prosurvival mechanisms i. endogenous prosurvival mechanisms ii. autophagy iii. induced prosurivival...
TRANSCRIPT

Prosurvival mechanisms
I. Endogenous prosurvival mechanismsII. AutophagyIII. Induced prosurivival mechanisms
MECHANISMS OF CARCINOGENESISSPRING SEMESTER
(Dean Tang, April 27, 2011)

Two Apoptotic Pathways

Prosurvival mechanisms: BH3-only proteins are tightly regulated
1. Like Egl-1, some BH3-only proteins (e.g., Puma, Noxa, Hrk, and Bim) are normally not expressed in some cell types and need transcriptional activation.
2. Some BH3-only proteins are expressed but are not lethal. For example, Bim in many cells is expressed but remains bound to tubulin or tubulin-based dynein motor complexes and therefore is not proapoptotic. Bid is expressed in many cells but generally needs a post-translational cleavage to generate proapoptotic tBid. Bad is abundant in many cells but inactive due to PHOSPHORYLATION and sequestration by 14-3-3 proteins.
BH3-onlyBCL-2
BH1-3pro-apoptoticproteins
adaptor caspaseanti-apoptoticBcl-2 proteins
BaxBak Apaf-1 Caspase-9
Cyto c
Caspase-3
Bad
Noxa

BAD: Phosphorylation sequesters it in the cytosol

*Phosphotidylinositol 3-kinase, or PI3K, is activated by multiple mitogens and survival factors. Prototypical examples include: NGF in neuronal cells, IL-3 and other cytokines in lymphocytes, PDGF and EGF in mesenchymal cells, and IGF-1 in epithelial cells.
*These tyrosine kinase receptors, as well as seven transmembrane G protein-coupled receptors and activated Ras, recruit PI3K to the membrane, which then catalyzes the transfer of phosphate from ATP to the D3 position of the inositol ring of membrane-localized phosphoinositides.
*The main products, PI3,4P and PI3,4,5P, function as signaling molecules that can activate TK, small G proteins, and ser/thr kinases such as atypical PKCs, pp70S6K, and c-Akt. Activation of c-Akt is the major survival function.
A Major Prosurvival Pathway: PI3K-Akt

*Akt (PKB) has a central kinase domain with ser/thr substrate specificities, an amino terminal PH (which mediates protein-lipid and/or protein-protein interactions), and a C-terminal hydrophobic and proline-rich domain.
*Direct binding of PI3,4,5P or related lipids to the PH domain in Akt leads to its translocation to plasma membrane. At this location and with PIP bound, Akt is phosphorylated (at Thr308 and Ser473) and activated by PKDs (3-phosphoinositide-dependent protein kinases). Integrin-linkedkinase (ILK) is also a PIP-dependent Akt kinase (Ser473).
*Activated Akt has three main apoptosis-related targets: Bad, caspase-9, and forkhead (FKH) transcription factors. Phosphorylation of Bad (at Ser136) and of caspase-9 inactivate these pro-apoptotic proteins. Phosphorylation of FKH and Bad sequesters them in the cytosol by binding to 14-3-3.
*Transfection with constitutively active Akt blocks apoptosis induced by deprivation, UV irradiation, matrix detachment, cell-cycle disruption, DNA damage, TGF, anti-Fas, etc.
*Conversely, trasnfection with dominant-negative alleles of Akt (catalytically inactive Akt, a mutant Akt consisting only of the PH domain) blocks the pro-survival functions of survival factors.
*PTEN is a lipid phosphatase that dephosphorylates PI3K-generated 3’-phosphorylated phosphotidylinositides.
*Mitogens and survival factors activate a parallel survival pathway: Raf-MEK-MAPK pathway. Ras-dependent survival is mediated by both PI3K and Raf-MEK-MAPK pathway.

Prosurvival mechanisms: BH3-only proteins are tightly regulated
1. Like Egl-1, some BH3-only proteins (e.g., Puma, Noxa, Hrk, and Bim) are normally not expressed in some cell types and need transcriptional activation.
2. Some BH3-only proteins are expressed but are not lethal. For example, Bim in many cells is expressed but remains bound to tubulin or tubulin-based dynein motor complexes and therefore is not proapoptotic. Bid is expressed in many cells but generally needs a post-translational cleavage to generate proapoptotic tBid. Bad is abundant in many cells but inactive due to PHOSPHORYLATION and sequestration by 14-3-3 proteins.
3. Bim undergoes biallelic deletion in mantle cell lymphoma and promoter methylation in other B-cell lymphomas. A few human lymphomas also have Noxa gene mutations. Bik/Nbk is mutated in renal cell carcinomas.
BH3-onlyBCL-2
BH1-3pro-apoptoticproteins
adaptor caspaseanti-apoptoticBcl-2 proteins
BaxBak Apaf-1 Caspase-9
Cyto c
Caspase-3
Bad
Noxa

BH3-only proteins may have physiological/prosurvival functions
*BID is required for myeloid homeostasis: Bid-deficient mice, as they age, spontaneously develop a myeloproliferative disorder resembling CMML (chronic myelomonocytic leukemia).
*Bid plays a role in DNA-damage response and in promoting cell survival (Zinkel SS, et al., Cell 122, 579-591, 2005; Kamer I et al., Cell 122, 593-603, 2005).
*Bad is involved in normal glycolysis and in promoting cell survival (Danial NN et al., Nature 424, 952-956, 2003; Seo et al., JBC 279, 42240-42249, 2004).
*Most bim-deficient mice die embryonically, suggesting Bim may also have a normal physiologicalfunction. Bim may have a role in regulating cytoskeletal integrity.
*bid-/- mice (Yin et al., Nature 400:886-891, 1999.) --- Animals develop normally --- Resistant to anti-Fas induced liver failure
*bim-/- mice (Bouillet et al., Science 286, 1735-1738, 1999.) --- A significant number of bim-null, and even some of the bim+/-
mice die in utero prior to E9.5 for unknown reasons. --- Bim deficiency causes an SLE (systematic lupus erythematosus)-
like autoimmune disease.

Anti-apoptotic Bcl-2 proteins function as endogenous prosurvival molecules
1. Bcl-2 and Bcl-xL are indispensable for stem cell development and are critical for the survival of stem cells and many mature cell types. Bcl-2 and/or Bcl-xL are overexpressed perhaps in all tumor cells.
2. Mcl-1 is critical for the survival of HSCs, neutrophils (not macrophages), fibroblasts, keratinocytes, and many other cell types. Mcl-1 is also overexpressed in several cancers.
3. Bcl-w is expressed in a majority of invasive gastric adenocarcinomas.
BH3-onlyBCL-2
BH1-3pro-apoptoticproteins
adaptor caspaseanti-apoptoticBcl-2 proteins
BaxBak Apaf-1 Caspase-9
Cyto c
Caspase-3
Bad
Noxa

*bcl-2-/- mice (Nakayama et al., Science 261: 1584, 1993; Veis et al., Cell 75:229-240, 1993) --- Defects in renal development (essential for kid. stem cells) --- Premature loss of B/T lymphoid cells: Immunodeficiency --- Defects in melanocyte development (essential for the survival of melanocyte
stem cells): Loss of hair pigmentation
Knockouts of anti-apoptotic Bcl-2 family proteins
*bcl-xL-/- mice (Motoyama et al., Science 267:1506-1510, 1995.) --- Animals die during embryogenesis --- Massive apoptosis in developing post-mitotic cells of the embryonic spinal cord,
brain stem, and DRG --- Extensive apoptosis is also observed within the liver and hematopoietic systems --- Haploinsufficiency for Bcl-xL causes thrombocytopenia (Mason KD et al., Cell 128,
1173-1186, 2007).
*mcl-1-/- mice --- defects in survival and implantation of zygote (Rinkenberger et al., G&D 14:23, 2000) --- in conditional KO, critical in early hematopoiesis and B/T lymphopoiesis
(Opferman JT et al., Nature 426, 671-676, 2003; Science 307, 1101-4, 2005)
*bcl-w-/- mice --- defects in spermatogenesis (Print CG et al., PNAS 95, 12424-31, 1998; Ross AJ
et al., Nat Genet. 18, 251-6, 1998)

Bcl-2 proteins control apoptosis
BH3-onlyBCL-2
BH1-3pro-apoptoticproteins
adaptor caspaseanti-apoptoticBcl-2 proteins
BaxBak Apaf-1 Caspase-9
Cyto c
Caspase-3
Bad
Noxa

Bax and/or Bak knockouts
*bak-/- mice (Lindsten et al., Mol. Cell 6:1389-1399, 2000) --- Animals develop normally --- bak-/- cells show normal apoptotic responses to both extrinsic
and intrinsic apoptotic stimuli *bax-/-bak-/- mice: --- bak-/- bax-/- mice die perinatally with <10% animals reaching adulthood --- Adult bak-/- bax-/- mice display multiple phenotypic abnormalities (retain
interdigital webs, imperforate vaginas, neurological abnormalities including deafness, and circling behavior, increased numbers of neuronal and hematopoietic progenitors; leading to elevated numbers of granulocytes and mature T/B lymphocytes and enlarged spleen and LN and infiltration of parenchymal organs; bak-/- bax-/- cells are resistant to
death induced by neglect or irradiation but sensitive to Fas.)
*bax-/- mice (Knudson et al., Science 270:96-99, 1995) --- Animals develop rather normally --- Increased numbers of sympathetic and motor neurons --- In vitro, bax-/- sympathetic neurons are resistant to cell death
upon trophic factor withdrawal --- Male homozygous mice are sterile due to a paradoxical increase
in the death of spermatogonia

*Unstimulated, Bax is monomeric and inactive in the cytosol, probably due to its concealed amino (or carboxy) termini. Bax is also normally “sequestered” in the cytosol as a result of binding to multiple proteins including 14-3-3 proteins, Ku70, the peptide humanin, HSP70, crystallins, andARC or Apoptosis Repressor with Caspase recruitment domain).
*Recently, Bax has been shown to be retro-translocated by Bcl-xL to block its proapoptotic effect(Edilich et al., Cell 145: 104-116, 2011).
*Different from Bax, most Bak is constitutively expressed in the mitochondria by associating with VDAC2.
*Bax and Bak modulate the UPR by a direct interaction with IRE1 (inositol-requiring enzyme 1)(Hetz et al., Science 312, 572, 2006)
•Bax and Bak play an important role in mitochondrial morphogenesis (Karbowski M, Norris KL, Cleland MM, Jeong SY, Youle RJ. Nature 443:658-62, 2006)
*Bak has neuroprotective effect (Fannjiang Y et al., Dev. Cell. 4, 575-585, 2003).
*In some cancer cells, Bax and Bak are mutated.
Bax and Bak may have non-apoptotic and physiological functions
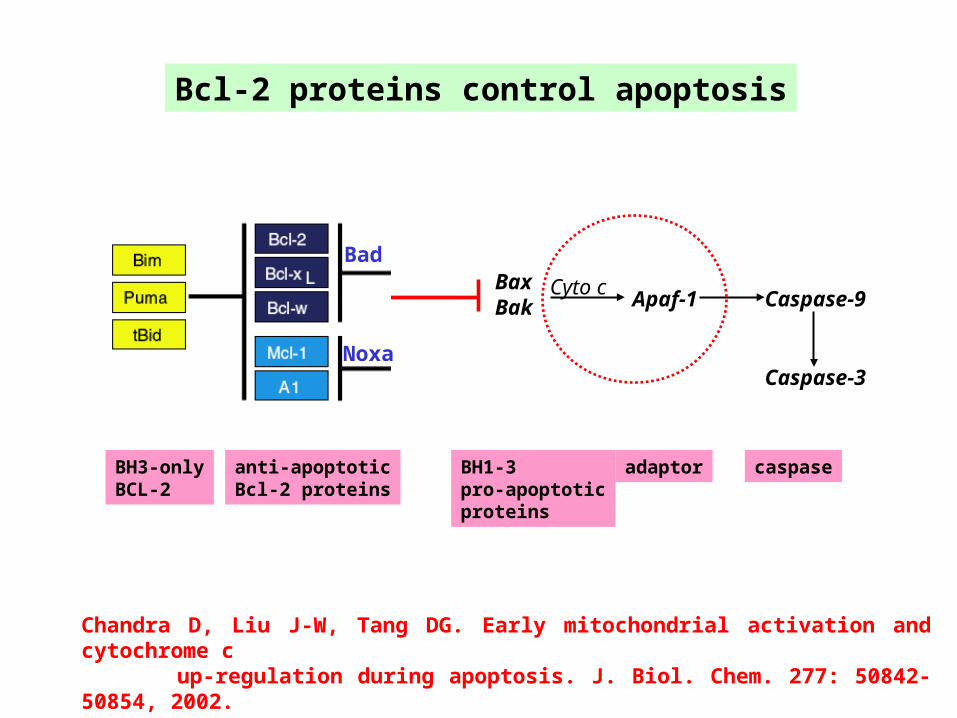
Bcl-2 proteins control apoptosis
BH3-onlyBCL-2
BH1-3pro-apoptoticproteins
adaptor caspaseanti-apoptoticBcl-2 proteins
BaxBak Apaf-1 Caspase-9
Cyto c
Caspase-3
Bad
Noxa
Chandra D, Liu J-W, Tang DG. Early mitochondrial activation and cytochrome c up-regulation during apoptosis. J. Biol. Chem. 277: 50842-50854, 2002.

Holo-cyto. c
-COOHApaf-1 CARD NOD WD-40 repeats (13)
NH2-
1 124899-109 415
Shi, Y., Mol. Cell, 9, 459-470, 2002

Holo-cyto. c
Apaf-1 -COOH CARD NOD WD-40 repeats (13)
NH2-
1 124899-109 415
Apoptosome Formation can be Reconstituted using Recombinant Proteins --- A Process that Requires (d)ATP
+
+(d)ATP(0.1-0.5 mM)

Holo-cyto. c
-COOHApaf-1 CARD NOD WD-40 repeats (13)
NH2-
1 124899-109 415
Physiological(mM) levelsof (deoxy)N
Caspase-9 activation
Chandra D, Bratton SB, Person MD, Tian Y, Martin AG, Ayers M, Fearnhead HO, Gandhi V, and Tang DG. Intracellular nucleotides act as critical prosurvival factors by binding to cytochrome c and inhibiting apoptosome. Cell 125: 1333-1346, 2006.

Holo-cyto. c
-COOHApaf-1 CARD NOD WD-40 repeats (13)
NH2-
1 124899-109 415
Cytochrome c
Apaf-1
Caspase-9
Caspase-3
Apoptosis
Apoptosome Physiological(mM) levelsof (deoxy)N

*Physiological levels of K+ also inhibits cytochrome c-dependent apoptosome formation (JBC, 276, 41985-990, 2001).
*Bao Q, Lu W, Rabinowitz JD, Shi Y. Calcium blocks formation of apoptosome by preventing nucleotide exchange in Apaf-1. Mol Cell. 2007 Jan 26;25(2):181-92.

Caspases are executioners of apoptosis
BH3-onlyBCL-2
BH1-3pro-apoptoticproteins
adaptor caspaseanti-apoptoticBcl-2 proteins
BaxBak Apaf-1 Caspase-9
Cyto c
Caspase-3
Bad
Noxa

Prodomain Large subunit Small subunit
Linker
p10p20p3-26
Prodomains:In executioner caspases: ~3kd In initiator caspases: 10-26 kd
Types of caspases:-Initiator caspases: caspase-2, -8, -9 and -10-Executioner Caspases: caspase-3, -6 and -7
Caspase Structure:
Caspase prodomains:
D. Chandra

Mol. Cell9, 459-470,2002

Caspase substrates
Proteosome components (Mol Cell 14, 81-93, 2004) Mitochondrial complex I p75 subunit (Cell 117, 773-786, 2004)
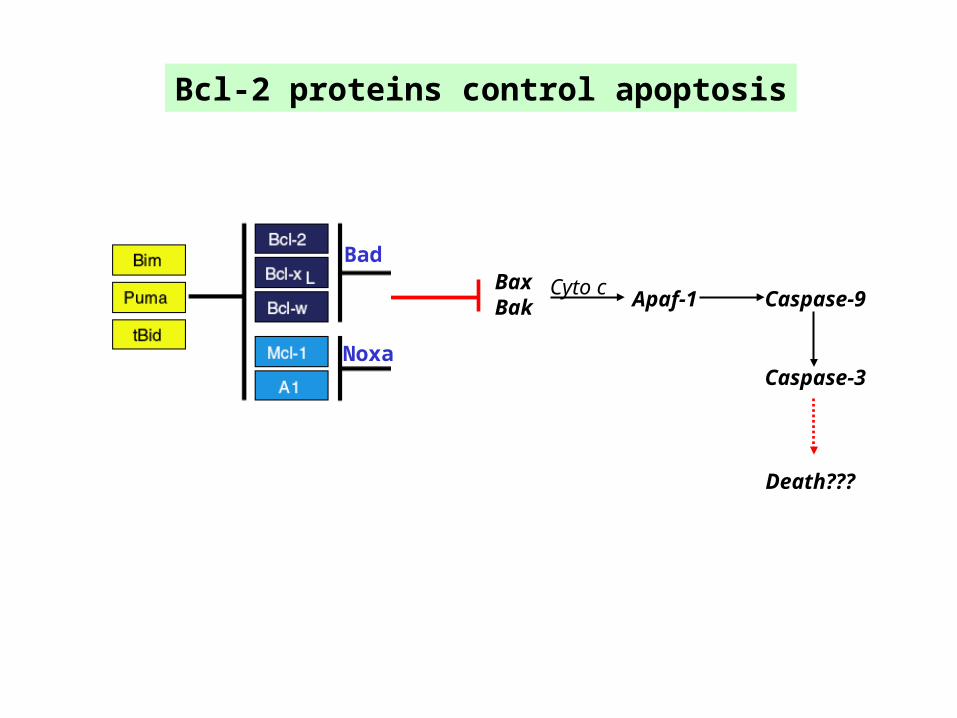
Bcl-2 proteins control apoptosis
BaxBak Apaf-1 Caspase-9
Cyto c
Caspase-3
Bad
Noxa
Death???

IAPs

Inhibitor-of-Apoptosis Proteins
604BIR1 BIR2 BIR3 RINGCARD
BIR1 BIR2 BIR3 RINGCARD618
BIR1 BIR2 BIR31403
BIR UBC4845
BIR142
1BIR1 BIR2 BIR3 RING
497XIAP
cIAP-1
NAIP
BRUCE
Survivin
cIAP-2
BIR3 RING236
ILP-2
BIR RING298
Livin
S. Bratton

IAPs

Shi, Y. Mol. Cell, 9, 459-470, 2002
IAPs function as critical prosurvival molecules
*IAPs (especially XIAP) are generally overexpreessedin multiple types of cancer cells
Smac (second mitochondrialactivator of caspases)
apoptotic stimulation

Caspases may have apoptosis-independent/physiological functions
Drosophila caspases (Cell 126, 583-596, 2006)--- Non-apoptotic caspase activity is required for proper sensory organ precursor development.
Caspase-1--- Promotes cell survival upon toxin challenge possibly through membrane repair (Cell 126,
1135-1145, 2006).--- Connects immunity (host defense) and apoptotic machinery (Bruey J-M., Cell 129, 45-56, 2007).
Caspase-3 --- Caspase-3 activation is involved in cancer cell invasion (Cancer Res, 65, 9121-9125, 2005).
Caspase-8 (Nature 419, 395, 2002; Genes & Dev 17, 883, 2003; Science 307, 1465, 2005)--- Targeted disruption of casp-8 in T-cell lineage leads to marked decrease in peripheral T-cells
and impaired T-cell response ex vivo to activation stimuli.--- Casp-8 ablation protected thymocytes and activated T-cells from CD95 ligation but not
anti-CD3-induced apoptosis, or apoptosis activated by the mitochondrial stimuli--- Casp-8 mutant mice were unable to mount an immune response to viral infection--- Casp-8 required for NF-kB activation by antigen receptor--- Casp-8 promotes cell motility and calpain activity (Cancer Res., 66, 4273-4278, 2006)--- However, loss of caspase-8 potentiates neuroblastoma metastasis (Nature 439, 95-99, 2006)--- Caspase-8 interacts with androgen receptor (AR) and regulates AR-driven gene expression
(Qi W et al., EMBO J. 26, 65-75, 2007)

Prosurvival mechanisms
I. Endogenous prosurvival mechanismsII. AutophagyIII. Induced prosurivival mechanisms

Autophagy*First recognized under EM early 1960s.*Also called macroautophagy: Self-eating to survive.*A unique form of membrane trafficking in which membrane compartments (autophagosomes) engulf both organelles and cytosolic macromolecules and deliver them to the lysosome for degradation.

Mizushima N. CDD, 12, 1535-1541, 2005

Autophagy*Molecular insights first came from studies in yeast, which expresses >20 autophagy-related genes (ATGs). *In yeast, two ubiquitin-like conjugation systems, Atg8-PE (phosphatidylethanolamine; Atg8 = LC3 in
mammals) and Atg12-Atg5, are involved in forming the initial autophagic vesicles.

Yorimitsu and Klionsky. CDD, 12, 1542-1552, 2005
Autophagosome nucleation requires a complex containingAtg6 (Beclin 1 in mammals) that recruits the class III PI3KVPS34 to generate PI3P. Expansion of autophagosome membranesinvolves Atg12 and Atg8 (or LC3).

Autophagy: A Major Prosurvival Mechanism
- In yeast, the primary function of autophagy is to maintain viability during starvation.- In mice, autophagy supports newborn survival at birth - in the absence of Atg5, mice do not survive neonatal starvation and die within a day after birth (Kuma et al., Nature 432, 1032, 2004).-In fact, autophagy-defective embryos fail to reach the blastocyct stage (Science 321:117, 2008).- In IL-3-dependent lymphocytes, cells without Bax and Bak survive for weeks due to autophagy - knockdown of Atg7 significantly reduces this survival (Lum et al., Cell 120, 237-48, 2005).- Autophagy-supported cell survival probably results from the products of self-degradation in lysosomes (e.g., amino acids), which support the TCA cycle to generate ATP.- Animal cells exhibit low levels of constitutive autophagy - a defense mechanism to eliminate aggregation-prone proteins (inclusion bodies) in, e.g., neurons and liver cells (absence of Atg5 and Atg7 in mouse brain induces neurodegenerative diseases (Hara T et al., Nature 441, 885, 2006; Komatsu M et al., Nature 441, 880, 2006).-Autophagy sequesters and kills the invading pathogens (Nakagawa I et al., Science 306, 1037, 2004).- Autophagy helps eliminate superfluous organelles (peroxisomes, mito., ER, etc) and maintain cellular homeostasis).- Defects in autophagy have been linked to liver diseases, neurodegeneration, Crohn’s disease, aging,
metabolic syndrome and cancer
*Indeed, autophagy functions as prosurvival mechanisms in cancer cells and mediate their resistance to therapeutics (Albedin MJ, et al., CDD, 14, 500-510, 2007; Katayama M et al., CDD, 14, 548-558, 2007; Amaravadi RK et al., JCI, 117:326-336, 2007).

Autophagy & Metabolism
Rabinowitz JD & White E. Autophagy & Metabolism. Science 330: 1344-1348, 2010.

Autophagy & Metabolism
Rabinowitz JD & White E. Autophagy & Metabolism. Science 330: 1344-1348, 2010.

Levine B and Yuan J. JCI, 115, 2679-2688, 2005Yoshimori T et al., Cell 128, 833-836, 2007
Autophagy (Type II PCD) & Apoptosis (Type I PCD)
*Autophagic cell death: nonapoptotic PCD (type II)*A large number of autophagosomes are present in dying cells: Cause-and-effect relationship unclear*The first example: mouse L929 fibroblasts treated with zVAD cells undergo nonapoptotic cell
death knockdown of Atg7 or Beclin-1 reduced cell death + zVAD treatment somehowcaused degradation of catalase resulting in increased ROS production followed by celldeath (Yu L et al., Science 304, 1500-1502, 2004).
*In Bax/Bak-DKO MEFs treated with chemicals such as etoposide cells die through ‘autophagy’ -cells do not die when Atg5 and Beclin-1 were silenced (Shimizu et al., NCB 6, 1221-1228, 2004).
*’Autophagy’ might act upstream of apoptosis: apoptosis of CD4 T cells by HIV envelope glyco-proteins inhibited when Beclin-1 and Atg7 were silenced (Espert L et al., JCI, 116,2161-72, 2006).
*Problems: 1) Autophagy is a morphological and descriptive term; 2) The Atg proteins may have autophagy-unrelated functions; 3) Truncated Atg5 can bind Bcl-xL in mitochondria and causes apoptosis (Yousefi S
et al., NCB 8, 1124-32, 2006). 4) Bcl-2 negatively regulates autophagy by binding to Beclin-1 (Pattingre S et al., Cell
122, 927-39, 2005). 5) ‘Autophagic cell death’ might be caused by excessive nonselective autophagy itself.

Yoshimori T et al., Cell 128, 833-836, 2007Qu X et al., Cell 128, 931-46, 2007

Prosurvival mechanisms
I. Endogenous prosurvival mechanismsII. AutophagyIII. Induced prosurivival mechanisms
Cell
Apoptotic stimulation
Survival Death

Two Apoptotic Pathways

Liu et al., ONCOGENE 2005
Induction of prosurvivalmolecules during apoptosis induction

Micheau O. & Tschopp J. Cell 114, 181-190, 2003
FasL (EMBO J. 23, 3175, 2004)--- FasL promotes tumor cell migration and invasion.
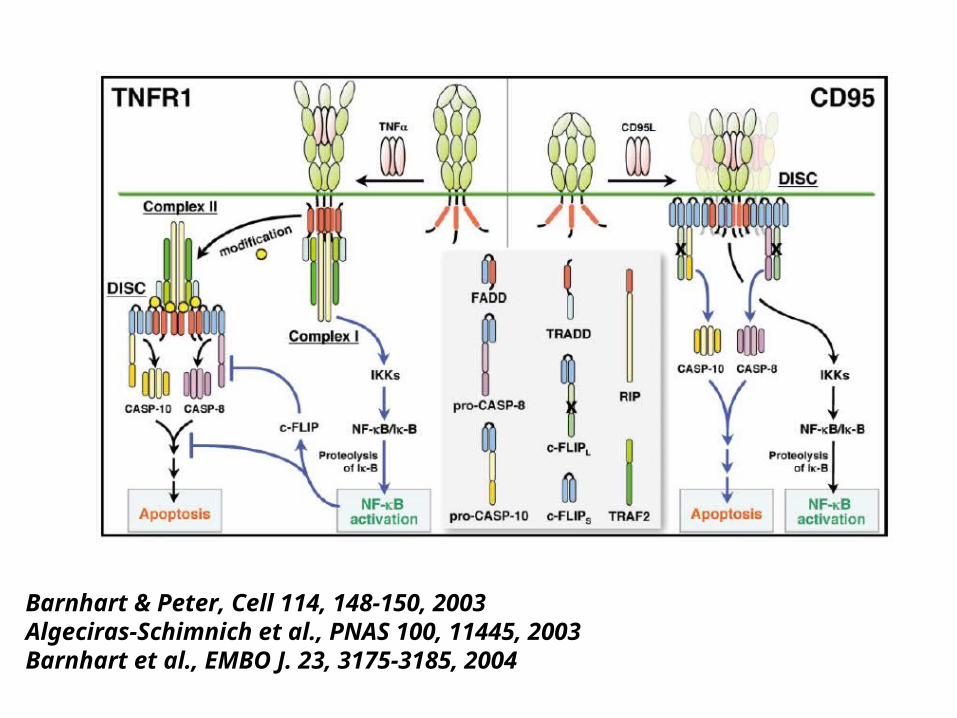
Barnhart & Peter, Cell 114, 148-150, 2003Algeciras-Schimnich et al., PNAS 100, 11445, 2003Barnhart et al., EMBO J. 23, 3175-3185, 2004

Prosurvival mechanisms
I. Endogenous prosurvival mechanismsII. AutophagyIII. Induced prosurivival mechanisms
Cell
Apoptotic stimulation
Survival Death