relationships between the renal handling of dmps and …faculty.mercer.edu/zalups_rk/pdf manuscript...
TRANSCRIPT

Relationships between the Renal Handling of DMPS and DMSA andthe Renal Handling of MercuryRudolfs K. Zalups* and Christy C. Bridges
Division of Basic Medical Sciences, 1550 College Street, Mercer University School of Medicine, Macon, Georgia 31207, United States
ABSTRACT: Within the body of this review, we provide updates on the mechanismsinvolved in the renal handling mercury (Hg) and the vicinal dithiol complexing/chelatingagents, 2,3-bis(sulfanyl)propane-1-sulfonate (known formerly as 2,3-dimercaptopropane-1-sulfonate, DMPS) and meso-2,3-bis(sulfanyl)succinate (known formerly as meso-2,3-dimercaptosuccinate, DMSA), with a focus on the therapeutic effects of these dithiolsfollowing exposure to different chemical forms of Hg. We begin by reviewing briefly some ofthe chemical properties of Hg, with an emphasis on the high bonding affinity betweenmercuric ions and reduced sulfur atoms, principally those contained in protein andnonprotein thiols. A discussion is provided on the current body of knowledge pertaining tothe handling of various mercuric species within the kidneys, focusing on the primary cellulartargets that take up and are affected adversely by these species of Hg, namely, proximaltubular epithelial cells. Subsequently, we provide a brief update on the current knowledge onthe handling of DMPS and DMSA in the kidneys. In particular, parallels are drawn betweenthe mechanisms participating in the uptake of various thiol S-conjugates of Hg in proximaltubular cells and mechanisms by which DMPS and DMSA gain entry into these target epithelial cells. Finally, we discuss factorsthat permit DMPS and DMSA to bind intracellular mercuric ions and mechanisms transporting DMPS and DMSA S-conjugatesof Hg out of proximal tubular epithelial cells into the luminal compartment of the nephron, and promoting urinary excretion.
■ CONTENTS
Introduction 1825Human Exposure to Mercury 1826Binding of Mercury in Biological Systems 1826Kidney As a Primary Target of Mercuric Species 1827Proximal Tubular Uptake and Transport of Mercury 1827Luminal Uptake ofMercury by Proximal Tubular Cells 1828
Role of γ-Glutamyltransferase 1828Presence and Formation of Mercuric Conju-gates in the Proximal Tubular Lumen 1828Cleavage Products of GSH S-conjugates ofMercuric ions as Transportable Molecules 1828Role of Cysteinylglycinase in the LuminalUptake of Mercuric Species 1829Cys S-Conjugates of Mercury as PrimaryTransportable Substrates 1829Potential Role of Molecular Mimicry 1830
Basolateral Uptake of Mercury by Proximal TubularCells 1830
Role of Organic Anion Transporters 1830Role of the Basolateral Dicarboxylate Trans-porter NaC3 1831
Elimination of Mercuric Species by Proximal TubularCells 1831Handling of DMPS and DMSA in the Kidneys 1831Complexation of DMPS and DMSA with MercuricSpecies in Vivo 1832Renal Extraction of Mercuric Species Mediated byDMPS and DMSA 1832
Conclusions 1834Author Information 1834
Funding 1834Abbreviations 1834References 1835
■ INTRODUCTIONMercury (Hg) is designated as a transitional metal in row VI ofgroup 12 (IIB) in the periodic table of elements. Elementalmercury (Hg0) has an atomic mass of 200.59 g mol−1and isunique among metals in that it exists as a liquid at roomtemperature. It has a high vapor pressure that causes this elementto vaporize readily at 100 kPa (equivalent to one standardatmosphere according to the International Union of Pure andApplied Chemistry (IUPAC)). Accordingly, this propertyfacilitates entry of Hg0 to critical cellular targets in humans andother animals subsequent to exposure by inhalation. Once insystemic circulation, Hg0 gains access into a number of criticalsensitive target cells due to its nonpolar, lipophilic characteristics.Moreover, Hg0 undergoes oxidation in circulation and/or in theinternal compartments of target cells, forming highly reactivemercuric species (Hg2+).1
In the majority of environmental, occupational, and domesticsettings, Hg exists as a cation having an oxidation state of either1+ (mercurous) or 2+ (mercuric). In both aqueous and
Received: April 25, 2012Published: June 5, 2012
Review
pubs.acs.org/crt
© 2012 American Chemical Society 1825 dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−1838

terrestrial environments, mercuric ions do not exist as free,unbound cations. Rather, they are bonded to a host of inorganicand organic nucleophiles. Thiol-containing biological moleculeshave an extremely high affinity to bind mercuric ions. Mercuriccompounds are by far the primary species of Hg found inenvironment settings.2,3
In addition to having a 2+ oxidation state, the mercuric ion canexist in states having a valence of 1+ or 2+, depending on whetherit is bound covalently to a carbon atom (generally of low-molecular-weight organic chemical groups, such as methyl, ethyl,or phenyl groups). Of the known organic mercuric compounds,the methylmercuric (CH3Hg
+) form (generally referred to assimply methylmercury), when it is bound to a monovalent anionsuch as chloride, is the form most widely disseminated in theenvironment.2−4 The preponderance of methylmercuric speciesin environmental settings can be attributed largely to the actionsof aquatic and terrestrial prokaryotic organisms capable ofmethylating inorganic (mercuric) mercury (Hg2+) to CH3Hg
+.
■ HUMAN EXPOSURE TO MERCURY
Since inorganic and organic species of Hg are present or derivedfrom numerous sources in environmental, occupational, anddomestic settings, it is nearly impossible for the generalpopulation throughout the world to avoid exposure to low levelsof some form(s) of Hg on a regular basis. For example,individuals that have had dental amalgams, which are generallycomposed of at least 50%Hg0, implanted into occlusal surfaces oftheir teeth as a reconstructive treatment used to treat dentalcaries, can be exposed to varied amounts Hg0 vapor by inhalationand/or ingestion during the act of mastication or as a result ofbruxism.2,3,5−8 It is important to note that Hg0 is still beingutilized today by dentists in the United States.Risk of human exposure to elevated levels of various mercuric
species continues to be of considerable concern due to continuedand pervasive atmospheric deposition of Hg associated with theindustrial use of fossil fuels containing Hg.3,4,9 Exposure tomercuric species at levels that have well-documented toxic effectsin humans has been documented in environmental, industrial,medical, educational, governmental, and domestic settings.3,4,9
However, exposure to air, soil, water, and/or ingestion of variousanimal species contaminated with any of the toxic forms of Hgaccounts for the principal means by which Hg gains access intohumans and other animals.There are a number of additional situations in which the
general public can be exposed to elevated or toxic levels of Hg.These include breathing in air containing Hg vapor from spills ofmetallic Hg (such as from broken thermometers) or from areasin proximity to incinerators that burn fossil fuels (especially coal)containing Hg. Ingestion of fish (especially predatory species)contaminated with significant amounts of methylmercury isanother way that the general public can be exposed to elevated ortoxic levels of Hg, particularly if the fish are consumedfrequently.2,3,10,11 Careless use of certain antiseptics, disinfec-tants, and antifungal agents containing inorganic or organicforms of Hg is an additional means by which humans have beenexposed to Hg, although public access to medicinal and domesticchemicals containing mercuric compounds has been reducedgreatly due to federal and state regulations and statutespertaining to the use of such chemicals.
■ BINDING OF MERCURY IN BIOLOGICAL SYSTEMS
A significant body of evidence indicates that the biologicalactions and activities of mercurous and mercuric forms of Hg aredefined almost exclusively by the complex bonding reactions thatoccur between the ionic forms of Hg and the sulfur atom(s) ofsulfhydryl (-SH) groups (and thiolate anions) present in a host ofbiomolecules, especially low molecular weight thiol-containingmolecules, such as glutathione (GSH), DL-homocysteine (Hcy)and L-cysteine (Cys). Although mercuric ions can bind to anynucleophilic functional group of biological molecules, the affinityconstant for a mercuric ion binding to a reduced sulfur atom is onthe order of 1015 to 1020.2 In contrast, affinity constants for a Hgatom binding to oxygen- or nitrogen-containing ligands, such ascarboxylate or amide groups, respectively, are at least 10 orders ofmagnitude lower.2 The thermodynamic stability of the bondsformed between inorganic mercuric ions and the reduced sulfuratom of the tripeptide glutathione (GSH) has been studied using13CNMR.12,13 It was demonstrated that when GSH andHg2+ arein aqueous solution in a molar ratio of 2:1, each mercuric ionforms thermodynamically stable, linear II coordinate covalent,binds with the sulfur atom of two molecules of GSH, throughouta range of pH from 1 to 14.12,13 Organic mercuric ions, such asthe methylmercuric ion (CH3Hg
+), form linear I coordinatecovalent complexes with thiol-containing molecules.Despite the thermodynamic stability of the (linear I or II)
coordinate covalent bonds formed between the ionic forms of Hgand thiol-containing molecules in aqueous solution, the bindingcharacteristics between a mercuric ion and the reduced sulfuratom of various thiol-containing molecules appear to be morelabile in living organisms, especially in mammals.2,12,13 Complexfactors such as thiol-competition and electrophilic substitutionreactions may be responsible for the labile nature of binding thatoccurs between mercuric ions and certain thiol-containingmolecules in specific cellular and tissue-compartments inmammals. To exemplify this point, the preponderance ofmercuric ions present in the plasma of blood have been shownto be bound to proteins possessing one or more reducedcysteinyl residues.14−17 Albumin is one such protein. It is themost abundant protein in plasma and possesses a single −SHgroup with which mercuric ions can interact and bind. Othernucleophilic domains of albumin (and/or other plasma proteinsand nonprotein thiols) may react and bind to mercuric ions withless affinity and with less thermodynamic stability than to SHgroups. Interestingly, though, mercuric ions bound to albuminand other plasma proteins appear to be bound to these proteinsfor relatively short periods. This idea is supported by findingsshowing that the plasma burden of Hg decreases rapidly afterexposure, while there is a concurrent rapid rate of non-endocytotic uptake of mercuric ions in the epithelial cells ofthe kidneys and liver.2,18
In addition to binding to sulfhydryl groups on proteins,inorganic and organic mercuric ions bind to the sulfur atom ofone or more endogenous nonprotein thiols (such as GSH, Hcy,and Cys). Evidence indicates that the thiol S-conjugates formedcan serve as transportable substrates of various membranetransporters in the brain, endothelial cells of the blood-brainbarrier, and epithelial cells in the small intestine, liver andkidneys.19−22 It also appears that once mercuric ions gain entryinto systemic circulation, they undergo one or more complexelectrophilic substitution reactions that involve transfer from thereduced sulfur atom(s) of plasma proteins to the reduced sulfur
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381826

atom of one or more types of low molecular weight, nonprotein,thiols mentioned above.Due to the nonpolar nature of Hg0, it has the capacity to
traverse the plasma membrane of cells by mechanisms that donot require specific membrane transporters.22 It should beemphasized that once Hg0 gains access to systemic circulation,the oxygen-rich environment favors oxidation of Hg0 to Hg2+.Additionally, oxidation of Hg0 likely also occurs within certaintarget cells, although the amount of oxidation of Hg0 in bothintracellular and/or extracellular compartments is not clear atpresent. However, it does appear that most of the Hg0 that entersinto systemic circulation eventually undergoes oxidation,forming one or more mercuric species.23
The tremendously high affinity between oxidized forms of Hgand the reduced sulfur atom of sulfhydryl (and sulfanyl) (SH)groups serves as the therapeutic basis of thiol-containingpharmacological agents, such as penicillamine, N-acetylpenicill-amine, dithioerythritol, dithiothreitol, 2,3-bis(sulfanyl)propanol(also known as British Anti-Lewisite or BAL), meso-2,3-bis(sulfanyl)succinate (known formerly as meso-2,3-dimercapto-succinate; DMSA), and (R,S)-2,3-bis(sulfanyl)propane-1-sulfo-nate (known formerly as 2,3-dimercaptopropane-1-sulfonateacid; DMPS), forming complexes or chelates with mercuric(and/or mercurous) ions. The formation of mercuric complexesor chelates with these molecules appears to secure Hg2+ or R-Hg+
(where R represents an organic group) in a manner that likelyshields the mercuric ion from interacting with biologicallyrelevant nucleophiles. In addition, the polar, negatively chargedvicinal dithiols, DMPS and DMSA, have an apparent additionaladvantage in that, at least in the kidneys, the native moleculesgain access into target epithelial cells by sharing, serendipitously,one or more membrane proteins capable of transporting certainthiol S-conjugates of Hg2+ (and the native compound) into thetarget cells.24−26 After gaining access to the intracellular milieu ofrenal proximal tubular epithelial cells, the binding affinitybetween molecules of DMPS or DMSA and mercuric ions isgreat enough to detach and secure mercuric ions that were boundto a host of intracellular thiols.2 The complexes formed by thebinding of DMPS or DMSA to intracellular mercuric ions resultsin the formation of negatively charged DMPS or DMSA S-conjugates that are water-soluble27,28 andmay be transported outof target cells by one or more transporters in the luminal plasmamembrane of proximal tubular epithelial cells. The negativecharge on the complexes that are transported into the tubularlumen not only promotes their solubility in an aqueous-basedluminal fluid but also impedes reabsorption back into peritubularcirculation.29
■ KIDNEY AS A PRIMARY TARGET OF MERCURICSPECIES
Most chemical species of Hg can induce toxic effects in a numberof tissues and organs, although the type and form of toxic effectsdepend greatly on the chemical species of Hg and the magnitude,duration, and route of exposure. The kidneys in mammals areparticularly vulnerable to the toxic effects of Hg, especiallymercurous and mercuric forms. Virtually all forms of Hg,including elemental, inorganic, and organic forms, can mediatenephrotoxic effects, depending on the conditions of exposure. Inexperimental animals and a host of in vitro experimentalconditions, mercuric species have been shown to have a greatpredilection to interact with, and be transported into, renaltubular epithelial cells.19−21,30,31
Systemic distributions of organic forms of Hg are more diffusethan that of inorganic forms. In addition to having toxic effects inthe kidneys, organic forms of Hg adversely affect cells in blood,placenta, fetal tissues, and organs and neural tissues, as well asothers.2,3,22,32 Differences in cellular mechanisms involved in thetransport and metabolism of inorganic and organic forms of Hg(in the various compartments of the body) are likely responsiblefor the disparity in organ system distribution, pattern ofbiological effect, and toxic potency of these forms of Hg.In the kidneys, both inorganic and organic forms of Hg
accumulate primarily in the cortex and outer stripe of the outermedulla.18,33−35 Autoradiographic and histochemical data36−43
and tubular microdissection data from mice, rats, and rabbits44,45
indicate that inorganic species of Hg are taken up almostexclusively by the convoluted and straight segments of bothcortical and juxtamedullary proximal tubules. Deposits of Hghave also been found in the renal proximal tubules of monkeysexposed to Hg0 originating from dental amalgams.5 Although thesegments of the proximal tubule appear to be the predominantsites wheremercuric ions are taken up and accumulated (as Hg2+-thiol complexes), there is insufficient data to exclude thepossibility that other segments of the nephron and/or collectingduct may also take up, accumulate, and transport inorganic and/or organic forms of Hg.Deposits of presumed inorganic Hg have also been identified
by various experimental techniques in the epithelial cells liningthe entire lengths of renal proximal tubules in rats and miceexposed to organic forms of Hg.34,39−41 Moreover, experimentalevidence indicates that a significant fraction of Hg in the kidneysof animals exposed to methylmercury (CH3Hg
+) is biotrans-formed to Hg2+ prior to or after it enters renal tubular epithelialcells.46−48 Additional support for this hypothesis comes fromdata demonstrating that intracellular conversion of methylmer-cury to inorganic Hg can occur, albeit by a currently unknownmechanism.49
■ PROXIMAL TUBULAR UPTAKE AND TRANSPORTOF MERCURY
One of the initial hypotheses regarding the mechanisms of howmercuric species gain access to proximal tubular epithelial cellsarose from the notion that filtered complexes of Hg-albumin cangain potential entry into proximal tubular cells via endocyto-sis.33,50,51 As mentioned above, albumin is the most abundantprotein in plasma that possesses a single free, unbound, SH group(on a terminal cysteinyl residue), which provides a high affinitybinding site for a mercuric ion.52 Previous data indicate that thelargest percentage of Hg in the plasma is bound to acid-precipitable proteins, such as albumin.14−17,53 Despite the sievingcoefficient for albumin being low in renal glomeruli of morehighly developed mammals, significant amounts (low gramquantities) of albumin are filtered during each day. Thus, the ideathat albumin-Hg complexes are filtered at the renal glomerulus isnot an unreasonable one.In a previous study, where rats were made proteinuric by
treatment with the proximal tubular toxicant gentamicin,significant amounts of inorganic Hg were excreted in the urineand were presumed to be conjugated to albumin.50 Assuming,that the permeability of albumin at the glomerular filtrationbarrier in these rats was unaltered by gentamicin and that the Hgin excreted urine was associated with albumin, the findings tendto support the hypothesis that some fraction of inorganic Hg inthe urine may have entered into the luminal compartment ofproximal tubules bound to albumin.
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381827

In a later study, Zalups and Barfuss33 attempted to implicate amercuric conjugate of albumin in the luminal uptake of inorganicHg by simultaneously evaluating the renal disposition ofintravenously administered 125I-albumin and 203Hg2+. Theirdata suggest that mercuric conjugates of albumin are not theprimary species of Hg taken up at the luminal membrane ofproximal tubular cells. However, endocytosis of a mercuricconjugate of albumin could not be excluded as it may play aminor role in this uptake.Data from a series of more recent studies provide much more
definitive information on the proximal tubular handling of Hg.These data indicate that there are at least two distinctmechanisms involved in the uptake of mercuric ions by proximaltubular epithelial cells. One of the mechanisms involvesmembrane proteins localized in the luminal plasma mem-brane,2,45,54−62 while the other involves transporters in thebasolateral plasma membrane.62−70
■ LUMINAL UPTAKE OF MERCURY BY PROXIMALTUBULAR CELLS
Role of γ-Glutamyltransferase. In vivo uptake of inorganicHg (and to a lesser extent organic forms of Hg) at the luminalplasma membrane of tubular epithelial cells in the kidneys hasbeen linked to the activity of γ-glutamyltransferase (γ-GT),2,54,59,71−74 which is presumed to be present exclusively inthe luminal (brush border) membrane of the epithelial cellslining both the pars convoluta and pars recta segments of theproximal tubule.75,76 The active site of γ-GT is present in theluminal compartment of the proximal tubule.76 The sole action ofthis enzyme is to cleave the γ-glutamylcysteine bond inmoleculesof GSH present in the tubular lumen, thereby formingcysteinylglycine and glutamate. Evidence implicating the activityof the enzyme in the renal tubular uptake of mercuric speciescomes mainly from in vivo experiments in which renal (andhepatic) γ-glutamyltransferase was inhibited irreversibly bypretreatment with L-(αS,5S)-α-amino-3-chloro-4,5-dihydro-5-isoxazoleacetic acid (acivicin). Pretreating animals with acivicindecreases the renal uptake and/or accumulation of Hg2+ andcauses significant increases in the urinary excretion of Hg2+ inmice71,77 and rats59,78,79 exposed to inorganic Hg or in miceexposed to methylmercury77 or Hg vapor.80 Enhanced urinaryexcretion of GSH has also been documented in acivicin-pretreated rats injected subsequently with inorganic Hg,providing additional support for the hypothesis that luminaldegradation of GSH in proximal tubules is indeed prevented bytreatment with acivicin.78
In two isolated perfused tubule studies, Cannon et al. providedirect evidence, linking the activity of γ-GT to the luminal uptakeof Hg2+, when delivered as GSH S-conjugates, in proximaltubular segments. These investigators demonstrated thatinhibition of the activity of γ-GT (by adding acivicin to theperfusing solution) decreases the luminal uptake (disappearanceflux, JD) and cellular accumulation of mercuric ions in theepithelial cells lining intact proximal tubular segments when themercuric ions were delivered to the luminal compartment asGSH S-conjugates of Hg2+.57,81
Collectively, the aforementioned in vivo and in vitro dataindicate that following exposure to inorganic forms of Hg, asignificant fraction of inorganic mercuric ions is taken up byproximal tubular epithelial cells by a luminal absorptivemechanism dependent on the actions of γ-GT.Presence and Formation ofMercuric Conjugates in the
Proximal Tubular Lumen. A major implication of the data
obtained during in vivo inhibition of γ-GT is that a pool ofmercuric ions is present in the luminal compartment of proximaltubules, as GSH S-conjugates, prior to the mercuric ions beingtaken up. Although it is not known exactly where these mercuricconjugates of GSH are formed or how they gain entry to theluminal compartment of proximal tubules, one must consider thepossibility that some of these complexes are formed outside thekidneys (either in systemic circulation or in cells of anotherorgan, such as hepatocytes of the liver) and are then deliveredinto the lumen of the proximal tubule via glomerular filtration.There are three reasons to suspect that this occurs. First, theformation of mercuric conjugates of GSH in the plasma ispossible because the concentration of GSH in plasma (of rats) isapproximately 10 μM,82 which provides a pool of GSH to formGSH S-conjugates with mercuric ions in plasma. Second,hepatocytes have been shown to generate significant amountsof GSH that end up in systemic circulation.76 As a result of thesignificant production of GSH in the liver and the fact that theliver accumulates Hg2+, there is the potential for GSH S-conjugates of Hg2+ being formed in hepatocytes and then beingtransported out into both biliary and sinusoidal compart-ments.83−86 Accordingly, it is possible that some GSH S-conjugates of Hg2+ formed in hepatocytes may enter the systemiccirculation along with GSH, where they can then be delivered tothe kidneys. Third, GSH and the corresponding mercuricconjugates are small enough to pass readily through theglomerular filtration barrier unimpeded.One must also consider that a significant pool of GSH S-
conjugates of Hg is actually formed in the lumen of the proximaltubule via mechanisms involving thiol-competition, which likelyresults as a consequence of the secretion of significant amounts ofGSH into the proximal tubular lumen. Supporting this notion aredata demonstrating that approximately 75% of the GSHsynthesized de novo in pars recta segments of proximal tubulesis subsequently secreted into the tubular lumen.87 This secretioncould potentially provide a high enough concentration of GSH inthe luminal compartment of the proximal tubule for thiol-competition to occur.Another possibility is that GSH S-conjugates of Hg that are
formed within proximal tubular cells are secreted into the tubularlumen by export transporters in the luminal plasma membrane.Once in the lumen, the GSH S-conjugates of Hg can then bedegraded readily by the tremendous abundance of γ-GT in theluminal plasma membrane. There are in vivo data from mice thattend to support this hypothesis.77 Localization of the multidrugresistance-associated protein, MRP2, in the kidneys also tends tosupport the possibility of luminal secretion of GSH S-conjugatesof both inorganic and organic mercuric ions. MRP2 has beenlocalized in the brush-border membrane of the epithelial cellslining S1, S2, and S3 proximal tubular segments of the rat and inthe luminal plasma membrane of human proximal tubularepithelial cells.88,89 This membrane transporter is a member ofthe ATP-binding cassette (ABC) superfamily of transportproteins and has been shown to be involved in the intracellularto extracellular transport of GSH S-conjugates at the canalicularmembrane of hepatocytes.90 On the basis of what is currentlyknown about the cellular location and function of MRP2, itseems reasonable to hypothesize that intracellular GSH S-conjugates of Hg2+ are also transported (in a secretory manner)by this protein in both proximal tubular epithelial cells andhepatocytes.
Cleavage Products of GSH S-conjugates of Mercuricions as Transportable Molecules. Considering that luminal
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381828

uptake of mercuric ions by proximal tubular cells is linked to theactivity of γ-GT and that GSH S-conjugates of Hg2+ are presentin the tubular lumen, the preponderance of the luminal uptake ofmercuric ions appears to involve the transport of some productformed by the actions of γ-GT. One such product is a mercuricconjugate of cysteinylglycine (CysGly), which may be trans-ported by one of the small peptide transport systems in theluminal plasma membrane. However, because of the high level ofactivity of dehydropeptidases (such as cysteinylglycinase) in theluminal membrane, one would predict that if there is transport ofthis mercuric conjugate along the proximal tubule in vivo, the rateof transport would be very low. On the basis of the high activitiesof both γ-GT and cysteinylglycinase, it is most likely that theprimary species of Hg2+ transported at the luminal membrane isan L-Cys S-conjugate of Hg2+, via one or more amino acidtransport systems. In fact, in vitro evidence indicating thatsequential enzymatic degradation of GSH to CysGly, and then toL-Cys, is possible while a mercuric ion remains bound to thesulfur atom of the cysteinyl residue (at the site of the−SH group)of GSH.91
Role of Cysteinylglycinase in the Luminal Uptake ofMercuric Species. Cannon et al. have examined the potentialfor luminal transport of CysGly S-conjugates of Hg2+ in isolatedperfused S2 segments of the rabbit proximal tubule.57,81 Theydemonstrated that inhibition of cysteinylglycinase with thedehydropeptidase-1 inhibitor, Cilastatin, caused significantreductions in the luminal uptake of Hg2+ when it was in theform of a CysGly S-conjugate. These findings support thehypothesis that when Hg2+ is conjugated to CysGly, much of theluminal absorption of Hg is linked to the actions of thedehydropeptidase-1 (i.e., cysteinylglycinase) that cleaves thepeptide bond in molecules of CysGly to yield Cys and L-glycine(Gly). Interestingly, however, inhibition of luminal dehydro-peptidases did not completely prevent the luminal uptake of Hgwhen it was in the form of a CysGly S-conjugate of Hg2+. Thesefindings indicate that, at least in isolated perfused proximaltubular segments, some transport of mercuric conjugates ofCysGly may actually occur at the luminal membrane, whileluminal dehydropeptidases are inhibited. However, before onecan make any definitive conclusions about potential transport ofmercuric conjugates of CysGly along the three sections ofproximal tubule in vivo, one must consider additional factors,such as potential heterogeneity in the handling of GSH andCysGly S-conjugates of Hg2+ along the entire proximal tubule. Infact, there are findings indicating significant axial heterogeneity inthe synthesis, secretion, and/or transport of GSH occurringalong the length of the rabbit proximal tubule.87,92
Cys S-Conjugates ofMercury as Primary TransportableSubstrates. Evidence from both in vivo and in vitro studiesprovides strong support for the hypothesis that Cys S-conjugatesof Hg2+ (in particular, Cys-S-Hg-S-Cys) are one of the principalchemical species of Hg2+ transported into proximal tubularepithelial cells at the luminal membrane. More specifically, in vivodata from rats demonstrate that the renal uptake and subsequentaccumulation of Hg2+34,61 as well as the level of renal tubularinjury and necrosis induced by Hg2+ are increased in rats whenHg2+ is administered intravenously as Cys-S-Hg-S-Cys.93 In vitrodata from renal brush-border membranes demonstrate thatmercuric ions gain entry into the intravesicular compartmentmore readily when they are in the form of Cys S-conjugates thanwhen they are in the form of GSH S-conjugates or even mercuriccomplexes of chloride, including HgCl2.
94
By far, one of the more convincing bodies of evidenceimplicating Cys-S-Hg-S-Cys as a transportable substrate at theluminal plasma membrane of proximal tubular cells comes fromstudies utilizing the isolated perfused tubule technique.57,81
Some of the advantages of this technique are that isolatedsegments of the rabbit proximal tubule are perfused through thelumen in vitro under physiological and biophysical conditionssimilar to those found in vivo in intact kidneys. The findings fromthese isolated perfused tubule studies demonstrate that the ratesof luminal uptake (disappearance flux, JD) of mercuric ions in S2segments of the rabbit proximal tubule are at least 2-fold greaterwhen the segments are perfused through the luminal compart-ment as Cys S-conjugates than as GSH S-conjugates or CysGly S-conjugates. Additionally, the findings from these studies stronglyindicate that Cys-S-Hg-S-Cys is taken up from the luminalcompartment of proximal tubular segments by one or moreamino acid transporters present in the luminal plasmamembrane.57,81 Transport studies performed in the presenceand absence of sodium (in both luminal and basolateralcompartments) provide additional evidence that the luminaluptake of Cys-S-Hg-S-Cys involves at least two separate aminoacid transport systems, one (or more) dependent on thepresence of extracellular sodium and the other not dependent onthe presence of extracellular sodium.Another set of experiments with isolated perfused S2 segments
of rabbit proximal tubules indicate that the luminal uptake ofCys-S-Hg-S-Cys may involve one or more of the same transportsystems utilized in the luminal uptake the disulfide amino acidcystine (Cys-S−S-Cys).57 These experiments demonstrated thataddition of 3 mM L-lysine (Lys) to a perfusate containing 20 μMHg2+ and 80 μM Cys reduced the net rate of luminal uptake ofHg2+ by approximately 50% in the same tubular segments.Interestingly, Schafer and Watkins had established previouslythat the presence of 3 mM L-lysine (3 mM) in the perfusateinhibits the luminal uptake of cystine (300 μM) by approximately50% in isolated perfused S2 segments of the rabbit proximaltubule.95 Their findings provide evidence suggesting that theluminal absorption of cystine occurs through a transporter sharedby the basic amino acid L-Lys. Taken together, the findings fromthese studies suggest that at least some fraction of the luminaluptake of cystine and Cys-S-Hg-S-Cys occurs by a commontransport system.In a more recent study, Zalups and colleagues tested the
hypothesis that the heterodimeric amino acid transporter, systemb0,+, is capable of individually transporting cystine and Cys-S-Hg-S-Cys.55 The rationale for this hypothesis comes in part from thefact that in the kidneys, system b0,+ is present exclusively in theluminal plasma membrane of proximal tubular cells and that thistransport system plays an important role in the sodium-independent luminal absorption of cystine and certain positivelycharged amino acids (such as L-lysine and L-arginine (Arg)).Using Madin−Darby canine kidney (MDCK) cells (which arederived from distal nephron segments and do not express systemb0,+) transfected stably with both rBAT and b0,+AT (the heavychain and light chain components, respectively, of system b0,+),these investigators demonstrated that MDCK cells expressing afunctional form of system b0,+ are capable of transporting notonly cystine (and the positively charged amino acids, L-Arg and L-Lys) but also the mercuric complex, Cys-S-Hg-S-Cys.In a subsequent study utilizing the same transfected MDCK
cells, Bridges and Zalups demonstrated that the Hcy S-conjugateof the Hg2+, Hcy-S-Hg-S-Hcy, can also be transported by systemb0,+ in a time- and concentration-dependent manner.56 In
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381829

addition, they demonstrated that the transfected MDCK cellswere more susceptible to the toxic effects of Hg2+ thannontransfected cells when the cells were exposed to Hcy-S-Hg-S-Hcy in a concentration-dependent manner. Thus, these twostudies provide the most current line of molecular evidenceindicating that system b0,+ plays a role in the luminal absorptionof both Cys-S-Hg-S-Cys and Hcy-S-Hg-S-Hcy.Transport systems, other than system b0,+, may also participate
in the luminal absorption of thiol S-conjugates of Hg2+ inproximal tubular cells. However, they have not yet beenidentified.Potential Role of Molecular Mimicry. On the basis of
common features between the structures of cystine and Cys-S-Hg-S-Cys, Zalups and colleagues have hypothesized that somecomponent of the absorptive luminal transport of Cys-S-Hg-S-Cys occurs by a mechanism involving molecular mimicry.20 Theypostulate that Cys-S-Hg-S-Cys can act as a functional mimic ofthe amino acid cystine at the site of one or more transporter(s)responsible for the luminal uptake of cystine. Similarly, theseinvestigators postulated that Hcy-S-Hg-S-Hcy may havecharacteristics similar to the disulfide homocystine (Hcy-S−S-Hcy), which also permits the mercuric conjugate, Hcy-S-Hg-S-Hcy, to be transported into proximal tubular epithelial cells bysystem b0,+.Molecular mimicry is not a novel notion. Clarkson has
discussed the nature by which ionic forms of Hg and some othermetals can form complexes with certain biological molecules thatcan serve, either in part or in entirety, as molecular mimics ofendogenous molecules at critical recognition sites of transportersof these endogenous molecules.23 For example, the complexformed between methylmercury and Cys has been postulated toserve as a mimic of the amino acid L-methionine (Met), as ameans to gain entry into the central nervous system via specificamino acid transporters.23,96 This hypothesis is supported, inpart, by findings from studies on the extracellular-to-intracellulartransport of the S-conjugate methylmercury (Cys-S-CH3Hg) inastrocytes97 and endothelial cells lining the blood−brainbarrier.98,99 Clarkson postulates that additional molecularmimics may form when GSH binds to Hg2+ or CH3Hg
+. Inparticular, it was postulated that the complex formed when twomolecules of GSH bind to Hg2+, forming the complex GSH-S-Hg-S-GSH, may behave as a functional molecular mimic ofglutathione disulfide (GS-SG).23
In recent years, the idea of molecular mimicry has becomesomewhat controversial. Studies using mass spectrometry, X-rayabsorption spectroscopy, and computational chemistry sug-gested that the structures of Cys-S-CH3Hg and Cys-S-Hg-S-Cysdid not possess enough similarity with the amino acids, Met andCys, respectively, to support a mechanism of molecularmimicry.100 The authors of this study reject the idea thatamino acid conjugates of mercuric ions can serve, in theirentirety, as mimics of endogenous substrates. Rather, theypropose that the ability of mercuric conjugates to be taken up byamino acid transporters may be due to transporter recognition ofthe amino acid component of the conjugate.
■ BASOLATERAL UPTAKE OF MERCURY BYPROXIMAL TUBULAR CELLS
Role of Organic Anion Transporters. In addition toluminal mechanisms participating in the uptake of mercuriccompounds, certain mercuric species are also taken up at thebasolateral membrane of proximal tubular epithelial cells.Experimental findings from rats indicate that approximately
40% of the dose of Hg2+ is present in the total renal mass of ratsduring the initial hour after intravenous administration of anontoxic dose of mercuric chloride.64,101−103 Moreover,approximately 40−60% of the renal burden of Hg can beattributed to a basolateral transport mechanism.59,62,64,70,104
Zalups and Minor were the first to demonstrate that renaltubular uptake of administered Hg2+ occurs when glomerularfiltration is reduced to negligible levels in one or both kidneys, byusing the well established stop-flow technique.62 Pretreatmentwith mannitol in combination with ureteral ligation causes anapproximate 40% decrease in the net renal uptake andaccumulation of Hg2+ during the initial hour after administrationof a non-nephrotoxic dose of mercuric chloride. In vivopretreatment with p-aminohippurate (PAH), which is acompetitive substrate specific to members of the renal organicanion transporter family,105−112 also significantly reduces theacute renal tubular uptake and accumulation of Hg2+.62,104 Infact, the combination of ureteral ligation and pretreatment withPAH has been shown to cause an approximate 85% reduction inthe net renal tubular uptake and accumulation of Hg2+ during thefirst hour after exposure to mercuric chloride. Overall, thesefindings indicate that the majority of the basolateral uptake ofHg2+ can be inhibited by PAH, which implicates one or more ofthe organic anion transporter proteins as a primary mechanism inthe basolateral uptake of Hg2+. Data from other recent studieshave confirmed that basolateral uptake of inorganic Hg occurs inthe kidney and that the primary mechanism involved is linked tothe activity of one or more members of the organic aniontransport system.59,62,64,101,102,104,113
Organic anion transporters have also been implicated in thebasolateral uptake of organic mercuric compounds. It has beendemonstrated that renal uptake and/or accumulation58 andtoxicity114 of administered methylmercury are reduced signifi-cantly in mice pretreated with probenecid, which is anothercompetitive inhibitor of organic anion transporters in renalproximal tubules.110
Two main organic anion transport proteins are present in thebasolateral membrane of proximal tubular cells, namely, theorganic anion transporter 1 (OAT1) and the other isoform, theorganic anion transporter 3 (OAT3).115−119 The predominantorganic anion transporter in the basolateral membrane appears tobe OAT1, although PAH (which is thought of as the classicalcompetitive inhibitor of organic anion transport in proximaltubular cells) can inhibit the transport-activity of both OAT1 andOAT3.In a series of recent studies, Zalups and colleagues examined
the kinetics and substrate specificity of thiol S-conjugates of Hg2+
and CH3Hg+ in MDCK cells transfected stably to express OAT1
and in oocytes from Xenopus laevis expressing OAT3.65−69,120
Using these cellular models, they demonstrated that Cys andHcyS-conjugates of Hg2+ and Cys, and Hcy and NAC S-conjugates ofCH3Hg
+ are all transportable substrates of OAT1. Interestingly,findings from another recent study indicate that the NAC S-conjugate of CH3Hg
+ is not transported efficiently in oocytesfrom Xenopus laevis microinjected with the cDNA for OAT3,indicating that this methylmercuric species is not a high affinitysubstrate of OAT3.121 Overall, evidence to date indicates thatOAT1 has a fairly broad range of substrate-specificity forbiologically relevant mercuric species. Moreover, the findingsfrom the aforementioned experiments clearly support a role forOAT1 (and perhaps OAT3) in the basolateral uptake ofmercuric species from peritubular blood.
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381830

Role of the Basolateral Dicarboxylate TransporterNaC3. In the 1960s, Clarkson and Magos122 demonstratedthat when animals are pretreated with the dicarboxylate maleate,reductions occur in the net renal accumulation of administeredHg2+. However, it is not clear whether the changes in the renaldisposition of Hg2+ are due to the inhibitory effects of maleate onrenal cellular metabolism123 or whether they are due to directeffects at the site of a transporter of one or more S-conjugates.These investigators also found that fumarate, which is an isomerof maleate, did not have the same effects as maleate, indicatingisomer-specific effects on the renal disposition of Hg2+.In the late 1990s, Zalups and Barfuss102 demonstrated that
pretreatment with small aliphatic dicarboxylates (made up of 4−6 carbon atoms) such as succinate, glutarate, or adipate (but notmalonate, which contains only three carbon atoms), inhibits therenal (basolateral) uptake of intravenously administered Hg2+ (asmercuric chloride) in a dose-dependent manner in normal ratsand in rats with ligated ureters. The inhibitory effects of thedicarboxylates on the renal tubular uptake, transport, andaccumulation of Hg2+ likely involves the synergistic activity ofboth the sodium-dependent dicarboxylate transporter NaC3 andthe OAT-transporters OAT1 and/or OAT3, which are allpresent in the basolateral membrane of proximal tubularepithelial cells.Organic anion transporters in the basolateral membrane
function as organic anion/dicarboxylate exchangers.106,109
Numerous data indicate that intracellular generation of α-ketoglutarate contributes to the creation of an intracellularchemical gradient favoring the movement of this dicarboxylateout of the cell. α-Ketoglutarate moves down a concentrationgradient out of proximal tubular cells at the basolateralmembrane by exchanging with extracellular organic anions(including some dicarboxylates) at the site of OAT1 andOAT3.124 Interestingly, a significant fraction of the α-ketoglutarate (and other dicarboxylates) that exits proximaltubular cells at one or both of the organic anion exchangers istaken back up into the cells at the basolateral membrane viaNaC3.125 The activity of this dicarboxylate cotransport systemappears to derive electromotive energy from the sodium-gradientgenerated by the activity of (Na++ K+)-stimulated ATPase, whichis also present in the basolateral membrane.Although mechanisms by which succinate, glutarate, and
adipate inhibit the renal tubular uptake of inorganic Hg2+ havenot been defined fully, it seems likely that an excess of any ofthese dicarboxylates in the extracellular compartment createscompetition for the sodium-dependent entry of α-ketoglutarateat the site of the NaC3. Thus, reduction in the basolateral uptakeof α-ketoglutarate likely causes a decrease in the intracellularconcentration of this dicarboxylate. As a consequence, therewould be transient decreases in the chemical gradient favoringthe movement of α-ketoglutarate out of proximal tubular cells inexchange for the uptake of a host of organic anions from theperitubular plasma. Consequently, by changing the activity ofNaC3, there would be corresponding changes in the kineticactivity of the organic anion transport system. This would resultin a decreased rate of uptake of organic anions and transportablemercuric species by OAT1 and OAT3.Since a number of small dicarboxylates are themselves organic
anions, an excess of any of these dicarboxylates in theextracellular compartment would likely lead to competitionwith whatever species of Hg that is putatively transported by theorganic anion transport system. This could explain the decreasedrate of uptake of Hg at the basolateral membrane detected in rats
pretreated with small dicarboxylates. There is evidence that bothadipate and glutarate but not succinate or malonate can competedirectly with α-ketoglutarate at the site of the organic aniontransport system.108,109,111,112
■ ELIMINATION OF MERCURIC SPECIES BYPROXIMAL TUBULAR CELLS
Once mercuric ions gain access to the intracellular compartmentof proximal tubular epithelial cells, they tend to be retained in thecells by complex bonding interactions with protein andnonprotein thiols.2 Despite the apparent intracellular retentionof mercuric ions, there appears to be some degree of luminalsecretion of certain mercuric species,77,126−128 adding to the poolof Hg excreted in the urine.129 This conclusion is derived fromrecent developments by Bridges and colleagues, who havedemonstrated in vivo that low-level, luminal secretion of Hg2+
occurs in proximal tubular cells via the action of the multidrugresistance-associated protein 2 (MRP2).128 The presence ofMRP2 in the luminal plasma membrane of proximal tubularepithelial cells has been demonstrated previously.88,89 Bridgesand colleagues have demonstrated that without the presence offunctional MRP2 protein in the luminal plasma membrane,movement of mercuric species from within proximal tubular cellsto the luminal compartment of the nephron is greatly diminished.These data come from in vivo metabolic studies using MRP2-deficient (TR−) and normal (Wistar) rats treated acutely with anon-nephrotoxic dose of HgCl2.In addition to MRP2, the luminal plasma membrane of
proximal tubular cells also contains the multidrug resistanceprotein 4 (MRP4), another member of the ABC protein family.However, preliminary unpublished findings of Bridges andcolleagues tend to indicate that MRP4 does not participate in theluminal secretion of various mercuric species.
■ HANDLING OF DMPS AND DMSA IN THE KIDNEYS
According to documentation from Heyl Chem,28 DMPS (brandname Dimaval) was first synthesized in 1951. While preciseinformation on when DMSA (brand name Chemet) was firstsynthesized is less clear, it became available for use in the 1960s.Unlike DMPS, DMSA has been approved for use (mainly forreducing the body-burden of lead (Pb)) by the United StatesFood and Drug Administration (FDA). Although DMPS has notbeen approved for use in the U.S., it appears to be widely usedinternationally. Both molecules are composed of a short, straight-chain, aliphatic carbon skeleton. DMPS has a propane skeletonand has two SH groups on carbons 2 and 3 and a sulfonate (SO−)group on carbon 1. By contrast, DMSA is a dicarboxylic acid withsuccinate serving as the carbon backbone of the molecule.Carbons 2 and 3 have a single SH group bound to them.Although these two vicinal dithiols are essentially two differenttypes of molecules due to the functional groups on the terminalcarbon atoms, they are both polar in nature and are water-soluble, allowing them to be administered easily to patients.27
Studies in human indicate that both compounds are capable ofreducing the body burden of mercury.130,131 DMPS is the morewater-soluble species of the two, and data from experimentalstudies in rats tend to indicate that DMPS is slightly moreeffective at extracting mercuric ions from the kidneys andreducing the body-burden of Hg than DMSA.35,126−128,132−134
In well-oxygenated, aqueous solutions containing electrolytes,the SH groups in both DMPS and DMSA can undergo oxidationforming mixed disulfides. One of the more common disulfides
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381831

formed involves the oxidation of both SH groups of DMPS orDMSA, with the subsequent formation of disulfide bondsbetween corresponding sulfur atoms of another molecule ofDMPS or DMSA, respectively. In addition, previous exper-imental evidence indicates that DMPS, and likely DMSA,undergo extensive oxidation and form one or more disulfideforms in vivo.135,136
Diamond and colleagues have shown that DMPS can besecreted from peritubular blood into the luminal compartment ofrenal tubules of the rat by the organic anion transport systemsthat transport PAH and other alkanesulfonates.137−139 They alsoshowed that DMPS is effective in reducing the renal burden ofHg2+ when administered in either a reduced or oxidized form,indicating that the organic anion transport systems in mammalsare likely capable of transporting reduced and/or oxidized formsof DMPS from the blood into renal tubular epithelial cells. Morerecently, Wright and colleagues provide molecular data fromoocytes expressing human OAT1 that add significant support tothe findings of Diamond and colleagues implicating one or moreorganic anion transporters in the basolateral uptake of reducedand/or oxidized forms of DMPS.24 Diamond and colleagues alsodemonstrated that oxidized forms of DMPS undergo reductionby a biotransformation reaction in proximal tubular epithelialcells by a mechanism that is currently unknown. Once in thereduced state, the vicinal thiols of DMPS can interact with andcompete for mercuric ions bound to intracellular thiols.It is presently unknown if the organic anion transporter
proteins play a role in the basolateral uptake of the dicarboxylatechelator, DMSA, in proximal tubular epithelial cells. There are,however, data showing dicarboxylates composed of five and sixcarbon atoms (such as glutarate (and its homologue α-ketoglutarate) and adipate) are important substrates (of theorganic anion transport system) that can be transportedbidirectionally.105,106,109 Moreover, it has been shown thatthese dicarboxylates can significantly affect the basolateral uptakeof mercuric species.102,105,106,109
Addition of one or more polar, negatively charged, functionalgroups on a short-chain carbon skeleton creates a substrate thathas the potential to be transported by one or more of the organicanion transporters (OAT1, 3, and 4) and the dicarboxylatetransporter, NaC3. OAT1 and 3 are localized exclusively in thebasolateral membrane of proximal tubular epithelial cells, whileOAT4 is found only in the apical membrane.124 As a result of theactivity of OAT1 and 3, DMPS and DMSA can gain easy accessto the intracellular environment of renal proximal tubularepithelial cells.Even though the pharmaceutical intent in the design of DMPS
and DMSA was likely to make the compounds water-soluble(more so than compounds available previously), these negativelycharged, vicinal dithiols serve as effective transportable substratesof the organic anion transport system in proximal tubular cells.Fortuitously, this enables these compounds to gain access to theintracellular milieu of the primary target cells that accumulateand are affected adversely by mercuric species.It should also be mentioned that as a result of the negative
potential of the intracellular environment established by theactivity of the Na+/K+-ATPase, movement of negatively charged,water-soluble molecules like DMPS and DMSA is impeded inmost somatic cells, including other epithelial cells, unless theypossess some form of organic anion transporter capable oftranslocating these negatively charged species from the blood.
■ COMPLEXATION OF DMPS AND DMSA WITHMERCURIC SPECIES IN VIVO
Currently, there is a paucity of information regarding the precisechemical structure of the complexes formed by DMPS or DMSAbinding to inorganic or organic mercuric ions in blood and inproximal tubular epithelial cells. On the basis of results from anumber of different methods of analysis, it appears that a varietyof potential structures may form. Recent X-ray absorptionspectroscopic data and density functional theory (DFT)calculations obtained from simple aqueous solutions containingvarious ratios of DMPS or DMSA and mercuric nitrate(Hg(NO3)2) tend to suggest that a single mercuric ion is notlikely to bond to the vicinal thiols on a single molecule of DMPSor DMSA.140 The authors of these findings postulate that at leasttwo DMPS or DMSA molecules must be involved in the bindingof one or two inorganic mercuric ion(s). Interestingly, one of thestructures postulated to form between a single inorganicmercuric ion and two molecules of either DMPS or DMSAinvolves tetrahedral binding, rather than linear II coordinatecovalent binding. Although these findings are interesting, theremay be an alternate species formed in vivo involving the bindingof the vicinal thiols of a single molecule of DMPS or DMSA to asingle inorganic mercuric ion. The potential species may involvethe binding of the vicinal sulfur atoms of a single molecule ofeither DMPS or DMSA to a single inorganic mercuric ion, whilethe single sulfur atom of twomolecules of Cys, Hcy, and/or GSHbind to the mercuric ion in a tetrahedral manner. Assuming thatCys andHcy are present in plasma at concentrations of at least 10μM82 and that the intracellular concentrations of GSH are in lowmillimolar concentrations,74,76,141−144 some form of thiolcompetition may promote intracellular formation of a mixedmercuric conjugate. From a purely chemical standpoint, theauthors of the spectroscopic data suggest that DMPS and DMSAare not well optimized for chelation of Hg owing to theobservation that neither compound forms a true chelate complexwith mercury.140,145 A chelate has been defined as a stable ringcomplex formed by the binding of a metal ion with two or morefunctional groups of another molecule.140 Therefore, DMPS andDMSA may be best described as complexing agents. Both ofthese vicinal thiols have proven to be extremely effective inreducing the renal and plasma burden of Hg, which makes thecompounds therapeutically effective despite not being optimizedchelators.A likely species of a DMPS or DMSA S-conjugate of Hg2+
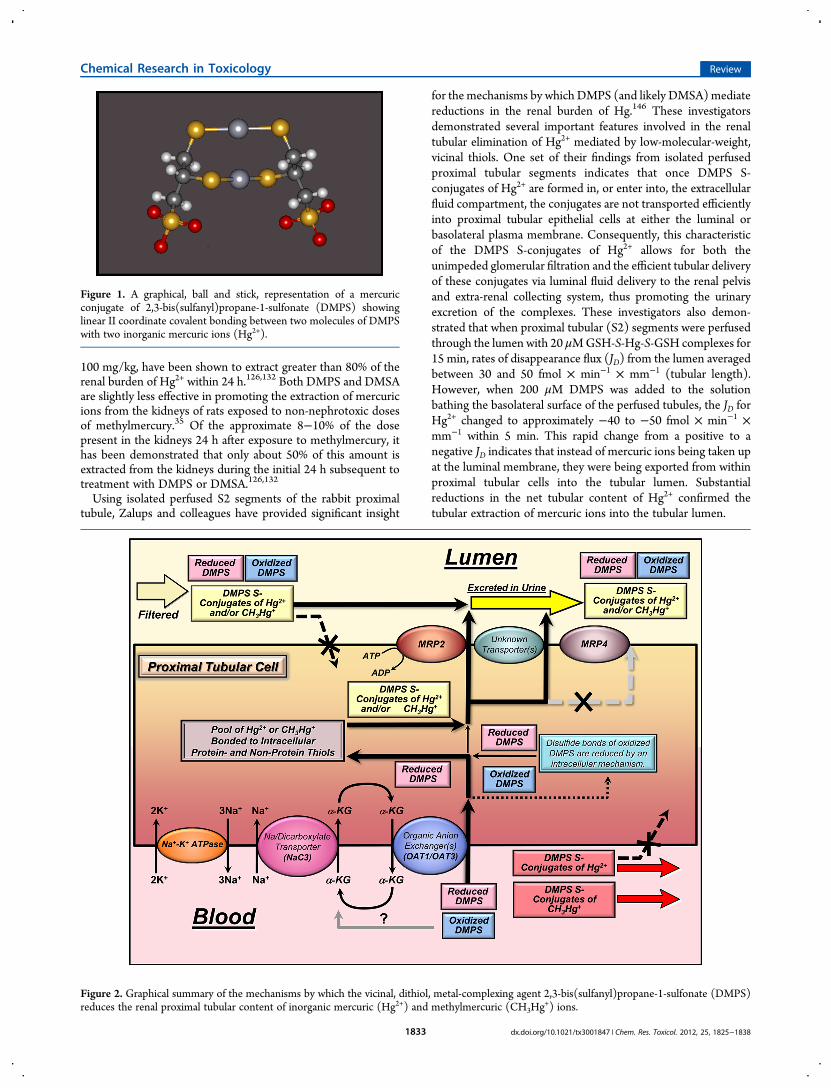
formed in both blood plasma and the intracellular compartmentof proximal tubular epithelial cells that promotes systemicelimination of Hg2+ is a complex formed by the binding of bothreduced sulfur atoms of twomolecules of either DMSA or DMPSwith one or two inorganic mercuric ions. Formation of such acomplex in aqueous solution has been demonstrated using X-rayabsorption spectroscopic data and density functional theory(DFT) calculations.140 An example of such a complex formed bytwo molecules of DMPS and two mercuric ions is illustrated inFigure 1.
■ RENAL EXTRACTION OF MERCURIC SPECIESMEDIATED BY DMPS AND DMSA
Numerous sets of data indicate that DMPS and DMSA canefficiently extract mercuric ions from the kidneys of rats, whichnormally accumulate as much as 50% or more of a single dose ofinorganic Hg within the first 24 h after exposure. Twointraperitoneal treatments of DMPS or DMSA, at a dose of
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381832
100 mg/kg, have been shown to extract greater than 80% of therenal burden of Hg2+ within 24 h.126,132 Both DMPS and DMSAare slightly less effective in promoting the extraction of mercuricions from the kidneys of rats exposed to non-nephrotoxic dosesof methylmercury.35 Of the approximate 8−10% of the dosepresent in the kidneys 24 h after exposure to methylmercury, ithas been demonstrated that only about 50% of this amount isextracted from the kidneys during the initial 24 h subsequent totreatment with DMPS or DMSA.126,132
Using isolated perfused S2 segments of the rabbit proximaltubule, Zalups and colleagues have provided significant insight
for the mechanisms by which DMPS (and likely DMSA)mediatereductions in the renal burden of Hg.146 These investigatorsdemonstrated several important features involved in the renaltubular elimination of Hg2+ mediated by low-molecular-weight,vicinal thiols. One set of their findings from isolated perfusedproximal tubular segments indicates that once DMPS S-conjugates of Hg2+ are formed in, or enter into, the extracellularfluid compartment, the conjugates are not transported efficientlyinto proximal tubular epithelial cells at either the luminal orbasolateral plasma membrane. Consequently, this characteristicof the DMPS S-conjugates of Hg2+ allows for both theunimpeded glomerular filtration and the efficient tubular deliveryof these conjugates via luminal fluid delivery to the renal pelvisand extra-renal collecting system, thus promoting the urinaryexcretion of the complexes. These investigators also demon-strated that when proximal tubular (S2) segments were perfusedthrough the lumen with 20 μMGSH-S-Hg-S-GSH complexes for15 min, rates of disappearance flux (JD) from the lumen averagedbetween 30 and 50 fmol × min−1 × mm−1 (tubular length).However, when 200 μM DMPS was added to the solutionbathing the basolateral surface of the perfused tubules, the JD forHg2+ changed to approximately −40 to −50 fmol × min−1 ×mm−1 within 5 min. This rapid change from a positive to anegative JD indicates that instead of mercuric ions being taken upat the luminal membrane, they were being exported from withinproximal tubular cells into the tubular lumen. Substantialreductions in the net tubular content of Hg2+ confirmed thetubular extraction of mercuric ions into the tubular lumen.
Figure 1. A graphical, ball and stick, representation of a mercuricconjugate of 2,3-bis(sulfanyl)propane-1-sulfonate (DMPS) showinglinear II coordinate covalent bonding between two molecules of DMPSwith two inorganic mercuric ions (Hg2+).
Figure 2. Graphical summary of the mechanisms by which the vicinal, dithiol, metal-complexing agent 2,3-bis(sulfanyl)propane-1-sulfonate (DMPS)reduces the renal proximal tubular content of inorganic mercuric (Hg2+) and methylmercuric (CH3Hg
+) ions.
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381833

More recent studies from our laboratory demonstrate that atleast some extraction of inorganic and organic mercuric ionsfrom within proximal tubular segments into the luminalextracellular compartment within the kidneys appears to bemediated by the MRP2. As mentioned above, this membraneprotein is a member of the ATP-binding cassette transporterfamily, which in the kidneys is located exclusively in the luminalmembrane of proximal tubular epithelial cells.89 This transporterhas been shown to mediate the transport of a broad range ofsubstrates into the lumen of proximal tubules.147−149 Moreover,recent sets of evidence indicate that MRP2 likely plays asignificant role in DMPS- and DMSA-mediated extraction ofmercuric ions from proximal tubular cells and contributes to thesubsequent urinary excretion of Hg.35,126,127 Figure 2 provides agraphical representation summarizing the known and putativemechanisms participating in the handling of DMPS and DMPSS-conjugates of Hg2+ and CH3Hg
+ by proximal tubular cells.Figure 2 shows that in humans and other mammals exposed toinorganic mercuric or methylmercuric forms of mercury and thentreated subsequently with DMPS, both DMPS (in reduced andoxidized forms) and DMPS S-conjugates of Hg2+ or CH3Hg
+ canbe present in the plasma of systemic blood shortly aftertreatment. During each passage of circulating blood through thekidneys, about 20−25% of the DMPS and the DMPS S-conjugates of Hg2+ or CH3Hg
+ in the plasma are filteredefficiently through the glomerular filtration barrier into theluminal compartment of proximal tubules. The remainingfraction of these compounds in plasma (approximately 75−80%) is delivered first to the basolateral compartment ofproximal tubules via peritubular capillary blood flow. In theluminal compartment of the proximal tubule and the remainderof the nephron, oxidized DMPS, reduced DMPS, or DMPS S-conjugates of Hg2+ or CH3Hg
+ are not absorbed efficientlybecause of the polar negative charge associated with the sulfonategroup of DMPS, thus promoting the urinary excretion of DMPSand mercuric complexes of DMPS. However, at the basolateralplasma membrane of proximal tubular cells, both oxidized andreduced forms of DMPS (not bonded tomercuric ions) are takenup efficiently into the proximal tubular epithelial cells by one orboth of the organic anion transporters present in the basolateralmembrane, namely, the organic transporter 1 (OAT1) and(OAT3).It should be mentioned that DMPS is taken up from the
extracellular basolateral compartment by OAT1 and/or OAT3by an outwardly driven intracellular to extracellular gradient forα-ketoglutarate, which is generated by intracellular metabolismand by transport back into the proximal tubular cells. One of thekey transporters linked to the activity of OAT1 and OAT3 andthe extracellular to intracellular transport of α-ketoglutarate is theNa+-dicarboxylate cotransporter, NaC3, which is also located inthe basolateral membrane. The electromotive force utilized bythis membrane transporter is derived from the actions of thesodium (Na+)−potassium (K+) ATPase in the basolateralmembrane, which pumps Na+ out of the proximal tubular cellsin an electrogenic manner, generating an extracellular tointracellular gradient of 140 mM to 10 mM, which favors themovement of Na+ back into the cells.Interestingly, evidence from isolated perfused segments of the
proximal tubule indicates that OAT1 and/or OAT3 are notinvolved in the basolateral uptake of DMPS S-conjugates of Hg2+.In fact, it appears that DMPS S-conjugates of Hg2+ are not takenup efficiently, or at all, from peritubular blood into proximaltubular epithelial cells.
Experimental evidence does indicate that the therapeuticactions of DMPS for extracting mercuric species from withinproximal tubular cells involve the following steps: (1) asmentioned above, DMPS (in a reduced and/or oxidized state)is taken up avidly at the basolateral membrane by OAT1 and/orOAT3. (2) Oxidized forms of DMPS taken up at the basolateralmembrane are reduced by an intracellular mechanism not yetknown. (3) Reduced forms of DMPS within the proximal tubularcells interact with, compete for, and then remove mercuric ionsbound to a host of potential intracellular thiol-containingmolecules, including large intracellular proteins, metallothio-neins (MT), glutathione (GSH), cysteine (Cys), and others. (4)After one or two mercuric ions bind to DMPS and a sufficientintracellular concentration of DMPS S-conjugates of Hg2+ orCH3Hg
+ has formed to generate a sufficient gradient favoring theoutward movement of these complexes, the conjugates aretransported out of the cells into the tubular lumen by themultidrug resistance-associated protein 2 (MRP2) and possiblyby another currently unknown transporter. Preliminary unpub-lished data indicate that MRP4 does not likely participate in theexport of mercuric conjugates of DMPS. (5) Finally, the DMPSS-conjugates of Hg2+ or CH3Hg
+ conjugates are excreted into theurine because they do not appear to be transportable substratesthat can be taken up by any segment of the nephron or collectingduct beyond the proximal tubule.
■ CONCLUSIONSAlthough a great deal of new information regarding mechanismsby which specific epithelial cells, particularly the proximal tubularepithelial cells in the kidneys handle various species of mercury, agreat deal of new information is needed to gain a more completeunderstanding of how mercuric species are handled in bothepithelial and nonepithelial target cells. Moreover, design of newmore effective, nontoxic, chelating or complexing agents isneeded to gain access to and extract mercuric ions from withintarget cells adversely affected by the various forms of mercury.Perhaps one of the more challenging tasks at hand is to designchemical compounds that can not only extract mercuric speciesfrom the kidney but also find a means to gain access through theblood−brain barrier to bind mercuric ions in neurons of theCNS. The compounds not only need to gain access to the targetneurons but also need to be designed to utilize one or moreneuronal membrane transporters to extract mercuric ions in amanner that can permit the formed mercuric complexes to gainaccess to systemic circulation, where they may be excreted by thekidneys and/or liver.
■ AUTHOR INFORMATIONCorresponding Author*Phone: 478-301-2559. Fax: 478-301-5489. E-mail: [email protected] of the contents of this review were supported, in part, bythe following grants awarded by the National Institute ofEnvironmental Health (NIEHS): ES05980 (to R.K.Z.),ES019991 (to C.C.B.), and ES015511 (to C.C.B.).NotesThe authors declare no competing financial interest.
■ ABBREVIATIONSABC, ATP-binding cassette; DMSA, meso-2,3-bis(sulfanyl)-succinate; DMPS, 2,3-bis(sulfanyl)propane-1-sulfonate; DFT,
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381834

density functional theory; CH3Hg+, methylmercury or methyl-
mercuric ion; Hg, mercury; Hg0, elemental mercury or metallicmercury; HgCl2, mercuric chloride; CysGly, cysteinylglycine;FDA, United States Food and Drug Administration; JD,disappearance flux; DL-Hcy, homocysteine; γ-GT, γ-glutamyl-transferase; GSH, glutathione; Hg+, mercurous ion; Hg2+,mercuric ion or Hg; Hg(NO3)2, mercuric nitrate; IUPAC,International Union of Pure and Applied Chemistry; NAC, N-acetylcysteine; MRP2, multidrug resistance-associated protein 2;MRP4, multidrug resistance-associated protein 4; NaC3,sodium-dependent dicarboxylate transporter 3; OAT1, organicanion transporter 1; OAT3, organic anion transporter 3; OAT4,organic anion transporter 4; SH, sulfhydryl or thiol group; SO−,sulfonate; PAH, p-aminohippurate; TR−, MRP2-deficient
■ REFERENCES(1) Zalups, R. K. (2000) Evidence for basolateral uptake of cadmium inthe kidneys of rats. Toxicol. Appl. Pharmacol. 164, 15−23.(2) Zalups, R. K. (2000) Molecular interactions with mercury in thekidney. Pharmacol. Rev. 52, 113−143.(3) ATSDR (1999) Toxicological Profile for Mercury, pp 1−617, UnitedStates Public Health Service/Agency for Toxic Substance Registry,Atlanta, GA.(4) Clarkson, T. W. (1993) Mercury: major issues in environmentalhealth. Environ. Health Perspect. 100, 31−38.(5) Danscher, G., Horsted-Bindslev, P., and Rungby, J. (1990) Tracesof mercury in organs from primates with amalgam fillings. Exp. Mol.Pathol. 52, 291−299.(6) George, G. N., Singh, S. P., Hoover, J., and Pickering, I. J. (2009)The chemical forms of mercury in aged and fresh dental amalgamsurfaces. Chem. Res. Toxicol. 22, 1761−1764.(7) Hahn, L. J., Kloiber, R., Leininger, R. W., Vimy, M. J., andLorscheider, F. L. (1990) Whole-body imaging of the distribution ofmercury released from dental fillings into monkey tissues. FASEB J. 4,3256−3260.(8) Hahn, L. J., Kloiber, R., Vimy, M. J., Takahashi, Y., and Lorscheider,F. L. (1989) Dental ″silver″ tooth fillings: a source of mercury exposurerevealed by whole-body image scan and tissue analysis. FASEB J. 3,2641−2646.(9) Fitzgerald, W. F., and Clarkson, T. W. (1991) Mercury andmonomethylmercury: present and future concerns. Environ. HealthPerspect. 96, 159−166.(10) Clarkson, T.W. (1972) The biological properties and distributionof mercury. Biochem. J. 130, 61P−63P.(11) Harris, H. H., Pickering, I. J., and George, G. N. (2003) Thechemical form of mercury in fish. Science 301, 1203.(12) Fuhr, B. J., and Rabenstein, D. L. (1973) Nuclear magneticresonance studies of the solution chemistry of metal complexes. IX. Thebinding of cadmium, zinc, lead, and mercury by glutathione. J. Am.Chem. Soc. 95, 6944−6950.(13) Rabenstein, D. L., Ed. (1989)Metal Complexes of Glutathione andTheir Biological Significance, Vol. 3, Wiley Press, New York.(14) Cember, H., Gallagher, P., and Faulkner, A. (1968) Distributionof mercury among blood fractions and serum proteins. Am. Ind. Hyg.Assoc. J. 29, 233−237.(15) Friedman, H. L. (1957) Relationship between chemical structureand biological activity in mercurial compounds. Ann. N.Y. Acad. Sci. 65,461−470.(16) Lau, S., and Sarkar, B. (1979) Inorganic mercury(II)-bindingcomponents in normal human blood serum. J. Toxicol. Environ. Health.5, 907−916.(17)Mussini, E. (1958) Bonds of mercurial diuretics to blood proteins.Boll. Soc. Ital. Biol. Sper. 34, 1588−1590.(18) Zalups, R. K. (1993) Early aspects of the intrarenal distribution ofmercury after the intravenous administration of mercuric chloride.Toxicology 79, 215−228.
(19) Aschner, M., and Aschner, J. L. (1990) Mercury neurotoxicity:mechanisms of blood-brain barrier transport.Neurosci. Biobehav. Rev. 14,169−176.(20) Bridges, C. C., and Zalups, R. K. (2005) Molecular and ionicmimicry and the transport of toxic metals. Toxicol. Appl. Pharmacol. 204,274−308.(21) Bridges, C. C., and Zalups, R. K. (2010) Transport of inorganicmercury and methylmercury in target tissues and organs. J. Toxicol.Environ. Health, Part B 13, 385−410.(22) Clarkson, T.W., andMagos, L. (2006) The toxicology of mercuryand its chemical compounds. Crit. Rev. Toxicol. 36, 609−662.(23) Clarkson, T. W. (1993) Molecular and ionic mimicry of toxicmetals. Annu. Rev. Pharmacol. Toxicol. 33, 545−571.(24) Islinger, F., Gekle, M., andWright, S. H. (2001) Interaction of 2,3-dimercapto-1-propane sulfonate with the human organic aniontransporter hOAT1. J. Pharmacol. Exp. Ther. 299, 741−747.(25) Rodiger, M., Zhang, X., Ugele, B., Gersdorff, N., Wright, S. H.,Burckhardt, G., and Bahn, A. (2010) Organic anion transporter 3(OAT3) and renal transport of the metal chelator 2,3-dimercapto-1-propanesulfonic acid (DMPS). Can. J. Physiol. Pharmacol. 88, 141−146.(26) Lungkaphin, A., Chatsudthipong, V., Evans, K. K., Groves, C. E.,Wright, S. H., and Dantzler, W. H. (2004) Interaction of the metalchelator DMPS with OAT1 and OAT3 in intact isolated rabbit renalproximal tubules. Am. J. Physiol. Renal Physiol. 286, F68−76.(27) Aposhian, H. V. (1983) DMSA and DMPS: water solubleantidotes for heavy metal poisoning. Annu. Rev. Pharmacol. Toxicol. 23,193−215.(28) Ruprecht, J. (2008)Dimaval, Heyl Chem-Pharm Fabrik G, Berlin.(29) Chang, I. J., Fischbach, B. V., Sile, S., and Golper, T. A. (2004)Extracorporeal Treatment of Poisoning, in The Kidney (Brenner, B. M.,Ed.) pp 2733−2757, Saunders, Philadelphia, PA.(30) Clarkson, T. W., Vyas, J. B., and Ballatori, N. (2007) Mechanismsof mercury disposition in the body. Am. J. Ind. Med. 50, 757−764.(31) Zalups, R. K., and Bridges, C. C. (2010) Molecular and CellularBiology of Mercury in the Kidneys, in Cellular and Molecular Biology ofMetals (Zalups, R. K., and Koropatnick, J., Ed.) pp 35−77, Taylor andFrancis, Boca Raton, FL.(32) Farina, M., Rocha, J. B., and Aschner, M. (2011) Mechanisms ofmethylmercury-induced neurotoxicity: evidence from experimentalstudies. Life Sci. 89, 555−563.(33) Zalups, R. K., and Barfuss, D. W. (1993) Intrarenal distribution ofinorganic mercury and albumin after coadministration. J. Toxicol.Environ. Health 40, 77−103.(34) Zalups, R. K., and Barfuss, D. W. (1995) Renal disposition ofmercury in rats after intravenous injection of inorganic mercury andcysteine. J. Toxicol. Environ. Health 44, 401−413.(35) Zalups, R. K., and Bridges, C. C. (2009) MRP2 involvement inrenal proximal tubular elimination of methylmercury mediated byDMPS or DMSA. Toxicol. Appl. Pharmacol. 235, 10−17.(36) Hultman, P., and Enestrom, S. (1986) Localization of mercury inthe kidney during experimental acute tubular necrosis studied by thecytochemical Silver Amplification method. Br. J. Exp. Pathol. 67, 493−503.(37) Hultman, P., and Enestrom, S. (1992) Dose-response studies inmurine mercury-induced autoimmunity and immune-complex disease.Toxicol. Appl. Pharmacol. 113, 199−208.(38) Hultman, P., Enestrom, S., and von Schenck, H. (1985) Renalhandling of inorganic mercury in mice. The early excretion phasefollowing a single intravenous injection of mercuric chloride studied bythe Silver Amplification method. Virchows Arch., B 49, 209−224.(39) Magos, L., Brown, A. W., Sparrow, S., Bailey, E., Snowden, R. T.,and Skipp, W. R. (1985) The comparative toxicology of ethyl- andmethylmercury. Arch. Toxicol. 57, 260−267.(40) Rodier, P. M., and Kates, B. (1988) Histological localization ofmethylmercury in mouse brain and kidney by emulsion autoradiographyof 203Hg. Toxicol. Appl. Pharmacol. 92, 224−234.(41) Rodier, P. M., Kates, B., and Simons, R. (1988) Mercurylocalization in mouse kidney over time: autoradiography versus silverstaining. Toxicol. Appl. Pharmacol. 92, 235−245.
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381835

(42) Taugner, R., Winkel, K. z., and Iravani, J. (1966) On thelocalization of mercuric chloride concentration in the rat kidney.Virchows Arch. Pathol. Anat. Physiol. Klin. Med. 340, 369−383.(43) Zalups, R. K. (1991) Autometallographic localization of inorganicmercury in the kidneys of rats: effect of unilateral nephrectomy andcompensatory renal growth. Exp. Mol. Pathol. 54, 10−21.(44) Zalups, R. K. (1991) Method for studying the in vivoaccumulation of inorganic mercury in segments of the nephron in thekidneys of rats treated with mercuric chloride. J. Pharmacol. Methods 26,89−104.(45) Zalups, R. K., and Barfuss, D. W. (1990) Accumulation ofinorganic mercury along the renal proximal tubule of the rabbit. Toxicol.Appl. Pharmacol. 106, 245−253.(46) Gage, J. C. (1964) Distribution and excretion of methyl andphenyl mercury salts. Br. J. Ind. Med. 21, 197−202.(47) Norseth, T., and Clarkson, T. W. (1970) Studies on thebiotransformation of 203Hg-labeled methyl mercury chloride in rats.Arch. Environ. Health 21, 717−727.(48) Norseth, T., and Clarkson, T. W. (1970) Biotransformation ofmethylmercury salts in the rat studied by specific determination ofinorganic mercury. Biochem. Pharmacol. 19, 2775−2783.(49) Dunn, J. D., and Clarkson, T. W. (1980) Does mercury exhalationsignal demethylation of methylmercury? Health Phys. 38, 411−414.(50) Madsen, K. M. (1980) Mercury accumulation in kidneylysosomes or proteinuric rats. Kidney Int. 18, 445−453.(51) Madsen, K. M., and Hansen, J. C. (1980) Subcellular distributionof mercury in the rat kidney cortex after exposure to mercuric chloride.Toxicol. Appl. Pharmacol. 54, 443−453.(52) Brown, D. L., and Shockley, P. (1982) Serum albumin: Structureand Characterization of Its Ligand Binding Sites, in Lipid-ProteinInteractions (Jost, P. V., and Griffith, P., Ed.) pp 25−68, John Wiley &Sons, New York.(53) Mussini, E. (1958) Distribution of mercurial diuretics in theorganism. Boll. Soc. Ital. Biol. Sper. 34, 1487−1490.(54) Baggett, J. M., and Berndt,W. O. (1986) The effect of depletion ofnonprotein sulfhydryls by diethyl maleate plus buthionine sulfoximineon renal uptake of mercury in the rat. Toxicol. Appl. Pharmacol. 83, 556−562.(55) Bridges, C. C., Bauch, C., Verrey, F., and Zalups, R. K. (2004)Mercuric conjugates of cysteine are transported by the amino acidtransporter system b(0,+): implications ofmolecular mimicry. J. Am. Soc.Nephrol. 15, 663−673.(56) Bridges, C. C., and Zalups, R. K. (2004) Homocysteine, systemb0,+ and the renal epithelial transport and toxicity of inorganic mercury.Am. J. Pathol. 165, 1385−1394.(57) Cannon, V. T., Zalups, R. K., and Barfuss, D. W. (2001) Aminoacid transporters involved in luminal transport of mercuric conjugates ofcysteine in rabbit proximal tubule. J. Pharmacol. Exp. Ther. 298, 780−789.(58) Tanaka, T., Naganuma, A., and Imura, N. (1992) Routes for renaltransport of methylmercury in mice. Eur. J. Pharmacol. 228, 9−14.(59) Zalups, R. K. (1995) Organic anion transport and action ofgamma-glutamyl transpeptidase in kidney linked mechanistically torenal tubular uptake of inorganic mercury. Toxicol. Appl. Pharmacol. 132,289−298.(60) Zalups, R. K., and Barfuss, D. W. (1993) Transport and toxicity ofmethylmercury along the proximal tubule of the rabbit. Toxicol. Appl.Pharmacol. 121, 176−185.(61) Zalups, R. K., and Barfuss, D. W. (1998) Participation of mercuricconjugates of cysteine, homocysteine, and N-acetylcysteine inmechanisms involved in the renal tubular uptake of inorganic mercury.J. Am. Soc. Nephrol. 9, 551−561.(62) Zalups, R. K., and Minor, K. H. (1995) Luminal and basolateralmechanisms involved in the renal tubular uptake of inorganic mercury. J.Toxicol. Environ. Health 46, 73−100.(63) Torres, A. M., Dnyanmote, A. V., Bush, K. T., Wu,W., and Nigam,S. K. (2011) Deletion of multispecific organic anion transporter Oat1/Slc22a6 protects against mercury-induced kidney injury. J. Biol. Chem.286, 26391−26395.
(64) Zalups, R. K. (1998) Basolateral uptake of inorganic mercury inthe kidney. Toxicol. Appl. Pharmacol. 151, 192−199.(65) Zalups, R. K., and Ahmad, S. (2004) Homocysteine and the renalepithelial transport and toxicity of inorganic mercury: role of basolateraltransporter organic anion transporter 1. J. Am. Soc. Nephrol. 15, 2023−2031.(66) Zalups, R. K., and Ahmad, S. (2005) Handling of cysteine S-conjugates of methylmercury in MDCK cells expressing human OAT1.Kidney Int. 68, 1684−1699.(67) Zalups, R. K., and Ahmad, S. (2005) Handling of thehomocysteine S-conjugate of methylmercury by renal epithelial cells:role of organic anion transporter 1 and amino acid transporters. J.Pharmacol. Exp. Ther. 315, 896−904.(68) Zalups, R. K., and Ahmad, S. (2005) Transport of N-acetylcysteine s-conjugates of methylmercury in Madin-Darby caninekidney cells stably transfected with human isoform of organic aniontransporter 1. J. Pharmacol. Exp. Ther. 314, 1158−1168.(69) Zalups, R. K., Aslamkhan, A. G., and Ahmad, S. (2004) Humanorganic anion transporter 1 mediates cellular uptake of cysteine-Sconjugates of inorganic mercury. Kidney Int. 66, 251−261.(70) Zalups, R. K., and Barfuss, D. W. (2002) Renal organic aniontransport system: a mechanism for the basolateral uptake of mercury-thiol conjugates along the pars recta of the proximal tubule. Toxicol.Appl. Pharmacol. 182, 234−243.(71) Tanaka, T., Naganuma, A., and Imura, N. (1990) Role of gamma-glutamyltranspeptidase in renal uptake and toxicity of inorganic mercuryin mice. Toxicology 60, 187−198.(72) Tanaka, T., Naganuma, A., Miura, N., and Imura, N. (1992) Roleof testosterone in gamma-glutamyltranspeptidase-dependent renalmethylmercury uptake in mice. Toxicol. Appl. Pharmacol. 112, 58−63.(73) Zalups, R. K., and Lash, L. H. (1994) Advances in understandingthe renal transport and toxicity of mercury. J. Toxicol. Environ. Health 42,1−44.(74) Zalups, R. K., and Lash, L. H. (1997) Depletion of glutathione inthe kidney and the renal disposition of administered inorganic mercury.Drug Metab. Dispos. 25, 516−523.(75) Meister, A. (1983) Transport and metabolism of glutathione andgamma-glutamyl amino acids. Biochem. Soc. Trans. 11, 793−794.(76) Meister, A., and Anderson, M. E. (1983) Glutathione. Annu. Rev.Biochem. 52, 711−760.(77) Tanaka-Kagawa, T., Naganuma, A., and Imura, N. (1993) Tubularsecretion and reabsorption of mercury compounds in mouse kidney. J.Pharmacol. Exp. Ther. 264, 776−782.(78) Berndt, W. O., Baggett, J. M., Blacker, A., and Houser, M. (1985)Renal glutathione and mercury uptake by kidney. Fundam. Appl. Toxicol.5, 832−839.(79) de Ceaurriz, J., Payan, J. P., Morel, G., and Brondeau,M. T. (1994)Role of extracellular glutathione and gamma-glutamyltranspeptidase inthe disposition and kidney toxicity of inorganic mercury in rats. J. Appl.Toxicol. 14, 201−206.(80) Kim, C. Y., Watanabe, C., and Satoh, H. (1995) Effects ofbuthionine sulfoximine (BSO) on mercury distribution after Hg(o)exposure. Toxicology 98, 67−72.(81) Cannon, V. T., Barfuss, D.W., and Zalups, R. K. (2000)Molecularhomology and the luminal transport of Hg2+ in the renal proximaltubule. J. Am. Soc. Nephrol. 11, 394−402.(82) Lash, L. H., and Jones, D. P. (1985) Distribution of oxidized andreduced forms of glutathione and cysteine in rat plasma. Arch. Biochem.Biophys. 240, 583−592.(83) Ballatori, N., and Clarkson, T. W. (1984) Dependence of biliarysecretion of inorganic mercury on the biliary transport of glutathione.Biochem. Pharmacol. 33, 1093−1098.(84) Ballatori, N., and Clarkson, T. W. (1985) Biliary secretion ofglutathione and of glutathione-metal complexes. Fundam. Appl. Toxicol.5, 816−831.(85) Ballatori, N., Gatmaitan, Z., and Truong, A. T. (1995) Impairedbiliary excretion and whole body elimination of methylmercury in ratswith congenital defect in biliary glutathione excretion. Hepatology 22,1469−1473.
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381836

(86) Ballatori, N. (2000) Molecular Mechanisms of Hepatic MetalTransport, in Molecular Biology and Toxicology of Metals (Zalups, R. K.,and Koropatnick, J., Ed.) pp 346−381, Taylor and Francis, London.(87) Parks, L. D., Zalups, R. K., and Barfuss, D. W. (1998)Heterogeneity of glutathione synthesis and secretion in the proximaltubule of the rabbit. Am. J. Physiol. 274, F924−931.(88) Schaub, T. P., Kartenbeck, J., Konig, J., Spring, H., Dorsam, J.,Staehler, G., Storkel, S., Thon, W. F., and Keppler, D. (1999) Expressionof the MRP2 gene-encoded conjugate export pump in human kidneyproximal tubules and in renal cell carcinoma. J. Am. Soc. Nephrol. 10,1159−1169.(89) Schaub, T. P., Kartenbeck, J., Konig, J., Vogel, O., Witzgall, R.,Kriz, W., and Keppler, D. (1997) Expression of the conjugate exportpump encoded by the mrp2 gene in the apical membrane of kidneyproximal tubules. J. Am. Soc. Nephrol. 8, 1213−1221.(90) Keppler, D., Leier, I., Jedlitschky, G., and Konig, J. (1998) ATP-dependent transport of glutathione S-conjugates by the multidrugresistance protein MRP1 and its apical isoform MRP2. Chem.-Biol.Interact. 111−112, 153−161.(91) Naganuma, A., Oda-Urano, N., Tanaka, T., and Imura, N. (1988)Possible role of hepatic glutathione in transport of methylmercury intomouse kidney. Biochem. Pharmacol. 37, 291−296.(92) Parks, L. D., Zalups, R. K., and Barfuss, D. W. (2000) Luminal andbasolateral membrane transport of glutathione in isolated perfused S(1),S(2), and S(3) segments of the rabbit proximal tubule. J. Am. Soc.Nephrol. 11, 1008−1015.(93) Zalups, R. K., and Barfuss, D. W. (1996) Nephrotoxicity ofinorganic mercury co-administrated with L-cysteine. Toxicology 109,15−29.(94) Zalups, R. K., and Lash, L. H. (1997) Binding of mercury in renalbrush-border and basolateral membrane-vesicles. Biochem. Pharmacol.53, 1889−1900.(95) Schafer, J. A., andWatkins, M. L. (1984) Transport of L-cystine inisolated perfused proximal straight tubules. Pflugers Arch. 401, 143−151.(96) Ballatori, N. (2002) Transport of toxic metals by molecularmimicry. Environ. Health Perspect. 110 (Suppl. 5), 689−694.(97) Aschner, M., Eberle, N. B., Goderie, S., and Kimelberg, H. K.(1990) Methylmercury uptake in rat primary astrocyte cultures: the roleof the neutral amino acid transport system. Brain Res. 521, 221−228.(98) Aschner, M., and Clarkson, T. W. (1989) Methyl mercury uptakeacross bovine brain capillary endothelial cells in vitro: the role of aminoacids. Pharmacol. Toxicol. 64, 293−297.(99) Kerper, L. E., Ballatori, N., and Clarkson, T. W. (1992)Methylmercury transport across the blood-brain barrier by an aminoacid carrier. Am. J. Physiol. 262, R761−765.(100) Hoffmeyer, R. E., Singh, S. P., Doonan, C. J., Ross, A. R., Hughes,R. J., Pickering, I. J., and George, G. N. (2006) Molecular mimicry inmercury toxicology. Chem. Res. Toxicol. 19, 753−759.(101) Zalups, R. K. (1998) Basolateral uptake of mercuric conjugatesof N-acetylcysteine and cysteine in the kidney involves the organic aniontransport system. J. Toxicol. Environ. Health, Part A 55, 13−29.(102) Zalups, R. K., and Barfuss, D. W. (1998) Small aliphaticdicarboxylic acids inhibit renal uptake of administered mercury. Toxicol.Appl. Pharmacol. 148, 183−193.(103) Zalups, R. K., and Diamond, G. L. (1987) Intrarenal distributionof mercury in the rat: effect of administered dose of mercuric chloride.Bull. Environ. Contam. Toxicol. 38, 67−72.(104) Zalups, R. K., and Barfuss, D. W. (1995) Pretreatment with p-aminohippurate inhibits the renal uptake and accumulation of injectedinorganic mercury in the rat. Toxicology 103, 23−35.(105) Dantzler, W. H., and Evans, K. K. (1996) Effect of alpha-KG inlumen on PAH transport by isolated perfused rabbit renal proximaltubules. Am. J. Physiol. 271, F521−526.(106) Dantzler, W. H., Evans, K. K., and Wright, S. H. (1995) Kineticsof interactions of para-aminohippurate, probenecid, cysteine conjugatesand N-acetyl cysteine conjugates with basolateral organic aniontransporter in isolated rabbit proximal renal tubules. J. Pharmacol. Exp.Ther. 272, 663−672.
(107) Ferrier, B., Martin, M., and Roch-Ramel, F. (1983) Effects of p-aminohippurate and pyrazinoate on the renal excretion of salicylate inthe rat: a micropuncture study. J. Pharmacol. Exp. Ther. 224, 451−458.(108) Pritchard, J. B. (1988) Coupled transport of p-aminohippurateby rat kidney basolateral membrane vesicles. Am. J. Physiol. 255, F597−604.(109) Pritchard, J. B., and Miller, D. S. (1993) Mechanisms mediatingrenal secretion of organic anions and cations. Physiol. Rev. 73, 765−796.(110) Shimomura, A., Chonko, A. M., and Grantham, J. J. (1981) Basisfor heterogeneity of para-aminohippurate secretion in rabbit proximaltubules. Am. J. Physiol. 240, F430−436.(111) Ullrich, K. J., Rumrich, G., Fritzsch, G., and Kloss, S. (1987)Contraluminal para-aminohippurate (PAH) transport in the proximaltubule of the rat kidney. I. Kinetics, influence of cations, anions, andcapillary preperfusion. Pflugers Arch. 409, 229−235.(112) Ullrich, K. J., Rumrich, G., Fritzsch, G., and Kloss, S. (1987)Contraluminal para-aminohippurate (PAH) transport in the proximaltubule of the rat kidney. II. Specificity: aliphatic dicarboxylic acids.Pflugers Arch. 408, 38−45.(113) Zalups, R. K. (1997) Enhanced renal outer medullary uptake ofmercury associated with uninephrectomy: implication of a luminalmechanism. J. Toxicol. Environ. Health 50, 173−194.(114) Ban, M., and de Ceaurriz, J. (1988) Probenecid-inducedprotection against acute hexachloro-1,3-butadiene and methyl mercurytoxicity to the mouse kidney. Toxicol. Lett. 40, 71−76.(115) Bhatnagar, V., Xu, G., Hamilton, B. A., Truong, D. M., Eraly, S.A., Wu, W., and Nigam, S. K. (2006) Analyses of 5′ regulatory regionpolymorphisms in human SLC22A6 (OAT1) and SLC22A8 (OAT3). J.Hum. Genet. 51, 575−580.(116) Nagle, M. A., Truong, D.M., Dnyanmote, A. V., Ahn, S. Y., Eraly,S. A., Wu, W., and Nigam, S. K. (2011) Analysis of three-dimensionalsystems for developing and mature kidneys clarifies the role of OAT1and OAT3 in antiviral handling. J. Biol. Chem. 286, 243−251.(117) Sweet, D. H., Miller, D. S., Pritchard, J. B., Fujiwara, Y., Beier, D.R., and Nigam, S. K. (2002) Impaired organic anion transport in kidneyand choroid plexus of organic anion transporter 3 (Oat3 (Slc22a8))knockout mice. J. Biol. Chem. 277, 26934−26943.(118) Vallon, V., Rieg, T., Ahn, S. Y., Wu,W., Eraly, S. A., andNigam, S.K. (2008) Overlapping in vitro and in vivo specificities of the organicanion transporters OAT1 and OAT3 for loop and thiazide diuretics. Am.J. Physiol. Renal Physiol. 294, F867−873.(119) Xu, G., Bhatnagar, V., Wen, G., Hamilton, B. A., Eraly, S. A., andNigam, S. K. (2005) Analyses of coding region polymorphisms in apicaland basolateral human organic anion transporter (OAT) genes [OAT1(NKT), OAT2, OAT3, OAT4, URAT (RST)]. Kidney Int. 68, 1491−1499.(120) Aslamkhan, A. G., Han, Y. H., Yang, X. P., Zalups, R. K., andPritchard, J. B. (2003) Human renal organic anion transporter 1-dependent uptake and toxicity of mercuric-thiol conjugates in Madin-Darby canine kidney cells. Mol. Pharmacol. 63, 590−596.(121) Koh, A. S., Simmons-Willis, T. A., Pritchard, J. B., Grassl, S. M.,and Ballatori, N. (2002) Identification of a mechanism by which themethylmercury antidotes N-acetylcysteine and dimercaptopropanesul-fonate enhance urinary metal excretion: transport by the renal organicanion transporter-1. Mol. Pharmacol. 62, 921−926.(122) Clarkson, T. W., and Magos, L. (1967) The effect of sodiummaleate on the renal deposition and excretion of mercury. Br. J.Pharmacol. Chemother. 31, 560−567.(123) Rogulski, J., and Angielski, S. (1963) Effect of maleic acid on thekidney. IV. Synthesis of amino acids in the kidney of maleate-treatedrats. Acta Biochim. Pol. 10, 133−139.(124)Wright, S. H., and Dantzler, W. H. (2004)Molecular and cellularphysiology of renal organic cation and anion transport. Physiol. Rev. 84,987−1049.(125) Pritchard, J. B., and Miller, D. S. (1991) Comparative insightsinto the mechanisms of renal organic anion and cation secretion. Am. J.Physiol. 261, R1329−1340.(126) Bridges, C. C., Joshee, L., and Zalups, R. K. (2008) MRP2 andthe DMPS- and DMSA-mediated elimination of mercury in TR(-) and
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381837

control rats exposed to thiol S-conjugates of inorganic mercury. Toxicol.Sci. 105, 211−220.(127) Bridges, C. C., Joshee, L., and Zalups, R. K. (2008) Multidrugresistance proteins and the renal elimination of inorganic mercurymediated by 2,3-dimercaptopropane-1-sulfonic acid and meso-2,3-dimercaptosuccinic acid. J. Pharmacol. Exp. Ther. 324, 383−390.(128) Bridges, C. C., Joshee, L., and Zalups, R. K. (2011) MRP2 andthe handling of mercuric ions in rats exposed acutely to inorganic andorganic species of mercury. Toxicol. Appl. Pharmacol. 251, 50−58.(129) Zalups, R. K., and Koropatnick, J. (2000) Temporal changes inmetallothionein gene transcription in rat kidney and liver: relationshipto content of mercury and metallothionein protein. J. Pharmacol. Exp.Ther. 295, 74−82.(130) Aposhian, H. V., Maiorino, R.M., Gonzalez-Ramirez, D., Zuniga-Charles, M., Xu, Z., Hurlbut, K. M., Junco-Munoz, P., Dart, R. C., andAposhian, M. M. (1995) Mobilization of heavy metals by newer,therapeutically useful chelating agents. Toxicology 97, 23−38.(131) Aposhian, H. V., Maiorino, R. M., Rivera, M., Bruce, D. C., Dart,R. C., Hurlbut, K. M., Levine, D. J., Zheng, W., Fernando, Q., Carter, D.,et al. (1992) Human studies with the chelating agents, DMPS and DMS.J. Toxicol. Clin. Toxicol. 30, 505−528.(132) Zalups, R. K. (1993) Influence of 2,3-dimercaptopropane-1-sulfonate (DMPS) and meso-2,3-dimercaptosuccinic acid (DMSA) onthe renal disposition of mercury in normal and uninephrectomized ratsexposed to inorganic mercury. J. Pharmacol. Exp. Ther. 267, 791−800.(133) Zalups, R. K., and Bridges, C. C. (2010) Seventy-five percentnephrectomy and the disposition of inorganic mercury in 2,3-dimercaptopropanesulfonic acid-treated rats lacking functional multi-drug-resistance protein 2. J. Pharmacol. Exp. Ther. 332, 866−875.(134) Zalups, R. K., Gelein, R. M., and Cernichiari, E. (1991) DMPS asa rescue agent for the nephropathy induced by mercuric chloride. J.Pharmacol. Exp. Ther. 256, 1−10.(135) Gabard, B., andWalser, R. (1979)Note on themetabolism of themercury chelating agent sodium 2,3-dimercaptopropane-1-sulfonate. J.Toxicol. Environ. Health 5, 759−764.(136) Maiorino, R. M., Weber, G. L., and Aposhian, H. V. (1986)Fluorometric determination of 2,3-dimercaptopropane-1-sulfonic acidand other dithiols by precolumn derivatization with bromobimane andcolumn liquid chromatography. J. Chromatogr. 374, 297−310.(137) Diamond, G. L., Klotzbach, J. M., and Stewart, J. R. (1988)Complexing activity of 2,3-dimercapto-1-propanesulfonate and itsdisulfide auto-oxidation product in rat kidney. J. Pharmacol. Exp. Ther.246, 270−274.(138) Klotzbach, J. M., and Diamond, G. L. (1988) Complexingactivity and excretion of 2,3-dimercapto-1-propane sulfonate in ratkidney. Am. J. Physiol. 254, F871−878.(139) Stewart, J. R., and Diamond, G. L. (1988) In vivo renal tubularsecretion and metabolism of the disulfide of 2,3-dimercaptopropane-1-sulfonate. Drug Metab. Dispos. 16, 189−195.(140) George, G. N., Prince, R. C., Gailer, J., Buttigieg, G. A., Denton,M. B., Harris, H. H., and Pickering, I. J. (2004) Mercury binding to thechelation therapy agents DMSA and DMPS and the rational design ofcustom chelators for mercury. Chem. Res. Toxicol. 17, 999−1006.(141) Lash, L. H., and Zalups, R. K. (1994) Activities of enzymesinvolved in renal cellular glutathione metabolism after uninephrectomyin the rat. Arch. Biochem. Biophys. 309, 129−138.(142) Zalups, R. K., Barfuss, D. W., and Lash, L. H. (1999)Relationships between alterations in glutathione metabolism and thedisposition of inorganic mercury in rats: effects of biliary ligation andchemically induced modulation of glutathione status. Chem.-Biol.Interact. 123, 171−195.(143) Zalups, R. K., Barfuss, D. W., and Lash, L. H. (1999) Dispositionof inorganic mercury following biliary obstruction and chemicallyinduced glutathione depletion: dispositional changes one hour after theintravenous administration of mercuric chloride. Toxicol. Appl.Pharmacol. 154, 135−144.(144) Zalups, R. K., and Lash, L. H. (1990) Effects of uninephrectomyand mercuric chloride on renal glutathione homeostasis. J. Pharmacol.Exp. Ther. 254, 962−970.
(145) Fu, J., Hoffmeyer, R. E., Pushie, M. J., Singh, S. P., Pickering, I. J.,and George, G. N. (2011) Towards a custom chelator for mercury:evaluation of coordination environments by molecular modeling. J. Biol.Inorg. Chem. 16, 15−24.(146) Zalups, R. K., Parks, L. D., Cannon, V. T., and Barfuss, D. W.(1998) Mechanisms of action of 2,3-dimercaptopropane-1-sulfonateand the transport, disposition, and toxicity of inorganic mercury inisolated perfused segments of rabbit proximal tubules. Mol. Pharmacol.54, 353−363.(147) Deeley, R. G., Westlake, C., and Cole, S. P. (2006)Transmembrane transport of endo- and xenobiotics by mammalianATP-binding cassette multidrug resistance proteins. Physiol. Rev. 86,849−899.(148) Zhou, S. F.,Wang, L. L., Di, Y. M., Xue, C. C., Duan,W., Li, C. G.,and Li, Y. (2008) Substrates and inhibitors of human multidrugresistance associated proteins and the implications in drug development.Curr. Med. Chem. 15, 1981−2039.(149) Aremu, D. A., Madejczyk, M. S., and Ballatori, N. (2008) N-acetylcysteine as a potential antidote and biomonitoring agent ofmethylmercury exposure. Environ. Health Perspect. 116, 26−31.
Chemical Research in Toxicology Review
dx.doi.org/10.1021/tx3001847 | Chem. Res. Toxicol. 2012, 25, 1825−18381838