resource acquisition and transport in vascular plants in vascular plants . overview: underground...
TRANSCRIPT

Chapter 36
Resource Acquisition and Transport in Vascular Plants

Overview: Underground Plants
• The success of plants depends on their ability to
gather and conserve resources from their
environment
– Many plants have become highly proficient in
acquiring resources that are limited in their
environments
– The transport of these materials is also critical
to the integrated functioning of the whole plant
• Diffusion, active transport, and bulk flow
work together to transfer water, minerals,
and sugars

Concept 36.1: Land plants acquire resources both above and below ground
• The algal ancestors of land plants absorbed water, minerals, and CO2
directly from the surrounding water
– Transport in these algae was relatively simple because each cell was
close to the surface of the organism, and thus the water
– The evolution of xylem and phloem in land plants made possible the
long-distance transport of water, minerals, and products of
photosynthesis to support extensive root
and shoot systems
• Today, evolution has resulting in many
mechanisms for acquiring sunlight,
CO2, and water, while at the same
time minimizing evaporative loss
of water
Fig. 36-2-3
H2O
H2Oand minerals
CO2 O2
O2
CO2
Sugar
Light

Shoot Architecture and Light Capture
• In shoot systems, stems serve as conduits for water and
nutrients, and also act as supporting structures for leaves
– Shoot systems can vary in terms of:
• Their form and arrangement of leaves
– Leaf size (1.3 mm – 20 m)
• The largest leaves are usually found in tropical
rainforests, where water is plentiful and
evaporative loss is not a problem
– Phyllotaxy: the arrangement of leaves on a stem
• The outgrowth of axillary buds
• The relative growth of stem length and thickness

Phyllotaxy
• Phyllotaxy is determined by the shoot apical meristem and is specific to
each species:
– Alternate (spiral) phyllotaxy: one leaf per node
– Opposite phyllotaxy: two leaves per node
– Whorled phyllotaxy: many leaves per node
• Most angiosperms have alternate phyllotaxy, with their leaves arranged in
an ascending spiral around the stem
– This allows each leaf to get the maximum exposure to light and
reduces shading of lower leaves by those above
• In environments where intense
sunlight can harm leaves, the
greater shading provided by
oppositely-arranged leaves may
be more advantageous

• Light absorption is affected by the leaf area index, the ratio of total upper
leaf surface of a plant divided by the surface area of land on which it grows
– Leaf area index values of up to 7 are common for many mature plants
– There is little agricultural benefit to leaf area indexes higher than 7
• Adding more leaves increases shading of lower leaves to the point
that they respire more than photosynthesize
• Under these circumstances, nonreproductive leaves or branches
undergo programmed
cell death, known as
self-pruning
Fig. 36-4
Ground area
covered by plant
Plant ALeaf area = 40%of ground area
(leaf area index = 0.4)
Plant BLeaf area = 80%of ground area
(leaf area index = 0.8)

• Leaf orientation also affects light absorption
– In low-light conditions, horizontal leaves capture sunlight
more effectively than vertical leaves
• In sunny regions, however, horizontal orientation may
expose upper leaves to intense light, resulting in injured
leaves and reduced photosythesis
– If leaves are vertical instead, light rays will essentially be
parallel to the leaf surfaces, so no leaf receives too much
light
• This arrangement also allows light to penetrate more
deeply to the lower leaves

• Other factors also contribute to the ecological success of plants
– Plants have a finite amount of energy they can devote to shoot
growth
• If they put most of their energy into branching, they have less
to devote towards growing tall and are thus at risk of being
shaded by taller plants
• If plants put all their energy into growing tall, they are not
optimally exploiting the resources above-ground
– Plant species also vary in stem thickness
• Most tall plants require thick stems that allow greater
vascular flow to leaves and more mechanical support
• In woody plants, stems become thicker through secondary
growth

Root Architecture and Acquisition of Water and Minerals
• Soil contains resources acquired by the root system
– The evolution of root branching allowed plants to obtain
water and minerals more effectively
• Taproot systems with numerous branches anchor plants
and are characteristic of most trees
• Most monocots do not reach tree-like heights because
their fibrous root systems do not anchor them as
strongly
– Evidence suggests that physiological mechanisms reduce
competition within the root system of a plant
• Ex) Cutting from the same plant develop fewer and
shorter roots in the presence of one another than they
did in the presence of cuttings from a different plant

Root Architecture and Acquisition of Water and Minerals
• The evolution of mutualistic associations between roots and fungi
called mycorrhizae was also a critical step in the successful
colonization of land by plants
– Mycorrhizal hyphae provide the fungus and plants roots with a
large surface area for absorbing water and minerals
• As much as 3 meters of
hyphae can extend from
each centimeter along a
root’s length, allowing
access to a greater
volume of soil than the
root alone could
penetrate
Fig. 36-5
2.5 mm

Concept 36.2: Transport occurs by short-distance diffusion or active transport and by long-distance bulk flow
• Transport begins with the absorption of resources by plant cells
– The movement of substances into and out of cells is
regulated by selective permeability of the plasma
membrane
• Solutes tend to diffuse down their electrochemical gradient, the
combined effect of:
– The solute’s concentration gradient
– The voltage (charge difference) across the membrane

Concept 36.2: Transport occurs by short-distance diffusion or active transport and by long-distance bulk flow
• Diffusion across a membrane is passive, while the pumping of
solutes across a membrane against their electrochemical
gradients is active and requires energy
– Whether or not the process requires energy, most solutes
pass through transport proteins embedded in the cell
membrane
• In some cases, transport proteins selectively bind a
solute on one side and the change shape, releasing the
solute on the opposite side
• Other transport proteins provide selective channels
across the membrane, some of which (gated channels)
open or close in response to stimuli

• The most important transport protein for active transport in plant cells is the
proton pump
– Proton pumps use energy from ATP to pump protons (H+) out of the
cell
– This movement results in an H+ gradient, with a higher H+
concentration outside the cell than inside
• This potential (stored) energy can be harnessed to do work as H+
flows back into the cell
– Movement of H+ out of the cell also makes the inside of the cell
negative in charge relative to the outside
• This charge separation across the membrane contributes to a
voltage called membrane potential
– Membrane potential is
another form of potential
energy that can be
harnessed to do
cellular work
Fig. 36-6
CYTOPLASM EXTRACELLULAR FLUID
ATP
H+
H+
H+
H+
H+
H+
H+
H+
H+
Proton pump
generates mem-
brane potential
and gradient.
+
+
+
+
+
_
_
_
_
_

• Plant cells use energy stored in the proton gradient and membrane potential
to drive the transport of many different solutes
– Ex) The membrane potential
generated by proton pumps
contributes to the absorption
of K+ by root cells
– Ex) In a form of cotransport, a
transport protein couples the
diffusion of one solute (H+)
with active transport of
another (NO3- or sucrose)
Fig. 36-7
CYTOPLASM EXTRACELLULAR FLUID
K+
K+
K+
K+
K+
K+
K+
Transport protein
_
_
_
_
+
_
+
+
+
+
(a) Membrane potential and cation uptake
NO 3?
NO3?
NO 3? NO
3?
NO3?
NO3
?
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
(b) Cotransport of an anion with H+
H+
_
_
_
_
_
_
+
+
+
+
+
+
H+ H+H+
H+
H+H+
H+
H+
H+
H+
H+ H+
_
_
_
_
_
_
+
+
+
+
+
+
(c) Cotransport of a neutral solute with H+
S
SS
S
S
S

Diffusion of Water (Osmosis)
• To survive, plants must balance water uptake and loss
– The net uptake of water by a cell is determined by the diffusion of water
across the membrane, a process called osmosis
• In animal cells, water will move from the solution with lower solute
concentration to higher solute concentration if the plasma
membrane is impermeable to the solute
• Because a plant cell has almost rigid cell walls, however, the
physical pressure of that wall pushing back against the cell adds
another factor that affects osmosis
– Water potential is a measurement that combines the effects of solute
concentration and pressure, which together determine the direction of
movement of water
• The term water potential refers to water’s potential energy (capacity
to perform work) when it moves from a region of higher water
potential to a region of lower water potential

• Water potential is abbreviated as Ψ (psi) and
measured in units of pressure called megapascals
(MPa)
• Ψ = 0 MPa for pure water at sea level (in a
container open to the atmosphere) and room
temperature
• One Mpa is equal to about 10 atmospheres of
pressure (~ 1 kg pressure/cm2)
• The internal pressure of a plant cell is ~0.5
MPa

How Solutes and Pressure Affect Water Potential
• Both pressure and solute concentration affect water potential:
Ψ = ΨS + ΨP
• The solute potential (ΨS) of a solution is proportional to the number of
dissolved molecules (molarity)
• Solute potential is also called osmotic potential because solutes
affect the direction of osmosis
• By definition, ΨS = 0 for pure water, since there are no dissolved
molecules
• As solutes are added, they bind to water molecules, reducing the
number of free water molecules
• This lowers the capacity of water to move and do work, meaning
that:
• Adding solutes always lowers water potential
• ΨS for a solution is always negative

How Solutes and Pressure Affect Water Potential
• Both pressure and solute concentration affect water potential:
Ψ = ΨS + ΨP
• Pressure potential (ΨP) is the physical pressure on a solution
• Unlike ΨS, ΨP can be positive or negative relative to atmospheric
pressure
• Turgor pressure is the pressure exerted by the plasma membrane
against the cell wall, and the cell wall against the protoplast (the living
part of the cell, including nucleus, cytoplasm, and plasma membrane)
• The contents of the living cell are usually under positive pressure

Measuring Water Potential
• Consider a U-shaped tube where the two arms are separated by a
membrane permeable only to water
• Keep in mind: water moves in the direction from higher water potential
to lower water potential
• The right arm contains 0.1M solution (ΨS = -0.23 MPa)
• The left arm contains pure water (ΨS = 0)
• There is no physical pressure because it is an
open system (ΨP = 0)
• Thus, Ψ = ΨS + ΨP = ΨS + 0 = ΨS =
-0.23 MPa for the right arm
• For the left arm, Ψ = ΨS + ΨP = 0 + 0 =
0 MPa
• Because water moves from regions of lower water
potential, the net water movement will be from the
left arm to the right arm
ψ = ?0.23 MPa
Fig. 36-8a
(a)
0.1 M
solution
Purewater
H2O
ψP = 0
ψS = 0ψP = 0
ψS = ?0.23
ψ = 0 MPa

• Physical pressure increases water potential
– Applying a positive physical pressure of
+0.23 MPa to the solution in the right arm raises
its water potential from a negative value to 0 MPa
Ψ = ΨS + ΨP = -0.23 + 0.23 = 0 MPa
– There is now no net flow of water between
this pressurized solution and the
compartment of pure water
• If we increase ΨP even further to +0.3 MPa in the
right arm, then the solution has a water potential of
+0.07 MPa
Ψ = ΨS + ΨP = -0.23 + 0.3 = +0.7 MPa
– This solution will actually lose water to a
compartment containing pure water
Fig. 36-8b
(b)Positive
pressure
H2O
ψP = 0.23
ψS = ?0.23ψP = 0
ψS = 0
ψ = 0 MPa ψ = 0 MPaFig. 36-8c
ψP =
ψS = ?0.23
(c)
Increasedpositivepressure
H2O
ψ = 0.07 MPa
ψP = 0
ψS = 0
ψ = 0 MPa
0.30

• Negative pressure (tension) decreases water potential
– A negative pressure potential of –0.30 MPa reduces the water potential
of the pure water compartment enough so that water is drawn from the
solution on the right side
Ψ = ΨS + ΨP = 0 + -0.30 = -0.30 MPa
Fig. 36-8d
(d)
Negativepressure(tension)
H2O
ψP = ?0.30
ψS =
ψP =
ψS = ?0.23
ψ = ?0.30 MPa ψ = ?0.23 MPa
0
0

• Water potential also affects uptake and loss of water by plant cells
– If a flaccid (limp) cell (ΨP = 0) is placed in an environment with a
higher solute concentration, the cell will lose water and undergo
plasmolysis (the protoplast shrinks and pulls away from the cell
wall)
Ψ = ΨS + ΨP = -0.9 + 0 = -0.9 MPa (sucrose solution)
Ψ = ΨS + ΨP = -0.7 + 0 = -0.7 MPa (initial flaccid cell)
Video: Plasmolysis
Fig. 36-9a
(a) Initial conditions: cellular ψ > environmental ψ
ψP = 0 ψS = ?0.9
ψP = 0 ψS = ?0.9
ψP = 0ψS = ?0.7
ψ = ?0.9 MPa
ψ = ?0.9 MPa
ψ = ?0.7 MPa0.4 M sucrose solution:
Plasmolyzed cell
Initial flaccid cell:

• If the same flaccid cell is placed in pure water (Ψ = 0), the
contents of the cell becomes turgid, swelling as water enters it
Ψ = ΨS + ΨP = 0 + 0 = 0 MPa (pure water)
Ψ = ΨS + ΨP = -0.7 + 0 = -0.7 MPa (initial flaccid cell)
Video: Turgid Elodea
Fig. 36-9b
ψP = 0ψS = ?0.7
Initial flaccid cell:
Pure water:ψP = 0ψS = 0
ψ = 0 MPa
ψ = ?0.7 MPa
ψP = 0.7ψS = ?0.7
ψ = 0 MPa
Turgid cell
(b) Initial conditions: cellular ψ < environmental ψ

• Turgor loss in plants causes wilting, during which
the leaves and stems droop as a result of water
loss
– Wilting can be reversed when the plant is
watered Fig. 36-10

Aquaporins: Facilitating Diffusion of Water
• Although water molecules are small enough to diffuse
across the phospholipid bilayer, their movement is too
rapid to be explained by passive transport
– Aquaporins are transport proteins in the cell
membrane that facilitate the passage of water
• These selective channels affect the rate at which
water diffuses down its water potential gradient
– The rate of water movement is likely regulated by
phosphorylation of these aquaporin proteins
• Phosphorylation can be induced by increases in
cytoplasmic calcium ions or decreases in
cytoplasmic pH

Three Major Pathways of Transport
• Transport is also regulated by the compartmental structure of plant cells
– The cell wall, cytosol, and vacuole are the 3 main compartments of
most mature plant cells
• Transport proteins in the plasma membrane regulate traffic of
molecules between the cytosol and the cell wall
• Transport proteins in
the vacuolar
membrane regulate
traffic of molecules
between the cytosol
and the vacuole
Fig. 36-11
Cell wall
Cytosol
Vacuole
Plasmodesma Vacuolar membranePlasma membrane
(a) Cell compartments Key
Transmembrane routeApoplast
SymplastApoplast
Symplast
Apoplast
Symplastic route
Apoplastic route
(b) Transport routes between cells

Three Major Pathways of Transport
• Ions can diffuse across tissues entirely through the continuum, known as the
apoplast, formed by cell walls, extracellular spaces, and the dead interiors
of tracheids and vessels
– The plasma membrane directly controls the traffic of molecules into and
out of the protoplast
• It is a barrier between two of the major compartments, the cell wall
and the cytosol
– The continuum formed
from the cytosol of cells
is collectively referred
to as the symplast
– Cytoplasmic channels
called plasmodesmata
connect the cytoplasm
of neighboring cells
Fig. 36-11
Cell wall
Cytosol
Vacuole
Plasmodesma Vacuolar membranePlasma membrane
(a) Cell compartments Key
Transmembrane routeApoplast
SymplastApoplast
Symplast
Apoplast
Symplastic route
Apoplastic route
(b) Transport routes between cells
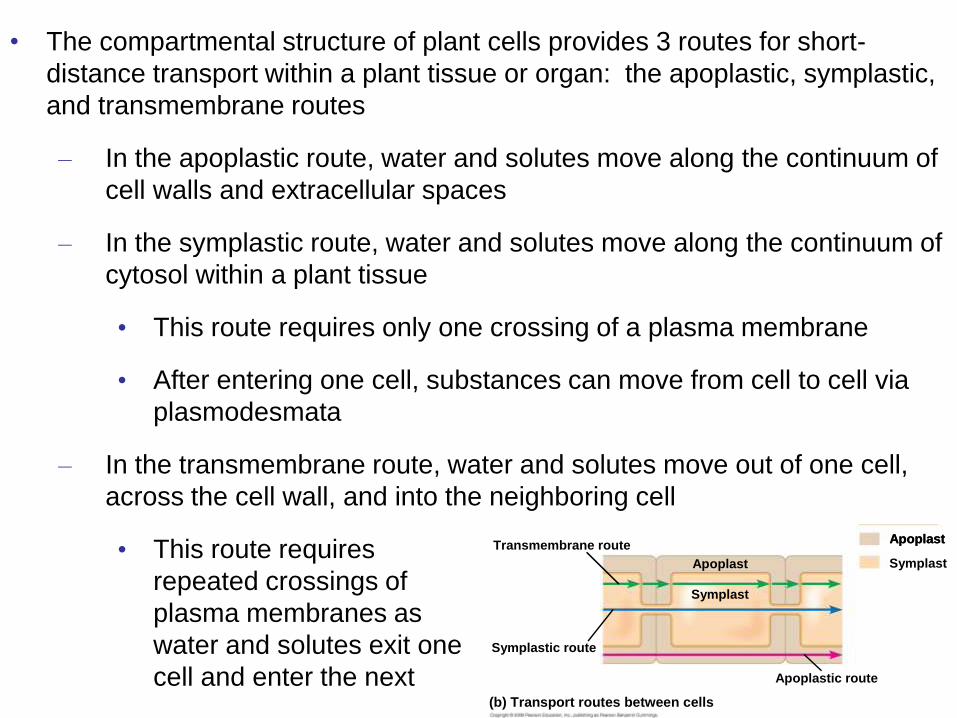
• The compartmental structure of plant cells provides 3 routes for short-
distance transport within a plant tissue or organ: the apoplastic, symplastic,
and transmembrane routes
– In the apoplastic route, water and solutes move along the continuum of
cell walls and extracellular spaces
– In the symplastic route, water and solutes move along the continuum of
cytosol within a plant tissue
• This route requires only one crossing of a plasma membrane
• After entering one cell, substances can move from cell to cell via
plasmodesmata
– In the transmembrane route, water and solutes move out of one cell,
across the cell wall, and into the neighboring cell
• This route requires
repeated crossings of
plasma membranes as
water and solutes exit one
cell and enter the next
Fig. 36-11
Cell wall
Cytosol
Vacuole
Plasmodesma Vacuolar membranePlasma membrane
(a) Cell compartments Key
Transmembrane routeApoplast
SymplastApoplast
Symplast
Apoplast
Symplastic route
Apoplastic route
(b) Transport routes between cells

Bulk Flow in Long-Distance Transport
• Efficient long distance (roots to stems and leaves) transport of fluid requires
bulk flow, the movement of a fluid driven by pressure
– Water and solutes move together through tracheids and vessel
elements of xylem, and sieve-tube elements of phloem
• Efficient movement is possible because mature tracheids and
vessel elements have no cytoplasm, and sieve-tube elements have
few organelles in their cytoplasm
• Bulk flow is also enhanced by the perforation plates at the ends of
vessel elements and the porous sieve plates connecting sieve-tube
elements
• Diffusion, active transport, and bulk flow act together to transport resources
throughout the whole plant
– Ex) Bulk flow due to a pressure difference is how long-distance
transport of sugars occurs from the phloem, but active transport of
sugar at the cellular level maintains this pressure difference

Concept 36.3: Water and minerals are transported from roots to shoots
• Even though they have no pumping mechanism, plants can move a large
volume of water from their roots to shoots
• Most water and mineral absorption occurs near root tips
• Here, the epidermal cell are permeable to water
• In addition, many of these epidermal cells are differentiated into
root hairs that account for much of the absorption of water by roots
• When soil solution (water and dissolved minerals) enters the roots, it first
flows into the hydrophilic walls of the epidermal cells and passes freely
along the extracellular spaces into the root cortex
• The exposure of these cortical cells to soil solution provides a greater
membrane surface area for absorption than the surface area of the
epidermal cells alone
• Active transport then allows roots to concentrate essential
minerals up to 100s of times higher than in the soil
Animation: Transport in Roots

• Before water and minerals can be transported to the rest of the plant, they
must enter the xylem of the vascular cylinder (stele)
• The endodermis (the innermost layer of cells in the root cortex)
surrounds the stele and acts as a last checkpoint for selective passage
of minerals from the cortex into the vascular tissue
• Water and minerals can reach the endodermis of the cortex in one of 2
ways:
• Through the symplast
(cytosolic continuum)
• Through the apoplast
(continuum of cell walls
and extracellular spaces)
Transport of Water and Minerals into the Xylem
Fig. 36-12a
Casparian strip
Plasma
membrane
Apoplastic
route
Symplastic
routeRoot
hair
Epidermis
Cortex
Endodermis
Vessels
(xylem)
Stele
(vascular
cylinder)

• Minerals already in the symplast when they reach the endodermis move
through plasmodesmata of the endodermal cells and into the stele
• Minerals that reach the endodermis via the apoblast, however, meet a
barrier called the Casparian strip, located in the walls of endodermal
cells
• It is made of suberin, a waxy material impervious to water and
dissolved minerals
• The strip forces water
and minerals in the
apoblast to cross the
plasma membrane of
the endodermal cells
and enter the stele via
the symplast
Transport of Water and Minerals into the Xylem
Fig. 36-12
Pathway along
apoplast
Casparian strip
Endodermal cell
Pathway
through
symplast
Casparian strip
Plasma
membrane
Apoplastic
route
Symplastic
routeRoot
hair
Epidermis
Cortex
Endodermis
Vessels
(xylem)
Stele
(vascular
cylinder)

• The endodermis, along with its Casparian strip, ensures that no minerals
can reach the root’s vascular tissue without crossing a selectively permeable
plasma membrane
• It also prevents solutes that have accumulated in the xylem of the stele
from leaking back into the soil solution
• Once in the xylem, endodermal cells discharge water and minerals into their
walls (apoblast), where it can pass into the xylem’s tracheids and vessel
elements
• The xylem vessels then
transport the water and
dissolved minerals upward
to the shoot system by
bulk flow
Fig. 36-12a
Casparian strip
Plasma
membrane
Apoplastic
route
Symplastic
routeRoot
hair
Epidermis
Cortex
Endodermis
Vessels
(xylem)
Stele
(vascular
cylinder)

Bulk Flow Driven by Negative Pressure in the Xylem
• Plants lose a large volume of water from
transpiration, the evaporation of water from a plant’s
surface
– Ex) A single maize plant transpires 60 L of water
during one growing season
• This water is replaced by the bulk flow of water
and minerals, called xylem sap, from the steles
of roots to the stems and leaves
– Is sap mainly pushed up from the roots, or pulled
up by the leaves?

Pushing Xylem Sap: Root Pressure
• At night, when transpiration is very low, root cells continue pumping mineral
ions into the xylem of the stele
– The accumulation of minerals lowers the water potential within the stele
• Because water flow towards areas of lower water potential, water
flows in from the root cortex and generates root pressure, a push
of xylem sap
– Root pressure sometimes results in guttation, the exudation of water
(not dew) droplets on tips or edges of leaves
• In most plants, root pressure
is relatively weak and
therefore only a minor
mechanism driving
xylem sap ascent
Fig. 36-13

Pulling Xylem Sap: The Transpiration-Cohesion-Tension Mechanism
• For the most part, xylem sap is pulled upward
by negative pressure in the xylem of leaves
– Transpiration provides the pull
– Cohesion of water due to hydrogen bonding
transmits this pull along the entire length of
the xylem to the roots

• Transpiration produces negative pressure (tension) in the leaf, which exerts
a pulling force on water in the xylem, pulling water into the leaf
• 1) Water vapor (blue dots) diffuses from moist air spaces of the leaf to
drier air outside via stomata
• This is due to the tendency of water to move toward areas of lower
water potential
• 2) Water vapor lost
by transpiration is
replaced by
evaporation from
the water film that
coats mesophyll
cells
Fig. 36-14
Cuticle Xylem
Upper
epidermis
Mesophyll
Lower
epidermis
Cuticle
Airspace
Microfibrils incell wall of
mesophyll cell
Stoma
Microfibril(cross section)
Waterfilm
Air-waterinterface

• 3) The evaporation of this water film causes the air-water interface to retreat
farther in to the cell wall and to become more curved
• This curvature increases the surface tension and the rate of
transpiration, pulling water from surrounding cells and air spaces
• 4) Water from the xylem is pulled into the surrounding cells and air spaces to
replace the water
that was lost
Fig. 36-14
Cuticle Xylem
Upper
epidermis
Mesophyll
Lower
epidermis
Cuticle
Airspace
Microfibrils incell wall of
mesophyll cell
Stoma
Microfibril(cross section)
Waterfilm
Air-waterinterface

• The transpirational pull on xylem sap is transmitted all the way from the
leaves to the root tips and even into the soil solution
• This pull is also facilitated by cohesion of water molecules to each other
and adhesion of water molecules to cell walls
• Cohesion of water due to H-bonding allows xylem sap to be pulled
from above without water
molecules separating
• Adhesion of water molecules
to the hydrophilic walls of
xylem cells, again due to
H-bonding, also helps
offset the downward force
of gravity
Animation: Transpiration
Animation: Water Transport
Fig. 36-15
Outside air ψ
= ?100.0 Mpa
Leaf ψ (air spaces)
= ?7.0 Mpa
Leaf ψ (cell walls)
= ?1.0 Mpa
Trunk xylem ψ
= ?0.8 Mpa
Trunk xylem ψ
= ?0.6 Mpa
Soil ψ
= ?0.3 Mpa
Xylemsap
Mesophyllcells
StomaStoma
Watermolecule
TranspirationAtmosphere
Adhesionby hydrogenbonding
Cellwall
Xylemcells
Cohesion andadhesion inthe xylem
Cohesionby hydrogenbonding
Watermolecule
Roothair
Soilparticle
WaterWater uptakefrom soil
Wate
r p
ote
nti
al g
rad
ien
t

• The formation of a water vapor pocket, called cavitation, can break the chain
of water molecules and hinder transpirational pull
• Cavitation occurs most often during drought or freezing
• Air bubbles resulting from cavitation expand and block water
channels of the xylem
• Small plants can refill their blocked vessels using root pressure in the
spring
• In trees, however, where root pressure is very weak (due to height),
a water vapor pocket usually can’t function in xylem sap transport
• Water can instead detour around the water vapor pocket
through pits between adjacent tracheids or vessels
• Also, as secondary growth adds a new layer of xylem each
year, the older layer of secondary xylem ceases to transport
water anyways

Xylem Sap Ascent by Bulk Flow: A Review
• The movement of xylem sap against gravity is
maintained by a combination of forces: transpiration,
cohesion, and tension
– Transpiration lowers water potential in leaves, and
this generates negative pressure (tension) that
pulls water up through the xylem
– There is no energy cost to bulk flow of xylem sap
• Absorption of sunlight drives most transpiration
by causing water to evaporate from the moist
cell walls of mesophyll cells

Concept 36.4: Stomata help regulate the rate of transpiration
• Leaves generally have broad surface areas and high surface-to-volume
ratios
– These characteristics increase photosynthesis (more light, CO2) but
also increase water loss through stomata (via transpiration)
• About 95% of the water a plant loses escapes through stomata
– The waxy cuticle limits water loss through the remaining leaf
surface
• Each stoma is flanked by a pair of guard cells, which control the
diameter of the stoma by
changing shape
– By opening and closing
stomata, guard cells
help balance the plant’s
environmental
requirements for
photosynthesis with
conservation of water
Fig. 36-16

Stomata: Major Pathways for Water Loss
• The amount of water lost by a leaf depends largely
on the number of stomata and the average size of
their pores
– Desert plants tend to have lower stomatal
densities than marsh plants
– Environmental condition can also influence
stomatal density
• Ex) High light exposure and low CO2 levels
during leaf development lead to increased
stomatal density in many species

Mechanisms of Stomatal Opening and Closing
• Changes in turgor pressure open and close stomata
– When guard cells are turgid as a result of uptake of water
via osmosis from nearby cells, stoma are open
– When the guard cells lose water and become flaccid, they
become less bowed and the pore closes
Fig. 36-17a
Guard cells turgid/Stoma open Guard cells flaccid/Stoma closed
Radially orientedcellulose microfibrils
Cellwall
VacuoleGuard cell
(a) Changes in guard cell shape and stomatal opening andclosing (surface view)

Mechanisms of Stomatal Opening and Closing
• Changes in turgor pressure result primarily from the reversible uptake and loss of potassium ions by the guard cells
– Stomata open when guard cells actively transport K+ ions from neighboring epidermal cells
• Energy for transport comes from cotransport of H+ ions (proton pump)
• As the guard cells accumulate K+ ions, this lowers water potential inside them and causes them to take up water and become turgid
– Stomatal closing results from loss of K+ from guard cells to neighboring cells
• This leads to an osmotic loss of water due to increased water potential (less solutes)
Fig. 36-17b
Guard cells turgid/Stoma open Guard cells flaccid/Stoma closed
(b) Role of potassium in stomatal opening and closing
H2O
H2OH2O
H2OH2O
H2O H2O
H2O
H2O
H2O
K+

Stimuli for Stomatal Opening and Closing
• Generally, stomata open during the day and close at night to minimize water
loss
– Stomatal opening at dawn is triggered by at least 3 cues:
• Light – stimulates guard cells to accumulate K+ and become turgid
– Triggered by the illumination of blue-light receptors in the plasma
membranes of guard cells
• This stimulates the activity of proton pumps (and in turn,
promotes K+ absorption)
• CO2 depletion as a result of photosynthesis – concentrations of CO2
decrease throughout the day
– This causes stomata to progressively increase in diameter (if
sufficient water is available)
• An internal “clock” in guard cells – ensures that stomata continue their
daily rhythm of opening and closing, even if a plant is kept in the dark
– All eukaryotic organisms have internal clocks; circadian rhythms
are 24-hour cycles

• Environmental stresses (ex: drought) can cause
stomata to close during the daytime
– If plants have a water deficiency guard cells may
lose turgor and close stomata
• In addition, a hormone produced in roots and
leaves in response to drought called abscisic
acid signals guard cell to close stomata
– This response reduces wilting but also
restricts CO2 absorption, thereby slowing
photosynthesis
• As a result, growth ceases, since turgor is
necessary for cell elongation

Effects of Transpiration on Wilting and Leaf Temperature
• Plants lose a large amount of water by transpiration
– Transpiration is greatest on days that are warm, sunny, dry,
and windy because these factors increase evaporation
• If the lost water is not replaced by sufficient transport of
water, the plant will lose water and wilt
– Transpiration can be beneficial on warm days, however,
since it also results in evaporative cooling, which can lower
the temperature of a leaf
• This cooling prevents denaturation of various enzymes
involved in photosynthesis and other metabolic
processes

Adaptations That Reduce Evaporative Water Loss
• Xerophytes are plants adapted to arid climates, like hot and dry deserts or
frozen regions where access to liquid water is problematic
– Many species of desert plants avoid drying out by completing their
short life cycles during the brief rainy seasons
• Longer-lived species have physiological or morphological
adaptations that allow them to withstand harsh desert conditions
– Cacti have highly reduced leaves that resist excessive water
loss, carrying out photosynthesis mainly in their stems
• The stems of these plants are fleshy because they store
water for use during prolonged dry periods
– Some plants use a specialized form of photosynthesis called
crassulacean acid metabolism (CAM) where stomatal gas
exchange occurs at night
• Because stomata remain closed during the day,
evaporative stresses are reduced

Concept 36.5: Sugars are transported from leaves and other sources to sites of use or storage
• Transpiration cannot meet all the long-distance
transport needs of plants
– Transport of sugars from leaves to roots (the
opposite direction of water and mineral
transport) requires another tissue called the
phloem
• The phloem transports these products of
photosynthesis the process of translocation

• In angiosperms, translocation occurs through specialized cells called sieve-
tube elements
– These cells are arranged end-to-end, forming long sieve tubes through
which sap can flow
• Phloem sap consists mainly of sugar (sucrose), along with smaller
amounts of amino acids, hormones, and minerals
– Phloem sap travels from a sugar source to a sugar sink
• A sugar source is an organ that is a net producer of sugar, such
as mature leaves
• A sugar sink is an organ that is a net consumer or storer of sugar,
such as roots, buds, stems, and fruits
– A storage organ, such as a tuber or bulb, can be both a sugar
sink in summer and sugar source in winter
– Sugar sinks usually receive sugar from the nearest sugar sources
• The upper leaves on a branch may export sugar to growing shoot
tips, whereas lower leaves may export sugar to roots

• Sugar must be loaded into sieve-tube elements before being
exposed to sinks
– Depending on the species, sugar may move by symplastic
(through plasmodesmata) or both symplastic and apoplastic
pathways
• In some plants, the walls of modified companion cells of
sieve-tube elements, known as transfer cells, have
ingrowths that enhance solute transfer between the apoblast
and symplast
Fig. 36-19a
Key
Apoplast
Symplast
Mesophyll cell
Cell walls (apoplast)
Plasma membrane
Plasmodesmata
Companion(transfer) cell
Sieve-tubeelement
Mesophyll cellBundle-sheath cell
Phloemparenchyma cell

• In many plants, phloem loading requires active transport because sucrose is
more concentrated in sieve-tube elements and companion cells than in
mesophyll
– Proton pumping and cotransport of sucrose and H+ enable the cells to
accumulate sucrose
• Once they reach the sink, sugar molecules diffuse down their concentration
gradient from the phloem to sink tissues
• The concentration of sugar is always lower in the sink because
unloaded is consumed rapidly
during growth and
metabolism or converted to
other polymers like starch
(storage plants)
Fig. 36-19b
High H+ concentration Cotransporter
Protonpump
Low H+ concentration
Sucrose
H+
H+ H+ATP
S
S

Bulk Flow by Positive Pressure: The Mechanism of Translocation in Angiosperms
• In studying angiosperms, researchers have concluded that sap
moves through a sieve tube by bulk flow driven by positive
pressure, known as pressure flow
– The building of pressure at the source end and reduction
of that pressure at the sink cause water to flow from
source to sink, carrying the sugar along
Animation: Translocation of Phloem Sap in Summer
Animation: Translocation of Phloem Sap in Spring

Bulk Flow by Positive Pressure: The Mechanism of Translocation in Angiosperms
• 1) Loading of sugar (green dots) into the sieve tube at the source reduces
water potential in these cells
– This causes the tube to take up water by osmosis
• 2) The uptake of water by sieve tubes generates a positive pressure that
forces sap to flow along the tube
• 3) This pressure is relieved by the
unloading of sugar, which in turn
causes loss of water at the sink via
osmosis
• 4) The xylem then recycles water from
sink to source
4
Fig. 36-20
3
2
1
1
2
34
Vessel(xylem)
Sieve tube(phloem)
Source cell(leaf) Loading of sugar
Uptake of water
Unloading of sugar
Water recycled
Sink cell(storageroot)
Sucrose
H2O
H2O
Bu
lk f
low
by n
eg
ati
ve p
res
su
re
H2O
Sucrose
Bu
lk f
low
by p
osit
ive p
ressu
re

Concept 36.6: The symplast is highly dynamic
• The symplast is a living tissue and is responsible for dynamic changes in
plant transport processes
– Plasmodesmata are dynamic in that they can change in permeability
and number
• They can open or close in response to changes in turgor pressure,
cytoplasmic calcium levels, or cytoplasmic pH
• Loss of function is also common during differentiation
– As a leaf matures from a sink (growing leaf) to a source
(mature leaf), its plasmodesmata either close or are eliminated
and phloem loading ceases
– In addition, plant viruses can cause plasmodesmata to dilate
• Plant viruses produce viral movement proteins that cause widening
of these pores to allow viral RNA to pass between cells

Electrical Signaling in the Phloem
• Rapid, long-distance electrical signaling through the phloem is
another dynamic feature of the symplast
– Studies have revealed that a stimulus in one part of a plant
can trigger an electrical signal in the phloem that affects
another part
• This can elicit changes in gene transcription,
respiration, photosynthesis, phloem unloading, or
hormonal levels
– This is similar to the function of nerves in that swift
electrical communication can occur between widely
separated organs

Phloem: An Information Superhighway
• The phloem is also a “superhighway” for systemic
transport of macromolecules and viruses
– Systemic changes are those that spread throughout
the body and affect many or all of the body’s systems
or organs
• In systemic transport, proteins and RNAs enter
sieve tubes through plasmodesmata, helping to
integrate the functions of the whole plant
– Ex) A defensive response to localized infection
in a plant organ sends signals through the
phloem that activate defense genes in
noninfected tissues

You should now be able to:
1. Describe how proton pumps function in
transport of materials across membranes
2. Define the following terms: osmosis, water
potential, flaccid, turgor pressure, turgid
3. Explain how aquaporins affect the rate of
water transport across membranes
4. Describe three routes available for short-
distance transport in plants

5. Relate structure to function in sieve-tube cells,
vessel cells, and tracheid cells
6. Explain how the endodermis functions as a
selective barrier between the root cortex and
vascular cylinder
7. Define and explain guttation
8. Explain this statement: “The ascent of xylem
sap is ultimately solar powered”

9. Describe the role of stomata and discuss
factors that might affect their density and
behavior
10. Trace the path of phloem sap from sugar
source to sugar sink; describe sugar loading
and unloading