rangtp is required for meiotic spindle organization and ... · journal of cell science 124,...
TRANSCRIPT

RanGTP is required for meiotic spindle organizationand the initiation of embryonic developmentin Drosophila
J. Cesario and K. S. McKim*Waksman Institute and Department of Genetics, Rutgers, the State University of New Jersey, 190 Frelinghuysen RD, Piscataway NJ 08854-8020,USA
*Author for correspondence ([email protected])
Accepted 4 July 2011Journal of Cell Science 124, 3797–3810� 2011. Published by The Company of Biologists Ltddoi: 10.1242/jcs.084855
SummaryRanGTP is important for chromosome-dependent spindle assembly in Xenopus extracts. Here we report on experiments to determine therole of the Ran pathway on microtubule dynamics in Drosophila oocytes and embryos. Females expressing a dominant-negative form of
Ran have fertility defects, suggesting that RanGTP is required for normal fertility. This is not, however, because of a defect inacentrosomal meiotic spindle assembly. Therefore, RanGTP does not appear to be essential or sufficient for the formation of theacentrosomal spindle. Instead, the most important function of the Ran pathway in spindle assembly appears to be in the tapering of
microtubules at the spindle poles, which might be through regulation of proteins such as TACC and the HURP homolog, Mars. Oneconsequence of this spindle organization defect is an increase in the nondisjunction of achiasmate chromosomes. However, the meioticdefects are not severe enough to cause the decreased fertility. Reductions in fertility occur because RanGTP has a role in microtubule
assembly that is not directly nucleated by the chromosomes. This includes microtubules nucleated from the sperm aster, which arerequired for pronuclear fusion. We propose that following nuclear envelope breakdown, RanGTP is released from the nucleus andcreates a cytoplasm that is activated for assembling microtubules, which is important for processes such as pronuclear fusion. Aroundthe chromosomes, however, RanGTP might be redundant with other factors such as the chromosome passenger complex.
Key words: Meiosis, Mitosis, Microtubule, Drosophila melanogaster, Acentrosomal spindle, Chromosome segregation
IntroductionRan is a member of the Ras family of small GTP-bindingproteins. It was originally discovered for its role in shuttlingproteins with nuclear localization sequences (NLS) into the
nucleus (Moore and Blobel, 1993). Ran cycles from an activestate, RanGTP, to an inactive state, RanGDP. The conversion ofRanGDP to RanGTP is stimulated by the chromatin bound
guanine nucleotide exchange factor RCC1 (Bischoff andPonstingl, 1991). Conversely, the conversion of RanGTP toRanGDP is facilitated by the cytoplasmic GTPase-activating
protein RanGAP (Bischoff et al., 1994). During G2, the nuclearenvelope creates a barrier where active RanGTP can only befound within the nucleus, because RCC1 is chromatin bound. Theimportin complex, which consists of importin a and importin b,
is capable of binding proteins with NLS and transporting theminto the nucleus. Once inside the nucleus, RanGTP bindsimportin b, releasing importin a and NLS containing proteins
(Clarke and Zhang, 2008).
Ran also has a role in spindle assembly by releasing spindleassembly factors from the repressive importin complex (Kalab
and Heald, 2008). The production of RanGTP near chromatin andconversion to RanGDP in the cytoplasm can lead to the formationof a gradient of active Ran that is capable of triggering
chromosome-mediated spindle assembly (Caudron et al., 2005).The role of RanGTP in chromosome-mediated spindle assemblyhas been most clearly shown by its activity in Xenopus laevis egg
extracts that lack centrosomes. Chromatin-mediated microtubuleassembly depends on the presence of RanGTP in Xenopus
extracts (Carazo-Salas et al., 1999). Similarly, depletion of RCC1results in a failure to form microtubule asters. Addition of
RanGTP to these RCC1-depleted eggs is sufficient to induce self-
organization of microtubule asters (Ohba et al., 1999). Disruptionof RanGTP levels also affects mitotic spindle assembly in
mammalian (Clarke and Zhang, 2008; Kalab et al., 2006),Drosophila melanogaster (Silverman-Gavrila and Wilde, 2006)
and Caenorhabditis elegans (Askjaer et al., 2002; Bamba et al.,2002) cells. These results suggest that RanGTP is a major
contributor to spindle assembly.
We have undertaken an analysis of Ran function in the
Drosophila oocyte because several aspects of oogenesis and
embryogenesis depend on microtubule dynamics (Dix and Raff,2007; Roth and Lynch, 2009). In Drosophila oocytes, as in many
oocytes, meiosis is acentrosomal. Spindle assembly occurswithout the guidance of the microtubule organizing centers at
the poles. In this situation, the chromosomes play an important
role in spindle assembly. Nuclear envelope breakdown (NEB) isfollowed by the accumulation of microtubules around the
chromosomes (Matthies et al., 1996; Theurkauf and Hawley,1992). The subsequent bundling and tapering of these
microtubules by motor proteins results in a bipolar spindle.
Thus, Drosophila oocyte chromosomes carry a signal thatpromotes spindle assembly when released into the cytoplasm
Research Article 3797
Journ
alof
Cell
Scie
nce

upon NEB. However, it is unclear, what are the components of
this signal.
Meiosis in Drosophila arrests at the first division (Theurkauf
et al., 1993). When the oocyte then moves down the oviduct, it
becomes activated and the two meiotic divisions are completed.
Independently, fertilization occurs and the sperm centriole
recruits microtubules that are required to bring together the
male and female pronuclei. Finally, the nuclear membranes of the
two nuclei fuse prior to the first mitotic division. All these events
depend on maternally contributed proteins and are thus a function
of the oocyte. We have examined the role of the Ran pathway in
these early developmental processes. We found that RanGTP has
a role in pronuclear fusion in the embryo, is active in promoting
microtubule assembly in the oocyte cytoplasm, but it might not
be required for their recruitment of microtubules by the meiotic
chromosomes.
ResultsGeneration of dominant ran mutants
Ran is required for mitosis (Silverman-Gavrila and Wilde, 2006)
and a mutation in the ran gene (G0075) causes lethality in
Drosophila (Peter et al., 2002). Furthermore, mutations in the
Drosophila RCC1 homolog, Bj1, cause lethality, and germline
clones do not make oocytes (Shi and Skeath, 2004) (K.S.M.,
unpublished results). Because these properties make it impossible
to study ran mutations in oocytes, we generated mutations
predicted to be dominant alleles of ran. Previous studies in a
variety of systems have characterized mutations that lock Ran in
either the GDP (inactive) or GTP (active) states (Kahana and
Cleveland, 1999; Trieselmann and Wilde, 2002). Because Ran is
highly conserved, these same changes can be made in
Drosophila. Transgenes were made by fusing the coding region
of the wild-type ran or mutant variants to three copies of the HA
epitope tag at the N-terminus. They were also put under the
control of the UASP promoter, which allows for inducible
germline expression regulated by a second transgene expressing
GAL4 (Rorth, 1998). To generate a GDP-locked mutant, the
P{w+; UASP:ranT24N} transgene was constructed with an amino
acid substitution of threonine to asparagine at position 24,
hereafter referred to as ranT24N. To generate a GTP-locked
mutant, the P{w+; UASP:ranQ69L} transgene was constructed
with an amino acid substitution of glutamine to leucine at
position 69, hereafter referred to as ranQ69L. For each allele, at
least three transgenic lines were examined for expression levels
and phenotypes. For each experiment, flies with these mutations
were compared with flies expressing a wild-type transgene
(P{w+; UASP:ran+}), hereafter referred to as ran+.
Expression of ranT24N has a dominant-negative effect
To determine whether expression of the mutant forms of ran
would cause lethality similar to the loss-of-function mutant, weexpressed the transgenes using P{tubP-GAL4}, which inducesubiquitous expression of UAS transgenes (Lee and Luo, 1999).
Expression of ran+ had no effect on viability and was able torescue the lethality of a ran mutation (Table 1). Furthermore,
dividing neuroblasts from third instar larvae exhibited properlyassembled spindles with no detectable abnormalities during
metaphase and anaphase (supplementary material Fig. S1). Thelocation of wild-type Ran was examined using antibodies to theHA epitope tag that was fused at the N-terminus of the ran
transgenes. Ran was nuclear during interphase and thenoverlapped with the spindle during metaphase and anaphase
(supplementary material Fig. S1). Similar to the localizationpattern in embryos (Trieselmann and Wilde, 2002), Ran was notdetected on the chromosomes. Unlike ran+, ubiquitous
expression of ranT24N or ranQ69L resulted in lethality of theembryos or at an early stage of larval development (Table 1).
These results suggest that expression of ranT24N or ranQ69L has adominant effect and disrupts the normal functioning of the Ranpathway.
Maternal expression of ranT24N and ranQ69L causes sterility
To examine the function of Ran in oogenesis and embryogenesis,
the ran transgenes were expressed using the P{GAL4:VP16-
nos.UTR}MVD1 driver, and then genetic assays were performed
to measure fertility and the frequency of X-chromosomenondisjunction. This driver typically overexpresses UASPtransgenes in oocytes (Jang et al., 2007; Van Doren et al.,
1998). Differences in expression levels between differentinsertion lines were assayed by western blotting and found to
be minimal (data not shown). When the wild-type ran transgenewas expressed, levels of fertility were normal (Table 2).
Therefore, expressing wild-type Ran does not have deleteriouseffects on embryonic development.
Expression of ranT24N in the female germline caused a drastic
reduction in fertility, with an average of only 6.8 progeny perfemale parent compared with 62.1 progeny per female expressing
wild-type Ran (Table 2). P{GAL4:VP16-nos.UTR}MVD1 alsodrives expression in the male germline. Males carrying this driverand ranT24N were sterile, suggesting that RanGTP has an essential
role in male meiosis. Expression of ranQ69L in the oocyte resultedin complete sterility (Table 2). These effects of ranT24N and
ranQ69L on fertility indicate that there is an important role forRanGTP in either meiosis, fertilization or the embryonicdivisions. Therefore, we examined oocytes and embryos in
more detail in order to determine why the ran mutants are sterile.
Table 1. Effect of ran mutants on viability
Transgene Progeny expressing Ran Progeny not expressing Ran
ran+ a 755 326ranT24N a 0 464ranQ69L a 0 1405ranG0075/Y; ran+ b 66 307
aEach transgene was expressed by crossing to P{tubP-GAL4}/TM6, Tb. Progeny expressing a ran transgene were Tb+. For each transgene at least twoindependent insertions were scored, both of which gave similar results. Stocks containing the ran mutant transgenes are viable and fertile, indicating that thelethality is specific and depends on the presence of the Gal4 driver.
branG0075/FM7 females were crossed to P{w+; UASP:ran+}/+; P{tubP-GAL4}/+ males. 25% of the progeny were expected to inherit both the driver andtransgene.
Journal of Cell Science 124 (22)3798
Journ
alof
Cell
Scie
nce
Two components of the Ran pathway, RCC1 and RanGAP,are present in the Drosophila oocyte
The Drosophila oocyte develops in a cyst along with 15 nurse
cells. Over the course of 3–5 days, the oocyte grows in size whilein a diplotene–diakinesis-like state, eventually receiving most ofthe nurse cell components by the time they reach stages 13 to 14.
Following NEB in the oocyte, microtubules accumulate aroundthe chromosomes, which are bundled together into a karyosome
(Matthies et al., 1996; Theurkauf and Hawley, 1992). This isfollowed by the extension of poles and lengthening of the spindle.
Our previous work has shown that the central spindle is importantfor organizing bipolarity (Jang et al., 2005), which can be
detected by staining for Subito, a kinesin 6 that localizes to theantiparallel microtubules of the central spindle. Meiosis arrests atmetaphase I until the oocyte passes down the oviduct and
becomes activated, at which point the two meiotic divisionsoccur.
A gradient of RanGTP with the concentration highest near thechromosomes can be established if RCC1 is enriched on the
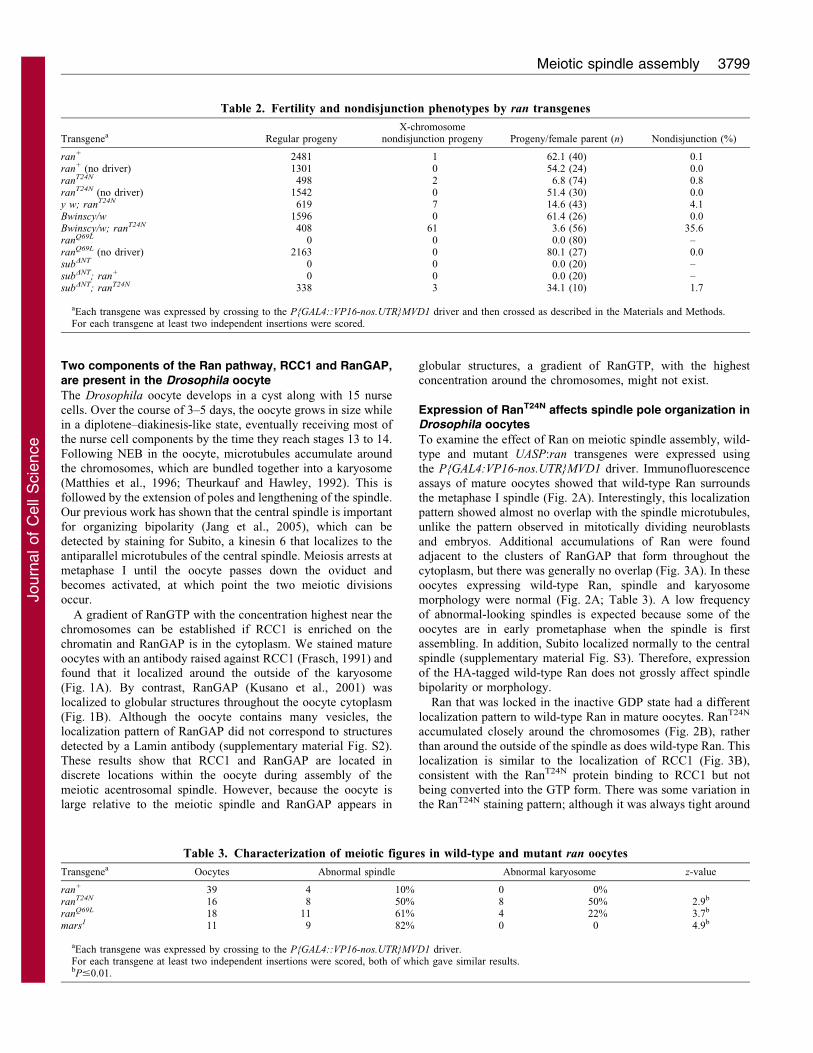
chromatin and RanGAP is in the cytoplasm. We stained matureoocytes with an antibody raised against RCC1 (Frasch, 1991) andfound that it localized around the outside of the karyosome
(Fig. 1A). By contrast, RanGAP (Kusano et al., 2001) waslocalized to globular structures throughout the oocyte cytoplasm
(Fig. 1B). Although the oocyte contains many vesicles, thelocalization pattern of RanGAP did not correspond to structures
detected by a Lamin antibody (supplementary material Fig. S2).These results show that RCC1 and RanGAP are located indiscrete locations within the oocyte during assembly of the
meiotic acentrosomal spindle. However, because the oocyte islarge relative to the meiotic spindle and RanGAP appears in
globular structures, a gradient of RanGTP, with the highestconcentration around the chromosomes, might not exist.
Expression of RanT24N affects spindle pole organization inDrosophila oocytes
To examine the effect of Ran on meiotic spindle assembly, wild-type and mutant UASP:ran transgenes were expressed usingthe P{GAL4:VP16-nos.UTR}MVD1 driver. Immunofluorescence
assays of mature oocytes showed that wild-type Ran surroundsthe metaphase I spindle (Fig. 2A). Interestingly, this localizationpattern showed almost no overlap with the spindle microtubules,unlike the pattern observed in mitotically dividing neuroblasts
and embryos. Additional accumulations of Ran were foundadjacent to the clusters of RanGAP that form throughout thecytoplasm, but there was generally no overlap (Fig. 3A). In these
oocytes expressing wild-type Ran, spindle and karyosomemorphology were normal (Fig. 2A; Table 3). A low frequencyof abnormal-looking spindles is expected because some of the
oocytes are in early prometaphase when the spindle is firstassembling. In addition, Subito localized normally to the centralspindle (supplementary material Fig. S3). Therefore, expression
of the HA-tagged wild-type Ran does not grossly affect spindlebipolarity or morphology.
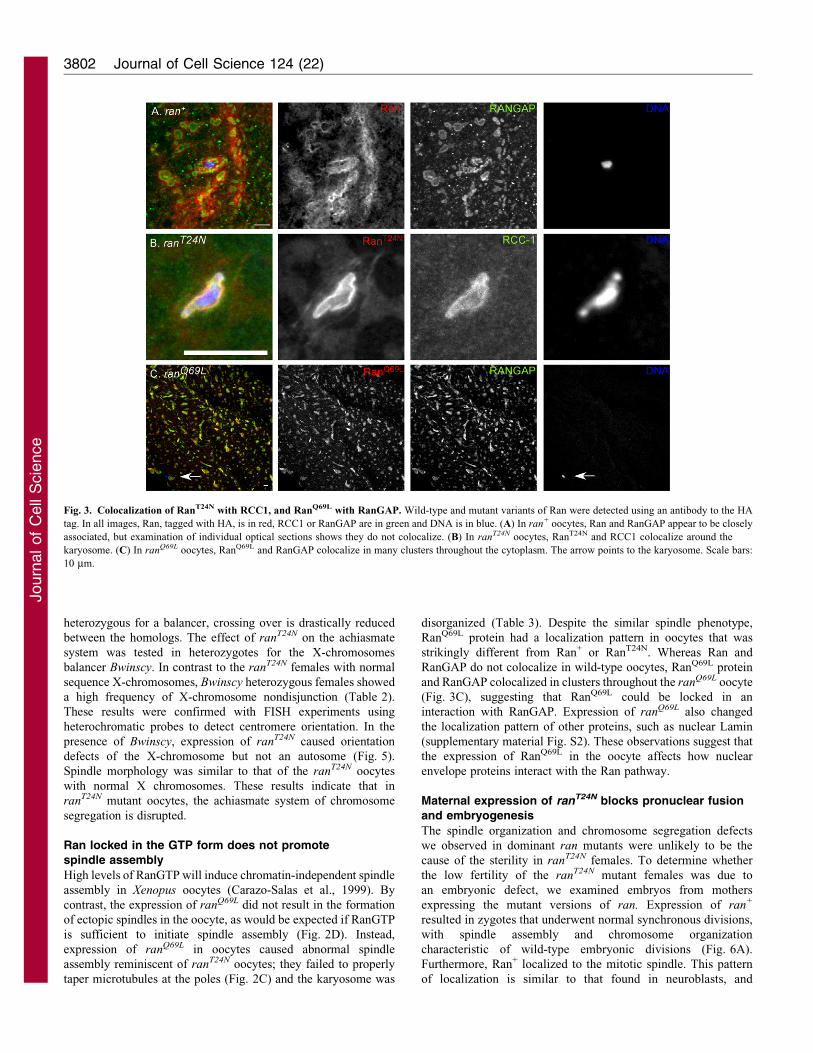
Ran that was locked in the inactive GDP state had a different
localization pattern to wild-type Ran in mature oocytes. RanT24N
accumulated closely around the chromosomes (Fig. 2B), ratherthan around the outside of the spindle as does wild-type Ran. Thislocalization is similar to the localization of RCC1 (Fig. 3B),
consistent with the RanT24N protein binding to RCC1 but notbeing converted into the GTP form. There was some variation inthe RanT24N staining pattern; although it was always tight around
Table 2. Fertility and nondisjunction phenotypes by ran transgenes
Transgenea Regular progenyX-chromosome
nondisjunction progeny Progeny/female parent (n) Nondisjunction (%)
ran+ 2481 1 62.1 (40) 0.1ran+ (no driver) 1301 0 54.2 (24) 0.0ranT24N 498 2 6.8 (74) 0.8ranT24N (no driver) 1542 0 51.4 (30) 0.0y w; ranT24N 619 7 14.6 (43) 4.1Bwinscy/w 1596 0 61.4 (26) 0.0Bwinscy/w; ranT24N 408 61 3.6 (56) 35.6ranQ69L 0 0 0.0 (80) –ranQ69L (no driver) 2163 0 80.1 (27) 0.0subDNT 0 0 0.0 (20) –subDNT; ran+ 0 0 0.0 (20) –subDNT; ranT24N 338 3 34.1 (10) 1.7
aEach transgene was expressed by crossing to the P{GAL4::VP16-nos.UTR}MVD1 driver and then crossed as described in the Materials and Methods.For each transgene at least two independent insertions were scored.
Table 3. Characterization of meiotic figures in wild-type and mutant ran oocytes
Transgenea Oocytes Abnormal spindle Abnormal karyosome z-value
ran+ 39 4 10% 0 0%ranT24N 16 8 50% 8 50% 2.9b
ranQ69L 18 11 61% 4 22% 3.7b
mars1 11 9 82% 0 0 4.9b
aEach transgene was expressed by crossing to the P{GAL4::VP16-nos.UTR}MVD1 driver.For each transgene at least two independent insertions were scored, both of which gave similar results.bP#0.01.
Meiotic spindle assembly 3799
Journ
alof
Cell
Scie
nce

the karyosome (Fig. 3B), in some images there was staining away
from DNA as well (Fig. 2B). Oocytes expressing ranT24N did not
appear to have a problem initiating the assembly of microtubules
around the chromosomes or building a bipolar spindle. However,
the ranT24N-expressing oocytes had an increased frequency of
abnormal spindle and karyosome organization (Table 3). The
microtubules were often not tapered at the spindle poles
(Fig. 2B). Furthermore, the chromosomes were frequently
disorganized and failed to condense into a single round or oval
karyosome (supplementary material Fig. S4). By contrast, Subito
localized correctly (supplementary material Fig. S3), suggesting
the central spindle was able to form in these mutants. These
results suggest that the Ran pathway has a role in organizing the
meiosis I spindle poles, but it might not be essential for initiating
chromosome-based microtubule assembly or for regulating
central spindle proteins like Subito.
Because RanGTP promotes spindle assembly through the
release of spindle assembly factors, we tested whether ranT24N
mutant oocytes showed evidence of downregulating proteins
known to be regulated by the Ran pathway. Mars is the
Drosophila homolog of HURP, a spindle assembly factor
regulated by RanGTP (Wilde, 2006) and has been shown to
have a role in the attachment of the centrosome to the mitotic
spindle during Drosophila embryogenesis (Tan et al., 2008; Yang
and Fan, 2008; Zhang et al., 2009). To study the role of Mars in
meiotic spindle assembly, we examined mar1, which is a null
allele that deletes part of the coding region (Tan et al., 2008).
Like ranT24N mutants, mature oocytes homozygous for the mars1
mutation were able to assemble a bipolar spindle but failed to
properly taper the microtubules at the poles (Fig. 2E). To test
whether these similarities could be the result of ranT24N mutant
oocytes failing to activate Mars, we stained ranT24N mutant
oocytes with an antibody against Mars (Tan et al., 2008). In wild-
type or ran+-expressing oocytes, Mars colocalized with tubulin,
except at the spindle poles and the central spindle (Fig. 4A). By
contrast, approximately 50% of oocytes expressing ranT24N failed
to localize Mars to the meiotic spindle (Fig. 4B–D), which was
significantly different from ran+ oocytes (z53.6, P#0.01).
Therefore, a Mars localization defect might contribute to the
spindle-tapering defect observed in ranT24N mutant oocytes.
These results are consistent with the conclusion that the RanGTP
pathway is not essential for the initiation of acentrosomal spindle
assembly in Drosophila oocytes, but might have a role in tapering
the poles.
Another spindle assembly factor regulated by the RanGTP
pathway is the microtubule-associated factor Transforming acidic
coiled-coil, or TACC (Kalab and Heald, 2008) (see Discussion).
TACC localizes to the poles of the meiotic (Cullen and Ohkura,
2001) and mitotic spindle (Giet et al., 2002) where it contributes
to the localization of Msps (Minispindles). To examine the
localization of TACC during female meiosis, we expressed a
GFP fusion gene under the control of a ubiquitin promoter
(Gergely et al., 2000). As expected, in wild-type oocytes TACC
localized to the poles of most metaphase I spindles (Fig. 4E,F).
There was some variation in this pattern, with TACC tending to
be less focused at the poles of shorter spindles. By contrast, in all
Fig. 1. Localization of RCC1 and RanGAP in mature (stage 14) oocytes. DNA is in blue and tubulin is in green. (A) Wild-type oocyte stained with RCC1
antibody (red). (B) Wild-type oocyte stained with RanGAP antibody (red). (C) A low magnification view of the same oocyte as in B showing the clusters of
RanGAP located throughout the ooplasm. The arrow points to the karyosome. Scale bars: 10 mm.
Journal of Cell Science 124 (22)3800
Journ
alof
Cell
Scie
nce

ranT24N oocytes TACC failed to show enrichment towards the
poles and was present next to the DNA (Fig. 4G,H). The failure
to properly localize both Mars and TACC in ranT24N mutant
oocytes is consistent with loss of RanGTP activity, but the
mislocalization of spindle assembly factors is not severe enough
to prevent spindle assembly.
Achiasmate chromosome segregation is abnormal in
ranT24N mutants
To determine whether the spindle organization defects in the ran
mutant females were associated with errors in chromosome
segregation, two genetic crosses were performed to measure the
frequency of X-chromosome nondisjunction. These experiments
measured the frequency of chiasmate and achiasmate chromosome
nondisjunction. Segregation of chiasmate chromosomes was
measured in females homozygous for normal sequence
X-chromosomes that were expected to have a crossover in
greater than 95% of meioses (Baker and Hall, 1976). In these
experiments, we found a low frequency of nondisjunction among
the few progeny from ranT24N-expressing mothers, showing that
X-chromosome segregation was not substantially affected by
loss of RanGTP (Table 2). These results are consistent with
fluorescence in situ hybridization (FISH) experiments, which
showed that ranT24N and ranQ69L mutants were able to properly
orient their homologous chromosomes at metaphase I
(supplementary material Fig. S5). Thus, although the spindles
are not properly tapered and the karyosome is disorganized in ran
mutants, this does not affect biorientation or segregation of
chiasmate chromosomes.
The achiasmate system in Drosophila females efficiently
segregates homologous chromosomes lacking a crossover
(Hawley and Theurkauf, 1993). For example, in a female
Fig. 2. Effect of Ran on spindle
morphology in mature oocytes. The
transgenes in these and all subsequent
experiments were expressed using the
P{GAL4::VP-nos.UTR}MVD1 driver.
DNA is in blue, Ran proteins are in red
and tubulin is in green. Ran was
detected using an antibody to the HA
tag fused to either wild-type ran
(A), ranT24N (B) or ranQ69L (C,D). The
images in A–C are high magnification
images centered on the karyosome.
The image in D is of the same oocyte
as in C but at lower magnification to
show the localization of mutant
RanQ69L in the oocyte cytoplasm. The
arrow in D points to the karyosome.
(E) In mars1 mutant oocytes, the
microtubules often fail to be properly
tapered at the spindle poles. Scale bars:
10 mm.
Meiotic spindle assembly 3801
Journ
alof
Cell
Scie
nce
heterozygous for a balancer, crossing over is drastically reduced
between the homologs. The effect of ranT24N on the achiasmate
system was tested in heterozygotes for the X-chromosomes
balancer Bwinscy. In contrast to the ranT24N females with normal
sequence X-chromosomes, Bwinscy heterozygous females showed
a high frequency of X-chromosome nondisjunction (Table 2).
These results were confirmed with FISH experiments using
heterochromatic probes to detect centromere orientation. In the
presence of Bwinscy, expression of ranT24N caused orientation
defects of the X-chromosome but not an autosome (Fig. 5).
Spindle morphology was similar to that of the ranT24N oocytes
with normal X chromosomes. These results indicate that in
ranT24N mutant oocytes, the achiasmate system of chromosome
segregation is disrupted.
Ran locked in the GTP form does not promote
spindle assembly
High levels of RanGTP will induce chromatin-independent spindle
assembly in Xenopus oocytes (Carazo-Salas et al., 1999). By
contrast, the expression of ranQ69L did not result in the formation
of ectopic spindles in the oocyte, as would be expected if RanGTP
is sufficient to initiate spindle assembly (Fig. 2D). Instead,
expression of ranQ69L in oocytes caused abnormal spindle
assembly reminiscent of ranT24N oocytes; they failed to properly
taper microtubules at the poles (Fig. 2C) and the karyosome was
disorganized (Table 3). Despite the similar spindle phenotype,
RanQ69L protein had a localization pattern in oocytes that was
strikingly different from Ran+ or RanT24N. Whereas Ran and
RanGAP do not colocalize in wild-type oocytes, RanQ69L protein
and RanGAP colocalized in clusters throughout the ranQ69L oocyte
(Fig. 3C), suggesting that RanQ69L could be locked in an
interaction with RanGAP. Expression of ranQ69L also changed
the localization pattern of other proteins, such as nuclear Lamin
(supplementary material Fig. S2). These observations suggest that
the expression of RanQ69L in the oocyte affects how nuclear
envelope proteins interact with the Ran pathway.
Maternal expression of ranT24N blocks pronuclear fusion
and embryogenesis
The spindle organization and chromosome segregation defects
we observed in dominant ran mutants were unlikely to be the
cause of the sterility in ranT24N females. To determine whether
the low fertility of the ranT24N mutant females was due to
an embryonic defect, we examined embryos from mothers
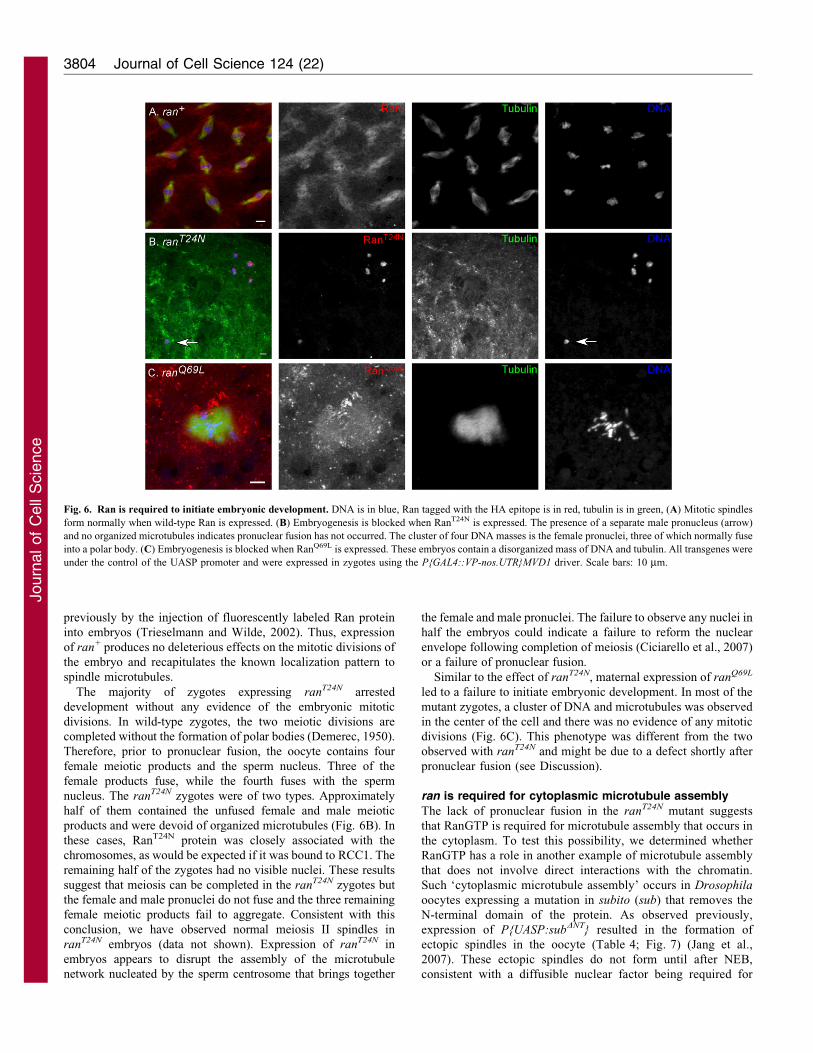
expressing the mutant versions of ran. Expression of ran+
resulted in zygotes that underwent normal synchronous divisions,
with spindle assembly and chromosome organization
characteristic of wild-type embryonic divisions (Fig. 6A).
Furthermore, Ran+ localized to the mitotic spindle. This pattern
of localization is similar to that found in neuroblasts, and
Fig. 3. Colocalization of RanT24N with RCC1, and RanQ69L with RanGAP. Wild-type and mutant variants of Ran were detected using an antibody to the HA
tag. In all images, Ran, tagged with HA, is in red, RCC1 or RanGAP are in green and DNA is in blue. (A) In ran+ oocytes, Ran and RanGAP appear to be closely
associated, but examination of individual optical sections shows they do not colocalize. (B) In ranT24N oocytes, RanT24N and RCC1 colocalize around the
karyosome. (C) In ranQ69L oocytes, RanQ69L and RanGAP colocalize in many clusters throughout the cytoplasm. The arrow points to the karyosome. Scale bars:
10 mm.
Journal of Cell Science 124 (22)3802
Journ
alof
Cell
Scie
nce
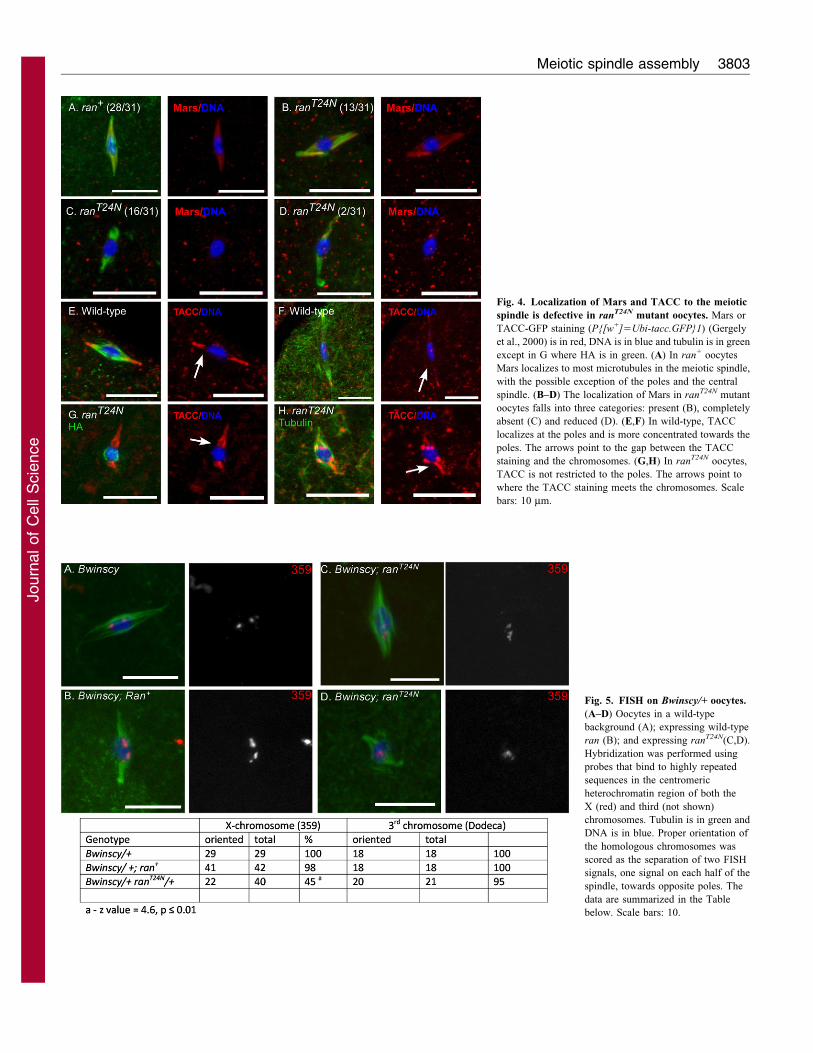
Fig. 4. Localization of Mars and TACC to the meiotic
spindle is defective in ranT24N mutant oocytes. Mars or
TACC-GFP staining (P{[w+]5Ubi-tacc.GFP}1) (Gergely
et al., 2000) is in red, DNA is in blue and tubulin is in green
except in G where HA is in green. (A) In ran+ oocytes
Mars localizes to most microtubules in the meiotic spindle,
with the possible exception of the poles and the central
spindle. (B–D) The localization of Mars in ranT24N mutant
oocytes falls into three categories: present (B), completely
absent (C) and reduced (D). (E,F) In wild-type, TACC
localizes at the poles and is more concentrated towards the
poles. The arrows point to the gap between the TACC
staining and the chromosomes. (G,H) In ranT24N oocytes,
TACC is not restricted to the poles. The arrows point to
where the TACC staining meets the chromosomes. Scale
bars: 10 mm.
Fig. 5. FISH on Bwinscy/+ oocytes.
(A–D) Oocytes in a wild-type
background (A); expressing wild-type
ran (B); and expressing ranT24N(C,D).
Hybridization was performed using
probes that bind to highly repeated
sequences in the centromeric
heterochromatin region of both the
X (red) and third (not shown)
chromosomes. Tubulin is in green and
DNA is in blue. Proper orientation of
the homologous chromosomes was
scored as the separation of two FISH
signals, one signal on each half of the
spindle, towards opposite poles. The
data are summarized in the Table
below. Scale bars: 10.
Meiotic spindle assembly 3803
Journ
alof
Cell
Scie
nce
previously by the injection of fluorescently labeled Ran protein
into embryos (Trieselmann and Wilde, 2002). Thus, expression
of ran+ produces no deleterious effects on the mitotic divisions of
the embryo and recapitulates the known localization pattern to
spindle microtubules.
The majority of zygotes expressing ranT24N arrested
development without any evidence of the embryonic mitotic
divisions. In wild-type zygotes, the two meiotic divisions are
completed without the formation of polar bodies (Demerec, 1950).
Therefore, prior to pronuclear fusion, the oocyte contains four
female meiotic products and the sperm nucleus. Three of the
female products fuse, while the fourth fuses with the sperm
nucleus. The ranT24N zygotes were of two types. Approximately
half of them contained the unfused female and male meiotic
products and were devoid of organized microtubules (Fig. 6B). In
these cases, RanT24N protein was closely associated with the
chromosomes, as would be expected if it was bound to RCC1. The
remaining half of the zygotes had no visible nuclei. These results
suggest that meiosis can be completed in the ranT24N zygotes but
the female and male pronuclei do not fuse and the three remaining
female meiotic products fail to aggregate. Consistent with this
conclusion, we have observed normal meiosis II spindles in
ranT24N embryos (data not shown). Expression of ranT24N in
embryos appears to disrupt the assembly of the microtubule
network nucleated by the sperm centrosome that brings together
the female and male pronuclei. The failure to observe any nuclei in
half the embryos could indicate a failure to reform the nuclear
envelope following completion of meiosis (Ciciarello et al., 2007)
or a failure of pronuclear fusion.
Similar to the effect of ranT24N, maternal expression of ranQ69L
led to a failure to initiate embryonic development. In most of the
mutant zygotes, a cluster of DNA and microtubules was observed
in the center of the cell and there was no evidence of any mitotic
divisions (Fig. 6C). This phenotype was different from the two
observed with ranT24N and might be due to a defect shortly after
pronuclear fusion (see Discussion).
ran is required for cytoplasmic microtubule assembly
The lack of pronuclear fusion in the ranT24N mutant suggests
that RanGTP is required for microtubule assembly that occurs in
the cytoplasm. To test this possibility, we determined whether
RanGTP has a role in another example of microtubule assembly
that does not involve direct interactions with the chromatin.
Such ‘cytoplasmic microtubule assembly’ occurs in Drosophila
oocytes expressing a mutation in subito (sub) that removes the
N-terminal domain of the protein. As observed previously,
expression of P{UASP:subDNT} resulted in the formation of
ectopic spindles in the oocyte (Table 4; Fig. 7) (Jang et al.,
2007). These ectopic spindles do not form until after NEB,
consistent with a diffusible nuclear factor being required for
Fig. 6. Ran is required to initiate embryonic development. DNA is in blue, Ran tagged with the HA epitope is in red, tubulin is in green, (A) Mitotic spindles
form normally when wild-type Ran is expressed. (B) Embryogenesis is blocked when RanT24N is expressed. The presence of a separate male pronucleus (arrow)
and no organized microtubules indicates pronuclear fusion has not occurred. The cluster of four DNA masses is the female pronuclei, three of which normally fuse
into a polar body. (C) Embryogenesis is blocked when RanQ69L is expressed. These embryos contain a disorganized mass of DNA and tubulin. All transgenes were
under the control of the UASP promoter and were expressed in zygotes using the P{GAL4::VP-nos.UTR}MVD1 driver. Scale bars: 10 mm.
Journal of Cell Science 124 (22)3804
Journ
alof
Cell
Scie
nce

their formation. Ectopic spindles cluster and form in many
regions of the mutant oocytes without direct contact with
chromosomes.
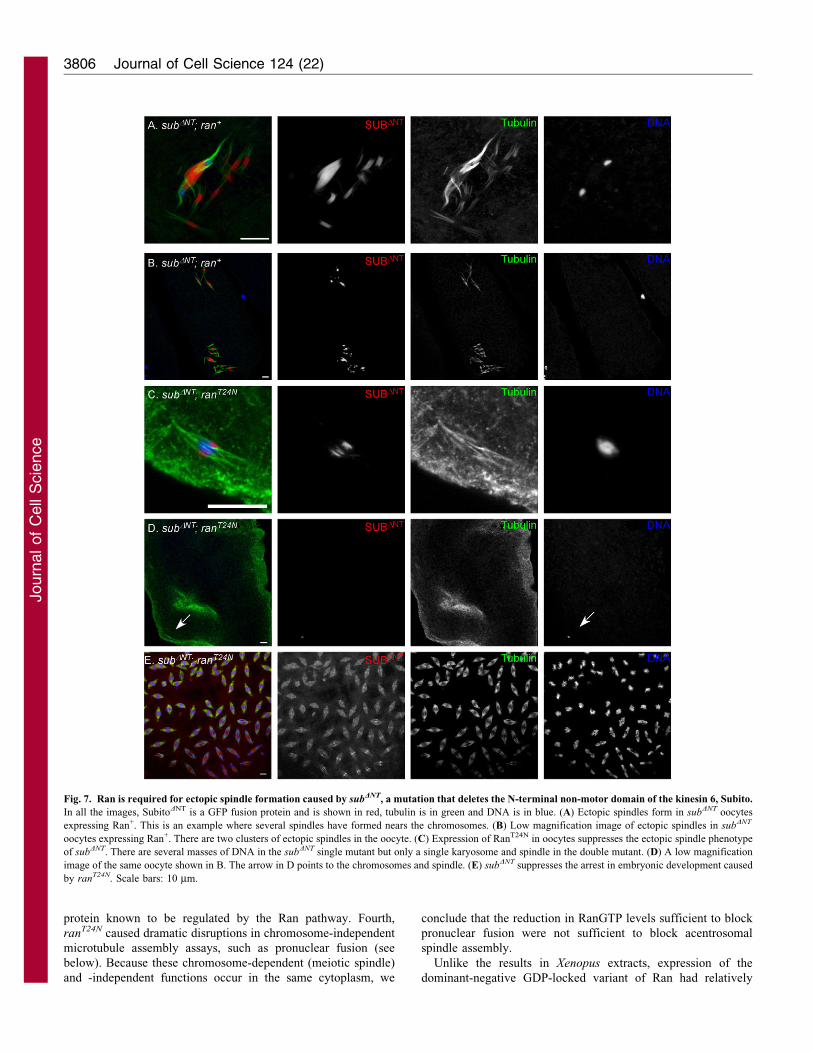
We tested whether RanGTP in the cytoplasm stimulates
microtubule assembly by constructing a double mutant with the
N-terminal deletion mutation using subDNT and ranT24N.
Typically, two to four clusters of ectopic spindles could be
observed in subDNT mutant oocytes, such as at the posterior tip
and the region near the karyosome (Fig. 7A,B). Any oocyte
containing more than one cluster of spindle formation was
considered to have the ectopic spindle phenotype. The frequency
of subDNT oocytes expressing ran+ with ectopic spindles was
similar to that with subDNT alone (96.8% and 97.9%, respectively;
Table 4). Strikingly, the dominant-negative mutation ranT24N
completely suppressed the ectopic spindle phenotype (Fig. 7C,D;
Table 4). The only spindle that formed in ranT24N; subDNT
oocytes was around the karyosome. These results suggest that
RanGTP is required for the interaction between SubitoDNT and
microtubules that occurs in the absence of the chromosomes. In
other words, RanGTP could be required for microtubule
assembly that does not depend on direct contacts with the
chromosomes. Another surprising finding was that the
suppression was reciprocal. The ranT24N; subDNT double mutant
had increased fertility relative to the two single mutants (Table 2)
and this correlated with an increased frequency of embryos
undergoing mitosis (Fig. 7E).
DiscussionThe Ran pathway has a variety of targets, which leads to effects
on kinetochores, centrosomes and microtubule-associated
proteins (Kalab and Heald, 2008). RanGTP is potentially an
important molecule for spindle assembly in acentrosomal oocytes
because it has been identified as a key factor for chromatin-
induced spindle formation in Xenopus extracts (Carazo-Salas
et al., 1999; Kalab et al., 1999; Karsenti and Vernos, 2001; Ohba
et al., 1999). Surprisingly, our results suggest that RanGTP might
be more important for microtubule assembly in other
circumstances, such as when centrosomes are present or when
microtubules assemble without direct contact with the
chromosomes.
Regulators of RanGTP, RCC1 and RanGAP, during meiosis
in Drosophila females
Diffusion of RanGTP from its source, the chromatin, into the
cytoplasm, where it is converted into RanGDP, can create a
gradient that regulates microtubule organization (Caudron et al.,
2005; Kalab et al., 2006). Drosophila oocytes contain two key
regulators of the Ran pathway in distinct locations. RCC1, as
expected, is located tightly around the karyosome in mature
oocytes. RanGAP localization is more complex than expected
because it is present in many clusters, possibly vesicles, within
the oocyte, suggesting that conversion of RanGTP to RanGDP
might be regulated and only occur in certain locations. This could
mean that a gradient of RanGTP (Clarke and Zhang, 2008; Kalab
and Heald, 2008) is not established in the oocyte. A candidate
protein responsible for generating the concentrations of RanGAP
is Ran binding protein 2 (RanBP2; also known as Nup358). This
protein is found within the nuclear envelope and binds to
RanGAP (Hutten et al., 2008). Following NEB, RanGAP could
be anchored to RanBP2-containing cytoplasmic vesicles.
Ran has an unusual localization pattern in oocytes;
concentrating around the outside of the spindle. By contrast,
Ran overlaps with the spindle in Drosophila mitotic cells (this
work) (Silverman-Gavrila and Wilde, 2006; Trieselmann and
Wilde, 2002). We have not determined whether these
concentrations of Ran are in the GDP or GTP state. However,
we can speculate on the basis of the localization patterns of wild-
type and mutant proteins. From this type of evidence,
Trieselmann and Wilde suggested that the bulk of Ran on the
embryonic spindle is in the GTP state. (Trieselmann and Wilde,
2002) Similarly, the bulk of the Ran localized around the outside
of the meiotic spindle might be in the GTP form. The pattern of
mutant RanQ69L staining suggests it enters RanGAP-containing
vesicles but does not leave because it is not hydrolyzed. Thus, the
wild-type Ran that localizes adjacent to the clusters of RanGAP
could be the GDP form of the protein that has left RanGAP-
containing vesicles.
RanGTP has a role in organizing the acentrosomal
spindle poles
RCC1 and RanGTP were found to be required for chromatin-
induced spindle assembly in Xenopus extracts (Carazo-Salas
et al., 1999; Kalab et al., 1999). In such extracts, expression of
RanT24N blocks spindle assembly (Ohba et al., 1999) and high
concentrations of RCC1 or expression of a GTP-locked form of
Ran leads to spindle formation in the absence of chromosomes
and centrosomes (Carazo-Salas et al., 1999).
Our analysis of RanGTP function in Drosophila oocytes is
based on these and numerous other studies in which expression of
the ranT24N mutation effectively reduces the concentration of
RanGTP. We believe that the ranT24N mutant had the desired
effect of reducing RanGTP production, for four reasons. First,
expression of RanT24N in somatic cells caused embryonic or early
larval lethality. Second, RanT24N localized tightly to the meiotic
chromosomes, consistent with the expectation that this form of
Ran remains bound to RCC1 because it has a low rate of GTP
exchange. The high affinity of RanT24N for RCC1 causes a
reduction in the production of RanGTP (Dasso et al., 1994).
Third, the spindle organization defects observed in ranT24N
oocytes were similar to defects seen in mars1 mutant oocytes, a
Table 4. Characterization of ectopic spindle phenotype in subDNT double mutants
Genotype Total oocytes Oocytes containing ectopic spindles Ectopic spindle (%)a
subDNT 93 91 98subDNT; ran+ 63 61 97ranT24N 16 0 0subDNT; ranT24N 28 0 0
Each genotype was expressed by crossing to P{GAL4::VP16-nos.UTR}MVD1 driver.aEctopic spindle (%) is equal to the number of oocytes with ectopic spindles divided by the total number of oocytes.
Meiotic spindle assembly 3805
Journ
alof
Cell
Scie
nce
protein known to be regulated by the Ran pathway. Fourth,
ranT24N caused dramatic disruptions in chromosome-independent
microtubule assembly assays, such as pronuclear fusion (see
below). Because these chromosome-dependent (meiotic spindle)
and -independent functions occur in the same cytoplasm, we
conclude that the reduction in RanGTP levels sufficient to block
pronuclear fusion were not sufficient to block acentrosomal
spindle assembly.
Unlike the results in Xenopus extracts, expression of the
dominant-negative GDP-locked variant of Ran had relatively
Fig. 7. Ran is required for ectopic spindle formation caused by subDNT, a mutation that deletes the N-terminal non-motor domain of the kinesin 6, Subito.
In all the images, SubitoDNT is a GFP fusion protein and is shown in red, tubulin is in green and DNA is in blue. (A) Ectopic spindles form in subDNT oocytes
expressing Ran+. This is an example where several spindles have formed nears the chromosomes. (B) Low magnification image of ectopic spindles in subDNT
oocytes expressing Ran+. There are two clusters of ectopic spindles in the oocyte. (C) Expression of RanT24N in oocytes suppresses the ectopic spindle phenotype
of subDNT. There are several masses of DNA in the subDNT single mutant but only a single karyosome and spindle in the double mutant. (D) A low magnification
image of the same oocyte shown in B. The arrow in D points to the chromosomes and spindle. (E) subDNT suppresses the arrest in embryonic development caused
by ranT24N. Scale bars: 10 mm.
Journal of Cell Science 124 (22)3806
Journ
alof
Cell
Scie
nce

mild effects on Drosophila oocyte spindle assembly andkaryosome organization. Meiosis I spindles were bipolar in
ranT24N oocytes. Indeed, reducing the RanGTP concentration inthe oocyte was not sufficient to severely affect either meioticdivision, because meiosis II spindles (data not shown) and femalemeiotic products could be seen in the embryos. The most
important defect was that the meiosis I spindle often had non-tapered poles, and was associated with the abnormal localizationof proteins necessary for pole formation such as Mars or TACC
(Cullen and Ohkura, 2001). These results are consistent withexperiments in embryos that found depletion of RanGTP causesdefects in spindle pole organization and chromosome
organization and congression (Silverman-Gavrila and Wilde,2006). Abnormal spindle morphology in oocytes could be thereason for the disorganized karyosome phenotype andnondisjunction of achiasmate chromosomes. Loss of RanGTP
could result in a failure to activate Aurora A, whichphosphorylates TACC (Barros et al., 2005; Kalab and Heald,2008). In embryos, TACC localization to the centrosomes
depends on phosphorylation by Aurora A (Barros et al., 2005).TACC initially binds all microtubules, but as the spindle matures,TACC is phosphorylated and localizes to the poles. Expression of
ranT24N in oocytes might cause a reduction in Aurora A activity,resulting in a failure to phosphorylate TACC and localize it to thepoles. Further studies are needed, however, because the role of
Aurora A in Drosophila female meiosis is not known. In addition,Mars might have a role in promoting the dephosphorylation ofTACC (Tan et al., 2008). Overall, RanGTP might have a specificrole in organizing spindle poles but might not be required for
chromosome-promoted spindle assembly in oocytes.
Expression of ranT24N did not block spindle assembly.Conversely, expression of the GTP-locked mutant, ranQ69L, did
not induce an uncoupling between spindle assembly and thechromosomes, as it does in Xenopus oocytes (Carazo-Salas et al.,1999). Thus, RanGTP might not be sufficient to initiate spindle
assembly in Drosophila oocytes. Surprisingly, ranQ69L oocytesshowed loss-of-function spindle phenotypes similar to ranT24N
mutant oocytes. Interestingly, manipulation of RanGTP levelswith T24N or Q69L mutations in mammals has similar
phenotypes. For example, the expression of either form of Ranin mouse oocytes resulted in similar meiosis II spindlephenotypes (Dumont et al., 2007). These results suggest that
the effects of manipulating RanGTP levels in an intact oocyte arenot easily predicted by experiments in Xenopus extracts. Otherfactors such as protein localization might play important roles in
regulating the Ran pathway. We also cannot rule out thepossibility that the Ran pathway functions differently in oocytemeiosis, such as if active Ran is not GTP dependent.
We suggest there could be two reasons for the similarity of the
ranQ69L and ranT24N phenotypes. First, expressing the GTP-locked ranQ69L mutation can inhibit the binding of RCC1 to thechromatin (Zhang et al., 2002), causing a reduction in RanGTP
near the chromatin. Alternatively, the phenotypes of the ranQ69L
mutant oocyte might be associated with defects in theorganization of membranes or vesicles. For example,
expression of the ranQ69L mutation caused Lamin, RanGAP andRanQ69L to colocalize in globular structures throughout theoocyte. Kramer and Hawley (Kramer and Hawley, 2003) have
proposed that the transmembrane protein Axs is a component of amembranous structure surrounding the meiotic spindle. With thecaveat that the link between membranous structures and spindle
assembly is not known, the ranQ69L mutation might cause defects inmembranous structures that have a role in spindle organization. Wehave found that Ran and Axs are closely associated, although at the
light microscope level it is difficult to determine if Ran is inside oroutside the Axs staining (J.C. and K.S.M., unpublished results).
Similar to the oocytes, the phenotype of the ranQ69L mutantzygotes might be associated with defects in membrane structure.
In ranQ69L mutants, a single cluster of DNA and microtubulescould be observed in the center of the zygote. A strikingly similarphenotype has been observed in dominant-negative Ketel
mutants; Ketel is the Drosophila homolog of importin-b(Timinszky et al., 2002; Tirian et al., 2000). In the Ketel
dominant mutants, meiosis I and II occur and the female andmale pronuclei come together, but they interact abnormally
because of defects in the nuclear envelopes. Subsequently, thefirst mitotic division fails and the chromosomes disintegratewithin a large aggregate of microtubules. Similar to the Ketel
mutant, ranQ69L could cause abnormal interactions amongnuclear envelope proteins in the zygote, causing a failure in thefirst mitotic division.
RanGTP is required for achiasmatechromosome segregation
There are two chromosome segregation mechanisms inDrosophila females. The first is the segregation of bivalentsconnected by chiasmata (Hawley, 1988), which is how most
chromosomes segregate. The second is of the chromosomes thatlack chiasmata. This includes the small fourth chromosome,which always lacks crossovers, and larger chromosomes, which
lack a crossover in approximately 5% of meioses. Homologouspairs can be forced into the achiasmate system with balancers thatsuppress crossing over. In all these cases, homologous
chromosomes segregate correctly even though they are notconnected by chiasmata. Expression of ranT24N had only mildeffects on chiasmate segregation, but had a severe effect on thesegregation of achiasmate X-chromosomes. These results suggest
that the spindle pole organization defects caused by low RanGTPlevels affect chromosome segregation.
RanGTP is required for chromosome independentmicrotubule assembly
Unlike assembly of the meiosis I spindle, expression of ranT24N
blocked two other types of microtubule assembly. First, ranT24N
mutants had a defect in the fusion of the female and male
pronuclei. This was the most probable cause of the fertilitydefect in ranT24N mutants. Several genes with roles inmicrotubule assembly are also required for pronuclear fusion,including subito (Giunta et al., 2002). This process depends on
the assembly of a microtubule array that is nucleated by thecentrosome donated by the sperm, and acts to draw the femalepronucleus towards the male pronucleus. Second, ranT24N
suppressed the formation of the ectopic spindles that form in aneomorphic subito mutant (subDNT) (Jang et al., 2007). Theformation of these spindles occurs after NEB, consistent with a
dependence on release of RanGTP from the nucleus. Both ofthese examples involve assembly of microtubules without directinteraction with the chromosomes and suggest that the assembly
and bundling of microtubules in the oocyte cytoplasm depend onRanGTP.
One characteristic of the ectopic spindles in subDNT mutants isthat they form in discrete clusters within the oocyte. Because
Meiotic spindle assembly 3807
Journ
alof
Cell
Scie
nce

RanGAP appears in clusters, it is possible that RanGTP is not in agradient or distributed evenly in the cytoplasm. Thus, aninteresting possibility is that the regions containing ectopicspindles are where the concentration of RanGTP-dependent
spindle assembly factors are at their highest.
These experiments revealed a surprising mutual suppression by
the subDNT and ranT24N mutations. Although both mutants havedecreased fertility, the double mutant is fertile. One interpretationis that RanGTP regulates Subito, and the subDNT mutation
bypasses the dependence on RanGTP. However, we found thatran mutations did not affect Subito localization or the formationof the central spindle. A more probable explanation is that there
are two independent spindle assembly pathways in the oocyte andthe loss of spindle assembly factors in ranT24N zygotes isbalanced by the enhanced spindle assembly activity present in thesubDNT mutant. Expression of ranT24N might suppress the sterility
phenotype of subDNT by abolishing ectopic spindles, whereassubDNT might suppress the reduced fertility phenotype of ranT24N
by overcoming the defects in microtubule assembly needed for
processes such as pro-nuclear fusion.
ConclusionsUsing dominant-negative mutations, we have found that RanGTP is
required for the fertility of Drosophila females. We found no evidencethat RanGTP is required, or sufficient, for the initiation of acentrosomalspindle assembly in Drosophila oocytes. We did detect a role inorganizing the spindle poles that could be explained by RanGTP
regulation of proteins such as Mars/Hurp, TACC and Aurora A. Thesedefects, however, would not be expected to have a severe effect onfertility. A similar conclusion was drawn from expressing a dominant-
negative form of Ran in mouse oocytes or when RCC1 was depletedfrom Xenopus oocytes (Dumont et al., 2007). By analyzing mutantssimilar to the ones we used here, only mild defects in meiosis I spindle
assembly were found, such as a delay establishing bipolarity. Thefailure to observe evidence supporting a role for RanGTP inacentrosomal spindle assembly might be explained by a predominant
chromosome-dependent pathway in oocytes involving thechromosome passenger complex (CPC). The CPC is required forchromosome-dependent spindle assembly in Xenopus egg extracts(Maresca et al., 2009; Sampath et al., 2004) and Drosophila oocytes
(Colombie et al., 2008) (S. Radford and K.S.M., unpublished results).
Our results suggest that, compared with chromosome-mediated
spindle assembly, RanGTP has a greater role in microtubuleorganization when centrosomes are present or when at a distancefrom the chromosomes. We suggest that NEB preceding the
assembly of the meiosis I spindle releases RanGTP into thecytoplasm, which results in a cytoplasm enriched for activespindle assembly factors. The restriction of RanGAP to vesicle-like structures could leave a considerable amount of RanGTP in
the cytoplasm. This activity has only a minor role in meioticspindle assembly, but is crucial for the early events ofembryogenesis. The microtubule array facilitating pronuclear
fusion assembles while the nuclear envelope is intact. Therefore,the oocyte might accumulate and store RanGTP during themeiotic divisions when there is no nuclear envelope in order to
support pronuclear fusion, when the nuclear envelope is intact.
Materials and MethodsGeneration and analysis of transgenic linesFull-length and substitution derivatives of ran were amplified by PCR. The cloneswere verified by sequencing and then the fragments were cloned into the pENTR4vector (Gateway). The fragment was then recombined using Clonase (Invitrogen)
into the pPHW vector that encodes three copies of the HA epitope at theN-terminus of the coding region in a pUASP backbone (Rorth, 1998). Amino acidsubstitutions were made by modifying the wild-type ran clone in pENTR4 usingthe Change-IT mutagenesis kit (USB) and the appropriate primers. For the ranT24N
transgene, an asparagine was substituted for a threonine at amino acid 24. For theranQ69L transgene, a leucine was substituted for a glutamine at amino acid 69.
For ubiquitous expression in somatic tissues, males carrying a ran transgene,P{UASP::ran}, were crossed to females carrying a GAL4 transgene with atubulin promoter (P{tubP-GAL4}) (Lee and Luo, 1999). A cross with the driverheterozygous to a balancer that provides a Tubby phenotype visible in larvae,P{tubP-GAL4}/T(2;3)B3, CyO: TM6B, Tb, results in two genotypes:P{UASP::ran}/P{tubP-GAL4} and P{UASP::ran}/T(2;3)B3, CyO: TM6B, Tb.The percentage survival was calculated as (Tb+ flies)/(total flies). For expressionin the germline and early embryo, males carrying a ran transgene were crossed tofemales carrying a GAL4 transgene with a nanos promoter, P{GAL4::VP16-nos.UTR}MVD1 (Van Doren et al., 1998). To measure fertility and chromosomesegregation during meiosis, females carrying a transgene and the nanos driverwere crossed to either y w/BSY or C(1;Y)v f B, c(4) ci eyR males. Thenondisjunction frequency was calculated as 2(BS R+B+ =)/[B+ R+BS =+2(BS
R+B+ =)].
Antibodies and immunofluorescence microscopy
Mature (stage 14) oocytes were collected from 50–200 yeast-fed females that wereaged 3–4 days by physical disruption in a common household blender (McKimet al., 2009; Theurkauf and Hawley, 1992). The oocytes were fixed in modifiedRobb’s medium and cacodylate–formaldehyde fixative for 8 minutes and then theirouter membranes were removed by rolling the oocytes between the frosted part ofa slide and a coverslip.
Embryos were collected by placing females and males in cages with grape juiceplates for 2 hours to enrich for those undergoing the syncytial divisions. Embryoswere removed from the grape juice plates with water and placed in 50% bleach for90 seconds to remove the chorion. They were then thoroughly washed with waterto remove all traces of bleach. The embryos were fixed using heptane andmethanol (Rothwell and Sullivan, 2000).
For squashed neuroblast preparations, the third instar larval brains weredissected in saline and the brains were fixed in 3.7% formaldehyde in 16 PBS for30 minutes. The brains were then placed in 45% acetic acid for 3 minutes beforetransferring to ,8 ml of 60% acetic acid on a siliconized coverslip where theywere firmly squashed between the coverslip and slide. The slides were brieflyfrozen in liquid nitrogen and the coverslips were flicked off with a razor blade. Theslides were placed in ethanol at 220 C̊ (chilled on dry ice) for 10 minutes, thentransferred to a slide chamber containing 0.1% Triton X-100 in PBS for 10minutes. Rubber cement was used to form wells on the slides and the preparationswere washed twice for 5 minutes each with PBS. The tissue was blocked with 1%BSA in PBS for 45 minutes.
Oocytes, embryos and neuroblasts were stained for DNA with Hoechst 33342 ata 1:1000 dilution (10 mg/ml solution) and for microtubules with mouse anti-a-tubulin monoclonal antibody DM1A (1:50), directly conjugated to FITC (Sigma)or rat anti-a-tubulin monoclonal antibody (1:75; Millipore). The primaryantibodies were rat anti-SUB antibody (used at 1:75) (Jang et al., 2005), ratanti-HA (Roche, clone 3F10: 1:25), rat anti-INCENP (1:500) (Wu et al., 2008),mouse anti-RCC1 (1:20) (Frasch, 1991), rabbit anti-RanGAP (1:800) (Kusanoet al., 2001), rabbit anti-Mars (Tan et al., 2008) and mouse anti-Lamin Dm0
(1:800) (Klapper et al., 1997). These primary antibodies were detected with eithera Cy3 or Cy5 secondary antibody preabsorbed against a range of mammalianserum proteins (Jackson Labs) and Drosophila embryos. TACC was detected usinga GFP fusion protein (Gergely et al., 2000). Images were collected on a Leica TCSSP2 confocal microscope with a 636, 1.3 NA lens. Images are maximumprojections of image stacks with the individual channels merged and then croppedin Adobe Photoshop.
Fluorescent in situ hybridization
Stage 14 oocytes were collected as described above and then processed for bothimmunofluorescence and fluorescent in situ hybridization (FISH) as describedpreviously (Dernburg, 2000; McKim et al., 2009). Oligonucleotidescorresponding to the satellite sequence AACAC for the second chromosomecentric heterochromatin or CCCGTACTCGGT (Dodeca) for the thirdchromosome centric heterochromatin were end-labeled with Cc3-dCTP or Cc5-dCTP (GE Healthcare) by Terminal Deoxynucleotidyl Transferase (Invitrogen).A probe to the 359 bp repeat on the X-chromosome was amplified by PCR andend labeled as described previously (Dernburg, 2000). Oocytes weresubsequently stained for microtubules and DNA as described above.
AcknowledgementsWe are grateful to Li Nguyen for technical assistance, DaimarkBennett for providing Mars antibodies and the mars1 mutant andJanet Jang for Fig. 2A. Some stocks used in this study were obtained
Journal of Cell Science 124 (22)3808
Journ
alof
Cell
Scie
nce

from the Bloomington Stock Center and some antibodies were fromDevelopmental Studies Hybridoma Bank at the University of Iowa,developed under the auspices of the National Institute of ChildHealth and Human Development.
FundingThis work was supported by a fellowship from the Busch foundation(to J.C.); and a grant from the National Institutes of Health [grantnumber GM 067142 to K.S.M.]. Deposited in PMC for release after12 months.
Supplementary material available online at
http://jcs.biologists.org/lookup/suppl/doi:10.1242/jcs.084855/-/DC1
ReferencesAskjaer, P., Galy, V., Hannak, E. and Mattaj, I. W. (2002). Ran GTPase cycle and
importins alpha and beta are essential for spindle formation and nuclear envelopeassembly in living Caenorhabditis elegans embryos. Mol. Biol. Cell 13, 4355-4370.
Baker, B. S. and Hall, J. C. (1976). Meiotic mutants: genetic control of meioticrecombination and chromosome segregation. In The Genetics and Biology of
Drosophila, vol. 1a (eds M. Ashburner and E. Novitski), pp. 351-434. New York:Academic Press.
Bamba, C., Bobinnec, Y., Fukuda, M. and Nishida, E. (2002). The GTPase Ranregulates chromosome positioning and nuclear envelope assembly in vivo. Curr. Biol.
12, 503-507.
Barros, T. P., Kinoshita, K., Hyman, A. A. and Raff, J. W. (2005). Aurora A activatesD-TACC-Msps complexes exclusively at centrosomes to stabilize centrosomalmicrotubules. J. Cell Biol. 170, 1039-1046.
Bischoff, F. R. and Ponstingl, H. (1991). Catalysis of guanine nucleotide exchange onRan by the mitotic regulator RCC1. Nature 354, 80-82.
Bischoff, F. R., Klebe, C., Kretschmer, J., Wittinghofer, A. and Ponstingl, H. (1994).RanGAP1 induces GTPase activity of nuclear Ras-related Ran. Proc. Natl. Acad. Sci.
USA 91, 2587-2591.
Carazo-Salas, R. E., Guarguaglini, G., Gruss, O. J., Segref, A., Karsenti, E. and
Mattaj, I. W. (1999). Generation of GTP-bound Ran by RCC1 is required forchromatin-induced mitotic spindle formation. Nature 400, 178-181.
Caudron, M., Bunt, G., Bastiaens, P. and Karsenti, E. (2005). Spatial coordination ofspindle assembly by chromosome-mediated signaling gradients. Science 309, 1373-1376.
Ciciarello, M., Mangiacasale, R. and Lavia, P. (2007). Spatial control of mitosis by theGTPase Ran. Cell Mol. Life Sci. 64, 1891-1914.
Clarke, P. R. and Zhang, C. (2008). Spatial and temporal coordination of mitosis byRan GTPase. Nat. Rev. Mol. Cell Biol. 9, 464-477.
Colombie, N., Cullen, C. F., Brittle, A. L., Jang, J. K., Earnshaw, W. C., Carmena,
M., McKim, K. and Ohkura, H. (2008). Dual roles of Incenp crucial to the assemblyof the acentrosomal metaphase spindle in female meiosis. Development 135, 3239-3246.
Cullen, C. F. and Ohkura, H. (2001). Msps protein is localized to acentrosomal polesto ensure bipolarity of Drosophila meiotic spindles. Nature Cell. Biol. 3, 637-642.
Dasso, M., Seki, T., Azuma, Y., Ohba, T. and Nishimoto, T. (1994). A mutant form ofthe Ran/TC4 protein disrupts nuclear function in Xenopus laevis egg extracts byinhibiting the RCC1 protein, a regulator of chromosome condensation. EMBO J. 13,5732-5744.
Demerec, M. (1950). Biology of Drosophila. 632p. New York: Hafner Publishing Co.
Dernburg, A. F. (2000). In situ hybridization to somatic chromosomes. In Drosophila
Protocols (ed. W. Sullivan, M. Ashburner and R. S. Hawley), pp. 25-55. Cold SpringHarbor, NY: Cold Spring Harbor Laboratory Press.
Dix, C. I. and Raff, J. W. (2007). Drosophila Spd-2 recruits PCM to the spermcentriole, but is dispensable for centriole duplication. Curr. Biol. 17, 1759-1764.
Dumont, J., Petri, S., Pellegrin, F., Terret, M. E., Bohnsack, M. T., Rassinier, P.,
Georget, V., Kalab, P., Gruss, O. J. and Verlhac, M. H. (2007). A centriole- andRanGTP-independent spindle assembly pathway in meiosis I of vertebrate oocytes. J.
Cell Biol. 176, 295-305.
Frasch, M. (1991). The maternally expressed Drosophila gene encoding the chromatin-binding protein BJ1 is a homolog of the vertebrate gene regulator of chromatincondensation, RCC1. EMBO J. 10, 1225-1236.
Gergely, F., Kidd, D., Jeffers, K., Wakefield, J. G. and Raff, J. W. (2000). D-TACC:a novel centrosomal protein required for normal spindle function in early Drosophila
embryo. EMBO J. 19, 241-252.
Giet, R., McLean, D., Descamps, S., Lee, M. J., Raff, J. W., Prigent, C. and Glover,
D. M. (2002). Drosophila Aurora A kinase is required to localize D-TACC tocentrosomes and to regulate astral microtubules. J. Cell Biol. 156, 437-451.
Giunta, K. L., Jang, J. K., Manheim, E. M., Subramanian, G. and McKim, K. S.
(2002). subito encodes a kinesin-like protein required for meiotic spindle poleformation in Drosophila melanogaster. Genetics 160, 1489-1501.
Hawley, R. S. (1988). Exchange and chromosomal segregation in eucaryotes. In Genetic
Recombination (eds R. Kucherlapati and G. Smith), pp. 497-527. Washington, D.C.:American Society of Microbiology.
Hawley, R. S. and Theurkauf, W. E. (1993). Requiem for distributive segregation:Achiasmate segregation in Drosophila females. Trends Genet. 9, 310-317.
Hutten, S., Flotho, A., Melchior, F. and Kehlenbach, R. H. (2008). The Nup358-RanGAP complex is required for efficient importin alpha/beta-dependent nuclearimport. Mol. Biol. Cell 19, 2300-2310.
Jang, J. K., Rahman, T. and McKim, K. S. (2005). The kinesin-like protein Subitocontributes to central spindle assembly and organization of the meiotic spindle inDrosophila oocytes. Mol. Biol. Cell 16, 4684-4694.
Jang, J. K., Rahman, T., Kober, V. S., Cesario, J. and McKim, K. S. (2007).Misregulation of the Kinesin-like protein Subito induces meiotic spindle formation inthe absence of chromosomes and centrosomes. Genetics 177, 267-280.
Kahana, J. A. and Cleveland, D. W. (1999). Beyond nuclear transport. Ran-GTP as adeterminant of spindle assembly. J. Cell Biol. 146, 1205-1210.
Kalab, P. and Heald, R. (2008). The RanGTP gradient – a GPS for the mitotic spindle.J. Cell Sci. 121, 1577-1586.
Kalab, P., Pu, R. T. and Dasso, M. (1999). The ran GTPase regulates mitotic spindleassembly. Curr. Biol. 9, 481-484.
Kalab, P., Pralle, A., Isacoff, E. Y., Heald, R. and Weis, K. (2006). Analysis of aRanGTP-regulated gradient in mitotic somatic cells. Nature 440, 697-701.
Karsenti, E. and Vernos, I. (2001). The mitotic spindle: a self-made machine. Science
294, 543-547.
Klapper, M., Exner, K., Kempf, A., Gehrig, C., Stuurman, N., Fisher, P. A. and
Krohne, G. (1997). Assembly of A- and B-type lamins studied in vivo with thebaculovirus system. J. Cell Sci. 110, 2519-2532.
Kramer, J. and Hawley, R. S. (2003). The spindle-associated transmembrane proteinAxs identifies a membranous structure ensheathing the meiotic spindle. Nat. Cell
Biol. 5, 261-263.
Kusano, A., Staber, C. and Ganetzky, B. (2001). Nuclear mislocalization ofenzymatically active RanGAP causes segregation distortion in Drosophila. Dev.
Cell 1, 351-361.
Lee, T. and Luo, L. (1999). Mosaic analysis with a repressible cell marker for studies ofgene function in neuronal morphogenesis. Neuron 22, 451-461.
Maresca, T. J., Groen, A. C., Gatlin, J. C., Ohi, R., Mitchison, T. J. and Salmon,
E. D. (2009). Spindle assembly in the absence of a RanGTP gradient requireslocalized CPC activity. Curr. Biol. 19, 1210-1215.
Matthies, H. J., McDonald, H. B., Goldstein, L. S. and Theurkauf, W. E. (1996).Anastral meiotic spindle morphogenesis: role of the non-claret disjunctional kinesin-like protein. J. Cell Biol. 134, 455-464.
McKim, K. S., Joyce, E. F. and Jang, J. K. (2009). Cytological analysis of meiosis infixed Drosophila ovaries. Methods Mol. Biol. 558, 197-216.
Moore, M. S. and Blobel, G. (1993). The GTP-binding protein Ran/TC4 is required forprotein import into the nucleus. Nature 365, 661-663.
Ohba, T., Nakamura, M., Nishitani, H. and Nishimoto, T. (1999). Self-organizationof microtubule asters induced in Xenopus egg extracts by GTP-bound Ran. Science
284, 1356-1358.
Peter, A., Schottler, P., Werner, M., Beinert, N., Dowe, G., Burkert, P., Mourkioti,
F., Dentzer, L., He, Y., Deak, P. et al. (2002). Mapping and identification ofessential gene functions on the X chromosome of Drosophila. EMBO. Rep. 3, 34-38.
Rorth, P. (1998). Gal4 in the Drosophila female germline. Mech. Dev. 78, 113-118.
Roth, S. and Lynch, J. A. (2009). Symmetry breaking during Drosophila oogenesis.Cold Spring Harb. Perspect Biol. 1, a001891.
Rothwell, W. F. and Sullivan, W. (2000). Fluorescent analysis of Drosophila embryos.In Drosophila Protocols (eds. W. Sullivan, M. Ashburner and R. S. Hawley), pp. 141-157. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Sampath, S. C., Ohi, R., Leismann, O., Salic, A., Pozniakovski, A. and Funabiki, H.
(2004). The chromosomal passenger complex is required for chromatin-inducedmicrotubule stabilization and spindle assembly. Cell 118, 187-202.
Shi, W. Y. and Skeath, J. B. (2004). The Drosophila RCC1 homolog, Bj1,regulates nucleocytoplasmic transport and neural differentiation during Drosophiladevelopment. Dev. Biol. 270, 106-121.
Silverman-Gavrila, R. V. and Wilde, A. (2006). Ran is required before metaphase forspindle assembly and chromosome alignment and after metaphase for chromosomesegregation and spindle midbody organization. Mol. Biol. Cell 17, 2069-2080.
Tan, S., Lyulcheva, E., Dean, J. and Bennett, D. (2008). Mars promotes dTACCdephosphorylation on mitotic spindles to ensure spindle stability. J. Cell Biol. 182,27-33.
Theurkauf, W. E. and Hawley, R. S. (1992). Meiotic spindle assembly in Drosophila
females: behavior of nonexchange chromosomes and the effects of mutations in thenod kinesin-like protein. J. Cell Biol. 116, 1167-1180.
Theurkauf, W. E., Alberts, B. M., Jan, Y. N. and Jongens, T. A. (1993). A central rolefor microtubules in the differentiation of Drosophila oocytes. Development 118,1169-1180.
Timinszky, G., Tirian, L., Nagy, F. T., Toth, G., Perczel, A., Kiss-Laszlo, Z., Boros,
I., Clarke, P. R. and Szabad, J. (2002). The importin-beta P446L dominant-negativemutant protein loses RanGTP binding ability and blocks the formation of intactnuclear envelope. J. Cell Sci. 115, 1675-1687.
Tirian, L., Puro, J., Erdelyi, M., Boros, I., Papp, B., Lippai, M. and Szabad, J.
(2000). The Ketel(D) dominant-negative mutations identify maternal function of theDrosophila importin-beta gene required for cleavage nuclei formation. Genetics 156,1901-1912.
Trieselmann, N. and Wilde, A. (2002). Ran localizes around the microtubule spindle invivo during mitosis in Drosophila embryos. Curr. Biol. 12, 1124-1129.
Meiotic spindle assembly 3809
Journ
alof
Cell
Scie
nce

Van Doren, M., Williamson, A. L. and Lehmann, R. (1998). Regulation of zygoticgene expression in Drosophila primordial germ cells. Curr. Biol. 8, 243-246.
Wilde, A. (2006). 0HURP on 0 we’re off to the kinetochore! J. Cell Biol. 173, 829-831.Wu, C., Singaram, V. and McKim, K. S. (2008). mei-38 is required for chromosome
segregation during meiosis in Drosophila females. Genetics 180, 61-72.Yang, C. P. and Fan, S. S. (2008). Drosophila mars is required for organizing
kinetochore microtubules during mitosis. Exp. Cell Res. 314, 3209-3220.
Zhang, C., Goldberg, M. W., Moore, W. J., Allen, T. D. and Clarke, P. R. (2002).Concentration of Ran on chromatin induces decondensation, nuclear envelopeformation and nuclear pore complex assembly. Eur. J. Cell Biol. 81, 623-633.
Zhang, G., Breuer, M., Forster, A., Egger-Adam, D. and Wodarz, A. (2009). Mars, aDrosophila protein related to vertebrate HURP, is required for the attachment ofcentrosomes to the mitotic spindle during syncytial nuclear divisions. J. Cell Sci. 122,535-545.
Journal of Cell Science 124 (22)3810
Journ
alof
Cell
Scie
nce