos con Populus trichocarpa, reporta la relación del PE con cambios en el flujo de
savia, concluyendo que la señal se propaga por el floema en cortas y largas distancias, y que es
posible correlacionarlos con cambios en las tasas fotosintéticas. Grams et al. (2007) observaron
incrementos en la conductancia estomática en Zea mays, después de una rehidratación, midiendo
simultáneamente la transmisión de un señal eléctrica a una velocidad de 1 cm.s-1. Señalamiento

eléctrico en especies leñosas con la alternancia de luz/oscuridad y disponibilidad hídrica han sido
reportado por Gil et al. (2009); Gurovich y Hermosilla, (2009); Oyarce y Gurovich, (2011); Ríos-
Rojas et al (2014, 2015). Haciendo uso del comportamiento eléctrico de plantas leñosas, y su
asociación con la respuesta al estrés, fue posible usar este tipo de señal como un indicador del
estado hídrico de la planta, en tiempo real, para ser recomendado en el manejo agronómico y
preciso del riego.
Materiales y Métodos
El experimento se desarrolló en la unidad de invernaderos de la Pontificia Universidad Católica
de Chile, en Santiago de Chile. Se contó con cuatro jaulas Faraday, en cada una de las cuales se
tenían 10 plantas de una especie diferente: Persea americana, Prunus domestica, Citrus limón y
Prunus persica. Las plantas se instalaron en materos de 20L, con un fondo de grava y perlita
como sustrato altamente drenante, para llegar rápidamente a estados de déficit. En 4 de cada
10 contenedores se instaló un sensor de capacitancia eléctrica que mide el contenido
volumétrico de agua en el suelo (ɵ, %). El registro de humedad se hizo cada 15 minutos.
Parámetros micro - ambientales fueron registrados mediante sensores integrados a una
estación meteorológica automática situada entre las dos jaulas Faraday. Una estación
meteorológica automática completa fue instalada en el sitio experimental. Dos electrodos
(electrodo de referencia y electrodo de medición, figura 1) se insertan en el tronco a 20 cm y 30
cm por encima del injerto. Cada circuito eléctrico está conectado a un amplificador multi -
voltímetro (Keithley, modelo 2701). En simultáneo se conectan dos electrodos de un prototipo
de sensor electrofisiológico, que permite hacer las mediciones de forma inalámbrica. El registro
de las mediciones del PE utiliza el software de utilidad EXCELinx®, y esta información se analizó
en formato gráfico utilizando un algoritmo desarrollado en el lenguaje de programación R®. Los
tratamientos consistieron en regar ccinco plantas cada 12 horas (2,4 L), mientras otras cinco se
someten a restricción hídrica. La condición de estrés fue verificada midiendo conductancia
estomática (Gs; Wm-2s-1), concentración interna de CO2 (Ci; µmolm-2s-1) y fotosíntesis (A; µmolm-
2s-1) con un IRGAS-CI340 (CID, Bio-Science Inc, 2014). Un análisis de Boxplot fue hecho para
identificar los cambios en Gs durante el período de riego y restricción. El programa R® fue usado
para ejecutar algoritmos que correlacionaran el PE con: Temperatura, radiación y humedad del
sustrato. Con el mismo software, se hizo la caracterización de la señal, encontrando las
frecuencias fundamentales (ciclo circadiano – alta frecuencia) programando el método de la
transformada rápida de Fourier (FFT, siglas en inglés; Duhamel y Vetterli, 1990). Ocho días
después de la restricción, la totalidad de las plantas se regaron y se inició un período de
recuperación, en el cual se midió la respuesta del PE. Un análisis completo de la señal eléctrica
se muestra en este trabajo sólo para las plantas de Persea americana y Prunus domestica, para
el período experimental comprendido entre el 10 de octubre y el 05 de noviembre de 2013. Una
prueba de Shapiro-Wilks fue realizada para probar la existencia de la no-normalidad de los

datos. Una prueba de Mann Whitney (Kasuya, 2001) fue hecha para determinar las diferencias
significativas entre tratamientos, usando las variables medidas para las condiciones de estrés.
Figura 1. Esquema de medición del potencial eléctrico en la planta.
Resultados y Discusión
La figura 2 muestra la correlación del PE para Persea americana con la temperatura (a), la
radiación (b) y la humedad del sustrato (c), para el período comprendido entre el 10 de octubre
y el 05 de noviembre de 2013. La figura 3 relaciona los resultados para Prunus domestica. Las
líneas punteadas verticales muestran el período de restricción hídrica. Los valores de PE
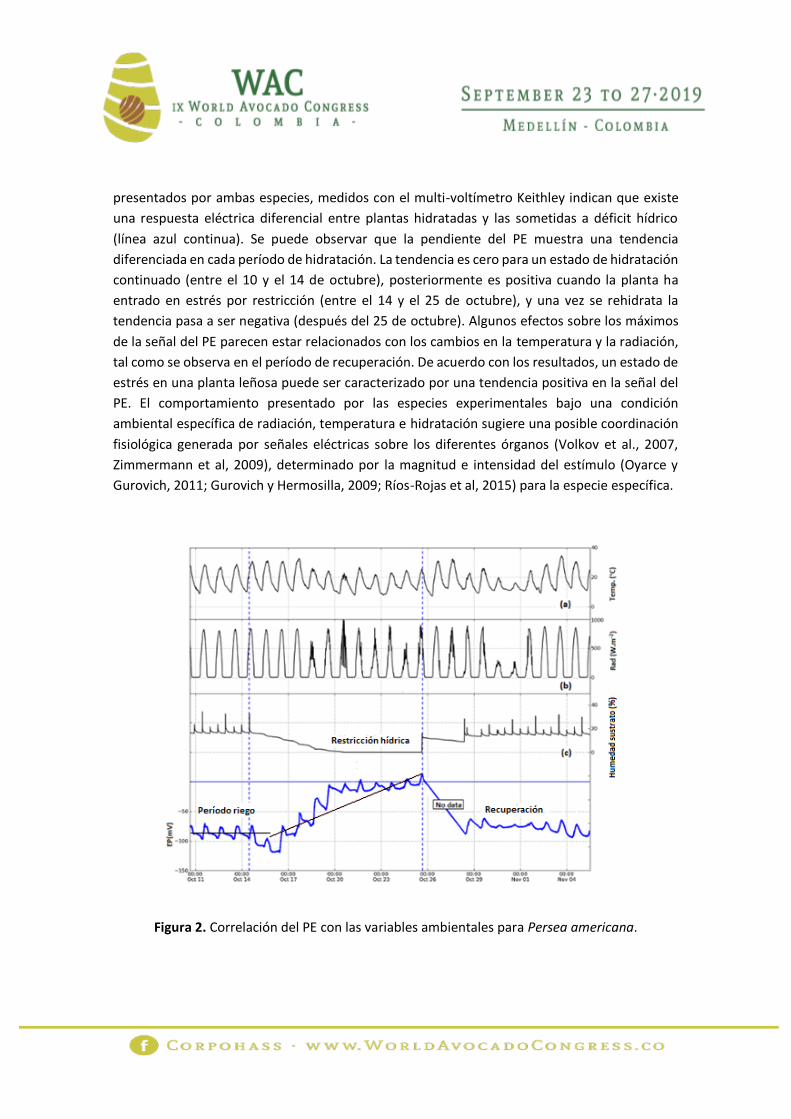
presentados por ambas especies, medidos con el multi-voltímetro Keithley indican que existe
una respuesta eléctrica diferencial entre plantas hidratadas y las sometidas a déficit hídrico
(línea azul continua). Se puede observar que la pendiente del PE muestra una tendencia
diferenciada en cada período de hidratación. La tendencia es cero para un estado de hidratación
continuado (entre el 10 y el 14 de octubre), posteriormente es positiva cuando la planta ha
entrado en estrés por restricción (entre el 14 y el 25 de octubre), y una vez se rehidrata la
tendencia pasa a ser negativa (después del 25 de octubre). Algunos efectos sobre los máximos
de la señal del PE parecen estar relacionados con los cambios en la temperatura y la radiación,
tal como se observa en el período de recuperación. De acuerdo con los resultados, un estado de
estrés en una planta leñosa puede ser caracterizado por una tendencia positiva en la señal del
PE. El comportamiento presentado por las especies experimentales bajo una condición
ambiental específica de radiación, temperatura e hidratación sugiere una posible coordinación
fisiológica generada por señales eléctricas sobre los diferentes órganos (Volkov et al., 2007,
Zimmermann et al, 2009), determinado por la magnitud e intensidad del estímulo (Oyarce y
Gurovich, 2011; Gurovich y Hermosilla, 2009; Ríos-Rojas et al, 2015) para la especie específica.
Figura 2. Correlación del PE con las variables ambientales para Persea americana.
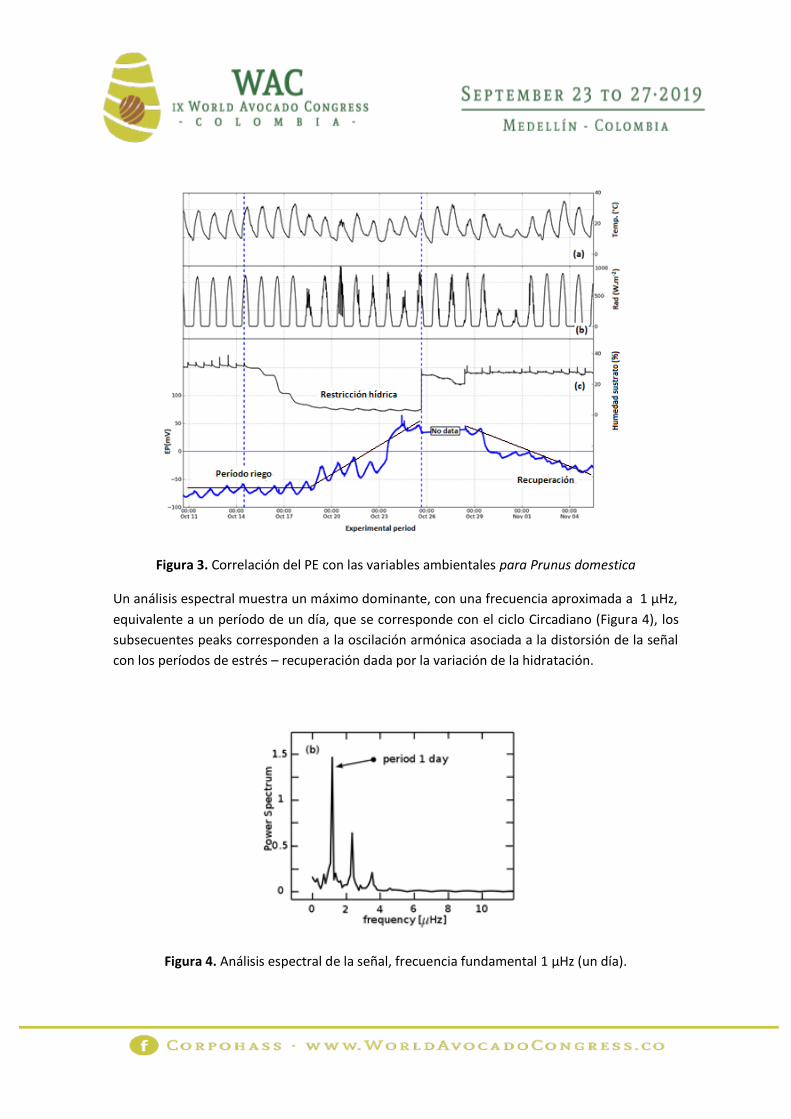
Figura 3. Correlación del PE con las variables ambientales para Prunus domestica
Un análisis espectral muestra un máximo dominante, con una frecuencia aproximada a 1 µHz,
equivalente a un período de un día, que se corresponde con el ciclo Circadiano (Figura 4), los
subsecuentes peaks corresponden a la oscilación armónica asociada a la distorsión de la señal
con los períodos de estrés – recuperación dada por la variación de la hidratación.
Figura 4. Análisis espectral de la señal, frecuencia fundamental 1 µHz (un día).

La respuesta para plantas de Persea americana, medidas con sensor electrofisiológico (línea
negra) y el sensor Keithley (línea roja), se muestran en la figura 5a. Los resultados exponen una
respuesta cíclica diaria (24 horas) en plantas bien regadas (panel izquierdo), siendo, 0,01 el valor
de la frecuencia más alta (Figura 5b,c); con un coeficiente de correlación entre los dos período-
gramas es de 0,79, [P<0.001; CI (intervalo de confianza): 0,75 – 0,82, al 95% de significancia];
esto indica que plantas estarían respondiendo a los estímulos ambientales incluidos la luz,
temperatura, radiación, etc. de ese periodo. Un segundo máximo, cercano a la frecuencia 0.02,
es detectado, con una frecuencia de menor magnitud, comparado con el espectro de la
frecuencia fundamental; correspondiendo a un comportamiento “típico” de las plantas de
Persea americana, ya reportado en Ríos-Rojas et al., (2014), para plantas bien regadas. Los
espectros resultantes de frecuencias de menor valor a 0,01 se refieren al período completo
analizado. En el caso del período con restricción hídrica (panel derecho), el multi-voltímetro
Keithley no expresa ciclos exactos de 24 horas, siendo la frecuencia dominante un ciclo de 1,1
días y la segunda frecuencia de 0,78 días (Figura 5b). Con el sensor electrofisiológico se registra
un ciclo de 0,5 días, precedido por un ciclo de 1,0 día (Figura 5c). La correlación para las señales
de los dos sensores es de 0,55, [P<0.001; CI: 0.47 – 0.62, 95%].
Figura 5. PE para Persea americana en período con restricción hídrica. (a) sensor Keithley (línea
roja), sensor electrofisiológico (línea negra); (b) Periodograma sensor Keithley. (c) Periodograma
sensor electrofisiológico. Panel izquierdo: plantas bien regadas; panel derecho: plantas en
restricción hídrica.
Con el fin de verificar el estado de estrés de las plantas experimentales, una vez ha pasado un
período de 23 días en restricción hídrica, se realizan mediciones de conductancia estomática en
tres horas distintas del día (9:00 h, 13:30 h y 17:30 h), durante un día, con estas mediciones se
busca verificar un cambio fisiológico, expresado en la respuesta del PE, por concepto de déficit
hídrico y no por cambios en la luz o la temperatura ambientales. En el período de riego se miden
10 plantas, que reciben el mismo tratamiento de hidratación; posteriormente 5 de las 10 plantas
son sometidas a restricción hídrica durante. Finalizado el período de restricción se realizan las

mediciones de Gs, en todas las plantas; observándose que la Gs se mantiene en un nivel
constante para las 5 plantas siempre regadas (“I”, figura 6a), mientras que, para las 5 plantas
sometidas a restricción hídrica, Gs disminuye significativamente (“NI”, figuras 6b).
Figura 6. Gs para Persea americana. (a) 10 plantas con riego. (b). 5 plantas con riego y 5
plantas sin riego.
La tabla 1a muestra las diferencias significativas para Persea americana para los valores de Gs,
A y Ci, para ambos tratamientos de disponibilidad hídrica a lo largo del día, con excepción de las
mediciones hechas para “A” a las 17:30 PM, coincidiendo con la hora de baja asimilación de CO2.
También para Prunus domestica (Tabla 1b), Gs y Ci presentan diferencias estadísticas
significativas entre tratamientos, excepto para A, cuya distribución no presenta diferencias.
Tabla 1. Prueba de Mann Whitney para A, Gs y Ci para (a) Persea Americana, (b) Prunus
domestica

Los resultados obtenidos en este período experimental son consistentes con los publicados en
Ríos-Rojas et al. (2014), donde un déficit hídrico prolongado genera un estrés en las plantas que
conllevan a un cambio en la tendencia del PE. Estos cambios en el PE pueden ser captados por
un sensor electrofisiológico que permita tomar medidas tempranas para mantener el buen
estado de las plantas leñosas. Plantas bien regadas presentan una respuesta cíclica, con un solo
máximo en el día (24 horas). En los períodos de restricción hídrica el ciclo se altera, ampliando
la frecuencia fundamental a más de 24 horas, verificado para ambas especies. Se observó que
el prototipo del sensor electrofisiológico amplificó adecuadamente la señal, tomando como
referencia la señal medida con el multi-voltímetro Keithley; un avance tecnológico en este
sentido permitiría instalar medidores de estrés fisiológico en los cultivos, haciendo un
seguimiento continuado, no destructivo que permita tomar decisiones tempranas sobre la
hidratación de las plantas. Las mediciones de Gs, permite evidenciar el efecto del estrés por
déficit hídrico, en tanto que plantas sometidas a restricción minimizan su Gs, a fin de evitar
grandes pérdidas de agua desde los estomas; éste comportamiento es claramente mostrado en
los resultados, y contrastado con plantas bien regadas. Es posible además observar en función
del PE que el proceso de recuperación después de la rehidratación no inicia de manera
instantánea, ésta podría estar relacionado a procesos como la transpiración y fotosíntesis que
tardan días en recuperarse después de una rehidratación (Flexas et al., 2009 and Zufferey et al.,
2011).
Conclusiones
Se presenta evidencia de la respuesta eléctrica de las plantas a estímulos ambientales. La
magnitud de la respuesta a estos estímulos, incluso a estrés por sequía pueden ser medidos en
las plantas, de manera no destructiva y en tiempo real. Las medidas del PE muestran el cambio
de pendiente en la señal, definido como un efecto del micro-ambiente sobre la planta, incluido
su estado hídrico, para las especies estudiadas, Persea americana y Prunus domestica; sin
embargo, estos resultados no son suficiente evidencia para generalizar a todas las especies
leñosas. La tecnología actual para realizar estas mediciones fisiológicas es usada sólo en
investigación, no está disponible para los productores por su alto costo o requerir de mano de
obra calificada para su operación, un equipo inalámbrico de menor tamaño sería apropiado para
realizar las mediciones del PE de manera continuada no destructiva, presentado en esta
investigación como un prototipo de sensor electrofisiológico. Cabe mencionar que el sensor
electrofisiológico no es recomendado como instrumento único para tomar decisiones en el
programa de riego; pues puede indicar cuándo la planta entra en estrés hídrico, más no puede
cuantificar el volumen de agua a aplicar a la planta, por lo cual no puede ser aislado de un sensor
que determine la humedad del suelo.

Referencias
Baluška F, Mancuso S. (2009). Plant neurobiology: From stimulus perception to adaptive behavior of plants, via integrated chemical and electrical signaling. Plant Signal Behav; 4: 475-76.
Bose J. C. (1926). The Nervous Mechanism of Plants. London, UK: Longmans, Green and Co; 123-34.
Brenner E, Stahlberg R, Mancuso S, Vivanco J, Baluška F, Van Volkenburgh E. (2006). Plant neurobiology: an integrated view of plant signaling. Trends Plant Sci; 8: 413-9.
Burdon-Sanderson J. 1872. Note on the electrical phenomena which accompany irritation of the leaf of Dionaea muscipula. Proc R Soc Lond; 21: 495-6.
CID Bio-Science Inc. Recuperado de: http://cid-inc.com/photosynthesis-system/ci-340-accessories. Consultado en enero, 2016.
Darwin C. (1896). The Power of Movements in Plants. New York. D. Appleton and Co; 625 pp.
Cabral E, Pecora P, Céspedes A, Bruno A, Costa E. (2011). The oscillatory bioelectrical signal from plants explained by a simulated electrical model and tested using Lempel–Ziv complexity. Comput Electron Agr; 76: 1-5.
Fromm J. (2006). Long-Distance Electrical Signaling and Physiological Functions in Higher Plants.Springer-Verlag Berlin Heidelberg.
Fromm J, Lautner S. (2007). Electrical signals and their physiological significance in plants. Plant Cell Environ; 30: 249-57.
Gurovich L, Hermosilla P. (2009). Electric signaling in fruit trees in response to water applications and light–darkness conditions. J Plant Physiol; 166: 290-300.
Heil M, Ton J. (2008). Review. Long-distance signaling in plant defence. Trends Plant Sci; 13: 264-72.
Kai L, Xi G, Fan L, Meng X, Gao X, Liu Q. (2011). The changes of electrical signals in corn at different temperatures. Proc. 3rd International Conference on Environmental Science and Information Application Technology (ESIAT). In; Procedia Environmental Sciences 2011; 10: 39-44. Berlin Heidelberg. Elsevier.
Kasuya, E. (2001). Mann‐Whitney U test when variances are unequal. Anim Behav 61:1247‐1249.
Khaliq M, Ouarda T. (2007). On the critical values of the standard normal homogeneity test (SHNT). Int JClimatol; 27:681-687
Mullendore D, Windt C, Van As H, Knoblaucha M. (2010). Sieve tube geometry in relation to phloem flow. Plant Cell; 22: 579-93.

Gibert D, Le Mouel J, Lambs L, Nicolin F, Perrier F. (2006). Sap flow and daily electric potential variations in a tree trunk. Plant Sci. 171:572–84.
Gil P., Gurovich L., Schaffer B., García N., Iturriaga R. (2009). Electrical signaling, stomatal conductance, ABA and ethylene content in avocado trees in response to root hypoxia. Plant Signal Behav. 4:100-8.
Grams T., Koziolek C., Lautner S., Matyssek R., Fromm J. (2007). Distinct roles of electric and hydraulic signals on the reaction of leaf gas exchange upon re-irrigation in Zea mays L. Plant Cell Environ. 30:79-84.
Gurovich L., Hermosilla P. (2009). Electric signalling in fruit trees in response to water applications and light–darkness conditions. Journal of Plant Physiology. 166:3: 290–300.
Gurovich L. (2012). Electrophysiology of Woody Plants, Electrophysiology - From Plants to Heart, Saeed Oraii (Ed.), ISBN: 978-953-51-0006-5, In Tech Publishers.
Mancuso E. (1999). Hydraulic and electrical transmission of wound-induced signals in Vitis vinifera. Australian Journal of Plant Physiology 26: 55 – 61.
Oyarce P., Gurovich L. (2010). Electrical signals in avocado trees. Responses to light and water availability conditions. Plant Signaling and Behavior 5: 34-41.
Oyarce P., Gurovich L. (2011). Evidence for the transmission of information through electric potentials in injured avocado trees. Journal of Plant Physiology. 168:2:103–108.
Pavlovic A., Mancuso S. (2011). Electrical signaling and photosynthesis Can they co-exist together? Plant Signaling & Behavior. 6:6, 840-842.
Ríos-Rojas L, Tapia F, Gurovich L. (2014). Electrophysiological assessment of water stress in fruit-bearing woody plants. J Plant Physiol; 171:799-806.
Ríos-Rojas L, Morales D, Alcalde J, Gurovich L. (2015). Use of plant woody species electrical potential for irrigation scheduling. Plant Signaling and Behavior. 10.
Rodríguez-Gamir J, Ancillo G, González M, Primo-Millo E, Iglesias D, Fomer G. (2011). Root signaling and modulation of stomata closure in flooded citrus seedlings. Plant Physiol Biochem; 49: 636-45.
Schroeder J, Hedrich R. (1989). Involvement of ion channels and active transport in osmoregulation and signaling of higher plant cells. Trends Biochem Sci; 14: 187-92.
Van Bel A, Ehlers K. (2005). Electrical signaling via plasmodesmata. In Plasmodesmata (ed. KJ Oparka). Annual Plant Reviews. Oxford, UK:Blackwell Publishing; 18: 263-78.
Volkov A., Lang L., Volkova-Gugeshashvili M. (2007). Electrical signaling in Aloe Vera induced by localized thermal stress. Bioelectrochemistry. 71:192–197

Volkov A. (2012). Plant electrophysiology. Signaling and responses. Springer - Verlag. Berlin Heidelberg. pp 390.
Volkov A., Wait, A., Wooten J., Markin V. (2012). Circadian rhythms in biologically closed electrical circuits of plants. Plant Signaling and Behavior; 168: 1753-60.
Wagner E, Lehner L, Veit J, Normann J, Albrechtová J. (2012). Biosystems analysis of plant development concerning photoperiodic flower Induction by hydro-electrochemical signal transduction. Plant Electrophysiology. Berlin Heidelberg: Springer Verlag.
Wang Z., Leng Q., Huang L., Zhao L., X, z., Hou R., Wan C. (2009). Monitoring system for electrical signals in plants in the greenhouse and its applications. biosystems engineering103:1–11.
Zimmermann M., Maischak H., Mithofer A., Boland W., Felle H. (2009). System Potentials, a Novel Electrical Long-Distance Apoplastic Signal in Plants, Induced by Wounding. Plant Physiology. 149:1593–16