protective role of arginase in a mouse model of · pdf fileprotective role of arginase in a...
TRANSCRIPT

of May 22, 2018.This information is current as
of ColitisProtective Role of Arginase in a Mouse Model
Amy Hacker, Robert A. Casero, Jr and Keith T. WilsonChaturvedi, Cinthia B. Drachenberg, Jean-Luc Boucher,
RupeshD. Mersey, Darren R. Blumberg, Raymond K. Cross, Alain P. Gobert, Yulan Cheng, Mahmood Akhtar, Benjamin
http://www.jimmunol.org/content/173/3/2109doi: 10.4049/jimmunol.173.3.2109
2004; 173:2109-2117; ;J Immunol
Referenceshttp://www.jimmunol.org/content/173/3/2109.full#ref-list-1
, 24 of which you can access for free at: cites 47 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2004 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 22, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 22, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Protective Role of Arginase in a Mouse Model of Colitis1
Alain P. Gobert,2*§ Yulan Cheng,*§ Mahmood Akhtar,* § Benjamin D. Mersey,*§
Darren R. Blumberg,* Raymond K. Cross,* Rupesh Chaturvedi,*§ Cinthia B. Drachenberg,†
Jean-Luc Boucher,¶ Amy Hacker,� Robert A. Casero, Jr.,� and Keith T. Wilson3*‡§
Arginase is the endogenous inhibitor of inducible NO synthase (iNOS), because both enzymes use the same substrate,L-arginase(Arg). Importantly, arginase synthesizes ornithine, which is metabolized by the enzyme ornithine decarboxylase (ODC) to producepolyamines. We investigated the role of these enzymes in theCitrobacter rodentium model of colitis. Arginase I, iNOS, and ODCwere induced in the colon during the infection, while arginase II was not up-regulated.L-Arg supplementation of wild-type miceor iNOS deletion significantly improved colitis, andL-Arg treatment of iNOS�/� mice led to an additive improvement. There wasa significant induction of IFN-�, IL-1, and TNF-� mRNA expression in colitis tissues that was markedly attenuated withL-Argtreatment or iNOS deletion. Treatment with the arginase inhibitor S-(2-boronoethyl)-L-cysteine worsened colitis in both wild-typeand iNOS�/� mice. Polyamine levels were increased in colitis tissues, and were further increased byL-Arg. In addition, in vivoinhibition of ODC with �-difluoromethylornithine also exacerbated the colitis. Taken together, these data indicate that arginaseis protective in C. rodentium colitis by enhancing the generation of polyamines in addition to competitive inhibition of iNOS.Modulation of the balance of iNOS and arginase, and of the arginase-ODC metabolic pathway may represent a new strategy forregulating intestinal inflammation. The Journal of Immunology, 2004, 173: 2109–2117.
D espite extensive study, there is uncertainty about the reg-ulation of inflammatory responses in human inflamma-tory bowel disease and in experimental colitis models. In
such models, there is strong evidence for a protective role of cy-clooxygenase-2 (1), TGF-�, and IL-10 (2), and a proinflammatoryeffect of IL-12 (3). However, in the case of NO, produced by theenzyme-inducible NO synthase (iNOS),4 there have been incon-sistent results concerning its role in colitis. For example, iNOSinhibition has been reported to either reduce (4) or increase (5) theseverity of trinitrobenzene sulfonic acid (TNBS) colitis, whileiNOS deletion has been shown to exacerbate acetic acid (6) orTNBS (7) colitis, but to improve dextran sulfate sodium (DSS)colitis (8, 9).
Evidence is accumulating that the activity of the enzyme argi-nase, which competes with iNOS for the common substrate, L-arginine (L-Arg), has important biologic effects on host innate im-
mune response (10–12) and epithelial cell functions (13, 14). Thecytoplasmic type I arginase (15) and the mitochondrial type IIarginase (16) catalyze the same conversion of L-Arg into ornithine,the latter being converted to polyamines by ornithine decarboxyl-ase (ODC). Thus, arginase is an inhibitor of NO synthesis and canalso modulate polyamine synthesis (17). Through these functions,arginase may have important biological effects in host mucosalimmune response, which have been demonstrated in vitro (11, 18).However, the role of arginase has not been studied in the gastro-intestinal mucosa in vivo. We hypothesized that arginase is anunrecognized regulator of mucosal inflammation and host-patho-gen interactions in the gut.
Citrobacter rodentium is the rodent equivalent of enteropatho-genic Escherichia coli (EPEC), which causes diarrhea in humans.C. rodentium colonizes the surface of colonic epithelial cells, re-sulting in signal transduction events, cytoskeletal rearrangements,formation of attaching and effacing lesions, epithelial hyperplasia,and a strong mucosal Th1 response (19, 20). Murine infectionprovides an ideal model, with reproducible histologic changes sim-ilar to human inflammatory bowel disease. The aim of this studywas to investigate the roles of iNOS, arginase, and ODC in C.rodentium colitis. There was marked up-regulation of iNOS, argi-nase I, and ODC in the colon of infected mice. Both wild-type(WT) and iNOS�/� mice supplemented with L-Arg had improve-ment in colitis, while disease was worsened when mice weretreated with S-(2-boronoethyl)-L-cysteine (BEC), a specific argi-nase inhibitor (21), or �-difluoromethylornithine (DFMO), anODC inhibitor. These findings suggest a protective role for argi-nase in the colon through the formation of polyamines.
Materials and MethodsBacteria
A WT strain of C. rodentium (DBS100), provided by D. Schauer (Massa-chusetts Institute of Technology, Cambridge, MA), was used. Before in-fection, C. rodentium were grown in Luria broth overnight, washed, andresuspended in PBS, and bacteria concentration was determined by OD andconfirmed by serial dilution and culture (18).
*Department of Medicine, Division of Gastroenterology, †Department of Pathology,and ‡Greenebaum Cancer Center, School of Medicine, University of Maryland, and§Veterans Affairs Maryland Health Care System, Baltimore, MD 21201; ¶Laboratoirede Chimie et Biochimie Pharmacologiques, Unite Mixte de Recherche 8601, CentreNational de la Recherche Scientifique, Universite Rene Descartes, Paris, France; and�Department of Oncology, Sidney Kimmel Comprehensive Cancer Center, School ofMedicine, Johns Hopkins University, Baltimore, MD 21231
Received for publication February 11, 2004. Accepted for publication May 21, 2004.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by grants from the National Institutes of Health (DK02469, DK56938, DK53620, and DK63626 to K.T.W., and CA51085 and CA98454to R.A.C.), the Office of Medical Research, Department of Veterans of Affairs (toK.T.W.), and the Crohn’s and Colitis Foundation of America (to K.T.W.).2 Current address: Unite de Microbiologie, Institut National de la RechercheAgronomique de Clermont-Ferrand-Theix, 63122 Saint-Genes-Champanelle, France.3 Address correspondence and reprint requests to Dr. Keith T. Wilson, Division of Gas-troenterology, School of Medicine, University of Maryland, 22 South Greene Street,Room N3W62, Baltimore, MD 21201. E-mail address: [email protected] Abbreviations used in this paper: iNOS, inducible NO synthase; BEC, S-(2-boro-noethyl)-L-cysteine; DFMO, �-difluoromethylornithine; DSS, dextran sulfate sodium;L-Arg, L-arginase; ODC, ornithine decarboxylase; TNBS, trinitrobenzene sulfonicacid; WT, wild type; EPEC, enteropathogenic Escherichia coli.
The Journal of Immunology
Copyright © 2004 by The American Association of Immunologists, Inc. 0022-1767/04/$02.00
by guest on May 22, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Mice and experimental design
All studies were approved by the University of Maryland (Baltimore, MD)Institutional Animal Care and Use Committee. WT and iNOS�/� C57BL/6mice were obtained from The Jackson Laboratory (Bar Harbor, ME), andbred in our facility. Eight-week-old mice were gavaged with C. rodentiumin 100 �l of PBS or PBS vehicle alone. Initial studies determined that aninoculum of 5 � 108 bacteria/mouse was required for consistent inductionof colitis. In some studies, mice were treated with 1% L-Arg (Sigma-Aldrich, St. Louis, MO), starting day 1 postinfection; 0.1% BEC (providedby J.-L.B.), starting day 3; or 2.5% DFMO (Ilex Oncology, San Antonio,TX), starting day 3, all at pH 7 in the drinking water. Daily weights wereobtained for each mouse. Water consumption was measured and found tobe similar between all experimental groups. Mice were sacrificed after 14days or when moribund. Blood samples were obtained by intracardiacpuncture, and colons were collected, weighed, and divided. Tissues snapfrozen in liquid nitrogen were used for RNA studies, Western blotting,enzyme activity assays, and polyamine measurements; tissues fixed in 10%buffered formalin were used for histology and immunohistochemistry;fresh tissues were homogenized in PBS; and colonization levels were de-termined by serial dilution and culture.
Histology
Sections (6 �m) were cut from paraffin sections and stained with H&E.Tissues were examined in a blinded manner by a pathologist (C.B.D.).Acute (neutrophilic) and chronic (lymphocytic) inflammation, and epithe-lial regenerative changes were each scored on a 0–4 scale, and the sum wasused as an index of histologic injury.
Immunohistochemistry
Immunohistochemistry was performed, as described (22, 23). PolyclonalAb to bovine liver arginase I (1/200; Research Diagnostics, Flanders, NJ)or to murine iNOS (1/400; BD Transduction Laboratories, Lexington, KY)were used for 1.5 h at room temperature. Sections were washed and incu-bated with goat biotinylated anti-rabbit secondary Ab (1/200; Vector Lab-oratories, Burlingame, CA) for 15 min. The Vectastain Elite ABC kit (Vec-tor Laboratories) was used as a peroxidase system. Color was developedusing 3,3�-diaminobenzidine, and hematoxylin was used as a counterstain.
Western blotting
Frozen colon samples were homogenized in lysis buffer containing pro-tease inhibitors (11). Protein concentrations of 14,000 � g supernatantswere measured (DC Protein Assay kit; Bio-Rad, Hercules, CA), and 100�g/lane was separated by SDS-PAGE and transferred onto Immobilon-Pmembranes (Millipore, Bedford, MA) by electroblotting. Membranes wereblocked overnight at room temperature with PBS containing 0.1% Tweenand 5% nonfat dry milk. Polyclonal Ab to arginase I (1/1000) or arginaseII (1/1000; provided by R. Iyer and S. Cederbaum, University of Califor-nia, Los Angeles, CA (11)) were used for 1 h at room temperature. Blotswere washed and incubated with donkey anti-rabbit Ab conjugated to HRP(1/2000; Amersham Biosciences, Piscataway, NJ). Chemiluminescencewas detected with SuperSignal West Pico chemiluminescent substrate(Pierce, Rockford, IL) and exposure to Hyperfilm ECL (AmershamBiosciences).
Determination of arginase activity
Protein supernatants were incubated with 0.1 �M L-[guanido-14C]Arg(NEN, Boston, MA) for 24 h at 37°C, and arginase activity assay wasperformed, as reported (10).
Determination of ODC activity and polyamine levels
Frozen tissues were homogenized in a solution of 25 mM Tris-HCl (pH7.4), 0.1 mM EDTA, and 2.5 mM DTT. Supernatants of 14,000 � g cen-trifugations were incubated at 37°C with 10 nmol of L-[1-14C]ornithine(NEN) for 24 h, and ODC activity was assayed, as reported (11). BacterialODC activity was measured after sonicating C. rodentium at log phase ofgrowth. Polyamine concentrations were measured in tissues by HPLC, asdescribed (24), and standardized to protein concentration. Data shown arethe total concentration of the biogenic polyamines putrescine, spermidine,and spermine.
mRNA analysis
Colon samples were homogenized in 1 ml of TRIzol reagent (InvitrogenLife Technologies, Grand Island, NY), and total RNA was isolated. Re-verse transcription, PCR cycle conditions, and primer sequences for iNOS,
arginase I, arginase II, ODC, and �-actin were as described (11). PCRproducts were run on 2% agarose gels with 0.4 �g/ml ethidium bromide.Bands were visualized under UV light and photographed with a digital geldocumentation system (EDAS 290; Kodak Digital Science, Rochester,NY). Real-time PCR was performed with a Bio-Rad iCycler iQ detectionsystem for iNOS, arginase I, arginase II, and ODC, and with an MJ Re-search DNA Engine Opticon 2 (Waltham, MA) for IFN-�, TNF-�, andIL-1. Fluorescein dye and SYBR green (Molecular Probes, Eugene, OR)were included in the PCR mix. For ODC, 3 pmol, and for iNOS, arginaseI, arginase II, IFN-�, TNF-�, IL-1, and �-actin, 7 pmol each of sense andantisense primers was used in each reaction. Sense and antisense primersequences for the cytokines were: IFN-�, 5�-GCCACGGCACAGTCATTGAA-3� and 5�-CGCCTTGCTGTTGCTGAAGA-3�; TNF-�, 5�-CTGTGAAGGGAATGGGTGTT-3� and 5�-GGTCACTGTCCCAGCATCTT3�;and IL-1, 5�-GCTGAAGGAGTTGCCAGAAA-3� and 5�-GTGCAAGTGACTCAGGGTGA-3�. One PCR cycle consisted of the following: 94°Cfor 30 s, 60°C for 30 s, and 72°C for 30 s. After standardization to thehousekeeping gene �-actin, relative expression of each gene comparedwith that in control animals was calculated according to the manufacturer’sinstructions.
Serum analysis
Serum was deproteinized with an equal volume of 6% 5-sulfosalicylic acid.Concentrations of L-Arg and total reactive nitrogen metabolites were de-termined in the serum, by HPLC analysis and chemiluminescence with anNO analyzer (NOA 280; Sievers, Boulder, CO), respectively, as previouslydescribed (18).
Statistical analysis
Data are expressed as the mean � SEM. When comparisons between mul-tiple groups were made, the Student-Newman-Keuls test was used, and forcomparisons between two groups, Student’s t test was used. For survivaldata, �2 or Fisher’s exact test was used as appropriate at each time point,and Cox hazard regression analysis was used to determine the effect ofL-Arg throughout. SAS (SAS Institute, Cary, NC) was used for the Coxregression, and Statview v. 5.01 for the Macintosh (SAS Institute) was usedfor all other analyses.
ResultsiNOS and arginase I are up-regulated in the colon of mice withC. rodentium colitis
As shown in Fig. 1A, expression of both arginase I and arginase IIwas very low in control tissues. Arginase I was increased in thecolon of C. rodentium-infected WT mice, while arginase II mRNAwas not induced. By real-time PCR, arginase I mRNA levels ofinfected mice were increased by 9.7 � 2.5-fold compared withuninfected mice (n � 6 for each; p � 0.01), while there was nosignificant change in arginase II mRNA levels from the low con-trol levels (data not shown). In addition, iNOS mRNA levels werealso found by RT-PCR (Fig. 1A) and by real-time PCR (data not
FIGURE 1. Colonic arginase and iNOS induction after C. rodentiuminfection for 14 days. A, arginase I, arginase II, and iNOS mRNA levelsassessed by RT-PCR. B, Western blotting for arginase I and II. In A and B,each lane represents tissue from a different mouse.
2110 ROLE OF ARGINASE IN COLITIS
by guest on May 22, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
shown) to be markedly increased in the colon of mice infected withC. rodentium, and absent in the colon of uninfected mice. Asshown in Fig. 1B, arginase I protein was present in the tissues ofinfected mice and absent from control, while arginase II levelswere very low in both control and infected mice.
Immunohistochemical detection of arginase I and iNOS
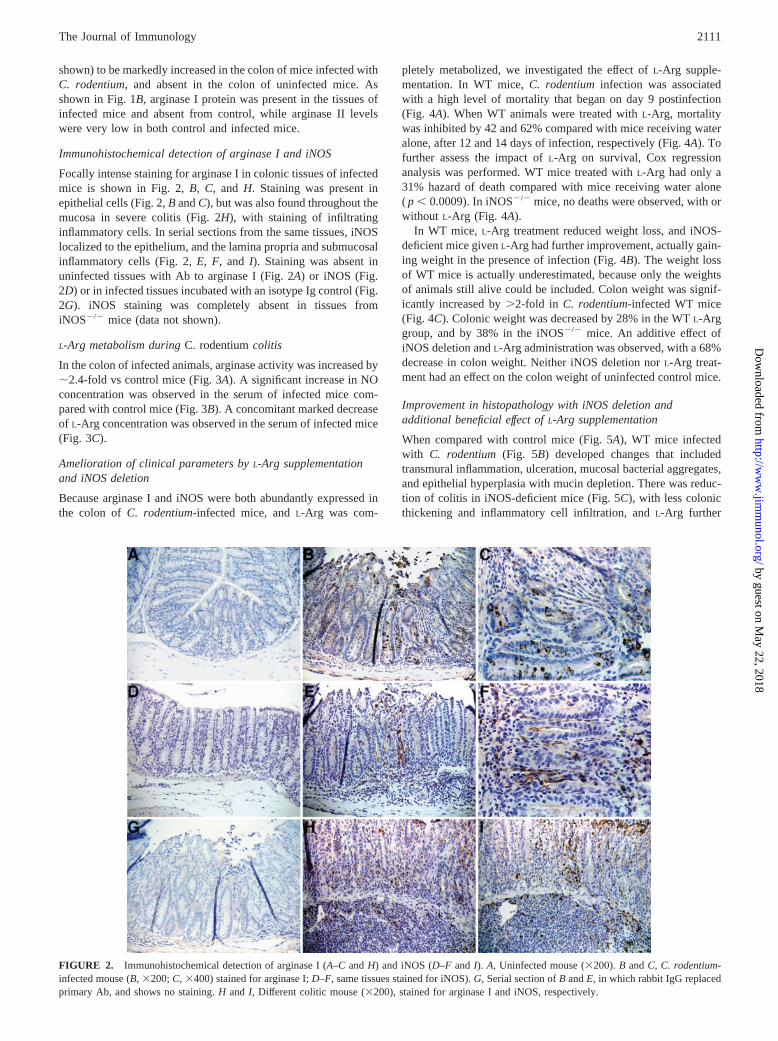
Focally intense staining for arginase I in colonic tissues of infectedmice is shown in Fig. 2, B, C, and H. Staining was present inepithelial cells (Fig. 2, B and C), but was also found throughout themucosa in severe colitis (Fig. 2H), with staining of infiltratinginflammatory cells. In serial sections from the same tissues, iNOSlocalized to the epithelium, and the lamina propria and submucosalinflammatory cells (Fig. 2, E, F, and I). Staining was absent inuninfected tissues with Ab to arginase I (Fig. 2A) or iNOS (Fig.2D) or in infected tissues incubated with an isotype Ig control (Fig.2G). iNOS staining was completely absent in tissues fromiNOS�/� mice (data not shown).
L-Arg metabolism during C. rodentium colitis
In the colon of infected animals, arginase activity was increased by�2.4-fold vs control mice (Fig. 3A). A significant increase in NOconcentration was observed in the serum of infected mice com-pared with control mice (Fig. 3B). A concomitant marked decreaseof L-Arg concentration was observed in the serum of infected mice(Fig. 3C).
Amelioration of clinical parameters by L-Arg supplementationand iNOS deletion
Because arginase I and iNOS were both abundantly expressed inthe colon of C. rodentium-infected mice, and L-Arg was com-
pletely metabolized, we investigated the effect of L-Arg supple-mentation. In WT mice, C. rodentium infection was associatedwith a high level of mortality that began on day 9 postinfection(Fig. 4A). When WT animals were treated with L-Arg, mortalitywas inhibited by 42 and 62% compared with mice receiving wateralone, after 12 and 14 days of infection, respectively (Fig. 4A). Tofurther assess the impact of L-Arg on survival, Cox regressionanalysis was performed. WT mice treated with L-Arg had only a31% hazard of death compared with mice receiving water alone( p � 0.0009). In iNOS�/� mice, no deaths were observed, with orwithout L-Arg (Fig. 4A).
In WT mice, L-Arg treatment reduced weight loss, and iNOS-deficient mice given L-Arg had further improvement, actually gain-ing weight in the presence of infection (Fig. 4B). The weight lossof WT mice is actually underestimated, because only the weightsof animals still alive could be included. Colon weight was signif-icantly increased by �2-fold in C. rodentium-infected WT mice(Fig. 4C). Colonic weight was decreased by 28% in the WT L-Arggroup, and by 38% in the iNOS�/� mice. An additive effect ofiNOS deletion and L-Arg administration was observed, with a 68%decrease in colon weight. Neither iNOS deletion nor L-Arg treat-ment had an effect on the colon weight of uninfected control mice.
Improvement in histopathology with iNOS deletion andadditional beneficial effect of L-Arg supplementation
When compared with control mice (Fig. 5A), WT mice infectedwith C. rodentium (Fig. 5B) developed changes that includedtransmural inflammation, ulceration, mucosal bacterial aggregates,and epithelial hyperplasia with mucin depletion. There was reduc-tion of colitis in iNOS-deficient mice (Fig. 5C), with less colonicthickening and inflammatory cell infiltration, and L-Arg further
FIGURE 2. Immunohistochemical detection of arginase I (A–C and H) and iNOS (D–F and I). A, Uninfected mouse (�200). B and C, C. rodentium-infected mouse (B, �200; C, �400) stained for arginase I; D–F, same tissues stained for iNOS). G, Serial section of B and E, in which rabbit IgG replacedprimary Ab, and shows no staining. H and I, Different colitic mouse (�200), stained for arginase I and iNOS, respectively.
2111The Journal of Immunology
by guest on May 22, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

ameliorated the inflammation, epithelial reactivity, and mucin de-pletion (Fig. 5D). Because individual scores for acute inflamma-tion, chronic inflammation, and epithelial reactivity had similarpatterns, a composite score is shown in Fig. 5E. iNOS deletion andiNOS deletion L-Arg inhibited the mean histologic injury scoreby 48 and 72%, respectively, compared with WT mice. L-Argtreatment of iNOS�/� mice resulted in a 46% improvement com-pared with water-treated iNOS�/� mice. It should be noted that themean histologic injury score in the WT mice is likely to be un-derestimated, due to the early mortality that precluded obtaininghistologic samples in some of the mice that would be expected tohave had high injury scores. In uninfected mice, no differenceswere observed between the colons of WT and iNOS�/� mice, withor without L-Arg treatment (data not shown).
C. rodentium colonization levels
Consistent with the decrease in histopathology, there was a sig-nificant reduction in C. rodentium colonization levels in the
iNOS�/� mice (WT mice, 8.3 � 3.0 � 109 CFU/g colon, n � 12;iNOS�/� mice, 5.6 � 3.3 � 106 CFU/g colon, n � 18; p � 0.01vs WT). However, L-Arg supplementation did not alter coloniza-tion levels in WT mice (WT L-Arg, 3.2 � 1.7 � 1010 CFU/g colon,n � 12), and actually increased levels in iNOS�/� mice comparedwith mutant mice receiving water alone (iNOS�/�
L-Arg, 2.8 �1.8 � 108 CFU/g colon, n � 13; p � 0.05 vs iNOS�/�), indicatingthat the clinical and histologic improvement with L-Arg is not sim-ply due to inhibition of bacterial growth by this amino acid.
FIGURE 3. Colonic arginase activity (A), serum NO concentration (B),and serum L-Arg concentration (C) in control (Ctrl) or C. rodentium-in-fected (C. rod) mice. n � 3 for Ctrl and n � 6 for C. rodentium. �, p �0.05; ���, p � 0.001.
FIGURE 4. Changes in survival (A), body weight (B), and colon weight(C) in WT and iNOS�/� mice infected with C. rodentium. Mice were givenL-Arg or water alone. Note that L-Arg treatment fully restored serum L-Arglevels in infected mice. n � 31 for WT infected with C. rodentium (f), n �32 for L-Arg-treated WT-infected mice (�), n � 23 for infected iNOS�/�
mice (Œ), and n � 20 for infected iNOS�/� mice treated with L-Arg (E).A–C, �, p � 0.05; ���, p � 0.001 vs day 0; §, p � 0.05; §§, p � 0.01; §§§,p � 0.001 vs WT water; #, p � 0.05 vs iNOS�/� without L-Arg.
2112 ROLE OF ARGINASE IN COLITIS
by guest on May 22, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

FIGURE 5. Histologic findings in H&E-stainedcolon of mice infected with C. rodentium. A, Un-infected WT, normal tissue. B, Infected WT withsevere colitis. C, Improvement in histologic dam-age in iNOS�/� mice. D, Further improvement inL-Arg-treated iNOS�/� mice. E, Histologic scoresin C. rodentium colitis. n � 19 for WT, n � 17 forWT L-Arg, n � 12 for iNOS�/�, and n � 11 foriNOS�/�
L-Arg. ��, p � 0.01 vs WT; §, p � 0.05vs iNOS�/�. All mice were inoculated with thesame amount of C. rodentium (5 � 108 CFU/mouse) and sacrificed on days 12–14; mice thatdied before day 12 were not included. Colonizationlevels for each experimental group are summarizedin Results.
2113The Journal of Immunology
by guest on May 22, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Induction of proinflammatory cytokines in colitis tissues isattenuated by L-Arg treatment or iNOS deletion
Because C. rodentium colitis has been strongly associated withactivation of the Th1 cytokine IFN-�, and the associated proin-flammatory cytokines TNF-� and IL-1 (19, 20), we sought to de-termine the relationship between the clinical and histologic effectsthat we observed with these immunologic parameters. Real-timePCR analysis demonstrated a marked increase in IFN-� (Fig. 6A),TNF-� (Fig. 6B), and IL-1 (Fig. 6C) in C. rodentium colitis tissuescompared with normal tissues, and a significant attenuation ofthese increases with either iNOS deletion or L-Arg treatment.
ODC activity is induced in the colon of C. rodentium-infectedmice
Because ornithine, the product of arginase, is metabolized by ODCto form polyamines, we investigated ODC expression in the colonof C. rodentium-infected WT mice. By real-time PCR, we found a2.5 � 0.5-fold increase of mRNA level in infected mice (n � 13)compared with control mice (n � 4; data not shown). However, a�40-fold increase of ODC activity was measured in the colon ofeither infected WT or WT L-Arg mice (Fig. 7A). This increase wasnot likely to be due to ODC activity from C. rodentium itself,because we measured bacterial ODC activity, and determined that itrepresented no more than 1% of the total ODC activity in the tissue.There was a 1.8 � 0.1-fold increase in colonic polyamines in C.rodentium-infected WT mice, and a significant, further increase of2.7 � 0.2-fold with L-Arg treatment (Fig. 7B), indicating that arginaseactivity was an important determinant of polyamine synthesis.
Exacerbation of C. rodentium colitis by arginase or ODCinhibition
To further demonstrate the beneficial effect of arginase and poly-amine formation, we conducted experiments with BEC andDFMO, inhibitors of arginase and ODC, respectively. In unin-fected control mice, BEC or DFMO treatment had no effect (TableI). However, there was a significant loss of survival in C. roden-tium-infected WT mice treated with BEC or with DFMO (Table I).In fact, the experiments needed to be terminated at 10 days afterinfection, because of the deaths and severe disease at this point.The colons of C. rodentium-BEC and C. rodentium-DFMO groupshad a greater increase in weight and histologic injury than those ofthe C. rodentium-water group (Table I). When compared with theinfected WT mice treated with water (Fig. 8A), the colons of bothBEC (Fig. 8B)- and DFMO (Fig. 8C)-treated mice showed markedtransmural inflammation and mucin depletion. The BEC-treated
FIGURE 6. Cytokine mRNA levels for IFN-� (A), TNF-� (B), and IL-1(C) in C. rodentium-infected mice compared with uninfected WT control.Mice were inoculated and sacrificed, as in Fig. 5. mRNA levels were de-termined by real-time PCR, as described in Materials and Methods. n �3–5 per group. ��, p � 0.01 vs WT uninfected control; §, p � 0.05; §§, p �0.01 vs WT C. rodentium. Uninfected iNOS�/� tissues had similar valuesas the WT uninfected tissues.
FIGURE 7. Colonic ODC activity (A) and polyamine concentrations(B) in C. rodentium-infected WT mice. n � 4 for control, n � 7 for C.rodentium, n � 5 for C. rodentium L-Arg. �, p � 0.05; ��, p � 0.01 vscontrol; §§, p � 0.01 vs C. rodentium.
2114 ROLE OF ARGINASE IN COLITIS
by guest on May 22, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

mice had substantial submucosal abscess formation, and theDFMO-treated mice exhibited mucosal and submucosal hemor-rhage, both indicative of severe acute inflammation.
To further confirm the importance of arginase, BEC was admin-istered to iNOS�/� C. rodentium-infected mice. When comparedwith iNOS�/� alone, BEC caused a significant worsening of colonhistologic injury scores (iNOS�/�: 2.42 � 0.46, n � 12 vsiNOS�/� BEC: 6.44 � 0.98, n � 8, p � 0.01) and colon weight(iNOS�/�: 0.36 � 0.01% of total body weight vs iNOS�/� BEC: 0.92 � 0.20%, p � 0.05).
DiscussionOur data demonstrate for the first time an important effect of ar-ginase in the intestine in vivo, namely a beneficial role of thearginase-ODC metabolic pathway in colitis. Arginase I and iNOSare two enzymes up-regulated in C. rodentium colitis that metab-olize L-Arg. Because iNOS deletion ameliorated colitis, and L-Argsupplementation was protective in both WT and iNOS�/� mice,our data indicate that arginase exerts its salutary effects by increas-ing polyamines in addition to inhibiting NO production. This wasconfirmed by the in vivo use of BEC, an arginase inhibitor. Theimprovement in animals fed L-Arg is most likely due to enhancedformation of polyamines, because an increase of polyamine con-centration was demonstrated in L-Arg-treated mice, and ODC in-hibition with DFMO worsened the disease.
The fact that L-Arg treatment resulted in improvement mostlikely derives from the systemic depletion of L-Arg that occurredin the C. rodentium-infected mice. We measured serum L-Argrather than tissue levels, because L-Arg is in constant flux betweenthe intracellular and extracellular space (25), making serum levelsa more reliable means to measure L-Arg availability. As an exam-ple of this, in mice with targeted deletion of arginase II, the met-abolic effect was measured in the serum in which significant ac-cumulation of L-Arg was demonstrated in these mice (26).
It should be noted that there was a marked (40-fold) increase inODC activity in C. rodentium colitis tissues, but only a 2-foldincrease in polyamine levels. This is due in part to the fact thatpolyamines are rapidly acetylated, which results in their efflux outof cells and excretion (27). Additionally, polyamine synthesis isdependent on availability of L-ornithine substrate for ODC, whichderives from L-Arg by the activity of arginase; our current datashow that arginase activity is increased only �2-fold in the colitistissues, so this can be rate limiting in polyamine synthesis. Anotherimportant point is that DFMO is an irreversible inhibitor of ODC(28), and ODC is absolutely required for polyamine synthesis.Therefore, at an effective dose of DFMO, supplementation of L-Arg or iNOS deletion would not be expected to reverse the effectof DFMO on polyamine synthesis or the exacerbation of colitis.
The potent inhibition of ODC by DFMO is also illustrated by themore severe exacerbation of colitis by DFMO than by BEC. It is
likely that C. rodentium infection has a direct effect on ODC induc-tion. Because we observed a 2.5-fold increase in ODC mRNA, thereis activation of mRNA expression, but because of the 40-fold increasein enzyme activity, it is likely that there may be posttranslationaleffects on ODC activity as well. Direct activation of ODC by C. ro-dentium is also supported by our findings of an 87-fold increase inODC activity in intestinal epithelial-6 cells stimulated in vitro with C.rodentium for 4 h (our unpublished data).
To our knowledge, this is also the first report of the activation ofarginase I in the gastrointestinal tract under pathophysiologic con-ditions. We have reported that arginase II mRNA and protein lev-els were up-regulated in the stomach of Helicobacter pylori-in-fected mice and humans, but arginase I was not induced in thesetissues (11). Murine macrophages and dendritic cells express ar-ginase I when stimulated by Th2 cytokines (29), LPS (30), orcAMP (15). Even though C. rodentium infection is a Th1-drivendisease, with no increase in Th2 cytokines (20) (our own results),arginase I expression was induced in this model. We have alsofound that in vitro, arginase I is induced in C. rodentium-stimu-lated murine macrophages (our unpublished data), consistent withexpression in lamina propria cells in our immunohistochemistrydata. It is possible that the selective expression of arginase I maybe due to: 1) a specific activation by C. rodentium and/or its re-leased factors; 2) a colon-specific expression; or 3) selective in-duction in an acute form of inflammation vs chronic inflammation.In studies in mouse DSS (31) and rat TNBS colitis tissues (ourunpublished data), we also found that arginase I, and not arginaseII, is up-regulated; therefore, the expression of arginase I in colitisis not specific to C. rodentium. Because it has been shown thatarginase I is up-regulated in a mouse model of acute immune com-plex-induced inflammation of the nephron (32) and in macro-phages from an acute wounded rat model (30), the expression ofarginase I is not specific to the colon. It is also unlikely that argi-nase I occurs only in acute inflammation, because we have ob-served increased expression of arginase I in human inflammatorybowel disease tissues (discussed below), in which there is achronic component to the disease and the histologic injury pattern.
It is likely that the main effect of L-Arg treatment is enhancedsubstrate availability for both arginase and iNOS. It should benoted that we demonstrated increased arginase activity in the co-litis tissues, while showing increased iNOS activity by measuringserum NO production. There is abundant published evidence thatiNOS enzymatic activity is increased in rodent models of colitisand in human inflammatory bowel disease and that this correlateswith iNOS expression; iNOS activity is regulated in vitro and invivo by enzyme abundance (see Ref. 33 for our review on thissubject). We measured serum NO production to yield a more sen-sitive means to determine the relationship of L-Arg consumption toiNOS activity. The fact that the histologic scores of infected WTL-Arg-treated mice are not significantly different from those of
Table I. Effect of 0.1% BEC or 2.5% DFMO on WT mice 10 days after inoculation with C. rodentium or PBS control
n Survival (%) Body Weight (% Total Body Weight) Colon Weight (% Total Body Weight) Histology Score
Control 7 100 102.6 � 1.6 0.51 � 0.04 0.43 � 0.05Control BEC 5 100 110.1 � 2.2 0.42 � 0.04 0.67 � 0.17Control DFMO 3 100 101.3 � 2.6 0.61 � 0.07 0.75 � 0.25C. rodentium 11 81.8 98.7 � 2.7 0.83 � 0.08a 3.82 � 1.01b
C. rodentium BEC 9 55.5 82.4 � 5.2a,c 1.25 � 0.07b,d 9.25 � 0.69b,d
C. rodentium DFMO 6 0b,d 78.6 � 0.4b,c 1.35 � 0.12b,d 9.54 � 0.68b,d
a p � 0.05 vs control.b p � 0.01 vs control.c p � 0.05 vs C. rodentium.d p � 0.01 vs C. rodentium.
2115The Journal of Immunology
by guest on May 22, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

water-treated mice could be due to an overproduction of NO, coun-teracting the beneficial effect of arginase metabolism and poly-amine synthesis. Because NO can inhibit ODC by S-nitrosylation(34), this could interfere with colonic ODC activity.
Polyamines have been shown to have numerous biological func-tions that may be relevant to amelioration of colitis, including in-hibition of monocyte activation and Th1 cytokine production (35),stimulation of epithelial restitution by enhancing cell migration(36) and proliferation (37), and regulation of apoptosis (38). It hasbeen demonstrated that polyamine production is essential for therepair of rat duodenal mucosa after stress (39), and that ODC ac-tivity is increased in the acetic acid colitis model (40). Arginaseactivity can also result in increased collagen production throughproline synthesis from L-ornithine by the enzyme ornithine ami-notransferase (25). Collagen production and polyamine synthesishave both been proposed as major events leading to wound healingby favoring fibroblast replication (41).
We have addressed the potential effect of modulation of poly-amine levels on immune response by measuring Th1 and proin-flammatory cytokines and demonstrating that the large increases inIFN-�, TNF-�, and IL-1 in C. rodentium colitis are ameliorated byeither L-Arg treatment or iNOS deletion. These findings may beattributable to enhanced substrate availability for polyamine syn-thesis in these groups; however, it should be recognized that thesecytokines are markers of inflammation and our results may simplyrepresent a correlation with the improvement of colitis. We havealso found that there is induction of epithelial apoptosis in C. ro-dentium colitis tissues by TUNEL assay (data not shown) and asignificant increase in apoptosis in vitro in intestinal epithelial-6cells stimulated with C. rodentium (our unpublished data). Theinteraction of epithelial apoptosis, proliferation, and cell migrationin response to C. rodentium with acute vs chronic infection islikely to be important and is a complex area that requires addi-tional investigation.
Similar to our findings, iNOS expression was shown to be up-regulated in epithelial and lamina propria cells of mice infectedwith C. rodentium in a recent report (42). However, there was noreduction in bacterial colonization or of colonic weight or histo-
logic injury in iNOS�/� mice in this study, although there was animprovement in survival that was not statistically significant (42).We attribute these differences from our results in the current studyof significant improvement of each of the above parameters iniNOS�/� mice to several possibilities, including their use of alower inoculum of C. rodentium, younger age of infected mice(3–4 wk vs 8 wk in our study), and specific pathogen-free housingvs conventional housing in our study. A second report showed nodifferences between WT and iNOS�/� mice (43), but in that studya different strain was used (E. coli attaching and effacing (eae)-deficient mutant complemented with eae from EPEC) and coloni-zation studies revealed �2 log orders less C. rodentium/g of tissuethan we recovered at day 14 in WT mice, indicating importantdifferences in their model. Our data show a correlation of de-creased colonization levels of iNOS�/� mice with decreased his-tologic injury, proinflammatory cytokine levels, and clinicalcourse. However, there is a more complex relationship betweencolonization and indicators of disease, because L-Arg led to im-provement of both WT and iNOS�/� mice without decreasingcolonization. Our present results are strengthened by the fact thatwe have obtained similar preliminary results in the DSS model ofcolitis (31). L-Arg improved, while BEC worsened clinical andhistologic features of colitis in both WT and iNOS�/� mice (31),indicating that the findings of a beneficial role of arginase are notlimited to one model of colitis.
In human inflammatory bowel disease, an increase in ODC ac-tivity has been described in ulcerative colitis and Crohn’s diseasein children (44), while a decrease in ODC activity has been re-ported in severe human colitis (45). Interestingly, a decrease inODC activity, but an increase in mucosal spermidine concentrationhas been reported in patients with ulcerative colitis (46). We havefound that expression of arginase I, arginase II, and ODC mRNAis up-regulated in both human ulcerative colitis and Crohn’s dis-ease tissues (our unpublished data), providing relevance for themurine colitis studies presented in this work. Additionally, induc-tion of arginase II by cAMP has been described in human colonicepithelial cells (47); we have observed that cAMP can also inducearginase I expression in colonic cell lines (our unpublished data).
FIGURE 8. Photomicrographs of H&E-stained colon of WT mice infected with C. rodentium for 10 days and treated with A, water; B, BEC; or C,DFMO.
2116 ROLE OF ARGINASE IN COLITIS
by guest on May 22, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

In summary, our data define an important in vivo role for argi-nase as a regulator of mucosal inflammation. In addition to syn-thesis of polyamines in the colonic mucosa, alterations in luminaluptake, as well as polyamine metabolism and efflux may be im-portant events in colitis. Chronic overproduction of polyaminescould contribute to the risk for colorectal cancer associated withcolitis due to the growth-promoting effects of polyamines and theoxidative stress that occurs with polyamine metabolism by poly-amine oxidase. Nonetheless, enhancement of arginase and ODCactivities may represent important new strategies for amelioratinginflammatory bowel disease.
References1. Morteau, O., S. G. Morham, R. Sellon, L. A. Dieleman, R. Langenbach,
O. Smithies, and R. B. Sartor. 2000. Impaired mucosal defense to acute colonicinjury in mice lacking cyclooxygenase-1 or cyclooxygenase-2. J. Clin. Invest.105:469.
2. Fuss, I. J., M. Boirivant, B. Lacy, and W. Strober. 2002. The interrelated roles ofTGF-� and IL-10 in the regulation of experimental colitis. J. Immunol. 168:900.
3. Neurath, M. F., I. Fuss, B. L. Kelsall, E. Stuber, and W. Strober. 1995. Antibodiesto interleukin 12 abrogate established experimental colitis in mice. J. Exp. Med.182:1281.
4. Kankuri, E., K. Vaali, R. G. Knowles, M. Lahde, R. Korpela, H. Vapaatalo, andE. Moilanen. 2001. Suppression of acute experimental colitis by a highly selec-tive inducible nitric-oxide synthase inhibitor. N-[3-(aminomethyl)benzyl]aceta-midine. J. Pharmacol. Exp. Ther. 298:1128.
5. Dikopoulos, N., A. K. Nussler, S. Liptay, M. Bachem, M. Reinshagen,M. Stiegler, R. M. Schmid, G. Adler, and H. Weidenbach. 2001. Inhibition ofnitric oxide synthesis by aminoguanidine increases intestinal damage in the acutephase of rat TNB-colitis. Eur. J. Clin. Invest. 31:234.
6. McCafferty, D. M., J. S. Mudgett, M. G. Swain, and P. Kubes. 1997. Induciblenitric oxide synthase plays a critical role in resolving intestinal inflammation.Gastroenterology 112:1022.
7. McCafferty, D. M., M. Miampamba, E. Sihota, K. A. Sharkey, and P. Kubes.1999. Role of inducible nitric oxide synthase in trinitrobenzene sulphonic acidinduced colitis in mice. Gut 45:864.
8. Hokari, R., S. Kato, K. Matsuzaki, M. Kuroki, A. Iwai, A. Kawaguchi, S. Nagao,T. Miyahara, K. Itoh, E. Sekizuka, et al. 2001. Reduced sensitivity of induciblenitric oxide synthase-deficient mice to chronic colitis. Free Radical Biol. Med.31:153.
9. Krieglstein, C. F., W. H. Cerwinka, F. S. Laroux, J. W. Salter, J. M. Russell,G. Schuermann, M. B. Grisham, C. R. Ross, and D. N. Granger. 2001. Regulationof murine intestinal inflammation by reactive metabolites of oxygen and nitrogen:divergent roles of superoxide and nitric oxide. J. Exp. Med. 194:1207.
10. Gobert, A. P., S. Daulouede, M. Lepoivre, J. L. Boucher, B. Bouteille, A. Buguet,R. Cespuglio, B. Veyret, and P. Vincendeau. 2000. L-arginine availability mod-ulates local nitric oxide production and parasite killing in experimental trypano-somiasis. Infect. Immun. 68:4653.
11. Gobert, A. P., Y. Cheng, J. Y. Wang, J. L. Boucher, R. K. Iyer, S. D. Cederbaum,R. A. Casero, Jr., J. C. Newton, and K. T. Wilson. 2002. Helicobacter pyloriinduces macrophage apoptosis by activation of arginase II. J. Immunol. 168:4692.
12. Huang, J., F. J. DeGraves, S. D. Lenz, D. Gao, P. Feng, D. Li, T. Schlapp, andB. Kaltenboeck. 2002. The quantity of nitric oxide released by macrophagesregulates Chlamydia-induced disease. Proc. Natl. Acad. Sci. USA 99:3914.
13. Ignarro, L. J., G. M. Buga, L. H. Wei, P. M. Bauer, G. Wu, and P. del Soldato.2001. Role of the arginine-nitric oxide pathway in the regulation of vascularsmooth muscle cell proliferation. Proc. Natl. Acad. Sci. USA 98:4202.
14. Buga, G. M., L. H. Wei, P. M. Bauer, J. M. Fukuto, and L. J. Ignarro. 1998.NG-hydroxy-L-arginine and nitric oxide inhibit Caco-2 tumor cell proliferation bydistinct mechanisms. Am. J. Physiol. 275:R1256.
15. Morris, S. M., Jr., D. Kepka-Lenhart, and L. C. Chen. 1998. Differential regula-tion of arginases and inducible nitric oxide synthase in murine macrophage cells.Am. J. Physiol. 275:E740.
16. Iyer, R. K., J. M. Bando, C. P. Jenkinson, J. G. Vockley, P. S. Kim, R. M. Kern,S. D. Cederbaum, and W. W. Grody. 1998. Cloning and characterization of themouse and rat type II arginase genes. Mol. Genet. Metab. 63:168.
17. Li, H., C. J. Meininger, J. R. Hawker, Jr., T. E. Haynes, D. Kepka-Lenhart,S. K. Mistry, S. M. Morris, Jr., and G. Wu. 2001. Regulatory role of arginase Iand II in nitric oxide, polyamine, and proline syntheses in endothelial cells.Am. J. Physiol. 280:E75.
18. Gobert, A. P., D. J. McGee, M. Akhtar, G. L. Mendz, J. C. Newton, Y. Cheng,H. L. Mobley, and K. T. Wilson. 2001. Helicobacter pylori arginase inhibitsnitric oxide production by eukaryotic cells: a strategy for bacterial survival. Proc.Natl. Acad. Sci. USA 98:13844.
19. Higgins, L. M., G. Frankel, I. Connerton, N. S. Goncalves, G. Dougan, andT. T. MacDonald. 1999. Role of bacterial intimin in colonic hyperplasia andinflammation. Science 285:588.
20. Higgins, L. M., G. Frankel, G. Douce, G. Dougan, and T. T. MacDonald. 1999.Citrobacter rodentium infection in mice elicits a mucosal Th1 cytokine responseand lesions similar to those in murine inflammatory bowel disease. Infect. Immun.67:3031.
21. Kim, N. N., J. D. Cox, R. F. Baggio, F. A. Emig, S. K. Mistry, S. L. Harper,D. W. Speicher, S. M. Morris, Jr., D. E. Ash, A. Traish, and D. W. Christianson.
2001. Probing erectile function: S-(2-boronoethyl)-L-cysteine binds to arginase asa transition state analogue and enhances smooth muscle relaxation in humanpenile corpus cavernosum. Biochemistry 40:2678.
22. Wilson, K. T., S. Fu, K. S. Ramanujam, and S. J. Meltzer. 1998. Increasedexpression of inducible nitric oxide synthase and cyclooxygenase-2 in Barrett’sesophagus and associated adenocarcinomas. Cancer Res. 58:2929.
23. Fu, S., K. S. Ramanujam, A. Wong, G. T. Fantry, C. B. Drachenberg, S. P. James,S. J. Meltzer, and K. T. Wilson. 1999. Increased expression and cellular local-ization of inducible nitric oxide synthase and cyclooxygenase 2 in Helicobacterpylori gastritis. Gastroenterology 116:1319.
24. Casero, R. A., Jr., P. Celano, S. J. Ervin, C. W. Porter, R. J. Bergeron, andP. R. Libby. 1989. Differential induction of spermidine/spermine N1-acetyltrans-ferase in human lung cancer cells by the bis(ethyl)polyamine analogues. CancerRes. 49:3829.
25. Wu, G., and S. M. Morris, Jr. 1998. Arginine metabolism: nitric oxide and be-yond. Biochem. J. 336:1.
26. Shi, O., S. M. Morris, Jr., H. Zoghbi, C. W. Porter, and W. E. O’Brien. 2001.Generation of a mouse model for arginase II deficiency by targeted disruption ofthe arginase II gene. Mol. Cell. Biol. 21:811.
27. Pegg, A. E., R. Wechter, R. Pakala, and R. J. Bergeron. 1989. Effect of N1,N12-bis(ethyl)spermine and related compounds on growth and polyamine acetylation,content, and excretion in human colon tumor cells. J. Biol. Chem. 264:11744.
28. Poulin, R., L. Lu, B. Ackermann, P. Bey, and A. E. Pegg. 1992. Mechanism ofthe irreversible inactivation of mouse ornithine decarboxylase by �-difluorometh-ylornithine: characterization of sequences at the inhibitor and coenzyme bindingsites. J. Biol. Chem. 267:150.
29. Munder, M., K. Eichmann, J. M. Moran, F. Centeno, G. Soler, and M. Modolell.1999. Th1/Th2-regulated expression of arginase isoforms in murine macrophagesand dendritic cells. J. Immunol. 163:3771.
30. Louis, C. A., J. S. Reichner, W. L. Henry, Jr., B. Mastrofrancesco, T. Gotoh,M. Mori, and J. E. Albina. 1998. Distinct arginase isoforms expressed in primary andtransformed macrophages: regulation by oxygen tension. Am. J. Physiol. 274:R775.
31. Cheng, Y., H. Xu, J. S. Forman, P. C. Panchal, D. R. Blumberg, R. Chaturvedi,F. I. Bussiere, C. B. Drachenberg, A. P. Gobert, and K. T. Wilson. 2003. Inhi-bition of colitis by the arginase-ODC pathway. Gastroenterology 124:A473.
32. Waddington, S. N., K. Mosley, H. T. Cook, F. W. Tam, and V. Cattell. 1998.Arginase AI is up-regulated in acute immune complex-induced inflammation.Biochem. Biophys. Res. Commun. 247:84.
33. Cross, R. K., and K. T. Wilson. 2003. Nitric oxide in inflammatory bowel disease.Inflamm. Bowel Dis. 9:179.
34. Bauer, P. M., G. M. Buga, J. M. Fukuto, A. E. Pegg, and L. J. Ignarro. 2001.Nitric oxide inhibits ornithine decarboxylase via S-nitrosylation of cysteine 360in the active site of the enzyme. J. Biol. Chem. 276:34458.
35. Zhang, M., T. Caragine, H. Wang, P. S. Cohen, G. Botchkina, K. Soda,M. Bianchi, P. Ulrich, A. Cerami, B. Sherry, and K. J. Tracey. 1997. Spermineinhibits proinflammatory cytokine synthesis in human mononuclear cells: a counter-regulatory mechanism that restrains the immune response. J. Exp. Med. 185:1759.
36. Rao, J. N., L. Li, V. A. Golovina, O. Platoshyn, E. D. Strauch, J. X. Yuan, andJ. Y. Wang. 2001. Ca2-RhoA signaling pathway required for polyamine-depen-dent intestinal epithelial cell migration. Am. J. Physiol. 280:C993.
37. Li, L., J. N. Rao, X. Guo, L. Liu, R. Santora, B. L. Bass, and J. Y. Wang. 2001.Polyamine depletion stabilizes p53 resulting in inhibition of normal intestinalepithelial cell proliferation. Am. J. Physiol. 281:C941.
38. Schipper, R. G., L. C. Penning, and A. A. Verhofstad. 2000. Involvement ofpolyamines in apoptosis: facts and controversies: effectors or protectors? Semin.Cancer Biol. 10:55.
39. Wang, J. Y., and L. R. Johnson. 1991. Polyamines and ornithine decarboxylaseduring repair of duodenal mucosa after stress in rats. Gastroenterology 100:333.
40. Yamada, T., K. Fujimoto, P. Tso, T. Fujimoto, T. S. Gaginella, andM. B. Grisham. 1992. Misoprostol accelerates colonic mucosal repair in aceticacid-induced colitis. J. Pharmacol. Exp. Ther. 260:313.
41. Shearer, J. D., J. R. Richards, C. D. Mills, and M. D. Caldwell. 1997. Differentialregulation of macrophage arginine metabolism: a proposed role in wound heal-ing. Am. J. Physiol. 272:E181.
42. Vallance, B. A., W. Deng, M. De Grado, C. Chan, K. Jacobson, and B. B. Finlay.2002. Modulation of inducible nitric oxide synthase expression by the attachingand effacing bacterial pathogen Citrobacter rodentium in infected mice. Infect.Immun. 70:6424.
43. Simmons, C. P., N. S. Goncalves, M. Ghaem-Maghami, M. Bajaj-Elliott,S. Clare, B. Neves, G. Frankel, G. Dougan, and T. T. MacDonald. 2002. Impairedresistance and enhanced pathology during infection with a noninvasive, attach-ing-effacing enteric bacterial pathogen, Citrobacter rodentium, in mice lackingIL-12 or IFN-�. J. Immunol. 168:1804.
44. Pillai, R. B., V. Tolia, R. Rabah, P. M. Simpson, R. Vijesurier, and C. H. Lin.1999. Increased colonic ornithine decarboxylase activity in inflammatory boweldisease in children. Dig. Dis. Sci. 44:1565.
45. Ricci, G., G. Stabellini, G. Bersani, G. Marangoni, P. Fabbri, G. Gentili, andV. Alvisi. 1999. Ornithine decarboxylase in colonic mucosa from patients withmoderate or severe Crohn’s disease and ulcerative colitis. Eur. J. Gastroenterol.Hepatol. 11:903.
46. Obayashi, M., I. Matsui-Yuasa, T. Matsumoto, A. Kitano, K. Kobayashi, andS. Otani. 1992. Polyamine metabolism in colonic mucosa from patients withulcerative colitis. Am. J. Gastroenterol. 87:736.
47. Wei, L. H., S. M. Morris, Jr., S. D. Cederbaum, M. Mori, and L. J. Ignarro. 2000.Induction of arginase II in human caco-2 tumor cells by cyclic AMP. Arch.Biochem. Biophys. 374:255.
2117The Journal of Immunology
by guest on May 22, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from