research article apoptosis effect of girinimbine isolated ... · research article apoptosis effect...
TRANSCRIPT

Hindawi Publishing CorporationEvidence-Based Complementary and Alternative MedicineVolume 2013 Article ID 689865 12 pageshttpdxdoiorg1011552013689865
Research ArticleApoptosis Effect of Girinimbine Isolated from Murraya koenigiion Lung Cancer Cells In Vitro
Syam Mohan1 Siddig Ibrahim Abdelwahab2 Shiau-Chuen Cheah34
Mohd Aspollah Sukari5 Suvitha Syam16
Noorasyikin Shamsuddin1 and Mohd Rais Mustafa3
1 Department of Pharmacy Faculty of Medicine University of Malaya 50603 Kuala Lumpur Malaysia2Medical Research Centre Jazan University PO Box 114 45 142 Jazan Saudi Arabia3 Centre for Natural Products and Drug Discovery (CENAR) Department of Pharmacology Faculty of MedicineUniversity of Malaya 50603 Kuala Lumpur Malaysia
4 Faculty of Medicine amp Health Sciences UCSI University No 1 Jalan Menara Gading UCSI Heights Cheras56000 Kuala Lumpur Malaysia
5 Department of Chemistry Faculty of Science University Putra Malaysia 43400 Serdang Malaysia6UPM-MAKNA Cancer Research Laboratory Institute of Bioscience University Putra Malaysia 43400 Serdang Malaysia
Correspondence should be addressed to SyamMohan syammohanmumedumy
Received 7 November 2012 Revised 20 January 2013 Accepted 6 February 2013
Academic Editor Weena Jiratchariyakul
Copyright copy 2013 SyamMohan et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Murraya koenigii Spreng has been traditionally claimed as a remedy for cancerThe current study investigated the anticancer effectsof girinimbine a carbazole alkaloid isolated from Murraya koenigii Spreng on A549 lung cancer cells in relation to apoptoticmechanistic pathway Girinimbine was isolated from Murraya koenigii Spreng The antiproliferative activity was assayed usingMTT and the apoptosis detection was done by annexin V and lysosomal stability assays Multiparameter cytotoxicity assays wereperformed to investigate the change in mitochondrial membrane potential and cytochrome c translocation ROS caspase andhuman apoptosis proteome profiler assays were done to investigate the apoptotic mechanism of cell deathTheMTT assay revealedthat the girinimbine induces cell death with an IC
50of 1901 120583M A significant induction of early phase of apoptosis was shown
by annexin V and lysosomal stability assays After 24 h treatment with 1901 120583M of girinimbine decrease in the nuclear area andincrease inmitochondrialmembrane potential and plasmamembrane permeability were readily visibleMoreover the translocationof cytochrome c alsowas observed Girinimbinemediates its antiproliferative and apoptotic effects through up- and downregulationof apoptotic and antiapoptotic proteins There was a significant involvement of both intrinsic and extrinsic pathways Moreoverthe upregulation of p53 as well as the cell proliferation repressor proteins p27 and p21 and the significant role of insulinIGF-1signaling were also identified Moreover the caspases 3 and 8 were found to be significantly activated Our results taken togetherindicated that girinimbine may be a potential agent for anticancer drug development
1 Introduction
Lung cancer is one of the leading causes of cancer relateddeaths worldwide which has a high incidence of recurrenceIt has been estimated that approximately 14 million arediagnosed every year and more than 1 million people dieannually 12 of which are new cases [1] Nonsmall cell lungcancer (NSCLC) constitutes majority of lung cancers whichcomprises more than 80 of total diagnoses [2] Although
chemotherapy and radiation therapy are available to treatNSCLC they are largely ineffective and highly toxic witha low survival profile [3] This toxicity and resistance tothe current chemotherapy made researchers focus on newdrug candidates targeting apoptosis a programmed celldeath as physiological process that provides an effectivenoninflammatory way to remove redundant or damagedcells from tissues thereby securing tissue homeostasis [4]A multitude of signals activated by variable triggers such
2 Evidence-Based Complementary and Alternative Medicine
NH
O
0
20
40
60
80
100
025 05 075 1 125 15 175 2
Cel
ls vi
abili
ty (
)
Sl no120583M
(1) Girinimbine(2) Paclitaxel 68 plusmn 103
1901 plusmn 101
(a)
(b)(c)
log 120583g mLminus1
IC50 plusmn SD (24h)
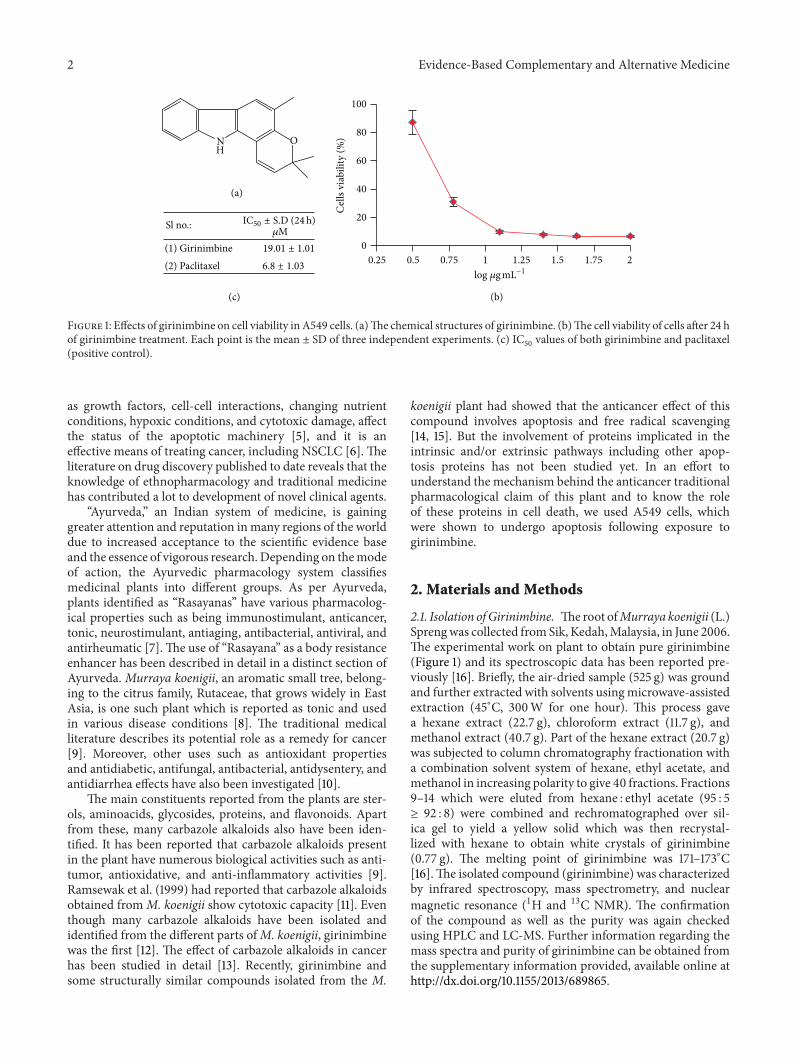
Figure 1 Effects of girinimbine on cell viability in A549 cells (a)The chemical structures of girinimbine (b)The cell viability of cells after 24 hof girinimbine treatment Each point is the mean plusmn SD of three independent experiments (c) IC
50values of both girinimbine and paclitaxel
(positive control)
as growth factors cell-cell interactions changing nutrientconditions hypoxic conditions and cytotoxic damage affectthe status of the apoptotic machinery [5] and it is aneffective means of treating cancer including NSCLC [6] Theliterature on drug discovery published to date reveals that theknowledge of ethnopharmacology and traditional medicinehas contributed a lot to development of novel clinical agents
ldquoAyurvedardquo an Indian system of medicine is gaininggreater attention and reputation inmany regions of the worlddue to increased acceptance to the scientific evidence baseand the essence of vigorous research Depending on themodeof action the Ayurvedic pharmacology system classifiesmedicinal plants into different groups As per Ayurvedaplants identified as ldquoRasayanasrdquo have various pharmacolog-ical properties such as being immunostimulant anticancertonic neurostimulant antiaging antibacterial antiviral andantirheumatic [7] The use of ldquoRasayanardquo as a body resistanceenhancer has been described in detail in a distinct section ofAyurveda Murraya koenigii an aromatic small tree belong-ing to the citrus family Rutaceae that grows widely in EastAsia is one such plant which is reported as tonic and usedin various disease conditions [8] The traditional medicalliterature describes its potential role as a remedy for cancer[9] Moreover other uses such as antioxidant propertiesand antidiabetic antifungal antibacterial antidysentery andantidiarrhea effects have also been investigated [10]
The main constituents reported from the plants are ster-ols aminoacids glycosides proteins and flavonoids Apartfrom these many carbazole alkaloids also have been iden-tified It has been reported that carbazole alkaloids presentin the plant have numerous biological activities such as anti-tumor antioxidative and anti-inflammatory activities [9]Ramsewak et al (1999) had reported that carbazole alkaloidsobtained fromM koenigii show cytotoxic capacity [11] Eventhough many carbazole alkaloids have been isolated andidentified from the different parts ofM koenigii girinimbinewas the first [12] The effect of carbazole alkaloids in cancerhas been studied in detail [13] Recently girinimbine andsome structurally similar compounds isolated from the M
koenigii plant had showed that the anticancer effect of thiscompound involves apoptosis and free radical scavenging[14 15] But the involvement of proteins implicated in theintrinsic andor extrinsic pathways including other apop-tosis proteins has not been studied yet In an effort tounderstand the mechanism behind the anticancer traditionalpharmacological claim of this plant and to know the roleof these proteins in cell death we used A549 cells whichwere shown to undergo apoptosis following exposure togirinimbine
2 Materials and Methods
21 Isolation ofGirinimbine Theroot ofMurraya koenigii (L)Sprengwas collected fromSik KedahMalaysia in June 2006The experimental work on plant to obtain pure girinimbine(Figure 1) and its spectroscopic data has been reported pre-viously [16] Briefly the air-dried sample (525 g) was groundand further extracted with solvents usingmicrowave-assistedextraction (45∘C 300W for one hour) This process gavea hexane extract (227 g) chloroform extract (117 g) andmethanol extract (407 g) Part of the hexane extract (207 g)was subjected to column chromatography fractionation witha combination solvent system of hexane ethyl acetate andmethanol in increasing polarity to give 40 fractions Fractions9ndash14 which were eluted from hexane ethyl acetate (95 5ge 92 8) were combined and rechromatographed over sil-ica gel to yield a yellow solid which was then recrystal-lized with hexane to obtain white crystals of girinimbine(077 g) The melting point of girinimbine was 171ndash173∘C[16]The isolated compound (girinimbine) was characterizedby infrared spectroscopy mass spectrometry and nuclearmagnetic resonance (1H and 13C NMR) The confirmationof the compound as well as the purity was again checkedusing HPLC and LC-MS Further information regarding themass spectra and purity of girinimbine can be obtained fromthe supplementary information provided available online athttpdxdoiorg1011552013689865
Evidence-Based Complementary and Alternative Medicine 3
22 Cell Culture A549 a human nonsmall cell lung cancercell line was purchased from ATCC (Rockville MD USA)and cultured in RPMI-1640 supplemented with 100 120583LmLfetal bovine serum 100UmL penicillin and 100mgmLstreptomycin sulfate Cells were maintained at 37∘C in ahumidified atmosphere of 5 CO
2 Cells were treated with
the girinimbine dissolved in DMSO while the untreatedcontrol cultures received only the vehicle (DMSO lt 1)
23 Cell Viability Assay Cells were seeded in 96-well platesat a density of 1 times 104 per well and treated with various con-centrations of girinimbine for 24 h Cells were then incubatedwith a medium containing 5mgmL 3-(45-dimethylthiazol-2-yl)-25-diphenyltetrazolium bromide (MTT) for another4 h The viable cell number was directly proportional toformazanproductionwhich thendissolved inDMSOandwasmeasured by spectrophotometry at 563 nm in a microplatereader (Tecan Infinite M 200 PRO Mannedorf Switzerland)[17]
24 Microscope Examination on Cellular Morphology Apop-tosis was monitored by annexin V labeling and fluorescencemicroscopy [18] Treated and untreated cells were washedwith PBS and then exposed with annexin V-fluorescein (BDPharmingen USA) for 15mins After a 488 nm excitationgreen fluorescence was visualized and recorded at 515 nmPhase contrast microscopic images from the same prepara-tions were also obtained for symptoms of apoptosis such ascell shrinkage ruffling and blebbing of cell membrane as wellas fragmentation of cells into small apoptotic bodies usinga fluorescent microscope (Nikon TE 2000U fluorescenceinverted microscope Tallahassee Florida)
25 Determination of Lysosomal Membrane Stability Cellswere assessed for lysosomal stability using acridine orange(AO) uptake assay Briefly cells were incubated with 7120583MAO (Sigma USA) and 12 120583M Hoechst 33342 (InvitrogenCarlsbad CA USA) diluted in culture media for 15minsat 37∘C and immediately observed and analyzed using theArrayScan HCS system (Cellomics PA USA) Acridineorange is ametachromatic fluorochrome and aweak base thatexhibits red fluorescence when highly concentrated in acidiclysosomes
26Measurement of Reactive Oxygen Species Generation Theproduction of intracellular reactive oxygen species (ROS)wasmeasured using 2101584071015840-dichlorofluorescein diacetate (DCFH-DA) [19] DCFH-DA passively enters the cell where itreacts with ROS to form the highly fluorescent compounddichlorofluorescein (DCF) Briefly 10mM DCFH-DA stocksolution (in methanol) was diluted 500-fold in HBSS withoutserum or other additives to yield a 20120583M working solutionAfter 24 h of exposure to girinimbine the cells in the 96-wellblack plate were washed twice with HBSS and then incubatedin 100 120583L working solution of DCFH-DA at 37∘C for 30minsFluorescence was then determined at 485 nm excitation and520 nm emission using a fluorescence microplate reader(Tecan Infinite M 200 PRO Mannedorf Switzerland)
27 Multiple Cytotoxicity Assay Cellomics MultiparameterCytotoxicity 3 Kit was used as described in detail previously[20]This kit enables simultaneousmeasurements in the samecell of six independent parameters that monitor cell healthincluding cell loss nuclear size and morphological changesmitochondrial membrane potential changes cytochrome crelease and changes in cell permeability Plates were analyzedusing the ArrayScan HCS system (Cellomics PA USA)
28 Image Acquisition and Cytometric Analysis Plates withstained cells were analyzed using the ArrayScan HCS sys-tem (Cellomics PA USA) This system is a computerizedautomated fluorescence imaging microscope that automat-ically identifies stained cells and reports the intensity anddistribution of fluorescence in individual cells The Array-Scan HCS system scans multiple fields in individual wellsto acquire and analyze images of single cells according todefined algorithms In each well 1000 cells were analyzedAutomatic focusing was performed in the nuclear channelto ensure focusing regardless of staining intensities in theother channels Images were acquired for each fluorescencechannel using suitable filters Images and data regardingintensity and texture of the fluorescence within each cell aswell as the average fluorescence of the cell population withinthe well were stored in a Microsoft SQL database for easyretrieval Data were captured extracted and analyzed withArrayScan II Data Acquisition and Data Viewer version 30(Cellomics)
29 HumanApoptosis Proteome Profiler Array To investigatethe pathways by which girinimbine induces apoptosis weperformed a determination of apoptosis-related proteinsusing the Proteome Profiler Array (RayBio Human Apop-tosis Antibody Array Kit RayBiotech USA) according tomanufacturerrsquos instructions In short the cells where treatedwith 19 120583M girinimbine 300 120583g proteins from each sam-ple and were incubated with the human apoptosis arrayovernight The apoptosis array data were quantified by scan-ning the membrane on a Biospectrum AC ChemiHR 40(UVP Upland CA) and analysis of the array image file wasperformed using image analysis software according to themanufacturerrsquos instruction
210 Measurement of Caspases 8 9 and 37 Activities Cas-pases 37 8 and 9 activity wasmeasured using luminescence-based assay Caspase-Glo 8 Assay Caspase-Glo 9 Assay andCaspase-Glo 37 Assay (Promega) 1 times 104 cells were culturedin 96-well culture plates in 50 120583L of RPMI 1640 supplementedwith 10 FBS and incubated for 24 h Cells then were treatedwith different concentrations of girinimbine and incubatedfor 24 h At the end of incubation 100 120583L of assay reagent wasadded to be incubated for 1 h at room temperature Lumines-cence wasmeasured using amicroplate reader (Tecan InfiniteM 200 PRO Mannedorf Switzerland)
211 Statistical Analysis From several independent measure-ments means and standard deviations were calculated Test-ing for significant differences between means were carried
4 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d) (e) (f)
Figure 2 Effects of girinimbine on the morphological changes of A549 cells after 24 h treatment Apoptosis was monitored by phase contrast((a) control (b) treatment 9120583M and (c) treatment 19120583M) and fluorescence microscopy ((d) control (e) treatment 9120583M and (f) treatment19120583M) Arrows indicate the cell shrinkage ruffling and blebbing of cell membrane while arrowheads indicate the annexin V binding tophosphatidylserine Magnification 20X
out using the one-way ANOVA and Dunnettrsquos post-test atprobabilities of error of 5 and 1
3 Results
31 Antipoliferative Activity The sigmoidal dose responsecurves of girinimbine in the end-point assays are shown inFigure 1 Cell viability was analyzed using the MTT assaywhich measures the metabolic activity of cell In the A549cells treated with girinimbine metabolic activity decreasedfollowed by 24 h treatment meanwhile in the control platecell viability and metabolism were not affected Figure 1(c)summarizes the IC
50values from MTT Meanwhile even at
380 120583M girinimbine could not exhibit any sign of toxicity inWRL-68 a normal (data not shown) which was employed inthis study to investigate the specificity of cytotoxicity
32 Apoptotic Mode of Cell Death Data obtained fromfluorescence detection of A549 cells together with the phasecontrast microscopic images are shown (Figure 2) Apoptosiswas clearly detected by the phosphatidylserine externaliza-tion on the treated cells Phase contrast microscopic picturesshowed a clear morphological change of treated cells whichwere observed after 24 h of treatment with girinimbine A549cells were seen to have shrunk in size and there was rufflingand blebbing of cell membranes thus suggesting that the cellswere undergoing apoptosis (Figure 2(c))
33 Girinimbine Initiates Lysosomal Membrane Permeabiliza-tion Recent studies have shown that lysosomal membrane
permeabilization (LMP) is an early and perhaps initiatingevent in apoptosis triggered by ligation of death recep-tors lysosomotropic agents oxidative stresses or serumwithdrawal [21 22] To evaluate the lysosome acidificationpattern the cells were analyzed by acridine orange staininga known pH indicator While untreated A549 cells dis-played strong granular acridine orange staining cells treatedwith girinimbine presented weak lysosome staining patternand cytoplasm acidification indicating lysosomal membranepermeabilization (Figures 3(a)ndash3(c)) Girinimbine induceddose-dependent decrease of fluorescence intensity of lyso-somal staining which was well compared with chloroquinea known inhibitor of lysosome function was included as apositive control (Figure 3(d))
34 ROS Assay Numerous investigations have documentedthat oxidative stress-mediated cellular changes are fre-quently induced in cells exposed to cytotoxic drugs UV orgamma irradiation [23 24] We examined whether girin-imbine affects the cellular levels of peroxide by measur-ing the changes in the fluorescence using DCF-DA Asshown in Figure 4(a) treatment with girinimbine markedlyincreased the DCF-DA-derived fluorescence (520 nm) Thisgirinimbine-mediated increase in fluorescence was markedlyinhibited by pretreatment with antioxidant ascorbic acidThen we next asked whether ROS generation induced bygirinimbine is directly associated with the induction ofapoptosis Chromatin condensation was measured in thecells which were pretreated with ascorbic acid using 12120583MHoechst 33342 The results shown in Figure 4(b) clearly
Evidence-Based Complementary and Alternative Medicine 5
(a) (b) (c)
200
100
0
Treatment
Fluo
resc
ent i
nten
sity
()
Chlo
roqu
ine25120583
M
Con
trol24
h
Giri
nim
bine
10120583
gm
L
Giri
nim
bine
5120583
gm
L
Giri
nim
bine
25120583
gm
L
(d)
Figure 3 Effects of girinimbine onA549 cells lysosome acidic environment Cells were incubatedwith different concentrations of girinimbinefor 24 h stained with acridine orange and analyzed by ArrayScan HCS system Control group corresponds to untreated cells Acridine orangeis a metachromatic fluorochrome and a weak base that exhibits red fluorescence when highly concentrated in acidic lysosomes (a) Normallysosome acidic environment (arrow)Notice theweak lysosome staining pattern and cytoplasm acidification (arrowhead) in cells treatedwithchloroquine (b) and girinimbine (c) indicating lysosomal membrane permeabilization Magnification 20X The mean fluorescent intensityproduced by the acridine orange was quantitatively measured (Figure 3(d))
exhibited that pretreatment with ascorbic acid could notprevent the apoptosis at various time periods Moreover thedata clearly shows the rapid decrease in the nuclear area andthe increase in fragmentation upon treatment with 19 120583Mgirinimbine (Figure 4(c))
35 Multiparameter Cytotoxicity Analysis This assay enablessimultaneousmeasurement of several cell-health parametersnuclear morphology DNA content cell membrane perme-ability and cytochrome c localization and release frommito-chondria Typical cytotoxic changes are illustrated in Figures5 and 6 Girinimbine induced decreases in cell numbernuclear area intensity and plasma membrane permeabilitywere significantly higher in the treated cells (119875 lt 001) andmitochondrial membrane potential reduction was observedsignificantly at 19120583M (119875 lt 001) The release of cytochromec was observed significantly in 9120583M and 19120583M with 119875 lt005 and 119875 lt 001 respectively These effects occurredmore rapidly and followed a dose-response pattern Cytotoxiceffects were considered to occur only when the rate of changeof fluorescence was distinctly greater than for the negativecontrols
36 Effect of Girinimbine on Apoptotic Markers After girin-imbine (19120583M) exposure A549 cells were lysed and apoptoticmarkers where screened using a protein array All majormarkers which are involved in both intrinsic and extrinsicpathways were induced on treatment As shown in Figure 7girinimbine treatment significantly increased the expressionof caspase 8 suggesting the activation of the death receptorpathway In addition the major protein involved in theextrinsic pathway such as Fas and FasL also regulates thetreatment Moreover the involvement of mitochondria in thecell death was evident by the regulation of the Bcl-2 familyof proteins such as Bad Bax Bcl-2 and Bim Besides theBcl-2 family member Bid was found to be cleaved as wellsuggesting a potential cross-talk between the death receptorand the mitochondrial pathway The treatment also resultedin a reduction in the level of expression of the inhibitorof apoptosis XIAP as well as survivin P53 as well as thecell proliferation repressor proteins p27 and p21 and theheat shock proteins such as antiapoptotic HSP60 HSP70and HSP27 which are a result of oxidative stress in the cellwere also induced InsulinIGF-1 signaling related proteinexpression has been observed on treatment Downregulation
6 Evidence-Based Complementary and Alternative Medicine
800850900950
100010501100115012001250130013501400
Control asaTreatment
24h 24h 8h 8h 4h 4h
Fluo
resc
ence
inte
nsity
(520
nm)
Girinimbine(19120583M)
Ascorbic acid
minus
minus minus minus minus
+ +
+
+ +
+
+ +
+ +
+
lowastlowast
lowast
lowastlowast
(a)
750
500
250
024h 12h 6h Control
Treatment
Fluo
resc
ent i
nten
sity
units
lowastlowast
lowastlowast
Girinimbine(19120583M)
Ascorbic acid
minus
minus
+
+
+
+
+
+
(b)
lowast
lowastlowastlowast
lowastlowast
Aver
age n
ucle
ar o
bjec
t siz
e
15
125
10
Girinimbine(19120583M)
Ascorbic acid
minus
minus
+
+
+
+
+
+
24h 12h 6h ControlTreatment
(c)
Figure 4 Effects of girinimbine on A549 cells ROS generation (a) DCF-fluorescence intensity after ascorbic acid and 19 120583M of girinimbineexposure at 4 8 and 24 h Rate of apoptosis in terms of fluorescent intensity (b) and average nuclear object size (c) of nucleus of the girinimbinetreatment was pretreatedwith 100mMascorbic acid Values aremeanplusmn SD from three independent experiments Triplicates of each treatmentgroup were used in each independent experiment The statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005 Fluorescent intensityunits directly represent the amount of ROS production
of antiapoptosis (IGF-I IGF-II IGFBP1 IGFBP2) and upreg-ulation of proapoptosis proteins (IGFBP3 IGFBP4) wereobserved
37 Caspase Depended Apoptosis in Girinimbine Treated A549Cells The involvement of the caspase cascade in the girin-imbine mediated cell death was confirmed The treatmentwith 19 120583M was significant at 119875 lt 001 for caspase 37and 9 with 8- and 5-fold increase respectively Meanwhilethe caspase 8 showed 10-fold increase at the maximumtreatment concentration (19 120583M 119875 lt 001) (Figure 8)These results are in parallel with the protein array andindicate that the signaling cascade leading to apoptosis in
girinimbine-treated cells involves both intrinsic and extrinsicpathways
4 Discussion
Apoptosis is a normal physiological process that plays avital role in numerous normal functions [4] Furthermoreit is an active physiological process causing cellular self-destruction that comprises specific morphological and bio-chemical changes in the nucleus and cytoplasm [25] Theinvolvement of an energy-dependent cascade of molecularevents makes the mechanism of apoptosis very highly com-plex and sophisticated [26] Apoptosis is regulated by two
Evidence-Based Complementary and Alternative Medicine 7
Hoechst 33342 Cell permeability Cytochrome c
Con
trol
Giri
nim
bine
Mitochondrial
membrane potential
Figure 5 Representative images of A549 cells treated withmedium alone and 19 120583Mof girinimbine and stained with Hoechst for nuclear cellpermeability dye mitochondrial membrane potential dye and cytochrome c The images from each row are obtained from the same field ofthe same treatment sample A549 produced a marked reduction in mitochondrial membrane potential and marked increases in membranepermeability and cytochrome c Magnification 20X
major pathways the extrinsic and the intrinsic pathways[27] Nevertheless there is evidence suggesting that boththe pathways are linked and that molecules involved in thepathways can influence one another [28]
The current study found that girinimbine a carbazolealkaloid from the roots of M koenigii can inhibit cellproliferation selectively in a dose dependent manner inA549 cells The morphological observation was conductedto explore whether the cytotoxic effect was related withthe apoptotic process and it was found that the cell deathinduced by girinimbine exhibited a clear morphologicalsign of apoptosis as this is an important property of acandidate anticancer drug [29] Followed by 24 h treatmentcell shrinkage ruffling and blebbing of cell membranes wereobserved These morphological features were more evidentwhile we analysed one of the key features of apoptosis whichis the change in plasma membrane structure by surfaceexposure of phosphatidylserine (PS) while the membraneintegrity remains unchallenged [30] The incorporation ofannexinV in to the same treated cells showed distinctive formof binding between the externalized PS and Annexin V whilecells from test flasks showedmore evidence of apoptosis thanthose grown under control conditions (Figure 2)
Translocation of lysosomal hydrolases from the lysosomallumen to the rest of the cell followed by the impairmentin the lysosomal membrane function is known as lysoso-mal membrane permeabilization (LMP) [31] Often thesehydrolases facilitate apoptosis by inducing mitochondrialouter-membrane permeabilization and caspase activation
[32] In our results the main LMP consequence such as weaklysosome acidification pattern and cytoplasmic acidificationwas clearly evident (Figure 3) Being aweak base chloroquineacts as lysosomotropic drug that raises intralysosomal pHHence we have monitored the release of liposomal contentby using chloroquine as a positive control Moreover in theFigure 3 it is very clear that cells treated with girinimbineresented a weak lysosome acidification pattern followed byan increase in cytoplasmic acidification Bearing in mind thesignificance of acidic lysosome environment for proteins andorganelles degradation it can be pointed out that autophagymay not the main event of cell death
Destabilization of the lysosomal membrane may becaused by increased production of ROS via massive peroxi-dation of membrane lipids [33] Moreover many studies hadshowed the relation between ROS and apoptosis induced incells exposed to cytotoxic drugs [34] Mitochondrial ROS areused as active mediators in the regulation of cell death [35]In our study the generation of ROS led to the upregulation ofcytoprotective protein markers HSP 60 70 and 27 Figure 4clearly shows that girinimbine exposure leads to increase ofROS accumulation but inhibition of ROSby ascorbic acid didnot prevent girinimbine-induced apoptosis indicating thatROS generation is not critical for the induction of apoptosisby girinimbine in A549 cells Moreover the treatment withgirinimbine significantly decreases the cell number nucleararea and cell membrane permeability as shown by themultiparameter apoptosis analysis In addition the complexrole of mitochondria in A549 cell apoptosis was investigated
8 Evidence-Based Complementary and Alternative Medicine
600
500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Total nuclear intensity
Giri19120583
M
Giri95120583
M
Giri475120583
M
(a)
Cell permeability500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(b)
350
300
250
200
150
100
50
0
Mitochondrial membrane potential
Con
trol
Pacli
taxe
l68120583
M
lowastlowastlowastlowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(c)
600700800
500400300200100
0
Cytochrome c
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowastlowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(d)
Figure 6 Quantitative analysis of girinimbine mediated apoptosis parameter Changes in total nuclear intensity (a) cell permeability(b) mitochondrial membrane potential (c) and cytochrome c localization (d) were measured simultaneously in A549 cells Followingtreatment with girinimbine we saw statistically significant increase in total nuclear intensity increased cell permeability loss ofmitochondrialmembrane potential and cytochrome c release frommitochondria Each experiment was performed at least two times Results are expressedas the means plusmn SD Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software (version40 GraphPad Software Inc San Diego CA) Statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
Mitochondria have been described to play a central role in theapoptotic process due to the mitochondrial proteins ability toactivate cellular apoptotic programs directly [36] Detectionof changes in mitochondrial membrane potential (Ψm) wascarried out as it is assumed that its disruption is the onsetofmitochondrial membrane transition pores (MPTP) forma-tion The present study revealed that girinimbine may act onmitochondria causing loss of Ψm and subsequent apoptosisThis kind of Ψm reduction and generation of MPTP mustfacilitate the relocalization of apoptogenic proteins from onesubcellular compartment such as mitochondria to cytoplasmto gain access to their substrates or interacting partners [37]
The multi cytotoxicity assay shows that girinimbine ind-uced cell death could be through the classical mitochondrial
pathway with cytochrome c release and caspase-dependentapoptosis The translocation of cytochrome c a componentof themitochondrial electron transfer chain was significantlyincreased upon girinimbine treatment In the mitochondriamediated apoptosis cytochrome c in turn binds to Apaf-1 to form a complex which triggers its oligomerization toform the apoptosome The caspase-9 holoenzyme is thenforms due to the binding of procaspase-9 to the apop-tosome and sequentially cleaves and activates the down-stream caspases such as caspase-3 [38] Our findings onthe upregulation of caspase 37 caspase 9 reduction in Ψmand release of cytochrome c to the cytosol substantiate thistheory and strongly support the involvement ofmitochondriain the apoptosis induced by girinimbine in A549 cells
Evidence-Based Complementary and Alternative Medicine 9
bad
bax
bcl-2
bcl-w
BID BI
MCa
spas
e 3Ca
spas
e 8IG
F-II IG
F-I HTR
AH
SP70
HSP
60H
SP27
FasL Fa
sD
R6Cy
to c
IGFB
P-1
IGFB
P-2 IG
FBP-
3IG
FBP-
4IG
FBP-
5IG
FBP-
6IG
F-1s
RLi
vin p
21 p
27 p53
SM
AC S
urvi
vin
sTN
F-R1 XI
AP
minus3
minus2
minus1
0
1
2
3
Relat
ive d
ensit
y di
ffere
nce t
o co
ntro
l (times10
5)
(a)
(b) (c)
A B C D E F G H I J K L M N1 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 82 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 83 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II4 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II5 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R16 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R17 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg Pos8 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg PosTNF-120572
TNF-120572 TNF-120573TNF-120573
(d)
Figure 7 Cells were lysed and protein arrays were performed Cells was treated with 19120583M girinimbine for 24 h and the whole cell proteinwas extracted Equal amount of (300 120583g) of protein from each sample was used for the assay Quantitative analysis in the arrays showeddifferences in the apoptotic markers (a) Representative images of the apoptotic protein array are shown for the control (b) treatment (c) andthe exact protein name of each dot in the array (d)
Suppression of apoptosis may promote the cancer develop-ment by inhibiting the chemotherapy as well as other forms ofinducers of cell death [39] Regulation of apoptosis has beencensoriously dependent on several genes which have beenidentified earlier and includes XIAP-a member of the IAPfamily It has the capacity to inhibit the activation of caspases3 7 and 9 [40] In our study we found that girinimbinetreatment of A549 cells caused inhibition of XIAP but SMAC
an antagonist to XIAP could be found to be slightly regulatedin this treatment concentration This is considered as a keyevent in the execution of cell death which is regulatedmainlyby proteins of the Bcl-2 family [41 42]
Critical issues in apoptosis include the dominance ofanti-versus proapoptotic Bcl-2 members [43] Girinimbine isfound to induce apoptosis in the A549 cells involvingdownregulated Bcl-2 expression This downregulation was
10 Evidence-Based Complementary and Alternative Medicine
3
2
1
0
times105
19 9 475 0 19 9 475 0 19 9 475 0Girinimbine treatment (120583M)
Lum
ines
cenc
e
Caspase 37Caspase 8Caspase 9
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast
Figure 8 Effect of girinimbine on activation of caspases 37 8 and 9Cells were treated with girinimbine at the indicated concentrationsfor 24 h and activity was determined by using a luminescentenzyme assay kit Each point represents the mean plusmn SD of threedifferent experiments Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software(version 40 GraphPad Software Inc San Diego CA) Statisticalsignificance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
concomitant with the upregulation of Bad Bax and Bimwhich inhibits antiapoptotic proteins and render the cellsmore susceptible to apoptogenic stimuli (Figure 7) Therelease of cytochrome c found in our study could be due tothe up regulated Bax since Bax in association with adeninenucleotide translocator within the permeability transitionpore complex will increase the mitochondrial membranepermeability and thereby discharge a number of apopto-genic molecules into the cytosol [44] In addition to thisupregulation of the tumor suppressor protein p53 which hasa critical role in regulation of the Bcl-2 and p27 and p21draws attention to the involvement of cell cycle arrest in theapoptosis process Alternatively we found the involvement ofextrinsic pathway proteins such as Fas and FasL upregulationand related upregulation of caspase 8 (Figure 7) Moreoverthe luminescent based caspase 8 assay had showed 10 folddifferences than the control Apart from this we foundthat there is a downregulation of Bid happening too Sincemitochondrial damage in the Fas pathway of apoptosis ismediated by the caspase 8 cleavage of Bid [45] the findingshere shows the cross talk between intrinsic and extrinsicpathways
The protein array results exhibited a significant reductionof both IGF-I and IGF-II IGF-I has been shown to exertstrongmitogenic and antiapoptotic effects in a variety of nor-mal and cancerous cells including lung cancer cell lines [46]Moreover the over expression of IGFBP3 is closely associatedwith the downregulation of IGF-I and its mitogenic activity[47] It is known that IGFBP-2 and IGFBP-1 are involved inthe regulation of cell migration apoptosis and cell growth
[48] Both are reported to be markedly overexpressed inmany tumors and tumor cell lines And it is assumed thatelevation of the IGFBP-2 production is part of a mechanismto compensate for the mitogenic and antiapoptotic effectsof tumor-derived IGFs [49] Our data clearly showed thedownregulation of IGFBP1 and IGFBP2 concomitant withupregulation of IGBP3
Our results provided a new insight into the mechanismof chemotherapeutic properties of Murraya koenigii Inaddition these study findings demonstrate that girinimbinemediates its apoptotic effects through both intrinsic andextrinsic pathway which is depended on caspase mediationMoreover the upregulation of p53 as well as the cell prolifer-ation repressor proteins p27 and p21 and the significant roleof insulinIGF-1 signaling were also identified These resultsindicate that girinimbine could be a candidate for a novelanticancer agentThereforemore in-depth in vitro and in vivostudies are currently going on in our laboratory
Conflict of Interests
The authors declare that they do not have any conflict ofinterests
Acknowledgment
The authors greatly acknowledge the financial support pro-vided by University of Malaya (Grant no HIR E0000002-20001)
References
[1] J K Field and S W Duffy ldquoLung cancer screening the wayforwardrdquo British Journal of Cancer vol 99 no 4 pp 557ndash5622008
[2] K A Yoon J H Park J Han et al ldquoA genome-wide associationstudy reveals susceptibility variants for nonsmall cell lungcancer in the Korean populationrdquo Human Molecular Geneticsvol 19 no 24 Article ID ddq421 pp 4948ndash4954 2010
[3] G Cartei F Cartei A Cantone et al ldquoCisplatin-cyclophos-phamide-mitomycin combination chemotherapy with support-ive care versus supportive care alone for treatment of metastaticnon- small-cell lung cancerrdquo Journal of the National CancerInstitute vol 85 no 10 pp 794ndash800 1993
[4] M O Hengartner ldquoThe biochemistry of apoptosisrdquoNature vol407 no 6805 pp 770ndash776 2000
[5] DHanahan andRAWeinberg ldquoThehallmarks of cancerrdquoCellvol 100 no 1 pp 57ndash70 2000
[6] J R Molina P Yang S D Cassivi S E Schild and A AAdjei ldquoNon-small cell lung cancer epidemiology risk factorstreatment and survivorshiprdquo Mayo Clinic Proceedings vol 83no 5 pp 584ndash594 2008
[7] S Diwanay D Chitre and B Patwardhan ldquoImmunoprotec-tion by botanical drugs in cancer chemotherapyrdquo Journal ofEthnopharmacology vol 90 no 1 pp 49ndash55 2004
[8] U M Thatte and S A Dahanukar ldquoRasayana Concept cluesfrom immunomodulatory therapyrdquo in An Annotated Bibliogra-phy of Indian Medicine S N Upadhyay Ed pp 141ndash148 1997
[9] A S Shah A S Wakade and A R Juvekar ldquoImmunomod-ulatory activity of methanolic extract of Murraya koenigii (L)
Evidence-Based Complementary and Alternative Medicine 11
Spreng leavesrdquo Indian Journal of Experimental Biology vol 46no 7 pp 505ndash509 2008
[10] R B Goswamia P K S Singha N Goswamib P Thomasd PU Devic and A K Pathakd ldquoStudies on antigenotoxic effect ofMurraya koenigii leavesrdquo International Journal of PharmaRecentResearch vol 2 pp 65ndash68 2010
[11] R S Ramsewak M G Nair G M Strasburg D L DeWittand J L Nitiss ldquoBiologically active carbazole alkaloids fromMurraya koenigiirdquo Journal of Agricultural and Food Chemistryvol 47 no 2 pp 444ndash447 1999
[12] D P Chakraborty B K Barman and P K Bose ldquoStructureof girinimbine a pyranocarbazole derivative isolated fromMurraya koenigii Sprengrdquo Scientific Culture vol 30 pp 445ndash448 1964
[13] L J M Rao K Ramalakshmi B B Borse and B RaghavanldquoAntioxidant and radical-scavenging carbazole alkaloids fromthe oleoresin of curry leaf (Murraya koenigii Spreng)rdquo FoodChemistry vol 100 no 2 pp 742ndash747 2007
[14] S L Wang B Cai C B Cui S Y Yan and C F Wu ldquoInductionof apoptosis by girinimbine in K562 cellrdquo Chinese Traditionaland Herbal Drugs vol 38 no 11 pp 1677ndash1681 2007
[15] S Syam A B Abdul M A Sukari S Mohan S I Abdelwahaband T S Wah ldquoThe growth suppressing effects of girinimbineon Hepg2 involve induction of apoptosis and cell cycle arrestrdquoMolecules vol 16 pp 7155ndash7170 2011
[16] A Bakar N Haslizawati M A Sukari et al ldquoChemical con-stituents from stem barks and roots of Murraya koenigii(Rutaceae)rdquo The Malaysian Journal of Analytical Sciences vol11 pp 173ndash176 2007
[17] S I A Wahab A B Abdul S M Mohan A S Al-ZubairiM M Elhassan and M Y Ibrahim ldquoBiological activities ofPereskia bleo extractsrdquo International Journal of Pharmacologyvol 5 no 1 pp 71ndash75 2009
[18] E Bossy-Wetzel and D R Green ldquoDetection of apoptosis byannexin V labelingrdquo Methods in Enzymology vol 322 pp 15ndash18 2000
[19] H Wang and J A Joseph ldquoQuantifying cellular oxidative stressby dichlorofluorescein assay usingmicroplate readerrdquo Free Rad-ical Biology and Medicine vol 27 no 5-6 pp 612ndash616 1999
[20] S C Cheah D R Appleton S T Lee M L Lam A H AHadi and M R Mustafa ldquoPanduratin a inhibits the growth ofA549 cells through induction of apoptosis and inhibition of NF-KappaB translocationrdquoMolecules vol 16 no 3 pp 2583ndash25982011
[21] L Foghsgaard DWissing DMauch et al ldquoCathepsin B acts asa dominant execution protease in tumor cell apoptosis inducedby tumor necrosis factorrdquo Journal of Cell Biology vol 153 no 5pp 999ndash1009 2001
[22] K Kagedal U Johansson and K Ollinger ldquoThe lysosomalprotease cathepsin D mediates apoptosis induced by oxidativestressrdquoThe FASEB journal vol 15 no 9 pp 1592ndash1594 2001
[23] M A Birch-Machin and H Swalwell ldquoHow mitochondriarecord the effects of UV exposure and oxidative stress usinghuman skin as a model tissuerdquo Mutagenesis vol 25 no 2 pp101ndash107 2010
[24] S Meister B Frey V R Lang et al ldquoCalcium channel blockerverapamil enhances endoplasmic reticulum stress and cell deathinduced by proteasome inhibition in myeloma cellsrdquoNeoplasiavol 12 no 7 pp 550ndash561 2010
[25] S Mohan A B Abdul S I Abdelwahab et al ldquoTyphoniumflagelliforme induces apoptosis in CEMss cells via activation
of caspase-9 PARP cleavage and cytochrome c release itsactivation coupled with G0G1 phase cell cycle arrestrdquo Journalof Ethnopharmacology vol 131 no 3 pp 592ndash600 2010
[26] S Elmore ldquoApoptosis a review of programmed cell deathrdquoToxicologic Pathology vol 35 no 4 pp 495ndash516 2007
[27] M Aberg M Johnell M Wickstrom and A Siegbahn ldquoTissueFactor FVIIa prevents the extrinsic pathway of apoptosis byregulation of the tumor suppressor Death-Associated ProteinKinase 1 (DAPK1)rdquoThrombosis Research vol 127 no 2 pp 141ndash148 2011
[28] M Nigam N Singh V Ranjan et al ldquoCentchroman mediatedapoptosis involves cross-talk between extrinsicintrinsic path-ways and oxidative regulationrdquo Life Sciences vol 87 no 23ndash26pp 750ndash758 2010
[29] O S Frankfurt and A Krishan ldquoApoptosis-based drug screen-ing and detection of selective toxicity to cancer cellsrdquo Anti-Cancer Drugs vol 14 no 7 pp 555ndash561 2003
[30] M van Engeland L J W Nieland F C S Ramaekers BSchutte and C P M Reutelingsperger ldquoAnnexin V-affinityassay a review on an apoptosis detection system based onphosphatidylserine exposurerdquo Cytometry vol 31 no 1 pp 1ndash91998
[31] G A Joanitti R B Azevedo and S M Freitas ldquoApoptosisand lysosome membrane permeabilization induction on breastcancer cells by an anticarcinogenic Bowman-Birk proteaseinhibitor from Vigna unguiculata seedsrdquo Cancer Letters vol293 no 1 pp 73ndash81 2010
[32] T Kirkegaard and M Jaattela ldquoLysosomal involvement in celldeath and cancerrdquo Biochimica et Biophysica ActamdashMolecularCell Research vol 1793 no 4 pp 746ndash754 2009
[33] A C Johansson H Appelqvist C Nilsson K Kagedal KRoberg and K Ollinger ldquoRegulation of apoptosis-associatedlysosomal membrane permeabilizationrdquo Apoptosis vol 15 no5 pp 527ndash540 2010
[34] J Wen K R You S Y Lee C H Song and D G KimldquoOxidative stress-mediated apoptosis the anticancer effect ofthe sesquiterpene lactone parthenoliderdquo Journal of BiologicalChemistry vol 277 no 41 pp 38954ndash38964 2002
[35] C Fleury B Mignotte and J L Vayssiere ldquoMitochondrialreactive oxygen species in cell death signalingrdquo Biochimie vol84 no 2-3 pp 131ndash141 2002
[36] A M Verhagen P G Ekert M Pakusch et al ldquoIdentificationof DIABLO a mammalian protein that promotes apoptosis bybinding to and antagonizing IAP proteinsrdquo Cell vol 102 no 1pp 43ndash53 2000
[37] N Ajenjo E Canonon I Sanchez-Perez et al ldquoSubcellularlocalization determines the protective effects of activated ERK2against distinct apoptogenic stimuli in myeloid leukemia cellsrdquoJournal of Biological Chemistry vol 279 no 31 pp 32813ndash328232004
[38] X Wang ldquoThe expanding role of mitochondria in apoptosisrdquoGenes and Development vol 15 no 22 pp 2922ndash2933 2001
[39] Y A Hannun ldquoApoptosis and the dilemma of cancer chemo-therapyrdquo Blood vol 89 no 6 pp 1845ndash1853 1997
[40] F L Scott J B Denault S J Riedl H Shin M Renatusand G S Salvesen ldquoXIAP inhibits caspase-3 and -7 usingtwo binding sites evolutionary conserved mechanism of IAPsrdquoEMBO Journal vol 24 no 3 pp 645ndash655 2005
[41] S Cory and J M Adams ldquoThe BCL2 family regulators of thecellular life-or-death switchrdquo Nature Reviews Cancer vol 2 no9 pp 647ndash656 2002
12 Evidence-Based Complementary and Alternative Medicine
[42] S B Bratton and GM Cohen ldquoDeath receptors leave a caspasefootprint that Smacs of XIAPrdquo Cell Death and Differentiationvol 10 no 1 pp 4ndash6 2003
[43] N Zamzami andGKroemer ldquoThemitochondrion in apoptosishow Pandorarsquos box opensrdquo Nature Reviews Molecular CellBiology vol 2 no 1 pp 67ndash71 2001
[44] I Marzo C Brenner N Zamzami et al ldquoBax and adeninenucleotide translocator cooperate in the mitochondrial controlof apoptosisrdquo Science vol 281 no 5385 pp 2027ndash2031 1998
[45] M D Esposti ldquoThe roles of BidrdquoApoptosis vol 7 no 5 pp 433ndash440 2002
[46] N E Allen A W Roddam D S Allen et al ldquoA prospectivestudy of serum insulin-like growth factor-I (IGF-I) IGF-IIIGF-binding protein-3 and breast cancer riskrdquo British Journalof Cancer vol 92 no 7 pp 1283ndash1287 2005
[47] L Buckbinder R Talbott S Velasco-Miguel et al ldquoInductionof the growth inhibitor IGF-binding protein 3 by p53rdquo Naturevol 377 no 6550 pp 646ndash649 1995
[48] E Ruoslahti ldquoFibronectin and its integrin receptors in cancerrdquoAdvances in Cancer Research vol 76 pp 18ndash20 1999
[49] B S Schutt M Langkamp U Rauschnabel M B Ranke andM W Elmlinger ldquoIntegrin-mediated action of insulin-likefactor binding protein-2 in tumor cellsrdquo Journal of MolecularEndocrinology vol 32 no 3 pp 859ndash868 2004
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
2 Evidence-Based Complementary and Alternative Medicine
NH
O
0
20
40
60
80
100
025 05 075 1 125 15 175 2
Cel
ls vi
abili
ty (
)
Sl no120583M
(1) Girinimbine(2) Paclitaxel 68 plusmn 103
1901 plusmn 101
(a)
(b)(c)
log 120583g mLminus1
IC50 plusmn SD (24h)
Figure 1 Effects of girinimbine on cell viability in A549 cells (a)The chemical structures of girinimbine (b)The cell viability of cells after 24 hof girinimbine treatment Each point is the mean plusmn SD of three independent experiments (c) IC
50values of both girinimbine and paclitaxel
(positive control)
as growth factors cell-cell interactions changing nutrientconditions hypoxic conditions and cytotoxic damage affectthe status of the apoptotic machinery [5] and it is aneffective means of treating cancer including NSCLC [6] Theliterature on drug discovery published to date reveals that theknowledge of ethnopharmacology and traditional medicinehas contributed a lot to development of novel clinical agents
ldquoAyurvedardquo an Indian system of medicine is gaininggreater attention and reputation inmany regions of the worlddue to increased acceptance to the scientific evidence baseand the essence of vigorous research Depending on themodeof action the Ayurvedic pharmacology system classifiesmedicinal plants into different groups As per Ayurvedaplants identified as ldquoRasayanasrdquo have various pharmacolog-ical properties such as being immunostimulant anticancertonic neurostimulant antiaging antibacterial antiviral andantirheumatic [7] The use of ldquoRasayanardquo as a body resistanceenhancer has been described in detail in a distinct section ofAyurveda Murraya koenigii an aromatic small tree belong-ing to the citrus family Rutaceae that grows widely in EastAsia is one such plant which is reported as tonic and usedin various disease conditions [8] The traditional medicalliterature describes its potential role as a remedy for cancer[9] Moreover other uses such as antioxidant propertiesand antidiabetic antifungal antibacterial antidysentery andantidiarrhea effects have also been investigated [10]
The main constituents reported from the plants are ster-ols aminoacids glycosides proteins and flavonoids Apartfrom these many carbazole alkaloids also have been iden-tified It has been reported that carbazole alkaloids presentin the plant have numerous biological activities such as anti-tumor antioxidative and anti-inflammatory activities [9]Ramsewak et al (1999) had reported that carbazole alkaloidsobtained fromM koenigii show cytotoxic capacity [11] Eventhough many carbazole alkaloids have been isolated andidentified from the different parts ofM koenigii girinimbinewas the first [12] The effect of carbazole alkaloids in cancerhas been studied in detail [13] Recently girinimbine andsome structurally similar compounds isolated from the M
koenigii plant had showed that the anticancer effect of thiscompound involves apoptosis and free radical scavenging[14 15] But the involvement of proteins implicated in theintrinsic andor extrinsic pathways including other apop-tosis proteins has not been studied yet In an effort tounderstand the mechanism behind the anticancer traditionalpharmacological claim of this plant and to know the roleof these proteins in cell death we used A549 cells whichwere shown to undergo apoptosis following exposure togirinimbine
2 Materials and Methods
21 Isolation ofGirinimbine Theroot ofMurraya koenigii (L)Sprengwas collected fromSik KedahMalaysia in June 2006The experimental work on plant to obtain pure girinimbine(Figure 1) and its spectroscopic data has been reported pre-viously [16] Briefly the air-dried sample (525 g) was groundand further extracted with solvents usingmicrowave-assistedextraction (45∘C 300W for one hour) This process gavea hexane extract (227 g) chloroform extract (117 g) andmethanol extract (407 g) Part of the hexane extract (207 g)was subjected to column chromatography fractionation witha combination solvent system of hexane ethyl acetate andmethanol in increasing polarity to give 40 fractions Fractions9ndash14 which were eluted from hexane ethyl acetate (95 5ge 92 8) were combined and rechromatographed over sil-ica gel to yield a yellow solid which was then recrystal-lized with hexane to obtain white crystals of girinimbine(077 g) The melting point of girinimbine was 171ndash173∘C[16]The isolated compound (girinimbine) was characterizedby infrared spectroscopy mass spectrometry and nuclearmagnetic resonance (1H and 13C NMR) The confirmationof the compound as well as the purity was again checkedusing HPLC and LC-MS Further information regarding themass spectra and purity of girinimbine can be obtained fromthe supplementary information provided available online athttpdxdoiorg1011552013689865
Evidence-Based Complementary and Alternative Medicine 3
22 Cell Culture A549 a human nonsmall cell lung cancercell line was purchased from ATCC (Rockville MD USA)and cultured in RPMI-1640 supplemented with 100 120583LmLfetal bovine serum 100UmL penicillin and 100mgmLstreptomycin sulfate Cells were maintained at 37∘C in ahumidified atmosphere of 5 CO
2 Cells were treated with
the girinimbine dissolved in DMSO while the untreatedcontrol cultures received only the vehicle (DMSO lt 1)
23 Cell Viability Assay Cells were seeded in 96-well platesat a density of 1 times 104 per well and treated with various con-centrations of girinimbine for 24 h Cells were then incubatedwith a medium containing 5mgmL 3-(45-dimethylthiazol-2-yl)-25-diphenyltetrazolium bromide (MTT) for another4 h The viable cell number was directly proportional toformazanproductionwhich thendissolved inDMSOandwasmeasured by spectrophotometry at 563 nm in a microplatereader (Tecan Infinite M 200 PRO Mannedorf Switzerland)[17]
24 Microscope Examination on Cellular Morphology Apop-tosis was monitored by annexin V labeling and fluorescencemicroscopy [18] Treated and untreated cells were washedwith PBS and then exposed with annexin V-fluorescein (BDPharmingen USA) for 15mins After a 488 nm excitationgreen fluorescence was visualized and recorded at 515 nmPhase contrast microscopic images from the same prepara-tions were also obtained for symptoms of apoptosis such ascell shrinkage ruffling and blebbing of cell membrane as wellas fragmentation of cells into small apoptotic bodies usinga fluorescent microscope (Nikon TE 2000U fluorescenceinverted microscope Tallahassee Florida)
25 Determination of Lysosomal Membrane Stability Cellswere assessed for lysosomal stability using acridine orange(AO) uptake assay Briefly cells were incubated with 7120583MAO (Sigma USA) and 12 120583M Hoechst 33342 (InvitrogenCarlsbad CA USA) diluted in culture media for 15minsat 37∘C and immediately observed and analyzed using theArrayScan HCS system (Cellomics PA USA) Acridineorange is ametachromatic fluorochrome and aweak base thatexhibits red fluorescence when highly concentrated in acidiclysosomes
26Measurement of Reactive Oxygen Species Generation Theproduction of intracellular reactive oxygen species (ROS)wasmeasured using 2101584071015840-dichlorofluorescein diacetate (DCFH-DA) [19] DCFH-DA passively enters the cell where itreacts with ROS to form the highly fluorescent compounddichlorofluorescein (DCF) Briefly 10mM DCFH-DA stocksolution (in methanol) was diluted 500-fold in HBSS withoutserum or other additives to yield a 20120583M working solutionAfter 24 h of exposure to girinimbine the cells in the 96-wellblack plate were washed twice with HBSS and then incubatedin 100 120583L working solution of DCFH-DA at 37∘C for 30minsFluorescence was then determined at 485 nm excitation and520 nm emission using a fluorescence microplate reader(Tecan Infinite M 200 PRO Mannedorf Switzerland)
27 Multiple Cytotoxicity Assay Cellomics MultiparameterCytotoxicity 3 Kit was used as described in detail previously[20]This kit enables simultaneousmeasurements in the samecell of six independent parameters that monitor cell healthincluding cell loss nuclear size and morphological changesmitochondrial membrane potential changes cytochrome crelease and changes in cell permeability Plates were analyzedusing the ArrayScan HCS system (Cellomics PA USA)
28 Image Acquisition and Cytometric Analysis Plates withstained cells were analyzed using the ArrayScan HCS sys-tem (Cellomics PA USA) This system is a computerizedautomated fluorescence imaging microscope that automat-ically identifies stained cells and reports the intensity anddistribution of fluorescence in individual cells The Array-Scan HCS system scans multiple fields in individual wellsto acquire and analyze images of single cells according todefined algorithms In each well 1000 cells were analyzedAutomatic focusing was performed in the nuclear channelto ensure focusing regardless of staining intensities in theother channels Images were acquired for each fluorescencechannel using suitable filters Images and data regardingintensity and texture of the fluorescence within each cell aswell as the average fluorescence of the cell population withinthe well were stored in a Microsoft SQL database for easyretrieval Data were captured extracted and analyzed withArrayScan II Data Acquisition and Data Viewer version 30(Cellomics)
29 HumanApoptosis Proteome Profiler Array To investigatethe pathways by which girinimbine induces apoptosis weperformed a determination of apoptosis-related proteinsusing the Proteome Profiler Array (RayBio Human Apop-tosis Antibody Array Kit RayBiotech USA) according tomanufacturerrsquos instructions In short the cells where treatedwith 19 120583M girinimbine 300 120583g proteins from each sam-ple and were incubated with the human apoptosis arrayovernight The apoptosis array data were quantified by scan-ning the membrane on a Biospectrum AC ChemiHR 40(UVP Upland CA) and analysis of the array image file wasperformed using image analysis software according to themanufacturerrsquos instruction
210 Measurement of Caspases 8 9 and 37 Activities Cas-pases 37 8 and 9 activity wasmeasured using luminescence-based assay Caspase-Glo 8 Assay Caspase-Glo 9 Assay andCaspase-Glo 37 Assay (Promega) 1 times 104 cells were culturedin 96-well culture plates in 50 120583L of RPMI 1640 supplementedwith 10 FBS and incubated for 24 h Cells then were treatedwith different concentrations of girinimbine and incubatedfor 24 h At the end of incubation 100 120583L of assay reagent wasadded to be incubated for 1 h at room temperature Lumines-cence wasmeasured using amicroplate reader (Tecan InfiniteM 200 PRO Mannedorf Switzerland)
211 Statistical Analysis From several independent measure-ments means and standard deviations were calculated Test-ing for significant differences between means were carried
4 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d) (e) (f)
Figure 2 Effects of girinimbine on the morphological changes of A549 cells after 24 h treatment Apoptosis was monitored by phase contrast((a) control (b) treatment 9120583M and (c) treatment 19120583M) and fluorescence microscopy ((d) control (e) treatment 9120583M and (f) treatment19120583M) Arrows indicate the cell shrinkage ruffling and blebbing of cell membrane while arrowheads indicate the annexin V binding tophosphatidylserine Magnification 20X
out using the one-way ANOVA and Dunnettrsquos post-test atprobabilities of error of 5 and 1
3 Results
31 Antipoliferative Activity The sigmoidal dose responsecurves of girinimbine in the end-point assays are shown inFigure 1 Cell viability was analyzed using the MTT assaywhich measures the metabolic activity of cell In the A549cells treated with girinimbine metabolic activity decreasedfollowed by 24 h treatment meanwhile in the control platecell viability and metabolism were not affected Figure 1(c)summarizes the IC
50values from MTT Meanwhile even at
380 120583M girinimbine could not exhibit any sign of toxicity inWRL-68 a normal (data not shown) which was employed inthis study to investigate the specificity of cytotoxicity
32 Apoptotic Mode of Cell Death Data obtained fromfluorescence detection of A549 cells together with the phasecontrast microscopic images are shown (Figure 2) Apoptosiswas clearly detected by the phosphatidylserine externaliza-tion on the treated cells Phase contrast microscopic picturesshowed a clear morphological change of treated cells whichwere observed after 24 h of treatment with girinimbine A549cells were seen to have shrunk in size and there was rufflingand blebbing of cell membranes thus suggesting that the cellswere undergoing apoptosis (Figure 2(c))
33 Girinimbine Initiates Lysosomal Membrane Permeabiliza-tion Recent studies have shown that lysosomal membrane
permeabilization (LMP) is an early and perhaps initiatingevent in apoptosis triggered by ligation of death recep-tors lysosomotropic agents oxidative stresses or serumwithdrawal [21 22] To evaluate the lysosome acidificationpattern the cells were analyzed by acridine orange staininga known pH indicator While untreated A549 cells dis-played strong granular acridine orange staining cells treatedwith girinimbine presented weak lysosome staining patternand cytoplasm acidification indicating lysosomal membranepermeabilization (Figures 3(a)ndash3(c)) Girinimbine induceddose-dependent decrease of fluorescence intensity of lyso-somal staining which was well compared with chloroquinea known inhibitor of lysosome function was included as apositive control (Figure 3(d))
34 ROS Assay Numerous investigations have documentedthat oxidative stress-mediated cellular changes are fre-quently induced in cells exposed to cytotoxic drugs UV orgamma irradiation [23 24] We examined whether girin-imbine affects the cellular levels of peroxide by measur-ing the changes in the fluorescence using DCF-DA Asshown in Figure 4(a) treatment with girinimbine markedlyincreased the DCF-DA-derived fluorescence (520 nm) Thisgirinimbine-mediated increase in fluorescence was markedlyinhibited by pretreatment with antioxidant ascorbic acidThen we next asked whether ROS generation induced bygirinimbine is directly associated with the induction ofapoptosis Chromatin condensation was measured in thecells which were pretreated with ascorbic acid using 12120583MHoechst 33342 The results shown in Figure 4(b) clearly
Evidence-Based Complementary and Alternative Medicine 5
(a) (b) (c)
200
100
0
Treatment
Fluo
resc
ent i
nten
sity
()
Chlo
roqu
ine25120583
M
Con
trol24
h
Giri
nim
bine
10120583
gm
L
Giri
nim
bine
5120583
gm
L
Giri
nim
bine
25120583
gm
L
(d)
Figure 3 Effects of girinimbine onA549 cells lysosome acidic environment Cells were incubatedwith different concentrations of girinimbinefor 24 h stained with acridine orange and analyzed by ArrayScan HCS system Control group corresponds to untreated cells Acridine orangeis a metachromatic fluorochrome and a weak base that exhibits red fluorescence when highly concentrated in acidic lysosomes (a) Normallysosome acidic environment (arrow)Notice theweak lysosome staining pattern and cytoplasm acidification (arrowhead) in cells treatedwithchloroquine (b) and girinimbine (c) indicating lysosomal membrane permeabilization Magnification 20X The mean fluorescent intensityproduced by the acridine orange was quantitatively measured (Figure 3(d))
exhibited that pretreatment with ascorbic acid could notprevent the apoptosis at various time periods Moreover thedata clearly shows the rapid decrease in the nuclear area andthe increase in fragmentation upon treatment with 19 120583Mgirinimbine (Figure 4(c))
35 Multiparameter Cytotoxicity Analysis This assay enablessimultaneousmeasurement of several cell-health parametersnuclear morphology DNA content cell membrane perme-ability and cytochrome c localization and release frommito-chondria Typical cytotoxic changes are illustrated in Figures5 and 6 Girinimbine induced decreases in cell numbernuclear area intensity and plasma membrane permeabilitywere significantly higher in the treated cells (119875 lt 001) andmitochondrial membrane potential reduction was observedsignificantly at 19120583M (119875 lt 001) The release of cytochromec was observed significantly in 9120583M and 19120583M with 119875 lt005 and 119875 lt 001 respectively These effects occurredmore rapidly and followed a dose-response pattern Cytotoxiceffects were considered to occur only when the rate of changeof fluorescence was distinctly greater than for the negativecontrols
36 Effect of Girinimbine on Apoptotic Markers After girin-imbine (19120583M) exposure A549 cells were lysed and apoptoticmarkers where screened using a protein array All majormarkers which are involved in both intrinsic and extrinsicpathways were induced on treatment As shown in Figure 7girinimbine treatment significantly increased the expressionof caspase 8 suggesting the activation of the death receptorpathway In addition the major protein involved in theextrinsic pathway such as Fas and FasL also regulates thetreatment Moreover the involvement of mitochondria in thecell death was evident by the regulation of the Bcl-2 familyof proteins such as Bad Bax Bcl-2 and Bim Besides theBcl-2 family member Bid was found to be cleaved as wellsuggesting a potential cross-talk between the death receptorand the mitochondrial pathway The treatment also resultedin a reduction in the level of expression of the inhibitorof apoptosis XIAP as well as survivin P53 as well as thecell proliferation repressor proteins p27 and p21 and theheat shock proteins such as antiapoptotic HSP60 HSP70and HSP27 which are a result of oxidative stress in the cellwere also induced InsulinIGF-1 signaling related proteinexpression has been observed on treatment Downregulation
6 Evidence-Based Complementary and Alternative Medicine
800850900950
100010501100115012001250130013501400
Control asaTreatment
24h 24h 8h 8h 4h 4h
Fluo
resc
ence
inte
nsity
(520
nm)
Girinimbine(19120583M)
Ascorbic acid
minus
minus minus minus minus
+ +
+
+ +
+
+ +
+ +
+
lowastlowast
lowast
lowastlowast
(a)
750
500
250
024h 12h 6h Control
Treatment
Fluo
resc
ent i
nten
sity
units
lowastlowast
lowastlowast
Girinimbine(19120583M)
Ascorbic acid
minus
minus
+
+
+
+
+
+
(b)
lowast
lowastlowastlowast
lowastlowast
Aver
age n
ucle
ar o
bjec
t siz
e
15
125
10
Girinimbine(19120583M)
Ascorbic acid
minus
minus
+
+
+
+
+
+
24h 12h 6h ControlTreatment
(c)
Figure 4 Effects of girinimbine on A549 cells ROS generation (a) DCF-fluorescence intensity after ascorbic acid and 19 120583M of girinimbineexposure at 4 8 and 24 h Rate of apoptosis in terms of fluorescent intensity (b) and average nuclear object size (c) of nucleus of the girinimbinetreatment was pretreatedwith 100mMascorbic acid Values aremeanplusmn SD from three independent experiments Triplicates of each treatmentgroup were used in each independent experiment The statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005 Fluorescent intensityunits directly represent the amount of ROS production
of antiapoptosis (IGF-I IGF-II IGFBP1 IGFBP2) and upreg-ulation of proapoptosis proteins (IGFBP3 IGFBP4) wereobserved
37 Caspase Depended Apoptosis in Girinimbine Treated A549Cells The involvement of the caspase cascade in the girin-imbine mediated cell death was confirmed The treatmentwith 19 120583M was significant at 119875 lt 001 for caspase 37and 9 with 8- and 5-fold increase respectively Meanwhilethe caspase 8 showed 10-fold increase at the maximumtreatment concentration (19 120583M 119875 lt 001) (Figure 8)These results are in parallel with the protein array andindicate that the signaling cascade leading to apoptosis in
girinimbine-treated cells involves both intrinsic and extrinsicpathways
4 Discussion
Apoptosis is a normal physiological process that plays avital role in numerous normal functions [4] Furthermoreit is an active physiological process causing cellular self-destruction that comprises specific morphological and bio-chemical changes in the nucleus and cytoplasm [25] Theinvolvement of an energy-dependent cascade of molecularevents makes the mechanism of apoptosis very highly com-plex and sophisticated [26] Apoptosis is regulated by two
Evidence-Based Complementary and Alternative Medicine 7
Hoechst 33342 Cell permeability Cytochrome c
Con
trol
Giri
nim
bine
Mitochondrial
membrane potential
Figure 5 Representative images of A549 cells treated withmedium alone and 19 120583Mof girinimbine and stained with Hoechst for nuclear cellpermeability dye mitochondrial membrane potential dye and cytochrome c The images from each row are obtained from the same field ofthe same treatment sample A549 produced a marked reduction in mitochondrial membrane potential and marked increases in membranepermeability and cytochrome c Magnification 20X
major pathways the extrinsic and the intrinsic pathways[27] Nevertheless there is evidence suggesting that boththe pathways are linked and that molecules involved in thepathways can influence one another [28]
The current study found that girinimbine a carbazolealkaloid from the roots of M koenigii can inhibit cellproliferation selectively in a dose dependent manner inA549 cells The morphological observation was conductedto explore whether the cytotoxic effect was related withthe apoptotic process and it was found that the cell deathinduced by girinimbine exhibited a clear morphologicalsign of apoptosis as this is an important property of acandidate anticancer drug [29] Followed by 24 h treatmentcell shrinkage ruffling and blebbing of cell membranes wereobserved These morphological features were more evidentwhile we analysed one of the key features of apoptosis whichis the change in plasma membrane structure by surfaceexposure of phosphatidylserine (PS) while the membraneintegrity remains unchallenged [30] The incorporation ofannexinV in to the same treated cells showed distinctive formof binding between the externalized PS and Annexin V whilecells from test flasks showedmore evidence of apoptosis thanthose grown under control conditions (Figure 2)
Translocation of lysosomal hydrolases from the lysosomallumen to the rest of the cell followed by the impairmentin the lysosomal membrane function is known as lysoso-mal membrane permeabilization (LMP) [31] Often thesehydrolases facilitate apoptosis by inducing mitochondrialouter-membrane permeabilization and caspase activation
[32] In our results the main LMP consequence such as weaklysosome acidification pattern and cytoplasmic acidificationwas clearly evident (Figure 3) Being aweak base chloroquineacts as lysosomotropic drug that raises intralysosomal pHHence we have monitored the release of liposomal contentby using chloroquine as a positive control Moreover in theFigure 3 it is very clear that cells treated with girinimbineresented a weak lysosome acidification pattern followed byan increase in cytoplasmic acidification Bearing in mind thesignificance of acidic lysosome environment for proteins andorganelles degradation it can be pointed out that autophagymay not the main event of cell death
Destabilization of the lysosomal membrane may becaused by increased production of ROS via massive peroxi-dation of membrane lipids [33] Moreover many studies hadshowed the relation between ROS and apoptosis induced incells exposed to cytotoxic drugs [34] Mitochondrial ROS areused as active mediators in the regulation of cell death [35]In our study the generation of ROS led to the upregulation ofcytoprotective protein markers HSP 60 70 and 27 Figure 4clearly shows that girinimbine exposure leads to increase ofROS accumulation but inhibition of ROSby ascorbic acid didnot prevent girinimbine-induced apoptosis indicating thatROS generation is not critical for the induction of apoptosisby girinimbine in A549 cells Moreover the treatment withgirinimbine significantly decreases the cell number nucleararea and cell membrane permeability as shown by themultiparameter apoptosis analysis In addition the complexrole of mitochondria in A549 cell apoptosis was investigated
8 Evidence-Based Complementary and Alternative Medicine
600
500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Total nuclear intensity
Giri19120583
M
Giri95120583
M
Giri475120583
M
(a)
Cell permeability500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(b)
350
300
250
200
150
100
50
0
Mitochondrial membrane potential
Con
trol
Pacli
taxe
l68120583
M
lowastlowastlowastlowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(c)
600700800
500400300200100
0
Cytochrome c
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowastlowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(d)
Figure 6 Quantitative analysis of girinimbine mediated apoptosis parameter Changes in total nuclear intensity (a) cell permeability(b) mitochondrial membrane potential (c) and cytochrome c localization (d) were measured simultaneously in A549 cells Followingtreatment with girinimbine we saw statistically significant increase in total nuclear intensity increased cell permeability loss ofmitochondrialmembrane potential and cytochrome c release frommitochondria Each experiment was performed at least two times Results are expressedas the means plusmn SD Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software (version40 GraphPad Software Inc San Diego CA) Statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
Mitochondria have been described to play a central role in theapoptotic process due to the mitochondrial proteins ability toactivate cellular apoptotic programs directly [36] Detectionof changes in mitochondrial membrane potential (Ψm) wascarried out as it is assumed that its disruption is the onsetofmitochondrial membrane transition pores (MPTP) forma-tion The present study revealed that girinimbine may act onmitochondria causing loss of Ψm and subsequent apoptosisThis kind of Ψm reduction and generation of MPTP mustfacilitate the relocalization of apoptogenic proteins from onesubcellular compartment such as mitochondria to cytoplasmto gain access to their substrates or interacting partners [37]
The multi cytotoxicity assay shows that girinimbine ind-uced cell death could be through the classical mitochondrial
pathway with cytochrome c release and caspase-dependentapoptosis The translocation of cytochrome c a componentof themitochondrial electron transfer chain was significantlyincreased upon girinimbine treatment In the mitochondriamediated apoptosis cytochrome c in turn binds to Apaf-1 to form a complex which triggers its oligomerization toform the apoptosome The caspase-9 holoenzyme is thenforms due to the binding of procaspase-9 to the apop-tosome and sequentially cleaves and activates the down-stream caspases such as caspase-3 [38] Our findings onthe upregulation of caspase 37 caspase 9 reduction in Ψmand release of cytochrome c to the cytosol substantiate thistheory and strongly support the involvement ofmitochondriain the apoptosis induced by girinimbine in A549 cells
Evidence-Based Complementary and Alternative Medicine 9
bad
bax
bcl-2
bcl-w
BID BI
MCa
spas
e 3Ca
spas
e 8IG
F-II IG
F-I HTR
AH
SP70
HSP
60H
SP27
FasL Fa
sD
R6Cy
to c
IGFB
P-1
IGFB
P-2 IG
FBP-
3IG
FBP-
4IG
FBP-
5IG
FBP-
6IG
F-1s
RLi
vin p
21 p
27 p53
SM
AC S
urvi
vin
sTN
F-R1 XI
AP
minus3
minus2
minus1
0
1
2
3
Relat
ive d
ensit
y di
ffere
nce t
o co
ntro
l (times10
5)
(a)
(b) (c)
A B C D E F G H I J K L M N1 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 82 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 83 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II4 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II5 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R16 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R17 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg Pos8 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg PosTNF-120572
TNF-120572 TNF-120573TNF-120573
(d)
Figure 7 Cells were lysed and protein arrays were performed Cells was treated with 19120583M girinimbine for 24 h and the whole cell proteinwas extracted Equal amount of (300 120583g) of protein from each sample was used for the assay Quantitative analysis in the arrays showeddifferences in the apoptotic markers (a) Representative images of the apoptotic protein array are shown for the control (b) treatment (c) andthe exact protein name of each dot in the array (d)
Suppression of apoptosis may promote the cancer develop-ment by inhibiting the chemotherapy as well as other forms ofinducers of cell death [39] Regulation of apoptosis has beencensoriously dependent on several genes which have beenidentified earlier and includes XIAP-a member of the IAPfamily It has the capacity to inhibit the activation of caspases3 7 and 9 [40] In our study we found that girinimbinetreatment of A549 cells caused inhibition of XIAP but SMAC
an antagonist to XIAP could be found to be slightly regulatedin this treatment concentration This is considered as a keyevent in the execution of cell death which is regulatedmainlyby proteins of the Bcl-2 family [41 42]
Critical issues in apoptosis include the dominance ofanti-versus proapoptotic Bcl-2 members [43] Girinimbine isfound to induce apoptosis in the A549 cells involvingdownregulated Bcl-2 expression This downregulation was
10 Evidence-Based Complementary and Alternative Medicine
3
2
1
0
times105
19 9 475 0 19 9 475 0 19 9 475 0Girinimbine treatment (120583M)
Lum
ines
cenc
e
Caspase 37Caspase 8Caspase 9
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast
Figure 8 Effect of girinimbine on activation of caspases 37 8 and 9Cells were treated with girinimbine at the indicated concentrationsfor 24 h and activity was determined by using a luminescentenzyme assay kit Each point represents the mean plusmn SD of threedifferent experiments Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software(version 40 GraphPad Software Inc San Diego CA) Statisticalsignificance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
concomitant with the upregulation of Bad Bax and Bimwhich inhibits antiapoptotic proteins and render the cellsmore susceptible to apoptogenic stimuli (Figure 7) Therelease of cytochrome c found in our study could be due tothe up regulated Bax since Bax in association with adeninenucleotide translocator within the permeability transitionpore complex will increase the mitochondrial membranepermeability and thereby discharge a number of apopto-genic molecules into the cytosol [44] In addition to thisupregulation of the tumor suppressor protein p53 which hasa critical role in regulation of the Bcl-2 and p27 and p21draws attention to the involvement of cell cycle arrest in theapoptosis process Alternatively we found the involvement ofextrinsic pathway proteins such as Fas and FasL upregulationand related upregulation of caspase 8 (Figure 7) Moreoverthe luminescent based caspase 8 assay had showed 10 folddifferences than the control Apart from this we foundthat there is a downregulation of Bid happening too Sincemitochondrial damage in the Fas pathway of apoptosis ismediated by the caspase 8 cleavage of Bid [45] the findingshere shows the cross talk between intrinsic and extrinsicpathways
The protein array results exhibited a significant reductionof both IGF-I and IGF-II IGF-I has been shown to exertstrongmitogenic and antiapoptotic effects in a variety of nor-mal and cancerous cells including lung cancer cell lines [46]Moreover the over expression of IGFBP3 is closely associatedwith the downregulation of IGF-I and its mitogenic activity[47] It is known that IGFBP-2 and IGFBP-1 are involved inthe regulation of cell migration apoptosis and cell growth
[48] Both are reported to be markedly overexpressed inmany tumors and tumor cell lines And it is assumed thatelevation of the IGFBP-2 production is part of a mechanismto compensate for the mitogenic and antiapoptotic effectsof tumor-derived IGFs [49] Our data clearly showed thedownregulation of IGFBP1 and IGFBP2 concomitant withupregulation of IGBP3
Our results provided a new insight into the mechanismof chemotherapeutic properties of Murraya koenigii Inaddition these study findings demonstrate that girinimbinemediates its apoptotic effects through both intrinsic andextrinsic pathway which is depended on caspase mediationMoreover the upregulation of p53 as well as the cell prolifer-ation repressor proteins p27 and p21 and the significant roleof insulinIGF-1 signaling were also identified These resultsindicate that girinimbine could be a candidate for a novelanticancer agentThereforemore in-depth in vitro and in vivostudies are currently going on in our laboratory
Conflict of Interests
The authors declare that they do not have any conflict ofinterests
Acknowledgment
The authors greatly acknowledge the financial support pro-vided by University of Malaya (Grant no HIR E0000002-20001)
References
[1] J K Field and S W Duffy ldquoLung cancer screening the wayforwardrdquo British Journal of Cancer vol 99 no 4 pp 557ndash5622008
[2] K A Yoon J H Park J Han et al ldquoA genome-wide associationstudy reveals susceptibility variants for nonsmall cell lungcancer in the Korean populationrdquo Human Molecular Geneticsvol 19 no 24 Article ID ddq421 pp 4948ndash4954 2010
[3] G Cartei F Cartei A Cantone et al ldquoCisplatin-cyclophos-phamide-mitomycin combination chemotherapy with support-ive care versus supportive care alone for treatment of metastaticnon- small-cell lung cancerrdquo Journal of the National CancerInstitute vol 85 no 10 pp 794ndash800 1993
[4] M O Hengartner ldquoThe biochemistry of apoptosisrdquoNature vol407 no 6805 pp 770ndash776 2000
[5] DHanahan andRAWeinberg ldquoThehallmarks of cancerrdquoCellvol 100 no 1 pp 57ndash70 2000
[6] J R Molina P Yang S D Cassivi S E Schild and A AAdjei ldquoNon-small cell lung cancer epidemiology risk factorstreatment and survivorshiprdquo Mayo Clinic Proceedings vol 83no 5 pp 584ndash594 2008
[7] S Diwanay D Chitre and B Patwardhan ldquoImmunoprotec-tion by botanical drugs in cancer chemotherapyrdquo Journal ofEthnopharmacology vol 90 no 1 pp 49ndash55 2004
[8] U M Thatte and S A Dahanukar ldquoRasayana Concept cluesfrom immunomodulatory therapyrdquo in An Annotated Bibliogra-phy of Indian Medicine S N Upadhyay Ed pp 141ndash148 1997
[9] A S Shah A S Wakade and A R Juvekar ldquoImmunomod-ulatory activity of methanolic extract of Murraya koenigii (L)
Evidence-Based Complementary and Alternative Medicine 11
Spreng leavesrdquo Indian Journal of Experimental Biology vol 46no 7 pp 505ndash509 2008
[10] R B Goswamia P K S Singha N Goswamib P Thomasd PU Devic and A K Pathakd ldquoStudies on antigenotoxic effect ofMurraya koenigii leavesrdquo International Journal of PharmaRecentResearch vol 2 pp 65ndash68 2010
[11] R S Ramsewak M G Nair G M Strasburg D L DeWittand J L Nitiss ldquoBiologically active carbazole alkaloids fromMurraya koenigiirdquo Journal of Agricultural and Food Chemistryvol 47 no 2 pp 444ndash447 1999
[12] D P Chakraborty B K Barman and P K Bose ldquoStructureof girinimbine a pyranocarbazole derivative isolated fromMurraya koenigii Sprengrdquo Scientific Culture vol 30 pp 445ndash448 1964
[13] L J M Rao K Ramalakshmi B B Borse and B RaghavanldquoAntioxidant and radical-scavenging carbazole alkaloids fromthe oleoresin of curry leaf (Murraya koenigii Spreng)rdquo FoodChemistry vol 100 no 2 pp 742ndash747 2007
[14] S L Wang B Cai C B Cui S Y Yan and C F Wu ldquoInductionof apoptosis by girinimbine in K562 cellrdquo Chinese Traditionaland Herbal Drugs vol 38 no 11 pp 1677ndash1681 2007
[15] S Syam A B Abdul M A Sukari S Mohan S I Abdelwahaband T S Wah ldquoThe growth suppressing effects of girinimbineon Hepg2 involve induction of apoptosis and cell cycle arrestrdquoMolecules vol 16 pp 7155ndash7170 2011
[16] A Bakar N Haslizawati M A Sukari et al ldquoChemical con-stituents from stem barks and roots of Murraya koenigii(Rutaceae)rdquo The Malaysian Journal of Analytical Sciences vol11 pp 173ndash176 2007
[17] S I A Wahab A B Abdul S M Mohan A S Al-ZubairiM M Elhassan and M Y Ibrahim ldquoBiological activities ofPereskia bleo extractsrdquo International Journal of Pharmacologyvol 5 no 1 pp 71ndash75 2009
[18] E Bossy-Wetzel and D R Green ldquoDetection of apoptosis byannexin V labelingrdquo Methods in Enzymology vol 322 pp 15ndash18 2000
[19] H Wang and J A Joseph ldquoQuantifying cellular oxidative stressby dichlorofluorescein assay usingmicroplate readerrdquo Free Rad-ical Biology and Medicine vol 27 no 5-6 pp 612ndash616 1999
[20] S C Cheah D R Appleton S T Lee M L Lam A H AHadi and M R Mustafa ldquoPanduratin a inhibits the growth ofA549 cells through induction of apoptosis and inhibition of NF-KappaB translocationrdquoMolecules vol 16 no 3 pp 2583ndash25982011
[21] L Foghsgaard DWissing DMauch et al ldquoCathepsin B acts asa dominant execution protease in tumor cell apoptosis inducedby tumor necrosis factorrdquo Journal of Cell Biology vol 153 no 5pp 999ndash1009 2001
[22] K Kagedal U Johansson and K Ollinger ldquoThe lysosomalprotease cathepsin D mediates apoptosis induced by oxidativestressrdquoThe FASEB journal vol 15 no 9 pp 1592ndash1594 2001
[23] M A Birch-Machin and H Swalwell ldquoHow mitochondriarecord the effects of UV exposure and oxidative stress usinghuman skin as a model tissuerdquo Mutagenesis vol 25 no 2 pp101ndash107 2010
[24] S Meister B Frey V R Lang et al ldquoCalcium channel blockerverapamil enhances endoplasmic reticulum stress and cell deathinduced by proteasome inhibition in myeloma cellsrdquoNeoplasiavol 12 no 7 pp 550ndash561 2010
[25] S Mohan A B Abdul S I Abdelwahab et al ldquoTyphoniumflagelliforme induces apoptosis in CEMss cells via activation
of caspase-9 PARP cleavage and cytochrome c release itsactivation coupled with G0G1 phase cell cycle arrestrdquo Journalof Ethnopharmacology vol 131 no 3 pp 592ndash600 2010
[26] S Elmore ldquoApoptosis a review of programmed cell deathrdquoToxicologic Pathology vol 35 no 4 pp 495ndash516 2007
[27] M Aberg M Johnell M Wickstrom and A Siegbahn ldquoTissueFactor FVIIa prevents the extrinsic pathway of apoptosis byregulation of the tumor suppressor Death-Associated ProteinKinase 1 (DAPK1)rdquoThrombosis Research vol 127 no 2 pp 141ndash148 2011
[28] M Nigam N Singh V Ranjan et al ldquoCentchroman mediatedapoptosis involves cross-talk between extrinsicintrinsic path-ways and oxidative regulationrdquo Life Sciences vol 87 no 23ndash26pp 750ndash758 2010
[29] O S Frankfurt and A Krishan ldquoApoptosis-based drug screen-ing and detection of selective toxicity to cancer cellsrdquo Anti-Cancer Drugs vol 14 no 7 pp 555ndash561 2003
[30] M van Engeland L J W Nieland F C S Ramaekers BSchutte and C P M Reutelingsperger ldquoAnnexin V-affinityassay a review on an apoptosis detection system based onphosphatidylserine exposurerdquo Cytometry vol 31 no 1 pp 1ndash91998
[31] G A Joanitti R B Azevedo and S M Freitas ldquoApoptosisand lysosome membrane permeabilization induction on breastcancer cells by an anticarcinogenic Bowman-Birk proteaseinhibitor from Vigna unguiculata seedsrdquo Cancer Letters vol293 no 1 pp 73ndash81 2010
[32] T Kirkegaard and M Jaattela ldquoLysosomal involvement in celldeath and cancerrdquo Biochimica et Biophysica ActamdashMolecularCell Research vol 1793 no 4 pp 746ndash754 2009
[33] A C Johansson H Appelqvist C Nilsson K Kagedal KRoberg and K Ollinger ldquoRegulation of apoptosis-associatedlysosomal membrane permeabilizationrdquo Apoptosis vol 15 no5 pp 527ndash540 2010
[34] J Wen K R You S Y Lee C H Song and D G KimldquoOxidative stress-mediated apoptosis the anticancer effect ofthe sesquiterpene lactone parthenoliderdquo Journal of BiologicalChemistry vol 277 no 41 pp 38954ndash38964 2002
[35] C Fleury B Mignotte and J L Vayssiere ldquoMitochondrialreactive oxygen species in cell death signalingrdquo Biochimie vol84 no 2-3 pp 131ndash141 2002
[36] A M Verhagen P G Ekert M Pakusch et al ldquoIdentificationof DIABLO a mammalian protein that promotes apoptosis bybinding to and antagonizing IAP proteinsrdquo Cell vol 102 no 1pp 43ndash53 2000
[37] N Ajenjo E Canonon I Sanchez-Perez et al ldquoSubcellularlocalization determines the protective effects of activated ERK2against distinct apoptogenic stimuli in myeloid leukemia cellsrdquoJournal of Biological Chemistry vol 279 no 31 pp 32813ndash328232004
[38] X Wang ldquoThe expanding role of mitochondria in apoptosisrdquoGenes and Development vol 15 no 22 pp 2922ndash2933 2001
[39] Y A Hannun ldquoApoptosis and the dilemma of cancer chemo-therapyrdquo Blood vol 89 no 6 pp 1845ndash1853 1997
[40] F L Scott J B Denault S J Riedl H Shin M Renatusand G S Salvesen ldquoXIAP inhibits caspase-3 and -7 usingtwo binding sites evolutionary conserved mechanism of IAPsrdquoEMBO Journal vol 24 no 3 pp 645ndash655 2005
[41] S Cory and J M Adams ldquoThe BCL2 family regulators of thecellular life-or-death switchrdquo Nature Reviews Cancer vol 2 no9 pp 647ndash656 2002
12 Evidence-Based Complementary and Alternative Medicine
[42] S B Bratton and GM Cohen ldquoDeath receptors leave a caspasefootprint that Smacs of XIAPrdquo Cell Death and Differentiationvol 10 no 1 pp 4ndash6 2003
[43] N Zamzami andGKroemer ldquoThemitochondrion in apoptosishow Pandorarsquos box opensrdquo Nature Reviews Molecular CellBiology vol 2 no 1 pp 67ndash71 2001
[44] I Marzo C Brenner N Zamzami et al ldquoBax and adeninenucleotide translocator cooperate in the mitochondrial controlof apoptosisrdquo Science vol 281 no 5385 pp 2027ndash2031 1998
[45] M D Esposti ldquoThe roles of BidrdquoApoptosis vol 7 no 5 pp 433ndash440 2002
[46] N E Allen A W Roddam D S Allen et al ldquoA prospectivestudy of serum insulin-like growth factor-I (IGF-I) IGF-IIIGF-binding protein-3 and breast cancer riskrdquo British Journalof Cancer vol 92 no 7 pp 1283ndash1287 2005
[47] L Buckbinder R Talbott S Velasco-Miguel et al ldquoInductionof the growth inhibitor IGF-binding protein 3 by p53rdquo Naturevol 377 no 6550 pp 646ndash649 1995
[48] E Ruoslahti ldquoFibronectin and its integrin receptors in cancerrdquoAdvances in Cancer Research vol 76 pp 18ndash20 1999
[49] B S Schutt M Langkamp U Rauschnabel M B Ranke andM W Elmlinger ldquoIntegrin-mediated action of insulin-likefactor binding protein-2 in tumor cellsrdquo Journal of MolecularEndocrinology vol 32 no 3 pp 859ndash868 2004
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 3
22 Cell Culture A549 a human nonsmall cell lung cancercell line was purchased from ATCC (Rockville MD USA)and cultured in RPMI-1640 supplemented with 100 120583LmLfetal bovine serum 100UmL penicillin and 100mgmLstreptomycin sulfate Cells were maintained at 37∘C in ahumidified atmosphere of 5 CO
2 Cells were treated with
the girinimbine dissolved in DMSO while the untreatedcontrol cultures received only the vehicle (DMSO lt 1)
23 Cell Viability Assay Cells were seeded in 96-well platesat a density of 1 times 104 per well and treated with various con-centrations of girinimbine for 24 h Cells were then incubatedwith a medium containing 5mgmL 3-(45-dimethylthiazol-2-yl)-25-diphenyltetrazolium bromide (MTT) for another4 h The viable cell number was directly proportional toformazanproductionwhich thendissolved inDMSOandwasmeasured by spectrophotometry at 563 nm in a microplatereader (Tecan Infinite M 200 PRO Mannedorf Switzerland)[17]
24 Microscope Examination on Cellular Morphology Apop-tosis was monitored by annexin V labeling and fluorescencemicroscopy [18] Treated and untreated cells were washedwith PBS and then exposed with annexin V-fluorescein (BDPharmingen USA) for 15mins After a 488 nm excitationgreen fluorescence was visualized and recorded at 515 nmPhase contrast microscopic images from the same prepara-tions were also obtained for symptoms of apoptosis such ascell shrinkage ruffling and blebbing of cell membrane as wellas fragmentation of cells into small apoptotic bodies usinga fluorescent microscope (Nikon TE 2000U fluorescenceinverted microscope Tallahassee Florida)
25 Determination of Lysosomal Membrane Stability Cellswere assessed for lysosomal stability using acridine orange(AO) uptake assay Briefly cells were incubated with 7120583MAO (Sigma USA) and 12 120583M Hoechst 33342 (InvitrogenCarlsbad CA USA) diluted in culture media for 15minsat 37∘C and immediately observed and analyzed using theArrayScan HCS system (Cellomics PA USA) Acridineorange is ametachromatic fluorochrome and aweak base thatexhibits red fluorescence when highly concentrated in acidiclysosomes
26Measurement of Reactive Oxygen Species Generation Theproduction of intracellular reactive oxygen species (ROS)wasmeasured using 2101584071015840-dichlorofluorescein diacetate (DCFH-DA) [19] DCFH-DA passively enters the cell where itreacts with ROS to form the highly fluorescent compounddichlorofluorescein (DCF) Briefly 10mM DCFH-DA stocksolution (in methanol) was diluted 500-fold in HBSS withoutserum or other additives to yield a 20120583M working solutionAfter 24 h of exposure to girinimbine the cells in the 96-wellblack plate were washed twice with HBSS and then incubatedin 100 120583L working solution of DCFH-DA at 37∘C for 30minsFluorescence was then determined at 485 nm excitation and520 nm emission using a fluorescence microplate reader(Tecan Infinite M 200 PRO Mannedorf Switzerland)
27 Multiple Cytotoxicity Assay Cellomics MultiparameterCytotoxicity 3 Kit was used as described in detail previously[20]This kit enables simultaneousmeasurements in the samecell of six independent parameters that monitor cell healthincluding cell loss nuclear size and morphological changesmitochondrial membrane potential changes cytochrome crelease and changes in cell permeability Plates were analyzedusing the ArrayScan HCS system (Cellomics PA USA)
28 Image Acquisition and Cytometric Analysis Plates withstained cells were analyzed using the ArrayScan HCS sys-tem (Cellomics PA USA) This system is a computerizedautomated fluorescence imaging microscope that automat-ically identifies stained cells and reports the intensity anddistribution of fluorescence in individual cells The Array-Scan HCS system scans multiple fields in individual wellsto acquire and analyze images of single cells according todefined algorithms In each well 1000 cells were analyzedAutomatic focusing was performed in the nuclear channelto ensure focusing regardless of staining intensities in theother channels Images were acquired for each fluorescencechannel using suitable filters Images and data regardingintensity and texture of the fluorescence within each cell aswell as the average fluorescence of the cell population withinthe well were stored in a Microsoft SQL database for easyretrieval Data were captured extracted and analyzed withArrayScan II Data Acquisition and Data Viewer version 30(Cellomics)
29 HumanApoptosis Proteome Profiler Array To investigatethe pathways by which girinimbine induces apoptosis weperformed a determination of apoptosis-related proteinsusing the Proteome Profiler Array (RayBio Human Apop-tosis Antibody Array Kit RayBiotech USA) according tomanufacturerrsquos instructions In short the cells where treatedwith 19 120583M girinimbine 300 120583g proteins from each sam-ple and were incubated with the human apoptosis arrayovernight The apoptosis array data were quantified by scan-ning the membrane on a Biospectrum AC ChemiHR 40(UVP Upland CA) and analysis of the array image file wasperformed using image analysis software according to themanufacturerrsquos instruction
210 Measurement of Caspases 8 9 and 37 Activities Cas-pases 37 8 and 9 activity wasmeasured using luminescence-based assay Caspase-Glo 8 Assay Caspase-Glo 9 Assay andCaspase-Glo 37 Assay (Promega) 1 times 104 cells were culturedin 96-well culture plates in 50 120583L of RPMI 1640 supplementedwith 10 FBS and incubated for 24 h Cells then were treatedwith different concentrations of girinimbine and incubatedfor 24 h At the end of incubation 100 120583L of assay reagent wasadded to be incubated for 1 h at room temperature Lumines-cence wasmeasured using amicroplate reader (Tecan InfiniteM 200 PRO Mannedorf Switzerland)
211 Statistical Analysis From several independent measure-ments means and standard deviations were calculated Test-ing for significant differences between means were carried
4 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d) (e) (f)
Figure 2 Effects of girinimbine on the morphological changes of A549 cells after 24 h treatment Apoptosis was monitored by phase contrast((a) control (b) treatment 9120583M and (c) treatment 19120583M) and fluorescence microscopy ((d) control (e) treatment 9120583M and (f) treatment19120583M) Arrows indicate the cell shrinkage ruffling and blebbing of cell membrane while arrowheads indicate the annexin V binding tophosphatidylserine Magnification 20X
out using the one-way ANOVA and Dunnettrsquos post-test atprobabilities of error of 5 and 1
3 Results
31 Antipoliferative Activity The sigmoidal dose responsecurves of girinimbine in the end-point assays are shown inFigure 1 Cell viability was analyzed using the MTT assaywhich measures the metabolic activity of cell In the A549cells treated with girinimbine metabolic activity decreasedfollowed by 24 h treatment meanwhile in the control platecell viability and metabolism were not affected Figure 1(c)summarizes the IC
50values from MTT Meanwhile even at
380 120583M girinimbine could not exhibit any sign of toxicity inWRL-68 a normal (data not shown) which was employed inthis study to investigate the specificity of cytotoxicity
32 Apoptotic Mode of Cell Death Data obtained fromfluorescence detection of A549 cells together with the phasecontrast microscopic images are shown (Figure 2) Apoptosiswas clearly detected by the phosphatidylserine externaliza-tion on the treated cells Phase contrast microscopic picturesshowed a clear morphological change of treated cells whichwere observed after 24 h of treatment with girinimbine A549cells were seen to have shrunk in size and there was rufflingand blebbing of cell membranes thus suggesting that the cellswere undergoing apoptosis (Figure 2(c))
33 Girinimbine Initiates Lysosomal Membrane Permeabiliza-tion Recent studies have shown that lysosomal membrane
permeabilization (LMP) is an early and perhaps initiatingevent in apoptosis triggered by ligation of death recep-tors lysosomotropic agents oxidative stresses or serumwithdrawal [21 22] To evaluate the lysosome acidificationpattern the cells were analyzed by acridine orange staininga known pH indicator While untreated A549 cells dis-played strong granular acridine orange staining cells treatedwith girinimbine presented weak lysosome staining patternand cytoplasm acidification indicating lysosomal membranepermeabilization (Figures 3(a)ndash3(c)) Girinimbine induceddose-dependent decrease of fluorescence intensity of lyso-somal staining which was well compared with chloroquinea known inhibitor of lysosome function was included as apositive control (Figure 3(d))
34 ROS Assay Numerous investigations have documentedthat oxidative stress-mediated cellular changes are fre-quently induced in cells exposed to cytotoxic drugs UV orgamma irradiation [23 24] We examined whether girin-imbine affects the cellular levels of peroxide by measur-ing the changes in the fluorescence using DCF-DA Asshown in Figure 4(a) treatment with girinimbine markedlyincreased the DCF-DA-derived fluorescence (520 nm) Thisgirinimbine-mediated increase in fluorescence was markedlyinhibited by pretreatment with antioxidant ascorbic acidThen we next asked whether ROS generation induced bygirinimbine is directly associated with the induction ofapoptosis Chromatin condensation was measured in thecells which were pretreated with ascorbic acid using 12120583MHoechst 33342 The results shown in Figure 4(b) clearly
Evidence-Based Complementary and Alternative Medicine 5
(a) (b) (c)
200
100
0
Treatment
Fluo
resc
ent i
nten
sity
()
Chlo
roqu
ine25120583
M
Con
trol24
h
Giri
nim
bine
10120583
gm
L
Giri
nim
bine
5120583
gm
L
Giri
nim
bine
25120583
gm
L
(d)
Figure 3 Effects of girinimbine onA549 cells lysosome acidic environment Cells were incubatedwith different concentrations of girinimbinefor 24 h stained with acridine orange and analyzed by ArrayScan HCS system Control group corresponds to untreated cells Acridine orangeis a metachromatic fluorochrome and a weak base that exhibits red fluorescence when highly concentrated in acidic lysosomes (a) Normallysosome acidic environment (arrow)Notice theweak lysosome staining pattern and cytoplasm acidification (arrowhead) in cells treatedwithchloroquine (b) and girinimbine (c) indicating lysosomal membrane permeabilization Magnification 20X The mean fluorescent intensityproduced by the acridine orange was quantitatively measured (Figure 3(d))
exhibited that pretreatment with ascorbic acid could notprevent the apoptosis at various time periods Moreover thedata clearly shows the rapid decrease in the nuclear area andthe increase in fragmentation upon treatment with 19 120583Mgirinimbine (Figure 4(c))
35 Multiparameter Cytotoxicity Analysis This assay enablessimultaneousmeasurement of several cell-health parametersnuclear morphology DNA content cell membrane perme-ability and cytochrome c localization and release frommito-chondria Typical cytotoxic changes are illustrated in Figures5 and 6 Girinimbine induced decreases in cell numbernuclear area intensity and plasma membrane permeabilitywere significantly higher in the treated cells (119875 lt 001) andmitochondrial membrane potential reduction was observedsignificantly at 19120583M (119875 lt 001) The release of cytochromec was observed significantly in 9120583M and 19120583M with 119875 lt005 and 119875 lt 001 respectively These effects occurredmore rapidly and followed a dose-response pattern Cytotoxiceffects were considered to occur only when the rate of changeof fluorescence was distinctly greater than for the negativecontrols
36 Effect of Girinimbine on Apoptotic Markers After girin-imbine (19120583M) exposure A549 cells were lysed and apoptoticmarkers where screened using a protein array All majormarkers which are involved in both intrinsic and extrinsicpathways were induced on treatment As shown in Figure 7girinimbine treatment significantly increased the expressionof caspase 8 suggesting the activation of the death receptorpathway In addition the major protein involved in theextrinsic pathway such as Fas and FasL also regulates thetreatment Moreover the involvement of mitochondria in thecell death was evident by the regulation of the Bcl-2 familyof proteins such as Bad Bax Bcl-2 and Bim Besides theBcl-2 family member Bid was found to be cleaved as wellsuggesting a potential cross-talk between the death receptorand the mitochondrial pathway The treatment also resultedin a reduction in the level of expression of the inhibitorof apoptosis XIAP as well as survivin P53 as well as thecell proliferation repressor proteins p27 and p21 and theheat shock proteins such as antiapoptotic HSP60 HSP70and HSP27 which are a result of oxidative stress in the cellwere also induced InsulinIGF-1 signaling related proteinexpression has been observed on treatment Downregulation
6 Evidence-Based Complementary and Alternative Medicine
800850900950
100010501100115012001250130013501400
Control asaTreatment
24h 24h 8h 8h 4h 4h
Fluo
resc
ence
inte
nsity
(520
nm)
Girinimbine(19120583M)
Ascorbic acid
minus
minus minus minus minus
+ +
+
+ +
+
+ +
+ +
+
lowastlowast
lowast
lowastlowast
(a)
750
500
250
024h 12h 6h Control
Treatment
Fluo
resc
ent i
nten
sity
units
lowastlowast
lowastlowast
Girinimbine(19120583M)
Ascorbic acid
minus
minus
+
+
+
+
+
+
(b)
lowast
lowastlowastlowast
lowastlowast
Aver
age n
ucle
ar o
bjec
t siz
e
15
125
10
Girinimbine(19120583M)
Ascorbic acid
minus
minus
+
+
+
+
+
+
24h 12h 6h ControlTreatment
(c)
Figure 4 Effects of girinimbine on A549 cells ROS generation (a) DCF-fluorescence intensity after ascorbic acid and 19 120583M of girinimbineexposure at 4 8 and 24 h Rate of apoptosis in terms of fluorescent intensity (b) and average nuclear object size (c) of nucleus of the girinimbinetreatment was pretreatedwith 100mMascorbic acid Values aremeanplusmn SD from three independent experiments Triplicates of each treatmentgroup were used in each independent experiment The statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005 Fluorescent intensityunits directly represent the amount of ROS production
of antiapoptosis (IGF-I IGF-II IGFBP1 IGFBP2) and upreg-ulation of proapoptosis proteins (IGFBP3 IGFBP4) wereobserved
37 Caspase Depended Apoptosis in Girinimbine Treated A549Cells The involvement of the caspase cascade in the girin-imbine mediated cell death was confirmed The treatmentwith 19 120583M was significant at 119875 lt 001 for caspase 37and 9 with 8- and 5-fold increase respectively Meanwhilethe caspase 8 showed 10-fold increase at the maximumtreatment concentration (19 120583M 119875 lt 001) (Figure 8)These results are in parallel with the protein array andindicate that the signaling cascade leading to apoptosis in
girinimbine-treated cells involves both intrinsic and extrinsicpathways
4 Discussion
Apoptosis is a normal physiological process that plays avital role in numerous normal functions [4] Furthermoreit is an active physiological process causing cellular self-destruction that comprises specific morphological and bio-chemical changes in the nucleus and cytoplasm [25] Theinvolvement of an energy-dependent cascade of molecularevents makes the mechanism of apoptosis very highly com-plex and sophisticated [26] Apoptosis is regulated by two
Evidence-Based Complementary and Alternative Medicine 7
Hoechst 33342 Cell permeability Cytochrome c
Con
trol
Giri
nim
bine
Mitochondrial
membrane potential
Figure 5 Representative images of A549 cells treated withmedium alone and 19 120583Mof girinimbine and stained with Hoechst for nuclear cellpermeability dye mitochondrial membrane potential dye and cytochrome c The images from each row are obtained from the same field ofthe same treatment sample A549 produced a marked reduction in mitochondrial membrane potential and marked increases in membranepermeability and cytochrome c Magnification 20X
major pathways the extrinsic and the intrinsic pathways[27] Nevertheless there is evidence suggesting that boththe pathways are linked and that molecules involved in thepathways can influence one another [28]
The current study found that girinimbine a carbazolealkaloid from the roots of M koenigii can inhibit cellproliferation selectively in a dose dependent manner inA549 cells The morphological observation was conductedto explore whether the cytotoxic effect was related withthe apoptotic process and it was found that the cell deathinduced by girinimbine exhibited a clear morphologicalsign of apoptosis as this is an important property of acandidate anticancer drug [29] Followed by 24 h treatmentcell shrinkage ruffling and blebbing of cell membranes wereobserved These morphological features were more evidentwhile we analysed one of the key features of apoptosis whichis the change in plasma membrane structure by surfaceexposure of phosphatidylserine (PS) while the membraneintegrity remains unchallenged [30] The incorporation ofannexinV in to the same treated cells showed distinctive formof binding between the externalized PS and Annexin V whilecells from test flasks showedmore evidence of apoptosis thanthose grown under control conditions (Figure 2)
Translocation of lysosomal hydrolases from the lysosomallumen to the rest of the cell followed by the impairmentin the lysosomal membrane function is known as lysoso-mal membrane permeabilization (LMP) [31] Often thesehydrolases facilitate apoptosis by inducing mitochondrialouter-membrane permeabilization and caspase activation
[32] In our results the main LMP consequence such as weaklysosome acidification pattern and cytoplasmic acidificationwas clearly evident (Figure 3) Being aweak base chloroquineacts as lysosomotropic drug that raises intralysosomal pHHence we have monitored the release of liposomal contentby using chloroquine as a positive control Moreover in theFigure 3 it is very clear that cells treated with girinimbineresented a weak lysosome acidification pattern followed byan increase in cytoplasmic acidification Bearing in mind thesignificance of acidic lysosome environment for proteins andorganelles degradation it can be pointed out that autophagymay not the main event of cell death
Destabilization of the lysosomal membrane may becaused by increased production of ROS via massive peroxi-dation of membrane lipids [33] Moreover many studies hadshowed the relation between ROS and apoptosis induced incells exposed to cytotoxic drugs [34] Mitochondrial ROS areused as active mediators in the regulation of cell death [35]In our study the generation of ROS led to the upregulation ofcytoprotective protein markers HSP 60 70 and 27 Figure 4clearly shows that girinimbine exposure leads to increase ofROS accumulation but inhibition of ROSby ascorbic acid didnot prevent girinimbine-induced apoptosis indicating thatROS generation is not critical for the induction of apoptosisby girinimbine in A549 cells Moreover the treatment withgirinimbine significantly decreases the cell number nucleararea and cell membrane permeability as shown by themultiparameter apoptosis analysis In addition the complexrole of mitochondria in A549 cell apoptosis was investigated
8 Evidence-Based Complementary and Alternative Medicine
600
500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Total nuclear intensity
Giri19120583
M
Giri95120583
M
Giri475120583
M
(a)
Cell permeability500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(b)
350
300
250
200
150
100
50
0
Mitochondrial membrane potential
Con
trol
Pacli
taxe
l68120583
M
lowastlowastlowastlowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(c)
600700800
500400300200100
0
Cytochrome c
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowastlowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(d)
Figure 6 Quantitative analysis of girinimbine mediated apoptosis parameter Changes in total nuclear intensity (a) cell permeability(b) mitochondrial membrane potential (c) and cytochrome c localization (d) were measured simultaneously in A549 cells Followingtreatment with girinimbine we saw statistically significant increase in total nuclear intensity increased cell permeability loss ofmitochondrialmembrane potential and cytochrome c release frommitochondria Each experiment was performed at least two times Results are expressedas the means plusmn SD Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software (version40 GraphPad Software Inc San Diego CA) Statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
Mitochondria have been described to play a central role in theapoptotic process due to the mitochondrial proteins ability toactivate cellular apoptotic programs directly [36] Detectionof changes in mitochondrial membrane potential (Ψm) wascarried out as it is assumed that its disruption is the onsetofmitochondrial membrane transition pores (MPTP) forma-tion The present study revealed that girinimbine may act onmitochondria causing loss of Ψm and subsequent apoptosisThis kind of Ψm reduction and generation of MPTP mustfacilitate the relocalization of apoptogenic proteins from onesubcellular compartment such as mitochondria to cytoplasmto gain access to their substrates or interacting partners [37]
The multi cytotoxicity assay shows that girinimbine ind-uced cell death could be through the classical mitochondrial
pathway with cytochrome c release and caspase-dependentapoptosis The translocation of cytochrome c a componentof themitochondrial electron transfer chain was significantlyincreased upon girinimbine treatment In the mitochondriamediated apoptosis cytochrome c in turn binds to Apaf-1 to form a complex which triggers its oligomerization toform the apoptosome The caspase-9 holoenzyme is thenforms due to the binding of procaspase-9 to the apop-tosome and sequentially cleaves and activates the down-stream caspases such as caspase-3 [38] Our findings onthe upregulation of caspase 37 caspase 9 reduction in Ψmand release of cytochrome c to the cytosol substantiate thistheory and strongly support the involvement ofmitochondriain the apoptosis induced by girinimbine in A549 cells
Evidence-Based Complementary and Alternative Medicine 9
bad
bax
bcl-2
bcl-w
BID BI
MCa
spas
e 3Ca
spas
e 8IG
F-II IG
F-I HTR
AH
SP70
HSP
60H
SP27
FasL Fa
sD
R6Cy
to c
IGFB
P-1
IGFB
P-2 IG
FBP-
3IG
FBP-
4IG
FBP-
5IG
FBP-
6IG
F-1s
RLi
vin p
21 p
27 p53
SM
AC S
urvi
vin
sTN
F-R1 XI
AP
minus3
minus2
minus1
0
1
2
3
Relat
ive d
ensit
y di
ffere
nce t
o co
ntro
l (times10
5)
(a)
(b) (c)
A B C D E F G H I J K L M N1 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 82 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 83 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II4 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II5 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R16 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R17 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg Pos8 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg PosTNF-120572
TNF-120572 TNF-120573TNF-120573
(d)
Figure 7 Cells were lysed and protein arrays were performed Cells was treated with 19120583M girinimbine for 24 h and the whole cell proteinwas extracted Equal amount of (300 120583g) of protein from each sample was used for the assay Quantitative analysis in the arrays showeddifferences in the apoptotic markers (a) Representative images of the apoptotic protein array are shown for the control (b) treatment (c) andthe exact protein name of each dot in the array (d)
Suppression of apoptosis may promote the cancer develop-ment by inhibiting the chemotherapy as well as other forms ofinducers of cell death [39] Regulation of apoptosis has beencensoriously dependent on several genes which have beenidentified earlier and includes XIAP-a member of the IAPfamily It has the capacity to inhibit the activation of caspases3 7 and 9 [40] In our study we found that girinimbinetreatment of A549 cells caused inhibition of XIAP but SMAC
an antagonist to XIAP could be found to be slightly regulatedin this treatment concentration This is considered as a keyevent in the execution of cell death which is regulatedmainlyby proteins of the Bcl-2 family [41 42]
Critical issues in apoptosis include the dominance ofanti-versus proapoptotic Bcl-2 members [43] Girinimbine isfound to induce apoptosis in the A549 cells involvingdownregulated Bcl-2 expression This downregulation was
10 Evidence-Based Complementary and Alternative Medicine
3
2
1
0
times105
19 9 475 0 19 9 475 0 19 9 475 0Girinimbine treatment (120583M)
Lum
ines
cenc
e
Caspase 37Caspase 8Caspase 9
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast
Figure 8 Effect of girinimbine on activation of caspases 37 8 and 9Cells were treated with girinimbine at the indicated concentrationsfor 24 h and activity was determined by using a luminescentenzyme assay kit Each point represents the mean plusmn SD of threedifferent experiments Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software(version 40 GraphPad Software Inc San Diego CA) Statisticalsignificance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
concomitant with the upregulation of Bad Bax and Bimwhich inhibits antiapoptotic proteins and render the cellsmore susceptible to apoptogenic stimuli (Figure 7) Therelease of cytochrome c found in our study could be due tothe up regulated Bax since Bax in association with adeninenucleotide translocator within the permeability transitionpore complex will increase the mitochondrial membranepermeability and thereby discharge a number of apopto-genic molecules into the cytosol [44] In addition to thisupregulation of the tumor suppressor protein p53 which hasa critical role in regulation of the Bcl-2 and p27 and p21draws attention to the involvement of cell cycle arrest in theapoptosis process Alternatively we found the involvement ofextrinsic pathway proteins such as Fas and FasL upregulationand related upregulation of caspase 8 (Figure 7) Moreoverthe luminescent based caspase 8 assay had showed 10 folddifferences than the control Apart from this we foundthat there is a downregulation of Bid happening too Sincemitochondrial damage in the Fas pathway of apoptosis ismediated by the caspase 8 cleavage of Bid [45] the findingshere shows the cross talk between intrinsic and extrinsicpathways
The protein array results exhibited a significant reductionof both IGF-I and IGF-II IGF-I has been shown to exertstrongmitogenic and antiapoptotic effects in a variety of nor-mal and cancerous cells including lung cancer cell lines [46]Moreover the over expression of IGFBP3 is closely associatedwith the downregulation of IGF-I and its mitogenic activity[47] It is known that IGFBP-2 and IGFBP-1 are involved inthe regulation of cell migration apoptosis and cell growth
[48] Both are reported to be markedly overexpressed inmany tumors and tumor cell lines And it is assumed thatelevation of the IGFBP-2 production is part of a mechanismto compensate for the mitogenic and antiapoptotic effectsof tumor-derived IGFs [49] Our data clearly showed thedownregulation of IGFBP1 and IGFBP2 concomitant withupregulation of IGBP3
Our results provided a new insight into the mechanismof chemotherapeutic properties of Murraya koenigii Inaddition these study findings demonstrate that girinimbinemediates its apoptotic effects through both intrinsic andextrinsic pathway which is depended on caspase mediationMoreover the upregulation of p53 as well as the cell prolifer-ation repressor proteins p27 and p21 and the significant roleof insulinIGF-1 signaling were also identified These resultsindicate that girinimbine could be a candidate for a novelanticancer agentThereforemore in-depth in vitro and in vivostudies are currently going on in our laboratory
Conflict of Interests
The authors declare that they do not have any conflict ofinterests
Acknowledgment
The authors greatly acknowledge the financial support pro-vided by University of Malaya (Grant no HIR E0000002-20001)
References
[1] J K Field and S W Duffy ldquoLung cancer screening the wayforwardrdquo British Journal of Cancer vol 99 no 4 pp 557ndash5622008
[2] K A Yoon J H Park J Han et al ldquoA genome-wide associationstudy reveals susceptibility variants for nonsmall cell lungcancer in the Korean populationrdquo Human Molecular Geneticsvol 19 no 24 Article ID ddq421 pp 4948ndash4954 2010
[3] G Cartei F Cartei A Cantone et al ldquoCisplatin-cyclophos-phamide-mitomycin combination chemotherapy with support-ive care versus supportive care alone for treatment of metastaticnon- small-cell lung cancerrdquo Journal of the National CancerInstitute vol 85 no 10 pp 794ndash800 1993
[4] M O Hengartner ldquoThe biochemistry of apoptosisrdquoNature vol407 no 6805 pp 770ndash776 2000
[5] DHanahan andRAWeinberg ldquoThehallmarks of cancerrdquoCellvol 100 no 1 pp 57ndash70 2000
[6] J R Molina P Yang S D Cassivi S E Schild and A AAdjei ldquoNon-small cell lung cancer epidemiology risk factorstreatment and survivorshiprdquo Mayo Clinic Proceedings vol 83no 5 pp 584ndash594 2008
[7] S Diwanay D Chitre and B Patwardhan ldquoImmunoprotec-tion by botanical drugs in cancer chemotherapyrdquo Journal ofEthnopharmacology vol 90 no 1 pp 49ndash55 2004
[8] U M Thatte and S A Dahanukar ldquoRasayana Concept cluesfrom immunomodulatory therapyrdquo in An Annotated Bibliogra-phy of Indian Medicine S N Upadhyay Ed pp 141ndash148 1997
[9] A S Shah A S Wakade and A R Juvekar ldquoImmunomod-ulatory activity of methanolic extract of Murraya koenigii (L)
Evidence-Based Complementary and Alternative Medicine 11
Spreng leavesrdquo Indian Journal of Experimental Biology vol 46no 7 pp 505ndash509 2008
[10] R B Goswamia P K S Singha N Goswamib P Thomasd PU Devic and A K Pathakd ldquoStudies on antigenotoxic effect ofMurraya koenigii leavesrdquo International Journal of PharmaRecentResearch vol 2 pp 65ndash68 2010
[11] R S Ramsewak M G Nair G M Strasburg D L DeWittand J L Nitiss ldquoBiologically active carbazole alkaloids fromMurraya koenigiirdquo Journal of Agricultural and Food Chemistryvol 47 no 2 pp 444ndash447 1999
[12] D P Chakraborty B K Barman and P K Bose ldquoStructureof girinimbine a pyranocarbazole derivative isolated fromMurraya koenigii Sprengrdquo Scientific Culture vol 30 pp 445ndash448 1964
[13] L J M Rao K Ramalakshmi B B Borse and B RaghavanldquoAntioxidant and radical-scavenging carbazole alkaloids fromthe oleoresin of curry leaf (Murraya koenigii Spreng)rdquo FoodChemistry vol 100 no 2 pp 742ndash747 2007
[14] S L Wang B Cai C B Cui S Y Yan and C F Wu ldquoInductionof apoptosis by girinimbine in K562 cellrdquo Chinese Traditionaland Herbal Drugs vol 38 no 11 pp 1677ndash1681 2007
[15] S Syam A B Abdul M A Sukari S Mohan S I Abdelwahaband T S Wah ldquoThe growth suppressing effects of girinimbineon Hepg2 involve induction of apoptosis and cell cycle arrestrdquoMolecules vol 16 pp 7155ndash7170 2011
[16] A Bakar N Haslizawati M A Sukari et al ldquoChemical con-stituents from stem barks and roots of Murraya koenigii(Rutaceae)rdquo The Malaysian Journal of Analytical Sciences vol11 pp 173ndash176 2007
[17] S I A Wahab A B Abdul S M Mohan A S Al-ZubairiM M Elhassan and M Y Ibrahim ldquoBiological activities ofPereskia bleo extractsrdquo International Journal of Pharmacologyvol 5 no 1 pp 71ndash75 2009
[18] E Bossy-Wetzel and D R Green ldquoDetection of apoptosis byannexin V labelingrdquo Methods in Enzymology vol 322 pp 15ndash18 2000
[19] H Wang and J A Joseph ldquoQuantifying cellular oxidative stressby dichlorofluorescein assay usingmicroplate readerrdquo Free Rad-ical Biology and Medicine vol 27 no 5-6 pp 612ndash616 1999
[20] S C Cheah D R Appleton S T Lee M L Lam A H AHadi and M R Mustafa ldquoPanduratin a inhibits the growth ofA549 cells through induction of apoptosis and inhibition of NF-KappaB translocationrdquoMolecules vol 16 no 3 pp 2583ndash25982011
[21] L Foghsgaard DWissing DMauch et al ldquoCathepsin B acts asa dominant execution protease in tumor cell apoptosis inducedby tumor necrosis factorrdquo Journal of Cell Biology vol 153 no 5pp 999ndash1009 2001
[22] K Kagedal U Johansson and K Ollinger ldquoThe lysosomalprotease cathepsin D mediates apoptosis induced by oxidativestressrdquoThe FASEB journal vol 15 no 9 pp 1592ndash1594 2001
[23] M A Birch-Machin and H Swalwell ldquoHow mitochondriarecord the effects of UV exposure and oxidative stress usinghuman skin as a model tissuerdquo Mutagenesis vol 25 no 2 pp101ndash107 2010
[24] S Meister B Frey V R Lang et al ldquoCalcium channel blockerverapamil enhances endoplasmic reticulum stress and cell deathinduced by proteasome inhibition in myeloma cellsrdquoNeoplasiavol 12 no 7 pp 550ndash561 2010
[25] S Mohan A B Abdul S I Abdelwahab et al ldquoTyphoniumflagelliforme induces apoptosis in CEMss cells via activation
of caspase-9 PARP cleavage and cytochrome c release itsactivation coupled with G0G1 phase cell cycle arrestrdquo Journalof Ethnopharmacology vol 131 no 3 pp 592ndash600 2010
[26] S Elmore ldquoApoptosis a review of programmed cell deathrdquoToxicologic Pathology vol 35 no 4 pp 495ndash516 2007
[27] M Aberg M Johnell M Wickstrom and A Siegbahn ldquoTissueFactor FVIIa prevents the extrinsic pathway of apoptosis byregulation of the tumor suppressor Death-Associated ProteinKinase 1 (DAPK1)rdquoThrombosis Research vol 127 no 2 pp 141ndash148 2011
[28] M Nigam N Singh V Ranjan et al ldquoCentchroman mediatedapoptosis involves cross-talk between extrinsicintrinsic path-ways and oxidative regulationrdquo Life Sciences vol 87 no 23ndash26pp 750ndash758 2010
[29] O S Frankfurt and A Krishan ldquoApoptosis-based drug screen-ing and detection of selective toxicity to cancer cellsrdquo Anti-Cancer Drugs vol 14 no 7 pp 555ndash561 2003
[30] M van Engeland L J W Nieland F C S Ramaekers BSchutte and C P M Reutelingsperger ldquoAnnexin V-affinityassay a review on an apoptosis detection system based onphosphatidylserine exposurerdquo Cytometry vol 31 no 1 pp 1ndash91998
[31] G A Joanitti R B Azevedo and S M Freitas ldquoApoptosisand lysosome membrane permeabilization induction on breastcancer cells by an anticarcinogenic Bowman-Birk proteaseinhibitor from Vigna unguiculata seedsrdquo Cancer Letters vol293 no 1 pp 73ndash81 2010
[32] T Kirkegaard and M Jaattela ldquoLysosomal involvement in celldeath and cancerrdquo Biochimica et Biophysica ActamdashMolecularCell Research vol 1793 no 4 pp 746ndash754 2009
[33] A C Johansson H Appelqvist C Nilsson K Kagedal KRoberg and K Ollinger ldquoRegulation of apoptosis-associatedlysosomal membrane permeabilizationrdquo Apoptosis vol 15 no5 pp 527ndash540 2010
[34] J Wen K R You S Y Lee C H Song and D G KimldquoOxidative stress-mediated apoptosis the anticancer effect ofthe sesquiterpene lactone parthenoliderdquo Journal of BiologicalChemistry vol 277 no 41 pp 38954ndash38964 2002
[35] C Fleury B Mignotte and J L Vayssiere ldquoMitochondrialreactive oxygen species in cell death signalingrdquo Biochimie vol84 no 2-3 pp 131ndash141 2002
[36] A M Verhagen P G Ekert M Pakusch et al ldquoIdentificationof DIABLO a mammalian protein that promotes apoptosis bybinding to and antagonizing IAP proteinsrdquo Cell vol 102 no 1pp 43ndash53 2000
[37] N Ajenjo E Canonon I Sanchez-Perez et al ldquoSubcellularlocalization determines the protective effects of activated ERK2against distinct apoptogenic stimuli in myeloid leukemia cellsrdquoJournal of Biological Chemistry vol 279 no 31 pp 32813ndash328232004
[38] X Wang ldquoThe expanding role of mitochondria in apoptosisrdquoGenes and Development vol 15 no 22 pp 2922ndash2933 2001
[39] Y A Hannun ldquoApoptosis and the dilemma of cancer chemo-therapyrdquo Blood vol 89 no 6 pp 1845ndash1853 1997
[40] F L Scott J B Denault S J Riedl H Shin M Renatusand G S Salvesen ldquoXIAP inhibits caspase-3 and -7 usingtwo binding sites evolutionary conserved mechanism of IAPsrdquoEMBO Journal vol 24 no 3 pp 645ndash655 2005
[41] S Cory and J M Adams ldquoThe BCL2 family regulators of thecellular life-or-death switchrdquo Nature Reviews Cancer vol 2 no9 pp 647ndash656 2002
12 Evidence-Based Complementary and Alternative Medicine
[42] S B Bratton and GM Cohen ldquoDeath receptors leave a caspasefootprint that Smacs of XIAPrdquo Cell Death and Differentiationvol 10 no 1 pp 4ndash6 2003
[43] N Zamzami andGKroemer ldquoThemitochondrion in apoptosishow Pandorarsquos box opensrdquo Nature Reviews Molecular CellBiology vol 2 no 1 pp 67ndash71 2001
[44] I Marzo C Brenner N Zamzami et al ldquoBax and adeninenucleotide translocator cooperate in the mitochondrial controlof apoptosisrdquo Science vol 281 no 5385 pp 2027ndash2031 1998
[45] M D Esposti ldquoThe roles of BidrdquoApoptosis vol 7 no 5 pp 433ndash440 2002
[46] N E Allen A W Roddam D S Allen et al ldquoA prospectivestudy of serum insulin-like growth factor-I (IGF-I) IGF-IIIGF-binding protein-3 and breast cancer riskrdquo British Journalof Cancer vol 92 no 7 pp 1283ndash1287 2005
[47] L Buckbinder R Talbott S Velasco-Miguel et al ldquoInductionof the growth inhibitor IGF-binding protein 3 by p53rdquo Naturevol 377 no 6550 pp 646ndash649 1995
[48] E Ruoslahti ldquoFibronectin and its integrin receptors in cancerrdquoAdvances in Cancer Research vol 76 pp 18ndash20 1999
[49] B S Schutt M Langkamp U Rauschnabel M B Ranke andM W Elmlinger ldquoIntegrin-mediated action of insulin-likefactor binding protein-2 in tumor cellsrdquo Journal of MolecularEndocrinology vol 32 no 3 pp 859ndash868 2004
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Evidence-Based Complementary and Alternative Medicine
(a) (b) (c)
(d) (e) (f)
Figure 2 Effects of girinimbine on the morphological changes of A549 cells after 24 h treatment Apoptosis was monitored by phase contrast((a) control (b) treatment 9120583M and (c) treatment 19120583M) and fluorescence microscopy ((d) control (e) treatment 9120583M and (f) treatment19120583M) Arrows indicate the cell shrinkage ruffling and blebbing of cell membrane while arrowheads indicate the annexin V binding tophosphatidylserine Magnification 20X
out using the one-way ANOVA and Dunnettrsquos post-test atprobabilities of error of 5 and 1
3 Results
31 Antipoliferative Activity The sigmoidal dose responsecurves of girinimbine in the end-point assays are shown inFigure 1 Cell viability was analyzed using the MTT assaywhich measures the metabolic activity of cell In the A549cells treated with girinimbine metabolic activity decreasedfollowed by 24 h treatment meanwhile in the control platecell viability and metabolism were not affected Figure 1(c)summarizes the IC
50values from MTT Meanwhile even at
380 120583M girinimbine could not exhibit any sign of toxicity inWRL-68 a normal (data not shown) which was employed inthis study to investigate the specificity of cytotoxicity
32 Apoptotic Mode of Cell Death Data obtained fromfluorescence detection of A549 cells together with the phasecontrast microscopic images are shown (Figure 2) Apoptosiswas clearly detected by the phosphatidylserine externaliza-tion on the treated cells Phase contrast microscopic picturesshowed a clear morphological change of treated cells whichwere observed after 24 h of treatment with girinimbine A549cells were seen to have shrunk in size and there was rufflingand blebbing of cell membranes thus suggesting that the cellswere undergoing apoptosis (Figure 2(c))
33 Girinimbine Initiates Lysosomal Membrane Permeabiliza-tion Recent studies have shown that lysosomal membrane
permeabilization (LMP) is an early and perhaps initiatingevent in apoptosis triggered by ligation of death recep-tors lysosomotropic agents oxidative stresses or serumwithdrawal [21 22] To evaluate the lysosome acidificationpattern the cells were analyzed by acridine orange staininga known pH indicator While untreated A549 cells dis-played strong granular acridine orange staining cells treatedwith girinimbine presented weak lysosome staining patternand cytoplasm acidification indicating lysosomal membranepermeabilization (Figures 3(a)ndash3(c)) Girinimbine induceddose-dependent decrease of fluorescence intensity of lyso-somal staining which was well compared with chloroquinea known inhibitor of lysosome function was included as apositive control (Figure 3(d))
34 ROS Assay Numerous investigations have documentedthat oxidative stress-mediated cellular changes are fre-quently induced in cells exposed to cytotoxic drugs UV orgamma irradiation [23 24] We examined whether girin-imbine affects the cellular levels of peroxide by measur-ing the changes in the fluorescence using DCF-DA Asshown in Figure 4(a) treatment with girinimbine markedlyincreased the DCF-DA-derived fluorescence (520 nm) Thisgirinimbine-mediated increase in fluorescence was markedlyinhibited by pretreatment with antioxidant ascorbic acidThen we next asked whether ROS generation induced bygirinimbine is directly associated with the induction ofapoptosis Chromatin condensation was measured in thecells which were pretreated with ascorbic acid using 12120583MHoechst 33342 The results shown in Figure 4(b) clearly
Evidence-Based Complementary and Alternative Medicine 5
(a) (b) (c)
200
100
0
Treatment
Fluo
resc
ent i
nten
sity
()
Chlo
roqu
ine25120583
M
Con
trol24
h
Giri
nim
bine
10120583
gm
L
Giri
nim
bine
5120583
gm
L
Giri
nim
bine
25120583
gm
L
(d)
Figure 3 Effects of girinimbine onA549 cells lysosome acidic environment Cells were incubatedwith different concentrations of girinimbinefor 24 h stained with acridine orange and analyzed by ArrayScan HCS system Control group corresponds to untreated cells Acridine orangeis a metachromatic fluorochrome and a weak base that exhibits red fluorescence when highly concentrated in acidic lysosomes (a) Normallysosome acidic environment (arrow)Notice theweak lysosome staining pattern and cytoplasm acidification (arrowhead) in cells treatedwithchloroquine (b) and girinimbine (c) indicating lysosomal membrane permeabilization Magnification 20X The mean fluorescent intensityproduced by the acridine orange was quantitatively measured (Figure 3(d))
exhibited that pretreatment with ascorbic acid could notprevent the apoptosis at various time periods Moreover thedata clearly shows the rapid decrease in the nuclear area andthe increase in fragmentation upon treatment with 19 120583Mgirinimbine (Figure 4(c))
35 Multiparameter Cytotoxicity Analysis This assay enablessimultaneousmeasurement of several cell-health parametersnuclear morphology DNA content cell membrane perme-ability and cytochrome c localization and release frommito-chondria Typical cytotoxic changes are illustrated in Figures5 and 6 Girinimbine induced decreases in cell numbernuclear area intensity and plasma membrane permeabilitywere significantly higher in the treated cells (119875 lt 001) andmitochondrial membrane potential reduction was observedsignificantly at 19120583M (119875 lt 001) The release of cytochromec was observed significantly in 9120583M and 19120583M with 119875 lt005 and 119875 lt 001 respectively These effects occurredmore rapidly and followed a dose-response pattern Cytotoxiceffects were considered to occur only when the rate of changeof fluorescence was distinctly greater than for the negativecontrols
36 Effect of Girinimbine on Apoptotic Markers After girin-imbine (19120583M) exposure A549 cells were lysed and apoptoticmarkers where screened using a protein array All majormarkers which are involved in both intrinsic and extrinsicpathways were induced on treatment As shown in Figure 7girinimbine treatment significantly increased the expressionof caspase 8 suggesting the activation of the death receptorpathway In addition the major protein involved in theextrinsic pathway such as Fas and FasL also regulates thetreatment Moreover the involvement of mitochondria in thecell death was evident by the regulation of the Bcl-2 familyof proteins such as Bad Bax Bcl-2 and Bim Besides theBcl-2 family member Bid was found to be cleaved as wellsuggesting a potential cross-talk between the death receptorand the mitochondrial pathway The treatment also resultedin a reduction in the level of expression of the inhibitorof apoptosis XIAP as well as survivin P53 as well as thecell proliferation repressor proteins p27 and p21 and theheat shock proteins such as antiapoptotic HSP60 HSP70and HSP27 which are a result of oxidative stress in the cellwere also induced InsulinIGF-1 signaling related proteinexpression has been observed on treatment Downregulation
6 Evidence-Based Complementary and Alternative Medicine
800850900950
100010501100115012001250130013501400
Control asaTreatment
24h 24h 8h 8h 4h 4h
Fluo
resc
ence
inte
nsity
(520
nm)
Girinimbine(19120583M)
Ascorbic acid
minus
minus minus minus minus
+ +
+
+ +
+
+ +
+ +
+
lowastlowast
lowast
lowastlowast
(a)
750
500
250
024h 12h 6h Control
Treatment
Fluo
resc
ent i
nten
sity
units
lowastlowast
lowastlowast
Girinimbine(19120583M)
Ascorbic acid
minus
minus
+
+
+
+
+
+
(b)
lowast
lowastlowastlowast
lowastlowast
Aver
age n
ucle
ar o
bjec
t siz
e
15
125
10
Girinimbine(19120583M)
Ascorbic acid
minus
minus
+
+
+
+
+
+
24h 12h 6h ControlTreatment
(c)
Figure 4 Effects of girinimbine on A549 cells ROS generation (a) DCF-fluorescence intensity after ascorbic acid and 19 120583M of girinimbineexposure at 4 8 and 24 h Rate of apoptosis in terms of fluorescent intensity (b) and average nuclear object size (c) of nucleus of the girinimbinetreatment was pretreatedwith 100mMascorbic acid Values aremeanplusmn SD from three independent experiments Triplicates of each treatmentgroup were used in each independent experiment The statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005 Fluorescent intensityunits directly represent the amount of ROS production
of antiapoptosis (IGF-I IGF-II IGFBP1 IGFBP2) and upreg-ulation of proapoptosis proteins (IGFBP3 IGFBP4) wereobserved
37 Caspase Depended Apoptosis in Girinimbine Treated A549Cells The involvement of the caspase cascade in the girin-imbine mediated cell death was confirmed The treatmentwith 19 120583M was significant at 119875 lt 001 for caspase 37and 9 with 8- and 5-fold increase respectively Meanwhilethe caspase 8 showed 10-fold increase at the maximumtreatment concentration (19 120583M 119875 lt 001) (Figure 8)These results are in parallel with the protein array andindicate that the signaling cascade leading to apoptosis in
girinimbine-treated cells involves both intrinsic and extrinsicpathways
4 Discussion
Apoptosis is a normal physiological process that plays avital role in numerous normal functions [4] Furthermoreit is an active physiological process causing cellular self-destruction that comprises specific morphological and bio-chemical changes in the nucleus and cytoplasm [25] Theinvolvement of an energy-dependent cascade of molecularevents makes the mechanism of apoptosis very highly com-plex and sophisticated [26] Apoptosis is regulated by two
Evidence-Based Complementary and Alternative Medicine 7
Hoechst 33342 Cell permeability Cytochrome c
Con
trol
Giri
nim
bine
Mitochondrial
membrane potential
Figure 5 Representative images of A549 cells treated withmedium alone and 19 120583Mof girinimbine and stained with Hoechst for nuclear cellpermeability dye mitochondrial membrane potential dye and cytochrome c The images from each row are obtained from the same field ofthe same treatment sample A549 produced a marked reduction in mitochondrial membrane potential and marked increases in membranepermeability and cytochrome c Magnification 20X
major pathways the extrinsic and the intrinsic pathways[27] Nevertheless there is evidence suggesting that boththe pathways are linked and that molecules involved in thepathways can influence one another [28]
The current study found that girinimbine a carbazolealkaloid from the roots of M koenigii can inhibit cellproliferation selectively in a dose dependent manner inA549 cells The morphological observation was conductedto explore whether the cytotoxic effect was related withthe apoptotic process and it was found that the cell deathinduced by girinimbine exhibited a clear morphologicalsign of apoptosis as this is an important property of acandidate anticancer drug [29] Followed by 24 h treatmentcell shrinkage ruffling and blebbing of cell membranes wereobserved These morphological features were more evidentwhile we analysed one of the key features of apoptosis whichis the change in plasma membrane structure by surfaceexposure of phosphatidylserine (PS) while the membraneintegrity remains unchallenged [30] The incorporation ofannexinV in to the same treated cells showed distinctive formof binding between the externalized PS and Annexin V whilecells from test flasks showedmore evidence of apoptosis thanthose grown under control conditions (Figure 2)
Translocation of lysosomal hydrolases from the lysosomallumen to the rest of the cell followed by the impairmentin the lysosomal membrane function is known as lysoso-mal membrane permeabilization (LMP) [31] Often thesehydrolases facilitate apoptosis by inducing mitochondrialouter-membrane permeabilization and caspase activation
[32] In our results the main LMP consequence such as weaklysosome acidification pattern and cytoplasmic acidificationwas clearly evident (Figure 3) Being aweak base chloroquineacts as lysosomotropic drug that raises intralysosomal pHHence we have monitored the release of liposomal contentby using chloroquine as a positive control Moreover in theFigure 3 it is very clear that cells treated with girinimbineresented a weak lysosome acidification pattern followed byan increase in cytoplasmic acidification Bearing in mind thesignificance of acidic lysosome environment for proteins andorganelles degradation it can be pointed out that autophagymay not the main event of cell death
Destabilization of the lysosomal membrane may becaused by increased production of ROS via massive peroxi-dation of membrane lipids [33] Moreover many studies hadshowed the relation between ROS and apoptosis induced incells exposed to cytotoxic drugs [34] Mitochondrial ROS areused as active mediators in the regulation of cell death [35]In our study the generation of ROS led to the upregulation ofcytoprotective protein markers HSP 60 70 and 27 Figure 4clearly shows that girinimbine exposure leads to increase ofROS accumulation but inhibition of ROSby ascorbic acid didnot prevent girinimbine-induced apoptosis indicating thatROS generation is not critical for the induction of apoptosisby girinimbine in A549 cells Moreover the treatment withgirinimbine significantly decreases the cell number nucleararea and cell membrane permeability as shown by themultiparameter apoptosis analysis In addition the complexrole of mitochondria in A549 cell apoptosis was investigated
8 Evidence-Based Complementary and Alternative Medicine
600
500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Total nuclear intensity
Giri19120583
M
Giri95120583
M
Giri475120583
M
(a)
Cell permeability500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(b)
350
300
250
200
150
100
50
0
Mitochondrial membrane potential
Con
trol
Pacli
taxe
l68120583
M
lowastlowastlowastlowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(c)
600700800
500400300200100
0
Cytochrome c
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowastlowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(d)
Figure 6 Quantitative analysis of girinimbine mediated apoptosis parameter Changes in total nuclear intensity (a) cell permeability(b) mitochondrial membrane potential (c) and cytochrome c localization (d) were measured simultaneously in A549 cells Followingtreatment with girinimbine we saw statistically significant increase in total nuclear intensity increased cell permeability loss ofmitochondrialmembrane potential and cytochrome c release frommitochondria Each experiment was performed at least two times Results are expressedas the means plusmn SD Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software (version40 GraphPad Software Inc San Diego CA) Statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
Mitochondria have been described to play a central role in theapoptotic process due to the mitochondrial proteins ability toactivate cellular apoptotic programs directly [36] Detectionof changes in mitochondrial membrane potential (Ψm) wascarried out as it is assumed that its disruption is the onsetofmitochondrial membrane transition pores (MPTP) forma-tion The present study revealed that girinimbine may act onmitochondria causing loss of Ψm and subsequent apoptosisThis kind of Ψm reduction and generation of MPTP mustfacilitate the relocalization of apoptogenic proteins from onesubcellular compartment such as mitochondria to cytoplasmto gain access to their substrates or interacting partners [37]
The multi cytotoxicity assay shows that girinimbine ind-uced cell death could be through the classical mitochondrial
pathway with cytochrome c release and caspase-dependentapoptosis The translocation of cytochrome c a componentof themitochondrial electron transfer chain was significantlyincreased upon girinimbine treatment In the mitochondriamediated apoptosis cytochrome c in turn binds to Apaf-1 to form a complex which triggers its oligomerization toform the apoptosome The caspase-9 holoenzyme is thenforms due to the binding of procaspase-9 to the apop-tosome and sequentially cleaves and activates the down-stream caspases such as caspase-3 [38] Our findings onthe upregulation of caspase 37 caspase 9 reduction in Ψmand release of cytochrome c to the cytosol substantiate thistheory and strongly support the involvement ofmitochondriain the apoptosis induced by girinimbine in A549 cells
Evidence-Based Complementary and Alternative Medicine 9
bad
bax
bcl-2
bcl-w
BID BI
MCa
spas
e 3Ca
spas
e 8IG
F-II IG
F-I HTR
AH
SP70
HSP
60H
SP27
FasL Fa
sD
R6Cy
to c
IGFB
P-1
IGFB
P-2 IG
FBP-
3IG
FBP-
4IG
FBP-
5IG
FBP-
6IG
F-1s
RLi
vin p
21 p
27 p53
SM
AC S
urvi
vin
sTN
F-R1 XI
AP
minus3
minus2
minus1
0
1
2
3
Relat
ive d
ensit
y di
ffere
nce t
o co
ntro
l (times10
5)
(a)
(b) (c)
A B C D E F G H I J K L M N1 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 82 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 83 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II4 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II5 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R16 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R17 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg Pos8 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg PosTNF-120572
TNF-120572 TNF-120573TNF-120573
(d)
Figure 7 Cells were lysed and protein arrays were performed Cells was treated with 19120583M girinimbine for 24 h and the whole cell proteinwas extracted Equal amount of (300 120583g) of protein from each sample was used for the assay Quantitative analysis in the arrays showeddifferences in the apoptotic markers (a) Representative images of the apoptotic protein array are shown for the control (b) treatment (c) andthe exact protein name of each dot in the array (d)
Suppression of apoptosis may promote the cancer develop-ment by inhibiting the chemotherapy as well as other forms ofinducers of cell death [39] Regulation of apoptosis has beencensoriously dependent on several genes which have beenidentified earlier and includes XIAP-a member of the IAPfamily It has the capacity to inhibit the activation of caspases3 7 and 9 [40] In our study we found that girinimbinetreatment of A549 cells caused inhibition of XIAP but SMAC
an antagonist to XIAP could be found to be slightly regulatedin this treatment concentration This is considered as a keyevent in the execution of cell death which is regulatedmainlyby proteins of the Bcl-2 family [41 42]
Critical issues in apoptosis include the dominance ofanti-versus proapoptotic Bcl-2 members [43] Girinimbine isfound to induce apoptosis in the A549 cells involvingdownregulated Bcl-2 expression This downregulation was
10 Evidence-Based Complementary and Alternative Medicine
3
2
1
0
times105
19 9 475 0 19 9 475 0 19 9 475 0Girinimbine treatment (120583M)
Lum
ines
cenc
e
Caspase 37Caspase 8Caspase 9
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast
Figure 8 Effect of girinimbine on activation of caspases 37 8 and 9Cells were treated with girinimbine at the indicated concentrationsfor 24 h and activity was determined by using a luminescentenzyme assay kit Each point represents the mean plusmn SD of threedifferent experiments Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software(version 40 GraphPad Software Inc San Diego CA) Statisticalsignificance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
concomitant with the upregulation of Bad Bax and Bimwhich inhibits antiapoptotic proteins and render the cellsmore susceptible to apoptogenic stimuli (Figure 7) Therelease of cytochrome c found in our study could be due tothe up regulated Bax since Bax in association with adeninenucleotide translocator within the permeability transitionpore complex will increase the mitochondrial membranepermeability and thereby discharge a number of apopto-genic molecules into the cytosol [44] In addition to thisupregulation of the tumor suppressor protein p53 which hasa critical role in regulation of the Bcl-2 and p27 and p21draws attention to the involvement of cell cycle arrest in theapoptosis process Alternatively we found the involvement ofextrinsic pathway proteins such as Fas and FasL upregulationand related upregulation of caspase 8 (Figure 7) Moreoverthe luminescent based caspase 8 assay had showed 10 folddifferences than the control Apart from this we foundthat there is a downregulation of Bid happening too Sincemitochondrial damage in the Fas pathway of apoptosis ismediated by the caspase 8 cleavage of Bid [45] the findingshere shows the cross talk between intrinsic and extrinsicpathways
The protein array results exhibited a significant reductionof both IGF-I and IGF-II IGF-I has been shown to exertstrongmitogenic and antiapoptotic effects in a variety of nor-mal and cancerous cells including lung cancer cell lines [46]Moreover the over expression of IGFBP3 is closely associatedwith the downregulation of IGF-I and its mitogenic activity[47] It is known that IGFBP-2 and IGFBP-1 are involved inthe regulation of cell migration apoptosis and cell growth
[48] Both are reported to be markedly overexpressed inmany tumors and tumor cell lines And it is assumed thatelevation of the IGFBP-2 production is part of a mechanismto compensate for the mitogenic and antiapoptotic effectsof tumor-derived IGFs [49] Our data clearly showed thedownregulation of IGFBP1 and IGFBP2 concomitant withupregulation of IGBP3
Our results provided a new insight into the mechanismof chemotherapeutic properties of Murraya koenigii Inaddition these study findings demonstrate that girinimbinemediates its apoptotic effects through both intrinsic andextrinsic pathway which is depended on caspase mediationMoreover the upregulation of p53 as well as the cell prolifer-ation repressor proteins p27 and p21 and the significant roleof insulinIGF-1 signaling were also identified These resultsindicate that girinimbine could be a candidate for a novelanticancer agentThereforemore in-depth in vitro and in vivostudies are currently going on in our laboratory
Conflict of Interests
The authors declare that they do not have any conflict ofinterests
Acknowledgment
The authors greatly acknowledge the financial support pro-vided by University of Malaya (Grant no HIR E0000002-20001)
References
[1] J K Field and S W Duffy ldquoLung cancer screening the wayforwardrdquo British Journal of Cancer vol 99 no 4 pp 557ndash5622008
[2] K A Yoon J H Park J Han et al ldquoA genome-wide associationstudy reveals susceptibility variants for nonsmall cell lungcancer in the Korean populationrdquo Human Molecular Geneticsvol 19 no 24 Article ID ddq421 pp 4948ndash4954 2010
[3] G Cartei F Cartei A Cantone et al ldquoCisplatin-cyclophos-phamide-mitomycin combination chemotherapy with support-ive care versus supportive care alone for treatment of metastaticnon- small-cell lung cancerrdquo Journal of the National CancerInstitute vol 85 no 10 pp 794ndash800 1993
[4] M O Hengartner ldquoThe biochemistry of apoptosisrdquoNature vol407 no 6805 pp 770ndash776 2000
[5] DHanahan andRAWeinberg ldquoThehallmarks of cancerrdquoCellvol 100 no 1 pp 57ndash70 2000
[6] J R Molina P Yang S D Cassivi S E Schild and A AAdjei ldquoNon-small cell lung cancer epidemiology risk factorstreatment and survivorshiprdquo Mayo Clinic Proceedings vol 83no 5 pp 584ndash594 2008
[7] S Diwanay D Chitre and B Patwardhan ldquoImmunoprotec-tion by botanical drugs in cancer chemotherapyrdquo Journal ofEthnopharmacology vol 90 no 1 pp 49ndash55 2004
[8] U M Thatte and S A Dahanukar ldquoRasayana Concept cluesfrom immunomodulatory therapyrdquo in An Annotated Bibliogra-phy of Indian Medicine S N Upadhyay Ed pp 141ndash148 1997
[9] A S Shah A S Wakade and A R Juvekar ldquoImmunomod-ulatory activity of methanolic extract of Murraya koenigii (L)
Evidence-Based Complementary and Alternative Medicine 11
Spreng leavesrdquo Indian Journal of Experimental Biology vol 46no 7 pp 505ndash509 2008
[10] R B Goswamia P K S Singha N Goswamib P Thomasd PU Devic and A K Pathakd ldquoStudies on antigenotoxic effect ofMurraya koenigii leavesrdquo International Journal of PharmaRecentResearch vol 2 pp 65ndash68 2010
[11] R S Ramsewak M G Nair G M Strasburg D L DeWittand J L Nitiss ldquoBiologically active carbazole alkaloids fromMurraya koenigiirdquo Journal of Agricultural and Food Chemistryvol 47 no 2 pp 444ndash447 1999
[12] D P Chakraborty B K Barman and P K Bose ldquoStructureof girinimbine a pyranocarbazole derivative isolated fromMurraya koenigii Sprengrdquo Scientific Culture vol 30 pp 445ndash448 1964
[13] L J M Rao K Ramalakshmi B B Borse and B RaghavanldquoAntioxidant and radical-scavenging carbazole alkaloids fromthe oleoresin of curry leaf (Murraya koenigii Spreng)rdquo FoodChemistry vol 100 no 2 pp 742ndash747 2007
[14] S L Wang B Cai C B Cui S Y Yan and C F Wu ldquoInductionof apoptosis by girinimbine in K562 cellrdquo Chinese Traditionaland Herbal Drugs vol 38 no 11 pp 1677ndash1681 2007
[15] S Syam A B Abdul M A Sukari S Mohan S I Abdelwahaband T S Wah ldquoThe growth suppressing effects of girinimbineon Hepg2 involve induction of apoptosis and cell cycle arrestrdquoMolecules vol 16 pp 7155ndash7170 2011
[16] A Bakar N Haslizawati M A Sukari et al ldquoChemical con-stituents from stem barks and roots of Murraya koenigii(Rutaceae)rdquo The Malaysian Journal of Analytical Sciences vol11 pp 173ndash176 2007
[17] S I A Wahab A B Abdul S M Mohan A S Al-ZubairiM M Elhassan and M Y Ibrahim ldquoBiological activities ofPereskia bleo extractsrdquo International Journal of Pharmacologyvol 5 no 1 pp 71ndash75 2009
[18] E Bossy-Wetzel and D R Green ldquoDetection of apoptosis byannexin V labelingrdquo Methods in Enzymology vol 322 pp 15ndash18 2000
[19] H Wang and J A Joseph ldquoQuantifying cellular oxidative stressby dichlorofluorescein assay usingmicroplate readerrdquo Free Rad-ical Biology and Medicine vol 27 no 5-6 pp 612ndash616 1999
[20] S C Cheah D R Appleton S T Lee M L Lam A H AHadi and M R Mustafa ldquoPanduratin a inhibits the growth ofA549 cells through induction of apoptosis and inhibition of NF-KappaB translocationrdquoMolecules vol 16 no 3 pp 2583ndash25982011
[21] L Foghsgaard DWissing DMauch et al ldquoCathepsin B acts asa dominant execution protease in tumor cell apoptosis inducedby tumor necrosis factorrdquo Journal of Cell Biology vol 153 no 5pp 999ndash1009 2001
[22] K Kagedal U Johansson and K Ollinger ldquoThe lysosomalprotease cathepsin D mediates apoptosis induced by oxidativestressrdquoThe FASEB journal vol 15 no 9 pp 1592ndash1594 2001
[23] M A Birch-Machin and H Swalwell ldquoHow mitochondriarecord the effects of UV exposure and oxidative stress usinghuman skin as a model tissuerdquo Mutagenesis vol 25 no 2 pp101ndash107 2010
[24] S Meister B Frey V R Lang et al ldquoCalcium channel blockerverapamil enhances endoplasmic reticulum stress and cell deathinduced by proteasome inhibition in myeloma cellsrdquoNeoplasiavol 12 no 7 pp 550ndash561 2010
[25] S Mohan A B Abdul S I Abdelwahab et al ldquoTyphoniumflagelliforme induces apoptosis in CEMss cells via activation
of caspase-9 PARP cleavage and cytochrome c release itsactivation coupled with G0G1 phase cell cycle arrestrdquo Journalof Ethnopharmacology vol 131 no 3 pp 592ndash600 2010
[26] S Elmore ldquoApoptosis a review of programmed cell deathrdquoToxicologic Pathology vol 35 no 4 pp 495ndash516 2007
[27] M Aberg M Johnell M Wickstrom and A Siegbahn ldquoTissueFactor FVIIa prevents the extrinsic pathway of apoptosis byregulation of the tumor suppressor Death-Associated ProteinKinase 1 (DAPK1)rdquoThrombosis Research vol 127 no 2 pp 141ndash148 2011
[28] M Nigam N Singh V Ranjan et al ldquoCentchroman mediatedapoptosis involves cross-talk between extrinsicintrinsic path-ways and oxidative regulationrdquo Life Sciences vol 87 no 23ndash26pp 750ndash758 2010
[29] O S Frankfurt and A Krishan ldquoApoptosis-based drug screen-ing and detection of selective toxicity to cancer cellsrdquo Anti-Cancer Drugs vol 14 no 7 pp 555ndash561 2003
[30] M van Engeland L J W Nieland F C S Ramaekers BSchutte and C P M Reutelingsperger ldquoAnnexin V-affinityassay a review on an apoptosis detection system based onphosphatidylserine exposurerdquo Cytometry vol 31 no 1 pp 1ndash91998
[31] G A Joanitti R B Azevedo and S M Freitas ldquoApoptosisand lysosome membrane permeabilization induction on breastcancer cells by an anticarcinogenic Bowman-Birk proteaseinhibitor from Vigna unguiculata seedsrdquo Cancer Letters vol293 no 1 pp 73ndash81 2010
[32] T Kirkegaard and M Jaattela ldquoLysosomal involvement in celldeath and cancerrdquo Biochimica et Biophysica ActamdashMolecularCell Research vol 1793 no 4 pp 746ndash754 2009
[33] A C Johansson H Appelqvist C Nilsson K Kagedal KRoberg and K Ollinger ldquoRegulation of apoptosis-associatedlysosomal membrane permeabilizationrdquo Apoptosis vol 15 no5 pp 527ndash540 2010
[34] J Wen K R You S Y Lee C H Song and D G KimldquoOxidative stress-mediated apoptosis the anticancer effect ofthe sesquiterpene lactone parthenoliderdquo Journal of BiologicalChemistry vol 277 no 41 pp 38954ndash38964 2002
[35] C Fleury B Mignotte and J L Vayssiere ldquoMitochondrialreactive oxygen species in cell death signalingrdquo Biochimie vol84 no 2-3 pp 131ndash141 2002
[36] A M Verhagen P G Ekert M Pakusch et al ldquoIdentificationof DIABLO a mammalian protein that promotes apoptosis bybinding to and antagonizing IAP proteinsrdquo Cell vol 102 no 1pp 43ndash53 2000
[37] N Ajenjo E Canonon I Sanchez-Perez et al ldquoSubcellularlocalization determines the protective effects of activated ERK2against distinct apoptogenic stimuli in myeloid leukemia cellsrdquoJournal of Biological Chemistry vol 279 no 31 pp 32813ndash328232004
[38] X Wang ldquoThe expanding role of mitochondria in apoptosisrdquoGenes and Development vol 15 no 22 pp 2922ndash2933 2001
[39] Y A Hannun ldquoApoptosis and the dilemma of cancer chemo-therapyrdquo Blood vol 89 no 6 pp 1845ndash1853 1997
[40] F L Scott J B Denault S J Riedl H Shin M Renatusand G S Salvesen ldquoXIAP inhibits caspase-3 and -7 usingtwo binding sites evolutionary conserved mechanism of IAPsrdquoEMBO Journal vol 24 no 3 pp 645ndash655 2005
[41] S Cory and J M Adams ldquoThe BCL2 family regulators of thecellular life-or-death switchrdquo Nature Reviews Cancer vol 2 no9 pp 647ndash656 2002
12 Evidence-Based Complementary and Alternative Medicine
[42] S B Bratton and GM Cohen ldquoDeath receptors leave a caspasefootprint that Smacs of XIAPrdquo Cell Death and Differentiationvol 10 no 1 pp 4ndash6 2003
[43] N Zamzami andGKroemer ldquoThemitochondrion in apoptosishow Pandorarsquos box opensrdquo Nature Reviews Molecular CellBiology vol 2 no 1 pp 67ndash71 2001
[44] I Marzo C Brenner N Zamzami et al ldquoBax and adeninenucleotide translocator cooperate in the mitochondrial controlof apoptosisrdquo Science vol 281 no 5385 pp 2027ndash2031 1998
[45] M D Esposti ldquoThe roles of BidrdquoApoptosis vol 7 no 5 pp 433ndash440 2002
[46] N E Allen A W Roddam D S Allen et al ldquoA prospectivestudy of serum insulin-like growth factor-I (IGF-I) IGF-IIIGF-binding protein-3 and breast cancer riskrdquo British Journalof Cancer vol 92 no 7 pp 1283ndash1287 2005
[47] L Buckbinder R Talbott S Velasco-Miguel et al ldquoInductionof the growth inhibitor IGF-binding protein 3 by p53rdquo Naturevol 377 no 6550 pp 646ndash649 1995
[48] E Ruoslahti ldquoFibronectin and its integrin receptors in cancerrdquoAdvances in Cancer Research vol 76 pp 18ndash20 1999
[49] B S Schutt M Langkamp U Rauschnabel M B Ranke andM W Elmlinger ldquoIntegrin-mediated action of insulin-likefactor binding protein-2 in tumor cellsrdquo Journal of MolecularEndocrinology vol 32 no 3 pp 859ndash868 2004
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 5
(a) (b) (c)
200
100
0
Treatment
Fluo
resc
ent i
nten
sity
()
Chlo
roqu
ine25120583
M
Con
trol24
h
Giri
nim
bine
10120583
gm
L
Giri
nim
bine
5120583
gm
L
Giri
nim
bine
25120583
gm
L
(d)
Figure 3 Effects of girinimbine onA549 cells lysosome acidic environment Cells were incubatedwith different concentrations of girinimbinefor 24 h stained with acridine orange and analyzed by ArrayScan HCS system Control group corresponds to untreated cells Acridine orangeis a metachromatic fluorochrome and a weak base that exhibits red fluorescence when highly concentrated in acidic lysosomes (a) Normallysosome acidic environment (arrow)Notice theweak lysosome staining pattern and cytoplasm acidification (arrowhead) in cells treatedwithchloroquine (b) and girinimbine (c) indicating lysosomal membrane permeabilization Magnification 20X The mean fluorescent intensityproduced by the acridine orange was quantitatively measured (Figure 3(d))
exhibited that pretreatment with ascorbic acid could notprevent the apoptosis at various time periods Moreover thedata clearly shows the rapid decrease in the nuclear area andthe increase in fragmentation upon treatment with 19 120583Mgirinimbine (Figure 4(c))
35 Multiparameter Cytotoxicity Analysis This assay enablessimultaneousmeasurement of several cell-health parametersnuclear morphology DNA content cell membrane perme-ability and cytochrome c localization and release frommito-chondria Typical cytotoxic changes are illustrated in Figures5 and 6 Girinimbine induced decreases in cell numbernuclear area intensity and plasma membrane permeabilitywere significantly higher in the treated cells (119875 lt 001) andmitochondrial membrane potential reduction was observedsignificantly at 19120583M (119875 lt 001) The release of cytochromec was observed significantly in 9120583M and 19120583M with 119875 lt005 and 119875 lt 001 respectively These effects occurredmore rapidly and followed a dose-response pattern Cytotoxiceffects were considered to occur only when the rate of changeof fluorescence was distinctly greater than for the negativecontrols
36 Effect of Girinimbine on Apoptotic Markers After girin-imbine (19120583M) exposure A549 cells were lysed and apoptoticmarkers where screened using a protein array All majormarkers which are involved in both intrinsic and extrinsicpathways were induced on treatment As shown in Figure 7girinimbine treatment significantly increased the expressionof caspase 8 suggesting the activation of the death receptorpathway In addition the major protein involved in theextrinsic pathway such as Fas and FasL also regulates thetreatment Moreover the involvement of mitochondria in thecell death was evident by the regulation of the Bcl-2 familyof proteins such as Bad Bax Bcl-2 and Bim Besides theBcl-2 family member Bid was found to be cleaved as wellsuggesting a potential cross-talk between the death receptorand the mitochondrial pathway The treatment also resultedin a reduction in the level of expression of the inhibitorof apoptosis XIAP as well as survivin P53 as well as thecell proliferation repressor proteins p27 and p21 and theheat shock proteins such as antiapoptotic HSP60 HSP70and HSP27 which are a result of oxidative stress in the cellwere also induced InsulinIGF-1 signaling related proteinexpression has been observed on treatment Downregulation
6 Evidence-Based Complementary and Alternative Medicine
800850900950
100010501100115012001250130013501400
Control asaTreatment
24h 24h 8h 8h 4h 4h
Fluo
resc
ence
inte
nsity
(520
nm)
Girinimbine(19120583M)
Ascorbic acid
minus
minus minus minus minus
+ +
+
+ +
+
+ +
+ +
+
lowastlowast
lowast
lowastlowast
(a)
750
500
250
024h 12h 6h Control
Treatment
Fluo
resc
ent i
nten
sity
units
lowastlowast
lowastlowast
Girinimbine(19120583M)
Ascorbic acid
minus
minus
+
+
+
+
+
+
(b)
lowast
lowastlowastlowast
lowastlowast
Aver
age n
ucle
ar o
bjec
t siz
e
15
125
10
Girinimbine(19120583M)
Ascorbic acid
minus
minus
+
+
+
+
+
+
24h 12h 6h ControlTreatment
(c)
Figure 4 Effects of girinimbine on A549 cells ROS generation (a) DCF-fluorescence intensity after ascorbic acid and 19 120583M of girinimbineexposure at 4 8 and 24 h Rate of apoptosis in terms of fluorescent intensity (b) and average nuclear object size (c) of nucleus of the girinimbinetreatment was pretreatedwith 100mMascorbic acid Values aremeanplusmn SD from three independent experiments Triplicates of each treatmentgroup were used in each independent experiment The statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005 Fluorescent intensityunits directly represent the amount of ROS production
of antiapoptosis (IGF-I IGF-II IGFBP1 IGFBP2) and upreg-ulation of proapoptosis proteins (IGFBP3 IGFBP4) wereobserved
37 Caspase Depended Apoptosis in Girinimbine Treated A549Cells The involvement of the caspase cascade in the girin-imbine mediated cell death was confirmed The treatmentwith 19 120583M was significant at 119875 lt 001 for caspase 37and 9 with 8- and 5-fold increase respectively Meanwhilethe caspase 8 showed 10-fold increase at the maximumtreatment concentration (19 120583M 119875 lt 001) (Figure 8)These results are in parallel with the protein array andindicate that the signaling cascade leading to apoptosis in
girinimbine-treated cells involves both intrinsic and extrinsicpathways
4 Discussion
Apoptosis is a normal physiological process that plays avital role in numerous normal functions [4] Furthermoreit is an active physiological process causing cellular self-destruction that comprises specific morphological and bio-chemical changes in the nucleus and cytoplasm [25] Theinvolvement of an energy-dependent cascade of molecularevents makes the mechanism of apoptosis very highly com-plex and sophisticated [26] Apoptosis is regulated by two
Evidence-Based Complementary and Alternative Medicine 7
Hoechst 33342 Cell permeability Cytochrome c
Con
trol
Giri
nim
bine
Mitochondrial
membrane potential
Figure 5 Representative images of A549 cells treated withmedium alone and 19 120583Mof girinimbine and stained with Hoechst for nuclear cellpermeability dye mitochondrial membrane potential dye and cytochrome c The images from each row are obtained from the same field ofthe same treatment sample A549 produced a marked reduction in mitochondrial membrane potential and marked increases in membranepermeability and cytochrome c Magnification 20X
major pathways the extrinsic and the intrinsic pathways[27] Nevertheless there is evidence suggesting that boththe pathways are linked and that molecules involved in thepathways can influence one another [28]
The current study found that girinimbine a carbazolealkaloid from the roots of M koenigii can inhibit cellproliferation selectively in a dose dependent manner inA549 cells The morphological observation was conductedto explore whether the cytotoxic effect was related withthe apoptotic process and it was found that the cell deathinduced by girinimbine exhibited a clear morphologicalsign of apoptosis as this is an important property of acandidate anticancer drug [29] Followed by 24 h treatmentcell shrinkage ruffling and blebbing of cell membranes wereobserved These morphological features were more evidentwhile we analysed one of the key features of apoptosis whichis the change in plasma membrane structure by surfaceexposure of phosphatidylserine (PS) while the membraneintegrity remains unchallenged [30] The incorporation ofannexinV in to the same treated cells showed distinctive formof binding between the externalized PS and Annexin V whilecells from test flasks showedmore evidence of apoptosis thanthose grown under control conditions (Figure 2)
Translocation of lysosomal hydrolases from the lysosomallumen to the rest of the cell followed by the impairmentin the lysosomal membrane function is known as lysoso-mal membrane permeabilization (LMP) [31] Often thesehydrolases facilitate apoptosis by inducing mitochondrialouter-membrane permeabilization and caspase activation
[32] In our results the main LMP consequence such as weaklysosome acidification pattern and cytoplasmic acidificationwas clearly evident (Figure 3) Being aweak base chloroquineacts as lysosomotropic drug that raises intralysosomal pHHence we have monitored the release of liposomal contentby using chloroquine as a positive control Moreover in theFigure 3 it is very clear that cells treated with girinimbineresented a weak lysosome acidification pattern followed byan increase in cytoplasmic acidification Bearing in mind thesignificance of acidic lysosome environment for proteins andorganelles degradation it can be pointed out that autophagymay not the main event of cell death
Destabilization of the lysosomal membrane may becaused by increased production of ROS via massive peroxi-dation of membrane lipids [33] Moreover many studies hadshowed the relation between ROS and apoptosis induced incells exposed to cytotoxic drugs [34] Mitochondrial ROS areused as active mediators in the regulation of cell death [35]In our study the generation of ROS led to the upregulation ofcytoprotective protein markers HSP 60 70 and 27 Figure 4clearly shows that girinimbine exposure leads to increase ofROS accumulation but inhibition of ROSby ascorbic acid didnot prevent girinimbine-induced apoptosis indicating thatROS generation is not critical for the induction of apoptosisby girinimbine in A549 cells Moreover the treatment withgirinimbine significantly decreases the cell number nucleararea and cell membrane permeability as shown by themultiparameter apoptosis analysis In addition the complexrole of mitochondria in A549 cell apoptosis was investigated
8 Evidence-Based Complementary and Alternative Medicine
600
500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Total nuclear intensity
Giri19120583
M
Giri95120583
M
Giri475120583
M
(a)
Cell permeability500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(b)
350
300
250
200
150
100
50
0
Mitochondrial membrane potential
Con
trol
Pacli
taxe
l68120583
M
lowastlowastlowastlowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(c)
600700800
500400300200100
0
Cytochrome c
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowastlowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(d)
Figure 6 Quantitative analysis of girinimbine mediated apoptosis parameter Changes in total nuclear intensity (a) cell permeability(b) mitochondrial membrane potential (c) and cytochrome c localization (d) were measured simultaneously in A549 cells Followingtreatment with girinimbine we saw statistically significant increase in total nuclear intensity increased cell permeability loss ofmitochondrialmembrane potential and cytochrome c release frommitochondria Each experiment was performed at least two times Results are expressedas the means plusmn SD Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software (version40 GraphPad Software Inc San Diego CA) Statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
Mitochondria have been described to play a central role in theapoptotic process due to the mitochondrial proteins ability toactivate cellular apoptotic programs directly [36] Detectionof changes in mitochondrial membrane potential (Ψm) wascarried out as it is assumed that its disruption is the onsetofmitochondrial membrane transition pores (MPTP) forma-tion The present study revealed that girinimbine may act onmitochondria causing loss of Ψm and subsequent apoptosisThis kind of Ψm reduction and generation of MPTP mustfacilitate the relocalization of apoptogenic proteins from onesubcellular compartment such as mitochondria to cytoplasmto gain access to their substrates or interacting partners [37]
The multi cytotoxicity assay shows that girinimbine ind-uced cell death could be through the classical mitochondrial
pathway with cytochrome c release and caspase-dependentapoptosis The translocation of cytochrome c a componentof themitochondrial electron transfer chain was significantlyincreased upon girinimbine treatment In the mitochondriamediated apoptosis cytochrome c in turn binds to Apaf-1 to form a complex which triggers its oligomerization toform the apoptosome The caspase-9 holoenzyme is thenforms due to the binding of procaspase-9 to the apop-tosome and sequentially cleaves and activates the down-stream caspases such as caspase-3 [38] Our findings onthe upregulation of caspase 37 caspase 9 reduction in Ψmand release of cytochrome c to the cytosol substantiate thistheory and strongly support the involvement ofmitochondriain the apoptosis induced by girinimbine in A549 cells
Evidence-Based Complementary and Alternative Medicine 9
bad
bax
bcl-2
bcl-w
BID BI
MCa
spas
e 3Ca
spas
e 8IG
F-II IG
F-I HTR
AH
SP70
HSP
60H
SP27
FasL Fa
sD
R6Cy
to c
IGFB
P-1
IGFB
P-2 IG
FBP-
3IG
FBP-
4IG
FBP-
5IG
FBP-
6IG
F-1s
RLi
vin p
21 p
27 p53
SM
AC S
urvi
vin
sTN
F-R1 XI
AP
minus3
minus2
minus1
0
1
2
3
Relat
ive d
ensit
y di
ffere
nce t
o co
ntro
l (times10
5)
(a)
(b) (c)
A B C D E F G H I J K L M N1 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 82 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 83 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II4 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II5 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R16 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R17 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg Pos8 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg PosTNF-120572
TNF-120572 TNF-120573TNF-120573
(d)
Figure 7 Cells were lysed and protein arrays were performed Cells was treated with 19120583M girinimbine for 24 h and the whole cell proteinwas extracted Equal amount of (300 120583g) of protein from each sample was used for the assay Quantitative analysis in the arrays showeddifferences in the apoptotic markers (a) Representative images of the apoptotic protein array are shown for the control (b) treatment (c) andthe exact protein name of each dot in the array (d)
Suppression of apoptosis may promote the cancer develop-ment by inhibiting the chemotherapy as well as other forms ofinducers of cell death [39] Regulation of apoptosis has beencensoriously dependent on several genes which have beenidentified earlier and includes XIAP-a member of the IAPfamily It has the capacity to inhibit the activation of caspases3 7 and 9 [40] In our study we found that girinimbinetreatment of A549 cells caused inhibition of XIAP but SMAC
an antagonist to XIAP could be found to be slightly regulatedin this treatment concentration This is considered as a keyevent in the execution of cell death which is regulatedmainlyby proteins of the Bcl-2 family [41 42]
Critical issues in apoptosis include the dominance ofanti-versus proapoptotic Bcl-2 members [43] Girinimbine isfound to induce apoptosis in the A549 cells involvingdownregulated Bcl-2 expression This downregulation was
10 Evidence-Based Complementary and Alternative Medicine
3
2
1
0
times105
19 9 475 0 19 9 475 0 19 9 475 0Girinimbine treatment (120583M)
Lum
ines
cenc
e
Caspase 37Caspase 8Caspase 9
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast
Figure 8 Effect of girinimbine on activation of caspases 37 8 and 9Cells were treated with girinimbine at the indicated concentrationsfor 24 h and activity was determined by using a luminescentenzyme assay kit Each point represents the mean plusmn SD of threedifferent experiments Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software(version 40 GraphPad Software Inc San Diego CA) Statisticalsignificance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
concomitant with the upregulation of Bad Bax and Bimwhich inhibits antiapoptotic proteins and render the cellsmore susceptible to apoptogenic stimuli (Figure 7) Therelease of cytochrome c found in our study could be due tothe up regulated Bax since Bax in association with adeninenucleotide translocator within the permeability transitionpore complex will increase the mitochondrial membranepermeability and thereby discharge a number of apopto-genic molecules into the cytosol [44] In addition to thisupregulation of the tumor suppressor protein p53 which hasa critical role in regulation of the Bcl-2 and p27 and p21draws attention to the involvement of cell cycle arrest in theapoptosis process Alternatively we found the involvement ofextrinsic pathway proteins such as Fas and FasL upregulationand related upregulation of caspase 8 (Figure 7) Moreoverthe luminescent based caspase 8 assay had showed 10 folddifferences than the control Apart from this we foundthat there is a downregulation of Bid happening too Sincemitochondrial damage in the Fas pathway of apoptosis ismediated by the caspase 8 cleavage of Bid [45] the findingshere shows the cross talk between intrinsic and extrinsicpathways
The protein array results exhibited a significant reductionof both IGF-I and IGF-II IGF-I has been shown to exertstrongmitogenic and antiapoptotic effects in a variety of nor-mal and cancerous cells including lung cancer cell lines [46]Moreover the over expression of IGFBP3 is closely associatedwith the downregulation of IGF-I and its mitogenic activity[47] It is known that IGFBP-2 and IGFBP-1 are involved inthe regulation of cell migration apoptosis and cell growth
[48] Both are reported to be markedly overexpressed inmany tumors and tumor cell lines And it is assumed thatelevation of the IGFBP-2 production is part of a mechanismto compensate for the mitogenic and antiapoptotic effectsof tumor-derived IGFs [49] Our data clearly showed thedownregulation of IGFBP1 and IGFBP2 concomitant withupregulation of IGBP3
Our results provided a new insight into the mechanismof chemotherapeutic properties of Murraya koenigii Inaddition these study findings demonstrate that girinimbinemediates its apoptotic effects through both intrinsic andextrinsic pathway which is depended on caspase mediationMoreover the upregulation of p53 as well as the cell prolifer-ation repressor proteins p27 and p21 and the significant roleof insulinIGF-1 signaling were also identified These resultsindicate that girinimbine could be a candidate for a novelanticancer agentThereforemore in-depth in vitro and in vivostudies are currently going on in our laboratory
Conflict of Interests
The authors declare that they do not have any conflict ofinterests
Acknowledgment
The authors greatly acknowledge the financial support pro-vided by University of Malaya (Grant no HIR E0000002-20001)
References
[1] J K Field and S W Duffy ldquoLung cancer screening the wayforwardrdquo British Journal of Cancer vol 99 no 4 pp 557ndash5622008
[2] K A Yoon J H Park J Han et al ldquoA genome-wide associationstudy reveals susceptibility variants for nonsmall cell lungcancer in the Korean populationrdquo Human Molecular Geneticsvol 19 no 24 Article ID ddq421 pp 4948ndash4954 2010
[3] G Cartei F Cartei A Cantone et al ldquoCisplatin-cyclophos-phamide-mitomycin combination chemotherapy with support-ive care versus supportive care alone for treatment of metastaticnon- small-cell lung cancerrdquo Journal of the National CancerInstitute vol 85 no 10 pp 794ndash800 1993
[4] M O Hengartner ldquoThe biochemistry of apoptosisrdquoNature vol407 no 6805 pp 770ndash776 2000
[5] DHanahan andRAWeinberg ldquoThehallmarks of cancerrdquoCellvol 100 no 1 pp 57ndash70 2000
[6] J R Molina P Yang S D Cassivi S E Schild and A AAdjei ldquoNon-small cell lung cancer epidemiology risk factorstreatment and survivorshiprdquo Mayo Clinic Proceedings vol 83no 5 pp 584ndash594 2008
[7] S Diwanay D Chitre and B Patwardhan ldquoImmunoprotec-tion by botanical drugs in cancer chemotherapyrdquo Journal ofEthnopharmacology vol 90 no 1 pp 49ndash55 2004
[8] U M Thatte and S A Dahanukar ldquoRasayana Concept cluesfrom immunomodulatory therapyrdquo in An Annotated Bibliogra-phy of Indian Medicine S N Upadhyay Ed pp 141ndash148 1997
[9] A S Shah A S Wakade and A R Juvekar ldquoImmunomod-ulatory activity of methanolic extract of Murraya koenigii (L)
Evidence-Based Complementary and Alternative Medicine 11
Spreng leavesrdquo Indian Journal of Experimental Biology vol 46no 7 pp 505ndash509 2008
[10] R B Goswamia P K S Singha N Goswamib P Thomasd PU Devic and A K Pathakd ldquoStudies on antigenotoxic effect ofMurraya koenigii leavesrdquo International Journal of PharmaRecentResearch vol 2 pp 65ndash68 2010
[11] R S Ramsewak M G Nair G M Strasburg D L DeWittand J L Nitiss ldquoBiologically active carbazole alkaloids fromMurraya koenigiirdquo Journal of Agricultural and Food Chemistryvol 47 no 2 pp 444ndash447 1999
[12] D P Chakraborty B K Barman and P K Bose ldquoStructureof girinimbine a pyranocarbazole derivative isolated fromMurraya koenigii Sprengrdquo Scientific Culture vol 30 pp 445ndash448 1964
[13] L J M Rao K Ramalakshmi B B Borse and B RaghavanldquoAntioxidant and radical-scavenging carbazole alkaloids fromthe oleoresin of curry leaf (Murraya koenigii Spreng)rdquo FoodChemistry vol 100 no 2 pp 742ndash747 2007
[14] S L Wang B Cai C B Cui S Y Yan and C F Wu ldquoInductionof apoptosis by girinimbine in K562 cellrdquo Chinese Traditionaland Herbal Drugs vol 38 no 11 pp 1677ndash1681 2007
[15] S Syam A B Abdul M A Sukari S Mohan S I Abdelwahaband T S Wah ldquoThe growth suppressing effects of girinimbineon Hepg2 involve induction of apoptosis and cell cycle arrestrdquoMolecules vol 16 pp 7155ndash7170 2011
[16] A Bakar N Haslizawati M A Sukari et al ldquoChemical con-stituents from stem barks and roots of Murraya koenigii(Rutaceae)rdquo The Malaysian Journal of Analytical Sciences vol11 pp 173ndash176 2007
[17] S I A Wahab A B Abdul S M Mohan A S Al-ZubairiM M Elhassan and M Y Ibrahim ldquoBiological activities ofPereskia bleo extractsrdquo International Journal of Pharmacologyvol 5 no 1 pp 71ndash75 2009
[18] E Bossy-Wetzel and D R Green ldquoDetection of apoptosis byannexin V labelingrdquo Methods in Enzymology vol 322 pp 15ndash18 2000
[19] H Wang and J A Joseph ldquoQuantifying cellular oxidative stressby dichlorofluorescein assay usingmicroplate readerrdquo Free Rad-ical Biology and Medicine vol 27 no 5-6 pp 612ndash616 1999
[20] S C Cheah D R Appleton S T Lee M L Lam A H AHadi and M R Mustafa ldquoPanduratin a inhibits the growth ofA549 cells through induction of apoptosis and inhibition of NF-KappaB translocationrdquoMolecules vol 16 no 3 pp 2583ndash25982011
[21] L Foghsgaard DWissing DMauch et al ldquoCathepsin B acts asa dominant execution protease in tumor cell apoptosis inducedby tumor necrosis factorrdquo Journal of Cell Biology vol 153 no 5pp 999ndash1009 2001
[22] K Kagedal U Johansson and K Ollinger ldquoThe lysosomalprotease cathepsin D mediates apoptosis induced by oxidativestressrdquoThe FASEB journal vol 15 no 9 pp 1592ndash1594 2001
[23] M A Birch-Machin and H Swalwell ldquoHow mitochondriarecord the effects of UV exposure and oxidative stress usinghuman skin as a model tissuerdquo Mutagenesis vol 25 no 2 pp101ndash107 2010
[24] S Meister B Frey V R Lang et al ldquoCalcium channel blockerverapamil enhances endoplasmic reticulum stress and cell deathinduced by proteasome inhibition in myeloma cellsrdquoNeoplasiavol 12 no 7 pp 550ndash561 2010
[25] S Mohan A B Abdul S I Abdelwahab et al ldquoTyphoniumflagelliforme induces apoptosis in CEMss cells via activation
of caspase-9 PARP cleavage and cytochrome c release itsactivation coupled with G0G1 phase cell cycle arrestrdquo Journalof Ethnopharmacology vol 131 no 3 pp 592ndash600 2010
[26] S Elmore ldquoApoptosis a review of programmed cell deathrdquoToxicologic Pathology vol 35 no 4 pp 495ndash516 2007
[27] M Aberg M Johnell M Wickstrom and A Siegbahn ldquoTissueFactor FVIIa prevents the extrinsic pathway of apoptosis byregulation of the tumor suppressor Death-Associated ProteinKinase 1 (DAPK1)rdquoThrombosis Research vol 127 no 2 pp 141ndash148 2011
[28] M Nigam N Singh V Ranjan et al ldquoCentchroman mediatedapoptosis involves cross-talk between extrinsicintrinsic path-ways and oxidative regulationrdquo Life Sciences vol 87 no 23ndash26pp 750ndash758 2010
[29] O S Frankfurt and A Krishan ldquoApoptosis-based drug screen-ing and detection of selective toxicity to cancer cellsrdquo Anti-Cancer Drugs vol 14 no 7 pp 555ndash561 2003
[30] M van Engeland L J W Nieland F C S Ramaekers BSchutte and C P M Reutelingsperger ldquoAnnexin V-affinityassay a review on an apoptosis detection system based onphosphatidylserine exposurerdquo Cytometry vol 31 no 1 pp 1ndash91998
[31] G A Joanitti R B Azevedo and S M Freitas ldquoApoptosisand lysosome membrane permeabilization induction on breastcancer cells by an anticarcinogenic Bowman-Birk proteaseinhibitor from Vigna unguiculata seedsrdquo Cancer Letters vol293 no 1 pp 73ndash81 2010
[32] T Kirkegaard and M Jaattela ldquoLysosomal involvement in celldeath and cancerrdquo Biochimica et Biophysica ActamdashMolecularCell Research vol 1793 no 4 pp 746ndash754 2009
[33] A C Johansson H Appelqvist C Nilsson K Kagedal KRoberg and K Ollinger ldquoRegulation of apoptosis-associatedlysosomal membrane permeabilizationrdquo Apoptosis vol 15 no5 pp 527ndash540 2010
[34] J Wen K R You S Y Lee C H Song and D G KimldquoOxidative stress-mediated apoptosis the anticancer effect ofthe sesquiterpene lactone parthenoliderdquo Journal of BiologicalChemistry vol 277 no 41 pp 38954ndash38964 2002
[35] C Fleury B Mignotte and J L Vayssiere ldquoMitochondrialreactive oxygen species in cell death signalingrdquo Biochimie vol84 no 2-3 pp 131ndash141 2002
[36] A M Verhagen P G Ekert M Pakusch et al ldquoIdentificationof DIABLO a mammalian protein that promotes apoptosis bybinding to and antagonizing IAP proteinsrdquo Cell vol 102 no 1pp 43ndash53 2000
[37] N Ajenjo E Canonon I Sanchez-Perez et al ldquoSubcellularlocalization determines the protective effects of activated ERK2against distinct apoptogenic stimuli in myeloid leukemia cellsrdquoJournal of Biological Chemistry vol 279 no 31 pp 32813ndash328232004
[38] X Wang ldquoThe expanding role of mitochondria in apoptosisrdquoGenes and Development vol 15 no 22 pp 2922ndash2933 2001
[39] Y A Hannun ldquoApoptosis and the dilemma of cancer chemo-therapyrdquo Blood vol 89 no 6 pp 1845ndash1853 1997
[40] F L Scott J B Denault S J Riedl H Shin M Renatusand G S Salvesen ldquoXIAP inhibits caspase-3 and -7 usingtwo binding sites evolutionary conserved mechanism of IAPsrdquoEMBO Journal vol 24 no 3 pp 645ndash655 2005
[41] S Cory and J M Adams ldquoThe BCL2 family regulators of thecellular life-or-death switchrdquo Nature Reviews Cancer vol 2 no9 pp 647ndash656 2002
12 Evidence-Based Complementary and Alternative Medicine
[42] S B Bratton and GM Cohen ldquoDeath receptors leave a caspasefootprint that Smacs of XIAPrdquo Cell Death and Differentiationvol 10 no 1 pp 4ndash6 2003
[43] N Zamzami andGKroemer ldquoThemitochondrion in apoptosishow Pandorarsquos box opensrdquo Nature Reviews Molecular CellBiology vol 2 no 1 pp 67ndash71 2001
[44] I Marzo C Brenner N Zamzami et al ldquoBax and adeninenucleotide translocator cooperate in the mitochondrial controlof apoptosisrdquo Science vol 281 no 5385 pp 2027ndash2031 1998
[45] M D Esposti ldquoThe roles of BidrdquoApoptosis vol 7 no 5 pp 433ndash440 2002
[46] N E Allen A W Roddam D S Allen et al ldquoA prospectivestudy of serum insulin-like growth factor-I (IGF-I) IGF-IIIGF-binding protein-3 and breast cancer riskrdquo British Journalof Cancer vol 92 no 7 pp 1283ndash1287 2005
[47] L Buckbinder R Talbott S Velasco-Miguel et al ldquoInductionof the growth inhibitor IGF-binding protein 3 by p53rdquo Naturevol 377 no 6550 pp 646ndash649 1995
[48] E Ruoslahti ldquoFibronectin and its integrin receptors in cancerrdquoAdvances in Cancer Research vol 76 pp 18ndash20 1999
[49] B S Schutt M Langkamp U Rauschnabel M B Ranke andM W Elmlinger ldquoIntegrin-mediated action of insulin-likefactor binding protein-2 in tumor cellsrdquo Journal of MolecularEndocrinology vol 32 no 3 pp 859ndash868 2004
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Evidence-Based Complementary and Alternative Medicine
800850900950
100010501100115012001250130013501400
Control asaTreatment
24h 24h 8h 8h 4h 4h
Fluo
resc
ence
inte
nsity
(520
nm)
Girinimbine(19120583M)
Ascorbic acid
minus
minus minus minus minus
+ +
+
+ +
+
+ +
+ +
+
lowastlowast
lowast
lowastlowast
(a)
750
500
250
024h 12h 6h Control
Treatment
Fluo
resc
ent i
nten
sity
units
lowastlowast
lowastlowast
Girinimbine(19120583M)
Ascorbic acid
minus
minus
+
+
+
+
+
+
(b)
lowast
lowastlowastlowast
lowastlowast
Aver
age n
ucle
ar o
bjec
t siz
e
15
125
10
Girinimbine(19120583M)
Ascorbic acid
minus
minus
+
+
+
+
+
+
24h 12h 6h ControlTreatment
(c)
Figure 4 Effects of girinimbine on A549 cells ROS generation (a) DCF-fluorescence intensity after ascorbic acid and 19 120583M of girinimbineexposure at 4 8 and 24 h Rate of apoptosis in terms of fluorescent intensity (b) and average nuclear object size (c) of nucleus of the girinimbinetreatment was pretreatedwith 100mMascorbic acid Values aremeanplusmn SD from three independent experiments Triplicates of each treatmentgroup were used in each independent experiment The statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005 Fluorescent intensityunits directly represent the amount of ROS production
of antiapoptosis (IGF-I IGF-II IGFBP1 IGFBP2) and upreg-ulation of proapoptosis proteins (IGFBP3 IGFBP4) wereobserved
37 Caspase Depended Apoptosis in Girinimbine Treated A549Cells The involvement of the caspase cascade in the girin-imbine mediated cell death was confirmed The treatmentwith 19 120583M was significant at 119875 lt 001 for caspase 37and 9 with 8- and 5-fold increase respectively Meanwhilethe caspase 8 showed 10-fold increase at the maximumtreatment concentration (19 120583M 119875 lt 001) (Figure 8)These results are in parallel with the protein array andindicate that the signaling cascade leading to apoptosis in
girinimbine-treated cells involves both intrinsic and extrinsicpathways
4 Discussion
Apoptosis is a normal physiological process that plays avital role in numerous normal functions [4] Furthermoreit is an active physiological process causing cellular self-destruction that comprises specific morphological and bio-chemical changes in the nucleus and cytoplasm [25] Theinvolvement of an energy-dependent cascade of molecularevents makes the mechanism of apoptosis very highly com-plex and sophisticated [26] Apoptosis is regulated by two
Evidence-Based Complementary and Alternative Medicine 7
Hoechst 33342 Cell permeability Cytochrome c
Con
trol
Giri
nim
bine
Mitochondrial
membrane potential
Figure 5 Representative images of A549 cells treated withmedium alone and 19 120583Mof girinimbine and stained with Hoechst for nuclear cellpermeability dye mitochondrial membrane potential dye and cytochrome c The images from each row are obtained from the same field ofthe same treatment sample A549 produced a marked reduction in mitochondrial membrane potential and marked increases in membranepermeability and cytochrome c Magnification 20X
major pathways the extrinsic and the intrinsic pathways[27] Nevertheless there is evidence suggesting that boththe pathways are linked and that molecules involved in thepathways can influence one another [28]
The current study found that girinimbine a carbazolealkaloid from the roots of M koenigii can inhibit cellproliferation selectively in a dose dependent manner inA549 cells The morphological observation was conductedto explore whether the cytotoxic effect was related withthe apoptotic process and it was found that the cell deathinduced by girinimbine exhibited a clear morphologicalsign of apoptosis as this is an important property of acandidate anticancer drug [29] Followed by 24 h treatmentcell shrinkage ruffling and blebbing of cell membranes wereobserved These morphological features were more evidentwhile we analysed one of the key features of apoptosis whichis the change in plasma membrane structure by surfaceexposure of phosphatidylserine (PS) while the membraneintegrity remains unchallenged [30] The incorporation ofannexinV in to the same treated cells showed distinctive formof binding between the externalized PS and Annexin V whilecells from test flasks showedmore evidence of apoptosis thanthose grown under control conditions (Figure 2)
Translocation of lysosomal hydrolases from the lysosomallumen to the rest of the cell followed by the impairmentin the lysosomal membrane function is known as lysoso-mal membrane permeabilization (LMP) [31] Often thesehydrolases facilitate apoptosis by inducing mitochondrialouter-membrane permeabilization and caspase activation
[32] In our results the main LMP consequence such as weaklysosome acidification pattern and cytoplasmic acidificationwas clearly evident (Figure 3) Being aweak base chloroquineacts as lysosomotropic drug that raises intralysosomal pHHence we have monitored the release of liposomal contentby using chloroquine as a positive control Moreover in theFigure 3 it is very clear that cells treated with girinimbineresented a weak lysosome acidification pattern followed byan increase in cytoplasmic acidification Bearing in mind thesignificance of acidic lysosome environment for proteins andorganelles degradation it can be pointed out that autophagymay not the main event of cell death
Destabilization of the lysosomal membrane may becaused by increased production of ROS via massive peroxi-dation of membrane lipids [33] Moreover many studies hadshowed the relation between ROS and apoptosis induced incells exposed to cytotoxic drugs [34] Mitochondrial ROS areused as active mediators in the regulation of cell death [35]In our study the generation of ROS led to the upregulation ofcytoprotective protein markers HSP 60 70 and 27 Figure 4clearly shows that girinimbine exposure leads to increase ofROS accumulation but inhibition of ROSby ascorbic acid didnot prevent girinimbine-induced apoptosis indicating thatROS generation is not critical for the induction of apoptosisby girinimbine in A549 cells Moreover the treatment withgirinimbine significantly decreases the cell number nucleararea and cell membrane permeability as shown by themultiparameter apoptosis analysis In addition the complexrole of mitochondria in A549 cell apoptosis was investigated
8 Evidence-Based Complementary and Alternative Medicine
600
500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Total nuclear intensity
Giri19120583
M
Giri95120583
M
Giri475120583
M
(a)
Cell permeability500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(b)
350
300
250
200
150
100
50
0
Mitochondrial membrane potential
Con
trol
Pacli
taxe
l68120583
M
lowastlowastlowastlowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(c)
600700800
500400300200100
0
Cytochrome c
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowastlowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(d)
Figure 6 Quantitative analysis of girinimbine mediated apoptosis parameter Changes in total nuclear intensity (a) cell permeability(b) mitochondrial membrane potential (c) and cytochrome c localization (d) were measured simultaneously in A549 cells Followingtreatment with girinimbine we saw statistically significant increase in total nuclear intensity increased cell permeability loss ofmitochondrialmembrane potential and cytochrome c release frommitochondria Each experiment was performed at least two times Results are expressedas the means plusmn SD Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software (version40 GraphPad Software Inc San Diego CA) Statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
Mitochondria have been described to play a central role in theapoptotic process due to the mitochondrial proteins ability toactivate cellular apoptotic programs directly [36] Detectionof changes in mitochondrial membrane potential (Ψm) wascarried out as it is assumed that its disruption is the onsetofmitochondrial membrane transition pores (MPTP) forma-tion The present study revealed that girinimbine may act onmitochondria causing loss of Ψm and subsequent apoptosisThis kind of Ψm reduction and generation of MPTP mustfacilitate the relocalization of apoptogenic proteins from onesubcellular compartment such as mitochondria to cytoplasmto gain access to their substrates or interacting partners [37]
The multi cytotoxicity assay shows that girinimbine ind-uced cell death could be through the classical mitochondrial
pathway with cytochrome c release and caspase-dependentapoptosis The translocation of cytochrome c a componentof themitochondrial electron transfer chain was significantlyincreased upon girinimbine treatment In the mitochondriamediated apoptosis cytochrome c in turn binds to Apaf-1 to form a complex which triggers its oligomerization toform the apoptosome The caspase-9 holoenzyme is thenforms due to the binding of procaspase-9 to the apop-tosome and sequentially cleaves and activates the down-stream caspases such as caspase-3 [38] Our findings onthe upregulation of caspase 37 caspase 9 reduction in Ψmand release of cytochrome c to the cytosol substantiate thistheory and strongly support the involvement ofmitochondriain the apoptosis induced by girinimbine in A549 cells
Evidence-Based Complementary and Alternative Medicine 9
bad
bax
bcl-2
bcl-w
BID BI
MCa
spas
e 3Ca
spas
e 8IG
F-II IG
F-I HTR
AH
SP70
HSP
60H
SP27
FasL Fa
sD
R6Cy
to c
IGFB
P-1
IGFB
P-2 IG
FBP-
3IG
FBP-
4IG
FBP-
5IG
FBP-
6IG
F-1s
RLi
vin p
21 p
27 p53
SM
AC S
urvi
vin
sTN
F-R1 XI
AP
minus3
minus2
minus1
0
1
2
3
Relat
ive d
ensit
y di
ffere
nce t
o co
ntro
l (times10
5)
(a)
(b) (c)
A B C D E F G H I J K L M N1 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 82 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 83 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II4 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II5 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R16 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R17 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg Pos8 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg PosTNF-120572
TNF-120572 TNF-120573TNF-120573
(d)
Figure 7 Cells were lysed and protein arrays were performed Cells was treated with 19120583M girinimbine for 24 h and the whole cell proteinwas extracted Equal amount of (300 120583g) of protein from each sample was used for the assay Quantitative analysis in the arrays showeddifferences in the apoptotic markers (a) Representative images of the apoptotic protein array are shown for the control (b) treatment (c) andthe exact protein name of each dot in the array (d)
Suppression of apoptosis may promote the cancer develop-ment by inhibiting the chemotherapy as well as other forms ofinducers of cell death [39] Regulation of apoptosis has beencensoriously dependent on several genes which have beenidentified earlier and includes XIAP-a member of the IAPfamily It has the capacity to inhibit the activation of caspases3 7 and 9 [40] In our study we found that girinimbinetreatment of A549 cells caused inhibition of XIAP but SMAC
an antagonist to XIAP could be found to be slightly regulatedin this treatment concentration This is considered as a keyevent in the execution of cell death which is regulatedmainlyby proteins of the Bcl-2 family [41 42]
Critical issues in apoptosis include the dominance ofanti-versus proapoptotic Bcl-2 members [43] Girinimbine isfound to induce apoptosis in the A549 cells involvingdownregulated Bcl-2 expression This downregulation was
10 Evidence-Based Complementary and Alternative Medicine
3
2
1
0
times105
19 9 475 0 19 9 475 0 19 9 475 0Girinimbine treatment (120583M)
Lum
ines
cenc
e
Caspase 37Caspase 8Caspase 9
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast
Figure 8 Effect of girinimbine on activation of caspases 37 8 and 9Cells were treated with girinimbine at the indicated concentrationsfor 24 h and activity was determined by using a luminescentenzyme assay kit Each point represents the mean plusmn SD of threedifferent experiments Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software(version 40 GraphPad Software Inc San Diego CA) Statisticalsignificance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
concomitant with the upregulation of Bad Bax and Bimwhich inhibits antiapoptotic proteins and render the cellsmore susceptible to apoptogenic stimuli (Figure 7) Therelease of cytochrome c found in our study could be due tothe up regulated Bax since Bax in association with adeninenucleotide translocator within the permeability transitionpore complex will increase the mitochondrial membranepermeability and thereby discharge a number of apopto-genic molecules into the cytosol [44] In addition to thisupregulation of the tumor suppressor protein p53 which hasa critical role in regulation of the Bcl-2 and p27 and p21draws attention to the involvement of cell cycle arrest in theapoptosis process Alternatively we found the involvement ofextrinsic pathway proteins such as Fas and FasL upregulationand related upregulation of caspase 8 (Figure 7) Moreoverthe luminescent based caspase 8 assay had showed 10 folddifferences than the control Apart from this we foundthat there is a downregulation of Bid happening too Sincemitochondrial damage in the Fas pathway of apoptosis ismediated by the caspase 8 cleavage of Bid [45] the findingshere shows the cross talk between intrinsic and extrinsicpathways
The protein array results exhibited a significant reductionof both IGF-I and IGF-II IGF-I has been shown to exertstrongmitogenic and antiapoptotic effects in a variety of nor-mal and cancerous cells including lung cancer cell lines [46]Moreover the over expression of IGFBP3 is closely associatedwith the downregulation of IGF-I and its mitogenic activity[47] It is known that IGFBP-2 and IGFBP-1 are involved inthe regulation of cell migration apoptosis and cell growth
[48] Both are reported to be markedly overexpressed inmany tumors and tumor cell lines And it is assumed thatelevation of the IGFBP-2 production is part of a mechanismto compensate for the mitogenic and antiapoptotic effectsof tumor-derived IGFs [49] Our data clearly showed thedownregulation of IGFBP1 and IGFBP2 concomitant withupregulation of IGBP3
Our results provided a new insight into the mechanismof chemotherapeutic properties of Murraya koenigii Inaddition these study findings demonstrate that girinimbinemediates its apoptotic effects through both intrinsic andextrinsic pathway which is depended on caspase mediationMoreover the upregulation of p53 as well as the cell prolifer-ation repressor proteins p27 and p21 and the significant roleof insulinIGF-1 signaling were also identified These resultsindicate that girinimbine could be a candidate for a novelanticancer agentThereforemore in-depth in vitro and in vivostudies are currently going on in our laboratory
Conflict of Interests
The authors declare that they do not have any conflict ofinterests
Acknowledgment
The authors greatly acknowledge the financial support pro-vided by University of Malaya (Grant no HIR E0000002-20001)
References
[1] J K Field and S W Duffy ldquoLung cancer screening the wayforwardrdquo British Journal of Cancer vol 99 no 4 pp 557ndash5622008
[2] K A Yoon J H Park J Han et al ldquoA genome-wide associationstudy reveals susceptibility variants for nonsmall cell lungcancer in the Korean populationrdquo Human Molecular Geneticsvol 19 no 24 Article ID ddq421 pp 4948ndash4954 2010
[3] G Cartei F Cartei A Cantone et al ldquoCisplatin-cyclophos-phamide-mitomycin combination chemotherapy with support-ive care versus supportive care alone for treatment of metastaticnon- small-cell lung cancerrdquo Journal of the National CancerInstitute vol 85 no 10 pp 794ndash800 1993
[4] M O Hengartner ldquoThe biochemistry of apoptosisrdquoNature vol407 no 6805 pp 770ndash776 2000
[5] DHanahan andRAWeinberg ldquoThehallmarks of cancerrdquoCellvol 100 no 1 pp 57ndash70 2000
[6] J R Molina P Yang S D Cassivi S E Schild and A AAdjei ldquoNon-small cell lung cancer epidemiology risk factorstreatment and survivorshiprdquo Mayo Clinic Proceedings vol 83no 5 pp 584ndash594 2008
[7] S Diwanay D Chitre and B Patwardhan ldquoImmunoprotec-tion by botanical drugs in cancer chemotherapyrdquo Journal ofEthnopharmacology vol 90 no 1 pp 49ndash55 2004
[8] U M Thatte and S A Dahanukar ldquoRasayana Concept cluesfrom immunomodulatory therapyrdquo in An Annotated Bibliogra-phy of Indian Medicine S N Upadhyay Ed pp 141ndash148 1997
[9] A S Shah A S Wakade and A R Juvekar ldquoImmunomod-ulatory activity of methanolic extract of Murraya koenigii (L)
Evidence-Based Complementary and Alternative Medicine 11
Spreng leavesrdquo Indian Journal of Experimental Biology vol 46no 7 pp 505ndash509 2008
[10] R B Goswamia P K S Singha N Goswamib P Thomasd PU Devic and A K Pathakd ldquoStudies on antigenotoxic effect ofMurraya koenigii leavesrdquo International Journal of PharmaRecentResearch vol 2 pp 65ndash68 2010
[11] R S Ramsewak M G Nair G M Strasburg D L DeWittand J L Nitiss ldquoBiologically active carbazole alkaloids fromMurraya koenigiirdquo Journal of Agricultural and Food Chemistryvol 47 no 2 pp 444ndash447 1999
[12] D P Chakraborty B K Barman and P K Bose ldquoStructureof girinimbine a pyranocarbazole derivative isolated fromMurraya koenigii Sprengrdquo Scientific Culture vol 30 pp 445ndash448 1964
[13] L J M Rao K Ramalakshmi B B Borse and B RaghavanldquoAntioxidant and radical-scavenging carbazole alkaloids fromthe oleoresin of curry leaf (Murraya koenigii Spreng)rdquo FoodChemistry vol 100 no 2 pp 742ndash747 2007
[14] S L Wang B Cai C B Cui S Y Yan and C F Wu ldquoInductionof apoptosis by girinimbine in K562 cellrdquo Chinese Traditionaland Herbal Drugs vol 38 no 11 pp 1677ndash1681 2007
[15] S Syam A B Abdul M A Sukari S Mohan S I Abdelwahaband T S Wah ldquoThe growth suppressing effects of girinimbineon Hepg2 involve induction of apoptosis and cell cycle arrestrdquoMolecules vol 16 pp 7155ndash7170 2011
[16] A Bakar N Haslizawati M A Sukari et al ldquoChemical con-stituents from stem barks and roots of Murraya koenigii(Rutaceae)rdquo The Malaysian Journal of Analytical Sciences vol11 pp 173ndash176 2007
[17] S I A Wahab A B Abdul S M Mohan A S Al-ZubairiM M Elhassan and M Y Ibrahim ldquoBiological activities ofPereskia bleo extractsrdquo International Journal of Pharmacologyvol 5 no 1 pp 71ndash75 2009
[18] E Bossy-Wetzel and D R Green ldquoDetection of apoptosis byannexin V labelingrdquo Methods in Enzymology vol 322 pp 15ndash18 2000
[19] H Wang and J A Joseph ldquoQuantifying cellular oxidative stressby dichlorofluorescein assay usingmicroplate readerrdquo Free Rad-ical Biology and Medicine vol 27 no 5-6 pp 612ndash616 1999
[20] S C Cheah D R Appleton S T Lee M L Lam A H AHadi and M R Mustafa ldquoPanduratin a inhibits the growth ofA549 cells through induction of apoptosis and inhibition of NF-KappaB translocationrdquoMolecules vol 16 no 3 pp 2583ndash25982011
[21] L Foghsgaard DWissing DMauch et al ldquoCathepsin B acts asa dominant execution protease in tumor cell apoptosis inducedby tumor necrosis factorrdquo Journal of Cell Biology vol 153 no 5pp 999ndash1009 2001
[22] K Kagedal U Johansson and K Ollinger ldquoThe lysosomalprotease cathepsin D mediates apoptosis induced by oxidativestressrdquoThe FASEB journal vol 15 no 9 pp 1592ndash1594 2001
[23] M A Birch-Machin and H Swalwell ldquoHow mitochondriarecord the effects of UV exposure and oxidative stress usinghuman skin as a model tissuerdquo Mutagenesis vol 25 no 2 pp101ndash107 2010
[24] S Meister B Frey V R Lang et al ldquoCalcium channel blockerverapamil enhances endoplasmic reticulum stress and cell deathinduced by proteasome inhibition in myeloma cellsrdquoNeoplasiavol 12 no 7 pp 550ndash561 2010
[25] S Mohan A B Abdul S I Abdelwahab et al ldquoTyphoniumflagelliforme induces apoptosis in CEMss cells via activation
of caspase-9 PARP cleavage and cytochrome c release itsactivation coupled with G0G1 phase cell cycle arrestrdquo Journalof Ethnopharmacology vol 131 no 3 pp 592ndash600 2010
[26] S Elmore ldquoApoptosis a review of programmed cell deathrdquoToxicologic Pathology vol 35 no 4 pp 495ndash516 2007
[27] M Aberg M Johnell M Wickstrom and A Siegbahn ldquoTissueFactor FVIIa prevents the extrinsic pathway of apoptosis byregulation of the tumor suppressor Death-Associated ProteinKinase 1 (DAPK1)rdquoThrombosis Research vol 127 no 2 pp 141ndash148 2011
[28] M Nigam N Singh V Ranjan et al ldquoCentchroman mediatedapoptosis involves cross-talk between extrinsicintrinsic path-ways and oxidative regulationrdquo Life Sciences vol 87 no 23ndash26pp 750ndash758 2010
[29] O S Frankfurt and A Krishan ldquoApoptosis-based drug screen-ing and detection of selective toxicity to cancer cellsrdquo Anti-Cancer Drugs vol 14 no 7 pp 555ndash561 2003
[30] M van Engeland L J W Nieland F C S Ramaekers BSchutte and C P M Reutelingsperger ldquoAnnexin V-affinityassay a review on an apoptosis detection system based onphosphatidylserine exposurerdquo Cytometry vol 31 no 1 pp 1ndash91998
[31] G A Joanitti R B Azevedo and S M Freitas ldquoApoptosisand lysosome membrane permeabilization induction on breastcancer cells by an anticarcinogenic Bowman-Birk proteaseinhibitor from Vigna unguiculata seedsrdquo Cancer Letters vol293 no 1 pp 73ndash81 2010
[32] T Kirkegaard and M Jaattela ldquoLysosomal involvement in celldeath and cancerrdquo Biochimica et Biophysica ActamdashMolecularCell Research vol 1793 no 4 pp 746ndash754 2009
[33] A C Johansson H Appelqvist C Nilsson K Kagedal KRoberg and K Ollinger ldquoRegulation of apoptosis-associatedlysosomal membrane permeabilizationrdquo Apoptosis vol 15 no5 pp 527ndash540 2010
[34] J Wen K R You S Y Lee C H Song and D G KimldquoOxidative stress-mediated apoptosis the anticancer effect ofthe sesquiterpene lactone parthenoliderdquo Journal of BiologicalChemistry vol 277 no 41 pp 38954ndash38964 2002
[35] C Fleury B Mignotte and J L Vayssiere ldquoMitochondrialreactive oxygen species in cell death signalingrdquo Biochimie vol84 no 2-3 pp 131ndash141 2002
[36] A M Verhagen P G Ekert M Pakusch et al ldquoIdentificationof DIABLO a mammalian protein that promotes apoptosis bybinding to and antagonizing IAP proteinsrdquo Cell vol 102 no 1pp 43ndash53 2000
[37] N Ajenjo E Canonon I Sanchez-Perez et al ldquoSubcellularlocalization determines the protective effects of activated ERK2against distinct apoptogenic stimuli in myeloid leukemia cellsrdquoJournal of Biological Chemistry vol 279 no 31 pp 32813ndash328232004
[38] X Wang ldquoThe expanding role of mitochondria in apoptosisrdquoGenes and Development vol 15 no 22 pp 2922ndash2933 2001
[39] Y A Hannun ldquoApoptosis and the dilemma of cancer chemo-therapyrdquo Blood vol 89 no 6 pp 1845ndash1853 1997
[40] F L Scott J B Denault S J Riedl H Shin M Renatusand G S Salvesen ldquoXIAP inhibits caspase-3 and -7 usingtwo binding sites evolutionary conserved mechanism of IAPsrdquoEMBO Journal vol 24 no 3 pp 645ndash655 2005
[41] S Cory and J M Adams ldquoThe BCL2 family regulators of thecellular life-or-death switchrdquo Nature Reviews Cancer vol 2 no9 pp 647ndash656 2002
12 Evidence-Based Complementary and Alternative Medicine
[42] S B Bratton and GM Cohen ldquoDeath receptors leave a caspasefootprint that Smacs of XIAPrdquo Cell Death and Differentiationvol 10 no 1 pp 4ndash6 2003
[43] N Zamzami andGKroemer ldquoThemitochondrion in apoptosishow Pandorarsquos box opensrdquo Nature Reviews Molecular CellBiology vol 2 no 1 pp 67ndash71 2001
[44] I Marzo C Brenner N Zamzami et al ldquoBax and adeninenucleotide translocator cooperate in the mitochondrial controlof apoptosisrdquo Science vol 281 no 5385 pp 2027ndash2031 1998
[45] M D Esposti ldquoThe roles of BidrdquoApoptosis vol 7 no 5 pp 433ndash440 2002
[46] N E Allen A W Roddam D S Allen et al ldquoA prospectivestudy of serum insulin-like growth factor-I (IGF-I) IGF-IIIGF-binding protein-3 and breast cancer riskrdquo British Journalof Cancer vol 92 no 7 pp 1283ndash1287 2005
[47] L Buckbinder R Talbott S Velasco-Miguel et al ldquoInductionof the growth inhibitor IGF-binding protein 3 by p53rdquo Naturevol 377 no 6550 pp 646ndash649 1995
[48] E Ruoslahti ldquoFibronectin and its integrin receptors in cancerrdquoAdvances in Cancer Research vol 76 pp 18ndash20 1999
[49] B S Schutt M Langkamp U Rauschnabel M B Ranke andM W Elmlinger ldquoIntegrin-mediated action of insulin-likefactor binding protein-2 in tumor cellsrdquo Journal of MolecularEndocrinology vol 32 no 3 pp 859ndash868 2004
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
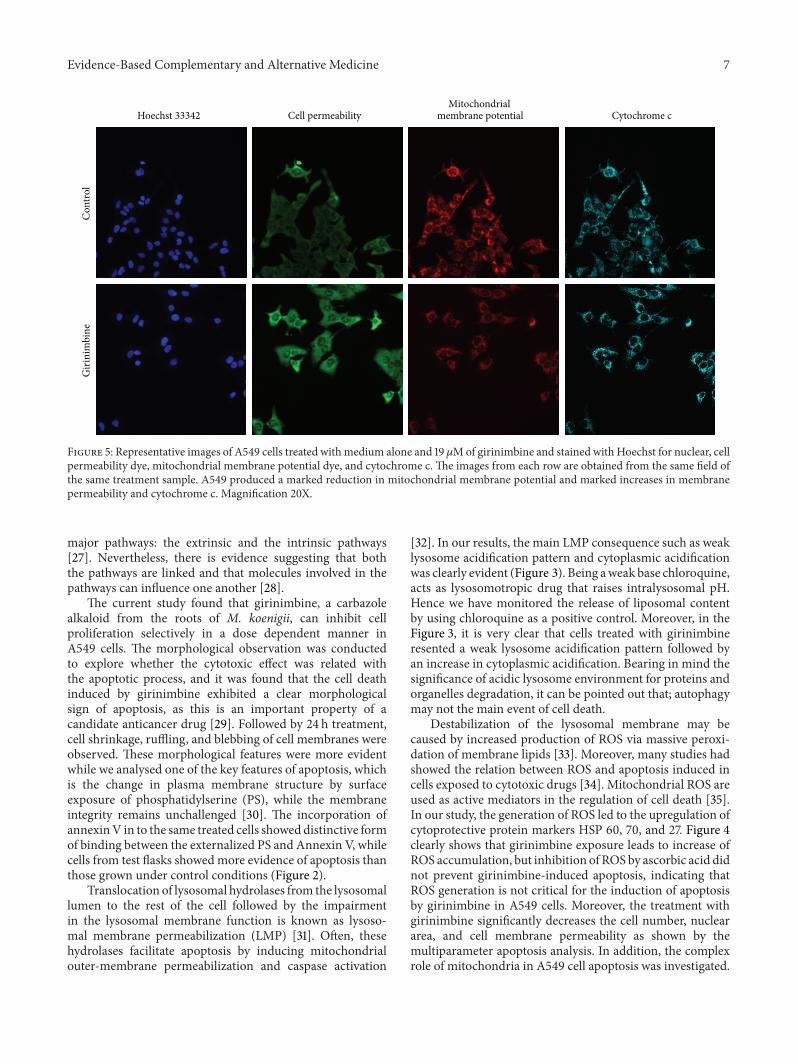
Evidence-Based Complementary and Alternative Medicine 7
Hoechst 33342 Cell permeability Cytochrome c
Con
trol
Giri
nim
bine
Mitochondrial
membrane potential
Figure 5 Representative images of A549 cells treated withmedium alone and 19 120583Mof girinimbine and stained with Hoechst for nuclear cellpermeability dye mitochondrial membrane potential dye and cytochrome c The images from each row are obtained from the same field ofthe same treatment sample A549 produced a marked reduction in mitochondrial membrane potential and marked increases in membranepermeability and cytochrome c Magnification 20X
major pathways the extrinsic and the intrinsic pathways[27] Nevertheless there is evidence suggesting that boththe pathways are linked and that molecules involved in thepathways can influence one another [28]
The current study found that girinimbine a carbazolealkaloid from the roots of M koenigii can inhibit cellproliferation selectively in a dose dependent manner inA549 cells The morphological observation was conductedto explore whether the cytotoxic effect was related withthe apoptotic process and it was found that the cell deathinduced by girinimbine exhibited a clear morphologicalsign of apoptosis as this is an important property of acandidate anticancer drug [29] Followed by 24 h treatmentcell shrinkage ruffling and blebbing of cell membranes wereobserved These morphological features were more evidentwhile we analysed one of the key features of apoptosis whichis the change in plasma membrane structure by surfaceexposure of phosphatidylserine (PS) while the membraneintegrity remains unchallenged [30] The incorporation ofannexinV in to the same treated cells showed distinctive formof binding between the externalized PS and Annexin V whilecells from test flasks showedmore evidence of apoptosis thanthose grown under control conditions (Figure 2)
Translocation of lysosomal hydrolases from the lysosomallumen to the rest of the cell followed by the impairmentin the lysosomal membrane function is known as lysoso-mal membrane permeabilization (LMP) [31] Often thesehydrolases facilitate apoptosis by inducing mitochondrialouter-membrane permeabilization and caspase activation
[32] In our results the main LMP consequence such as weaklysosome acidification pattern and cytoplasmic acidificationwas clearly evident (Figure 3) Being aweak base chloroquineacts as lysosomotropic drug that raises intralysosomal pHHence we have monitored the release of liposomal contentby using chloroquine as a positive control Moreover in theFigure 3 it is very clear that cells treated with girinimbineresented a weak lysosome acidification pattern followed byan increase in cytoplasmic acidification Bearing in mind thesignificance of acidic lysosome environment for proteins andorganelles degradation it can be pointed out that autophagymay not the main event of cell death
Destabilization of the lysosomal membrane may becaused by increased production of ROS via massive peroxi-dation of membrane lipids [33] Moreover many studies hadshowed the relation between ROS and apoptosis induced incells exposed to cytotoxic drugs [34] Mitochondrial ROS areused as active mediators in the regulation of cell death [35]In our study the generation of ROS led to the upregulation ofcytoprotective protein markers HSP 60 70 and 27 Figure 4clearly shows that girinimbine exposure leads to increase ofROS accumulation but inhibition of ROSby ascorbic acid didnot prevent girinimbine-induced apoptosis indicating thatROS generation is not critical for the induction of apoptosisby girinimbine in A549 cells Moreover the treatment withgirinimbine significantly decreases the cell number nucleararea and cell membrane permeability as shown by themultiparameter apoptosis analysis In addition the complexrole of mitochondria in A549 cell apoptosis was investigated
8 Evidence-Based Complementary and Alternative Medicine
600
500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Total nuclear intensity
Giri19120583
M
Giri95120583
M
Giri475120583
M
(a)
Cell permeability500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(b)
350
300
250
200
150
100
50
0
Mitochondrial membrane potential
Con
trol
Pacli
taxe
l68120583
M
lowastlowastlowastlowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(c)
600700800
500400300200100
0
Cytochrome c
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowastlowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(d)
Figure 6 Quantitative analysis of girinimbine mediated apoptosis parameter Changes in total nuclear intensity (a) cell permeability(b) mitochondrial membrane potential (c) and cytochrome c localization (d) were measured simultaneously in A549 cells Followingtreatment with girinimbine we saw statistically significant increase in total nuclear intensity increased cell permeability loss ofmitochondrialmembrane potential and cytochrome c release frommitochondria Each experiment was performed at least two times Results are expressedas the means plusmn SD Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software (version40 GraphPad Software Inc San Diego CA) Statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
Mitochondria have been described to play a central role in theapoptotic process due to the mitochondrial proteins ability toactivate cellular apoptotic programs directly [36] Detectionof changes in mitochondrial membrane potential (Ψm) wascarried out as it is assumed that its disruption is the onsetofmitochondrial membrane transition pores (MPTP) forma-tion The present study revealed that girinimbine may act onmitochondria causing loss of Ψm and subsequent apoptosisThis kind of Ψm reduction and generation of MPTP mustfacilitate the relocalization of apoptogenic proteins from onesubcellular compartment such as mitochondria to cytoplasmto gain access to their substrates or interacting partners [37]
The multi cytotoxicity assay shows that girinimbine ind-uced cell death could be through the classical mitochondrial
pathway with cytochrome c release and caspase-dependentapoptosis The translocation of cytochrome c a componentof themitochondrial electron transfer chain was significantlyincreased upon girinimbine treatment In the mitochondriamediated apoptosis cytochrome c in turn binds to Apaf-1 to form a complex which triggers its oligomerization toform the apoptosome The caspase-9 holoenzyme is thenforms due to the binding of procaspase-9 to the apop-tosome and sequentially cleaves and activates the down-stream caspases such as caspase-3 [38] Our findings onthe upregulation of caspase 37 caspase 9 reduction in Ψmand release of cytochrome c to the cytosol substantiate thistheory and strongly support the involvement ofmitochondriain the apoptosis induced by girinimbine in A549 cells
Evidence-Based Complementary and Alternative Medicine 9
bad
bax
bcl-2
bcl-w
BID BI
MCa
spas
e 3Ca
spas
e 8IG
F-II IG
F-I HTR
AH
SP70
HSP
60H
SP27
FasL Fa
sD
R6Cy
to c
IGFB
P-1
IGFB
P-2 IG
FBP-
3IG
FBP-
4IG
FBP-
5IG
FBP-
6IG
F-1s
RLi
vin p
21 p
27 p53
SM
AC S
urvi
vin
sTN
F-R1 XI
AP
minus3
minus2
minus1
0
1
2
3
Relat
ive d
ensit
y di
ffere
nce t
o co
ntro
l (times10
5)
(a)
(b) (c)
A B C D E F G H I J K L M N1 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 82 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 83 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II4 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II5 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R16 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R17 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg Pos8 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg PosTNF-120572
TNF-120572 TNF-120573TNF-120573
(d)
Figure 7 Cells were lysed and protein arrays were performed Cells was treated with 19120583M girinimbine for 24 h and the whole cell proteinwas extracted Equal amount of (300 120583g) of protein from each sample was used for the assay Quantitative analysis in the arrays showeddifferences in the apoptotic markers (a) Representative images of the apoptotic protein array are shown for the control (b) treatment (c) andthe exact protein name of each dot in the array (d)
Suppression of apoptosis may promote the cancer develop-ment by inhibiting the chemotherapy as well as other forms ofinducers of cell death [39] Regulation of apoptosis has beencensoriously dependent on several genes which have beenidentified earlier and includes XIAP-a member of the IAPfamily It has the capacity to inhibit the activation of caspases3 7 and 9 [40] In our study we found that girinimbinetreatment of A549 cells caused inhibition of XIAP but SMAC
an antagonist to XIAP could be found to be slightly regulatedin this treatment concentration This is considered as a keyevent in the execution of cell death which is regulatedmainlyby proteins of the Bcl-2 family [41 42]
Critical issues in apoptosis include the dominance ofanti-versus proapoptotic Bcl-2 members [43] Girinimbine isfound to induce apoptosis in the A549 cells involvingdownregulated Bcl-2 expression This downregulation was
10 Evidence-Based Complementary and Alternative Medicine
3
2
1
0
times105
19 9 475 0 19 9 475 0 19 9 475 0Girinimbine treatment (120583M)
Lum
ines
cenc
e
Caspase 37Caspase 8Caspase 9
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast
Figure 8 Effect of girinimbine on activation of caspases 37 8 and 9Cells were treated with girinimbine at the indicated concentrationsfor 24 h and activity was determined by using a luminescentenzyme assay kit Each point represents the mean plusmn SD of threedifferent experiments Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software(version 40 GraphPad Software Inc San Diego CA) Statisticalsignificance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
concomitant with the upregulation of Bad Bax and Bimwhich inhibits antiapoptotic proteins and render the cellsmore susceptible to apoptogenic stimuli (Figure 7) Therelease of cytochrome c found in our study could be due tothe up regulated Bax since Bax in association with adeninenucleotide translocator within the permeability transitionpore complex will increase the mitochondrial membranepermeability and thereby discharge a number of apopto-genic molecules into the cytosol [44] In addition to thisupregulation of the tumor suppressor protein p53 which hasa critical role in regulation of the Bcl-2 and p27 and p21draws attention to the involvement of cell cycle arrest in theapoptosis process Alternatively we found the involvement ofextrinsic pathway proteins such as Fas and FasL upregulationand related upregulation of caspase 8 (Figure 7) Moreoverthe luminescent based caspase 8 assay had showed 10 folddifferences than the control Apart from this we foundthat there is a downregulation of Bid happening too Sincemitochondrial damage in the Fas pathway of apoptosis ismediated by the caspase 8 cleavage of Bid [45] the findingshere shows the cross talk between intrinsic and extrinsicpathways
The protein array results exhibited a significant reductionof both IGF-I and IGF-II IGF-I has been shown to exertstrongmitogenic and antiapoptotic effects in a variety of nor-mal and cancerous cells including lung cancer cell lines [46]Moreover the over expression of IGFBP3 is closely associatedwith the downregulation of IGF-I and its mitogenic activity[47] It is known that IGFBP-2 and IGFBP-1 are involved inthe regulation of cell migration apoptosis and cell growth
[48] Both are reported to be markedly overexpressed inmany tumors and tumor cell lines And it is assumed thatelevation of the IGFBP-2 production is part of a mechanismto compensate for the mitogenic and antiapoptotic effectsof tumor-derived IGFs [49] Our data clearly showed thedownregulation of IGFBP1 and IGFBP2 concomitant withupregulation of IGBP3
Our results provided a new insight into the mechanismof chemotherapeutic properties of Murraya koenigii Inaddition these study findings demonstrate that girinimbinemediates its apoptotic effects through both intrinsic andextrinsic pathway which is depended on caspase mediationMoreover the upregulation of p53 as well as the cell prolifer-ation repressor proteins p27 and p21 and the significant roleof insulinIGF-1 signaling were also identified These resultsindicate that girinimbine could be a candidate for a novelanticancer agentThereforemore in-depth in vitro and in vivostudies are currently going on in our laboratory
Conflict of Interests
The authors declare that they do not have any conflict ofinterests
Acknowledgment
The authors greatly acknowledge the financial support pro-vided by University of Malaya (Grant no HIR E0000002-20001)
References
[1] J K Field and S W Duffy ldquoLung cancer screening the wayforwardrdquo British Journal of Cancer vol 99 no 4 pp 557ndash5622008
[2] K A Yoon J H Park J Han et al ldquoA genome-wide associationstudy reveals susceptibility variants for nonsmall cell lungcancer in the Korean populationrdquo Human Molecular Geneticsvol 19 no 24 Article ID ddq421 pp 4948ndash4954 2010
[3] G Cartei F Cartei A Cantone et al ldquoCisplatin-cyclophos-phamide-mitomycin combination chemotherapy with support-ive care versus supportive care alone for treatment of metastaticnon- small-cell lung cancerrdquo Journal of the National CancerInstitute vol 85 no 10 pp 794ndash800 1993
[4] M O Hengartner ldquoThe biochemistry of apoptosisrdquoNature vol407 no 6805 pp 770ndash776 2000
[5] DHanahan andRAWeinberg ldquoThehallmarks of cancerrdquoCellvol 100 no 1 pp 57ndash70 2000
[6] J R Molina P Yang S D Cassivi S E Schild and A AAdjei ldquoNon-small cell lung cancer epidemiology risk factorstreatment and survivorshiprdquo Mayo Clinic Proceedings vol 83no 5 pp 584ndash594 2008
[7] S Diwanay D Chitre and B Patwardhan ldquoImmunoprotec-tion by botanical drugs in cancer chemotherapyrdquo Journal ofEthnopharmacology vol 90 no 1 pp 49ndash55 2004
[8] U M Thatte and S A Dahanukar ldquoRasayana Concept cluesfrom immunomodulatory therapyrdquo in An Annotated Bibliogra-phy of Indian Medicine S N Upadhyay Ed pp 141ndash148 1997
[9] A S Shah A S Wakade and A R Juvekar ldquoImmunomod-ulatory activity of methanolic extract of Murraya koenigii (L)
Evidence-Based Complementary and Alternative Medicine 11
Spreng leavesrdquo Indian Journal of Experimental Biology vol 46no 7 pp 505ndash509 2008
[10] R B Goswamia P K S Singha N Goswamib P Thomasd PU Devic and A K Pathakd ldquoStudies on antigenotoxic effect ofMurraya koenigii leavesrdquo International Journal of PharmaRecentResearch vol 2 pp 65ndash68 2010
[11] R S Ramsewak M G Nair G M Strasburg D L DeWittand J L Nitiss ldquoBiologically active carbazole alkaloids fromMurraya koenigiirdquo Journal of Agricultural and Food Chemistryvol 47 no 2 pp 444ndash447 1999
[12] D P Chakraborty B K Barman and P K Bose ldquoStructureof girinimbine a pyranocarbazole derivative isolated fromMurraya koenigii Sprengrdquo Scientific Culture vol 30 pp 445ndash448 1964
[13] L J M Rao K Ramalakshmi B B Borse and B RaghavanldquoAntioxidant and radical-scavenging carbazole alkaloids fromthe oleoresin of curry leaf (Murraya koenigii Spreng)rdquo FoodChemistry vol 100 no 2 pp 742ndash747 2007
[14] S L Wang B Cai C B Cui S Y Yan and C F Wu ldquoInductionof apoptosis by girinimbine in K562 cellrdquo Chinese Traditionaland Herbal Drugs vol 38 no 11 pp 1677ndash1681 2007
[15] S Syam A B Abdul M A Sukari S Mohan S I Abdelwahaband T S Wah ldquoThe growth suppressing effects of girinimbineon Hepg2 involve induction of apoptosis and cell cycle arrestrdquoMolecules vol 16 pp 7155ndash7170 2011
[16] A Bakar N Haslizawati M A Sukari et al ldquoChemical con-stituents from stem barks and roots of Murraya koenigii(Rutaceae)rdquo The Malaysian Journal of Analytical Sciences vol11 pp 173ndash176 2007
[17] S I A Wahab A B Abdul S M Mohan A S Al-ZubairiM M Elhassan and M Y Ibrahim ldquoBiological activities ofPereskia bleo extractsrdquo International Journal of Pharmacologyvol 5 no 1 pp 71ndash75 2009
[18] E Bossy-Wetzel and D R Green ldquoDetection of apoptosis byannexin V labelingrdquo Methods in Enzymology vol 322 pp 15ndash18 2000
[19] H Wang and J A Joseph ldquoQuantifying cellular oxidative stressby dichlorofluorescein assay usingmicroplate readerrdquo Free Rad-ical Biology and Medicine vol 27 no 5-6 pp 612ndash616 1999
[20] S C Cheah D R Appleton S T Lee M L Lam A H AHadi and M R Mustafa ldquoPanduratin a inhibits the growth ofA549 cells through induction of apoptosis and inhibition of NF-KappaB translocationrdquoMolecules vol 16 no 3 pp 2583ndash25982011
[21] L Foghsgaard DWissing DMauch et al ldquoCathepsin B acts asa dominant execution protease in tumor cell apoptosis inducedby tumor necrosis factorrdquo Journal of Cell Biology vol 153 no 5pp 999ndash1009 2001
[22] K Kagedal U Johansson and K Ollinger ldquoThe lysosomalprotease cathepsin D mediates apoptosis induced by oxidativestressrdquoThe FASEB journal vol 15 no 9 pp 1592ndash1594 2001
[23] M A Birch-Machin and H Swalwell ldquoHow mitochondriarecord the effects of UV exposure and oxidative stress usinghuman skin as a model tissuerdquo Mutagenesis vol 25 no 2 pp101ndash107 2010
[24] S Meister B Frey V R Lang et al ldquoCalcium channel blockerverapamil enhances endoplasmic reticulum stress and cell deathinduced by proteasome inhibition in myeloma cellsrdquoNeoplasiavol 12 no 7 pp 550ndash561 2010
[25] S Mohan A B Abdul S I Abdelwahab et al ldquoTyphoniumflagelliforme induces apoptosis in CEMss cells via activation
of caspase-9 PARP cleavage and cytochrome c release itsactivation coupled with G0G1 phase cell cycle arrestrdquo Journalof Ethnopharmacology vol 131 no 3 pp 592ndash600 2010
[26] S Elmore ldquoApoptosis a review of programmed cell deathrdquoToxicologic Pathology vol 35 no 4 pp 495ndash516 2007
[27] M Aberg M Johnell M Wickstrom and A Siegbahn ldquoTissueFactor FVIIa prevents the extrinsic pathway of apoptosis byregulation of the tumor suppressor Death-Associated ProteinKinase 1 (DAPK1)rdquoThrombosis Research vol 127 no 2 pp 141ndash148 2011
[28] M Nigam N Singh V Ranjan et al ldquoCentchroman mediatedapoptosis involves cross-talk between extrinsicintrinsic path-ways and oxidative regulationrdquo Life Sciences vol 87 no 23ndash26pp 750ndash758 2010
[29] O S Frankfurt and A Krishan ldquoApoptosis-based drug screen-ing and detection of selective toxicity to cancer cellsrdquo Anti-Cancer Drugs vol 14 no 7 pp 555ndash561 2003
[30] M van Engeland L J W Nieland F C S Ramaekers BSchutte and C P M Reutelingsperger ldquoAnnexin V-affinityassay a review on an apoptosis detection system based onphosphatidylserine exposurerdquo Cytometry vol 31 no 1 pp 1ndash91998
[31] G A Joanitti R B Azevedo and S M Freitas ldquoApoptosisand lysosome membrane permeabilization induction on breastcancer cells by an anticarcinogenic Bowman-Birk proteaseinhibitor from Vigna unguiculata seedsrdquo Cancer Letters vol293 no 1 pp 73ndash81 2010
[32] T Kirkegaard and M Jaattela ldquoLysosomal involvement in celldeath and cancerrdquo Biochimica et Biophysica ActamdashMolecularCell Research vol 1793 no 4 pp 746ndash754 2009
[33] A C Johansson H Appelqvist C Nilsson K Kagedal KRoberg and K Ollinger ldquoRegulation of apoptosis-associatedlysosomal membrane permeabilizationrdquo Apoptosis vol 15 no5 pp 527ndash540 2010
[34] J Wen K R You S Y Lee C H Song and D G KimldquoOxidative stress-mediated apoptosis the anticancer effect ofthe sesquiterpene lactone parthenoliderdquo Journal of BiologicalChemistry vol 277 no 41 pp 38954ndash38964 2002
[35] C Fleury B Mignotte and J L Vayssiere ldquoMitochondrialreactive oxygen species in cell death signalingrdquo Biochimie vol84 no 2-3 pp 131ndash141 2002
[36] A M Verhagen P G Ekert M Pakusch et al ldquoIdentificationof DIABLO a mammalian protein that promotes apoptosis bybinding to and antagonizing IAP proteinsrdquo Cell vol 102 no 1pp 43ndash53 2000
[37] N Ajenjo E Canonon I Sanchez-Perez et al ldquoSubcellularlocalization determines the protective effects of activated ERK2against distinct apoptogenic stimuli in myeloid leukemia cellsrdquoJournal of Biological Chemistry vol 279 no 31 pp 32813ndash328232004
[38] X Wang ldquoThe expanding role of mitochondria in apoptosisrdquoGenes and Development vol 15 no 22 pp 2922ndash2933 2001
[39] Y A Hannun ldquoApoptosis and the dilemma of cancer chemo-therapyrdquo Blood vol 89 no 6 pp 1845ndash1853 1997
[40] F L Scott J B Denault S J Riedl H Shin M Renatusand G S Salvesen ldquoXIAP inhibits caspase-3 and -7 usingtwo binding sites evolutionary conserved mechanism of IAPsrdquoEMBO Journal vol 24 no 3 pp 645ndash655 2005
[41] S Cory and J M Adams ldquoThe BCL2 family regulators of thecellular life-or-death switchrdquo Nature Reviews Cancer vol 2 no9 pp 647ndash656 2002
12 Evidence-Based Complementary and Alternative Medicine
[42] S B Bratton and GM Cohen ldquoDeath receptors leave a caspasefootprint that Smacs of XIAPrdquo Cell Death and Differentiationvol 10 no 1 pp 4ndash6 2003
[43] N Zamzami andGKroemer ldquoThemitochondrion in apoptosishow Pandorarsquos box opensrdquo Nature Reviews Molecular CellBiology vol 2 no 1 pp 67ndash71 2001
[44] I Marzo C Brenner N Zamzami et al ldquoBax and adeninenucleotide translocator cooperate in the mitochondrial controlof apoptosisrdquo Science vol 281 no 5385 pp 2027ndash2031 1998
[45] M D Esposti ldquoThe roles of BidrdquoApoptosis vol 7 no 5 pp 433ndash440 2002
[46] N E Allen A W Roddam D S Allen et al ldquoA prospectivestudy of serum insulin-like growth factor-I (IGF-I) IGF-IIIGF-binding protein-3 and breast cancer riskrdquo British Journalof Cancer vol 92 no 7 pp 1283ndash1287 2005
[47] L Buckbinder R Talbott S Velasco-Miguel et al ldquoInductionof the growth inhibitor IGF-binding protein 3 by p53rdquo Naturevol 377 no 6550 pp 646ndash649 1995
[48] E Ruoslahti ldquoFibronectin and its integrin receptors in cancerrdquoAdvances in Cancer Research vol 76 pp 18ndash20 1999
[49] B S Schutt M Langkamp U Rauschnabel M B Ranke andM W Elmlinger ldquoIntegrin-mediated action of insulin-likefactor binding protein-2 in tumor cellsrdquo Journal of MolecularEndocrinology vol 32 no 3 pp 859ndash868 2004
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
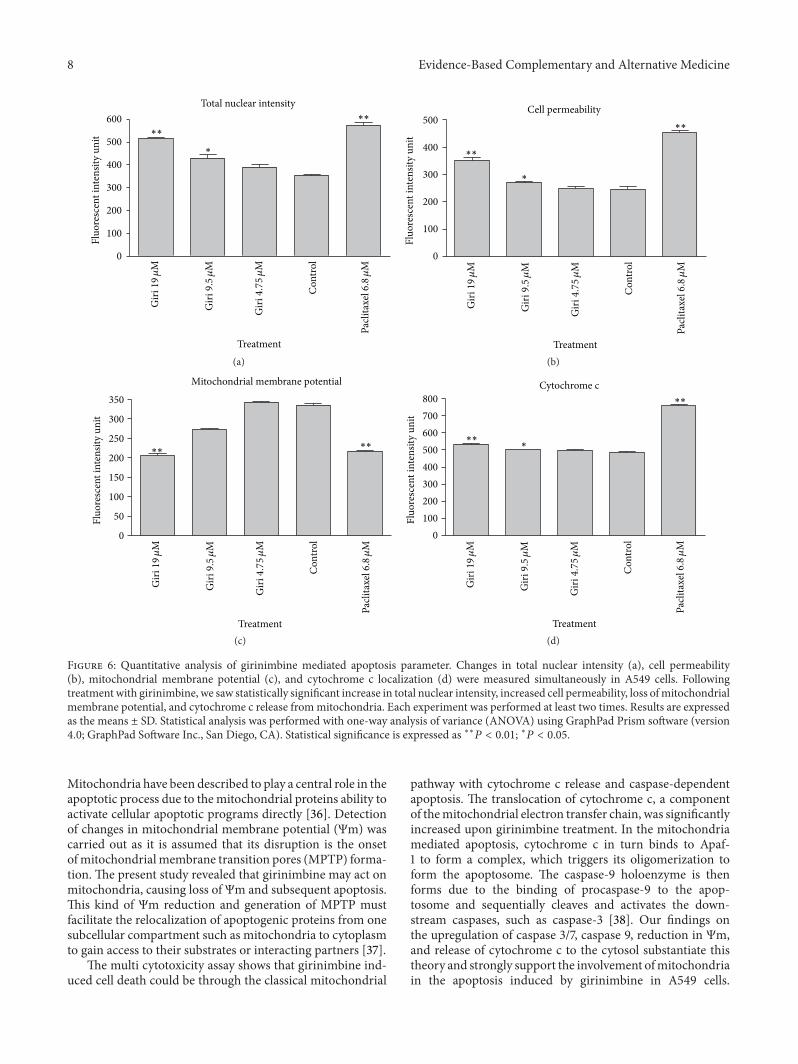
8 Evidence-Based Complementary and Alternative Medicine
600
500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Total nuclear intensity
Giri19120583
M
Giri95120583
M
Giri475120583
M
(a)
Cell permeability500
400
300
200
100
0
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowast
lowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(b)
350
300
250
200
150
100
50
0
Mitochondrial membrane potential
Con
trol
Pacli
taxe
l68120583
M
lowastlowastlowastlowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(c)
600700800
500400300200100
0
Cytochrome c
Con
trol
Pacli
taxe
l68120583
M
lowastlowast
lowastlowast
lowast
Treatment
Fluo
resc
ent i
nten
sity
unit
Giri19120583
M
Giri95120583
M
Giri475120583
M
(d)
Figure 6 Quantitative analysis of girinimbine mediated apoptosis parameter Changes in total nuclear intensity (a) cell permeability(b) mitochondrial membrane potential (c) and cytochrome c localization (d) were measured simultaneously in A549 cells Followingtreatment with girinimbine we saw statistically significant increase in total nuclear intensity increased cell permeability loss ofmitochondrialmembrane potential and cytochrome c release frommitochondria Each experiment was performed at least two times Results are expressedas the means plusmn SD Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software (version40 GraphPad Software Inc San Diego CA) Statistical significance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
Mitochondria have been described to play a central role in theapoptotic process due to the mitochondrial proteins ability toactivate cellular apoptotic programs directly [36] Detectionof changes in mitochondrial membrane potential (Ψm) wascarried out as it is assumed that its disruption is the onsetofmitochondrial membrane transition pores (MPTP) forma-tion The present study revealed that girinimbine may act onmitochondria causing loss of Ψm and subsequent apoptosisThis kind of Ψm reduction and generation of MPTP mustfacilitate the relocalization of apoptogenic proteins from onesubcellular compartment such as mitochondria to cytoplasmto gain access to their substrates or interacting partners [37]
The multi cytotoxicity assay shows that girinimbine ind-uced cell death could be through the classical mitochondrial
pathway with cytochrome c release and caspase-dependentapoptosis The translocation of cytochrome c a componentof themitochondrial electron transfer chain was significantlyincreased upon girinimbine treatment In the mitochondriamediated apoptosis cytochrome c in turn binds to Apaf-1 to form a complex which triggers its oligomerization toform the apoptosome The caspase-9 holoenzyme is thenforms due to the binding of procaspase-9 to the apop-tosome and sequentially cleaves and activates the down-stream caspases such as caspase-3 [38] Our findings onthe upregulation of caspase 37 caspase 9 reduction in Ψmand release of cytochrome c to the cytosol substantiate thistheory and strongly support the involvement ofmitochondriain the apoptosis induced by girinimbine in A549 cells
Evidence-Based Complementary and Alternative Medicine 9
bad
bax
bcl-2
bcl-w
BID BI
MCa
spas
e 3Ca
spas
e 8IG
F-II IG
F-I HTR
AH
SP70
HSP
60H
SP27
FasL Fa
sD
R6Cy
to c
IGFB
P-1
IGFB
P-2 IG
FBP-
3IG
FBP-
4IG
FBP-
5IG
FBP-
6IG
F-1s
RLi
vin p
21 p
27 p53
SM
AC S
urvi
vin
sTN
F-R1 XI
AP
minus3
minus2
minus1
0
1
2
3
Relat
ive d
ensit
y di
ffere
nce t
o co
ntro
l (times10
5)
(a)
(b) (c)
A B C D E F G H I J K L M N1 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 82 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 83 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II4 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II5 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R16 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R17 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg Pos8 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg PosTNF-120572
TNF-120572 TNF-120573TNF-120573
(d)
Figure 7 Cells were lysed and protein arrays were performed Cells was treated with 19120583M girinimbine for 24 h and the whole cell proteinwas extracted Equal amount of (300 120583g) of protein from each sample was used for the assay Quantitative analysis in the arrays showeddifferences in the apoptotic markers (a) Representative images of the apoptotic protein array are shown for the control (b) treatment (c) andthe exact protein name of each dot in the array (d)
Suppression of apoptosis may promote the cancer develop-ment by inhibiting the chemotherapy as well as other forms ofinducers of cell death [39] Regulation of apoptosis has beencensoriously dependent on several genes which have beenidentified earlier and includes XIAP-a member of the IAPfamily It has the capacity to inhibit the activation of caspases3 7 and 9 [40] In our study we found that girinimbinetreatment of A549 cells caused inhibition of XIAP but SMAC
an antagonist to XIAP could be found to be slightly regulatedin this treatment concentration This is considered as a keyevent in the execution of cell death which is regulatedmainlyby proteins of the Bcl-2 family [41 42]
Critical issues in apoptosis include the dominance ofanti-versus proapoptotic Bcl-2 members [43] Girinimbine isfound to induce apoptosis in the A549 cells involvingdownregulated Bcl-2 expression This downregulation was
10 Evidence-Based Complementary and Alternative Medicine
3
2
1
0
times105
19 9 475 0 19 9 475 0 19 9 475 0Girinimbine treatment (120583M)
Lum
ines
cenc
e
Caspase 37Caspase 8Caspase 9
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast
Figure 8 Effect of girinimbine on activation of caspases 37 8 and 9Cells were treated with girinimbine at the indicated concentrationsfor 24 h and activity was determined by using a luminescentenzyme assay kit Each point represents the mean plusmn SD of threedifferent experiments Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software(version 40 GraphPad Software Inc San Diego CA) Statisticalsignificance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
concomitant with the upregulation of Bad Bax and Bimwhich inhibits antiapoptotic proteins and render the cellsmore susceptible to apoptogenic stimuli (Figure 7) Therelease of cytochrome c found in our study could be due tothe up regulated Bax since Bax in association with adeninenucleotide translocator within the permeability transitionpore complex will increase the mitochondrial membranepermeability and thereby discharge a number of apopto-genic molecules into the cytosol [44] In addition to thisupregulation of the tumor suppressor protein p53 which hasa critical role in regulation of the Bcl-2 and p27 and p21draws attention to the involvement of cell cycle arrest in theapoptosis process Alternatively we found the involvement ofextrinsic pathway proteins such as Fas and FasL upregulationand related upregulation of caspase 8 (Figure 7) Moreoverthe luminescent based caspase 8 assay had showed 10 folddifferences than the control Apart from this we foundthat there is a downregulation of Bid happening too Sincemitochondrial damage in the Fas pathway of apoptosis ismediated by the caspase 8 cleavage of Bid [45] the findingshere shows the cross talk between intrinsic and extrinsicpathways
The protein array results exhibited a significant reductionof both IGF-I and IGF-II IGF-I has been shown to exertstrongmitogenic and antiapoptotic effects in a variety of nor-mal and cancerous cells including lung cancer cell lines [46]Moreover the over expression of IGFBP3 is closely associatedwith the downregulation of IGF-I and its mitogenic activity[47] It is known that IGFBP-2 and IGFBP-1 are involved inthe regulation of cell migration apoptosis and cell growth
[48] Both are reported to be markedly overexpressed inmany tumors and tumor cell lines And it is assumed thatelevation of the IGFBP-2 production is part of a mechanismto compensate for the mitogenic and antiapoptotic effectsof tumor-derived IGFs [49] Our data clearly showed thedownregulation of IGFBP1 and IGFBP2 concomitant withupregulation of IGBP3
Our results provided a new insight into the mechanismof chemotherapeutic properties of Murraya koenigii Inaddition these study findings demonstrate that girinimbinemediates its apoptotic effects through both intrinsic andextrinsic pathway which is depended on caspase mediationMoreover the upregulation of p53 as well as the cell prolifer-ation repressor proteins p27 and p21 and the significant roleof insulinIGF-1 signaling were also identified These resultsindicate that girinimbine could be a candidate for a novelanticancer agentThereforemore in-depth in vitro and in vivostudies are currently going on in our laboratory
Conflict of Interests
The authors declare that they do not have any conflict ofinterests
Acknowledgment
The authors greatly acknowledge the financial support pro-vided by University of Malaya (Grant no HIR E0000002-20001)
References
[1] J K Field and S W Duffy ldquoLung cancer screening the wayforwardrdquo British Journal of Cancer vol 99 no 4 pp 557ndash5622008
[2] K A Yoon J H Park J Han et al ldquoA genome-wide associationstudy reveals susceptibility variants for nonsmall cell lungcancer in the Korean populationrdquo Human Molecular Geneticsvol 19 no 24 Article ID ddq421 pp 4948ndash4954 2010
[3] G Cartei F Cartei A Cantone et al ldquoCisplatin-cyclophos-phamide-mitomycin combination chemotherapy with support-ive care versus supportive care alone for treatment of metastaticnon- small-cell lung cancerrdquo Journal of the National CancerInstitute vol 85 no 10 pp 794ndash800 1993
[4] M O Hengartner ldquoThe biochemistry of apoptosisrdquoNature vol407 no 6805 pp 770ndash776 2000
[5] DHanahan andRAWeinberg ldquoThehallmarks of cancerrdquoCellvol 100 no 1 pp 57ndash70 2000
[6] J R Molina P Yang S D Cassivi S E Schild and A AAdjei ldquoNon-small cell lung cancer epidemiology risk factorstreatment and survivorshiprdquo Mayo Clinic Proceedings vol 83no 5 pp 584ndash594 2008
[7] S Diwanay D Chitre and B Patwardhan ldquoImmunoprotec-tion by botanical drugs in cancer chemotherapyrdquo Journal ofEthnopharmacology vol 90 no 1 pp 49ndash55 2004
[8] U M Thatte and S A Dahanukar ldquoRasayana Concept cluesfrom immunomodulatory therapyrdquo in An Annotated Bibliogra-phy of Indian Medicine S N Upadhyay Ed pp 141ndash148 1997
[9] A S Shah A S Wakade and A R Juvekar ldquoImmunomod-ulatory activity of methanolic extract of Murraya koenigii (L)
Evidence-Based Complementary and Alternative Medicine 11
Spreng leavesrdquo Indian Journal of Experimental Biology vol 46no 7 pp 505ndash509 2008
[10] R B Goswamia P K S Singha N Goswamib P Thomasd PU Devic and A K Pathakd ldquoStudies on antigenotoxic effect ofMurraya koenigii leavesrdquo International Journal of PharmaRecentResearch vol 2 pp 65ndash68 2010
[11] R S Ramsewak M G Nair G M Strasburg D L DeWittand J L Nitiss ldquoBiologically active carbazole alkaloids fromMurraya koenigiirdquo Journal of Agricultural and Food Chemistryvol 47 no 2 pp 444ndash447 1999
[12] D P Chakraborty B K Barman and P K Bose ldquoStructureof girinimbine a pyranocarbazole derivative isolated fromMurraya koenigii Sprengrdquo Scientific Culture vol 30 pp 445ndash448 1964
[13] L J M Rao K Ramalakshmi B B Borse and B RaghavanldquoAntioxidant and radical-scavenging carbazole alkaloids fromthe oleoresin of curry leaf (Murraya koenigii Spreng)rdquo FoodChemistry vol 100 no 2 pp 742ndash747 2007
[14] S L Wang B Cai C B Cui S Y Yan and C F Wu ldquoInductionof apoptosis by girinimbine in K562 cellrdquo Chinese Traditionaland Herbal Drugs vol 38 no 11 pp 1677ndash1681 2007
[15] S Syam A B Abdul M A Sukari S Mohan S I Abdelwahaband T S Wah ldquoThe growth suppressing effects of girinimbineon Hepg2 involve induction of apoptosis and cell cycle arrestrdquoMolecules vol 16 pp 7155ndash7170 2011
[16] A Bakar N Haslizawati M A Sukari et al ldquoChemical con-stituents from stem barks and roots of Murraya koenigii(Rutaceae)rdquo The Malaysian Journal of Analytical Sciences vol11 pp 173ndash176 2007
[17] S I A Wahab A B Abdul S M Mohan A S Al-ZubairiM M Elhassan and M Y Ibrahim ldquoBiological activities ofPereskia bleo extractsrdquo International Journal of Pharmacologyvol 5 no 1 pp 71ndash75 2009
[18] E Bossy-Wetzel and D R Green ldquoDetection of apoptosis byannexin V labelingrdquo Methods in Enzymology vol 322 pp 15ndash18 2000
[19] H Wang and J A Joseph ldquoQuantifying cellular oxidative stressby dichlorofluorescein assay usingmicroplate readerrdquo Free Rad-ical Biology and Medicine vol 27 no 5-6 pp 612ndash616 1999
[20] S C Cheah D R Appleton S T Lee M L Lam A H AHadi and M R Mustafa ldquoPanduratin a inhibits the growth ofA549 cells through induction of apoptosis and inhibition of NF-KappaB translocationrdquoMolecules vol 16 no 3 pp 2583ndash25982011
[21] L Foghsgaard DWissing DMauch et al ldquoCathepsin B acts asa dominant execution protease in tumor cell apoptosis inducedby tumor necrosis factorrdquo Journal of Cell Biology vol 153 no 5pp 999ndash1009 2001
[22] K Kagedal U Johansson and K Ollinger ldquoThe lysosomalprotease cathepsin D mediates apoptosis induced by oxidativestressrdquoThe FASEB journal vol 15 no 9 pp 1592ndash1594 2001
[23] M A Birch-Machin and H Swalwell ldquoHow mitochondriarecord the effects of UV exposure and oxidative stress usinghuman skin as a model tissuerdquo Mutagenesis vol 25 no 2 pp101ndash107 2010
[24] S Meister B Frey V R Lang et al ldquoCalcium channel blockerverapamil enhances endoplasmic reticulum stress and cell deathinduced by proteasome inhibition in myeloma cellsrdquoNeoplasiavol 12 no 7 pp 550ndash561 2010
[25] S Mohan A B Abdul S I Abdelwahab et al ldquoTyphoniumflagelliforme induces apoptosis in CEMss cells via activation
of caspase-9 PARP cleavage and cytochrome c release itsactivation coupled with G0G1 phase cell cycle arrestrdquo Journalof Ethnopharmacology vol 131 no 3 pp 592ndash600 2010
[26] S Elmore ldquoApoptosis a review of programmed cell deathrdquoToxicologic Pathology vol 35 no 4 pp 495ndash516 2007
[27] M Aberg M Johnell M Wickstrom and A Siegbahn ldquoTissueFactor FVIIa prevents the extrinsic pathway of apoptosis byregulation of the tumor suppressor Death-Associated ProteinKinase 1 (DAPK1)rdquoThrombosis Research vol 127 no 2 pp 141ndash148 2011
[28] M Nigam N Singh V Ranjan et al ldquoCentchroman mediatedapoptosis involves cross-talk between extrinsicintrinsic path-ways and oxidative regulationrdquo Life Sciences vol 87 no 23ndash26pp 750ndash758 2010
[29] O S Frankfurt and A Krishan ldquoApoptosis-based drug screen-ing and detection of selective toxicity to cancer cellsrdquo Anti-Cancer Drugs vol 14 no 7 pp 555ndash561 2003
[30] M van Engeland L J W Nieland F C S Ramaekers BSchutte and C P M Reutelingsperger ldquoAnnexin V-affinityassay a review on an apoptosis detection system based onphosphatidylserine exposurerdquo Cytometry vol 31 no 1 pp 1ndash91998
[31] G A Joanitti R B Azevedo and S M Freitas ldquoApoptosisand lysosome membrane permeabilization induction on breastcancer cells by an anticarcinogenic Bowman-Birk proteaseinhibitor from Vigna unguiculata seedsrdquo Cancer Letters vol293 no 1 pp 73ndash81 2010
[32] T Kirkegaard and M Jaattela ldquoLysosomal involvement in celldeath and cancerrdquo Biochimica et Biophysica ActamdashMolecularCell Research vol 1793 no 4 pp 746ndash754 2009
[33] A C Johansson H Appelqvist C Nilsson K Kagedal KRoberg and K Ollinger ldquoRegulation of apoptosis-associatedlysosomal membrane permeabilizationrdquo Apoptosis vol 15 no5 pp 527ndash540 2010
[34] J Wen K R You S Y Lee C H Song and D G KimldquoOxidative stress-mediated apoptosis the anticancer effect ofthe sesquiterpene lactone parthenoliderdquo Journal of BiologicalChemistry vol 277 no 41 pp 38954ndash38964 2002
[35] C Fleury B Mignotte and J L Vayssiere ldquoMitochondrialreactive oxygen species in cell death signalingrdquo Biochimie vol84 no 2-3 pp 131ndash141 2002
[36] A M Verhagen P G Ekert M Pakusch et al ldquoIdentificationof DIABLO a mammalian protein that promotes apoptosis bybinding to and antagonizing IAP proteinsrdquo Cell vol 102 no 1pp 43ndash53 2000
[37] N Ajenjo E Canonon I Sanchez-Perez et al ldquoSubcellularlocalization determines the protective effects of activated ERK2against distinct apoptogenic stimuli in myeloid leukemia cellsrdquoJournal of Biological Chemistry vol 279 no 31 pp 32813ndash328232004
[38] X Wang ldquoThe expanding role of mitochondria in apoptosisrdquoGenes and Development vol 15 no 22 pp 2922ndash2933 2001
[39] Y A Hannun ldquoApoptosis and the dilemma of cancer chemo-therapyrdquo Blood vol 89 no 6 pp 1845ndash1853 1997
[40] F L Scott J B Denault S J Riedl H Shin M Renatusand G S Salvesen ldquoXIAP inhibits caspase-3 and -7 usingtwo binding sites evolutionary conserved mechanism of IAPsrdquoEMBO Journal vol 24 no 3 pp 645ndash655 2005
[41] S Cory and J M Adams ldquoThe BCL2 family regulators of thecellular life-or-death switchrdquo Nature Reviews Cancer vol 2 no9 pp 647ndash656 2002
12 Evidence-Based Complementary and Alternative Medicine
[42] S B Bratton and GM Cohen ldquoDeath receptors leave a caspasefootprint that Smacs of XIAPrdquo Cell Death and Differentiationvol 10 no 1 pp 4ndash6 2003
[43] N Zamzami andGKroemer ldquoThemitochondrion in apoptosishow Pandorarsquos box opensrdquo Nature Reviews Molecular CellBiology vol 2 no 1 pp 67ndash71 2001
[44] I Marzo C Brenner N Zamzami et al ldquoBax and adeninenucleotide translocator cooperate in the mitochondrial controlof apoptosisrdquo Science vol 281 no 5385 pp 2027ndash2031 1998
[45] M D Esposti ldquoThe roles of BidrdquoApoptosis vol 7 no 5 pp 433ndash440 2002
[46] N E Allen A W Roddam D S Allen et al ldquoA prospectivestudy of serum insulin-like growth factor-I (IGF-I) IGF-IIIGF-binding protein-3 and breast cancer riskrdquo British Journalof Cancer vol 92 no 7 pp 1283ndash1287 2005
[47] L Buckbinder R Talbott S Velasco-Miguel et al ldquoInductionof the growth inhibitor IGF-binding protein 3 by p53rdquo Naturevol 377 no 6550 pp 646ndash649 1995
[48] E Ruoslahti ldquoFibronectin and its integrin receptors in cancerrdquoAdvances in Cancer Research vol 76 pp 18ndash20 1999
[49] B S Schutt M Langkamp U Rauschnabel M B Ranke andM W Elmlinger ldquoIntegrin-mediated action of insulin-likefactor binding protein-2 in tumor cellsrdquo Journal of MolecularEndocrinology vol 32 no 3 pp 859ndash868 2004
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 9
bad
bax
bcl-2
bcl-w
BID BI
MCa
spas
e 3Ca
spas
e 8IG
F-II IG
F-I HTR
AH
SP70
HSP
60H
SP27
FasL Fa
sD
R6Cy
to c
IGFB
P-1
IGFB
P-2 IG
FBP-
3IG
FBP-
4IG
FBP-
5IG
FBP-
6IG
F-1s
RLi
vin p
21 p
27 p53
SM
AC S
urvi
vin
sTN
F-R1 XI
AP
minus3
minus2
minus1
0
1
2
3
Relat
ive d
ensit
y di
ffere
nce t
o co
ntro
l (times10
5)
(a)
(b) (c)
A B C D E F G H I J K L M N1 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 82 Pos Pos Neg Neg Blank Blank bad bax bcl-2 bcl-w BID BIM Caspase 3 Caspase 83 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II4 CD40 CD40L cIAP-2 Cyto c DR6 Fas FasL Blank HSP27 HSP60 HSP70 HTRA IGF-I IGF-II5 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R16 IGFBP1 IGFBP2 IGFBP3 IGFBP4 IGFBP5 IGFBP6 IGF-1sR Livin p21 p27 p53 SMAC Survivin sTNF-R17 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg Pos8 sTNF-R2 TRAIL-1 TRAIL-2 TRAIL-3 TRAIL-4 XIAP Blank Blank Neg Neg Neg PosTNF-120572
TNF-120572 TNF-120573TNF-120573
(d)
Figure 7 Cells were lysed and protein arrays were performed Cells was treated with 19120583M girinimbine for 24 h and the whole cell proteinwas extracted Equal amount of (300 120583g) of protein from each sample was used for the assay Quantitative analysis in the arrays showeddifferences in the apoptotic markers (a) Representative images of the apoptotic protein array are shown for the control (b) treatment (c) andthe exact protein name of each dot in the array (d)
Suppression of apoptosis may promote the cancer develop-ment by inhibiting the chemotherapy as well as other forms ofinducers of cell death [39] Regulation of apoptosis has beencensoriously dependent on several genes which have beenidentified earlier and includes XIAP-a member of the IAPfamily It has the capacity to inhibit the activation of caspases3 7 and 9 [40] In our study we found that girinimbinetreatment of A549 cells caused inhibition of XIAP but SMAC
an antagonist to XIAP could be found to be slightly regulatedin this treatment concentration This is considered as a keyevent in the execution of cell death which is regulatedmainlyby proteins of the Bcl-2 family [41 42]
Critical issues in apoptosis include the dominance ofanti-versus proapoptotic Bcl-2 members [43] Girinimbine isfound to induce apoptosis in the A549 cells involvingdownregulated Bcl-2 expression This downregulation was
10 Evidence-Based Complementary and Alternative Medicine
3
2
1
0
times105
19 9 475 0 19 9 475 0 19 9 475 0Girinimbine treatment (120583M)
Lum
ines
cenc
e
Caspase 37Caspase 8Caspase 9
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast
Figure 8 Effect of girinimbine on activation of caspases 37 8 and 9Cells were treated with girinimbine at the indicated concentrationsfor 24 h and activity was determined by using a luminescentenzyme assay kit Each point represents the mean plusmn SD of threedifferent experiments Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software(version 40 GraphPad Software Inc San Diego CA) Statisticalsignificance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
concomitant with the upregulation of Bad Bax and Bimwhich inhibits antiapoptotic proteins and render the cellsmore susceptible to apoptogenic stimuli (Figure 7) Therelease of cytochrome c found in our study could be due tothe up regulated Bax since Bax in association with adeninenucleotide translocator within the permeability transitionpore complex will increase the mitochondrial membranepermeability and thereby discharge a number of apopto-genic molecules into the cytosol [44] In addition to thisupregulation of the tumor suppressor protein p53 which hasa critical role in regulation of the Bcl-2 and p27 and p21draws attention to the involvement of cell cycle arrest in theapoptosis process Alternatively we found the involvement ofextrinsic pathway proteins such as Fas and FasL upregulationand related upregulation of caspase 8 (Figure 7) Moreoverthe luminescent based caspase 8 assay had showed 10 folddifferences than the control Apart from this we foundthat there is a downregulation of Bid happening too Sincemitochondrial damage in the Fas pathway of apoptosis ismediated by the caspase 8 cleavage of Bid [45] the findingshere shows the cross talk between intrinsic and extrinsicpathways
The protein array results exhibited a significant reductionof both IGF-I and IGF-II IGF-I has been shown to exertstrongmitogenic and antiapoptotic effects in a variety of nor-mal and cancerous cells including lung cancer cell lines [46]Moreover the over expression of IGFBP3 is closely associatedwith the downregulation of IGF-I and its mitogenic activity[47] It is known that IGFBP-2 and IGFBP-1 are involved inthe regulation of cell migration apoptosis and cell growth
[48] Both are reported to be markedly overexpressed inmany tumors and tumor cell lines And it is assumed thatelevation of the IGFBP-2 production is part of a mechanismto compensate for the mitogenic and antiapoptotic effectsof tumor-derived IGFs [49] Our data clearly showed thedownregulation of IGFBP1 and IGFBP2 concomitant withupregulation of IGBP3
Our results provided a new insight into the mechanismof chemotherapeutic properties of Murraya koenigii Inaddition these study findings demonstrate that girinimbinemediates its apoptotic effects through both intrinsic andextrinsic pathway which is depended on caspase mediationMoreover the upregulation of p53 as well as the cell prolifer-ation repressor proteins p27 and p21 and the significant roleof insulinIGF-1 signaling were also identified These resultsindicate that girinimbine could be a candidate for a novelanticancer agentThereforemore in-depth in vitro and in vivostudies are currently going on in our laboratory
Conflict of Interests
The authors declare that they do not have any conflict ofinterests
Acknowledgment
The authors greatly acknowledge the financial support pro-vided by University of Malaya (Grant no HIR E0000002-20001)
References
[1] J K Field and S W Duffy ldquoLung cancer screening the wayforwardrdquo British Journal of Cancer vol 99 no 4 pp 557ndash5622008
[2] K A Yoon J H Park J Han et al ldquoA genome-wide associationstudy reveals susceptibility variants for nonsmall cell lungcancer in the Korean populationrdquo Human Molecular Geneticsvol 19 no 24 Article ID ddq421 pp 4948ndash4954 2010
[3] G Cartei F Cartei A Cantone et al ldquoCisplatin-cyclophos-phamide-mitomycin combination chemotherapy with support-ive care versus supportive care alone for treatment of metastaticnon- small-cell lung cancerrdquo Journal of the National CancerInstitute vol 85 no 10 pp 794ndash800 1993
[4] M O Hengartner ldquoThe biochemistry of apoptosisrdquoNature vol407 no 6805 pp 770ndash776 2000
[5] DHanahan andRAWeinberg ldquoThehallmarks of cancerrdquoCellvol 100 no 1 pp 57ndash70 2000
[6] J R Molina P Yang S D Cassivi S E Schild and A AAdjei ldquoNon-small cell lung cancer epidemiology risk factorstreatment and survivorshiprdquo Mayo Clinic Proceedings vol 83no 5 pp 584ndash594 2008
[7] S Diwanay D Chitre and B Patwardhan ldquoImmunoprotec-tion by botanical drugs in cancer chemotherapyrdquo Journal ofEthnopharmacology vol 90 no 1 pp 49ndash55 2004
[8] U M Thatte and S A Dahanukar ldquoRasayana Concept cluesfrom immunomodulatory therapyrdquo in An Annotated Bibliogra-phy of Indian Medicine S N Upadhyay Ed pp 141ndash148 1997
[9] A S Shah A S Wakade and A R Juvekar ldquoImmunomod-ulatory activity of methanolic extract of Murraya koenigii (L)
Evidence-Based Complementary and Alternative Medicine 11
Spreng leavesrdquo Indian Journal of Experimental Biology vol 46no 7 pp 505ndash509 2008
[10] R B Goswamia P K S Singha N Goswamib P Thomasd PU Devic and A K Pathakd ldquoStudies on antigenotoxic effect ofMurraya koenigii leavesrdquo International Journal of PharmaRecentResearch vol 2 pp 65ndash68 2010
[11] R S Ramsewak M G Nair G M Strasburg D L DeWittand J L Nitiss ldquoBiologically active carbazole alkaloids fromMurraya koenigiirdquo Journal of Agricultural and Food Chemistryvol 47 no 2 pp 444ndash447 1999
[12] D P Chakraborty B K Barman and P K Bose ldquoStructureof girinimbine a pyranocarbazole derivative isolated fromMurraya koenigii Sprengrdquo Scientific Culture vol 30 pp 445ndash448 1964
[13] L J M Rao K Ramalakshmi B B Borse and B RaghavanldquoAntioxidant and radical-scavenging carbazole alkaloids fromthe oleoresin of curry leaf (Murraya koenigii Spreng)rdquo FoodChemistry vol 100 no 2 pp 742ndash747 2007
[14] S L Wang B Cai C B Cui S Y Yan and C F Wu ldquoInductionof apoptosis by girinimbine in K562 cellrdquo Chinese Traditionaland Herbal Drugs vol 38 no 11 pp 1677ndash1681 2007
[15] S Syam A B Abdul M A Sukari S Mohan S I Abdelwahaband T S Wah ldquoThe growth suppressing effects of girinimbineon Hepg2 involve induction of apoptosis and cell cycle arrestrdquoMolecules vol 16 pp 7155ndash7170 2011
[16] A Bakar N Haslizawati M A Sukari et al ldquoChemical con-stituents from stem barks and roots of Murraya koenigii(Rutaceae)rdquo The Malaysian Journal of Analytical Sciences vol11 pp 173ndash176 2007
[17] S I A Wahab A B Abdul S M Mohan A S Al-ZubairiM M Elhassan and M Y Ibrahim ldquoBiological activities ofPereskia bleo extractsrdquo International Journal of Pharmacologyvol 5 no 1 pp 71ndash75 2009
[18] E Bossy-Wetzel and D R Green ldquoDetection of apoptosis byannexin V labelingrdquo Methods in Enzymology vol 322 pp 15ndash18 2000
[19] H Wang and J A Joseph ldquoQuantifying cellular oxidative stressby dichlorofluorescein assay usingmicroplate readerrdquo Free Rad-ical Biology and Medicine vol 27 no 5-6 pp 612ndash616 1999
[20] S C Cheah D R Appleton S T Lee M L Lam A H AHadi and M R Mustafa ldquoPanduratin a inhibits the growth ofA549 cells through induction of apoptosis and inhibition of NF-KappaB translocationrdquoMolecules vol 16 no 3 pp 2583ndash25982011
[21] L Foghsgaard DWissing DMauch et al ldquoCathepsin B acts asa dominant execution protease in tumor cell apoptosis inducedby tumor necrosis factorrdquo Journal of Cell Biology vol 153 no 5pp 999ndash1009 2001
[22] K Kagedal U Johansson and K Ollinger ldquoThe lysosomalprotease cathepsin D mediates apoptosis induced by oxidativestressrdquoThe FASEB journal vol 15 no 9 pp 1592ndash1594 2001
[23] M A Birch-Machin and H Swalwell ldquoHow mitochondriarecord the effects of UV exposure and oxidative stress usinghuman skin as a model tissuerdquo Mutagenesis vol 25 no 2 pp101ndash107 2010
[24] S Meister B Frey V R Lang et al ldquoCalcium channel blockerverapamil enhances endoplasmic reticulum stress and cell deathinduced by proteasome inhibition in myeloma cellsrdquoNeoplasiavol 12 no 7 pp 550ndash561 2010
[25] S Mohan A B Abdul S I Abdelwahab et al ldquoTyphoniumflagelliforme induces apoptosis in CEMss cells via activation
of caspase-9 PARP cleavage and cytochrome c release itsactivation coupled with G0G1 phase cell cycle arrestrdquo Journalof Ethnopharmacology vol 131 no 3 pp 592ndash600 2010
[26] S Elmore ldquoApoptosis a review of programmed cell deathrdquoToxicologic Pathology vol 35 no 4 pp 495ndash516 2007
[27] M Aberg M Johnell M Wickstrom and A Siegbahn ldquoTissueFactor FVIIa prevents the extrinsic pathway of apoptosis byregulation of the tumor suppressor Death-Associated ProteinKinase 1 (DAPK1)rdquoThrombosis Research vol 127 no 2 pp 141ndash148 2011
[28] M Nigam N Singh V Ranjan et al ldquoCentchroman mediatedapoptosis involves cross-talk between extrinsicintrinsic path-ways and oxidative regulationrdquo Life Sciences vol 87 no 23ndash26pp 750ndash758 2010
[29] O S Frankfurt and A Krishan ldquoApoptosis-based drug screen-ing and detection of selective toxicity to cancer cellsrdquo Anti-Cancer Drugs vol 14 no 7 pp 555ndash561 2003
[30] M van Engeland L J W Nieland F C S Ramaekers BSchutte and C P M Reutelingsperger ldquoAnnexin V-affinityassay a review on an apoptosis detection system based onphosphatidylserine exposurerdquo Cytometry vol 31 no 1 pp 1ndash91998
[31] G A Joanitti R B Azevedo and S M Freitas ldquoApoptosisand lysosome membrane permeabilization induction on breastcancer cells by an anticarcinogenic Bowman-Birk proteaseinhibitor from Vigna unguiculata seedsrdquo Cancer Letters vol293 no 1 pp 73ndash81 2010
[32] T Kirkegaard and M Jaattela ldquoLysosomal involvement in celldeath and cancerrdquo Biochimica et Biophysica ActamdashMolecularCell Research vol 1793 no 4 pp 746ndash754 2009
[33] A C Johansson H Appelqvist C Nilsson K Kagedal KRoberg and K Ollinger ldquoRegulation of apoptosis-associatedlysosomal membrane permeabilizationrdquo Apoptosis vol 15 no5 pp 527ndash540 2010
[34] J Wen K R You S Y Lee C H Song and D G KimldquoOxidative stress-mediated apoptosis the anticancer effect ofthe sesquiterpene lactone parthenoliderdquo Journal of BiologicalChemistry vol 277 no 41 pp 38954ndash38964 2002
[35] C Fleury B Mignotte and J L Vayssiere ldquoMitochondrialreactive oxygen species in cell death signalingrdquo Biochimie vol84 no 2-3 pp 131ndash141 2002
[36] A M Verhagen P G Ekert M Pakusch et al ldquoIdentificationof DIABLO a mammalian protein that promotes apoptosis bybinding to and antagonizing IAP proteinsrdquo Cell vol 102 no 1pp 43ndash53 2000
[37] N Ajenjo E Canonon I Sanchez-Perez et al ldquoSubcellularlocalization determines the protective effects of activated ERK2against distinct apoptogenic stimuli in myeloid leukemia cellsrdquoJournal of Biological Chemistry vol 279 no 31 pp 32813ndash328232004
[38] X Wang ldquoThe expanding role of mitochondria in apoptosisrdquoGenes and Development vol 15 no 22 pp 2922ndash2933 2001
[39] Y A Hannun ldquoApoptosis and the dilemma of cancer chemo-therapyrdquo Blood vol 89 no 6 pp 1845ndash1853 1997
[40] F L Scott J B Denault S J Riedl H Shin M Renatusand G S Salvesen ldquoXIAP inhibits caspase-3 and -7 usingtwo binding sites evolutionary conserved mechanism of IAPsrdquoEMBO Journal vol 24 no 3 pp 645ndash655 2005
[41] S Cory and J M Adams ldquoThe BCL2 family regulators of thecellular life-or-death switchrdquo Nature Reviews Cancer vol 2 no9 pp 647ndash656 2002
12 Evidence-Based Complementary and Alternative Medicine
[42] S B Bratton and GM Cohen ldquoDeath receptors leave a caspasefootprint that Smacs of XIAPrdquo Cell Death and Differentiationvol 10 no 1 pp 4ndash6 2003
[43] N Zamzami andGKroemer ldquoThemitochondrion in apoptosishow Pandorarsquos box opensrdquo Nature Reviews Molecular CellBiology vol 2 no 1 pp 67ndash71 2001
[44] I Marzo C Brenner N Zamzami et al ldquoBax and adeninenucleotide translocator cooperate in the mitochondrial controlof apoptosisrdquo Science vol 281 no 5385 pp 2027ndash2031 1998
[45] M D Esposti ldquoThe roles of BidrdquoApoptosis vol 7 no 5 pp 433ndash440 2002
[46] N E Allen A W Roddam D S Allen et al ldquoA prospectivestudy of serum insulin-like growth factor-I (IGF-I) IGF-IIIGF-binding protein-3 and breast cancer riskrdquo British Journalof Cancer vol 92 no 7 pp 1283ndash1287 2005
[47] L Buckbinder R Talbott S Velasco-Miguel et al ldquoInductionof the growth inhibitor IGF-binding protein 3 by p53rdquo Naturevol 377 no 6550 pp 646ndash649 1995
[48] E Ruoslahti ldquoFibronectin and its integrin receptors in cancerrdquoAdvances in Cancer Research vol 76 pp 18ndash20 1999
[49] B S Schutt M Langkamp U Rauschnabel M B Ranke andM W Elmlinger ldquoIntegrin-mediated action of insulin-likefactor binding protein-2 in tumor cellsrdquo Journal of MolecularEndocrinology vol 32 no 3 pp 859ndash868 2004
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Evidence-Based Complementary and Alternative Medicine
3
2
1
0
times105
19 9 475 0 19 9 475 0 19 9 475 0Girinimbine treatment (120583M)
Lum
ines
cenc
e
Caspase 37Caspase 8Caspase 9
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowastlowast
lowast
Figure 8 Effect of girinimbine on activation of caspases 37 8 and 9Cells were treated with girinimbine at the indicated concentrationsfor 24 h and activity was determined by using a luminescentenzyme assay kit Each point represents the mean plusmn SD of threedifferent experiments Statistical analysis was performed with one-way analysis of variance (ANOVA) using GraphPad Prism software(version 40 GraphPad Software Inc San Diego CA) Statisticalsignificance is expressed as lowastlowast119875 lt 001 lowast119875 lt 005
concomitant with the upregulation of Bad Bax and Bimwhich inhibits antiapoptotic proteins and render the cellsmore susceptible to apoptogenic stimuli (Figure 7) Therelease of cytochrome c found in our study could be due tothe up regulated Bax since Bax in association with adeninenucleotide translocator within the permeability transitionpore complex will increase the mitochondrial membranepermeability and thereby discharge a number of apopto-genic molecules into the cytosol [44] In addition to thisupregulation of the tumor suppressor protein p53 which hasa critical role in regulation of the Bcl-2 and p27 and p21draws attention to the involvement of cell cycle arrest in theapoptosis process Alternatively we found the involvement ofextrinsic pathway proteins such as Fas and FasL upregulationand related upregulation of caspase 8 (Figure 7) Moreoverthe luminescent based caspase 8 assay had showed 10 folddifferences than the control Apart from this we foundthat there is a downregulation of Bid happening too Sincemitochondrial damage in the Fas pathway of apoptosis ismediated by the caspase 8 cleavage of Bid [45] the findingshere shows the cross talk between intrinsic and extrinsicpathways
The protein array results exhibited a significant reductionof both IGF-I and IGF-II IGF-I has been shown to exertstrongmitogenic and antiapoptotic effects in a variety of nor-mal and cancerous cells including lung cancer cell lines [46]Moreover the over expression of IGFBP3 is closely associatedwith the downregulation of IGF-I and its mitogenic activity[47] It is known that IGFBP-2 and IGFBP-1 are involved inthe regulation of cell migration apoptosis and cell growth
[48] Both are reported to be markedly overexpressed inmany tumors and tumor cell lines And it is assumed thatelevation of the IGFBP-2 production is part of a mechanismto compensate for the mitogenic and antiapoptotic effectsof tumor-derived IGFs [49] Our data clearly showed thedownregulation of IGFBP1 and IGFBP2 concomitant withupregulation of IGBP3
Our results provided a new insight into the mechanismof chemotherapeutic properties of Murraya koenigii Inaddition these study findings demonstrate that girinimbinemediates its apoptotic effects through both intrinsic andextrinsic pathway which is depended on caspase mediationMoreover the upregulation of p53 as well as the cell prolifer-ation repressor proteins p27 and p21 and the significant roleof insulinIGF-1 signaling were also identified These resultsindicate that girinimbine could be a candidate for a novelanticancer agentThereforemore in-depth in vitro and in vivostudies are currently going on in our laboratory
Conflict of Interests
The authors declare that they do not have any conflict ofinterests
Acknowledgment
The authors greatly acknowledge the financial support pro-vided by University of Malaya (Grant no HIR E0000002-20001)
References
[1] J K Field and S W Duffy ldquoLung cancer screening the wayforwardrdquo British Journal of Cancer vol 99 no 4 pp 557ndash5622008
[2] K A Yoon J H Park J Han et al ldquoA genome-wide associationstudy reveals susceptibility variants for nonsmall cell lungcancer in the Korean populationrdquo Human Molecular Geneticsvol 19 no 24 Article ID ddq421 pp 4948ndash4954 2010
[3] G Cartei F Cartei A Cantone et al ldquoCisplatin-cyclophos-phamide-mitomycin combination chemotherapy with support-ive care versus supportive care alone for treatment of metastaticnon- small-cell lung cancerrdquo Journal of the National CancerInstitute vol 85 no 10 pp 794ndash800 1993
[4] M O Hengartner ldquoThe biochemistry of apoptosisrdquoNature vol407 no 6805 pp 770ndash776 2000
[5] DHanahan andRAWeinberg ldquoThehallmarks of cancerrdquoCellvol 100 no 1 pp 57ndash70 2000
[6] J R Molina P Yang S D Cassivi S E Schild and A AAdjei ldquoNon-small cell lung cancer epidemiology risk factorstreatment and survivorshiprdquo Mayo Clinic Proceedings vol 83no 5 pp 584ndash594 2008
[7] S Diwanay D Chitre and B Patwardhan ldquoImmunoprotec-tion by botanical drugs in cancer chemotherapyrdquo Journal ofEthnopharmacology vol 90 no 1 pp 49ndash55 2004
[8] U M Thatte and S A Dahanukar ldquoRasayana Concept cluesfrom immunomodulatory therapyrdquo in An Annotated Bibliogra-phy of Indian Medicine S N Upadhyay Ed pp 141ndash148 1997
[9] A S Shah A S Wakade and A R Juvekar ldquoImmunomod-ulatory activity of methanolic extract of Murraya koenigii (L)
Evidence-Based Complementary and Alternative Medicine 11
Spreng leavesrdquo Indian Journal of Experimental Biology vol 46no 7 pp 505ndash509 2008
[10] R B Goswamia P K S Singha N Goswamib P Thomasd PU Devic and A K Pathakd ldquoStudies on antigenotoxic effect ofMurraya koenigii leavesrdquo International Journal of PharmaRecentResearch vol 2 pp 65ndash68 2010
[11] R S Ramsewak M G Nair G M Strasburg D L DeWittand J L Nitiss ldquoBiologically active carbazole alkaloids fromMurraya koenigiirdquo Journal of Agricultural and Food Chemistryvol 47 no 2 pp 444ndash447 1999
[12] D P Chakraborty B K Barman and P K Bose ldquoStructureof girinimbine a pyranocarbazole derivative isolated fromMurraya koenigii Sprengrdquo Scientific Culture vol 30 pp 445ndash448 1964
[13] L J M Rao K Ramalakshmi B B Borse and B RaghavanldquoAntioxidant and radical-scavenging carbazole alkaloids fromthe oleoresin of curry leaf (Murraya koenigii Spreng)rdquo FoodChemistry vol 100 no 2 pp 742ndash747 2007
[14] S L Wang B Cai C B Cui S Y Yan and C F Wu ldquoInductionof apoptosis by girinimbine in K562 cellrdquo Chinese Traditionaland Herbal Drugs vol 38 no 11 pp 1677ndash1681 2007
[15] S Syam A B Abdul M A Sukari S Mohan S I Abdelwahaband T S Wah ldquoThe growth suppressing effects of girinimbineon Hepg2 involve induction of apoptosis and cell cycle arrestrdquoMolecules vol 16 pp 7155ndash7170 2011
[16] A Bakar N Haslizawati M A Sukari et al ldquoChemical con-stituents from stem barks and roots of Murraya koenigii(Rutaceae)rdquo The Malaysian Journal of Analytical Sciences vol11 pp 173ndash176 2007
[17] S I A Wahab A B Abdul S M Mohan A S Al-ZubairiM M Elhassan and M Y Ibrahim ldquoBiological activities ofPereskia bleo extractsrdquo International Journal of Pharmacologyvol 5 no 1 pp 71ndash75 2009
[18] E Bossy-Wetzel and D R Green ldquoDetection of apoptosis byannexin V labelingrdquo Methods in Enzymology vol 322 pp 15ndash18 2000
[19] H Wang and J A Joseph ldquoQuantifying cellular oxidative stressby dichlorofluorescein assay usingmicroplate readerrdquo Free Rad-ical Biology and Medicine vol 27 no 5-6 pp 612ndash616 1999
[20] S C Cheah D R Appleton S T Lee M L Lam A H AHadi and M R Mustafa ldquoPanduratin a inhibits the growth ofA549 cells through induction of apoptosis and inhibition of NF-KappaB translocationrdquoMolecules vol 16 no 3 pp 2583ndash25982011
[21] L Foghsgaard DWissing DMauch et al ldquoCathepsin B acts asa dominant execution protease in tumor cell apoptosis inducedby tumor necrosis factorrdquo Journal of Cell Biology vol 153 no 5pp 999ndash1009 2001
[22] K Kagedal U Johansson and K Ollinger ldquoThe lysosomalprotease cathepsin D mediates apoptosis induced by oxidativestressrdquoThe FASEB journal vol 15 no 9 pp 1592ndash1594 2001
[23] M A Birch-Machin and H Swalwell ldquoHow mitochondriarecord the effects of UV exposure and oxidative stress usinghuman skin as a model tissuerdquo Mutagenesis vol 25 no 2 pp101ndash107 2010
[24] S Meister B Frey V R Lang et al ldquoCalcium channel blockerverapamil enhances endoplasmic reticulum stress and cell deathinduced by proteasome inhibition in myeloma cellsrdquoNeoplasiavol 12 no 7 pp 550ndash561 2010
[25] S Mohan A B Abdul S I Abdelwahab et al ldquoTyphoniumflagelliforme induces apoptosis in CEMss cells via activation
of caspase-9 PARP cleavage and cytochrome c release itsactivation coupled with G0G1 phase cell cycle arrestrdquo Journalof Ethnopharmacology vol 131 no 3 pp 592ndash600 2010
[26] S Elmore ldquoApoptosis a review of programmed cell deathrdquoToxicologic Pathology vol 35 no 4 pp 495ndash516 2007
[27] M Aberg M Johnell M Wickstrom and A Siegbahn ldquoTissueFactor FVIIa prevents the extrinsic pathway of apoptosis byregulation of the tumor suppressor Death-Associated ProteinKinase 1 (DAPK1)rdquoThrombosis Research vol 127 no 2 pp 141ndash148 2011
[28] M Nigam N Singh V Ranjan et al ldquoCentchroman mediatedapoptosis involves cross-talk between extrinsicintrinsic path-ways and oxidative regulationrdquo Life Sciences vol 87 no 23ndash26pp 750ndash758 2010
[29] O S Frankfurt and A Krishan ldquoApoptosis-based drug screen-ing and detection of selective toxicity to cancer cellsrdquo Anti-Cancer Drugs vol 14 no 7 pp 555ndash561 2003
[30] M van Engeland L J W Nieland F C S Ramaekers BSchutte and C P M Reutelingsperger ldquoAnnexin V-affinityassay a review on an apoptosis detection system based onphosphatidylserine exposurerdquo Cytometry vol 31 no 1 pp 1ndash91998
[31] G A Joanitti R B Azevedo and S M Freitas ldquoApoptosisand lysosome membrane permeabilization induction on breastcancer cells by an anticarcinogenic Bowman-Birk proteaseinhibitor from Vigna unguiculata seedsrdquo Cancer Letters vol293 no 1 pp 73ndash81 2010
[32] T Kirkegaard and M Jaattela ldquoLysosomal involvement in celldeath and cancerrdquo Biochimica et Biophysica ActamdashMolecularCell Research vol 1793 no 4 pp 746ndash754 2009
[33] A C Johansson H Appelqvist C Nilsson K Kagedal KRoberg and K Ollinger ldquoRegulation of apoptosis-associatedlysosomal membrane permeabilizationrdquo Apoptosis vol 15 no5 pp 527ndash540 2010
[34] J Wen K R You S Y Lee C H Song and D G KimldquoOxidative stress-mediated apoptosis the anticancer effect ofthe sesquiterpene lactone parthenoliderdquo Journal of BiologicalChemistry vol 277 no 41 pp 38954ndash38964 2002
[35] C Fleury B Mignotte and J L Vayssiere ldquoMitochondrialreactive oxygen species in cell death signalingrdquo Biochimie vol84 no 2-3 pp 131ndash141 2002
[36] A M Verhagen P G Ekert M Pakusch et al ldquoIdentificationof DIABLO a mammalian protein that promotes apoptosis bybinding to and antagonizing IAP proteinsrdquo Cell vol 102 no 1pp 43ndash53 2000
[37] N Ajenjo E Canonon I Sanchez-Perez et al ldquoSubcellularlocalization determines the protective effects of activated ERK2against distinct apoptogenic stimuli in myeloid leukemia cellsrdquoJournal of Biological Chemistry vol 279 no 31 pp 32813ndash328232004
[38] X Wang ldquoThe expanding role of mitochondria in apoptosisrdquoGenes and Development vol 15 no 22 pp 2922ndash2933 2001
[39] Y A Hannun ldquoApoptosis and the dilemma of cancer chemo-therapyrdquo Blood vol 89 no 6 pp 1845ndash1853 1997
[40] F L Scott J B Denault S J Riedl H Shin M Renatusand G S Salvesen ldquoXIAP inhibits caspase-3 and -7 usingtwo binding sites evolutionary conserved mechanism of IAPsrdquoEMBO Journal vol 24 no 3 pp 645ndash655 2005
[41] S Cory and J M Adams ldquoThe BCL2 family regulators of thecellular life-or-death switchrdquo Nature Reviews Cancer vol 2 no9 pp 647ndash656 2002
12 Evidence-Based Complementary and Alternative Medicine
[42] S B Bratton and GM Cohen ldquoDeath receptors leave a caspasefootprint that Smacs of XIAPrdquo Cell Death and Differentiationvol 10 no 1 pp 4ndash6 2003
[43] N Zamzami andGKroemer ldquoThemitochondrion in apoptosishow Pandorarsquos box opensrdquo Nature Reviews Molecular CellBiology vol 2 no 1 pp 67ndash71 2001
[44] I Marzo C Brenner N Zamzami et al ldquoBax and adeninenucleotide translocator cooperate in the mitochondrial controlof apoptosisrdquo Science vol 281 no 5385 pp 2027ndash2031 1998
[45] M D Esposti ldquoThe roles of BidrdquoApoptosis vol 7 no 5 pp 433ndash440 2002
[46] N E Allen A W Roddam D S Allen et al ldquoA prospectivestudy of serum insulin-like growth factor-I (IGF-I) IGF-IIIGF-binding protein-3 and breast cancer riskrdquo British Journalof Cancer vol 92 no 7 pp 1283ndash1287 2005
[47] L Buckbinder R Talbott S Velasco-Miguel et al ldquoInductionof the growth inhibitor IGF-binding protein 3 by p53rdquo Naturevol 377 no 6550 pp 646ndash649 1995
[48] E Ruoslahti ldquoFibronectin and its integrin receptors in cancerrdquoAdvances in Cancer Research vol 76 pp 18ndash20 1999
[49] B S Schutt M Langkamp U Rauschnabel M B Ranke andM W Elmlinger ldquoIntegrin-mediated action of insulin-likefactor binding protein-2 in tumor cellsrdquo Journal of MolecularEndocrinology vol 32 no 3 pp 859ndash868 2004
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Evidence-Based Complementary and Alternative Medicine 11
Spreng leavesrdquo Indian Journal of Experimental Biology vol 46no 7 pp 505ndash509 2008
[10] R B Goswamia P K S Singha N Goswamib P Thomasd PU Devic and A K Pathakd ldquoStudies on antigenotoxic effect ofMurraya koenigii leavesrdquo International Journal of PharmaRecentResearch vol 2 pp 65ndash68 2010
[11] R S Ramsewak M G Nair G M Strasburg D L DeWittand J L Nitiss ldquoBiologically active carbazole alkaloids fromMurraya koenigiirdquo Journal of Agricultural and Food Chemistryvol 47 no 2 pp 444ndash447 1999
[12] D P Chakraborty B K Barman and P K Bose ldquoStructureof girinimbine a pyranocarbazole derivative isolated fromMurraya koenigii Sprengrdquo Scientific Culture vol 30 pp 445ndash448 1964
[13] L J M Rao K Ramalakshmi B B Borse and B RaghavanldquoAntioxidant and radical-scavenging carbazole alkaloids fromthe oleoresin of curry leaf (Murraya koenigii Spreng)rdquo FoodChemistry vol 100 no 2 pp 742ndash747 2007
[14] S L Wang B Cai C B Cui S Y Yan and C F Wu ldquoInductionof apoptosis by girinimbine in K562 cellrdquo Chinese Traditionaland Herbal Drugs vol 38 no 11 pp 1677ndash1681 2007
[15] S Syam A B Abdul M A Sukari S Mohan S I Abdelwahaband T S Wah ldquoThe growth suppressing effects of girinimbineon Hepg2 involve induction of apoptosis and cell cycle arrestrdquoMolecules vol 16 pp 7155ndash7170 2011
[16] A Bakar N Haslizawati M A Sukari et al ldquoChemical con-stituents from stem barks and roots of Murraya koenigii(Rutaceae)rdquo The Malaysian Journal of Analytical Sciences vol11 pp 173ndash176 2007
[17] S I A Wahab A B Abdul S M Mohan A S Al-ZubairiM M Elhassan and M Y Ibrahim ldquoBiological activities ofPereskia bleo extractsrdquo International Journal of Pharmacologyvol 5 no 1 pp 71ndash75 2009
[18] E Bossy-Wetzel and D R Green ldquoDetection of apoptosis byannexin V labelingrdquo Methods in Enzymology vol 322 pp 15ndash18 2000
[19] H Wang and J A Joseph ldquoQuantifying cellular oxidative stressby dichlorofluorescein assay usingmicroplate readerrdquo Free Rad-ical Biology and Medicine vol 27 no 5-6 pp 612ndash616 1999
[20] S C Cheah D R Appleton S T Lee M L Lam A H AHadi and M R Mustafa ldquoPanduratin a inhibits the growth ofA549 cells through induction of apoptosis and inhibition of NF-KappaB translocationrdquoMolecules vol 16 no 3 pp 2583ndash25982011
[21] L Foghsgaard DWissing DMauch et al ldquoCathepsin B acts asa dominant execution protease in tumor cell apoptosis inducedby tumor necrosis factorrdquo Journal of Cell Biology vol 153 no 5pp 999ndash1009 2001
[22] K Kagedal U Johansson and K Ollinger ldquoThe lysosomalprotease cathepsin D mediates apoptosis induced by oxidativestressrdquoThe FASEB journal vol 15 no 9 pp 1592ndash1594 2001
[23] M A Birch-Machin and H Swalwell ldquoHow mitochondriarecord the effects of UV exposure and oxidative stress usinghuman skin as a model tissuerdquo Mutagenesis vol 25 no 2 pp101ndash107 2010
[24] S Meister B Frey V R Lang et al ldquoCalcium channel blockerverapamil enhances endoplasmic reticulum stress and cell deathinduced by proteasome inhibition in myeloma cellsrdquoNeoplasiavol 12 no 7 pp 550ndash561 2010
[25] S Mohan A B Abdul S I Abdelwahab et al ldquoTyphoniumflagelliforme induces apoptosis in CEMss cells via activation
of caspase-9 PARP cleavage and cytochrome c release itsactivation coupled with G0G1 phase cell cycle arrestrdquo Journalof Ethnopharmacology vol 131 no 3 pp 592ndash600 2010
[26] S Elmore ldquoApoptosis a review of programmed cell deathrdquoToxicologic Pathology vol 35 no 4 pp 495ndash516 2007
[27] M Aberg M Johnell M Wickstrom and A Siegbahn ldquoTissueFactor FVIIa prevents the extrinsic pathway of apoptosis byregulation of the tumor suppressor Death-Associated ProteinKinase 1 (DAPK1)rdquoThrombosis Research vol 127 no 2 pp 141ndash148 2011
[28] M Nigam N Singh V Ranjan et al ldquoCentchroman mediatedapoptosis involves cross-talk between extrinsicintrinsic path-ways and oxidative regulationrdquo Life Sciences vol 87 no 23ndash26pp 750ndash758 2010
[29] O S Frankfurt and A Krishan ldquoApoptosis-based drug screen-ing and detection of selective toxicity to cancer cellsrdquo Anti-Cancer Drugs vol 14 no 7 pp 555ndash561 2003
[30] M van Engeland L J W Nieland F C S Ramaekers BSchutte and C P M Reutelingsperger ldquoAnnexin V-affinityassay a review on an apoptosis detection system based onphosphatidylserine exposurerdquo Cytometry vol 31 no 1 pp 1ndash91998
[31] G A Joanitti R B Azevedo and S M Freitas ldquoApoptosisand lysosome membrane permeabilization induction on breastcancer cells by an anticarcinogenic Bowman-Birk proteaseinhibitor from Vigna unguiculata seedsrdquo Cancer Letters vol293 no 1 pp 73ndash81 2010
[32] T Kirkegaard and M Jaattela ldquoLysosomal involvement in celldeath and cancerrdquo Biochimica et Biophysica ActamdashMolecularCell Research vol 1793 no 4 pp 746ndash754 2009
[33] A C Johansson H Appelqvist C Nilsson K Kagedal KRoberg and K Ollinger ldquoRegulation of apoptosis-associatedlysosomal membrane permeabilizationrdquo Apoptosis vol 15 no5 pp 527ndash540 2010
[34] J Wen K R You S Y Lee C H Song and D G KimldquoOxidative stress-mediated apoptosis the anticancer effect ofthe sesquiterpene lactone parthenoliderdquo Journal of BiologicalChemistry vol 277 no 41 pp 38954ndash38964 2002
[35] C Fleury B Mignotte and J L Vayssiere ldquoMitochondrialreactive oxygen species in cell death signalingrdquo Biochimie vol84 no 2-3 pp 131ndash141 2002
[36] A M Verhagen P G Ekert M Pakusch et al ldquoIdentificationof DIABLO a mammalian protein that promotes apoptosis bybinding to and antagonizing IAP proteinsrdquo Cell vol 102 no 1pp 43ndash53 2000
[37] N Ajenjo E Canonon I Sanchez-Perez et al ldquoSubcellularlocalization determines the protective effects of activated ERK2against distinct apoptogenic stimuli in myeloid leukemia cellsrdquoJournal of Biological Chemistry vol 279 no 31 pp 32813ndash328232004
[38] X Wang ldquoThe expanding role of mitochondria in apoptosisrdquoGenes and Development vol 15 no 22 pp 2922ndash2933 2001
[39] Y A Hannun ldquoApoptosis and the dilemma of cancer chemo-therapyrdquo Blood vol 89 no 6 pp 1845ndash1853 1997
[40] F L Scott J B Denault S J Riedl H Shin M Renatusand G S Salvesen ldquoXIAP inhibits caspase-3 and -7 usingtwo binding sites evolutionary conserved mechanism of IAPsrdquoEMBO Journal vol 24 no 3 pp 645ndash655 2005
[41] S Cory and J M Adams ldquoThe BCL2 family regulators of thecellular life-or-death switchrdquo Nature Reviews Cancer vol 2 no9 pp 647ndash656 2002
12 Evidence-Based Complementary and Alternative Medicine
[42] S B Bratton and GM Cohen ldquoDeath receptors leave a caspasefootprint that Smacs of XIAPrdquo Cell Death and Differentiationvol 10 no 1 pp 4ndash6 2003
[43] N Zamzami andGKroemer ldquoThemitochondrion in apoptosishow Pandorarsquos box opensrdquo Nature Reviews Molecular CellBiology vol 2 no 1 pp 67ndash71 2001
[44] I Marzo C Brenner N Zamzami et al ldquoBax and adeninenucleotide translocator cooperate in the mitochondrial controlof apoptosisrdquo Science vol 281 no 5385 pp 2027ndash2031 1998
[45] M D Esposti ldquoThe roles of BidrdquoApoptosis vol 7 no 5 pp 433ndash440 2002
[46] N E Allen A W Roddam D S Allen et al ldquoA prospectivestudy of serum insulin-like growth factor-I (IGF-I) IGF-IIIGF-binding protein-3 and breast cancer riskrdquo British Journalof Cancer vol 92 no 7 pp 1283ndash1287 2005
[47] L Buckbinder R Talbott S Velasco-Miguel et al ldquoInductionof the growth inhibitor IGF-binding protein 3 by p53rdquo Naturevol 377 no 6550 pp 646ndash649 1995
[48] E Ruoslahti ldquoFibronectin and its integrin receptors in cancerrdquoAdvances in Cancer Research vol 76 pp 18ndash20 1999
[49] B S Schutt M Langkamp U Rauschnabel M B Ranke andM W Elmlinger ldquoIntegrin-mediated action of insulin-likefactor binding protein-2 in tumor cellsrdquo Journal of MolecularEndocrinology vol 32 no 3 pp 859ndash868 2004
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 Evidence-Based Complementary and Alternative Medicine
[42] S B Bratton and GM Cohen ldquoDeath receptors leave a caspasefootprint that Smacs of XIAPrdquo Cell Death and Differentiationvol 10 no 1 pp 4ndash6 2003
[43] N Zamzami andGKroemer ldquoThemitochondrion in apoptosishow Pandorarsquos box opensrdquo Nature Reviews Molecular CellBiology vol 2 no 1 pp 67ndash71 2001
[44] I Marzo C Brenner N Zamzami et al ldquoBax and adeninenucleotide translocator cooperate in the mitochondrial controlof apoptosisrdquo Science vol 281 no 5385 pp 2027ndash2031 1998
[45] M D Esposti ldquoThe roles of BidrdquoApoptosis vol 7 no 5 pp 433ndash440 2002
[46] N E Allen A W Roddam D S Allen et al ldquoA prospectivestudy of serum insulin-like growth factor-I (IGF-I) IGF-IIIGF-binding protein-3 and breast cancer riskrdquo British Journalof Cancer vol 92 no 7 pp 1283ndash1287 2005
[47] L Buckbinder R Talbott S Velasco-Miguel et al ldquoInductionof the growth inhibitor IGF-binding protein 3 by p53rdquo Naturevol 377 no 6550 pp 646ndash649 1995
[48] E Ruoslahti ldquoFibronectin and its integrin receptors in cancerrdquoAdvances in Cancer Research vol 76 pp 18ndash20 1999
[49] B S Schutt M Langkamp U Rauschnabel M B Ranke andM W Elmlinger ldquoIntegrin-mediated action of insulin-likefactor binding protein-2 in tumor cellsrdquo Journal of MolecularEndocrinology vol 32 no 3 pp 859ndash868 2004
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom