projecting forest stand structures using stand dynamics principles
TRANSCRIPT

Copyright, 1991, David R. Larsen
Projecting Forest Stand Structures Using Stand Dynamics
Principles: An Adaptive Approach
by
David Rolf Larsen
A dissertation submitted in partial ful�llment
of the requirements for the degree of
Doctor of Philosophy
University of Washington
1991
Approved by
(Chairperson of Supervisory Committee)
Program Authorized
to O�er Degree
Date


In presenting this dissertation in partial ful�llment of the requirements of the Doctoral
degree at the University of Washington, I agree that the Library shall make its
copies freely available for inspection. I further agree that extensive copying of this
dissertation is allowable only for scholarly purposes, consistent with \fair use" as
prescribed in the U. S. Copyright Law. Requests for copying and reproduction of
this dissertation may be referred to University Micro�lms, 300 North Zeeb Road,
Ann Arbor, Michigan 48106, to whom the author has granted \the right to reproduce
and sell (a) copies of the manuscript in micro�lm and/or (b) printed copies of the
manuscript from micro�lm."
Signature
Date


University of Washington
Abstract
Projecting Forest Stand Structures Using Stand Dynamics
Principles: An Adaptive Approach
by David Rolf Larsen
Chairperson of Supervisory Committee
Professor Chadwick D. Oliver
College of Forest Resources
A silvicultural systems can be as a progression of stand structures through which
a given stand moves through it's life. Silviculture practiced with this view requires
a knowledge of the potential stand structures that a given stand can obtain and
understand the treatment required to move through the desired set of stand structures.
This view of silviculture di�ers from that of traditional silviculture in expectation and
application. Traditional silviculture focuses on the operations that forester do in stand
whereas this approach focuses on the change in stand structures. Operations simple
become a tool to implement the desired stand structure change. Tools needed to
practice silviculture in this manor are presented and developed.

ACKNOWLEDGMENTS
The author wishes to express sincere appreciation to D. Chadwick D. Oliver for
his patent guidance and friendship. I also wish to thank my son Carl V. Larsen
for his tolerance during the years of graduate school and foregone activities. I
would like to also thank my fellow students for there friendship and support.
I would especially like to thank Dean Berg, J. Ren�ee Brooks, Jo~ao Batista,
Ann Camp, Glen Galloway, John Kershaw, Jim McCarter, Mark Petruncio,
Gerardo Segura, Lhakpa Sherpa and Wieger Schaap.
vi

Chapter 1
INTRODUCTION
\It is just as true for the sowing of wild trees as for Xylotrophia, their
planting, transplanting, pruning, removal of sprouts and other forms of
care and treatment, are not products of our minds, but unquestionably
those of our forefathers in `ancient' times. These ideas are plain to see in
their writings. They have been known and applied ever since the beginning
of time. Indeed we willingly acknowledge that this science has now been
developed to a considerably higher level, and both understood and imple-
mented to a fuller and more certain extent than ever before."
Hannss Carl von Carlowitz 1713. Sylvicultura Oeconomica p. 254. (The
�rst European book written entirely on forestry)
\The whole of science is nothing more than a re�nement of everyday think-
ing. It is for this reason that the critical thinking of the physicist cannot
possibly be restricted to the examination of concepts of this own �eld. He
cannot proceed without considering critically a much more di�cult prob-
lem, the problem of analyzing the nature of everyday thinking."
A. Einstien 1936.
The following dissertation proposes that theories of forest stand dynamics can be
used to plan silvicultural systems. Silvicultural systems are described as the change
in stand structure through the life of a stand. Along with this description, a tool is

2
presented to aid in visualization and understanding of the processes of stand develop-
ment and dynamics. The proposed approach does not change traditional silviculture,
but provides a exible way of thinking about silviculture for both traditional and
non-traditional objectives.
1.1 Silvicultural Practice
Silviculture is the practice of manipulating stands to change the existing stand struc-
ture to a more \desirable" one. While having this generalized goal, most silviculturists
focus on the operations to manipulate stands rather than on the development of stand
structure. These operations have been repeated and re�ned so that silviculture has
been good at doing operations that implement common silvicultural systems to pro-
duce stands that meet traditional objectives. The silvicultural systems tend to be
named for regeneration methods, since these methods have one of the largest impacts
on the resultant stand structure (Troup, 1952; Matthews, 1989).
Alternatively, silvicultural systems can be described as changes in stand structure
through which a stand can reach a desired structure. Stand structure in this context
is much more than simple tree size distributions. It includes the size distributions,
spatial distributions, species, densities, and history of the stand.
A silvicultural system can be considered the sequence of structures through which
a given stand must progress to achieve a stand of the desired character at the desired
time. The practice of silviculture then becomes de�ning and achieving the sequence
of forest structures that will attain the goal with the least intervention and in an ac-
ceptable time. Interventions used to implement the silviculture system are operations
such as regeneration, thinning, pruning, fertilization, and others. These operations
are viewed as tools to implement silvicultural systems, not the focus of silviculture.
For this approach to silviculture to be e�ective, forest managers must have a good
understanding of how manipulations to a tree's environment will change the way it

3
will respond.
Historically, people have assumed that a stand continuously recycles through var-
ious structures found in the previous stands on the same area (Roth, 1925). The
assumption is that, if left alone, a stand on a given area will attain the same set of
structures in the future, which it had in the past.
These assumptions are not valid in most cases. Stand structures are the product
of the speci�c site and the sequence of events (Oliver and Larson, 1990). A given
stand has a range of possible structures that can be achieved, and the one realized
depends on the speci�c sequence of events that occur. Sequential stands on a given
area often develop to quite di�erent structures. Stands will only repeat the same
structures held by a previous stand if the same sequence of events are experienced.
Alternatively, stands may achieve common structures by di�erent sequences; however,
the structures before and after the common structure may quite di�erent.
Additionally, people often assume that the products desired from the forest will
be the same in the future as they are now. This assumption, again may not be true.
Some current products will continue in demand, but new resources are continually
being found and used (Perlin, 1991).
Silviculture systems have often been designed to accomplish the objective of wood
production. If changing objectives and a changing balance among objectives are ex-
pected in the future of forest management, then a more exible type of silviculture
is needed. This exible silviculture requires a thorough knowledge of the processes of
stand dynamics|a knowledge that can be learned by years of experience or through
tools that allow the application of the principles of stand dynamics to speci�c sit-
uations. This approach expands both the ability to deal with traditional and non-
traditional problems and the context of what are feasible solutions.
Forestry in the United States has developed under a resource rich environment.
Foresters and the public are increasingly critical of management decisions with respect
to both timber and other resources produced. These changing conditions have forced

4
the consideration of new objectives in forest management. Forest managers are seeing
objectives change faster than research into how to achieve objectives.
A exible silvicultural tool is needed to explore the consequences of innovative
silvicultural prescriptions on future stand structures. Such a tool could be created
by combining general theories on stand and tree dynamics with a monitoring system
and a growth model. The proposed approach requires a broad view of growth models
and their use. Most current models are based on regional average trends, and these
models are designed to predict regional averages. They are also based on historical
conditions, and implicit to their predictions is the assumption that those conditions
will continue. Furthermore, they implicitly assume that treatments sampled are the
treatments that will be applied. The approach to stand projection proposed in this
paper is to predict the trends of individual stands or groups of stands based on the
theories of tree growth and stand development and the past growth of the subject
stand. This approach may provide less precise growth estimates, but will provide the
exibility to explore various alternative treatments. Figure 1.1 is a Venn diagram of
the relationship between adaptive models of stand dynamics and growth and yield
models.
1.2 Adaptive silviculture
Traditionally, research silviculturists develop a collection of \standard" prescriptions
for the types of stands usually desired. These \standard" prescriptions are based on
treatment experiments. Given these experiments, a regimented management schedule
is developed. These silvicultural precripttions are taught by analogy. Silviculturists
need only determine if the stand to be managed will respond in the same manner as
the experimental stand, and if so, apply the operational schedule. These \standard"
prescriptions were developed in a time when there were few trained silviculturists
and there were a lot of unskilled labor. The prescriptions needed to be simple to

5
All possible ways to model forests
Adaptive Stand Dynamics ModelsGrowth and Yield Models
Figure 1.1: The relationship between adaptive stand dynamics models and growthyield models. Much of the approach is similar; however, there are di�erences inapproach, objective and results, as described in this paper.

6
understand and implement.
Silvicultural systems have been described as the method of regenerating, tend-
ing, and harvesting stands. Silviculturists group systems by regeneration method:
clear-cutting, shelterwood cutting, successive regeneration cuttings, selection cutting
coppice cutting, and others (Troup, 1952; Matthews, 1989).
Forestry schools have produced many well trained silviculturists in the twentieth
century, creating an opportunity to increase exibility in silvicultural decisions. Silvi-
culturists must be knowledgable in stand dynamics for a more exible approach to be
successful. Knowledge of stand dynamics can be obtained either through experience
or by careful study of stand histories (Oliver, 1978). Fortunately, several relationships
of stands and trees can be generalized to allow the description of the process of stand
development (Oliver and Larson, 1990). The use of these generalization allow a way to
anticipate the response of a stand to the current condition or any anticipated changed
condition. These generalizations can be applied without a computer by individuals
well versed in the subtleties of the theories. Alternatively, these generalizations can
be computerized in a model which include the subtleties.
Silviculturists desire prediction of the relative growth of a stand, correctly re-
ecting the biological relationships among the trees. A model should incorporate
the species mix found within a stand and provide predictions of the structure of a
stand, after it is treated in ways very di�erent than any stands found in the existing
landscape. This type of model can be called an \adaptive stand dynamics model"
(Larsen, 1991). These models, while predicting the growth of trees within a stand the
same as forest growth models, di�er in objective, design, and assumptions. \Adaptive
stand dynamics models" do not replace traditional growth and yield models but are a
complementary tool. \Adaptive stand dynamics models" explore the forest structure
change given the current conditions and any change to those conditions. This model
is focused not on volume production but on change in the gross dimensions of the
trees (e.g. height, spatial pattern, crown size) and how these dimensions are related

7
to change in various parts of the trees.
The use of stand dynamics to describe silvicultural systems provides an alter-
native way of viewing silviculture practices. It does so by shifting from a product
and treatment (anthropocentric) approach to a stand structure development (arbor-
centric) approach. Relatively few relationships are needed to represent the major
elements of stand dynamics theory adequately. In addition, reasonable estimate of
silvicultural systems can be made for creating stand structures that do not currently
exist on the landscape. Further, these relatively few principles have been reported in
widely varying forest types in many di�erent parts of the world (Oliver, 1992).
By describing the individual parts of traditional silviculture in terms of their
e�ect on stand structures, di�erences between the proposed approach and traditional
silviculture become apparent. The major methods of changing a forest are harvesting
(a silvicultural operation), regeneration, thinning, pruning, and fertilization.
Harvesting and regeneration are the most drastic changes to a forest, since the
old forest is removed and a new forest is started. Regeneration is the manipulation
in which the base spatial pattern is de�ned and the future alternative relative size of
the trees is determined. The base spatial pattern is de�ned because the locations of
trees are de�ned and those locations can not be moved throughout their lives. Future
relative sizes of the trees are also de�ned by the timing of their establishment. For
example, a regeneration process that establishes trees on a uniform spacing at one
time will produce a stand of very uniform trees, whereas a regeneration process that
varies the spacing and the times of establishment will produce a stand of very di�erent
tree sizes.
Thinnings are the second most widely used treatment and are a means of modifying
the environment of residual trees. As stated above, the options are not as wide as
in regeneration because trees can only be removed; the base spatial pattern has been
set by the regeneration process. Additionally, it is very important to consider the
tree's past history, when thinning around a given tree. This history is expressed in

8
the tree's present crown size and any deformations caused by damaging agents. These
damages are a detriment in wood production, but they may be an asset, if ones goal is
to manage for cavity nesting birds. When considering thinning one should determine
the amount of growing space being made available to the residual trees, how quickly
will their crowns be able to utilize that released space, and how long the trees will have
additional space available. These determinations have two important consequences.
If the crowns take a very long time to utilize the released space because of previous
conditions, the thinning may not provide the bene�t required (Siemon et al., 1980;
Oliver and Murray, 1983; Oliver et al., 1986; O'Hara, 1989).
Pruning is a treatment that has been utilized to a lesser extent in silviculture.
Prunings are the direct manipulation of a given tree's crown. E�ective pruning does
not kill the tree or have an a�ect on the live crown. E�ective pruning is again a
balance between the amount of the subject tree's crown removed and the amount of
foliage on the competitors crowns. Thinnings usually accompany pruning treatments
(Lethpere, 1957; Mar:M�oller, 1960; Staebler, 1964) because the pruning treatments
are much less e�ective with out thinning.
Fertilizers also a�ect the crowns of trees by increasing the amount of foliage, the
density of foliage per unit area, and the production per unit of the foliage. Fertilization
has the e�ect of increasing tree growth for the period that the trees can maintain the
increased nutrients within the tree (Brix, 1983; Vose and Allen, 1988; Vose, 1988).
These traditional silvicultural treatments become tools to change the tree or the
environment around trees. The traditional approach to silviculture focuses on the
treatment of the present structure. The focus should be on the present structure,
the future structure, and the conditions needed to change the present stand into a
desired future stand structure. In a real sense, silviculture is about choices: �rst,
which features of the present stand should be favored so stand development will
produce a future stand of the desired structure. The other choice is that of time.
A treatment can sometimes reduce the time a stand takes to reach a given future

9
structure.
1.3 Stand Structure
Silviculture is the management of a complex of spatially arrayed trees and their various
parts that can be manipulated. Stand structure is the three-dimensional arrangement
of the various tree components (e.g. stems, foliage, etc.) of a forest stand. Character
means something more general: the range of structures that are neighbors in the
distribution of possible structures. Some characters are named, such as \Old Growth"
character. Many potential characters have no name but can still be used to describe
a range of stand structures. Because the above de�nition of structure forms multiple,
related continua, it is not possible to categorize the structures logically. A method
of describing a modal structure for a target stand may be to describe a distribution
for each of the components of the description. This structure de�nition is easier to
explain in terms of a set of model parameters used to generate the range of stands of
the target character.
The elements used to describe structure should include species, density or inten-
sity (number per unit space), pattern both by species and by all trees (on a scale from
uniform, to random, to clumped), size (distributions of height, diameters, and vol-
umes), and some indication of stand history. This sort of description is quite involved
but is important to de�ne exactly what is meant by a given structure with common
reference. Few people can visualize a forest structure from these measures directly;
however, a graphical display system can be used to visualize a given stand structure
exactly.
A quanti�ed and graphically displayed stand structure can provide a common
language for resource managers from di�erent �elds to describe the attributes they
require from the stands that silviculturists are managing. Much of the confusion
of what is required for di�erent resource objectives can be greatly reduced if stand

10
structures are used in this way.
Forest character is related to the human perception of forest structures. When
a human, especially a trained forester, views a forest, he observes many components
that de�ne the arrangement of trees. Character relates to the human ability to di�er-
entiate between various structures. Many unique stand structures may not be distin-
guishable by human perception. Classi�cation of structures into character types for
communication purposes could be useful, but the boundaries are not distinct. The
spectrum of stand structures is continuous and there are many stand structures that
are half way between one character and another. A method to deal with the problem
of indistinct character de�nitions is to de�ne character as a distribution of target
structures. Distributions are hard for many people to visualize; so again, the idea of
a viewer that displays the target structure is suggested, and the range of structures
is de�ned through simulation. Humans have the ability to resolve complicated 3-D
images and distill relationships from these images and di�erences between them.
1.4 Adaptive approach to silviculture
Adaptive silviculture is the description of stand structures combined with the knowl-
edge of stand development, thus providing a exible, adaptable approach to silvicul-
ture. This view of forest management is based on the ideas of adaptive management.
These ideas were �rst stated in natural resource management by Carl Walters (1986)
and in forestry by Gordon Baskerville (1985) and further expanded by Chadwick
Oliver and the author (Larsen, 1991; Oliver, 1992).
Managing forests with adaptive silviculture requires that each stand treatment be
considered as a experiment. The stand is measured periodically before and after treat-
ment to determine if the treatment has accomplished the desired (\hypothesized")
e�ect. If the desired e�ect is not obtained, the stand's management is \adapted" to
use a di�erent treatment or to a di�erent expectation. The \adapted" management

11
may vary from accepting the unexpected stand behavior, to supplemental treatments
to accomplish the desired stand behavior, to modifying the stand dynamics model to
re ect the observed stand behavior. This process is repeated throughout the life of
the stand.
Silviculture in the adaptive management framework is the manipulation of stand
development patterns to produce desired stand structures. Alternatively, silvicultural
systems can be viewed as the manipulation of a stand structure in space and time
through the life of a stand. Silviculturists practicing this adaptive silviculture require
a thorough knowledge of stand dynamics. Adaptive models of stand dynamics can be
a tool to assist silviculturists.
The Venn diagram in Figure 2.1 illustrates the relationship between adaptive
silviculture and traditional silviculture. Traditional silviculture is a subset of the
possible ways of forests developing. Adaptive silviculture includes all of traditional
and some non-traditional silviculture. In the diagram, non-traditional silviculture
extends outside adaptive management because there may be possible ways for a forest
to develop that are not accounted from by adaptive silviculture.
1.5 Adaptive growth models
Stands grow by relatively few principles. Stand growth is the aggregation of the
growth of each individual tree in the stand. The location and size of each tree a�ects
the environment of each neighbor tree. By projecting the development of each tree
while accounting for the in uence of each neighboring tree, the stand growth can be
determined as the summation of change of each tree's attribute of interest. Through
aggregation of the attributes that describe the components of stand structure, the
various structural characteristics can be described.
Di�erences between stands include spacing, time of initiation, and the characteris-
tics of individual trees. Characteristics of individual trees are a�ected by site quality,

12
All possible ways of forest development
Adaptive Silviculture
Traditional Silviculture
Non-Traditional Silviculture
Figure 1.2: Venn diagram of the relationship of adaptive silviculture to traditional sil-viculture. Adaptive silviculture includes all of traditional silviculture. Non-traditionalsilviculture extends outside adaptive silviculture because adaptive silviculture maynot account for all possible ways that a forest develops.

13
which a�ects height growth rate, canopy thickness, and species. Species a�ect the
height growth rates, response to disturbances (including the regeneration mechanism),
and tree form (height growth rate, crown length, crown width, and shade tolerance).
With these principles, stand structure can be projected by knowing the spatial
arrangement of trees, site characteristics, and characteristics of the species. The same
model forms are used rather than developing a new model forms for each species and
site. The site and growth forms are adjusted to the particular location. The model's
predictions will be more accurate when there is accurate and complete user collected
information.
In appreciation of silviculturists who often work with little information, this type
of model can project stand growth in new areas with relatively little information and
many assumptions. As the stand grows, the actual stand structure can be compared
to the projected structures; and this information can be added to the information
base for the particular location. In this way, the model constantly is improving in an
iterative or adaptive manner.
Silviculturists can project stand structures for new areas or new stand structures
without waiting for long term permanent plot. This modeling approach may not
produce as precise estimates of volume as modeling approaches designed to produce
precise volume estimates, but it will allow a relative estimate of a treatment a�ect
for the decisions that must be made immediately.
Adaptive models of stand dynamics are designed to make maximum use of diverse
information. Under the proposed scenario, a user would have information and data
from various sources such as published equations, stem analysis from a stand, stand
measurements, repeat measurement from the same stand, and/or growth equations
from forest-wide inventories. The models should make maximum use of whatever
information or data is available with in the framework proposed for the model.

14
1.6 Scope of the dissertation
This dissertation will present a conceptual framework within which to view silvicul-
ture. The design of a tool is presented to visualize and explore the consequences
of treatments or disturbances on a speci�c stand. One example of such a tool will
be presented as well as the application of that tool to three speci�c stands. The
strengths and weaknesses of the current example are discussed. The consequences
of them on management decisions will also be presented along with some ideas for
potential future work.
This study uses a theoretical approach as opposed to the experimental approach
to science. A theoretical approach explores the consequences of a given set of assump-
tions, not the truth of the assumptions. If the results of the theory do not agree with
observation then the assumptions must be questioned. This deductive approach is the
generalization of relationships after years of observation and experimentation. This
approach is di�erent from an experimental approach which manipulates part of the
real system and records the consequences of those manipulations. In this inductive
approach, research experiments are perform concerning a given question, granted,
many of the ideas come form deduction of published literature. After a number of
experiments, the general results of the experiments are then summarized. The two
approaches ask di�erent questions and provide di�erent answers.

Chapter 2
ADAPTIVE SILVICULTURE
\The whole of science is nothing more than a re�nement of everyday think-
ing. It is for this reason that the critical thinking of the physicist cannot
possibly be restricted to the examination of concepts of this own �eld. He
cannot proceed without considering critically a much more di�cult prob-
lem, the problem of analyzing the nature of everyday thinking."
A. Einstien 1936.
The use of stand dynamics to describe silvicultural systems provides an alter-
native way of viewing silviculture practices. It does so by shifting from a product
and treatment (anthropocentric) approach to a stand structure development (arbor-
centric) approach. Relatively few relationships are needed to represent the major
elements of stand dynamics theory adequately. In addition, reasonable estimate of
silvicultural systems can be made for creating stand structures that do not currently
exist on the landscape. Further, these relatively few principles have been reported in
widely varying forest types in many di�erent parts of the world (Oliver, 1992).
By describing the individual parts of traditional silviculture in terms of their
e�ect on stand structures, di�erences between the proposed approach and traditional
silviculture become apparent. The major methods of changing a forest are harvesting
(a silvicultural operation), regeneration, thinning, pruning, and fertilization.
Harvesting and regeneration are the most drastic changes to a forest, since the
old forest is removed and a new forest is started. Regeneration is the manipulation
in which the base spatial pattern is de�ned and the future alternative relative size of

16
the trees is determined. The base spatial pattern is de�ned because the locations of
trees are de�ned and those locations can not be moved throughout their lives. Future
relative sizes of the trees are also de�ned by the timing of their establishment. For
example, a regeneration process that establishes trees on a uniform spacing at one
time will produce a stand of very uniform trees, whereas a regeneration process that
varies the spacing and the times of establishment will produce a stand of very di�erent
tree sizes.
Thinnings are the second most widely used treatment and are a means of modifying
the environment of residual trees. As stated above, the options are not as wide as
in regeneration because trees can only be removed; the base spatial pattern has been
set by the regeneration process. Additionally, it is very important to consider the
tree's past history, when thinning around a given tree. This history is expressed in
the tree's present crown size and any deformations caused by damaging agents. These
damages are a detriment in wood production, but they may be an asset, if ones goal is
to manage for cavity nesting birds. When considering thinning one should determine
the amount of growing space being made available to the residual trees, how quickly
will their crowns be able to utilize that released space, and how long the trees will have
additional space available. These determinations have two important consequences.
If the crowns take a very long time to utilize the released space because of previous
conditions, the thinning may not provide the bene�t required (Siemon et al., 1980;
Oliver and Murray, 1983; Oliver et al., 1986; O'Hara, 1989).
Pruning is a treatment that has been utilized to a lesser extent in silviculture.
Prunings are the direct manipulation of a given tree's crown. E�ective pruning does
not kill the tree or have an a�ect on the live crown. E�ective pruning is again a
balance between the amount of the subject tree's crown removed and the amount of
foliage on the competitors crowns. Thinnings usually accompany pruning treatments
(Lethpere, 1957; Mar:M�oller, 1960; Staebler, 1964) because the pruning treatments
are much less e�ective with out thinning.

17
Fertilizers also a�ect the crowns of trees by increasing the amount of foliage, the
density of foliage per unit area, and the production per unit of the foliage. Fertilization
has the e�ect of increasing tree growth for the period that the trees can maintain the
increased nutrients within the tree (Brix, 1983; Vose and Allen, 1988; Vose, 1988).
These traditional silvicultural treatments become tools to change the tree or the
environment around trees. The traditional approach to silviculture focuses on the
treatment of the present structure. The focus should be on the present structure,
the future structure, and the conditions needed to change the present stand into a
desired future stand structure. In a real sense, silviculture is about choices: �rst,
which features of the present stand should be favored so stand development will
produce a future stand of the desired structure. The other choice is that of time.
A treatment can sometimes reduce the time a stand takes to reach a given future
structure.
2.1 Stand Structure
Silviculture is the management of a complex of spatially arrayed trees and their various
parts that can be manipulated. Stand structure is the three-dimensional arrangement
of the various tree components (e.g. stems, foliage, etc.) of a forest stand. Character
means something more general: the range of structures that are neighbors in the
distribution of possible structures. Some characters are named, such as \Old Growth"
character. Many potential characters have no name but can still be used to describe
a range of stand structures. Because the above de�nition of structure forms multiple,
related continua, it is not possible to categorize the structures logically. A method
of describing a modal structure for a target stand may be to describe a distribution
for each of the components of the description. This structure de�nition is easier to
explain in terms of a set of model parameters used to generate the range of stands of
the target character.

18
The elements used to describe structure should include species, density or inten-
sity (number per unit space), pattern both by species and by all trees (on a scale from
uniform, to random, to clumped), size (distributions of height, diameters, and vol-
umes), and some indication of stand history. This sort of description is quite involved
but is important to de�ne exactly what is meant by a given structure with common
reference. Few people can visualize a forest structure from these measures directly;
however, a graphical display system can be used to visualize a given stand structure
exactly.
A quanti�ed and graphically displayed stand structure can provide a common
language for resource managers from di�erent �elds to describe the attributes they
require from the stands that silviculturists are managing. Much of the confusion
of what is required for di�erent resource objectives can be greatly reduced if stand
structures are used in this way.
Forest character is related to the human perception of forest structures. When
a human, especially a trained forester, views a forest, he observes many components
that de�ne the arrangement of trees. Character relates to the human ability to di�er-
entiate between various structures. Many unique stand structures may not be distin-
guishable by human perception. Classi�cation of structures into character types for
communication purposes could be useful, but the boundaries are not distinct. The
spectrum of stand structures is continuous and there are many stand structures that
are half way between one character and another. A method to deal with the problem
of indistinct character de�nitions is to de�ne character as a distribution of target
structures. Distributions are hard for many people to visualize; so again, the idea of
a viewer that displays the target structure is suggested, and the range of structures
is de�ned through simulation. Humans have the ability to resolve complicated 3-D
images and distill relationships from these images and di�erences between them.

19
2.2 Adaptive approach to silviculture
Adaptive silviculture is the description of stand structures combined with the knowl-
edge of stand development, thus providing a exible, adaptable approach to silvicul-
ture. This view of forest management is based on the ideas of adaptive management.
These ideas were �rst stated in natural resource management by Carl Walters (1986)
and in forestry by Gordon Baskerville (1985) and further expanded by Chadwick
Oliver and the author (Larsen, 1991; Oliver, 1992).
Managing forests with adaptive silviculture requires that each stand treatment be
considered as a experiment. The stand is measured periodically before and after treat-
ment to determine if the treatment has accomplished the desired (\hypothesized")
e�ect. If the desired e�ect is not obtained, the stand's management is \adapted" to
use a di�erent treatment or to a di�erent expectation. The \adapted" management
may vary from accepting the unexpected stand behavior, to supplemental treatments
to accomplish the desired stand behavior, to modifying the stand dynamics model to
re ect the observed stand behavior. This process is repeated throughout the life of
the stand.
Silviculture in the adaptive management framework is the manipulation of stand
development patterns to produce desired stand structures. Alternatively, silvicultural
systems can be viewed as the manipulation of a stand structure in space and time
through the life of a stand. Silviculturists practicing this adaptive silviculture require
a thorough knowledge of stand dynamics. Adaptive models of stand dynamics can be
a tool to assist silviculturists.
The Venn diagram in Figure 2.1 illustrates the relationship between adaptive
silviculture and traditional silviculture. Traditional silviculture is a subset of the
possible ways of forests developing. Adaptive silviculture includes all of traditional
and some non-traditional silviculture. In the diagram, non-traditional silviculture
extends outside adaptive management because there may be possible ways for a forest

20
All possible ways of forest development
Adaptive Silviculture
Traditional Silviculture
Non-Traditional Silviculture
Figure 2.1: Venn diagram of the relationship of adaptive silviculture to traditional sil-viculture. Adaptive silviculture includes all of traditional silviculture. Non-traditionalsilviculture extends outside adaptive silviculture because adaptive silviculture maynot account for all possible ways that a forest develops.
to develop that are not accounted from by adaptive silviculture.
2.3 Adaptive growth models
Stands grow by relatively few principles. Stand growth is the aggregation of the
growth of each individual tree in the stand. The location and size of each tree a�ects
the environment of each neighbor tree. By projecting the development of each tree
while accounting for the in uence of each neighboring tree, the stand growth can be
determined as the summation of change of each tree's attribute of interest. Through
aggregation of the attributes that describe the components of stand structure, the

21
various structural characteristics can be described.
Di�erences between stands include spacing, time of initiation, and the characteris-
tics of individual trees. Characteristics of individual trees are a�ected by site quality,
which a�ects height growth rate, canopy thickness, and species. Species a�ect the
height growth rates, response to disturbances (including the regeneration mechanism),
and tree form (height growth rate, crown length, crown width, and shade tolerance).
With these principles, stand structure can be projected by knowing the spatial
arrangement of trees, site characteristics, and characteristics of the species. The same
model forms are used rather than developing a new model forms for each species and
site. The site and growth forms are adjusted to the particular location. The model's
predictions will be more accurate when there is accurate and complete user collected
information.
In appreciation of silviculturists who often work with little information, this type
of model can project stand growth in new areas with relatively little information and
many assumptions. As the stand grows, the actual stand structure can be compared
to the projected structures; and this information can be added to the information
base for the particular location. In this way, the model constantly is improving in an
iterative or adaptive manner.
Silviculturists can project stand structures for new areas or new stand structures
without waiting for long term permanent plot. This modeling approach may not
produce as precise estimates of volume as modeling approaches designed to produce
precise volume estimates, but it will allow a relative estimate of a treatment a�ect
for the decisions that must be made immediately.
Adaptive models of stand dynamics are designed to make maximum use of diverse
information. Under the proposed scenario, a user would have information and data
from various sources such as published equations, stem analysis from a stand, stand
measurements, repeat measurement from the same stand, and/or growth equations
from forest-wide inventories. The models should make maximum use of whatever

22
information or data is available with in the framework proposed for the model.

Chapter 3
THEORY
\Mathematics is a useful vehicle for expression and manipulation; but the
heart of the theory is elsewhere."
Sir Arthur Eddington (Physics Professor at Cambridge, consider a author-
ity on general relativity)
This chapter will review three topics that relate to the development of the quan-
titative description of forest stand dynamics and the use of that description in a
conceptual model. First, a discussion is presented of a quantitative method for de-
scribing stand structure. Second, diagnostic criteria are discussed that are useful
tools for evaluating current and future stand conditions and growth model inputs and
outputs. Third, three related growth models are described that provided ideas used
in the current approach.
3.1 Ways to describe forest structure
There are several components to forest stand structure. These include size distri-
bution, spatial distribution, density or intensity, species, and stand history. Each
structure descriptor will be discussed in detail.
3.1.1 Size and size distribution
Size (e.g. diameter at breast height, height, crown length, crown width, and foliage
area) and size distribution are the most common methods for describing the structure

24
Relative Rank
Per
cent
Hei
ght
0.0 0.2 0.4 0.6 0.8 1.0
0.0
0.2
0.4
0.6
0.8
1.0
Helena Plots - Height ordered by height
Figure 3.1: A standardized rank order plot of a size distribution. This can be a usefulway to compare distributions.
of forest stands (Knox et al., 1989; Ford, 1975). Typically the data are presented
as a frequency histogram. This provides visual presentation of the distribution that
allows the the investigator to grasp shape, skewness, and kurtosis quickly; however,
the presentation may be confusing if histograms are not at the same scale.
There are a number of methods to measure this inequality. Benjamin and Hard-
wick (1986) list four measures; Coe�cient of variation, Gini coe�cient, skewness
and kurtosis. The coe�cient of variation is a dimensionless ratio to compare relative
variability of di�erent populations. It disregards asymmetry. Coe�cient of variation
V is calculated by equation 3.1.
V =s
�x100; (3.1)
where s is the standard deviation and �x is the mean.
The size distribution can also be plotted as the rank order of the population versus

25
the size of each tree. This plot produces a shape similar to an inverse cumulative curve.
With this type of curve it is very easy detect subtle di�erence in the change of the size
distribution. Another method of displaying size structure is the standardized rank
order plot (Figure 3.1).
Another type of plot presents the cumulative proportion of the population (relative
rank) versus the cumulative proportion of size (relative size, the size of the current
individual divided by the largest individual). This approach removes the actual size
from the plot and presents the relationship between the sizes in the distribution
(Figure 3.2). All sizes in the stand have equality If a distribution plots as a 45o line
(i.e. an increase of one unit in rank is equal to one unit in relative size.) A distribution
which plots above or below this line has inequality and is called a Lorenz curve.
The Gini coe�cient (Dixon et al., 1987; Weiner, 1984; Weiner and Solbrig, 1984) is
best described in terms of the above-mentioned standardized rank order plot. On this
plot the curve of the size distribution is called the Lorenz curve. The Gini coe�cient
was �rst used by economists to describe the inequality in the distribution of wealth in
societies. The Gini coe�cient G describes the area between the line of equality and
the Lorenz curve (Figure 3.2)
G =
nXi=1
nXj=1
j xi � xj j
2n2�x; (3.2)
where n is the number of individuals in the sample, i and j are indices that extend
from 1 to n (i.e. i = 1; 2; 3 : : : n and j = 1; 2; 3 : : : n ), x is the vector of observations
and �x is the mean of those observations.
The Gini coe�cient provides a good measure of the amount of inequality but
no information about the shape of the inequalities. Skewness is a statistic of the
asymmetry of a distribution and kurtosis is a statistic of shape and are described in
terms of the �rst four moments about the mean for a population. In general, the rth
moment mr about the mean �x is

26
Size
Cumulative Proportion of Population
ofProportionCumulative Line of Absoulte Equality
Lorenz Curve
Figure 3.2: The area between the Lorenz curve and the 45o line is the area de�nedas the Gini coe�cient of inequality.
Coefficient of variation = 0.5345225 Coefficient of skewness = 3.741657 Coefficient of excess = -0.01041667
Coefficient of variation = 0.3333333 Coefficient of skewness = 0
Coefficient of excess = 0.3068182
Coefficient of variation = 0.2672612 Coefficient of skewness = -3.741657
Coefficient of excess = 1.333333
Figure 3.3. An illustration of Skewness for three distributions

27
Coefficient of variation = 0.4472136 Coefficient of skewness = 0
Coefficient of excess = 0.5357143
Coefficient of variation = 0.3333333 Coefficient of skewness = 0
Coefficient of excess = 0.3068182
Coefficient of variation = 0.2773501 Coefficient of skewness = 0
Coefficient of excess = 0.2166667
Figure 3.4. An illustration of kurtosis for three distributions
mr =1
n
nXi=1
(xi � �x)r; (3.3)
where n is the number of observations x is a vector of observations and r is a integer
value (i.e. 2; 3; 4 ).
The coe�cient of skewness for a population g1 is de�ned as
g1 =m3
m23=2
; (3.4)
where mr is de�ned in equations 3.3.
Figure 3.3 presents several distributions and their related coe�cient of skewness.
The coe�cient of kurtosis for a population g2 is de�ned as
g2 =m4
m22; (3.5)
This term is centered on the value 3; therefore, for ease of interpretation the
coe�cient of excess is de�ned as g2 � 3.

28
* * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * ** * * * * * * * * * * * * * *
Uniform
*
*
*
*
*
*
*
**
*
*
* *
*
**
*
*
*
**
**
*
*
*
*
*
*
*
**
***
*
**
*
*
*
*
*
*
*
**
**
*
*
*
*
** *
*
**
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
**
*
* *
* *
* **
** *
*
*
*
**
*
*
**
*
**
*
*
**
*
*
* *
**
*
*
*
**
*
*
*
**
*
*
**
*
**
*
*
*
* **
*
*
**
*
*
*
**
*
**
**
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
***
* **
*
*
*
*
*
**
*
**
***
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
* *
*
**
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
Random
**
***
** **
****
****
****
****
****
****
********
****
****
****
****
****
****
****
***
********
**
****
********
****
****
****
****
****
****
****
****
********
* ***
*** *
****
***
****
*
****
****
** **
****
*** *
****
****
****
****
**
****
****
****** **
****
****
********
** **
****
****
Clustered
Figure 3.5: a. A \uniform" pattern, b. a \random" or Poisson pattern, and c. a\clustered" pattern
Figure 3.4 presents several distributions and their related coe�cient of excess.
Each of these measures describes a di�erent part of the distribution. For visualization
of a distribution, the histogram or order plot is most useful. The other measures
provide a quantitative measure of several di�erent aspects of the distribution.
3.1.2 Density
One of the most common measures of stand structure is \density", by foresters or
\intensity", by statisticians. This measure is de�ned as the number of individuals per
unit area. This measure of stand structure seems very simple compared to the other
measures, but it is very important to assess the other measures properly. One of the
more interesting ways to describe forest structure is through size/density relationships
(See Diagnostic criteria in section 3.2).

29
Table 3.1: Table of values for spatial patterns for the three example plot shown abovein Figure 3.5.
Statistic Uniform Random Clustered
Number 229 250 236
Skellam-Moore 175.01 85.48 18.59
Clark & Evans 1.74 1.01 0.41
Brown 0.99 0.51 0.34
Pielou 0.63 1.09 2.81
Hopkins' F 0.26 1.02 11.36
Hopkins' N 0.19 0.53 0.85
Holgate F 0.08 0.11 0.30
Holgate N 0.37 0.39 0.62
3.1.3 Spatial pattern
The spatial pattern in a forest stand has been viewed as a two-dimensional point
process. Given a pattern of points in two-dimensions, one question that statisticians
have asked is whether the pattern is randomly distributed or not. Point patterns
can assume a range of patterns from \uniform", to \random", to \clustered" (Ripley,
1981; Diggle, 1983) (Figure 3.5.). If a pattern represents tree locations in a forest,
a \random" pattern is called a Poisson forest, as the points are Poisson distributed.
Many tests for nonrandomness have been developed for just such questions. In most
of the following tests, they determine if the pattern is Poisson distributed. First, a
number of �rst order tests will be examined. Table 3.1 lists the values for the above
tests for the three patterns in Figure 3.5. The speci�c of how to calculate each index
can be found in Appendix A.
� Skellam-Moore

30
Both Skellam (1952) and Moore (1954) developed an approach to look at mapped
point patterns at a time when average quadrate samples were common. Their
main question is the validity of randomness of pattern assumption for the
quadrate samples? Skellam-Moore is a test that for the Poisson assumption
is �22m. It was noted by Pielou (1959) noted that estimates of intensity (n=A)
are poor using distance methods and that independent estimate of intensity
from quadrates should be used .
� Clark and Evans
Clark and Evans (1954) were attempting to develop a general index of spatial
pattern. They noted that the assumption of random spatial plant distribution
is not valid of observed plant patterns. Their method was a test of randomness
for populations of known density and they state the measure uses only the
nearest neighbor, it ignores the majority of the spatial relationships to other
plants. With the index of aggregation, if the spatial pattern is random, R = 1;
clustered, R! 0; and uniform, R! some arbitrary upper limit.
� Brown
The Brown index of aggregation is the geometric mean of squared distances over
the arithmetic mean and is independent of area. The index G ranges from 0 to
1 with G! 0 for \clustered" pattern and G! 1 for \uniform" pattern (Brown
and Rothery, 1978).
� Hopkins'
The Hopkins' index of aggregation is the ratio of the distance from a randomly
chosen point to its nearest neighbor tree and the distance from that nearest
neighbor tree to its nearest neighbor.

31
HopF =
mXi=1
d2(p�ti)i
mXi=1
d2(t�ti)i
; (3.6)
where d(p�ti)i is the distance from a random point to it's nearest neighbor in-
dividual i and d(t�ti)i is the distance from individual i to it's nearest neighbor
individual. This test has an F distribution with F (2m; 2m) (Hopkins, 1954).
Byth and Ripley (1980) presented a standardized index based on this test as:
HopN =1
m
mXi=1
24 d2(p�ti)i(d2(p�ti)i + d2(t�ti)i)
35 ; (3.7)
This index has a Normal null distribution, N(12; 112m). As HopN ! 0, it indi-
cates a more \uniform" pattern; and as HopN ! 1, it indicates a more \clus-
tered" pattern.
� Holgate
The Holgate index is based on the ratio of the distance from a random point
to its nearest neighbor and the distance from the same random point to the
next nearest neighbor (Holgate, 1964). The Holgate indices have the same null
distributions as the Hopkins' indices.
� Pielou
Pielou (1954) noted the value of the di�erence between tree to tree distances,
point to tree distances, and intensity estimates as separate properties of a spatial
pattern. The bias is introduced by using the same sample for estimating density
is discussed above.

32
Pielou (1959) de�nes an index of nonrandomness that has the following range �
is 1 if for a \random" pattern, �! 0 for \uniform" patterns and �! arbitrary
upper limit for \clustered" patterns.
Demonstrates the relative range of value for three very di�erent patterns. The
Hopkins' N index will be used in this paper because it is scaled between 0 and 1, the
index seems to be the most well behaved in extreme conditions, and it is relatively
easy to calculate.
The spatial indices discussed so far are all �rst order (i.e. measures use the distance
to the nearest neighbor. The Holgate index does incorporate distance to the second
nearest neighbor but does not exploit higher order information. There are a set of
statistics that exploit more of the information by evaluating the relationship of a set of
points to all other individuals. A few such statistics are Ripley's K, and semi-variance.
� Ripley's K
This second-order test looks at the relationship of each point to all other points
(Ripley, 1981; Diggle, 1983; Tomppo, 1986). If the point pattern is Poisson, the
cumulative distribution of a number of individuals within a given distance will
follow �t2, where t is the maximum distance observed.
This statistic provides more information about the underlying process than the
previously mentioned statistics. If K(t) > �t2 the process is \clustered" and if
K(t) < �t2 the process is \uniform." Additionally, the size and range of the
deviations from �t2 can be determined about the process.
� Semi-variance
Semi-variance is the basis for many geostatistic tools and is a di�erence between
all points a given distance apart. (Palmer, 1988). This concept may be applied
in one, two, or more dimensions; here the two dimensional case is discussed.

33
This statistic must be applied to a value that can be measured as a continuous
variable across the study area. This technique has potential for resource vari-
ables such as nitrogen N or water content of the soil as well as aggregated stand
variables at landscape scales. Palmer (1988) plots several hypothetical process
patterns and the resultant semivariogram as well as several scales of plant data.
While these measures provide more information about the underlying spatial pat-
tern the calculation time and the added value of the information may not be worth
the added computation time required.
3.1.4 Species
The relative number of individuals in each of species has also been used as a measure of
the structure of plant communities. In the pollenology literature the relative amounts
of the various species is the main descriptor of forest structure (Von Post, 1946; Henry
and Swan, 1974; Whitmore, 1982; Winkler, 1985; Brubaker, 1986; Liu, 1990; Ford,
1990). Additionally, models have been built to predict the change in the species
composition of a plant communities (Botkin et al., 1972a; Botkin et al., 1972b; Urban,
1990). As with the size distributions, species can be described with distributions;
however, the species are categorical and do not have an inherent order. Methods of
ordering species by tolerance, abundance, or serial stage have been used.
3.1.5 History
Forests develop as a set of distribuance, regeneration, and mortality events interacting
with the above elements of stand structure. When discussing history; however, the
logic may become circuitous, because di�erent histories can interact with similar
patterns or distributions or species and produce very di�erent stand structures. This
is not to say the processes are not predictable; but if one ignores the history of a
forest, the assumptions about the stand development may be in error.

34
A number of studies have demonstrated through stand reconstruction how the
history of a stand interacts with the pattern, size distribution, and species to create
the current forest structure (Oliver and Stephens, 1977; Oliver et al., 1985; Oliver
and Larson, 1990).
3.2 Diagnostic criteria
Silviculturists have developed many relationships that help assess the condition of a
stand and its potential for treatment. The relationships are called diagnostic criteria
(Oliver, 1992) and are a familiar way of examining a stand for many silviculturists.
Model builders can utilize this experience by building growth models that present the
results of forest simulations in these familiar ways.
Diagnostic criteria can be grouped into three categories, each with a speci�c di-
agnostic use. The categories include stocking or stand density measures, growth or
change measures, and condition or vigor measures.
3.2.1 Stocking and stand density
Stocking and stand density are two related concepts which sometimes are confused
as the same thing, stocking refers to the relative density of a given stand compared
to some standard stand. Usually, the standard is a \normal" stand but the concept
of normality is not widely use today. Stand density is some average stand measure
per unit area (Bickford, 1957; West, 1983).
Spacing in relation to diameter
Rules of spacing in relation to diameter are often referred to as the \D plus rule of
thumb" and were presented by Matthews (1935). He related spacing, diameter, and
basal area as:

35
S =c � dbhpBA
; (3.8)
where S is the spacing of the stand, c is a constant to correct for the units and �
(Matthews reported a c value of 185 for english units.), dbh is the mean stand diameter
at breast height, and BA is the stand basal area per unit area ( acres or hectares ).
This rule is sometimes used as an aid in thinning a stand to a constant basal area
per unit area. The \D rule" can be plotted over time to indicate the need to thin a
stand.
Density indices
A large number of density measures, formed as indices, include basal area per unit
area, Curtis's relative density (Curtis, 1971), crown competition factor (Krajicek
et al., 1961), and Drew and Flewelling's maximum line (Drew and Flewelling, 1979).
These are summarized in West (1983), including their formulas and mathematical
relations to each other. All measures provide ways to scale a stand's density from
zero to a species' \maximum." Each measure emphasizes a di�erent component of
stand density and therefore explains a slightly di�erent part of the story.
Spacing in relation to height
Spacing|top-height ratios are the ratio of the tree spacing to tree height and have
been applied with the average spacing of the stand and the top-height (i.e. the average
height of some arbitrary number of the largest trees in the stand). These measures are
argued to be better than diameter-based measures of stocking since height is reported
to be less a�ected by stand density. Wilson (1946) reported the following relationship
of spacing to height
TPA =A
(b � ht)2 ; (3.9)

36
where TPA is the trees per unit area, A is the square units per unit area (e.g square
feet per acre or square meters per hectare), b is a fraction of height and ht is the top
height for the stand. Wilson advocated thinning stands to a given value of b. This
measure can be applied as the average spacing of a tree with its competitors and its
height to provide an index of an individual tree's growing space and an idea of the
range of density conditions within the stand.
Spacing in relation to volume
Drew and Flewelling's (1979) maximum size-density line provides an example of
this type of relationship. The advantage of volume is that it integrates diameter
and height into a single measure; however, it has the disadvantage of being tied to
a speci�c volume equation. An interesting variation is the relation of bole area to
number of trees (Lexen, 1943). Bole area provides a measure more closely related to
stem respiration. With sapwood taper equation now available (Maguire and Hann,
1987), sapwood volume relationships may prove of interest.
Density Management Diagrams
All of the above relationships can be plotted in log-log space to produce density
management diagrams. Density management diagrams are plots of trees per unit
area (e.g. trees per acre or trees per hectare) versus average size (e.g. mean diameter,
mean height, mean volume). The movement of a stand through this space provides
information about a stand's relative position in terms of a hypothetical maximum size-
density relationship and the rate at which the stand is approaching that maximum.
These relationships were �rst described by Reineke (1933) in which the average
of some size component of the stand (e.g.. diameter (Reineke, 1933; Long et al.,
1988; McCarter and Long, 1986), height (Wilson, 1951), or volume (Drew and
Flewelling, 1979)) is plotted against the average number of stems per unit area. These
are probably the most developed of the diagnostic criteria, with the largest body

37
of literature, including two growth models based entirely on assumed trajectories
through this space (Smith and Hann, 1984; Lloyd and Harms, 1986). These plots
present the mean trend of the the size variable, but provide no information about the
character of the underlying distribution.
3.2.2 Growth or change
Growth or change is probably the measure of most interest to silviculturists managing
a forest stand. Because growth or change is the means by which a stand moves from
the current condition to another, presumably more desirable, condition.
Diameter Growth
Diameter is the most accessible of the tree dimensions because it is easy to measure.
Diameter at breast height has no particular biological signi�cance. Many people
have advocated measuring diameters at proportions of tree height (e.g. 10 percent
of tree height), however, this may not be logistically convenient. Diameter growth
is the easiest of the tree's dimensional changes to remeasure. With paint a semi-
permanent mark can be placed on the stem for diameter remeasurement. Additionally,
increment cores can easily be collected to determine radial increment. These growth
measurements can be plotted over time to determine rate of diameter change.
Years per unit measure (e.g., years per inch or years per centimeter) is a measure of
the diameter growth rate of an individual tree. By analyzing the trends in individual
tree diameter growth rate, one can assess how well the tree has been competing in the
past and whether the rate will increase or decrease. The diameter growth can also be
plotted in cumulative form, presenting diameter over time. This plotting can be easily
done in the �eld by marking, radial increment versus time on graph paper. When
observing diameter growth a decline is expected as a consequence of the geometry of
placing the new wood around an ever increasing core. This must be accounted for in
any interpretation of diameter growth.

38
Height Growth
Height is one of the most di�cult dimensions of a tree to measure because the view
of the tree tops become obscured as stands grow taller. Height is one of the more
biologically signi�cant dimensions, since it is directly related to a tree's competitive
status. Height growth is usually determined in two ways: One is repeat measurement
of individual tree heights. This method often yields poor estimates of height growth
except when the trees are short. Large variances are usually observed, even when the
same person remeasures the heights. The second method is to measurement of height
growth from stem analysis. This is usually the preferred method; however, the tree
is destroyed in the process.
Volume Growth
Traditionally, volume growth is the feature of a stand that most interest foresters.
Volume is a variable that is an estimate as opposed to variable that is actually mea-
sured. Stem volume is the feature of a stand that is most closely related to timber
products. Under a timber management objective, stand volume growth can be a mis-
leading statistic in that stand volume growth can be added to stems that will make
useful products or to stems that can not be harvested. A stand of tree with less total
volume but on a fewer stem will usually be more valuable.
3.2.3 Condition or vigor
The third set of diagnostic criteria focus on the condition or vigor of a stand and
how that stand might respond to a thinning. These include measures such as height-
diameter ratio, leaf area index, crown closure, and sway period.

39
Height-Diameter Ratios
Height-diameter ratios are the ratio of a tree's height to it's diameter in the same
units. This tool can be used to track the stability of a stand of trees over time and
providing a indication of the time when the stand will become unthinnable (Wilson,
1946; Wilson, 1951). Traditionally, silviculturists have considered trees with height-
diameter ratios greater than 100 as unstable for thinning.
Leaf area index
Leaf area index is a standardized measure of the amount of leaf surface area per unit of
ground surface area. Leaf area index can be a very useful indicator of a stand's ability
to respond to thinning and/or fertilization (Vose and Allen, 1988). The problem with
leaf area index is that to date there is no reliable, easy method of measurement.
Currently the most common technique is to predict leaf area from the tree's diameter
or sapwood basal area and species. Another technique is to measure the relative light
interception of the canopy and use these values to estimate leaf area index. These
techniques are highly variable; and a reliable, fast e�cient method is needed to allow
the widespread use of this index.
Live crown ratio
Live crown ratio is a estimate of the amount of crown on a individual tree. It can
be used in determining the vigor of a tree. This is a measure that is used widely by
growth models and some volume equations (Walters, 1986; Valenti and Cao, 1986)
Crown Closure
Crown closure is an estimate of the proportion of sky covered by foliage observed from
beneath the canopy. This measure can be estimated from spherical densiometers in the
�eld or from hemispherical photographs in the o�ce. Crown closure is an imprecise

40
estimate of the amount of leaves in a stand. This is not as useful as LAI because
it only indicates the presents or absences of leafs with little information about the
number if present. It is, however, useful for determining site utilization.
Sway period
Sway period is a measure of the periodicity of the sway of a tree. This is a biome-
chanical property of a tree. The theoretical period can be calculated as
T = k �M � L3 (3.10)
where T is the period of sway, k a constant of proportionality, M is the mass of a
weight on the beam at height L. This relation was reported by Sugden (1962) as a
method of crown classi�cation for determination of stand competition. Sugden sug-
gested using this method to determine the amount of foliage weight on a tree. Others
have questioned the biological reason for tree stem form and have used mechanical
support arguments to explain tree form (Wilson and Archer, 1979; McMahon and
Kronauer, 1976).
3.3 Stand dynamics and growth models for management
Silviculturists use growth models to predict stand change and how disturbances will
e�ect that change. Silviculturists usually describe stands in terms of stand structure
using the elements of the spatial arrangement, the relative sizes, species, and density
in intuitive, if not quantitative, terms. How these elements change over time provides
the other important element of stand structure | history.
All models are designed to provide a speci�c type of answer. If a model is used
for a purpose other that the one that it was originally designed for the user should
reexamine the all the assumptions and relationship built into the model and access
the consequences of those assumption on the current problem.

41
3.3.1 Related models
Related models have had an in uence on the approach to silvicultural modeling de-
scribed in this paper. These three models have taken approaches that are quite
di�erent than that of the majority of forest growth models. The �rst is the TASS
model (Mitchell, 1975), second a stand dynamics demonstration model CROGRO
(Fellows, Sprague and Baskerville, 1983) designed to teach the principles of crown
development to students, and the third is the many publications of models for the the
management of Scots pine (Pukkala, 1987; Pukkala and T. Kolstr�om, 1987; Pukkala,
1988; Pukkala, 1989a; Pukkala, 1989b; Pukkala, 1990).
The TASS model
The Tree And Stand Simulator (TASS) is a distance dependent, forest growth model
that uses a three-dimensional description of a part of the stand to simulate tree
growth and crown interaction. Th amount of foliage on a given tree determines the
trees height growth and in turn branch growth. The resultant stem increment is
allocated over the stem. A description of the functions in the TASS model are as
follows.
This model has been calibrated to a larger amount of permanent sample plot
(PSP) data for Douglas-�r (Pseudotsuga menziesii (Mirb.) Franco ) and versions
are being prepared for western hemlock (Tsuga heterophylla (Raf.) Sarg.) and sitka
spruce (Picea sitchensis (Bong.) Carr. ). The exibility of the TASS approach is
that many treatment e�ects can be simulated for pure stand of calibrated species
(Mitchell, 1975). The TASS models have been used to generate managed stand yield
tables for British Columbia (Mitchell and Cameron, 1985). The ability to mix species
currently has been envisioned but not implemented. Many of the equations of this
model can be found in Appendix B.

42
Crown Length
Maximum Crown Length
Height
Maximum Branch Length
Foliage volume
Figure 3.6. Diagram of the crown description for the TASS model
The CROGRO model
While this growth model had much less e�ort expended in its construction it is no
less interesting in concept. The CROGRO model was built as a tool to teach crown
development to students (Fellows et al., 1983). The approach made a number of
assumptions about crown interactions. It was calibrated with regional growth and
yield information and was evaluated by presentation to students, foresters, and aca-
demics. One of the most interesting features is that the model results are displayed
in two-dimensional computer drawing of the trees and their crowns.
Height growth is the driving function of this model. The actual height growth
is determined by reducing the potential height growth by the ratio of the projected
vertical cross-sectional area of the crown to the optimal vertical cross-sectional area
of the crown. The equations for this model can be found in Appendix B.
The driving functions are based on local yield curves. The estimate of growth
is modi�ed by the vertical projection of crown area and this is modi�ed by tree

43
Maximum Branch Length
Branch Base and TipHeight difference between Branch Angle
Maximum Crown Radius
Figure 3.7. Diagram of the crown description for the CROGRO model
interaction. Figure 3.8 illustrates the basic parameters of crown shape in this model.
The functions are rate curves derived from the weibull function �t to the height-age
curve.
This approach was found acceptable for the design purpose. The dynamics of
open grown crowns were not well represented; however the essential dynamics of
stand grown trees were acceptably represented.
Pukkala's Model
Pukkala has build a spatial growth model called MikroMikko for Scots pine (Pinus
sylestris L.) in Finland. Average stand characteristics were used to generate a tree list

44
for a spatial pattern. In Pukkala (1989b), two methods were presented for generating
diameters from a spatial pattern. The �rst method assumed that trees with many
close neighbors will be small and trees with few close neighbors will be large. The
second method used the local spatial pattern to predict a diameter distribution and
then this distribution is sampled to determine the diameter for the subject tree. Then
the spatial pattern and the predicted diameter were used to predict tree height. The
equation for diameter and height can be found in Appendix B.
Pukkala's model used these relationships in the following sequence.
1. Stand average statistics such as density and total stand basal area are speci�ed
for the stand;
2. The tree coordinates are generated as a realization of a suitable spatial process;
3. The diameters are predicted from the spatial pattern;
4. The heights are then predicted as a function of the spatial pattern and the
diameter.
The output of these models are displayed in two and three-dimensional displays
showing also statically predicted crown dimensions. This model has been extensively
tested against non-spatial models built from the same data. Pukkala (1987) has
shown that the non-spatial models may be considerably in error, if a stand has large
amounts of variability.
Pukkala's has put these models to interesting uses. In one study, Pukkala explored
the e�ects of roads for thinning equipment placed through a stand at 20 meter intervals
(Pukkala, 1989b). If the remainder of the stand was thinned uniformly, the model
predicted that the trees along the thinning road would have higher growth than the
internal trees within the strip. By experimenting with these simulations he devised a

45
n
i = 1
i
4
3
2
1 Θ
Θ
ΘΘ
θΣ
5 meters
Figure 3.8: Diagram of the spatial competition measure in Pukkala's (1987) growthmodel.

46
thinning proportion between the skid roads that equalized growth among all trees in
the residual stand.
Another model presented by Pukkala (1987), along with work currently in progress
Pukkala (Personal communication, 1990), explored the spatial relationship of an over-
story canopy and the initiation and survival of tree seedlings.

Chapter 4
METHODS
\Seek simplicity, and distrust it"
Alfred North Whitehead
\... our position is that modeling is an art; that the �rst task is to de�ne
objectives; the second to select a consistent view of the system; and only
later, and if appropriate, to use a mathematical description."
Mike West and Je� Harrison, 1989. Bayesian Forecasting and Dynamic
Models
Stand management for a variety of objectives is di�cult when the di�erent ob-
jectives are expressed in diverse measures (e.g., numbers of roosting pairs of birds,
board foot volume for timber, acre-feet of water yield for drainage basins. Stand
structure can be the common language among the various disciplines that deal with
stands (Oliver, 1992). Foresters can produce forests that will meet the multi-resource
goals, if the range of stand structures and the timing of those structures can be de-
scribed for the diverse objectives. Management with the stand structure approach
requires a through knowledge of stand development and how management can e�ect
that development.
Adaptive models of stand dynamics are based on the idea that theories of forest
stand dynamics can be used to de�ne the relationships for projection of stand struc-
tures. These general relationships can then be calibrated to given stand conditions
to \localize" the predictions. Adaptive models of stand dynamics are less precise at

48
estimating volume, although they can accurately describe stand structure and the
change in that structure over time.
Adaptive models of forest growth can be built in many forms. An individual tree
model of stand dynamics will be presented in the current example. The individual tree
model assumes that each tree responds to its immediate environment. In individual
tree models, it is easier to describe the expected biological relationships, to de�ne the
e�ect of disturbances, and to re ect species di�erences. There are disadvantages to
the individual tree approach as well. Individual tree models have more parameters to
de�ne, which makes the models more di�cult to test. A whole stand model has the
advantage of being easier to parameterize and test data, faster to run, and if stand
average estimates are all that is needed they may provide the information needed.
When designing any type of model, there are facts that e�ect the performance of
the model whether admitted or not. These facts are especially important to consider
in the design process:
� Models are nothing more than an explicit statement of a person's or group of
people's ideas about a system;
� All models are biased by the modeler in the parts they choose to include and
exclude;
� A modeling approach's success depends on attempts by several modelers the
determine overall feasibility of the approach.
4.1 Use of an adaptive stand dynamics model
An adaptive stand dynamics model is designed to utilize whatever data is collected
from the stand of interest and information on the species' age-height trends. This
procedure allows the general tree and stand dynamics trends to be adjusted to the

49
local site and species speci�c conditions or to expected growth patterns. The following
are the steps need to set up the model:
� Data gathered from the stand or group of stands of interest are used to parame-
terize the functions in the model. This tree list should include species, diameters
at breast height, total tree height, height to crown base, average crown width,
and a stem map of the trees. Obviously this information is not always avail-
able or obtainable. The stem map is use to calculate an Hopkins' index. An
alternative method would be to measure a number of distances between random
points and the nearest neighbor tree (e.g. 30 distances) and then from those
same thirty trees to their nearest neighbor tree. If the stand is mapped and
the random sample points determined in the parameterization, it is less likely
to be biased. Crown measurements are very important in this type of model to
de�ne the parameters properly, since the growth of a given tree is a function
of crown size. Only general theoretical trends are built into the model|no re-
gional trends. This means that the results are very dependent on the data that
the user collects. The advantage of this design is the exibility; however, this
places more responsibility on the model user. Flexibility means the ability to
adjust the growth trends within the model to re ect the expect growth patterns
for the stand and species.
� The height growth trends must also be de�ned for each species considered. Two
approaches can be taken to obtaining this information. The �rst is to take
data from stem analyses of trees representing dominant or co-dominant growth
patterns for the various species. This is rather labor intensive and needs to be
done when the results are likely to di�er from existing age-height curves. The
other approach is to use existing regional age-height curves for the species and
site.

50
Value Cell - End result of current model.
Decision Cell - input information is used to decide on course of action.
Deterministic Cell - item will always produce the same output for the same input.
Probablistic Cell - item measured with error or a random number.
Figure 4.1. The de�nitions for the shapes in an in uence diagram.
These data and information are used by a parameterization routines to determine
the various parameters estimates for the model. The data and information described
here are the variables needed to parameterize the model presented in the example. If
the functions were changed to have other variables, the data and information require-
ments would change.
Using these parameter estimates, projections are made for the stand; then, alter-
native management scenarios can be considered. The topic of selection of management
scenarios using this growth model will be left for future work.
4.2 Components of an adaptive stand dynamics model
Figure 4.1 presents the de�nitions for the four basic shapes used in in uence diagrams
(Shachter, 1986). These diagrams are acyclic (i.e. loops are not allowed in in uence

51
diagrams). If a decision box is entered with only one outlet, the previous procedure
is continued until the exit condition is obtained. Another description method is
mathematical notation where the pertinent variables will be describe at the time of
presentation. Then each model is described verbally.
4.2.1 Component models for stand generation
The �rst set of submodels generate a simulated stand from input parameters. The
models have components for the prediction of spatial pattern, static height pattern,
crown size, and diameters of the trees. A simulation approach is used to make the
stands grow, for two reasons. First, the growth models are a combination of determin-
istic and probabilistic techniques. The deterministic part describes the mean trend
for the relationship, the probabilistic part adds variation similar to that observed in
the input data. This type of model is designed to capture the general behavior of
the stand structure, not to accurately predict a speci�c tree. Second, the proposed
approach is to explore a di�erent technique. The aim is to describe the stand dy-
namics behavior of the speci�c stand, not precise yield estimates of a speci�c stand.
The emphasis in on de�ning a range of outcomes. Figure 4.2. illustrates the order in
which the submodels are executed.
4.2.2 Generation Process
Spatial pattern
The process of generating a suitable spatial pattern has problems associated with it.
Several people have developed methods to generate spatial patterns. The problem
is that one speci�c resultant spatial pattern can be generated by several possible
generating processes; therefore, simply knowing the spatial pattern does not provide
information about underlying generating processes. The approach used in this study
is to group patterns into two types, stands started as plantations and stands started as

52
Tree List
Foliage
Diameter Crown SizeHeight
Species
SpatialPattern
Figure 4.2. The in uence diagram for the generation process
ParametersSpatial
Process
PatternTest Pattern
Lattice
ProcessNeyman-Scott
ProcessDetermine
Origin of Stand
Figure 4.3. The in uence diagram for the pattern generation process.
natural regeneration. This distinction that should be rather easily made by foresters.
Once classi�ed, two di�erent generating processes are used. These two processes
use similar parameters but make di�erent basic assumptions about the underlying
pattern. The process of calculating the appropriate parameters involves an iterative
routine in which starting parameters are chosen. A simulated stand is generated and
compared to the input pattern. To make this comparison, a statistic must be used.
The statistic chosen for this study is Hopkins' N , it is easy to interpret, it is

53
11
11
1
11
11 1
1 1 11
11 1
1 1 1
2
2
2
22
22 2 2
2 2 2 22
22
22 2
2
3
3
3
3
33
33 3
3 3 33 3 3
3 3 33
3
44
44
4
44
4
4 44 4 4
4 4 4 4 44 4
55
55
55
5
55
55
55 5
55
5 5 5 5
(A) Lattice
Amount of Distance Variation
Hop
kin’
s N
5 10 15 20
0.0
0.2
0.4
0.6
0.8
1.0
11
1
1 1 1 1 1 1 1 11 1 1
1 11
11 1
2
2
22
2 22
22 2 2 2 2
2 2 22
22 2
33
3
3
33
33 3
33 3 3
3 3 33
3 33
44
44 4
44
44 4
4 4 4
4 4 44 4
4
4
55
55
5 5
5
5 55 5
5 5 55 5 5
5 55
(B) Neyman-Scott
Amount of Distance Variation
Hop
kin’
s N
5 10 15 20
0.0
0.2
0.4
0.6
0.8
1.0
Figure 4.4: The range of Hopkins' N for various pattern generation parameters. The(A) lattice process and the (B) Neyman-Scott process. These diagrams illustratethe e�ect of varying the input parameters to the two generating processes and theresultant hopkins indices generated as a outcome.
rather easy to calculate so can be calculated at each step of the iteration process, and
it provides reasonably stable results. A diagram of the submodel is in Figure 4.3.
If a plantation origin is selected, a lattice process is used to generate the spatial
pattern. The lattice process assumes the trees are arranged on a lattice pattern with
variation from that pattern. The nominal spacing is determined from the density of
trees and the area of the sample plot. A grid of points is establish with a random
starting point. These points are called the \parent points" and are the intended
location of the trees. From each parent point a random azimuth and distance within
a speci�ed range are chosen to establish the location of the child point. Trees are
assigned to children points only. The parameters for a lattice process are the spacing
of the parent points, the variation range for each child point and the number of child
points. In plantations the number of child points is usually one. Additionally all
points are tested to be within the range of the simulated plot. This process seems

54
to work well for Hopkins' values from 0 to 0.5 as seen in Figure 4.4(A). The main
method of changing the Hopkins' value of the simulated stand is by changing the
variation range parameter.
If a natural stand origin is selected, a Neyman-Scott process is used. Neyman and
Scott (1972) were astronomers, who developed this process to generate patterns for
simulated galactic clusters. The generating process is based on a random or Poisson
point pattern. The random or Poisson pattern is generated by a binomial distribution
of two uniform random numbers for the required number of parent points. From these
parent points, a random azimuth and random distance, within a range limit, is applied
to produce each child point. Additionally, all points are tested for the range of the
simulated plot. This process works well for Hopkins' values from 0.5 to 1.0 as seen
in Figure 4.4(B). The parameters to match the Hopkins' index can be determined
by changing the variation range and the number of child points associated with one
parent point. Figure 4.4 is a graph of how the parameters can be varied to generate
di�erent spatial patterns.
Species
The next step in the tree list generation process is to assign species to each point in
the generated pattern. The parameters for this submodel are the relative proportion
of each species on the sample plot. From these proportions, each point is randomly
assigned a species in the same overall proportion as the original sample plot. The
assumption is that the number of of individuals of a given species in the simulated
stand will be in the same proportion as in the current stand. This is probably only
valid if the age of the generated stand is near the age of the observed stand. A better
approach would be to grow a stand from a assumed regeneration event; however, the
number of assumptions and time to calculate this approach would make the result a
dubious improvement. Figure 4.5 presents the in uence diagram for this submodel.

55
Tree DataInput
For Species
RelativeSpecies / TreeProbabilities
Figure 4.5. The in uence diagram for the species generation process.
Height / TreeDistributionBeta
Random Number
ParametersHeight
Figure 4.6. The in uence diagram for the height generation process.
Height
Heights are generated from the parameters for a four parameter beta distribution
�(a; b;min;max) by species. A beta distribution was choosen because of its exibil-
ity. Beta distributions can �t most unimodal distributional shapes and some bimodal
shapes. For these reasons less assumptions regarding the underlying shape are re-
quired. A range is speci�ed for the heights of the stand to be generated and the
heights are randomly drawn from this beta distribution and assigned to each tree
in the tree list. Again the assumption involved with this approach are only valid
for generated stands of an age near the observed stand age. Figure 4.6 presents the
in uence diagram for the height generation submodel.
Diameters
At this point in the ow of the generation process (Figure 4.7), the pattern, species,
height, have been determined; and diameter, crown width, height to crown base, and

56
Height
Dbh / TreeDiametersCalculate
Tree
ParametersDiameter
Random Number
Figure 4.7. The in uence diagram for the diameter generation process.
amount of foliage are left to be determined.
For the generation process, diameters are predicted as a direct relationship to
height. A height/diameter equation is parameterized for the input data and is used
in the inverted form to predict diameter. This done to allow the �tting of a linearized
form to the input diameter-height data.
di =
"ln(hti � bht)� b0
b1
# 1
b2
; (4.1)
where di is the diameter at breast height for the ith tree in the tree list, hti is the
height of the ith tree in the tree list and bht is the breast height for the measurement
system (i.e. english or metric). This equation is applied with random error determined
from the input data to mimic relationships found in the original data. The residual
from the above �t diameter-height equation are estimated as a beta distribution. A
random draw from this distribution is added to the deterministic prediction from the
above equation. The resultant diameter-height values mimic the original distribution
of the input data.

57
Height to
Height toCrown Base
Crown Base
Equation
Tree
Crown Width
Heights
Height/DiameterHeight/DiameterTree
Parameters
Calculate
Crown Width
Crown Base
Random Number
Parameters
Figure 4.8. The in uence diagram for the crown generation process.
Crown size
Crown size for the generation process is predicted from diameter using the following
equation. The inverse of this relationship has been used for years in aerial photogram-
metry as a method of determining volumes for sample plots on aerial photographs
(Paine and McCadden, 1988). The relationship is of the form:
cwi = b0 + b1db2i ; (4.2)
where di is the diameter for the ith tree, cwi is the crown width for the ith tree in
the tree list, and the b's are the regression coe�cients.
Crown length is determined as a deterministic ratio to crown width for the species.
Again a better method to determine that crown widths and crown lengths is to grow
the stand from an inital state. But the generation process is designed to produce a
simulated stand of a similar character to the input stand.
Figure 4.8 is the in uence diagram for the height to crown base prediction in the
generation submodel.

58
Foliage
The foliage calculation function has three basic components for determination of
foliage amount per tree. These three parts are the calculation of the surface area
of a geometric solid of the dimensions of the tree crown, the conversion of the surface
area of the solid to foliage area, and the calculation of the variation in these functions.
They are diagrammed in Figure 4.9.
The idea of calculating foliage amount from the surface area of a solid is from
Maguire (1989). In this paper, Maguire discusses the relationships of gross crown
dimensions to to sapwood area at crown base. Sapwood area at crown base has been
shown to be a good estimator of leaf area in Douglas-�r (Grier and Waring, 1974;
Waring et al., 1982; Whitehead et al., 1984; Espinosa Bancalari et al., 1987) and in
other species (Rogers and Hinckley, 1979; Dean and Long, 1986; Keane and Weetman,
1987).
The �rst part is to calculate the surface area of a geometric solid. The solid can
be any shape from a paraboloid, to a cone, to a neiloid. The di�erences between the
various areas are negligible compared to the variation added, so the formula for the
lateral surface area of a cone was used.
The conversion factor allows the assumption that foliage amount is related to
surface area in some proportion. A conversion factor of one is assumed in the current
example. As better methods of measuring canopy foliage are developed, this type of
factor will become important in distinguishing between foliage conditions.
An amount of variation was added to the predictions to indicate the assumed
uncertainty of the relationship. A variation of �20% of the surface area was added as
a random normal to the foliage amount in the current example. The other equations
can be found in Appendix C.
SA � N(dSA; :2 �dSA); (4.3)

59
Parameters
FoliageFoliage / Tree
Foliage areaSurface area to
Proportion
Calculate
DistributionBeta Error
Surface AreaCalculate
VariationProportion
TreeCrown Length
TreeCrown Width
Random Number
Foliage
Figure 4.9. The in uence diagram for the foliage generation process.

60
Upon completion of the generation process, a tree list has been created that has
a similar statistical character as the originally sampled stand. This generated stand
is used as input to the growth submodel of the model.
4.2.3 Component models for growth
The growth submodel is the portion of the model that changes the list of tree variables
from a given time to the next time step. An in uence diagram of the relationship
of the components is given in Figure 4.10. From the diagram it will be noted that
the process is much less dependent on the calculations in the other submodels at the
current time step. The process is very dependent on the values in the starting time
tree list.
The philosophy of the growth submodel is based on the idea that the rate of change
of an individual tree on a given site is dependent on that tree's height and crown size.
Three factors in uence the rate of change of a tree: the maximum potential rate of
change for the species on the site, the ability of the tree to produce photosynthate (a
function of the size of the crown), and limits on the size of the tree (analogous to the
respirative cost for increasing size). This model uses these simple concepts about the
change of trees and aggregates the trees into stands. This section will now examine
the strengths and weaknesses of the approach.
Height Growth
Height growth is one of the more complex functions within the submodel. It is
designed to use readily available data assuming an interpretation of that data, to
predict height growth of a tree. Figure 4.11 is the in uence diagram for the height
growth submodel.
The height growth submodel has three parts; a rate �, a modifying function to
determine the role of relative foliage amount, and a modifying function to determine
the role of relative tree size in height growth. The rate value � is the periodic rate

61
Tree ListEnding
FoliageCalculate
Regeneration
Diameter Growth
Crown Change
Height Growth
Tree ListStarting
Figure 4.10: The in uence diagram for the growth process. Note that only the crownchange part depends on the values of other parts. All parts depend on the startingconditions.
DerivativeWeibull
Parameter
HeightCalculate
Exponential
Parameter
HeightEnding
Height Growth
Function
Function
Height GrowthSize
Height GrowthFoliage
Tree ListStarting
Figure 4.11. The in uence diagram for the height growth process.

62
Height growth and Height growth rate
Time (yr)
Hei
ght g
row
th (
m)
0 50 100 150
0.0
0.2
0.4
0.6
Time
010
2030
40H
eigh
t (m
)Figure 4.12: Example height-age curve with the base curve (solid line) and the �rstderivative of the curve, which is the height growth rate curve (dotted line).
of growth at the in ection point of a height-age curve for the species and site. This
represents the period in a tree's life for which it is growing at the fastest rate. This
point is determined by �nding the maximum of the �rst derivative of the height-age
function. This point is used as the maximum growth rate (Figure 4.12).
The �rst modifying function predicts the e�ect of foliage amounts other than the
amount of foliage on a tree growing at the maximum rate. First, a tree with a full
crown (crown length equal to tree height) of the size of the maximum rate fol� is
assumed to grow at the maximum rate �. Further, additional foliage is not assumed
to increase the tree's height growth rate. Trees with less foliage than the maximum

63
Foliage Ratio
Mod
ifer
0.0 0.2 0.4 0.6 0.8 1.0
0.0
0.2
0.4
0.6
0.8
1.0
Height Ratio
Mod
ifer
1.0 2.0 3.0 4.0 5.0
0.0
0.2
0.4
0.6
0.8
1.0
Figure 4.13: Example foliage ratio and height ratio functions for the height growthsubmodel
rate amount fol� will grow as a full crowned tree of that crown length. The main
e�ect of this part of the equation is to reduce growth on trees with foliage amounts
less than the maximum rate amount fol�. Figure 4.13a is a graph of the shape of the
multiplicative e�ect of this part of the equation.
The second modifying function de�nes the e�ect of trees growing larger or the
upper asymptote of the height/age curve. This part of the equation assumes that
height growth rate will slow down as trees become larger relative to the size of a
tree at the maximum rate size h�. This part of the submodel de�nes the asymptotic
behavior of the resultant height-age curve. The equation for this submodel is of the
form:
�h = �
"fol
fol�
#a1266664hmax
0BBBB@0@ h
h�� age�b
1Ac
c
0BBBB@exp
��
h
h��age�
b
�c!hh�� age�
1CCCCA1CCCCA =�
377775 ; (4.4)
where �h is the change in a tree's height, � is the rate at the in ection point of the

64
Ending
EndingCrown Dimensions
Starting
ParametersCrown Width
Crown WidthCalculate
Crown Width
Crown Base
New Crown BaseHeight to
No RecessionRepeat until
RecessionProbability of
CompetitorsLAI in
Cumulative
Random Number
ParametersCrown
Figure 4.14. The in uence diagram for the crown change in the growth process.
height-age curve, and the two modifying equation described above; where age� is the
age at the in ection point of the height-age curve and hmax is the maximum expected
height for the site.
This submodel, while a bit cumbersome, has a rational appeal. The rate � is
the site speci�c maximum growth rate for trees of that species. All site di�erences
are incorporated through this term. Once a growth rate is speci�ed, height growth
has less of an e�ect on treatment decisions. Also the �rst modifying function may
be thought of as the photosynthetic potential for the tree and the second modifying
function as the respirative cost of the tree becoming larger. These functions are tied
to height growth because the model focuses on tree dimensions that relate to stand
structure changes. Here, implicit assumptions are made concerning the allocation of
photosynthate with in a tree.
Crown Change
The crown change submodel is the one submodel that is dependent on the current
cycle's prediction of tree height as one of the dimensions of the crown length change.
Figure 4.14 is an in uence diagram of the crown change function.

65
Competitor
Subject Tree
Non-Competitor
θ
θ
Figure 4.15. Illustration of determination of competitor trees.
A vertical pro�le of the leaf area index of all trees considered competitors of
the subject tree is determined by summing the proportional amounts of each tree's
vertical foliage pro�le into a cumulative vertical pro�le. The equations to accomplish
this summations are as follows:
Lk =mXj=1
GroundXk=Canopy Top
�jA�(aj; bj) Ij;k; (4.5)
where Lk is the cumulative leaf area index of competitors for each tree at level k from
the top of the canopy, m is the number of trees considered to be competitors to the
subject tree, � is the amount of leaf area on tree j, and A is the area associated with
the maximum distance of the m competitors to each subject tree. Beta probability
density functions (p.d.f.) �() with parameters aj and bj are for the species of com-
petitor tree j and are used to distribute the amount of leaf area � along the crown
length of competitor tree j. Ij;k indicates whether level k is within the crown length
of competitor tree j.

66
Leaf Area IndexCumulative
Height
Competitors
Competitors
Subject Tree
Non-Competitor
θ
θ
θθ
θ
Figure 4.16: The method of determining the cumulative leaf area index in competitors
For all competitor trees, a cumulative leaf area index pro�le from the top of the
canopy is constructed (Figure 4.15). A probabilistic prediction is based on the current
leaf area competition at the crown base for the crown base changing one height unit
(e.g., 0.3 meters in the current example). The process is stopped for that subject tree
for that cycle if the equation predicts no change. If the equation predicts a crown
change then the process is repeated until no change is predicted, and the latest crown
base value becomes the crown base in the new tree list.
Given this cumulative leaf area pro�le the probability of the crown recession of
one height unit is determined by:
P (i; k) = 0:5 +1
�arctan [�(Lk � LAI)] ; (4.6)
where P (i; k) is the probability of the crown receding at height k to height k+1 on tree
i, which is predicted by an arc tangent function (Figure 4.17). The LAI determines

67
the location of the in ection point on the \x" axis. This LAI is the level of leaf area
index in competitors at which the foliage on the subject tree dies.
Regeneration
Seedling regeneration is predicted from a distribution describing the number of seedlings
per time step for a given species. A random variate is drawn from this distribution
for the number of seedlings to consider for regeneration for the time step. A cumu-
lative leaf area index pro�le is calculated for the seedling location, just as for the
crown change function, and probability of survival is predicted for the seedling. If the
seedling survives, it is added to the tree list as a live tree. Figure 4.18 is an in uence
diagram of the regeneration process.
Mortality
Tree mortality is usually one of the weakest links in forest growth models. Trees
can die for many reasons or from many causes. In general, trees die from suppression
through competition, windthrow, breakage, and attack by insects or disease. Pest- and
disease-caused mortality are host- and site-speci�c and require special submodels of
those species. Breakage and windthrow are functions of the soils, species, combination
of the physical dimensions of the tree, and local wind environment.
Suppression is relatively the easiest to predict and is a function of reduced growth
from competition with neighboring trees. It is the only mortality form predicted in
this model. There is no explicit mortality function in the current version of the growth
model. A tree dies when a tree's crown recedes above tree height. This calculation
only represents one form of suppression mortality.

68
Leaf area index in competitors above point of consideration
Pro
babi
lity
P(i,
k)
0 2 4 6 8 10
0.0
0.2
0.4
0.6
0.8
1.0
Figure 4.17: A plot of an example arc tangent function for predicting crown recession,� = 4 for this example

69
at EndingNew Trees
Survival
Test Seedling
for Seedlingsx, y locations
Generate
CompetitorsLAI in
Cumulative
ParametersPattern
ParametersRegeneration
Tree ListStarting
Figure 4.18. The in uence diagram for the regeneration growth process.
Stem Increment
Stem increment should follow a function of the form described by Long et al. (1981)
and graphed in Figure 4.19. It is assumed that a crown of a given size will produce
the same area increment at crown base regardless of the amount of stem below the
crown. Using this assumption, a prediction of the area increment at crown base Ahcb
is determined. This equation is of the form:
�Ahcb = b1fol: (4.7)
The relative distance between the crown base and breast height on the stem is
used in this equation to predict the reduction in the amount of area at breast height
increment Abh with distance from the crown base:
�Abh =�
4d2t�1 +
"�Ahcb
1
1 + b1 e(�b2 hcb
h )
!#; (4.8)
where dt�1 is the diameter of the tree at the last time step, hcb is the height to crown
base and h is the total tree height and hcb=h is a bole ratio. The area increment is
then converted to diameter and added to starting time diameter:

70
Relative Area IncrementRelative Area Increment
HeightHeightRelativeRelative
Crown base
Crown base
Figure 4.19: The assumed shape of the stem area increment, similar to functionsdescribed by Long et al. (1981).
dt = 2
s�Adbh
�+ dt�1; (4.9)
Stem increment calculated with this approach re ects the changing size of the
crown.
4.2.4 Assumption of the Current Approach
There are many assumptions that a�ect the behavior and results of an adaptive stand
dynamics model. Here the assumptions are stated to clarify both thinking about the
model and expectations of the output.
� Stand responses are the aggregate of an individual tree's response to its imme-
diate environment.

71
DiameterEnding
Dbh Area
Crown BaseArea At Calculate
FunctionLogistic
Tree ListStarting
ParametersDiameter
Figure 4.20. The in uence diagram for the diameter growth process.
� Tree height increment is a function of crown size and tree size and the potential
growth rate for a species on a speci�c site.
� Crown size, as the productive part of a plant, is the dominant force changing a
tree's size.
� Stem area increment at crown base is a function of crown size regardless of the
tree size.
� Stem increment at other points on the stem fellow a function similar to Long et
al. (1981) (Figure 4.19).
� Leaf area is related in some proportion to the surface area of a geometric solid
representing a crown shape of that species (Maguire and Hann, 1987).
These assumptions are used to guide construction of model components.
4.3 Presentation of Model Output
An often overlooked part of growth models is type of output. Output should convey
useful information and be exible enough so the user can request information in a
desired form. Many forest growth models are built by modelers who, by virtue of
their interest and training, are most interested in the numerical output; however,

72
users relate more to graphical displays of the same information. Many people can
grasp graphical displayed relationships much quicker than tabular output. A growth
models that presents output in many imagitive ways could bring the results of grwoth
models to many more people.
Over the years, silviculturists have developed many relationships that help access
the condition of a stand and the potential for treatment of that stand. The relation-
ships are called diagnostic criteria (Oliver and Larson, 1990) and are familiar ways of
examining a stand for many silviculturists. Once a person is accustomed to analyzing
stands using these tools, a background of experience arises that aids the manager
in using of the tools. Model builders can utilize this experience by building growth
models that present the results of forest simulations in these terms.
4.4 Example System
\I cannot really imagine any other law of thought than that our pictures
should be clearly and unambiguously imaginable."
L. Boltzmann 1897
4.4.1 Objective
This chapter presents the detail of an adaptive stand dynamics model. There are many
ways to implement these ideas, some that work better than others. This dissertation
presents the ideas with the intent of conveying the concepts, rather than the details
of a particular implementation.
4.4.2 Design
This model is designed to use a \Run and Display" approach as opposed to a \Display
as you Run" approach. The growth submodel is \Run," and output is stored in an
output �le or data structure. Then, a separate display part of the model will read

73
that �le or data structure. This approach has several advantages. Any time step
in the growth sequence can be easily viewed; plots over time are easily made; and,
when many batch runs are needed this task is much easier. In \Display as You
Run" approach, the program plots the current values of the projected tree list. The
run usually must be reinitiated and rerun to show previous states of the stand; and
over-time plots are not available until the end of the run.
This model uses a simulation approach; input tree data are distilled into a set of
parameters that describe a stand. A simulated stand is then generated from these
parameter sets and used as the initial condition for model runs. Because of the
stochastic nature of the models, the output will di�er with each run with identical
input. If averages are desired, several runs must be averaged to obtain estimates of
the predicted mean and range.
4.4.3 Components of the models
The \GENERATE" program is a set of routines that produces a tree list from a
parameter set. As stated in chapter 4 the program steps through the routine in the
following order.
The function to generate the random numbers used in the C routines is the \ran3"
function from Numerical Recipes (Press et al., 1988), and inside S the \runif" function
is used.
Pattern { The �rst step is to generate a spatial pattern for the simulated stand.
One of two generating processes is used. If the stand is natural in origin then a
Neyman-Scott process is used; otherwise a lattice processe is used.
� Neyman-Scott process generates a pattern of random \parent points" which
are invisible and used as the origins for a cluster of \children points". The
algorithm is given a number of \children" and a maximum variation dis-
tance. For each \child point" a random azimuth and random distance is

74
used to determine the location of the `child point." This process is used
for Hopkins' indices between 0.5 and 1.0.
� Lattice process generates a nominally spaced grid of points with a random
start as the \parent points". These are used as the origin for the \children
points". Again the algorithm is given a number of \children" and the
maximum variation distance. Random azimuth and distance is determine
to locate the child point. The lattice process is used to generate patterns
with Hopkins' indices between 0.0 and 0.5.
Species { The species subroutine generates a species for each point in the previously
generated pattern. This is done randomly based on the relative proportion of
the species found in the original input data set.
Height { The initial heights are generated as random variates from a beta distri-
bution, with the parameter estimated from the input data. The range of the
generated data can be changed to allow for the generation of stands that di�er
in size from the input data.
Diameters { The diameters are generated using height/diameter equations inverted
to predict diameters from heights. An error component is added that mimics
the error in the original input data.
Crown Width { The crown width equations are exponential functions which use the
diameters to predict crown widths. An error component is added re ecting the
errors within the original data.
Crown Base { The height to crown base is a deterministic ratio between the width
and the length of the crown.

75
Foliage { The amount of foliage is determined as a ratio of the crown surface area to
foliage area. This ratio is currently assumed to be one, but in the near future
could be measured.
The \GROW" program takes a generated tree list or a previously grown tree list
and a set of tree parameters to grow the tree list through one or more time steps.
The program can start at any existing year in the list and project the list from that
time forward.
Height growth { The height growth subroutine predicts the change in height which
is added to the height at the previous time. The height change is a function of
the crown size and tree size at the previous time.
Crown change { The crown change subroutine �rst builds a cumulative leaf area pro-
�le of all competitors for each subject tree. From this information, a prediction
of the probability of the crown base receding is made; and the crown recedes if
a random uniform number drawn is greater than the probability. If the crown
recedes the process is iterated until the crown does not recede.
Diameter growth { The diameter growth of a tree is predicted as a function of the
crown size. For example, if the trees crown increases, diameter growth increases.
The area increment at the crown base is �rst predicted; then, the ratio between
the area increment at crown base and the area increment at breast height is
determined. The area increment at breast height is then converted into diameter
increment and added to the previous diameter.
Regeneration { The amount of regeneration is determined �rst by randomly drawing
a number of potential seedlings to test per unit area for the next time step. Each
seedling is assigned a coordinate, and the competition around that coordinate is

76
determined. The seedling is established and added to the tree list with a small
random height if the competition is less than the establishment threshold.
Mortality { Mortality is not explicitly modeled but mortality is determined by the
trees inability to grow because of loss of crown.
Utilities are a set of subroutines used to manipulate the tree list to simulate
stand treatments. These include thinning and pruning. The potential for a fertilizer
subroutine exists but is not implemented in the current work.
Thinning { This subroutine simply identi�es trees pointed to on a tree map and
removes them.
Prune { This subroutine removes the lower part of tree crowns in either variable or
�xed lifts.
The set parameter routines are used to take an input tree list and build the
parameters needed to run the model. These routines can be used with many types
of data or information. The parameters are in two major groups: the plot level
parameters and the species-speci�c tree level parameters. Plot level parameters relate
to the density or intensity, spatial pattern, species mix, type of measurement units
and time step length. Tree level parameters describe distributions of tree variables in
the original input data, growth rates of the input growth data, and assumed crown
recession parameters.
Hopkins { The Hopkins' index of non-randomness is used to determine the spatial
pattern of input data. This index was chosen because of the relative ease of
calculation and the logical range for ease of interpretation. The Hopkins' index
is determined for the input data and is used as a target for the generated spatial
patterns.

77
Set plot parameters { This routine queries the user for the general plot level param-
eters of the original input stand. It also queries for labeling and measurement
unit information.
Set species parameters { This routine steps the user through the setting of species
speci�c parameters. Parameters can set through assumptions while others are
�t to input data when ever possible. First, diameter/height equations are �t to
the input data for use by the generating functions. The relative abundance of
various species are noted; the height distribution and the distribution of errors
for each equation are estimated, with the beta distribution function using the
moments method. Next, the crown routine is called and the user is asked to
set a crown shape and width parameter visually for the species. Figure 5.1 is
an example of the visual screen that can be changed interactively. The height
growth routine is called next. It is a rather involved routine and is explained in
the next section. Then, a crown width relationship is �t to the input data for
use in stand generation. Finally, parameters that cannot be determined from
the input data are added with reasonable values to complete the parameter set.
This routine must be repeated for each species in the stand.
Set height growth parameters { The height growth functions are one of the key
functions within the model. The function �rst reads the input height/age data
for the species. These data can be from published height/age curves, stem
analysis data, or repeat measurement data from permanent plots. A guided
�tting technique is used because these data do not always extend over the
range of the time for which predictions will be made. The routine requires as
input height/age pairs and a target asymptote value. A exible cumulative
Weibull model is plotted with starting values �t to the data. The user can
vary the parameters until the �t \looks good". When the technique was tried,
statistically �t models worked well when complete or near complete data were

78
Distance (ft)
Hei
ght (
ft)
0 20 40 60
020
4060 Shape = 0.8
Crown Width = 0.66
Crown Length = 42
Figure 4.21: Example of the set crown parameter display. The user can interactivelyset the three parameters.

79
11
1
1
1
1
1
1
11
1
22
2
2
2
2
2
2
2
2
2
3
3
3 3
3
3
3
3
3
33
4
4
4
4
4
4
4
4
4
4
4
5
5
5
5
5
5
5
55
55
6
6
6
6
6
6
6
6
6
66
7
7
7
7
7
7
7
7
7
77
8
8
8
8
8
8
8
8
88
8
9
9
9
9
9
9
9
9
9
99
Height - Age
0 50 100 150
010
2030
40
Tree age (yr)
Hei
ght (
m)
Figure 4.22: Example of the set height parameter display. The number line are theheight age trajectories for nine trees and the solid line is the equation �t to thesedata.
available but were hard to guide to the desired asymptote when complete data
are not available.
Once the age/height curve is determined, its �rst derivative with respect to
height is calculated and the maximum found (see Figure 4.12). This maximum
is the potential height growth rate for the species on the given site. The data are
now divided at this maximum point and functions �t to the parts using a guided
technique. This technique allows use of surrogate variables to substitute for the
original age axis. This surrogate substitution is accomplished by expressing the
age as a ratio to the maximum rate age. The foliage ratio and size ratio are
then expressed within the same range.

80
Feet
0 20 40 60 80 100
020
4060
8010
0
Fee
t*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
***
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
*
* *
*
* **
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
**
**
*
**
**
*
*
*
*
*
*
*
* **
*
* *
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
*
***
*
Stand Map of Hugo Peak Stand at age 45
Figure 4.23: Example stand map from the growth model. The circle represent thecrown diameters.
Display is a set of routines used to display the output of a growth model run.
Some routines have been constructed to date; but, as stated earlier, others can be
constructed as well. The display scale units are in either English units (inches and
feet) or metric units (centimeters and meters).
Map { The map routine plots the x, y coordinates of each tree stem at a given time
and represents the diameter of the crown with a circle. This is in map view, or
from above the stand. Figure 5.3 is a sample output of this routine.
Identify map items { This routine allows the interactive identi�cation of trees within
the map. When using X-windows the user can point to trees and click. The

81
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 45
Figure 4.24. Example stand pro�le from the growth model.
tree number will appear near the tree. The numbers of all trees are printed if
the routine is run with other graphics interfaces.
Pro�le { Pro�le is a routine that will present a side or pro�le view of the stand from
either the x or y side and over any speci�ed range. Trees are plotted using a
tree outline de�ned in the set crown routine. Figure 5.4 is an example of the
stand pro�le plot.
Pro�les over time { This routine allows the user to view an individual tree over the
entire growth run. This view is useful to observe the history of height growth
and crown change visually. Figures 5.5 and 5.6 are examples of the pro�le over
time routine.

82
Hugo Peak Stand - Tree 188
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
Hugo Peak Stand - Tree 117
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
Figure 4.25. Example individual tree pro�les from the growth model.

83
Hugo Peak Stand - Tree 107
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
Figure 4.26. Example individual tree pro�le from the growth model.
Plot variables { This routine simply plots any variable in the tree list against any
other variable in the tree list. The routines allows the user the look at relation-
ships between variables such diameter and height. Figure 5.7 is an example of
the variable plotting function.
Plot variable over time { This routine produces a time trace for each tree of the
variable requested. Figure 5.8 is an example plot over time.
Histogram variables { This routine plots a histogram of the speci�ed variable at the
speci�ed time. Figure 5.9 is an example of the histogram.
Plot leaf competition { This routine displays the cumulative leaf area index pro�le
used to predict crown recession and the current tree pro�le. If a user steps
through the time steps or tree numbers, they can see how the crown of an
individual tree interacts with the leaf area of the competitors. Figure 5.10 is an

84
*
**
*
**
* *
*
*
**
* *
**
*
*
* *
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
**
**
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
**
**
*
*
*
*
*
*
**
**
*
*
*
*
*
*
**
*
*
*
*
*
* *
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
**
*
*
**
*
*
*
*
*
**
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
**
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
**
*
* *
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
**
***
*
*
**
*
*
**
*
*
**
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
* *
*
*
*
** **
*
*
**
*
**
* *
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
**
*
Diameter at breast height (in)
Hei
ght (
ft)
0 2 4 6 8 10 12
020
4060
80
Hugo Peak Stand 28 - Diameter vs. Height
Figure 4.27. Example plot of diameter versus height.

85
Time (5 yr)
Hei
ght (
ft)
2 4 6 8 10 12
020
4060
8010
0
Hugo Peak Stand - Height vs. Time
Figure 4.28. Example plot of individual tree heights over time.

86
20 40 60 80
05
1015
20
Height (ft)
Num
ber
Hugo Peak Stand 28 - Height
Figure 4.29. Example histogram of heights.

87
Cumulative LAI in competitors (sq ft)
Hei
ght (
ft)
0.0 0.5 1.0 1.5
010
2030
4050
Distance (ft)
Hei
ght (
ft)
-5 0 5 10
010
2030
4050
LAI profile of tree 386 year 1
Figure 4.30: Example cumulative leaf area index pro�le. The x axis on the left plotis the cumulative leaf area index (LAI). The x axis on the right plot is distance.
example of the plot of leaf area competition.
Plot variable rank order { This routine displays an ordered plot of the trees in a
stand versus a variable. The order variable need not be the plotted variable.
As described earlier, this plot is one way of looking at stand structure. Figure
5.11 is an example of ordered plots of variables.
Density management space { The density management relation is a common tool
used by foresters to evaluate the condition of a stand and the change within the
stand. This routine allows the plotting the relation of trees per unit area and
the quadratic mean diameter of the stand. Maximum lines are species speci�c
and so are not routinely plotted; however, a line of speci�ed stand density index

88
Relative Rank
Hei
ght (
ft)
0.0 0.2 0.4 0.6 0.8 1.0
020
4060
Hugo Peak Stand 28 - Height ordered by height
Figure 4.31. Example plot of rank order of the heights.

89
*
**
**
**
*****
********
Trees per acre
Qua
drat
ic M
ean
Dia
met
er
1 10 100 1000
15
1050
100
Hugo Peak Stand 28 - Density Management Diagram
Figure 4.32: Example size{density plot over time. The point represent the locationof the stand a successive points in time. The solid line is an assumed maximum forDouglas-�r of SDI 595.
can be overlayed to provide a reference point as done in Figure 5.12. Figure
5.12 is the track of a stand of Douglas-�r (Pseudotsuga menziesii) and the stand
density index (SDI) 595 (The assumed maximum for that species (Drew and
Flewelling, 1979)) for reference. Points indicate locations of the stand at each
�ve year time step.
Structure plots { A set of multi-plots allow many plots to be viewed at various times.
These include change in a set of reverse ordered tree heights with time, the
change in spatial index over time, change in species mix over time, and change in
the height distribution over time. In Figure 5.13 a few of the largest tree at time

90
one remain the largest tree though out the model run. Figure 5.14 illustrates
a large range in the density of the stand; however, the spatial character as
described by the Hopkins' index does not change. Figure 5.15 illustrates the
relationship among the three species in the stand. In this particular run only one
species Douglas-�r (df) survived to the end of the run. Figure 5.16 illustrates
the height distributions for the stand.
Tree means { This is a non-graphical routine to report the means and variances for
all the tree list variables.
One feature of the current model is the method of conveying the output to a user.
The method is based on the assumption that graphical information can be recognized
and comprehended faster than tabular information. There is a three layered approach
to model output display. The �rst level is a \picture" of the stand at the various time
steps. The second level is traditional plot displays such as scatter plots, histograms,
and function plots. Thirdly, the tabulated data are available for question, which arise
from examination of the graphical data.
4.4.4 Description of implementation
This model building e�ort has been an evolution through several computational plat-
forms. Initially the programming was implemented in the C++ programming lan-
guage on a IBM compatible computer. This program worked well in some respects
but, had drawbacks. The �rst problem was to build a graphical interface for the
display of the model output. Using a low level graphics library supplied with the
compiler. Implementation of the program was straight-forward and the concept of
classes the C++ language make the building of the routines relatively fast and easy.
This programming approach was abandoned for three reasons. First, the implementa-
tion grew and the programs became di�cult to debug because of the memory required
by both the debugging software and the program. As the complexity of particular

91
..............................................................................................................................................................................................................................................................................................................................................................................................................................
0100
300
0 20 40
Tim
e 1
.... .. ..... . .... ... .. ... .. ... ..... . .. ....... ... . ... .. .. .. .. ... .. .. ... ..... .. ... ... .. ... ... .......... .... ... ..... .... .... ... ......... . ....... ..... ...... ...... ..... .... .... ..... .. .... .. ... .. ... ... ....... ........ ..................... ....... ......... ....... ... ... ............... ... ... ........ ... ......... .......... ...... ..... ...................... ..... ...... .. ..... ....... ........ .... ........................... .......
0100
3000 20 50
Tim
e 2
.... .. .. ... . .... ... .. ... .. ... .. ... ... ....... ... .... .. .. .. .. ... .. ..... ....... ... ... .. .. . ... .... .. .... .... ... ... .. .... .. .. .. . . ... ..... . .... ... .. ... ... ... ..... .. .... .. .. ... . ...... .. .. . .. ... .. . .. ..... .. ... .. . .. ... ... ... .. .. .. ..... .. ..... ... .. ... .. .... .. ....... .. . .. . ...... ... .. .... ... .... ... . ... ..... .. ... ........ ..... .. .. ... . ..... .... .. ....... ... ... .. .... . ... .. ... .. ...... ........ ....... ........ ............. .........
0100
300
0 20 60
Tim
e 3
.. .. .... .. .. .... ... .. ... ..... .. .. . ... . ... .. . .. .. . .. .... . . ... .. .. .. . .. .... . .... . ... .. .. .... . ... .. .. .. .... .. .. .. .. .. .... .. .. .. .. .... ... .... . ... ... .. .. ... ..... .. .... .. .. ... . . .. ... .. .. . .. ... .. . .. ..... .. ... .. . .. . .. . .. ... .. .. ....... .. . .... ... .. ... .. .... .. ... ..... .. .. . ...... ..... ... .... .... ... .... .. ... .. ... ... ..... ..... .... ... . ......... .. ....... ... ..... ..... ... .. ..... ...... . ... ... . ........................... ..........
0100
300
0 40
Tim
e 4
.. . . .. .. .... ... . .. . .. .. . ..... .. .. . ... .... ... .. .. . .. ... . .. ... .. .. ... .. .... . ... . . ..... .. .... . ... . ... ...... .. .. .. .. .. . ... .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .... . .. . . .. ... .. .. . .. ... .. . .. .. ... .. ... .. . .. . .. . .. ... .. .. .. ..... .. . .... ... .. ... .. .... .. ... ..... .. .. . ...... ..... ... ....... . ... .... . . ... .. ... ... ..... ..... .... .... ..... .... .. ....... ... ..... ..... ... .. ... .. .. .... . ... .... .............................. .......
0100
300
0 40 80
Tim
e 5
.. . . ...... .. ... . ... .. .. . ... .. .... . ... .... .. . .. .. . .. ... . . . ... .. .. ... .. .... . ... . . ..... .. . ... . ... . ... . ..... .. .. .. .. .. . ... .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... .. . .. .. ... .. ... .. . .. . .. . .. ... .. .. .. ..... .. . .... ... .. ... .. .... .. . .. ..... .. .. . ...... ... .. ... ........ ... .... .. ... .. ........... ..... .... .... ..... .... .. ....... ... ..... ..... ... .. ..... ...... . ... .... ........... ................... .......
0100
300
0 40 80
Tim
e 6
.. . . ...... .. .... .. . .. .. . .. . .. .. .. . ... .... .. . .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . .... ... . ... . ... . ... . ..... .. .. .. .. .. . ... .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .... . .. . . .. ... .. .. . .. ... ... .. .. ... .. ... .. . .. . .. . .. ... .. .. ....... .. . .... ... .. ... .. .... .. . .. ..... .. .. . ...... ..... ... . ... .... ... .... .. ... ... .......... ..... .... ... . ..... .... .. ....... ... ..... ..... ... .. ..... .. ... .. ... .... .............................. .......
0100
300
0 40 80
Tim
e 7
.. . . ...... .. .... .. . .. .. . .. ... .. .. . ... . ... ... .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . .... ... . ... . ... . ... . ..... .. .. .. .. .. . ... .. .. .. .. .... . .. .... . ... ... .. .. ... .... . .. .... .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ... .. . .. . .. . .. ... .. .. .. ..... .. . .... ... .. ... .. .... .. . .. .... . .. .. . ...... ..... ... ........ ... .... .. ... .. ........... ..... .... ... . ..... .... .. ....... ... ... .. ..... ... .. ... .. ...... . ... ... . ....... ..............................
0100
300
0 40 80
Tim
e 8
.. . .... ... .. .... .. ... ... .. ... .. .. .... . ... ... .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . .... ... . ... . ... . ... . ..... .. .. .. .. .. . ... .. .. .. .. .... . .. ... . . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... .. . .. ..... .. ..... . .. . .. . .. ... .. .. ....... .. . .... ... .. ... .. .... .. . .. .... . .. .. . ...... ..... ... ........ ... .... .. ... ... .......... ..... .... ... ......... ... ....... ... ..... ..... ... .. ... .. ....... ... ... . ........... ................... .......
0100
300
0 40 80
Tim
e 9
.. . . .. . ... . . .... .. ... ... .. . .. .... .. .. . ... ... .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . .... ... . ... . ... . ... . ..... .. .. .. .. .. . ... .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ..... . .. . .. . .. ... .. .. ....... .. . .... ... .. ... .. .... .. . .. ..... .. .. . ...... ..... ... .... .... ... .... .. ... .. ........... ..... .... .... ......... .. ....... ... ..... ..... ... .. ... .. ....... ... .... .....................................
0100
300
0 40 80
Tim
e 10
.. . . .. .. .... . ... .. ... .. . .. . .. . ... .. .. . ... ... .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . ... .. .. .. .. .... . .. ... . . ... ... .. .. ... ..... .. .. .. .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ..... . .. . .. . .. ... .. .. ....... .. . .... ... .. ... .. ... ... . .. ..... .. ... ...... ..... ... ........ ... .... .. ... .. ...... ..... ..... .... ... . ......... .. ....... ... ..... ..... ... .. ... .. ...... . ... .... ... ............ ............... .......
Relative ht
0100
300
0 40 80
Tim
e 11
.. . . .. . ... ... ... .. . .. .. . .. ... . ... .. ... ... ... .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ..... . .. . .. . .. ... .. .. .. ....... ..... ..... ... .. .... .. . .. .... . .. .. . ...... ..... ... ....... . ... .... .. ... .. ...... ..... ..... .... ... . ..... .... .. ....... ... ..... ..... ... .. ... .. ...... . ... .... ....... .... ..........................
0100
300
0 40 80
Tim
e 12
.. . . .. .... .. . ... .. ... .. . .. ... .. .. .. ... ... ... .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... .. . .. ..... .. ... .. . .. . .. . .. ... .. .. .......... .... ... .. ... .. .... .. . ...... . .. .. . ...... ..... ... ........ ... .... .. ... .. ...... ..... ..... .... ... . ......... .. ....... ... ..... .... . ... .. .......... .. ...... . .............................. .......
0100
300
0 40 80
Tim
e 13
.. . . .. ...... . ... .. ... .. . .. ... . ... ..... ... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... .... . .. .... .... . .. . . .. ... .. .. . .. ... .. . .. .. ... .. ... .. . .. . .. . .. ... .. .. .. ..... .. . .... ... .. ... .. ... ... . .. .... . .. .. . ...... ..... ... ........ ... .... .. ... ........ ..... ..... .... .... ........ ... ....... ... ... .. ..... ... .. ... .. .. ..... ... .... ....... .... ................ ..........
0100
300
0 40 100
Tim
e 14
.. . . .. .... .. . ... .. . .. .. . .. ... . ... . .... ... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .. .. .... . .. . . .. ... .. .. . .. ... ... .. .. ... .. ... .. . .. . .. . .. ... .. .. .. ..... .. . .... ... .. ... .. ... ... . .. .... . .. .. . ...... ..... ... ....... . ... .... .. ... ............. ..... .... .... ..... ...... ....... ... ..... ..... ... .. ..... ..... .. ...... . .....................................
0100
300
0 40 100
Tim
e 15
.. . . .. .... .. . ... .. ... .. . .. ... .... .. ... ... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ... .. . .. . .. . .. ... .. .. .. ........ .... ... .. ... .. .... .. . ...... . .. .. . ...... ..... ... . ....... ... .... .. ... .. ........... ..... ....... .......... .. ....... ... ..... ..... ... .. ..... .. ..... ... .... .....................................
0100
300
0 40 100
Tim
e 16
.. .. .. . ... .. . ... .. ... .. . .. ... .. .. .. ... ... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ... .. . .. . .. . .. ... .. .. .......... .... ... .. ... .. .... .. . ....... .. .. . ...... ..... ... ........ ... .... . . ... .. ...... ..... ..... .... .... ........... ... .... ... ..... ..... ... .. ..... ..... .. ....... ... ........................... .......
0100
300
0 40 100
Tim
e 17
.. .. .. . ... ... ... .. . .. .. . .. ... .. .. .. ... ... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... .. . .. .. ... .. ..... . .. . .. . .. ... .. .. ....... .. ..... ... .. ... .. .... .. . .. ..... .. ... ...... ..... ... ....... . ... .... .. ... ... .......... ..... ....... .......... .. .......... ... .. ..... ... .. ... .. .. .... . ... .... ........................... ..........
Rank
0100
300
0 40 100
Tim
e 18
.... .. . ...... ... .. ... .. . .. ... .. .. .. ...... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . .... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ..... . .. . .. . .. ... .. .. ....... .. . .... ... .. ... .. ... ... . .. .... . .. .. . ...... ..... .... ....... ... .... .. ... .. ...... ..... ..... .... .... ........... ....... ... ..... .... . ... .. ............ ... .... ............................ .........
0100
300
0 40 100
Tim
e 19
.... .. . ... .. . ... .. ... .. . .. ... .. .. .. ... ... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . .... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .. .. .... . .. . . .. ... .. .. . .. ... ... .. ..... .. ..... . .. . .. . .. ... .. .. .. ........ .... ..... ... .. .... .. . ....... .. ... ......... .. ... ........ ... .... .. ... ............. ..... ....... .......... .. ....... ... ..... ..... ... .. ..... ..... .. ....... ....... ..............................
0100
300
0 40 100
Tim
e 20
Figu
re4.33:
Exam
ple
oford
eredheigh
tsplot
overtim
e.Thexaxisisthereverse
ofsize
rank(th
elargest
treeis1)
andtheyaxisistherelative
heigh
tpercen
t(th
elargest
treeis100).
Tree
ranksare
based
ontim
e1;
they
change
order
inthexaxis
with
time.

92
*
*
**
** *
** *
*
**
**
* *
*
*
*
*
*
*
**
**
**
*
*
*
*
*
**
**
*
*
*****
****
**
* *
*
*
*
*
*
**
*
*
*
**
**
*
*
* * **
*
*
* **
**
*
*
**
*
*
**
*
*
**
*
*
**
**
*
**
*
*
*
*
*
*
*
**
**
***
***
*
*
*
*
*
*
* **
**
* *
**
*
*
*
*
**
*
*
**
*
* *
*
*
*
* *
*
**
***
*
*
*
*
*
*
*
*
*
* *
**
* *
*
*
*
**
*
*
*
**
*
*
**
*
*
* *
*
** *
*
*
* *****
*
*
**
** ***
**
***
*
*
*
**
* *
*
*
*
***
**
*
* *
* *
**
**
*
**
* **
*
* *
*
*
*
**
****
*
*
*
*
*
* *
*
* **
*
*** *
*
*
*
*
*
*
**
*
*
*
**
*
**
* **
**
*
* ***
*
*
***
**
*
****
*
**
** ** *
**
*
**
**
**
****
*
**
**
*
*
*
* **
**
*
*
*
*
**
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
***
*
**
*
*
* **
*
*
*
**
*
*
***
*
**
* *
**
*
*
**
* *
* *
*
*
*
**
***
*
*
0 40 80
040
80F
eet
Feet
Hopkins = 0.1839014
Time 1
*
*
****
**
**
*
*
*
*
*
*
*
* *
**
*
*****
**
** *
**
*
*
*
**
*
*
*
* *
* **
*
*
*
**
*
*
**
*
**
*
*
*
**
*
*
*
*
*
*
***
* **
* **
*
**
*
*
*
*
*
* *
*
*
*
*
**
*
*
**
*
*
**
*
*
*
*
*
*
*
**
***
*
***
*
* **
**
***
*
*
*
*
***
*
*
*
* *
***
*
***
*
**
***
*
*
*
*
** *
*
*
*
*
**
*
*
*
*
* **
**
* ***
*
**
****
*
***
**
** *
**
***
*
*
*
*
**
*
*
**
*
* *
*
*
*
*
*
*
*
*
*
* **
*
*
*
***
**
*
*
* **
*
* *
* *
*
*
**
**
*
*
0 40 80
040
80
Hopkins = 0.1983205
Time 2
***
***
*
*
*
*
* *
*
****
**
* ****
**
*
*
*
* **
*
**
*
**
**
*
* **
*
*
*
*
**
*
* * *
**
*
*
*
*
*
*
*
*
**
*
*
** *
**
*
*
*
*
*
***
**
**
**
***
*
*
*
***
*
*
*
* *
***
*
**
*
**
* *
*
*
*
*
**
*
*
*
*
**
**
*
*
* ***
*
**
****
*
**
**
****
**
*
**
**
*
**
*
* *
*
*
** *
*
*
* *
*
*
*
**
**
*
**
*
*
*
**
*
0 40 80
040
80
Hopkins = 0.1945829
Time 3
*
***
*
*
* *****
*
****
**
*
* **
*
*
* ** * **
**
**
*
*
**
*
*
**
*
*
**
*
* *
**
*
*
****
**
**
***
*
*
*
**
*
*
*
* *
* *
**
*
**
*
* *
*
**
*
*
*
**
**
*
***
**
***
*
**
**
* **
**
**
*
*
**
***
*
*
* *
*
*
**
*
*
*
* **
0 40 80
040
80
Hopkins = 0.1963809
Time 4
*
*
*
* ***
*
*
****
*
*
* *
*
*
* * * **
**
*
*
*
*
*
**
**
**
**
*
****
**
***
*
*
*
***
* *
*
**
*
** *
*
** *
*
**
**
*
***
***
*
*
**
**
* ***
**
*
*
**
**
*
**
*
*
**
*
**
0 40 80
040
80
Hopkins = 0.2062234
Time 5
*
*
* ***
*
*
****
*
* *
** * **
**
*
*
*
**
**
**
**
*
***
**
***
*
*
*
***
* *
*
**
*
**
*
** *
**
**
*
***
***
** * *
**
*
*
**
**
*
**
*
*
**
**
0 40 80
040
80
Hopkins = 0.1889682
Time 6
*
*
* ***
*
*
****
* *
** * *
**
*
*
*
**
**
**
**
*
*
*
**
***
*
*
***
*
**
*
*
*
** *
**
**
*
***
**
** *
**
*
*
*
***
* *
**
**
0 40 80
040
80
Hopkins = 0.1966938
Time 7
** *
**
*
*
****
*
**
**
*
*
*
**
**
**
**
*
*
*
**
**
*
*
*
*
*
**
*
*
** *
**
**
*
***
**
**
** ***
* *
***
0 40 80
040
80
Hopkins = 0.1935857
Time 8
** *
**
*
*
**
*
*
**
**
*
*
*
**
**
*
**
*
*
*
**
**
*
*
*
* **
*
*
** *
*
**
*
***
**
**
** ** *
***
0 40 80
040
80
Hopkins = 0.1941776
Time 9
** *
*
*
*
**
*
*
**
**
*
*
*
**
**
**
*
*
*
**
**
*
*
*
* **
*
*
** *
**
*
***
**
**
* ** *
**
0 40 80
040
80
Hopkins = 0.1770236
Time 10
** *
*
*
*
**
*
*
**
*
*
*
*
**
**
**
*
*
*
**
**
*
*
* **
*
*
** *
**
*
***
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.2111597
Time 11
** *
*
*
*
**
*
*
**
*
*
*
*
**
**
**
*
*
*
**
**
*
*
* **
*
*
** *
****
*
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.1850428
Time 12
**
*
*
**
*
*
**
*
*
*
*
**
**
**
*
*
*
**
**
*
*
* **
*
*
** *
****
*
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.1952518
Time 13
**
*
*
**
*
*
**
*
*
*
*
**
**
**
*
*
*
**
**
*
*
* **
*
*
** *
****
*
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.2009852
Time 14
**
*
*
**
*
*
**
*
**
**
**
**
*
*
*
**
**
*
*
* **
*
*
**
****
*
**
*
* ** *
**
0 40 80
040
80Hopkins = 0.1879953
Time 15
**
*
*
**
*
*
**
*
**
**
**
**
*
*
*
**
**
*
*
* **
*
*
**
****
*
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.1777561
Time 16
**
*
*
**
*
*
**
*
**
**
**
**
*
*
*
**
**
*
*
* **
*
**
****
*
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.1806601
Time 17
**
*
*
**
*
*
**
*
**
**
**
**
*
*
*
**
**
*
*
* **
*
**
****
*
**
*
** *
**
0 40 80
040
80
Hopkins = 0.1916783
Time 18
**
*
*
**
*
*
**
*
**
**
**
**
*
*
*
**
**
*
*
* **
*
**
****
*
**
*
** *
**
0 40 80
040
80
Hopkins = 0.1994442
Time 19
**
*
*
**
*
*
**
*
**
**
**
**
*
*
*
**
**
*
*
* **
*
**
****
*
**
*
** *
**
0 40 80
040
80
Hopkins = 0.2002466
Time 20
Figure 4.34: Example of spatial pattern plot over time. The Hopkins' index for eachpattern is printed under the map plot.

93
df wh ch
020
0
Time 1
df wh ch
010
020
0
Time 2
df wh ch
050
150
Time 3
df wh ch
040
100
Time 4
df wh ch
040
80
Time 5
df wh ch
040
80
Time 6
df wh ch
040
80
Time 7
df wh ch
020
60
Time 8
df wh ch
020
50
Time 9
df wh ch
020
40
Time 10
df wh ch
020
40
Time 11
df wh ch
020
40
Time 12
df wh ch
020
40
Time 13
df wh ch
020
40
Time 14
df wh ch
020
40
Time 15
df wh ch
020
40
Time 16
df wh ch
020
40
Time 17
df wh ch
020
40
Time 18
df wh ch
020
40
Time 19
df wh ch
020
40
Time 20
Figure 4.35: Example of species barchart plot over time. The x axis are the variousspecies found on the plot and the y axis is the number of individual in each species.

94
0 20 40
020
60
Height (ft)
Num
ber
Time 1
10 30 50
020
Height (ft)
Num
ber
Time 2
10 30 50 70
010
25
Height (ft)
Num
ber
Time 3
10 30 50 70
05
10
Height (ft)
Num
ber
Time 4
20 40 60 80
05
15
Height (ft)
Num
ber
Time 5
20 40 60 80
05
15
Height (ft)
Num
ber
Time 6
40 60 80
05
10
Height (ft)
Num
ber
Time 7
60 70 80 90
05
15
Height (ft)
Num
ber
Time 8
80 90 100
04
8
Height (ft)N
umbe
r
Time 9
85 95 105
04
8
Height (ft)
Num
ber
Time 10
90 100 110
04
812
Height (ft)
Num
ber
Time 11
95 105 115
04
812
Height (ft)
Num
ber
Time 12
100 110 120
05
1015
Height (ft)
Num
ber
Time 13
105 115 125
05
10
Height (ft)
Num
ber
Time 14
110 120 130
04
812
Height (ft)N
umbe
r
Time 15
115 125
04
812
Height (ft)
Num
ber
Time 16
120 130
04
812
Height (ft)
Num
ber
Time 17
125 135
04
812
Height (ft)
Num
ber
Time 18
130 140
04
812
Height (ft)
Num
ber
Time 19
130 140
05
1015
Height (ft)
Num
ber
Time 20
Figure 4.36: Example of histogram of heights plotted over time. The x axis is theheight of the height classes in feet and the y axis is the number of individuals in eachclass.

95
programs increased, the amount of time to debug the program increased exponen-
tially. The size of the programs and the run times on the 386 system became large
and long, respectively. The models were converted to run on a Sun Sparcstation 1.
The DOS software was left at the state of development at the time of the conversion.
The goal is to demonstrate the viability of the approach and not to produce a
commercial product. Additionally, no C++ compilation software was available on the
Sun Sparcstation used. The S statistical package provided all the graphics capability
required and the exibility to implement the model through S user functions, linked
C functions, and stand-alone C programs.
This approach provided a good development environment; for example, a function
or routine that would take a week to code in C or C++ would take about a day in
S. The conversion from C++ version to the S version took approximately a month.
The S version of the model is fragile, requiring a knowledge of S and Unix. One very
nice feature of S is that it is an interactive environment; if the user would like to plot
data in a di�erent way, it is a relatively easy task to write a new function to make
the plot. S is one of several interactive packages that would be suitable for this type
of work. The real advantage is that the package handles the graphics user interface,
allowing the modeler to concentrate on the relationships and behavior of the model.
To implement a system based on graphical displays, a good graphical user interface
is needed. In the current project both interfaces developed from a low level routines
and interfaces developed from high level routine were used. Both system seem to
work well within the limitation of their respective computer. They provide a easy
method to view the output from the growth model and an easy method to visualize
the predictions.

Chapter 5
EXAMPLE SYSTEM
\I cannot really imagine any other law of thought than that our pictures
should be clearly and unambiguously imaginable."
L. Boltzmann 1897
5.1 Objective
This chapter presents the detail of an adaptive stand dynamics model. There are many
ways to implement these ideas, some that work better than others. This dissertation
presents the ideas with the intent of conveying the concepts, rather than the details
of a particular implementation.
5.2 Design
This model is designed to use a \Run and Display" approach as opposed to a \Display
as you Run" approach. The growth submodel is \Run," and output is stored in an
output �le or data structure. Then, a separate display part of the model will read
that �le or data structure. This approach has several advantages. Any time step
in the growth sequence can be easily viewed; plots over time are easily made; and,
when many batch runs are needed this task is much easier. In \Display as You
Run" approach, the program plots the current values of the projected tree list. The
run usually must be reinitiated and rerun to show previous states of the stand; and
over-time plots are not available until the end of the run.

97
This model uses a simulation approach; input tree data are distilled into a set of
parameters that describe a stand. A simulated stand is then generated from these
parameter sets and used as the initial condition for model runs. Because of the
stochastic nature of the models, the output will di�er with each run with identical
input. If averages are desired, several runs must be averaged to obtain estimates of
the predicted mean and range.
5.3 Components of the models
The \GENERATE" program is a set of routines that produces a tree list from a
parameter set. As stated in chapter 4 the program steps through the routine in the
following order.
The function to generate the random numbers used in the C routines is the \ran3"
function from Numerical Recipes (Press et al., 1988), and inside S the \runif" function
is used.
Pattern { The �rst step is to generate a spatial pattern for the simulated stand.
One of two generating processes is used. If the stand is natural in origin then a
Neyman-Scott process is used; otherwise a lattice processe is used.
� Neyman-Scott process generates a pattern of random \parent points" which
are invisible and used as the origins for a cluster of \children points". The
algorithm is given a number of \children" and a maximum variation dis-
tance. For each \child point" a random azimuth and random distance is
used to determine the location of the `child point." This process is used
for Hopkins' indices between 0.5 and 1.0.
� Lattice process generates a nominally spaced grid of points with a random
start as the \parent points". These are used as the origin for the \children
points". Again the algorithm is given a number of \children" and the

98
maximum variation distance. Random azimuth and distance is determine
to locate the child point. The lattice process is used to generate patterns
with Hopkins' indices between 0.0 and 0.5.
Species { The species subroutine generates a species for each point in the previously
generated pattern. This is done randomly based on the relative proportion of
the species found in the original input data set.
Height { The initial heights are generated as random variates from a beta distri-
bution, with the parameter estimated from the input data. The range of the
generated data can be changed to allow for the generation of stands that di�er
in size from the input data.
Diameters { The diameters are generated using height/diameter equations inverted
to predict diameters from heights. An error component is added that mimics
the error in the original input data.
Crown Width { The crown width equations are exponential functions which use the
diameters to predict crown widths. An error component is added re ecting the
errors within the original data.
Crown Base { The height to crown base is a deterministic ratio between the width
and the length of the crown.
Foliage { The amount of foliage is determined as a ratio of the crown surface area to
foliage area. This ratio is currently assumed to be one, but in the near future
could be measured.
The \GROW" program takes a generated tree list or a previously grown tree list
and a set of tree parameters to grow the tree list through one or more time steps.

99
The program can start at any existing year in the list and project the list from that
time forward.
Height growth { The height growth subroutine predicts the change in height which
is added to the height at the previous time. The height change is a function of
the crown size and tree size at the previous time.
Crown change { The crown change subroutine �rst builds a cumulative leaf area pro-
�le of all competitors for each subject tree. From this information, a prediction
of the probability of the crown base receding is made; and the crown recedes if
a random uniform number drawn is greater than the probability. If the crown
recedes the process is iterated until the crown does not recede.
Diameter growth { The diameter growth of a tree is predicted as a function of the
crown size. For example, if the trees crown increases, diameter growth increases.
The area increment at the crown base is �rst predicted; then, the ratio between
the area increment at crown base and the area increment at breast height is
determined. The area increment at breast height is then converted into diameter
increment and added to the previous diameter.
Regeneration { The amount of regeneration is determined �rst by randomly drawing
a number of potential seedlings to test per unit area for the next time step. Each
seedling is assigned a coordinate, and the competition around that coordinate is
determined. The seedling is established and added to the tree list with a small
random height if the competition is less than the establishment threshold.
Mortality { Mortality is not explicitly modeled but mortality is determined by the
trees inability to grow because of loss of crown.
Utilities are a set of subroutines used to manipulate the tree list to simulate

100
stand treatments. These include thinning and pruning. The potential for a fertilizer
subroutine exists but is not implemented in the current work.
Thinning { This subroutine simply identi�es trees pointed to on a tree map and
removes them.
Prune { This subroutine removes the lower part of tree crowns in either variable or
�xed lifts.
The set parameter routines are used to take an input tree list and build the
parameters needed to run the model. These routines can be used with many types
of data or information. The parameters are in two major groups: the plot level
parameters and the species-speci�c tree level parameters. Plot level parameters relate
to the density or intensity, spatial pattern, species mix, type of measurement units
and time step length. Tree level parameters describe distributions of tree variables in
the original input data, growth rates of the input growth data, and assumed crown
recession parameters.
Hopkins { The Hopkins' index of non-randomness is used to determine the spatial
pattern of input data. This index was chosen because of the relative ease of
calculation and the logical range for ease of interpretation. The Hopkins' index
is determined for the input data and is used as a target for the generated spatial
patterns.
Set plot parameters { This routine queries the user for the general plot level param-
eters of the original input stand. It also queries for labeling and measurement
unit information.
Set species parameters { This routine steps the user through the setting of species
speci�c parameters. Parameters can set through assumptions while others are

101
�t to input data when ever possible. First, diameter/height equations are �t to
the input data for use by the generating functions. The relative abundance of
various species are noted; the height distribution and the distribution of errors
for each equation are estimated, with the beta distribution function using the
moments method. Next, the crown routine is called and the user is asked to
set a crown shape and width parameter visually for the species. Figure 5.1 is
an example of the visual screen that can be changed interactively. The height
growth routine is called next. It is a rather involved routine and is explained in
the next section. Then, a crown width relationship is �t to the input data for
use in stand generation. Finally, parameters that cannot be determined from
the input data are added with reasonable values to complete the parameter set.
This routine must be repeated for each species in the stand.
Set height growth parameters { The height growth functions are one of the key
functions within the model. The function �rst reads the input height/age data
for the species. These data can be from published height/age curves, stem
analysis data, or repeat measurement data from permanent plots. A guided
�tting technique is used because these data do not always extend over the
range of the time for which predictions will be made. The routine requires as
input height/age pairs and a target asymptote value. A exible cumulative
Weibull model is plotted with starting values �t to the data. The user can
vary the parameters until the �t \looks good". When the technique was tried,
statistically �t models worked well when complete or near complete data were
available but were hard to guide to the desired asymptote when complete data
are not available.
Once the age/height curve is determined, its �rst derivative with respect to
height is calculated and the maximum found (see Figure 4.12). This maximum
is the potential height growth rate for the species on the given site. The data are

102
Distance (ft)
Hei
ght (
ft)
0 20 40 60
020
4060 Shape = 0.8
Crown Width = 0.66
Crown Length = 42
Figure 5.1: Example of the set crown parameter display. The user can interactivelyset the three parameters.

103
11
1
1
1
1
1
1
11
1
22
2
2
2
2
2
2
2
2
2
3
3
3 3
3
3
3
3
3
33
4
4
4
4
4
4
4
4
4
4
4
5
5
5
5
5
5
5
55
55
6
6
6
6
6
6
6
6
6
66
7
7
7
7
7
7
7
7
7
77
8
8
8
8
8
8
8
8
88
8
9
9
9
9
9
9
9
9
9
99
Height - Age
0 50 100 150
010
2030
40
Tree age (yr)
Hei
ght (
m)
Figure 5.2: Example of the set height parameter display. The number line are theheight age trajectories for nine trees and the solid line is the equation �t to thesedata.

104
now divided at this maximum point and functions �t to the parts using a guided
technique. This technique allows use of surrogate variables to substitute for the
original age axis. This surrogate substitution is accomplished by expressing the
age as a ratio to the maximum rate age. The foliage ratio and size ratio are
then expressed within the same range.
Display is a set of routines used to display the output of a growth model run.
Some routines have been constructed to date; but, as stated earlier, others can be
constructed as well. The display scale units are in either English units (inches and
feet) or metric units (centimeters and meters).
Map { The map routine plots the x, y coordinates of each tree stem at a given time
and represents the diameter of the crown with a circle. This is in map view, or
from above the stand. Figure 5.3 is a sample output of this routine.
Identify map items { This routine allows the interactive identi�cation of trees within
the map. When using X-windows the user can point to trees and click. The
tree number will appear near the tree. The numbers of all trees are printed if
the routine is run with other graphics interfaces.
Pro�le { Pro�le is a routine that will present a side or pro�le view of the stand from
either the x or y side and over any speci�ed range. Trees are plotted using a
tree outline de�ned in the set crown routine. Figure 5.4 is an example of the
stand pro�le plot.
Pro�les over time { This routine allows the user to view an individual tree over the
entire growth run. This view is useful to observe the history of height growth
and crown change visually. Figures 5.5 and 5.6 are examples of the pro�le over
time routine.

105
Feet
0 20 40 60 80 100
020
4060
8010
0
Fee
t
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
***
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
*
* *
*
* **
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
**
**
*
**
**
*
*
*
*
*
*
*
* **
*
* *
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
*
***
*
Stand Map of Hugo Peak Stand at age 45
Figure 5.3: Example stand map from the growth model. The circle represent thecrown diameters.

106
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 45
Figure 5.4. Example stand pro�le from the growth model.

107
Hugo Peak Stand - Tree 188
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
Hugo Peak Stand - Tree 117
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
Figure 5.5. Example individual tree pro�les from the growth model.

108
Hugo Peak Stand - Tree 107
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
Figure 5.6. Example individual tree pro�le from the growth model.
Plot variables { This routine simply plots any variable in the tree list against any
other variable in the tree list. The routines allows the user the look at relation-
ships between variables such diameter and height. Figure 5.7 is an example of
the variable plotting function.
Plot variable over time { This routine produces a time trace for each tree of the
variable requested. Figure 5.8 is an example plot over time.
Histogram variables { This routine plots a histogram of the speci�ed variable at the
speci�ed time. Figure 5.9 is an example of the histogram.
Plot leaf competition { This routine displays the cumulative leaf area index pro�le
used to predict crown recession and the current tree pro�le. If a user steps
through the time steps or tree numbers, they can see how the crown of an
individual tree interacts with the leaf area of the competitors. Figure 5.10 is an

109
*
**
*
**
* *
*
*
**
* *
**
*
*
* *
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
**
**
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
**
**
*
*
*
*
*
*
**
**
*
*
*
*
*
*
**
*
*
*
*
*
* *
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
**
*
*
**
*
*
*
*
*
**
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
**
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
**
*
* *
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
**
***
*
*
**
*
*
**
*
*
**
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
* *
*
*
*
** **
*
*
**
*
**
* *
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
**
*
Diameter at breast height (in)
Hei
ght (
ft)
0 2 4 6 8 10 12
020
4060
80
Hugo Peak Stand 28 - Diameter vs. Height
Figure 5.7. Example plot of diameter versus height.

110
Time (5 yr)
Hei
ght (
ft)
2 4 6 8 10 12
020
4060
8010
0
Hugo Peak Stand - Height vs. Time
Figure 5.8. Example plot of individual tree heights over time.

111
20 40 60 80
05
1015
20
Height (ft)
Num
ber
Hugo Peak Stand 28 - Height
Figure 5.9. Example histogram of heights.

112
Cumulative LAI in competitors (sq ft)
Hei
ght (
ft)
0.0 0.5 1.0 1.5
010
2030
4050
Distance (ft)
Hei
ght (
ft)
-5 0 5 10
010
2030
4050
LAI profile of tree 386 year 1
Figure 5.10: Example cumulative leaf area index pro�le. The x axis on the left plotis the cumulative leaf area index (LAI). The x axis on the right plot is distance.
example of the plot of leaf area competition.
Plot variable rank order { This routine displays an ordered plot of the trees in a
stand versus a variable. The order variable need not be the plotted variable.
As described earlier, this plot is one way of looking at stand structure. Figure
5.11 is an example of ordered plots of variables.
Density management space { The density management relation is a common tool
used by foresters to evaluate the condition of a stand and the change within the
stand. This routine allows the plotting the relation of trees per unit area and
the quadratic mean diameter of the stand. Maximum lines are species speci�c
and so are not routinely plotted; however, a line of speci�ed stand density index

113
Relative Rank
Hei
ght (
ft)
0.0 0.2 0.4 0.6 0.8 1.0
020
4060
Hugo Peak Stand 28 - Height ordered by height
Figure 5.11. Example plot of rank order of the heights.

114
*
**
**
**
*****
********
Trees per acre
Qua
drat
ic M
ean
Dia
met
er
1 10 100 1000
15
1050
100
Hugo Peak Stand 28 - Density Management Diagram
Figure 5.12: Example size{density plot over time. The point represent the locationof the stand a successive points in time. The solid line is an assumed maximum forDouglas-�r of SDI 595.
can be overlayed to provide a reference point as done in Figure 5.12. Figure
5.12 is the track of a stand of Douglas-�r (Pseudotsuga menziesii) and the stand
density index (SDI) 595 (The assumed maximum for that species (Drew and
Flewelling, 1979)) for reference. Points indicate locations of the stand at each
�ve year time step.
Structure plots { A set of multi-plots allow many plots to be viewed at various times.
These include change in a set of reverse ordered tree heights with time, the
change in spatial index over time, change in species mix over time, and change in
the height distribution over time. In Figure 5.13 a few of the largest tree at time

115
one remain the largest tree though out the model run. Figure 5.14 illustrates
a large range in the density of the stand; however, the spatial character as
described by the Hopkins' index does not change. Figure 5.15 illustrates the
relationship among the three species in the stand. In this particular run only one
species Douglas-�r (df) survived to the end of the run. Figure 5.16 illustrates
the height distributions for the stand.
Tree means { This is a non-graphical routine to report the means and variances for
all the tree list variables.
One feature of the current model is the method of conveying the output to a user.
The method is based on the assumption that graphical information can be recognized
and comprehended faster than tabular information. There is a three layered approach
to model output display. The �rst level is a \picture" of the stand at the various time
steps. The second level is traditional plot displays such as scatter plots, histograms,
and function plots. Thirdly, the tabulated data are available for question, which arise
from examination of the graphical data.
5.4 Description of implementation
This model building e�ort has been an evolution through several computational plat-
forms. Initially the programming was implemented in the C++ programming lan-
guage on a IBM compatible computer. This program worked well in some respects
but, had drawbacks. The �rst problem was to build a graphical interface for the
display of the model output. Using a low level graphics library supplied with the
compiler. Implementation of the program was straight-forward and the concept of
classes the C++ language make the building of the routines relatively fast and easy.
This programming approach was abandoned for three reasons. First, the implementa-
tion grew and the programs became di�cult to debug because of the memory required

116
..............................................................................................................................................................................................................................................................................................................................................................................................................................
0100
300
0 20 40
Tim
e 1
.... .. ..... . .... ... .. ... .. ... ..... . .. ....... ... . ... .. .. .. .. ... .. .. ... ..... .. ... ... .. ... ... .......... .... ... ..... .... .... ... ......... . ....... ..... ...... ...... ..... .... .... ..... .. .... .. ... .. ... ... ....... ........ ..................... ....... ......... ....... ... ... ............... ... ... ........ ... ......... .......... ...... ..... ...................... ..... ...... .. ..... ....... ........ .... ........................... .......
0100
300
0 20 50
Tim
e 2
.... .. .. ... . .... ... .. ... .. ... .. ... ... ....... ... .... .. .. .. .. ... .. ..... ....... ... ... .. .. . ... .... .. .... .... ... ... .. .... .. .. .. . . ... ..... . .... ... .. ... ... ... ..... .. .... .. .. ... . ...... .. .. . .. ... .. . .. ..... .. ... .. . .. ... ... ... .. .. .. ..... .. ..... ... .. ... .. .... .. ....... .. . .. . ...... ... .. .... ... .... ... . ... ..... .. ... ........ ..... .. .. ... . ..... .... .. ....... ... ... .. .... . ... .. ... .. ...... ........ ....... ........ ............. .........
0100
300
0 20 60
Tim
e 3
.. .. .... .. .. .... ... .. ... ..... .. .. . ... . ... .. . .. .. . .. .... . . ... .. .. .. . .. .... . .... . ... .. .. .... . ... .. .. .. .... .. .. .. .. .. .... .. .. .. .. .... ... .... . ... ... .. .. ... ..... .. .... .. .. ... . . .. ... .. .. . .. ... .. . .. ..... .. ... .. . .. . .. . .. ... .. .. ....... .. . .... ... .. ... .. .... .. ... ..... .. .. . ...... ..... ... .... .... ... .... .. ... .. ... ... ..... ..... .... ... . ......... .. ....... ... ..... ..... ... .. ..... ...... . ... ... . ........................... ..........
0100
300
0 40
Tim
e 4
.. . . .. .. .... ... . .. . .. .. . ..... .. .. . ... .... ... .. .. . .. ... . .. ... .. .. ... .. .... . ... . . ..... .. .... . ... . ... ...... .. .. .. .. .. . ... .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .... . .. . . .. ... .. .. . .. ... .. . .. .. ... .. ... .. . .. . .. . .. ... .. .. .. ..... .. . .... ... .. ... .. .... .. ... ..... .. .. . ...... ..... ... ....... . ... .... . . ... .. ... ... ..... ..... .... .... ..... .... .. ....... ... ..... ..... ... .. ... .. .. .... . ... .... .............................. .......
0100
300
0 40 80
Tim
e 5
.. . . ...... .. ... . ... .. .. . ... .. .... . ... .... .. . .. .. . .. ... . . . ... .. .. ... .. .... . ... . . ..... .. . ... . ... . ... . ..... .. .. .. .. .. . ... .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... .. . .. .. ... .. ... .. . .. . .. . .. ... .. .. .. ..... .. . .... ... .. ... .. .... .. . .. ..... .. .. . ...... ... .. ... ........ ... .... .. ... .. ........... ..... .... .... ..... .... .. ....... ... ..... ..... ... .. ..... ...... . ... .... ........... ................... .......
0100
300
0 40 80
Tim
e 6
.. . . ...... .. .... .. . .. .. . .. . .. .. .. . ... .... .. . .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . .... ... . ... . ... . ... . ..... .. .. .. .. .. . ... .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .... . .. . . .. ... .. .. . .. ... ... .. .. ... .. ... .. . .. . .. . .. ... .. .. ....... .. . .... ... .. ... .. .... .. . .. ..... .. .. . ...... ..... ... . ... .... ... .... .. ... ... .......... ..... .... ... . ..... .... .. ....... ... ..... ..... ... .. ..... .. ... .. ... .... .............................. .......
0100
300
0 40 80
Tim
e 7
.. . . ...... .. .... .. . .. .. . .. ... .. .. . ... . ... ... .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . .... ... . ... . ... . ... . ..... .. .. .. .. .. . ... .. .. .. .. .... . .. .... . ... ... .. .. ... .... . .. .... .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ... .. . .. . .. . .. ... .. .. .. ..... .. . .... ... .. ... .. .... .. . .. .... . .. .. . ...... ..... ... ........ ... .... .. ... .. ........... ..... .... ... . ..... .... .. ....... ... ... .. ..... ... .. ... .. ...... . ... ... . ....... ..............................
0100
300
0 40 80
Tim
e 8
.. . .... ... .. .... .. ... ... .. ... .. .. .... . ... ... .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . .... ... . ... . ... . ... . ..... .. .. .. .. .. . ... .. .. .. .. .... . .. ... . . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... .. . .. ..... .. ..... . .. . .. . .. ... .. .. ....... .. . .... ... .. ... .. .... .. . .. .... . .. .. . ...... ..... ... ........ ... .... .. ... ... .......... ..... .... ... ......... ... ....... ... ..... ..... ... .. ... .. ....... ... ... . ........... ................... .......
0100
300
0 40 80
Tim
e 9
.. . . .. . ... . . .... .. ... ... .. . .. .... .. .. . ... ... .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . .... ... . ... . ... . ... . ..... .. .. .. .. .. . ... .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ..... . .. . .. . .. ... .. .. ....... .. . .... ... .. ... .. .... .. . .. ..... .. .. . ...... ..... ... .... .... ... .... .. ... .. ........... ..... .... .... ......... .. ....... ... ..... ..... ... .. ... .. ....... ... .... .....................................
0100
300
0 40 80
Tim
e 10
.. . . .. .. .... . ... .. ... .. . .. . .. . ... .. .. . ... ... .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . ... .. .. .. .. .... . .. ... . . ... ... .. .. ... ..... .. .. .. .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ..... . .. . .. . .. ... .. .. ....... .. . .... ... .. ... .. ... ... . .. ..... .. ... ...... ..... ... ........ ... .... .. ... .. ...... ..... ..... .... ... . ......... .. ....... ... ..... ..... ... .. ... .. ...... . ... .... ... ............ ............... .......
Relative ht
0100
300
0 40 80
Tim
e 11
.. . . .. . ... ... ... .. . .. .. . .. ... . ... .. ... ... ... .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ..... . .. . .. . .. ... .. .. .. ....... ..... ..... ... .. .... .. . .. .... . .. .. . ...... ..... ... ....... . ... .... .. ... .. ...... ..... ..... .... ... . ..... .... .. ....... ... ..... ..... ... .. ... .. ...... . ... .... ....... .... ..........................
0100
300
0 40 80
Tim
e 12
.. . . .. .... .. . ... .. ... .. . .. ... .. .. .. ... ... ... .. .. . .. ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... .. . .. ..... .. ... .. . .. . .. . .. ... .. .. .......... .... ... .. ... .. .... .. . ...... . .. .. . ...... ..... ... ........ ... .... .. ... .. ...... ..... ..... .... ... . ......... .. ....... ... ..... .... . ... .. .......... .. ...... . .............................. .......
0100
300
0 40 80
Tim
e 13
.. . . .. ...... . ... .. ... .. . .. ... . ... ..... ... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... .... . .. .... .... . .. . . .. ... .. .. . .. ... .. . .. .. ... .. ... .. . .. . .. . .. ... .. .. .. ..... .. . .... ... .. ... .. ... ... . .. .... . .. .. . ...... ..... ... ........ ... .... .. ... ........ ..... ..... .... .... ........ ... ....... ... ... .. ..... ... .. ... .. .. ..... ... .... ....... .... ................ ..........
0100
300
0 40 100T
ime 14
.. . . .. .... .. . ... .. . .. .. . .. ... . ... . .... ... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .. .. .... . .. . . .. ... .. .. . .. ... ... .. .. ... .. ... .. . .. . .. . .. ... .. .. .. ..... .. . .... ... .. ... .. ... ... . .. .... . .. .. . ...... ..... ... ....... . ... .... .. ... ............. ..... .... .... ..... ...... ....... ... ..... ..... ... .. ..... ..... .. ...... . .....................................
0100
300
0 40 100
Tim
e 15
.. . . .. .... .. . ... .. ... .. . .. ... .... .. ... ... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ... .. . .. . .. . .. ... .. .. .. ........ .... ... .. ... .. .... .. . ...... . .. .. . ...... ..... ... . ....... ... .... .. ... .. ........... ..... ....... .......... .. ....... ... ..... ..... ... .. ..... .. ..... ... .... .....................................
0100
300
0 40 100
Tim
e 16
.. .. .. . ... .. . ... .. ... .. . .. ... .. .. .. ... ... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ... .. . .. . .. . .. ... .. .. .......... .... ... .. ... .. .... .. . ....... .. .. . ...... ..... ... ........ ... .... . . ... .. ...... ..... ..... .... .... ........... ... .... ... ..... ..... ... .. ..... ..... .. ....... ... ........................... .......
0100
300
0 40 100
Tim
e 17
.. .. .. . ... ... ... .. . .. .. . .. ... .. .. .. ... ... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . . ... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... .. . .. .. ... .. ..... . .. . .. . .. ... .. .. ....... .. ..... ... .. ... .. .... .. . .. ..... .. ... ...... ..... ... ....... . ... .... .. ... ... .......... ..... ....... .......... .. .......... ... .. ..... ... .. ... .. .. .... . ... .... ........................... ..........
Rank
0100
300
0 40 100
Tim
e 18
.... .. . ...... ... .. ... .. . .. ... .. .. .. ...... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . .... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .... .. .. . .. . . .. ... .. .. . .. ... ... .. .. ... .. ..... . .. . .. . .. ... .. .. ....... .. . .... ... .. ... .. ... ... . .. .... . .. .. . ...... ..... .... ....... ... .... .. ... .. ...... ..... ..... .... .... ........... ....... ... ..... .... . ... .. ............ ... .... ............................ .........
0100
300
0 40 100
Tim
e 19
.... .. . ... .. . ... .. ... .. . .. ... .. .. .. ... ... ... .. .. ... ... . . . ... .. .. ... .. .. .. . ... . . .... ... . ... . ... . ... . ..... .. .. .. .. .. . . .. .. .. .. .. .... . .. .... . ... ... .. .. ... ..... .. .. .. .... . .. . . .. ... .. .. . .. ... ... .. ..... .. ..... . .. . .. . .. ... .. .. .. ........ .... ..... ... .. .... .. . ....... .. ... ......... .. ... ........ ... .... .. ... ............. ..... ....... .......... .. ....... ... ..... ..... ... .. ..... ..... .. ....... ....... ..............................
0100
300
0 40 100T
ime 20
Figu
re5.13:
Exam
ple
oford
eredheigh
tsplot
overtim
e.Thexaxisisthereverse
ofsize
rank(th
elargest
treeis1)
andtheyaxisistherelative
heigh
tpercen
t(th
elargest
treeis100).
Tree
ranksare
based
ontim
e1;
they
change
order
inthexaxis
with
time.

117
*
*
**
** *
** *
*
**
**
* *
*
*
*
*
*
*
**
**
**
*
*
*
*
*
**
**
*
*
*****
****
**
* *
*
*
*
*
*
**
*
*
*
**
**
*
*
* * **
*
*
* **
**
*
*
**
*
*
**
*
*
**
*
*
**
**
*
**
*
*
*
*
*
*
*
**
**
***
***
*
*
*
*
*
*
* **
**
* *
**
*
*
*
*
**
*
*
**
*
* *
*
*
*
* *
*
**
***
*
*
*
*
*
*
*
*
*
* *
**
* *
*
*
*
**
*
*
*
**
*
*
**
*
*
* *
*
** *
*
*
* *****
*
*
**
** ***
**
***
*
*
*
**
* *
*
*
*
***
**
*
* *
* *
**
**
*
**
* **
*
* *
*
*
*
**
****
*
*
*
*
*
* *
*
* **
*
*** *
*
*
*
*
*
*
**
*
*
*
**
*
**
* **
**
*
* ***
*
*
***
**
*
****
*
**
** ** *
**
*
**
**
**
****
*
**
**
*
*
*
* **
**
*
*
*
*
**
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
***
*
**
*
*
* **
*
*
*
**
*
*
***
*
**
* *
**
*
*
**
* *
* *
*
*
*
**
***
*
*
0 40 80
040
80F
eet
Feet
Hopkins = 0.1839014
Time 1
*
*
****
**
**
*
*
*
*
*
*
*
* *
**
*
*****
**
** *
**
*
*
*
**
*
*
*
* *
* **
*
*
*
**
*
*
**
*
**
*
*
*
**
*
*
*
*
*
*
***
* **
* **
*
**
*
*
*
*
*
* *
*
*
*
*
**
*
*
**
*
*
**
*
*
*
*
*
*
*
**
***
*
***
*
* **
**
***
*
*
*
*
***
*
*
*
* *
***
*
***
*
**
***
*
*
*
*
** *
*
*
*
*
**
*
*
*
*
* **
**
* ***
*
**
****
*
***
**
** *
**
***
*
*
*
*
**
*
*
**
*
* *
*
*
*
*
*
*
*
*
*
* **
*
*
*
***
**
*
*
* **
*
* *
* *
*
*
**
**
*
*
0 40 80
040
80
Hopkins = 0.1983205
Time 2
***
***
*
*
*
*
* *
*
****
**
* ****
**
*
*
*
* **
*
**
*
**
**
*
* **
*
*
*
*
**
*
* * *
**
*
*
*
*
*
*
*
*
**
*
*
** *
**
*
*
*
*
*
***
**
**
**
***
*
*
*
***
*
*
*
* *
***
*
**
*
**
* *
*
*
*
*
**
*
*
*
*
**
**
*
*
* ***
*
**
****
*
**
**
****
**
*
**
**
*
**
*
* *
*
*
** *
*
*
* *
*
*
*
**
**
*
**
*
*
*
**
*
0 40 80
040
80
Hopkins = 0.1945829
Time 3
*
***
*
*
* *****
*
****
**
*
* **
*
*
* ** * **
**
**
*
*
**
*
*
**
*
*
**
*
* *
**
*
*
****
**
**
***
*
*
*
**
*
*
*
* *
* *
**
*
**
*
* *
*
**
*
*
*
**
**
*
***
**
***
*
**
**
* **
**
**
*
*
**
***
*
*
* *
*
*
**
*
*
*
* **
0 40 80
040
80
Hopkins = 0.1963809
Time 4
*
*
*
* ***
*
*
****
*
*
* *
*
*
* * * **
**
*
*
*
*
*
**
**
**
**
*
****
**
***
*
*
*
***
* *
*
**
*
** *
*
** *
*
**
**
*
***
***
*
*
**
**
* ***
**
*
*
**
**
*
**
*
*
**
*
**
0 40 80
040
80
Hopkins = 0.2062234
Time 5
*
*
* ***
*
*
****
*
* *
** * **
**
*
*
*
**
**
**
**
*
***
**
***
*
*
*
***
* *
*
**
*
**
*
** *
**
**
*
***
***
** * *
**
*
*
**
**
*
**
*
*
**
**
0 40 80
040
80
Hopkins = 0.1889682
Time 6
*
*
* ***
*
*
****
* *
** * *
**
*
*
*
**
**
**
**
*
*
*
**
***
*
*
***
*
**
*
*
*
** *
**
**
*
***
**
** *
**
*
*
*
***
* *
**
**
0 40 80
040
80
Hopkins = 0.1966938
Time 7
** *
**
*
*
****
*
**
**
*
*
*
**
**
**
**
*
*
*
**
**
*
*
*
*
*
**
*
*
** *
**
**
*
***
**
**
** ***
* *
***
0 40 80
040
80
Hopkins = 0.1935857
Time 8
** *
**
*
*
**
*
*
**
**
*
*
*
**
**
*
**
*
*
*
**
**
*
*
*
* **
*
*
** *
*
**
*
***
**
**
** ** *
***
0 40 80
040
80
Hopkins = 0.1941776
Time 9
** *
*
*
*
**
*
*
**
**
*
*
*
**
**
**
*
*
*
**
**
*
*
*
* **
*
*
** *
**
*
***
**
**
* ** *
**
0 40 80
040
80
Hopkins = 0.1770236
Time 10
** *
*
*
*
**
*
*
**
*
*
*
*
**
**
**
*
*
*
**
**
*
*
* **
*
*
** *
**
*
***
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.2111597
Time 11
** *
*
*
*
**
*
*
**
*
*
*
*
**
**
**
*
*
*
**
**
*
*
* **
*
*
** *
****
*
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.1850428
Time 12
**
*
*
**
*
*
**
*
*
*
*
**
**
**
*
*
*
**
**
*
*
* **
*
*
** *
****
*
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.1952518
Time 13
**
*
*
**
*
*
**
*
*
*
*
**
**
**
*
*
*
**
**
*
*
* **
*
*
** *
****
*
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.2009852
Time 14
**
*
*
**
*
*
**
*
**
**
**
**
*
*
*
**
**
*
*
* **
*
*
**
****
*
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.1879953
Time 15
**
*
*
**
*
*
**
*
**
**
**
**
*
*
*
**
**
*
*
* **
*
*
**
****
*
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.1777561
Time 16
**
*
*
**
*
*
**
*
**
**
**
**
*
*
*
**
**
*
*
* **
*
**
****
*
**
*
* ** *
**
0 40 80
040
80
Hopkins = 0.1806601
Time 17
**
*
*
**
*
*
**
*
**
**
**
**
*
*
*
**
**
*
*
* **
*
**
****
*
**
*
** *
**
0 40 80
040
80
Hopkins = 0.1916783
Time 18
**
*
*
**
*
*
**
*
**
**
**
**
*
*
*
**
**
*
*
* **
*
**
****
*
**
*
** *
**
0 40 80
040
80
Hopkins = 0.1994442
Time 19
**
*
*
**
*
*
**
*
**
**
**
**
*
*
*
**
**
*
*
* **
*
**
****
*
**
*
** *
**
0 40 80
040
80
Hopkins = 0.2002466
Time 20
Figure 5.14: Example of spatial pattern plot over time. The Hopkins' index for eachpattern is printed under the map plot.

118
df wh ch
020
0
Time 1
df wh ch
010
020
0
Time 2
df wh ch
050
150
Time 3
df wh ch
040
100
Time 4
df wh ch
040
80
Time 5
df wh ch
040
80
Time 6
df wh ch
040
80
Time 7
df wh ch
020
60
Time 8
df wh ch
020
50
Time 9
df wh ch
020
40
Time 10
df wh ch
020
40
Time 11
df wh ch
020
40
Time 12
df wh ch
020
40
Time 13
df wh ch
020
40Time 14
df wh ch
020
40
Time 15
df wh ch
020
40
Time 16
df wh ch
020
40
Time 17
df wh ch
020
40
Time 18
df wh ch
020
40
Time 19
df wh ch
020
40Time 20
Figure 5.15: Example of species barchart plot over time. The x axes are the variousspecies found on the plot and the y axes is the number of individuals in each species.df = Douglas-�r, wh = western hemlock, and ch = cherry.

119
0 20 40
020
60
Height (ft)
Num
ber
Time 1
10 30 50
020
Height (ft)
Num
ber
Time 2
10 30 50 70
010
25
Height (ft)
Num
ber
Time 3
10 30 50 70
05
10
Height (ft)
Num
ber
Time 4
20 40 60 80
05
15
Height (ft)
Num
ber
Time 5
20 40 60 80
05
15
Height (ft)
Num
ber
Time 6
40 60 80
05
10
Height (ft)
Num
ber
Time 7
60 70 80 90
05
15
Height (ft)
Num
ber
Time 8
80 90 100
04
8
Height (ft)
Num
ber
Time 9
85 95 105
04
8
Height (ft)
Num
ber
Time 10
90 100 110
04
812
Height (ft)
Num
ber
Time 11
95 105 115
04
812
Height (ft)
Num
ber
Time 12
100 110 120
05
1015
Height (ft)
Num
ber
Time 13
105 115 125
05
10
Height (ft)
Num
ber
Time 14
110 120 130
04
812
Height (ft)
Num
ber
Time 15
115 125
04
812
Height (ft)
Num
ber
Time 16
120 130
04
812
Height (ft)
Num
ber
Time 17
125 135
04
812
Height (ft)
Num
ber
Time 18
130 140
04
812
Height (ft)
Num
ber
Time 19
130 140
05
1015
Height (ft)
Num
ber
Time 20
Figure 5.16: Example of histogram of heights plotted over time. The x axis is theheight of the height classes in feet and the y axis is the number of individuals in eachclass.

120
by both the debugging software and the program. As the complexity of particular
programs increased, the amount of time to debug the program increased exponen-
tially. The size of the programs and the run times on the 386 system became large
and long, respectively. The models were converted to run on a Sun Sparcstation 1.
The DOS software was left at the state of development at the time of the conversion.
The goal is to demonstrate the viability of the approach and not to produce a
commercial product. Additionally, no C++ compilation software was available on the
Sun Sparcstation used. The S statistical package provided all the graphics capability
required and the exibility to implement the model through S user functions, linked
C functions, and stand-alone C programs.
This approach provided a good development environment; for example, a function
or routine that would take a week to code in C or C++ would take about a day in
S. The conversion from C++ version to the S version took approximately a month.
The S version of the model is fragile, requiring a knowledge of S and Unix. One very
nice feature of S is that it is an interactive environment; if the user would like to plot
data in a di�erent way, it is a relatively easy task to write a new function to make
the plot. S is one of several interactive packages that would be suitable for this type
of work. The real advantage is that the package handles the graphics user interface,
allowing the modeler to concentrate on the relationships and behavior of the model.
To implement a system based on graphical displays, a good graphical user interface
is needed. In the current project both interfaces developed from a low level routines
and interfaces developed from high level routine were used. Both system seem to
work well within the limitation of their respective computer. They provide a easy
method to view the output from the growth model and an easy method to visualize
the predictions.

Chapter 6
RESULTS AND DISCUSSION
\It is much more important to discover you are wrong and rectify that
situation than it is to go on inde�nitely."
John C. Merriam 1927. What Science can do for Forestry, Report of the
Conference on Commercial Forestry
This chapter presents the result of this model building exercise. This chapter
covers an illustration of the use of the model with various data sets, a discussion of
the evaluation process with examples of height growth patterns, and a discussion of
the advantages and disadvantage of the present modeling approach.
6.1 Problems in analyzes of the present type
A model with no way to check its performance is of little use; and an \adaptive stand
dynamics model" needs to be tested! \Adaptive stand dynamics models" are designed
to work with general stand dynamics relationships, parameterized to a speci�c stand
for projection of that stand's structure. They are designed for use where the stands
being projected are periodically remeasured. One of the best methods of evaluating
is to begin the projection from a time in the past, with information based on stand
reconstruction, and project the stand to the current time. Evaluating the di�erences
between projected and actual values should allow one to determine how well the model
is adapting to the current growth trend. This method would however, only indicate
how well the parameters were built, not how well the model will do in the future.

122
Another method is to select parameters of interest and to plot the change in param-
eters as more information is added to the growth sequence. Stable parameters would
indicate that the early projections are reasonable and the treatments selected will
probably develop as predicted. Unstable parameters would indicate that treatment
decisions made on those projections are probably in error and should be reevaluated.
Still another method is to compare mean trends of various diagnostic criteria of the
actual stand and the projected stand. Diagnostic criteria are common measures used
by silviculturists to determine stand condition (e.g. height-diameter ratios, density
management diagrams, spacing-top height ratios, and others). This comparison will
help determine if the parameters are reasonable and consistent with the behavior of
the modeled system. This evaluation is less precise, since it depends mainly on the
experience of the manager concerning the \reasonable" range of the various diagnostic
criteria; however, it can increase the con�dence of the model user.
This model is not a best statistical �t to the data, but a guided evaluation of
probable trends of the stand evaluated from many di�erent points of view. The
advantage of the approach is the exibility of the modeling system; the disadvantage
is the uncertainty of the validity of the prediction.
When considering testing the model, a quandary developed. Traditional models
have two parts to examine; the model forms and the parameters. Usually, the model
forms chosen by the model builder are assumed to be the best available. Then, the
testing process is to determine how well the parameters �t for those model forms
predict an independent data set. Adaptive stand dynamics models do not follow this
logic because there are no �xed parameters. Parameters adapt to observed growth
trends in the subject stand up to the current point, and future growth is strongly
a�ected by the assumption build into the model form and the assumption incorporated
by the user during parameterization.

123
6.2 Approach to demonstration and evaluation of the adaptive stand
dynamics model
The approach followed in this chapter to accomplish the stated goal will be as follows:
� Present the data used to test the ability to calibrate the model.
� Discuss the process of parameterization
� Check height growth projections
� Show example model runs
6.3 Example data sets
Three data sets are presented to illustrate the process of building a set of parameters
for a given stand. Each set has distinguishing features that present various chal-
lenges to the process. While none of the sets are remeasurment plots, they all have
reconstruction or stem analysis data.
6.3.1 Hugo Ridge Stand 28 Data Set
The Hugo ridge stand is in the southeast corner of the northeast quarter of the
southeast quarter of section 28, T16N, R4E. The stand was planted with Douglas-�r
(Pseudotsuga menziesii) in 1969 to approximately an 8'x8' foot spacing and a larger
amount of volunteer western hemlock (Tsuga heterophylla (Raf.) Sarg.), and Douglas-
�r had seeded in. The crown bases of dominant tree had receded approximately 10
feet.
6.3.2 Data description
The data were collected in the fall of 1989 by the author. A 2500 square foot plot
was laid out within the stand. All �eld measurement were taken in English units.

124
All live and dead trees were mapped using the interpoint distance method (Rohlf
and Archie, 1978). In all, 105 living trees and 244 dead trees were measured. Dead
trees were removed from the plot after their heights were measured. The diameter
at breast height, tree height, and height to live crown base were measured on all the
living trees.
A core set of thirteen trees was chosen roughly within the center 900 ft2 of the plot.
All dead and living branches were measured on these trees. On the dead branches the
following variables were measured: height to branch, branch angle, branch azimuth,
branch length. All these items were measured on the living branches plus green length
(length containing living foliage), green width, and green depth. A black and white
photograph of each green branch was taken. The branches were removed, the trees
were felled, and each whorl was removed for stem analysis. In the laboratory, annual
radial growth of the stem on each side of each whorl was measured.
For this analysis, a tree list was prepared containing species, x and y coordinates,
diameter at breast height, tree height, height to crown base, and crown width. The
tree list for this data set was complete except for crown widths, which were subsampled
on the stem analysis trees. A list of height/age values was created from the discs for
all stem analysis trees. Table 6.1 list the means and ranges of the input data. The
units are expressed in English units to illustrate the exibility built into the model.
Other data are in either metric or English units.
6.4 Bethel Ridge Data Set
The Bethel Ridge stand is at an undetermined location on Bethel Ridge in the Uni-
versity of Washington's Charles Lathrop Pack forest. The data were collected by
L. C. Kuiper and recorded on an unpublished drafting sheet. A description of the
techniques used is published in Kuiper (1988). This was an approximately 60 year
old Douglas-�r stand developed, and from natural regeneration after cutting of an old

125
Table 6.1. Description of the input data set for the Hugo Peak Stand
Mean Variance Minimum Maximum
Diameter (in.) 2.52 5.24 0.00 8.38
Height (ft.) 25.47 182.39 3.30 55.00
Crown Base Height (ft.) 10.16 6.82 2.00 15.00
Crown Width (ft.) 0.49 3.29 0.00 9.00
Height/Diameter 163.73 3896.39 68.02 442.11
growth forest.
6.4.1 Data description
The data were collected in May, 1984, and recorded on a single page drafting sheet.
A stem map of the plot and the crown projections were drawn. Two stand pro�les
were shown through the plot as well as a table of data with species, social position,
age, diameter at breast height, total height, live crown ratio, crown length, and crown
width. The species on the plot were Douglas-�r (Pseudotsuga menziesii) and western
hemlock (Tsuga heterophylla). Additionally, ten trees were dissected to provide height
growth patterns. Measurements were in metric units. Crown lengths and ages were
subsampled. Table 6.2 lists the means and ranges of the data for the Bethel Ridge
stand.
6.5 Helena Data Set
These data are from a set of experimental plots where four rows of each species were
planted. The species were cherrybark oak (Quercus falcata var. pagodifolia Ell.),
American sycamore (Platanus occidentalis L.), yellow poplar (Liriodendron tulipifera
L.) and eastern cottonwood (Populus deltoides Bart. ex Marsh.). The cottonwood
experienced very high mortality so that one side of one oak row had no competition.

126
Table 6.2. Description of the input data set for the Bethel Ridge Stand
Mean Variance Minimum Maximum
Diameter (cm.) 27.85 94.43 6.50 53.00
Height (m.) 28.83 20.25 12.50 34.80
Crown Base Height (m.) 10.53 12.33 2.70 17.40
Crown Width (m.) 4.02 2.07 1.20 9.60
Height/Diameter 101.17 573.02 56.79 185.45
6.5.1 Data description
These plots are in Lee County Arkansas, on Crowley's ridge in the middle of an
alluvial ood plain in Northeast Arkansas. The soils are a silt loam and the site index
is 90 to 110 for cherrybark oak and 110 plus for sycamore. The stand was planted
by the Chicago Mill company during the 1959-60 season. Three species were planted
on an 8'x8' spacing in alternating blocks of four rows of each species. The eastern
cottonwood did not survive, so the resulting stand consisted of four rows of cherry
bark oak, four rows of sycamore, four empty rows, and then a repeat. This stand
was analyzed and reported in two papers (Clatterbuck et al., 1987; Oliver et al.,
1990). The data used in this study included height-diameter pairs from the stand
and height/age curves for both species. Table 6.3 lists the means and ranges for the
Helena plots input data.
6.6 Parameterization process
The parameterization process, as described in the previous chapter, uses a set of
data to build a parameter set that can be used to make growth projections with the
model. The �rst step is to de�ne the maximum height growth curve for the plot,
because height growth modi�ers reduce this maximum height growth. The routine
reads the data set and �ts a tentative cumulative weibull equation to the height data.

127
Table 6.3. Description of the input data set for the Helena plots
Mean Variance Minimum Maximum
Diameter (in.) 8.19 13.82 2.60 16.80
Height (ft.) 65.70 446.83 23.00 103.00
Crown Base Height { { { {
Crown Width { { { {
Height/Diameter 102.46 370.89 65.68 139.46
These tentative, �ts are seldom adequate unless the height growth data are from the
full range of potential height-age pairs. The function is then plotted and the ranges
and parameters can be changed to allow the users to de�ne a curve , which they feel
represents the expected height-age curve. For the current runs a maximum age of 150
was used.
6.6.1 Hugo Peak stand
In �tting these data the curve can be bent to �t the observed data very closely; how-
ever, the curve would then reach the asymptote more quickly than experience suggests
occurs in Douglas-�r. The parameters and ranges shown in Figure 6.1 appeared to
be a good compromise. Figures 6.2 and 6.3 are the use of the same routine with
published height-age data of each species (McArdle et al., 1949; Barnes, 1949) for an
appropriate site for Hugo Peak.

128
111111111111111111
1
22222
22222222222222222
333333333333333
3333
3
44444
4444444444444
4
5555555555555
5
5
66666
666666666
6
77777777777
88888
88888888888
8
999999999999999999
9
Hugo Peak Stand 28
0 50 100 150
Tree age (yr)
020
4060
8010
012
0H
eigh
t (ft)
Figure 6.1: Height-age curve �t to stem analysis data for Hugo Peak Stand. Thesolid line is the assumed height growth curve and the numbers are the trajectories ofindividual trees.

129
1
1
1
1
1
1
1
1
1
11
11
1 1 1
Height - Age Curve (McArdle, 1949)
0 50 100 150
Tree Age (yr)
020
4060
8010
012
014
0
Hei
ght (
ft)
Figure 6.2: Height-age curve �t to published height-age data for Douglas-�r (McArdle1949). The line with ones in is the McArdle height/age curve and the solid line is theassumed height/age curve.

130
1
1
1
1
1
11
Height - age Curve (Barnes, 1949)
0 50 100 150
Tree Age (yr)
020
4060
8010
012
014
0
Hei
ght (
ft)
Figure 6.3: Height-age curve �t to published height-age data for western hemlock(Barnes, 1949). The line with the ones in it is the Barnes height/age curve and thesolid line is the assumed height/age curve

131
Height growth and Height growth rate
Time (yr)
Hei
ght g
row
th (
m)
0 50 100 150
0.0
0.2
0.4
0.6
010
2030
40H
eigh
t (m
)
Figure 6.4: Height-age curve �t to stem analysis data for Bethel Ridge Stand. Thenumber lines are the height/age trajectories for individual tree and the solid line isthe assumed height/age curve.
6.6.2 Bethel Ridge stand
The Bethel Ridge data were only for Douglas-�r. There were only two western hem-
lock trees and these were not analyzed. Figure 6.4 is a �t below the maximum and is
closer to the solution given by the �tting routine. In the runs in the following chapter,
a �t along the maximum of tree 9 was used.

132
11111111111111111111111111
2222222222222222222222
2
33333333333333333333333
3
4444444444444444444444444
555555555555555555555555
5
Helena Plots -- Sycamore Data
0 50 100 150
Tree Age (yr)
020
4060
8010
012
014
0
Hei
ght (
ft)
Figure 6.5: Height-age curve �t to sycamore stem analysis data for the Helena plots.The numbered line contains the trajectories for individual trees and the solid line isthe assumed height/age curve.
6.6.3 Helena plots
The data set for the Helena Plots included stem analysis for both cherrybark oak
and for American sycamore. Figure 6.5 is the plot of the chosen height-age curve for
sycamore with the data and Figure 6.6 is the plot for the chosen height-age curve for
cherrybark oak.

133
11111111111111111111
22222222222222222222
2
333333333333333333333
444444444444444444444
55555555555555555555
666666
6666666666666
Helena Plots -- Cherrybark Oak
0 50 100 150
Time (yr)
020
4060
8010
012
014
0
Hei
ght (
ft)
Figure 6.6: Height-age curve �t to cherrybark oak stem analysis data for the Helenaplots. The numbered lines contain the trajectories for individual trees and the solidline is the assumed height/age curve.

134
6.7 Evaluation of predicted height growth patterns
As an example, the predicted height growth patterns will be compared with the height-
age data in this section evaluate the ability of the model system to conform to the
observed data. Since the height-age curve de�ned in the parameterization process is
a maximum rate, the maximum simulated height growth is compared to the observed
height age trends.
The Hugo Peak run indicates good conformity. The Bethel Ridge run also indicates
that the observed data and the simulated (Figure 6.103) give similar results. The
Height/age data were collected for both species on the Helena plots and reasonable
agreement can be observed between the observed data and the simulated maximum
height (Figure 6.104 and 6.105).

135
Hugo Peak Stand - Check maximum height growth
Time (yr)
Hei
ght (
ft)
0 10 20 30 40 50 60
050
100
150
11111111111111111
11
2222222222
222222222222
333333333333333
33333
444444444444444
4444
5555555555
55555
66666666666666
6
77777777777
88888888888
888
88
8
99999999999999
9999
9
Figure 6.7: Comparison of the observed height growth and the maximum heightgrowth in the Hugo Peak simulation. The numbered lines contain the trajectories ofindividual trees and the solid line is the maximum predicted height growth from thecalibrated model. The other lines are the maximum growth predicted for the otherspecies.

136
Bethal Ridge Stand - Check maximum height growth
Time (yr)
Hei
ght (
m)
0 10 20 30 40 50 60
010
2030
40
11
1
1
1
1
1
1
11 1
22
2
2
2
2
2
2
22
2
3
3
3 3
3
3
3
3
3
33
44
4
4
4
4
4
44
44
5
5
5
5
5
5
55
55
5
6
6
6
6
6
6
6
66
66
7
7
7
7
7
7
7
7
77
7
8
8
8
8
8
8
8
88
88
9
9
9
9
9
9
9
9
99
9
Figure 6.8: Comparison of the observed height growth and the maximum heightgrowth in the Bethel Ridge simulation. The numbered lines contain the trajectoriesof individual trees and the solid line is the maximum predicted height growth fromthe calibrated model. The other lines are the maximum growth predicted for theother species.

137
Helena Plots - Check maximum height growth
Time (yr)
Hei
ght (
ft)
0 10 20 30 40 50 60
050
100
150
111111111111111111111
11
11
1
22222222222222222
22
222
2
3333333333333333333
333
33
444444444444444
44444
44
44
4
555555555555555555
555
555
5
Figure 6.9: Comparison of the observed height growth and the maximum heightgrowth in the American sycamore in Helena Plots simulation. The numbered linescontain the trajectories of individual trees and the solid line is the maximum predictedheight growth for sycamore from the calibrated model. The dashed lines are themaximum growth predicted for cherrybark oak.

138
Helena Plots - Check maximum height growth
Time (yr)
Hei
ght (
ft)
0 10 20 30 40 50 60
050
100
150
11
11111111111
11
111
11
2222222222222222
2222
2
33333333333
33333333
33
444444444444
44
44444
44
55555555555
55555
55
55
66
66
66
666666
666
66
66
Figure 6.10: Comparison of the observed height growth and the maximum heightgrowth in the cherrybark oak in the Helena Plots simulation. The numbered linescontain the trajectories of individual trees and the solid line is the maximum predictedheight growth for sycamore from the calibrated model. The dashed lines are themaximum growth predicted for cherrybark oak.

139
6.8 Example model runs
Several example runs are presented to illustrate the dynamic nature of the current
model. All runs are 60 years long and start from a stand with heights that are appro-
priate to a stand at 5 years old. These runs have no intermediate operations and only
make assumptions of the initial condition. The lack of other combinations was done
for brevity. The six scenarios presented require approximately 100 pages of �gures to
illustrate the model behavior. The model runs include four variations for the Hugo
Peak stand: assumptions of natural origin, no regeneration and natural origin with
regeneration; and planted origin, no regeneration and planted origin with regenera-
tion. These variations illustrate the range of stand structures that are produced from
these simple assumptions. Additionally, the Bethel Ridge stand and the Helena plot
are run from the original parameters to illustrate parameterization of the model to
di�erent types of data. No edge correction is used in the current simulations, so the
plot may be viewed as a stand grown in a �eld.
6.8.1 Hugo Peak Stand, natural origin, full density
This run of the growth model for the Hugo Peak stand uses the original density at the
time of measurement to generate the pattern. The stand is assumed to have a spatial
pattern as if the stand were naturally regenerated. Figure 6.7 illustrates four selected
stand maps at time 1 (the initial time period; (i.e. the generated stand), time 5 (25
years after the initial time), time 9 (45 years after the initial time), and time 12 (60
years after the initial time). The plots illustrate the large amounts of crown surface
area around the time of crown closure and the thinning of the center of the stand.
Figure 6.8 presents stem pro�les for part of the stand. The entire stand was not
shown for clarity. Times of the pro�les are the same as those in the stand maps (it is
interesting to note the longer crown lengths around the time of crown closure).
Three individual trees were selected to illustrate the height, crown, and diameter

140
Feet
0 20 40 60 80 100
020
4060
8010
0
Fee
t
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
***
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
*
* *
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
**
**
*
**
**
*
*
*
*
*
*
*
* **
*
* *
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
Stand Map of Hugo Peak Stand at age 5
Feet
0 20 40 60 80 100
020
4060
8010
0
Fee
t
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
***
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
*
* *
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
**
**
*
**
**
*
*
*
*
*
*
*
* **
*
* *
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
***
*
Stand Map of Hugo Peak Stand at age 25
Feet
0 20 40 60 80 100
020
4060
8010
0
Fee
t
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
***
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
*
* *
*
* **
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
**
**
*
**
**
*
*
*
*
*
*
*
* **
*
* *
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
*
***
*
Stand Map of Hugo Peak Stand at age 45
Feet
0 20 40 60 80 100
020
4060
8010
0
Fee
t
*
*
**
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
***
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
* *
* **
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
**
*
**
**
*
*
*
*
*
*
* **
*
* *
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
*
**
Stand Map of Hugo Peak Stand at age 60
Figure 6.11: Stand maps at starting year, 25 years after start, 45 years after start,and 60 years after start

141
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 5
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 25
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 45
Distance (ft)
Hei
ght (
ft)
0 50 100
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 60
Figure 6.12: Stand pro�le at starting year, 25 years after start, 45 years after start,and 60 years after start

142
dynamics of these trees (Figures 6.9, 6.10, and 6.11). They were chosen to be a
dominant, co-dominant, and intermediate to suppressed tree within the stand. The
diameter growth plot is stylized since the diameter eccentricity is not known. Each
line denotes 5 years of growth in inches.
The next series of plots present individual tree variables for all trees alive at the
end of the 60 year growth period (Figures 6.12-6.17). The lower graph in the total
mean growth and the mean growth for each species. Tree variables presented are
diameter at breast height, tree height, height to crown base, crown width, foliage leaf
area, and height/diameter ratios (in the height growth plot (Figure 6.13) note the
di�erent heights for the di�erent species).
The ordered plots of the stand are a way of viewing two aspects of the stand
structure (Figure 6.18). The stand is ordered by relative size (height in this case) and
the stand is plotted at each time with the initial tree order. The change of a tree in
relation to other trees in similar relative positions with in the stand can be seen.
The plot of histograms over time illustrates the changes in height distributions
(Figure 6.19). A bimodal distribution developed with time.
The plot of the relative number of species over time indicates changes in the
species composition (Figure 6.20). This plot changes little except when regeneration
is allowed.
The density management diagram uses a diagnostic criteria to analysis the behav-
ior of the growth model (Figure 6.21). In this case the stand increases in size with
no mortality until the end of the run. Plot of the mean annual increment and the
periodic annual increment indicates that the stand is near culmination. The volume
equations used in the model are very simple and may not indicate true volume for
the stand.
The plot of foliage sum is designed to present the behavior of the total leaf area
for the stand over time (Figure 6.22). The lower plot is of the relationship of growth
over growing stock.

143
Hugo Peak Stand - Tree 188
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 188
Rings are 5 years apart in EnglishRadius (in)
Rad
ius
(in)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.13: Tree pro�le and diameter growth for a dominant tree. The radii are ininches. Each concentric circle denotes 5 years of growth.

144
Hugo Peak Stand - Tree 117
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 117
Rings are 5 years apart in EnglishRadius (in)
Rad
ius
(in)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.14: Tree pro�le and diameter growth for a co-dominant tree. The radii arein inches. Each concentric circle denotes 5 years of growth.

145
Hugo Peak Stand - Tree 107
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 107
Rings are 5 years apart in EnglishRadius (in)
Rad
ius
(in)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.15: Tree pro�le and diameter growth for an intermediate tree. The radiiare in inches. Each concentric circle denotes 5 years of growth. The reason that thistree regains its crown is because there is no minimum crown size below which a treecannot survive.

146
Time (5 yr)
Dia
met
er a
t bre
ast h
eigh
t (in
)
2 4 6 8 10 12
010
2030
40
Hugo Peak Stand - Diameter vs. Time
Time (5 yr)
Dia
met
er a
t bre
ast h
eigh
t (in
)
2 4 6 8 10 12
02
46
810
1214
Hugo Peak Stand - Diameter vs. Time
Totaldfwhch
Figure 6.16: Diameter growth for each tree over time (upper plot) and the meandiameter growth of each species (lower plot). The x axis is 5 year time steps and they axis is diameter at breast height in inches. df = Douglas-�r, wh = western hemlock,and ch = cherry.

147
Time (5 yr)
Hei
ght (
ft)
2 4 6 8 10 12
020
4060
8010
0
Hugo Peak Stand - Height vs. Time
Time (5 yr)
heig
ht (
ft)
2 4 6 8 10 12
2040
6080
100
Hugo Peak Stand - Height vs. Time
Totaldfwhch
Figure 6.17: Height growth for each tree over time (upper plot) and the mean heightgrowth of each species (lower plot). The x axis is in 5 year time steps and the y axisis height in feet. df = Douglas-�r, wh = western hemlock, and ch = cherry.

148
Time (5 yr)
Hei
ght t
o cr
own
base
(ft)
2 4 6 8 10 12
020
4060
8010
0
Hugo Peak Stand - Height to crown base vs. Time
Time (5 yr)
Hei
ght t
o cr
own
base
(ft)
2 4 6 8 10 12
020
4060
80
Hugo Peak Stand - Height to crown base vs. Time
Totaldfwhch
Figure 6.18: Height to crown base change for each tree over time (upper plot) andthe mean height to crown base change for each species (lower plot). The x axis is in5 year time steps and the y axis is height to crown base in feet.

149
Time (5 yr)
Cro
wn
Wid
th (
ft)
2 4 6 8 10 12
010
2030
40
Hugo Peak Stand - Crown width vs. Time
Time
cw
2 4 6 8 10 12
510
1520
Hugo Peak Stand - cw vs. Time
Totaldfwhch
Figure 6.19: Crown width for each tree over time (upper plot) and the mean crownwidth for each species (lower plot). The x axis is in 5 year time steps and the y axisis crown width in feet.

150
Time (5 yr)
Fol
iage
sur
face
are
a (s
q ft)
2 4 6 8 10 12
010
0020
0030
0040
0050
0060
00
Hugo Peak Stand - Foliage surface area vs. Time
Time (5 yr)
Fol
aiag
e su
rfac
e ar
ea (
sq ft
)
2 4 6 8 10 12
020
040
060
080
010
0012
0014
00
Hugo Peak Stand - Foliage surface area vs. Time
Totaldfwhch
Figure 6.20: Foliage leaf area for each tree over time (upper plot) and the mean foliageleaf area for each species (lower plot). The x axis is in 5 year time steps and the yaxis is foliage amount in square feet.

151
Time (5 yr)
Hei
ght-
Dia
met
er (
Per
cent
)
2 4 6 8 10 12
020
0040
0060
0080
00
Hugo Peak Stand - Height-diameter vs. Time
Time (5 yr)
Hei
ght-
diam
eter
(P
erce
nt)
2 4 6 8 10 12
100
150
200
250
300
350
Hugo Peak Stand - Height-diameter vs. Time
Totaldfwhch
Figure 6.21: Height/diameter ratio change for each tree over time (upper plot) andthe mean height/diameter ratio for each species (lower plot). The x axis is in 5 yeartime steps and the y axis is height-diameter ratio.

152
..........
.
...
.
.
.
.
..
..
.
..
..
.
.
.
.
.
.....
.
..........
.
.
.
..
........
.
.
.
.
..
.
.
.
..
..
.......
.
..
.
.
.
..
.
.
.
.
.
.
.
.
.
.
.
.
..
.
.....
..
.
.............
..
.
.
.
.
..
.
....
..
..
..
.
.
.
.
.
...
......
.
..
..
..
.
..
..
.
.
.
.
.
.
.........
.
.
...
.
.
..
.
.
.
.
...
.
.
...........
..
.
.
.
.
.
.
.
.
.
.
.
....
.
.
.
...
..
..
.
.
.......
.
.
.
.
.
...
..
.
.
.
.
.
.
.
.
.
......
.
.
.
.
.
...
.
.
.
.
..
.
.
.
..
.
.
.
.
.
.
.
..
...........
.
.
.
..
.
..
.
.
......
..
.
..
.
........
.
.
.
..
.
..
.
...
..
.............
.
........
.
..
.
.
...
..........................
0 100 200 300 400
02
46
Time 1
..............................................................................................................................................................................................................................................................................................................................................................................................................................
0 100 200 300 400
05
1015
20
Time 2
.
.....................................................................................................................................................................................................................
...
....................................................................................................................................
.
................................................................
0 100 200 300 400
010
2030
Time 3
.
.
...................................................
.
............................
.
.
...
...........................................................................................................................................................................................................
.
...
.
..
.
...............................................
.
...
.
.
.
.............................
.................................
0 100 200 300 400
010
2030
40
Time 4
..................................................................................
.
.
......................................................................................................................................................
..
......................................................
.
......................................................
.
.
..
.
.
....................................
.
..........................
Rel
ativ
e ht
0 100 200 300 400
010
2030
4050
60
Time 5
.............................................................................................................................................................................................................................................................................................................................................
.
.
.
..
.
...
.
.
.
.
...
.
.
........................................................
..
.....
0 100 200 300 400
020
4060
Time 6
.............................................................................................................................................................................................................................................................................................................................................
.
.
.
.........
.
.
..
.
.........................................................
..
....
.
0 100 200 300 400
020
4060
80
Time 7
....................................
..........................................................................
.
..............................................................................................................................................................
.
.
.............................
................................
.
..
.
.........
.
...
.
........
.
..........
.
.
.........
.
.
.
...................
.
.
.
..
..
....
.
0 100 200 300 400
020
4060
80
Time 8
..............................................................................................................
................................................................................................
.................................................................
.............................................................
.
..
.
.........
.
...
.
........
.
..........
.
.
.........
.
.
.
.
..................
.
.
.
.
.
..
....
.
0 100 200 300 400
020
4060
80
Time 9
.......................................
...................................................................
....................................................................................................
.......................................
.......................................................................................
.
..
.
........
.
.
...
.
........
.
..........
.
.
.........
.
.
.
.
....
.
...
.
....
..
.
..
.
.
.
.
.
..
....
.
Rank
0 100 200 300 400
020
4060
8010
0
Time 10
...........................................
....................................................................................................
.........................................................
..............................
...........................................................
...........................................
.
..
.
........
.
.
.
.
.
.
...
.
....
.
....
.
..
...
.
.
..
.
.
.
.
.
..
.
.
.
.
....
.
...
.
.
..
.
..
.
.
.
.
.
.
.
.
..
....
.
0 100 200 300 400
020
4060
8010
0
Time 11
............................................................
................................................................................................................
.
.....................................................................
..........................................................................................
.
..
.
........
.
.
.
.
.
.
.
..
.
..
..
.
...
.
.
.
.
...
.
.
...
.
.
.
.
..
.
.
.
.
....
.
...
.
.
..
.
.
.
.
.
.
.
.
.
.
.
..
.
.
.
.
.
0 100 200 300 400
020
4060
8010
0Time 12
Figure 6.22. Plot of ordered trees over time. The order time is time 2.

153
3 4 5 6 7
020
4060
8010
0
Height (ft)
Num
ber
Time 1
10 12 14 16 18 200
2040
6080
Height (ft)
Num
ber
Time 2
15 20 25 30
020
4060
8010
0
Height (ft)
Num
ber
Time 3
25 30 35 40 45
020
4060
8010
0
Height (ft)
Num
ber
Time 4
35 40 45 50 55 60
020
4060
8010
0
Height (ft)
Num
ber
Time 5
45 50 55 60 65 70
020
4060
8010
0
Height (ft)
Num
ber
Time 6
60 65 70 75 80
020
4060
8010
0
Height (ft)
Num
ber
Time 7
65 70 75 80 85
020
4060
8010
0
Height (ft)
Num
ber
Time 8
70 75 80 85 90 95
020
4060
8012
0
Height (ft)
Num
ber
Time 9
80 85 90 95 100
050
100
150
Height (ft)
Num
ber
Time 10
85 90 95 100
050
100
150
Height (ft)
Num
ber
Time 11
90 95 100 110
050
100
150
Height (ft)
Num
ber
Time 12
Figure 6.23. Histograms of tree height over time.

154
df wh ch
010
020
030
0
Time 1
df wh ch
010
020
030
0
Time 2
df wh ch
010
020
030
0
Time 3
df wh ch
010
020
030
0
Time 4
df wh ch
010
020
030
0
Time 5
df wh ch
010
020
030
0
Time 6
df wh ch
010
020
030
0
Time 7
df wh ch
010
020
030
0
Time 8
df wh ch
010
020
030
0
Time 9
df wh ch
010
020
030
0
Time 10
df wh ch
010
020
030
0
Time 11
df wh ch
010
020
030
0
Time 12
Figure 6.24. Histogram of species number over time.

155
*
**
*
*******
*
Trees per acre
Qua
drat
ic M
ean
Dia
met
er
1 10 100 1000
0.1
0.5
1.0
5.0
50.0
Hugo Peak Stand - Density Management Diagram
Time (5 yr)
Mai
and
Pai
(cu
bic
ft)
2 4 6 8 10 12
050
010
0015
0020
0025
00
Hugo Peak Stand - Mai and Pai vs. Time
Figure 6.25: Density management diagram for the plot and a plot of mean annualincrement and periodic annual increment.

156
Time (5 yr)
Fol
iage
Sum
(sq
ft)
2 4 6 8 10 12
010
0000
2000
0030
0000
4000
0050
0000
Hugo Peak Stand - Foliage Sum vs. Time
Growing Stock (cubic ft)
Gro
wth
(cu
bic
ft)
0 20000 40000 60000
050
010
0015
0020
0025
00
Hugo Peak Stand - Growth over Growing Stock
Figure 6.26: A plot of the stand foliage sum over time and a plot of growth overgrowing stock.

157
6.8.2 Hugo Peak Stand, natural origin, full density, with regeneration
This run of the model makes the same assumptions of a natural origin at the density
of the original stand but allows regeneration to enter the stand. Actually, very little
regeneration enters and stays in this run because the initial density is very high. The
main di�erence can be seen on the plot of the number of individuals by species (Figure
6.36). The dynamics of the minor species in the �rst few periods is di�erent than the
�rst run. Additionally, in the density management diagram one can see the ingrowth
of trees; however, the stand ends up in about the same position as in the �rst run.

158
Feet
0 20 40 60 80 100
020
4060
8010
0
Fee
t
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
***
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
*
* *
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
**
**
*
**
**
*
*
*
*
*
*
*
* **
*
* *
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
Stand Map of Hugo Peak Stand at age 5
Feet
0 20 40 60 80 100
020
4060
8010
0
Fee
t
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
***
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
*
* *
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
**
**
*
**
**
*
*
*
*
*
*
*
* **
*
* *
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
***
*
**
*
**
*
*
*
*
*
**
*
*
*
**
*
*
**
*
**
*
*
**
*
*
Stand Map of Hugo Peak Stand at age 25
Feet
0 20 40 60 80 100
020
4060
8010
0
Fee
t
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
***
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
** *
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
**
*
**
**
*
*
*
*
*
*
*
* **
*
* *
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
***
**
Stand Map of Hugo Peak Stand at age 45
Feet
0 20 40 60 80 100
020
4060
8010
0
Fee
t
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
***
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
** *
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
** *
* **
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
**
*
**
**
*
*
*
*
*
*
**
* *
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
*
**
Stand Map of Hugo Peak Stand at age 60
Figure 6.27: Stand maps at starting year, 25 years after start, 45 years after start,and 60 years after start.

159
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 5
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 25
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 45
Distance (ft)
Hei
ght (
ft)
0 50 100
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 60
Figure 6.28: Stand pro�le at starting year, 25 years after start, 45 years after start,and 60 years after start.

160
Hugo Peak Stand - Tree 188
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 188
Rings are 5 years apart in inchesRadius (in)
Rad
ius
(in)
-10 -5 0 5 10
-10
-50
510
Figure 6.29: Tree pro�le and diameter growth for a dominant tree. The radii are ininches and the concentric circles denote 5 year radial growth.

161
Hugo Peak Stand - Tree 117
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 117
Rings are 5 years apart in inchesRadius (in)
Rad
ius
(in)
-10 -5 0 5 10
-10
-50
510
Figure 6.30: Tree pro�le and diameter growth for a co-dominant tree. The radii arein inches and the concentric circles denote 5 year radial growth.

162
Hugo Peak Stand - Tree 107
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 107
Rings are 5 years apart in inchesRadius (in)
Rad
ius
(in)
-10 -5 0 5 10
-10
-50
510
Figure 6.31: Tree pro�le and diameter growth for an intermediate tree. The radii arein inches and the concentric circles denote 5 year radial growth.

163
Time (5 yr)
Dia
met
er a
t bre
ast h
eigh
t (in
)
2 4 6 8 10 12
05
1015
2025
Hugo Peak Stand - Diameter vs. Time
Time (5 yr)
Dia
met
er a
t bre
ast h
eigh
t (in
)
2 4 6 8 10 12
02
46
810
1214
Hugo Peak Stand - Diameter vs. Time
Totaldfwhch
Figure 6.32: Diameter growth for each tree over time (upper plot) and the meandiameter growth of each species (lower plot). df = Douglas-�r, wh = western hemlock,and ch = cherry.

164
Time (5 yr)
Hei
ght (
ft)
2 4 6 8 10 12
020
4060
8010
0
Hugo Peak Stand - Height vs. Time
Time (5 yr)
Hei
ght (
ft)
2 4 6 8 10 12
2040
6080
100
Hugo Peak Stand - Height vs. Time
Totaldfwhch
Figure 6.33: Height growth for each tree over time (upper plot) and the mean heightgrowth of each species (lower plot).

165
Time (5 yr)
Hei
ght t
o cr
own
base
(ft)
2 4 6 8 10 12
020
4060
8010
0
Hugo Peak Stand - Height to crown base vs. Time
Time (5 yr)
Hei
ght t
o cr
own
base
(ft)
2 4 6 8 10 12
020
4060
80
Hugo Peak Stand - Height to crown base vs. Time
Totaldfwhch
Figure 6.34: Height to crown base change for each tree over time (upper plot) andthe mean height to crown base change for each species (lower plot).

166
Time (5 yr)
Cro
wn
wid
th (
ft)
2 4 6 8 10 12
010
2030
40
Hugo Peak Stand - Crown width vs. Time
Time (5 yr)
Cro
wn
wid
th (
ft)
2 4 6 8 10 12
510
1520
Hugo Peak Stand - Crown width vs. Time
Totaldfwhch
Figure 6.35: Crown width for each tree over time (upper plot) and the mean crownwidth change for each species (lower plot).

167
Time (5 yr)
Fol
iage
sur
face
are
a (s
q ft)
2 4 6 8 10 12
010
0020
0030
0040
00
Hugo Peak Stand - Foliage surface area vs. Time
Time (5 yr)
Fol
iage
sur
face
are
a (s
q ft)
2 4 6 8 10 12
020
040
060
080
010
0012
0014
00
Hugo Peak Stand - Foliage surface area vs. Time
Totaldfwhch
Figure 6.36: Foliage leaf area change for each tree over time (upper plot) and themean foliage leaf area for each species (lower plot).

168
Time (5 yr)
Hei
ght/d
iam
eter
(P
erce
nt)
2 4 6 8 10 12
020
0040
0060
0080
00
Hugo Peak Stand - Height/diameter vs. Time
Time (5 yr)
Hei
ght/d
iam
eter
(P
erce
nt)
2 4 6 8 10 12
8010
012
014
016
0
Hugo Peak Stand - Height/diameter vs. Time
Totaldfwhch
Figure 6.37: Height/diameter ratio change for each tree over time (upper plot) andthe mean height/diameter change for each species (lower plot). The reason for thereduction in height/diameter ratios is because of the mortality in high height/diameterratio trees.

169
3 4 5 6 7
020
4060
8010
0
Height (ft)
Num
ber
Time 1
0 5 10 15 200
5010
015
020
025
0
Height (ft)
Num
ber
Time 2
0 10 20 30
010
020
030
0
Height (ft)
Num
ber
Time 3
0 10 20 30 40 50
050
100
150
200
250
Height (ft)
Num
ber
Time 4
0 10 20 30 40 50 60
050
100
150
200
Height (ft)
Num
ber
Time 5
0 20 40 60
050
100
150
Height (ft)
Num
ber
Time 6
0 20 40 60 80
010
020
030
0
Height (ft)
Num
ber
Time 7
50 60 70 80 90
050
100
150
200
250
300
Height (ft)
Num
ber
Time 8
60 70 80 90
050
100
150
Height (ft)
Num
ber
Time 9
80 85 90 95 100
050
100
150
Height (ft)
Num
ber
Time 10
85 90 95 100
050
100
150
Height (ft)
Num
ber
Time 11
85 90 95 100 110
050
100
150
Height (ft)
Num
ber
Time 12
Figure 6.38. Histograms of tree height over time.

170
df wh ch
010
020
030
0
Time 1
df wh ch
010
030
0
Time 2
df wh ch
010
030
050
0
Time 3
df wh ch
010
030
0
Time 4
df wh ch
010
020
030
0
Time 5
df wh ch
010
020
030
0
Time 6
df wh ch
010
020
030
0Time 7
df wh ch
010
020
030
0
Time 8
df wh ch
010
020
030
0
Time 9
df wh ch
010
020
030
0
Time 10
df wh ch
010
020
030
0
Time 11
df wh ch
010
020
030
0Time 12
Figure 6.39. Histogram of species number over time.
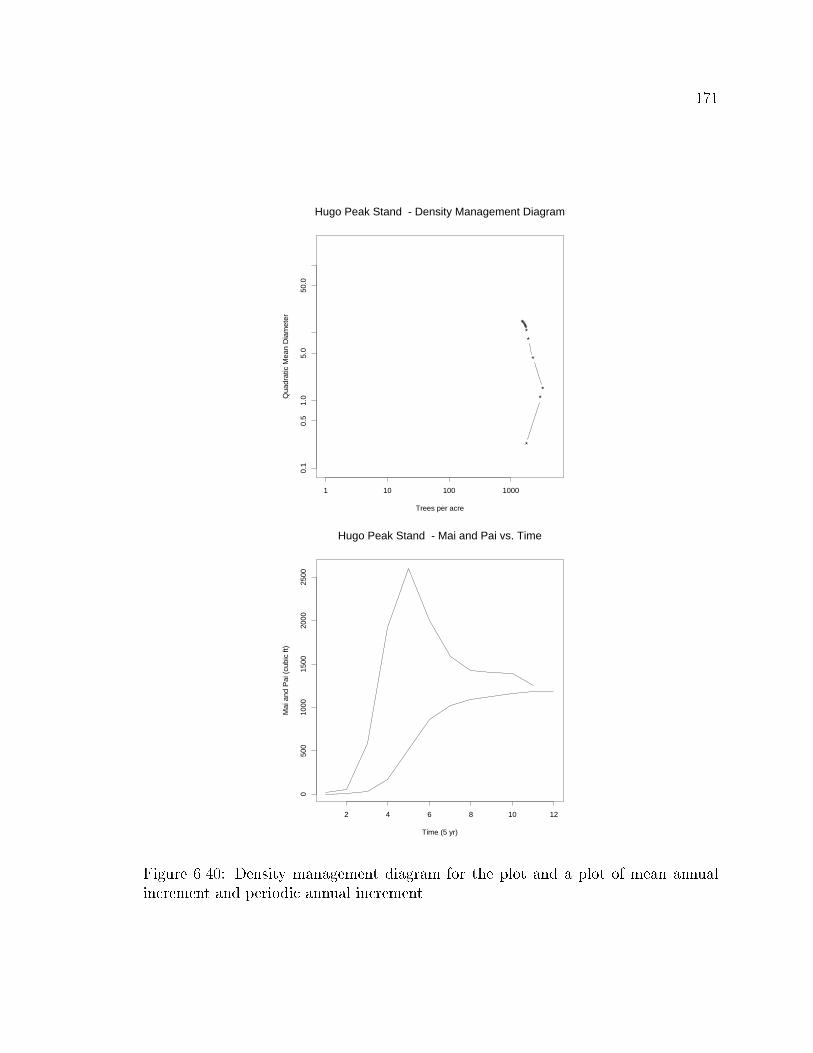
171
*
**
*
*******
*
Trees per acre
Qua
drat
ic M
ean
Dia
met
er
1 10 100 1000
0.1
0.5
1.0
5.0
50.0
Hugo Peak Stand - Density Management Diagram
Time (5 yr)
Mai
and
Pai
(cu
bic
ft)
2 4 6 8 10 12
050
010
0015
0020
0025
00
Hugo Peak Stand - Mai and Pai vs. Time
Figure 6.40: Density management diagram for the plot and a plot of mean annualincrement and periodic annual increment.

172
Time (5 yr)
Fol
iage
Sum
(sq
ft)
2 4 6 8 10 12
010
0000
2000
0030
0000
4000
0050
0000
Hugo Peak Stand - Foliage Sum vs. Time
Growing Stock (cubic ft)
Gro
wth
(cu
bic
ft)
0 20000 40000 60000
050
010
0015
0020
0025
00
Hugo Peak Stand - Growth over Growing Stock
Figure 6.41: A plot of the stand foliage sum over time and a plot of growth overgrowing stock.

173
6.8.3 Hugo Peak Stand, planted origin, 200 trees per acre
In this run, the simulation represents the original intention of the planted stand. It
assumes that 200 trees per acre were planted and the stand is grown with no other
regeneration allowed to enter the stand. The di�erences are very evident in the stand
maps and pro�les (Figures 6.39 and 6.40). The crowns maintain longer lengths and
the widths are more uniform.
Another di�erence is that the density management diagram shows no mortality
and the mean annual increment and periodic annual increment are much further from
culmination that in the �rst two runs (Figure 6.53).

174
Feet
20 40 60 80
2040
6080
Fee
t
* * * * ** *
* * * * * * *
* * * ** * *
* * ** * * *
* * * * * **
* * * * **
Stand Map of Hugo Peak Stand at age 5
feet
20 40 60 80
2040
6080
Fee
t
* * * * ** *
* * * * * * *
* * * ** * *
* * ** * * *
* * * * * **
* * * * **
Stand Map of Hugo Peak Stand at age 25
Feet
20 40 60 80
2040
6080
Fee
t
* * * * ** *
* * * * * * *
* * * ** * *
* * ** * * *
* * * * * **
* * * * **
Stand Map of Hugo Peak Stand at age 45
Feet
20 40 60 80
2040
6080
Fee
t
* * * * ** *
* * * * * * *
* * * ** * *
* * ** * * *
* * * * * **
* * * * **
Stand Map of Hugo Peak Stand at age 60
Figure 6.42: Stand maps at starting year, 25 years after start, 45 years after start,and 60 years after start

175
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
80
Stand Profile of Hugo Peak Stand at age 5
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
80
Stand Profile of Hugo Peak Stand at age 25
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
80
Stand Profile of Hugo Peak Stand at age 45
Distance (ft)
Hei
ght (
ft)
0 50 100
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 60
Figure 6.43: Stand pro�le at starting year, 25 years after start, 45 years after start,and 60 years after start

176
Hugo Peak Stand - Tree 16
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 16
Rings are 5 years apart in inchesRadius (in)
Rad
ius
(in)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.44: Tree pro�le and diameter growth for a dominant tree. The radii are ininches and the concentric circles denote �ve year radial growth.

177
Hugo Peak Stand - Tree 18
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 18
Rings are 5 years apart in inchesRadius (in)
Rad
ius
(in)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.45: Tree pro�le and diameter growth for a co-dominant tree. The radii arein inches and the concentric circles denote �ve year radial growth.

178
Hugo Peak Stand - Tree 4
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 4
Rings are 5 years apart in inchesRadius (in)
Rad
ius
(in)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.46: Tree pro�le and diameter growth for an intermediate tree. The radiiare in inches and the concentric circles denote �ve year radial growth. In the currentversion there is not threshold for crown size below with a tree cannot survive.

179
Time (5 yr)
Dia
met
er a
t bre
ast h
eigh
t (in
)
2 4 6 8 10 12
010
2030
4050
Hugo Peak Stand - Diameter vs. Time
Time (5 yr)
Dia
met
er a
t bre
ast h
eigh
t (in
)
2 4 6 8 10 12
010
2030
Hugo Peak Stand - Diameter vs. Time
Totaldfwhch
Figure 6.47: Diameter growth for each tree over time ( upper plot) and the meandiameter growth of each species (lower plot). df = Douglas-�r, wh = western hemlock,and ch = cherry.

180
Time (5 yr)
Hei
ght (
ft)
2 4 6 8 10 12
020
4060
8010
0
Hugo Peak Stand - Height vs. Time
Time (5 yr)
Hei
ght (
ft)
2 4 6 8 10 12
2040
6080
100
Hugo Peak Stand - Height vs. Time
Totaldfwhch
Figure 6.48: Height growth for each tree over time (upper plot) and the mean heightgrowth of each species (lower plot).

181
Time (5 yr )
Hei
ght t
o cr
own
base
(ft)
2 4 6 8 10 12
020
4060
80
Hugo Peak Stand - Height to crown base vs. Time
Time (5 yr)
Hei
ght t
o cr
own
base
(ft)
2 4 6 8 10 12
020
4060
80
Hugo Peak Stand - Height to crown base vs. Time
Totaldfwhch
Figure 6.49: Height to crown base change for each tree over time (upper plot) andthe mean height to crown base change for each species (lower plot).

182
Time (5 yr )
Cro
wn
wid
th (
ft)
2 4 6 8 10 12
010
2030
4050
60
Hugo Peak Stand - Crown width vs. Time
Time (5 yr)
Cro
wn
wid
th (
ft)
2 4 6 8 10 12
1020
3040
Hugo Peak Stand - Crown width vs. Time
Totaldfwhch
Figure 6.50: Crown width for each tree over time (upper plot) and the mean crownwidth for each species (lower plot).

183
Time(5 yr)
Fol
iage
sur
face
are
a (s
q ft)
2 4 6 8 10 12
020
0040
0060
0080
0010
000
Hugo Peak Stand - Foliage surface area vs. Time
Time (5 yr)
Fol
iage
sur
face
are
a (s
q ft)
2 4 6 8 10 12
010
0020
0030
0040
00
Hugo Peak Stand - Foliage surface area vs. Time
Totaldfwhch
Figure 6.51: Foliage leaf area for each tree over time (upper plot) and the mean foliageleaf area for each species (lower plot).

184
Time (5 yr)
Hei
ght/d
iam
eter
(P
erce
nt)
2 4 6 8 10 12
020
0040
0060
00
Hugo Peak Stand - Height/diameter vs. Time
In english unitsTime
hd
2 4 6 8 10 12
020
040
060
080
010
0012
0014
00
Hugo Peak Stand - hd vs. Time
Totaldfwhch
Figure 6.52: Height/diameter ratio change for each tree over time (upper plot) andthe mean height/diameter ratio for each species (lower plot).

185
.....
....
.....
....
.....
....
.
...
....
.
.
.
.
..
Rank
Rel
ativ
e ht
0 10 20 30 40
02
46
Time 1
......................................
...
Rank
Rel
ativ
e ht
0 10 20 30 400
510
1520
Time 2
.....
.........
......
...........
....
.
..
...
Rank
Rel
ativ
e ht
0 10 20 30 40
010
2030
Time 3
.............
.......
...........
....
.
.
.
...
Rank
Rel
ativ
e ht
0 10 20 30 40
010
2030
40
Time 4
...........
.........
...........
....
.
..
...
Rank
Rel
ativ
e ht
0 10 20 30 40
010
2030
4050
60
Time 5
................
..............
.....
.
.
....
Rank
Rel
ativ
e ht
0 10 20 30 40
020
4060
Time 6
....
....
.....................
.....
.
.
.
....
Rank
Rel
ativ
e ht
0 10 20 30 40
020
4060
80
Time 7
....
...............................
.
.
....
Rank
Rel
ativ
e ht
0 10 20 30 40
020
4060
80
Time 8
................
................
....
.
....
Rank
Rel
ativ
e ht
0 10 20 30 40
020
4060
80
Time 9
....
...............................
.
.
....
Rank
Rel
ativ
e ht
0 10 20 30 40
020
4060
8010
0
Time 10
...............................
....
.
.
....
Rank
Rel
ativ
e ht
0 10 20 30 40
020
4060
8010
0
Time 11
...................................
.
.
....
Rank
Rel
ativ
e ht
0 10 20 30 40
020
4060
8010
0
Time 12
Figure 6.53. Plot of ordered trees over time. The order time is time 1

186
4.0 5.0 6.0
01
23
45
Height (ft)
Num
ber
Time 1
12 14 16 18 20
02
46
810
Height (ft)
Num
ber
Time 2
20 25 30
02
46
810
12
Height (ft)
Num
ber
Time 3
30 35 40 45
02
46
810
12
Height (ft)
Num
ber
Time 4
40 45 50 55
02
46
810
Height (ft)
Num
ber
Time 5
50 55 60 65 70
02
46
810
12
Height (ft)
Num
ber
Time 6
60 65 70 75 80
05
1015
Height (ft)
Num
ber
Time 7
70 75 80 85
05
1015
Height (ft)N
umbe
r
Time 8
75 80 85 90
05
1015
Height (ft)
Num
ber
Time 9
80 85 90 95 100
05
1015
Height (ft)
Num
ber
Time 10
85 90 95 100 105
05
1015
Height (ft)
Num
ber
Time 11
90 95 100 110
05
1015
Height (ft)
Num
ber
Time 12
Figure 6.54. Histograms of tree height over time.

187
df wh ch
010
2030
Time 1
df wh ch0
1020
30
Time 2
df wh ch
010
2030
Time 3
df wh ch
010
2030
Time 4
df wh ch
010
2030
Time 5
df wh ch
010
2030
Time 6
df wh ch
010
2030
Time 7
df wh ch
010
2030
Time 8
df wh ch
010
2030
Time 9
df wh ch
010
2030
Time 10
df wh ch
010
2030
Time 11
df wh ch
010
2030
Time 12
Figure 6.55. Histogram of species number over time.

188
*
**
*
*
*******
Trees per acre
Qua
drat
ic M
ean
Dia
met
er
1 10 100 1000
0.1
0.5
1.0
5.0
50.0
Hugo Peak Stand - Density Management Diagram
In english unitsTime
Mai
and
Pai
2 4 6 8 10 12
050
010
0015
00
Hugo Peak Stand - Mai and Pai vs. Time
Figure 6.56: Density management diagram for the plot and a plot of mean annualincrement and periodic annual increment.
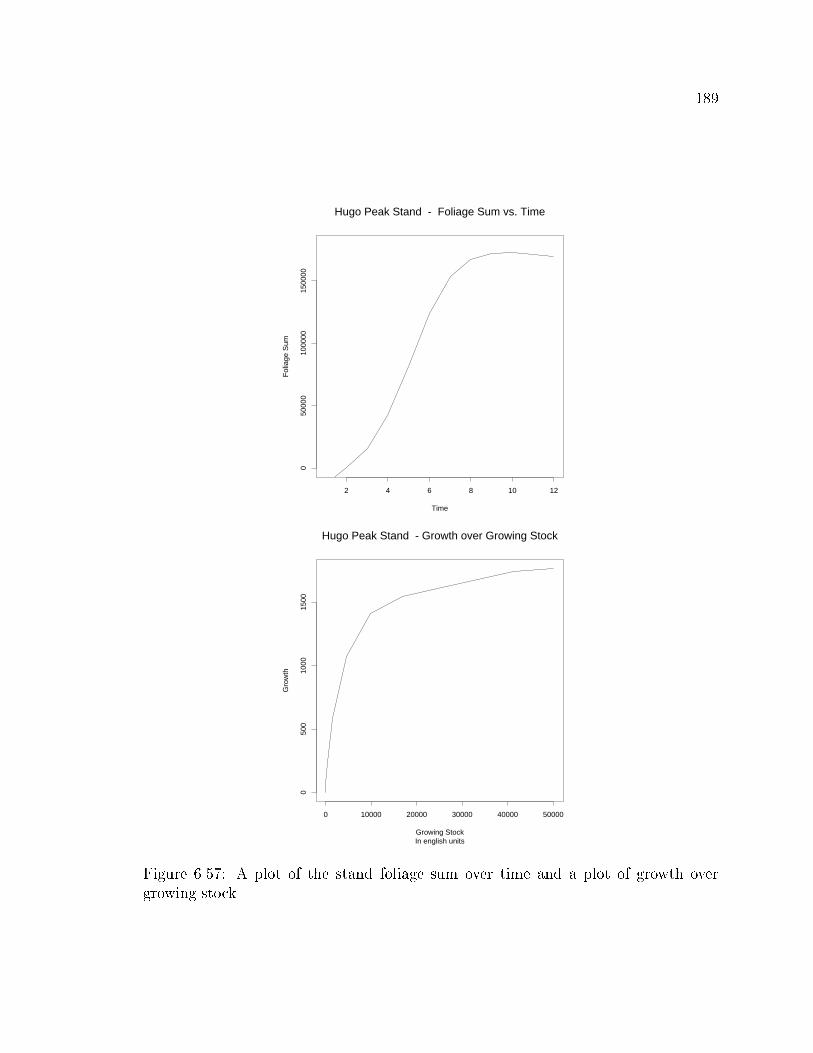
189
Time
Fol
iage
Sum
2 4 6 8 10 12
050
000
1000
0015
0000
Hugo Peak Stand - Foliage Sum vs. Time
In english unitsGrowing Stock
Gro
wth
0 10000 20000 30000 40000 50000
050
010
0015
00
Hugo Peak Stand - Growth over Growing Stock
Figure 6.57: A plot of the stand foliage sum over time and a plot of growth overgrowing stock.

190
6.8.4 Hugo Peak Stand, planted origin, 200 trees per acre with regeneration
This model run represents the actual sequence of events that the original stand under-
went and produces a stand most similar to the actual. In this run, a stand is planted
to 200 trees per acre and regeneration is allowed. The surviving regeneration is near
the edge of the stand, producing a \doughnut" e�ect in the stand maps later in the
growth period (Figure 6.55).
Some interesting species dynamics can be seen in the plots of the mean growth
by species over time. The height histograms over time show a pattern that is similar
to what might be seen in the original stand (Figure 6.66). The species mix changes
even greater because of the di�erential survival parameters for the di�erent species
(Figure 6.67). The ingrowth is apparent in the density management diagram (Figure
6.68). The solid line in this diagram is a stand density index of 595, which is con-
sidered the maximum for Douglas-�r. The model is predicting densities higher that
observed, but the behavior of the stand has been simulated with only the simplest of
mortality assumptions. Additionally, this stand is even further than the other runs
from culmination.
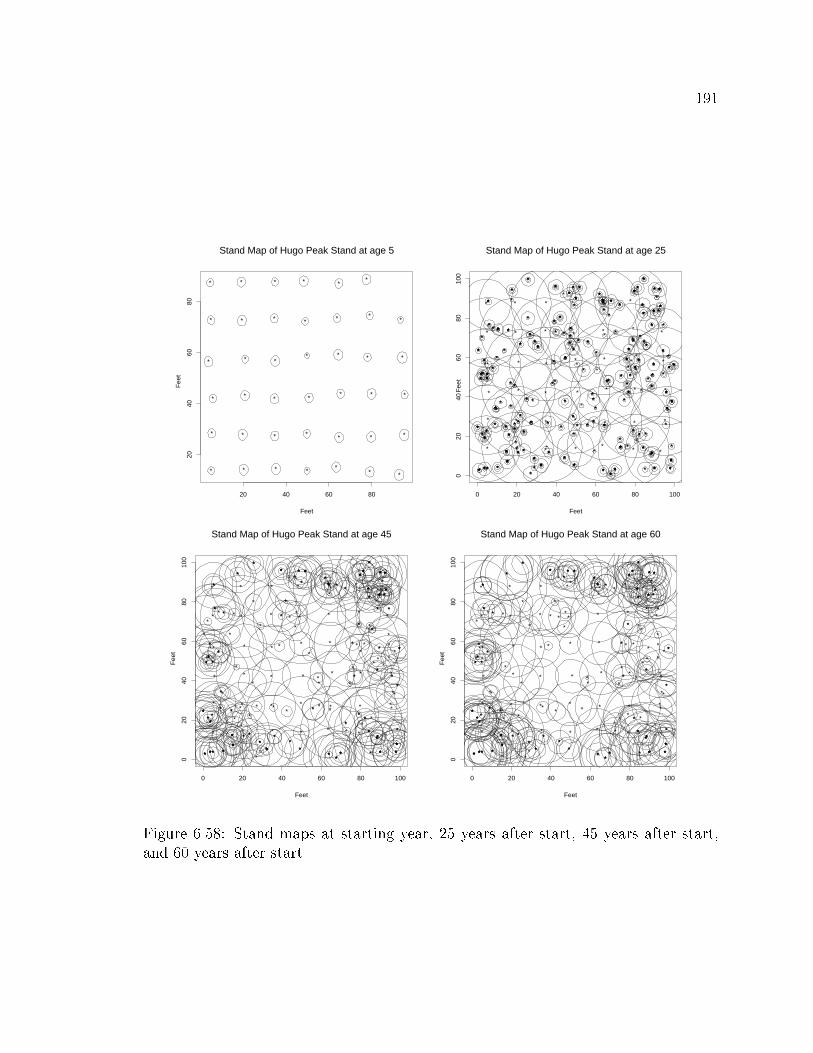
191
Feet
20 40 60 80
2040
6080
Fee
t
* * * * ** *
* * * * * * *
* * * ** * *
* * ** * * *
* * * * * **
* * * * **
Stand Map of Hugo Peak Stand at age 5
Feet
0 20 40 60 80 100
020
4060
8010
0F
eet
* * * * * * *
* * * * * * *
* * * * * * *
* * ** * * *
* * * * * * *
* * * * * *
*
*
*
*
*
*
*
*
*
* *
*
*
**
*
**
*
*
*
**
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
***
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
***
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
***
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
**
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
***
*
*
*
*
*
*
*
*
*
*
**
*
* *
*
*
*
**
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
*
*
* *
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
**
*
* *
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
***
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
*
Stand Map of Hugo Peak Stand at age 25
Feet
0 20 40 60 80 100
020
4060
8010
0
Fee
t
* * * * * * *
* * * * * * *
* * * * * * *
* * ** * * *
* * * * * * *
* * * * * *
*
*
*
*
*
*
* *
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
***
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
***
*
**
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
* *
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
***
*
*
**
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
* *
**
*
**
*
*
*
*
*
*
*
*
*
**
**
*
*
*
*
*
*
*
*
*
*
*
***
*
*
*
*
*
** *
*
*
*
*
*
Stand Map of Hugo Peak Stand at age 45
Feet
0 20 40 60 80 100
020
4060
8010
0
Fee
t
* * * * * * *
* * * * * * *
* * * * * * *
* * ** * * *
* * * * * * *
* * * * * *
*
*
*
*
*
*
* *
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
***
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
**
*
*
*
*
*
***
*
**
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
***
*
*
**
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
**
** *
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
***
*
*
*
** *
*
*
*
*
Stand Map of Hugo Peak Stand at age 60
Figure 6.58: Stand maps at starting year, 25 years after start, 45 years after start,and 60 years after start.

192
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
80
Stand Profile of Hugo Peak Stand at age 5
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 25
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 45
Distance (ft)
Hei
ght (
ft)
0 50 100
020
4060
8010
0
Stand Profile of Hugo Peak Stand at age 60
Figure 6.59: Stand pro�le at starting year, 25 years after start, 45 years after start,and 60 years after start.

193
Hugo Peak Stand - Tree 37
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 37
Rings are 5 years apart in inchesRadius (in)
Rad
ius
(in)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.60: Tree pro�le and diameter growth for a dominant tree. The radii are ininches and the concentric circles denote �ve year radial growth.

194
Hugo Peak Stand - Tree 26
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 26
Rings are 5 years apart in inchesRadius (in)
Rad
ius
(in)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.61: Tree pro�le and diameter growth for a co-dominant tree. The radii arein inches and the concentric circles denote �ve year radial growth.

195
Hugo Peak Stand - Tree 24
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 24
Rings are 5 years apart in inchesRadius (in)
Rad
ius
(in)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.62: Tree pro�le and diameter growth for an intermediate tree. The radii arein inches and the concentric circles denote �ve year radial growth.

196
Time (5 yr)
Dia
met
er a
t bre
ast h
eigh
t (in
)
2 4 6 8 10 12
010
2030
40
Hugo Peak Stand - Diameter vs. Time
Time (5 yr)
Dia
met
er a
t bre
ast h
eigh
t (in
)
2 4 6 8 10 12
02
46
810
1214
Hugo Peak Stand - Diameter vs. Time
Totaldfwhch
Figure 6.63: Diameter growth for each tree over time (upper plot) and the meandiameter growth of each species (lower plot). df = Douglas-�r, wh = western hemlock,and ch = cherry.

197
Time (5 yr)
Hei
ght (
ft)
2 4 6 8 10 12
020
4060
8010
0
Hugo Peak Stand - Height vs. Time
Time (5 yr)
Hei
ght (
ft)
2 4 6 8 10 12
020
4060
80
Hugo Peak Stand - Height vs. Time
Totaldfwhch
Figure 6.64: Height growth for each tree over time (upper plot) and the mean heightgrowth of each species (lower plot).

198
Time (5 yr)
Hei
ght t
o cr
own
base
(ft)
2 4 6 8 10 12
020
4060
80
Hugo Peak Stand - Height to crown base vs. Time
Time (5 yr)
Hei
ght t
o cr
own
base
(ft)
2 4 6 8 10 12
020
4060
Hugo Peak Stand - Height to crown base vs. Time
Totaldfwhch
Figure 6.65: Height to crown base change for each tree over time (upper plot) andthe mean height to crown base change for each species (lower plot).

199
Time (5 yr)
Cro
wn
wid
th (
ft)
2 4 6 8 10 12
010
2030
4050
Hugo Peak Stand - Crown width vs. Time
Time (5 yr)
Cro
wn
wid
th (
ft)
2 4 6 8 10 12
510
15
Hugo Peak Stand - Crown width vs. Time
Totaldfwhch
Figure 6.66: Crown width for each tree over time (upper plot) and the mean crownwidth for each species (lower plot).

200
Time (5 yr)
Fol
iage
sur
face
are
a (s
q ft)
2 4 6 8 10 12
020
0040
0060
0080
00
Hugo Peak Stand - Foliage surface area vs. Time
Time (5 yr)
Fol
iage
sur
face
are
a (s
q ft)
2 4 6 8 10 12
050
010
0015
00
Hugo Peak Stand - Foliage surface area vs. Time
Totaldfwhch
Figure 6.67: Foliage leaf area for each tree over time (upper plot) and the mean foliageleaf area for each species (lower plot).

201
Time (5 yr)
Hei
ght/d
iam
eter
(P
erce
nt)
2 4 6 8 10 12
020
0040
0060
00
Hugo Peak Stand - Height/diameter vs. Time
Figure 6.68. Height/diameter ratio change for each tree over time.

202
4.0 5.0 6.0
01
23
45
Height (ft)
Num
ber
Time 1
0 5 10 15 20
010
020
030
0
Height (ft)
Num
ber
Time 2
0 10 20 30
020
040
060
0
Height (ft)
Num
ber
Time 3
0 10 20 30 40 50
020
040
060
0
Height (ft)
Num
ber
Time 4
0 10 30 50
010
020
030
040
0
Height (ft)
Num
ber
Time 5
0 20 40 60
050
100
150
200
Height (ft)
Num
ber
Time 6
0 20 40 60 80
020
4060
8012
0
Height (ft)
Num
ber
Time 7
0 20 40 60 80
020
4060
80
Height (ft)N
umbe
r
Time 8
0 20 40 60 80 100
020
4060
80
Height (ft)
Num
ber
Time 9
20 40 60 80 100
020
4060
80
Height (ft)
Num
ber
Time 10
20 40 60 80 100
020
4060
80
Height (ft)
Num
ber
Time 11
40 60 80 100 120
020
4060
80
Height (ft)
Num
ber
Time 12
Figure 6.69. Histograms of tree height over time.

203
df wh ch
010
2030
Time 1
df wh ch
050
100
150
Time 2
df wh ch
010
020
030
0
Time 3
df wh ch
010
020
030
0
Time 4
df wh ch
010
020
030
0
Time 5
df wh ch
050
150
250
Time 6
df wh ch
050
150
250
Time 7
df wh ch
050
150
250
Time 8
df wh ch
050
150
Time 9
df wh ch
050
100
200
Time 10
df wh ch
050
100
200
Time 11
df wh ch
050
100
200
Time 12
Figure 6.70. Histogram of species number over time.

204
*
* *
*
*
*
**
****
Trees per acre
Qua
drat
ic M
ean
Dia
met
er
1 10 100 1000
0.1
0.5
1.0
5.0
50.0
Hugo Peak Stand - Density Management Diagram
Time (5 yr)
Mai
and
Pai
(cu
bic
ft)
2 4 6 8 10 12
050
010
0015
0020
0025
00
Hugo Peak Stand - Mai and Pai vs. Time
Figure 6.71: Density management diagram for the plot and a plot of mean annualincrement and periodic annual increment.

205
Time (5 yr)
Fol
iage
Sum
(sq
ft)
2 4 6 8 10 12
010
0000
2000
0030
0000
4000
00
Hugo Peak Stand - Foliage Sum vs. Time
Growing Stock (cubic ft)
Gro
wth
(cu
bic
ft)
0 10000 20000 30000 40000 50000 60000
050
010
0015
0020
0025
00
Hugo Peak Stand - Growth over Growing Stock
Figure 6.72: A plot of the stand foliage sum over time and a plot of growth overgrowing stock.

206
6.8.5 Bethel Ridge Stand
This stand is included as an example of a stand measured in metric units with similar
conditions to the Hugo Peak stand, but a little older. The example was run with no
regeneration allowed. The behavior of the stand follow the general pattern found in
the Hugo peak runs. Again the maps show the thinning of the crowns in the center
of the stand because of the greater amount of competition. Interestingly, the height
distributions remain relatively consistent shape throughout the entire run.
The Bethel Ridge stand was include to illustrate several points. First, the model
can be parameterized with metric data as easily as with English units. Additionally,
it was included to illustrate the type of data that can be used to produce parameter
estimates.

207
Meters
0 10 20 30 40 50
010
2030
4050
Met
ers
*
*
*
*
* *
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
***
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
*
* *
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
**
Stand Map of Bethel Ridge Stand at age 5
Meters
0 10 20 30 40 50
010
2030
4050
Met
ers
*
*
*
*
* *
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
***
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
*
* *
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
**
Stand Map of Bethel Ridge Stand at age 25
Meters
0 10 20 30 40 50
010
2030
4050
Met
ers
*
*
*
*
* *
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
***
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
*
* *
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
**
Stand Map of Bethel Ridge Stand at age 45
Meters
0 10 20 30 40 50
010
2030
4050
Met
ers
*
*
*
*
* *
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
***
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
* *
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
*
**
*
**
*
*
*
* *
*
* **
*
*
*
*
*
*
*
*
*
*
*
*
**
*
*
*
*
*
**
*
*
*
*
**
Stand Map of Bethel Ridge Stand at age 60
Figure 6.73: Stand maps at starting year, 25 years after start, 45 years after start,and 60 years after start.

208
Distance (m)
Hei
ght (
m)
-20 0 20 40 60
010
2030
4050
Stand Profile of Bethel Ridge Stand at age 5
Distance (m)
Hei
ght (
m)
-20 0 20 40 60
010
2030
4050
Stand Profile of Bethel Ridge Stand at age 25
Distance (m)
Hei
ght (
m)
-20 0 20 40 60
010
2030
4050
Stand Profile of Bethel Ridge Stand at age 45
Distance (m)
Hei
ght (
m)
-20 0 20 40 60
010
2030
4050
Stand Profile of Bethel Ridge Stand at age 60
Figure 6.74: Stand pro�le at starting year, 25 years after start, 45 years after start,and 60 years after start.

209
Bethel Ridge Stand - Tree 201
Time (5 yr)
Hei
ght (
m)
010
2030
4050
60
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 201
Rings are 5 years apart in centimetersRadius (cm)
Rad
ius
(cm
)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.75: Tree pro�le and diameter growth for a dominant tree. The radii are incentimeters. The concentric circle denote �ve year radial growth.

210
Bethel Ridge Stand - Tree 205
Time (5 yr)
Hei
ght (
m)
010
2030
4050
60
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 205
Rings are 5 years apart in centimetersRadius (cm)
Rad
ius
(cm
)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.76: Tree pro�le and diameter growth for a co-dominant tree. The radii arein centimeters. The concentric circle denote �ve year radial growth.

211
Bethel Ridge Stand - Tree 153
Time (5 yr)
Hei
ght (
m)
010
2030
4050
60
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 153
Rings are 5 years apart in centimetersRadius (cm)
Rad
ius
(cm
)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.77: Tree pro�le and diameter growth for an intermediate tree. The radii arein centimeters. The concentric circle denote �ve year radial growth.

212
Time (5 yr)
Dia
met
er a
t bre
ast h
eigh
t (cm
)
2 4 6 8 10 12
010
2030
40
Bethel Ridge Stand - Diameter vs. Time
Time (5 yr)
Dia
met
er a
t bre
ast h
eigh
t (cm
)
2 4 6 8 10 12
05
1015
20
Bethel Ridge Stand - Diameter vs. Time
Totalpsmetshe
Figure 6.78: Diameter growth for each tree over time (upper plot) and the meandiameter growth of each species (lower plot). psme = Douglas-�r, and tshe = westernhemlock.

213
Time (5 yr)
Hei
ght (
m)
2 4 6 8 10 12
1020
30
Bethel Ridge Stand - Height vs. Time
Time (5 yr)
Hei
ght (
m)
2 4 6 8 10 12
1020
30
Bethel Ridge Stand - Height vs. Time
Totalpsmetshe
Figure 6.79: Height growth for each tree over time (upper plot) and the mean heightgrowth of each species (lower plot).

214
Time (5 yr)
Hei
ght t
o cr
own
base
(m
)
2 4 6 8 10 12
010
2030
Bethel Ridge Stand - Height to crown base vs. Time
Time (5 yr)
Hei
ght t
o cr
own
base
(m
)
2 4 6 8 10 12
05
1015
2025
30
Bethel Ridge Stand - Height to crown base vs. Time
Totalpsmetshe
Figure 6.80: Height to crown base change for each tree over time (upper plot) andthe mean height to crown base change for each species (lower plot).

215
Time (5 yr)
Cro
wn
wid
th (
m)
2 4 6 8 10 12
510
1520
Bethel Ridge Stand - Crown width vs. Time
Time (5 yr)
Cro
wn
wid
th (
m)
2 4 6 8 10 12
24
68
Bethel Ridge Stand - Crown width vs. Time
Totalpsmetshe
Figure 6.81: Crown width for each tree over time (upper plot) and the mean crownwidth for each species (lower plot).

216
Time (5 yr)
Fol
iage
sur
face
are
a (s
q m
)
2 4 6 8 10 12
020
040
060
080
010
00
Bethel Ridge Stand - Foliage surface area vs. Time
Time (5 yr)
Foi
lage
sur
face
are
a (s
q m
)
2 4 6 8 10 12
5010
015
020
0
Bethel Ridge Stand - Foliage surface area vs. Time
Totalpsmetshe
Figure 6.82: Foliage leaf area for each tree over time (upper plot) and the mean foliageleaf area for each species (lower plot).

217
Time (5 yr)
Hei
ght/d
iam
eter
(P
erce
nt)
2 4 6 8 10 12
200
400
600
800
1000
Bethel Ridge Stand - Height/diameter vs. Time
Time (5 yr)
Hei
ght/d
iam
eter
(P
erce
nt)
2 4 6 8 10 12
200
300
400
500
600
Bethel Ridge Stand - Height/diameter vs. Time
Totalpsmetshe
Figure 6.83: Height/diameter ratio change for each tree over time (upper plot) andthe mean height/diameter ratio for each species (lower plot).

218
.
.
.
........
.
.
.............
.
.....
.
.
.
.......
.....
.
.
.
.
........
.
.
.........
.......
...
................
.
.........
.
.
...
.....
.
..
............
......
.
.
.
.
.
.
.
.
.............................................................
.
.
.
...............
.
...
Rank
Rel
ativ
e ht
0 50 100 150 200
01
23
4
Time 1
....................................................................................................................................................................................................................................
Rank
Rel
ativ
e ht
0 50 100 150 200
02
46
8
Time 2
.
..........................................................
.
..........................................................................
.
.
...........................
.............................................................
.
...
Rank
Rel
ativ
e ht
0 50 100 150 200
02
46
810
12
Time 3
.
..........................................................
.
......................................
.
...................
...........................
.....................
.
.............................................................
Rank
Rel
ativ
e ht
0 50 100 150 200
05
1015
Time 4
...........................................................
...........................................................................
.........................
.
.........................
.
.
.
........................
................
Rank
Rel
ativ
e ht
0 50 100 150 200
05
1015
20
Time 5
.................................................................
.................................
.
....................................
..................................................
.
.
.
........................................
Rank
Rel
ativ
e ht
0 50 100 150 200
05
1015
20
Time 6
........................................
..........................................................
.
........................................
..............................................
.
.
.
........................................
Rank
Rel
ativ
e ht
0 50 100 150 200
05
1015
2025
Time 7
..........................................
....................
..........................................
.......................................
.................................
..........................
..........................
RankR
elat
ive
ht
0 50 100 150 200
05
1015
2025
30
Time 8
...............................................
.......................
..................................
.......................................
..............................................................
.......................
Rank
Rel
ativ
e ht
0 50 100 150 200
05
1015
2025
30
Time 9
...........................................................
............................................
..........................................
................................
...................................................
Rank
Rel
ativ
e ht
0 50 100 150 200
010
2030
Time 10
..............................................................
.........................................
....................................................
..............................................................
...........
Rank
Rel
ativ
e ht
0 50 100 150 200
010
2030
Time 11
........................................................................................................
...................................................
.
........................................................................
Rank
Rel
ativ
e ht
0 50 100 150 200
010
2030
Time 12
Figure 6.84. Plot of ordered trees over time. The order time is time 1.

219
2.0 3.0 4.0
010
2030
Height (m)
Num
ber
Time 1
5 6 7 8 90
2040
60
Height (m)
Num
ber
Time 2
8 9 10 11 12 13
020
4060
Height (m)
Num
ber
Time 3
12 13 14 15 16 17
010
2030
4050
60
Height (m)
Num
ber
Time 4
16 17 18 19 20
020
4060
Height (m)
Num
ber
Time 5
19 20 21 22 23 24
010
2030
4050
60
Height (m)
Num
ber
Time 6
23 24 25 26 27
010
2030
4050
60
Height (m)
Num
ber
Time 7
25 26 27 28 29
020
4060
Height (m)
Num
ber
Time 8
28 29 30 31 32
010
2030
4050
60
Height (m)
Num
ber
Time 9
30 31 32 33 34
020
4060
Height (m)
Num
ber
Time 10
33 34 35 36
020
4060
80
Height (m)
Num
ber
Time 11
35 36 37 38
020
4060
80
Height (m)
Num
ber
Time 12
Figure 6.85. Histograms of tree height over time.

220
psme tshe
050
100
150
200
Time 1
psme tshe
050
100
150
200
Time 2
psme tshe
050
100
150
200
Time 3
psme tshe
050
100
150
200
Time 4
psme tshe
050
100
150
200
Time 5
psme tshe
050
100
150
200
Time 6
psme tshe
050
100
150
200
Time 7
psme tshe
050
100
150
200
Time 8
psme tshe
050
100
150
200
Time 9
psme tshe
050
100
150
200
Time 10
psme tshe
050
100
150
200
Time 11
psme tshe
050
100
150
200
Time 12
Figure 6.86. Histogram of species number over time.

221
*
*
*
*
********
Trees per hectare
Qua
drat
ic M
ean
Dia
met
er
1 10 100 1000 10000
15
1050
100
500
Bethel Ridge Stand - Density Management Diagram
Time (5 yr )
Mai
and
Pai
(cu
bic
m)
2 4 6 8 10 12
020
040
060
080
0
Bethel Ridge Stand - Mai and Pai vs. Time
Figure 6.87: Density management diagram for the plot and a plot of mean annualincrement and periodic annual increment.
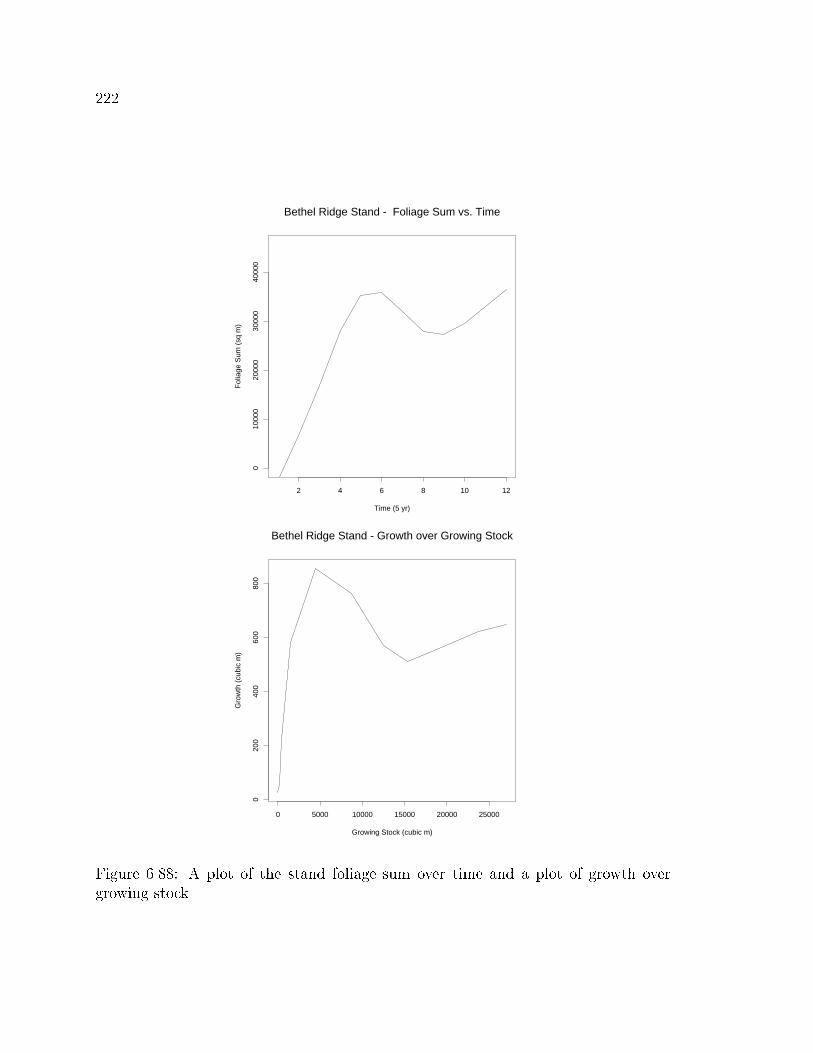
222
Time (5 yr)
Fol
iage
Sum
(sq
m)
2 4 6 8 10 12
010
000
2000
030
000
4000
0
Bethel Ridge Stand - Foliage Sum vs. Time
Growing Stock (cubic m)
Gro
wth
(cu
bic
m)
0 5000 10000 15000 20000 25000
020
040
060
080
0
Bethel Ridge Stand - Growth over Growing Stock
Figure 6.88: A plot of the stand foliage sum over time and a plot of growth overgrowing stock.

223
6.8.6 Helena Plots
The Helena plots illustrate the use of the model with data from a very di�erent
area and very di�erent tree species. The idea is to simulate the stand's history to
try to predict a stand that appeared similar to the one measured. The attempt was
somewhat successful. The assumption of a constant crown length to crown width ratio
is not applicable here, and more dynamic crown assumptions are needed. Outside of
this assumption, the model appears to work reasonably with these data.
In this run, the events of the original experiment were simulated. Four row of
Cherrybark oak were established next to four rows of american sycamore and then
four empty rows representing the space in which cottonwood were planted and died.
The run did re ect the superior height growth of the american sycamore but did
not clearly re ect the di�erential size pattern observed in the original stands (Oliver
et al., 1990).
The height growth patterns di�erentiated between the two species but not enough
variation was input into each species height growth pattern. The cherrybark oak
experienced a higher mortality rate than the sycamore which is re ected in the species
histograms in Figure 6.102.

224
Feet
0 20 40 60 80 100
2040
6080
Fee
t
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * *
Stand Map of Helena plots
Feet
0 20 40 60 80 100
2040
6080
Fee
t
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * * *
* * * * * * * *
Stand Map of Helena plots
Feet
0 20 40 60 80 100
2040
6080
Fee
t
* * * * * * * *
* * * * * * * * *
* * * * * * * *
* * * * * * *
* * * * * * * *
* * * * * * * *
* * * * * * * * *
* * * * * * *
* * * * * * * * *
* * * * * * * *
* * * * * * * * *
* * * * * * * *
Stand Map of Helena plots
Feet
0 20 40 60 80 100
2040
6080
Fee
t
* * * * * * * *
* * * * * * * * *
* * * * * * * *
* * * * * * *
* * * * * * * *
* * * * * * * *
* * * * * * * * *
* * * * * * *
* * * * * * * * *
* * * * * * *
* * * * * * * * *
* * * * * * * *
Stand Map of Helena plots
Figure 6.89: Stand maps at starting year, 25 years after start, 45 years after start,and 60 years after start. The four rows on the left are cherrybark oak the four centerrows are sycamore and the row on the far right is again cherrybark oak.

225
Distance (ft)
Hei
ght (
ft)
-20 0 20 40 60 80 100 120
020
4060
8010
0
Stand Profile of Helena plots
Distance (ft)
Hei
ght (
ft)
0 50 100
020
4060
8010
0
Stand Profile of Helena plots
Distance (ft)
Hei
ght (
ft)
0 50 100 150
020
4060
8010
012
014
0
Stand Profile of Helena plots
Distance (ft)
Hei
ght (
ft)
0 50 100 150
050
100
150
Stand Profile of Helena plots
Figure 6.90: Stand pro�le at starting year, 25 years after start, 45 years after start,and 60 years after start. The four rows on the left are cherrybark oak the four centerrows are sycamore and the row on the far right is again cherrybark oak.

226
Helena plots - Tree 62
Time (5 yr)
Hei
ght (
ft)
050
100
150
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 62
Rings are 5 years apart in inchesRadius (in)
Rad
ius
(in)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.91: Tree pro�le and diameter growth for a dominant tree. This tree is asycamore.

227
Helena plots - Tree 75
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
014
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 75
Rings are 5 years apart in inchesRadius (in)
Rad
ius
(in)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.92: Tree pro�le and diameter growth for a co-dominant tree. This tree is ancherrybark oak.
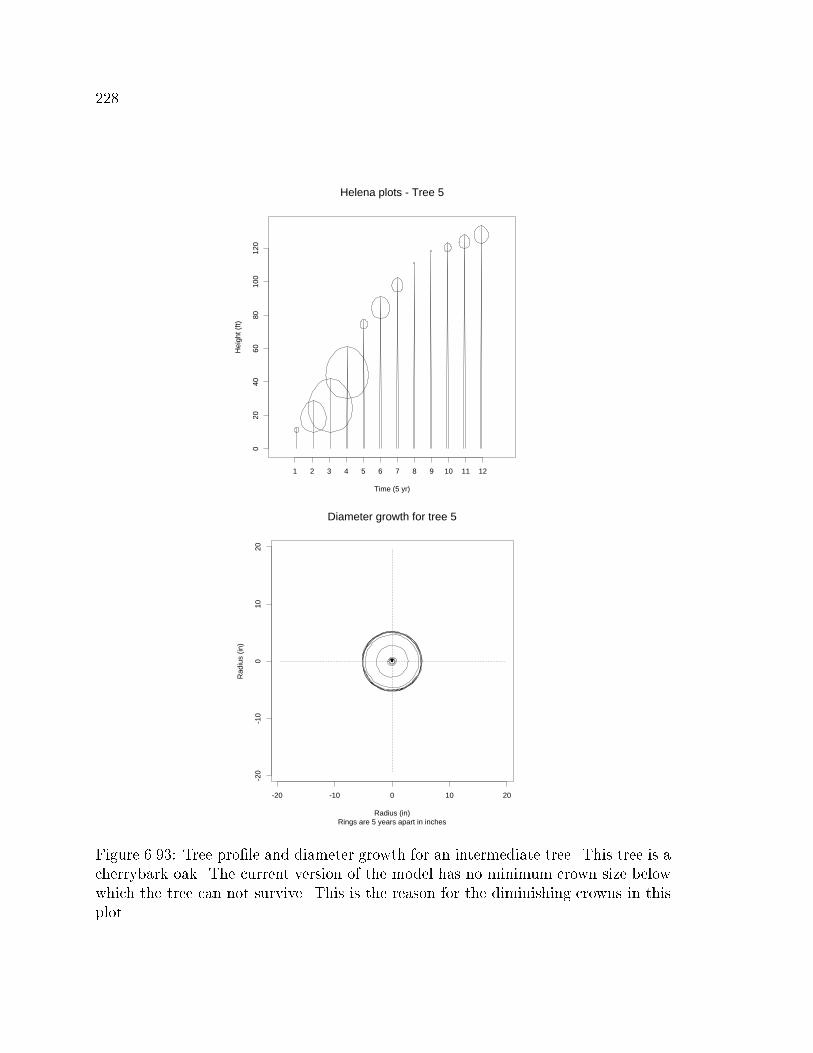
228
Helena plots - Tree 5
Time (5 yr)
Hei
ght (
ft)
020
4060
8010
012
0
1 2 3 4 5 6 7 8 9 10 11 12
************
Diameter growth for tree 5
Rings are 5 years apart in inchesRadius (in)
Rad
ius
(in)
-20 -10 0 10 20
-20
-10
010
20
Figure 6.93: Tree pro�le and diameter growth for an intermediate tree. This tree is acherrybark oak. The current version of the model has no minimum crown size belowwhich the tree can not survive. This is the reason for the diminishing crowns in thisplot.

229
Time (5 yr)
Dia
met
er a
t bre
ast h
eigh
t (in
)
2 4 6 8 10 12
010
2030
40
Helena plots - Diameter vs. Time
Time (5 yr)
Dia
met
er a
t bre
ast h
eigh
t (in
)
2 4 6 8 10 12
510
1520
25
Helena Plots - Diameter vs. Time
Totalsyccbo
Figure 6.94: Diameter growth for each tree over time (upper plot) and the meandiameter growth of each species (lower plot). syc = American sycamore, and cbo =cherrybark oak.

230
Time (5 yr)
Hei
ght (
ft)
2 4 6 8 10 12
2040
6080
100
120
140
160
Helena plots - Height vs. Time
Time (5 yr)
Hei
ght (
ft)
2 4 6 8 10 12
2040
6080
100
120
140
Helena Plots - Height vs. Time
Totalsyccbo
Figure 6.95: Height growth for each tree over time (upper plot) and the mean heightgrowth of each species (lower plot).
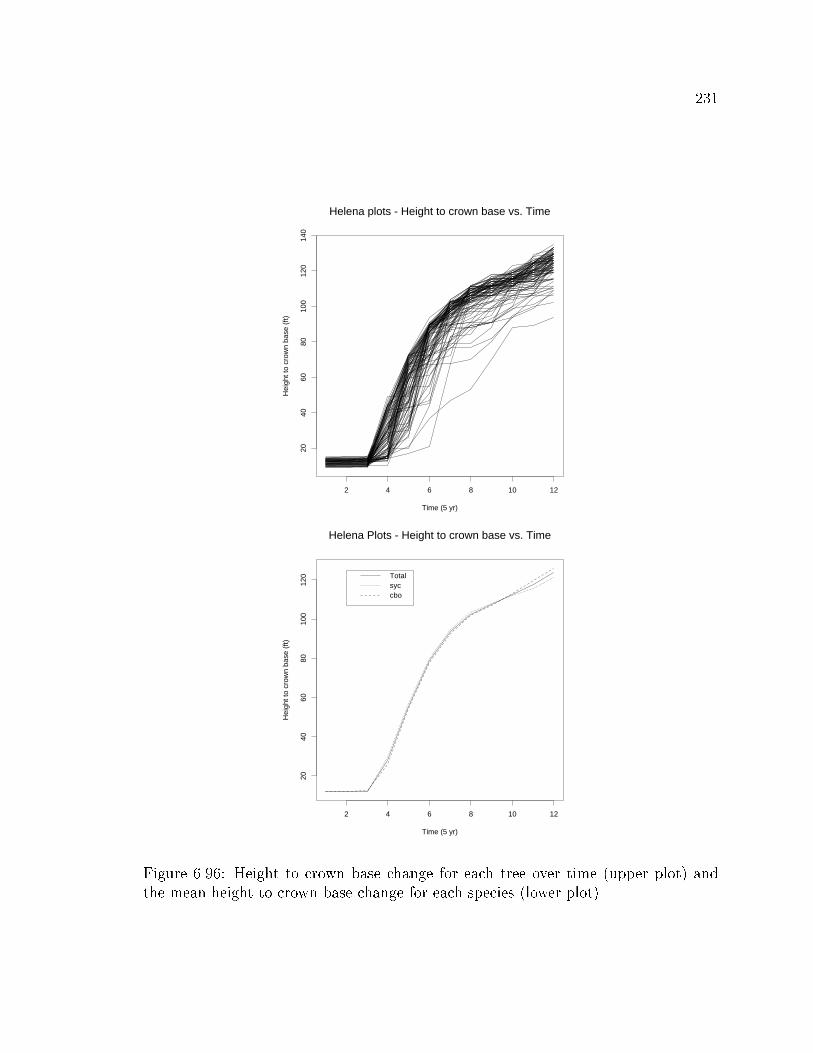
231
Time (5 yr)
Hei
ght t
o cr
own
base
(ft)
2 4 6 8 10 12
2040
6080
100
120
140
Helena plots - Height to crown base vs. Time
Time (5 yr)
Hei
ght t
o cr
own
base
(ft)
2 4 6 8 10 12
2040
6080
100
120
Helena Plots - Height to crown base vs. Time
Totalsyccbo
Figure 6.96: Height to crown base change for each tree over time (upper plot) andthe mean height to crown base change for each species (lower plot).

232
Time (5 yr)
Cro
wn
wid
th (
ft)
2 4 6 8 10 12
010
2030
4050
60
Helena plots - Crown width vs. Time
Time (5 yr)
Cro
wn
wid
th (
ft)
2 4 6 8 10 12
1020
3040
Helena Plots - Crown width vs. Time
Totalsyccbo
Figure 6.97: Crown width for each tree over time (upper plot) and the mean crownwidth for each species (lower plot).
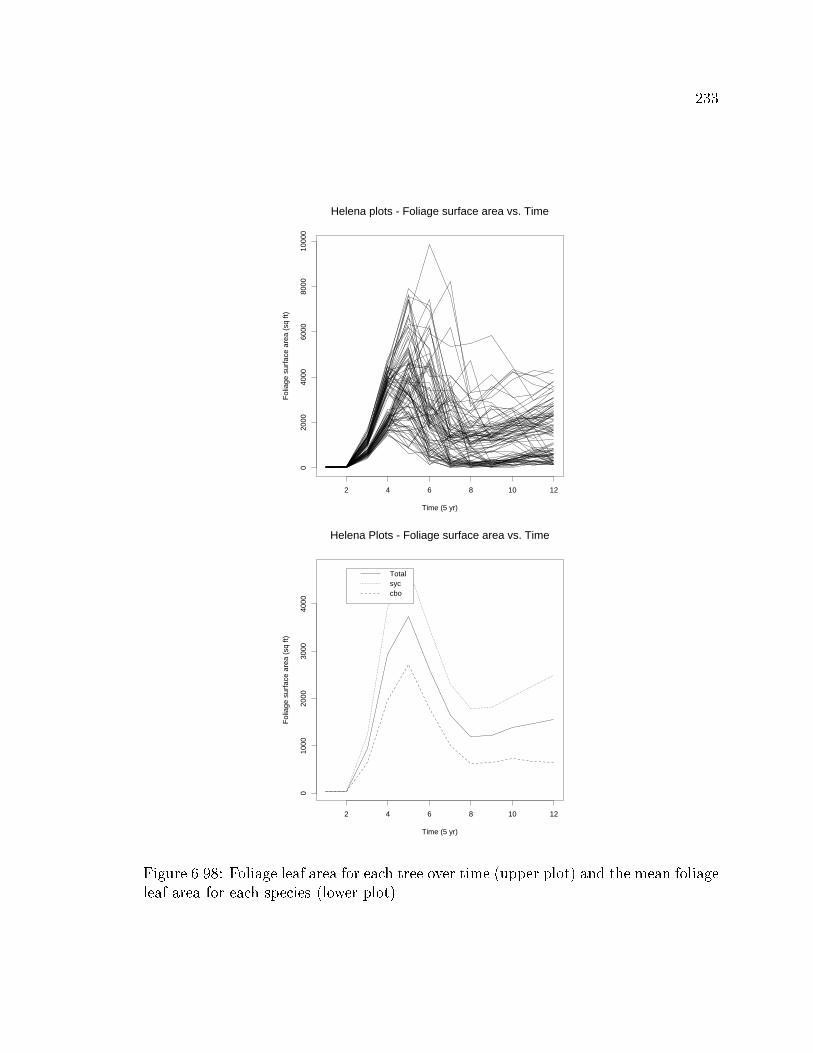
233
Time (5 yr)
Fol
iage
sur
face
are
a (s
q ft)
2 4 6 8 10 12
020
0040
0060
0080
0010
000
Helena plots - Foliage surface area vs. Time
Time (5 yr)
Fol
iage
sur
face
are
a (s
q ft)
2 4 6 8 10 12
010
0020
0030
0040
00
Helena Plots - Foliage surface area vs. Time
Totalsyccbo
Figure 6.98: Foliage leaf area for each tree over time (upper plot) and the mean foliageleaf area for each species (lower plot).

234
Time (5 yr)
Hei
ght/d
iam
eter
(P
erce
nt)
2 4 6 8 10 12
100
200
300
400
Helena plots - Height/diameter vs. Time
Time (5 yr)
Hei
ght/d
iam
eter
(P
erce
nt)
2 4 6 8 10 12
100
150
200
250
300
350
Helena Plots - Height/diameter vs. Time
Totalsyccbo
Figure 6.99: Height/diameter ratio change for each tree over time (upper plot) andthe mean height/diameter ratio for each species (lower plot).

235
.....
.
....
...
.........
......
.
.
.
....
.......
.
.
..
.
.....
....
.......
..
................
.
.....
.........
........
...
Rank
Rel
ativ
e ht
0 20 40 60 80
05
1015
20Time 1
..........................................................................................................
.
Rank
Rel
ativ
e ht
0 20 40 60 800
1020
3040
50
Time 2
..........................
................
.
.
..
.
.....
..........
..............................
...............
Rank
Rel
ativ
e ht
0 20 40 60 80
020
4060
Time 3
.......................................
...
.
.
..
.
.....
....
....................................
...............
Rank
Rel
ativ
e ht
0 20 40 60 80
020
4060
80
Time 4
...........................
...............
.
.
..
.
.....
....
...................................................
Rank
Rel
ativ
e ht
0 20 40 60 80
020
4060
8010
0
Time 5
...................................
.......
.
.
..
.
.....
....
....................................
...............
Rank
Rel
ativ
e ht
0 20 40 60 80
020
4060
8010
0
Time 6
..........................................
.
.
..
.
.....
....
................................
............
.
.....
.
Rank
Rel
ativ
e ht
0 20 40 60 80
020
4060
8010
0
Time 7
..........................................
.
.
..
.
.....
....................................
............
.
.....
.
Rank
Rel
ativ
e ht
0 20 40 60 80
020
4060
8010
0
Time 8
..........................................
.
.
...
.....
.........................
......
.....
..
...
.
......
.
..
.
..
.
Rank
Rel
ativ
e ht
0 20 40 60 80
020
4060
8012
0
Time 9
..........................................
.
.
...
.....
.........................
.
..........
..
...
.
......
.
..
.
..
.
Rank
Rel
ativ
e ht
0 20 40 60 80
020
4060
8012
0
Time 10
..........................................
.
....
.....
.........................
.
....
.
.....
..
...
.
......
.
..
.
..
.
Rank
Rel
ativ
e ht
0 20 40 60 80
050
100
150
Time 11
..........................................
.
...
.
.
.
...
.........................
.
....
.
.....
..
...
.
......
.
..
.
..
.
Rank
Rel
ativ
e ht
0 20 40 60 80
050
100
150
Time 12
Figure 6.100: Plot of ordered trees over time. The order time is time 2. Note the twolevels apparent in these plots

236
12 14 16 18 20
05
1015
2025
Height (ft)
Num
ber
Time 1
25 30 35 40 45 50
02
46
810
1214
Height (ft)
Num
ber
Time 2
40 50 60 70
05
1015
20
Height (ft)
Num
ber
Time 3
60 70 80 90
05
1015
2025
Height (ft)
Num
ber
Time 4
75 85 95 105
05
1015
Height (ft)
Num
ber
Time 5
90 95 105 115
05
1015
Height (ft)
Num
ber
Time 6
100 110 120
05
1015
Height (ft)
Num
ber
Time 7
110 120 130
05
1015
20
Height (ft)N
umbe
r
Time 8
115 125 135
05
1015
20
Height (ft)
Num
ber
Time 9
125 135 145
05
1015
2025
Height (ft)
Num
ber
Time 10
130 135 140 145 150
05
1015
20
Height (ft)
Num
ber
Time 11
135 145 155
05
1015
2025
30
Height (ft)
Num
ber
Time 12
Figure 6.101. Histograms of tree height over time. Note the bimodal distribution.

237
syc cbo
010
2030
4050
60
Time 1
syc cbo0
1020
3040
5060
Time 2
syc cbo
010
2030
4050
60
Time 3
syc cbo
010
2030
4050
60
Time 4
syc cbo
010
2030
4050
60
Time 5
syc cbo
010
2030
4050
Time 6
syc cbo
010
2030
4050
Time 7
syc cbo
010
2030
4050
Time 8
syc cbo
010
2030
4050
Time 9
syc cbo
010
2030
4050
Time 10
syc cbo
010
2030
4050
Time 11
syc cbo
010
2030
4050
Time 12
Figure 6.102. Histogram of species number over time.

238
***
*
********
Trees per acre
Qua
drat
ic M
ean
Dia
met
er
1 10 100 1000
0.1
0.5
1.0
5.0
50.0
Helena plots - Density Management Diagram
Time (5 yr)
Mai
and
Pai
(cu
bic
ft)
2 4 6 8 10 12
050
010
0015
0020
0025
00
Helena plots - Mai and Pai vs. Time
Figure 6.103: Density management diagram for the plot and a plot of mean annualincrement and periodic annual increment.

239
Time (5 yr)
Fol
iage
Sum
(sq
ft)
2 4 6 8 10 12
010
0000
2000
0030
0000
4000
00
Helena plots - Foliage Sum vs. Time
Growing Stock (cubic ft)
Gro
wth
(cu
bic
ft)
0 10000 20000 30000 40000 50000 60000
050
010
0015
0020
0025
00
Helena plots - Growth over Growing Stock
Figure 6.104: A plot of the stand foliage sum over time and a plot of growth overgrowing stock.

240
6.9 Discussion of the approach
The adaptive model of stand dynamics illustrates the potential of this approach; it
has many interesting advantages and disadvantages. Advantages include exibility,
ease of use with very di�erent types of input information, and ease of application
for adaptive silviculture. Disadvantages include an inability to represent some of
the subtler behavior exhibited by the modeled stands. Some of this is because of
the generalizations made by the approach and some of it is because of incompletely
speci�ed submodels. These submodels can be corrected. This inability to explain �ne
detail is to be expected and can be easily accommodated in the adaptive management
approach.
The main advantage of the approach to modeling is exibility. This exibility
however, carries the responsibility of carefully evaluating the output from any sim-
ulations. For example, a new graph of stand values was generated from stand data
within minutes. The exibility is in part because of the statistical package used for
display and in part because of the models modular design. The modularity and ex-
ibility �t well into the adaptive management approach. As functions, output, other
desired information are found lacking, they can be incorporated into future versions
of the model.
Another advantage is the graphical output, which can help a person evaluate a lot
of information quickly. The graphical model interface has been e�ective, and parts
of the graphical interface have been adapted for use with other forest growth models.
Graphical user output is not limited to adaptive models of stand dynamics; but the
output is very useful in accomplishing the objectives of adaptive silviculture.
A third advantage is the relatively few relationships needed to explain behaviors
exhibited in model runs. Twelve deterministic parameters per species are needed to
run this model, and another �fteen stochastic parameters are used to simulate the
static and error distributions in the input data.

241
The predictions reproduce the stands' maximum height growth behavior well. The
current data did not have any other measurements over time to compare to the pre-
dictions. This adaptive stand dynamics model is designed to utilize information from
expert opinions, to single time measurements, to repeated measurements. Only the
single time measurements are used in this dissertation. While adequate to parame-
terize the model and make simulations, the current data are not the best. Repeated
measurements would provide even more useful information that would allow better
parameter sets and more complete evaluation of the model output as the stand be-
comes older.
There are numerous areas of the model that can be improved. A de�nition of
the dynamics of the crown width to crown length is needed to produce more realistic
crowns. This would improve the predictions of hardwood species and possibly conifers.
The current static relationship of crown length to crown width may provide over
estimates of crown size.
The question of mortality should also be addressed. The current method based on
crown competition seems plausible in the early developmental stage of the stand, but
is inadequate in later development. Another area of improvement is the relationship
of foliage area to stem increment. Both this and the previous question would require
independent experiments to determine the best approach to satisfy these questions.
Additional improvement beyond these is probably not be needed, since it would
be beyond the scope of the questions that this type of model would be able to analyze.
These more detailed question would be better answered with detailed models focused
at speci�c questions and from which generalized behaviors can be determined for
stand level models presented here.

Chapter 7
A LARGER FOREST MANAGEMENT SYSTEM
\There are no \bigger secrets" because the moment a secret is revealed, it
seems little."
Umberto Eco, 1988, \Foucault's Pendulum", Chapter 118
The adaptive growth model presented here is only part of a larger forest manage-
ment system. The design of the adaptive stand dynamics model implies that the forest
manager will be monitoring a stand over time and using the additional information
collected at successive measurements to update the parameters of the growth model.
This type of model is designed for managers interested in the long term management
of forest stands.
Along with monitoring, a larger forest management system could integrate many of
the decision making processes into a system of modular pieces, which could be updated
or changed as the the needs of the managers change. One proposed con�guration is
diagrammed in Figure 7.1.
7.1 A proposed larger forest management system
In Figure 7.1, the adaptive growth model is represented by the box in the upper right
corner. All work presented throughout this dissertation would belong in this box.
Inventory and Monitoring - This is probably the most valuable component for all
other components to work well. If managers do not have a good inventory upon
which to base decisions and a monitoring system to provide the information to

243
Evaluator Info.Traditional
Info.Spatial
GIS
Database
SuccessManagement
DocumentorPlan
Management
EvaluatorPattern
LandscapeScheduling
Harvest
EvaluatorAlternative
Management
ModelGrowth
Monitoring&
Inventory
EvaluatorSuccessSystem
Figure 7.1. One possible con�guration of a larger forest management system.

244
evaluate decisions previously made, decisions will be inaccurate. This informa-
tion can also be used to determine the time at which decisions are required. The
monitoring can be keyed to the times in stand development that would probably
require treatment (thinning, fertilization, and others), as opposed to regularly
space intervals in time. Either method should work as long as the monitoring
is consisted, rational, and well documented.
Database - This component of the system provides a data handling function for all
other components. With all components interacting, information about stands
and the relationships between stands must be well organized to be accessible
and useful. The database component provides this function in the forest man-
agement system. The box is divided into two parts, because information about
forests are in both spatial and list form; and a database that handles one form
well seldom handle the others well. A connection between the two databases is
necessary to have a fully functional system.
Alternative treatment generation - Silviculture should be the practice of taking a
set of stands and, with the minimum manipulation, moving the stand to a
set of conditions that meet the management goals for the ownership. When
considering alternative treatments, how is the appropriate treatment chosen?
One approach is to �lter through combinations of alternatives for the stand
conditions, developing possible operations and their cost. Alternatives that can
be generated in this way may have equivalent utility to the manager but have
very di�erent timing of treatments or provide very di�erent stand structure and
landscape patterns. A method of evaluating the selected pattern in terms of
these broader issues is also needed.
Documentation - This is very important in accessing the success of a forest man-
agement system. A recurrent problem in forest management is that stand pre-

245
scriptions are made, �led, and seldom referred to again. The management of
a stand following a prescription should have anticipated outcomes that can be
checked consequently, a good system of record keeping is used. A system of
record keeping should be easy to use, provide useful information to the people
collecting the information, and organized for easy future retrieval.
Success of Management - This component implements a management system idea
in which the performance of the chosen management alternative is evaluated.
This comparison of predicted behavior of the stand and the actual behavior of
the stand provides a needed check with reality. If the the behaviors are similar,
con�dence in the prediction is increased. If the behaviors are di�erent, a decision
must be made whether the actual behavior is acceptable or new operations are
and management direction is needed. The analyzes must be done on a routine
basis, not just when a problem is perceived.
Success of the System - This component could be designed to monitor the per-
formance of the components of the management systems. If portions of the
management system consistently have poor behavior in many di�erent stands,
that part of the system would be a candidate for improvement.
These are a few ways this type of model can be used in the context of a larger
system. Many other project need to be completed to have a working system.
7.2 Some statistical aspects designed into the current approach
The adaptive stand dynamics model has many potential aspects designed or, provided
for, than there is room or time to test or demonstrate in this dissertation. The model
has many of the same elements as traditional forest growth models but it emphasizes
di�erent things. Below is a review of how this type of model di�ers and it describes
why the proposed approach may be useful:

246
� Maximum use from diverse types of information is desired. Under the proposed
scenario, a user would have information and data from such various sources such
as published equations, stem analysis from a stand, stand measurements, repeat
measurements from the same stand, and growth equations from forest-wide
inventories. The models should make maximum use of whatever information or
data are available within the framework proposed for the model.
� A systematic method for updating the model with varying degrees of memory
of the the past values of parameters. When the established parameters are
considered by the user to be good, the model should be able to have a strong
memory. If the parameters of the model are known with less certainty, then
weak memory would be desired as the new information will have more value for
prediction that the old parameter estimates (Berger, 1985; Goldberg, 1989).
� A method of producing an estimate of the cumulative variance from the various
model components is important. Growth models have been built and used that
predict average values with little information about the range of the outcomes
likely to be encountered. This range of outcomes is at least as important that
the mean values. Because one must know something about the likelihood of a
particular outcome to accurately calculate a risk function for decision making.
7.2.1 How a Bayesian Approach may be useful in Forestry
Many desired features of the adaptive stand dynamics model do not �t easily within
a standard statistical framework. Bayesian statistics permits the systematic biasing
of equations with relevant additional information external to the modeling data set.
Information outside a given data set is known and can improve growth model predic-
tions. The equation is biased through the assumption of prior frequency distributions
about some components of the equation. In the case where \prior" assumptions pro-

247
vide no new information, the result is the same obtained from unbiased methods.
Bayesian statistics have had a poor reputation since the methods can be easily mis-
used. Additionally, the methods have not been as widely taught as the more common
\frequentist" methods (Berger, 1985); however, the results of using a Bayesian ap-
proach can be quite useful when the priors are formulated in a systematic and rational
way.
Foresters are often faced with a collection of information and data that di�er in
time, resolution, and quality. Information in this context is compiled data or equa-
tions such as height-age curves, height-diameter curves, height growth equations, and
site index equations. Data are individual measurements taken from forest samples
(Baskerville and Moore, 1988). The Bayesian approach permits foresters to take ad-
vantage of all information and data available, while accounting for di�erences in time
and quality. Other advantages include the ability to use assumed error distributions
to simulate ranges of potential outcomes of the models, as well as the ability to inte-
grate new information and data easily through the assumed prior distribution. The
biggest disadvantage to Bayesian statistics is the inclusion of known bias, since bias
can be abused. This disadvantage can be overcome with careful and conscientious
de�nition of prior distributions.
The present approach is designed to utilize a Bayesian approach; however, the
approach can only be fully tested with repeatedly measured data of the type required
by the model. This type of data are rare because of limited measurements taken
by foresters and because of inconsistent record keeping when good measurements are
taken.

Chapter 8
CONCLUSION
\All is clear, limpid; the eye rests on the whole and on the parts and sees
how the parts have conspired to make the whole; it perceives the center
where the lymph ows, the breath, the root of the whys ..."
Umberto Eco, 1988, \Foucault's Pendulum", Chapter 120
The practice of silviculture is changing. Silviculturists in the past have focused
on the speci�c operations done to existing stands. Rather, silviculturists can focusing
on the development of stand structure and the events that must occur to produce
speci�c structures. With this subtle change in thinking, the silviculturist shifts from
a human centered focus to a tree and stand centered focus. In the latter approach,
operations become a means not an end. The shift allows a broadening of the perceived
possibilities in multi-objective forest management.
Silviculturists are integrating more objectives in the practice of silviculture; there-
fore, they need better projections of the stand structure that will develop and how
treatments will change stand structures. They also need to be able to communicate
those changes in stand structure to others. The starting point for a silviculturist is
a thorough knowledge of the principles of stand dynamics. These principles provide
a conceptual framework within which to examine stands and their anticipate change.
While the principles are few and relatively simple, the interaction of the principles
can produce a wide variety of stand structures. Silviculturists can bene�t from tools
that apply the principles to speci�c situations. A model using the principles of stand
dynamics is presented to illustrate a this type of tool.

249
An stand dynamics model is designed to apply these principles to a speci�c stand
and to predict change in stand structure. It is assumed that changes in stand structure
can be described as the aggregation of responses of an individual plants to their
immediate environments. It is further assumed that the environment of each plant is
in uenced by the neighboring plants and the conditions of the site. Each plant has
inherent potential that is set by it's genetic makeup. This makeup also in uences
how a plant responds to its immediate environment. Stand dynamics then becomes
the de�nition of the important relationships and important environmental in uences.
This approach can be applied to any plant species that has height and a crown mass.
8.1 Models of stand dynamics and related models
The stand dynamics model incorporates may features from other growth models.
Many of the relationship come from the ideas presented at the end of chapter 3. The
TASS growth model is the dominant in uence in the current approach. A relationship
between crown size and height growth can be found in TASS. Additionally, the concept
of using crown size to predict stem increment can be found in TASS.
The CROGRO model suggested the potential of graphical model output. Graphi-
cal model output will become increasingly important. Forest growth models will have
methods of easily producing a wide variety of graphical outputs and exibility to
produce new outputs as the occasion requires.
The models of Pukkala (Pukkala, 1987; Pukkala and T. Kolstr�om, 1987; Pukkala,
1988; Pukkala, 1989a; Pukkala, 1989b; Pukkala, 1990) demonstrate the possible uses
of spatial growth models. Pukkala illustrated many creative uses of spatial growth
models. These include studying possible ways of arranging the spatial variation after
thinning to ameliorate the e�ects of trails produced for thinning equipment (Pukkala,
1989b). Another study examines the e�ect of overstory residual trees on the growth
and development of natural regeneration (Pukkala and Kuulivainen, 1987). These

250
examples point to the potential of using spatial models to answer questions that have
not been attempted in the past.
8.2 Structural indices and diagnostic criteria
Several structural indices and diagnostic criteria have been presented in the theory
chapter to provide ways to measure stand structure quantitatively. These measures
are needed to build these models. Many di�erent measures have been present to
illustrate the fact that each measure di�erentiates speci�c aspects of the data. Some
are more robust than others, but they each describe an aspect of the spatial character.
The diagnostic criteria also are tools to describe speci�c aspects of a stand's con-
dition, structure, or change. Because silviculturists have developed an understanding
of the range and dynamics of these tools, diagnostic criteria are useful for evaluating
growth model output. All these measures can be used to evaluated the output of any
forest growth model.
8.3 Adaptive silviculture
In the adaptive approach to silviculture, a sequence of expected stand structures is
de�ned. These expectations can be thought of as hypotheses. At each point in the
future at which a structure has been de�ned, the stand is compared to the expected
stand structure. Con�dence in the model's predictions increases if the two structures
agree. If the two structure disagree, both management and expectation of future
structures are adjusted and the process is repeated. These process is analogous to
hypothesizing and testing in science.
The thrust of this dissertation is that stand dynamics, while complex, is very
understandable. Stand dynamics can be reproduced with relatively simple models,
and these models can be useful in a forest management. The examples in this study
produced results that reproduced the stand dynamics behavior in the sampled stands.

251
The models predicted reasonable behaviors for the near future. The models should
work well for the designed use in adaptive silviculture. They can also adjust to
changing growth conditions that are not originally predicted.
Many ideas in this dissertation are not exclusive to adaptive model of stand dy-
namics. They would work equally well with traditional forest growth models or other
type of models. Monitoring growth of a speci�c stand, using that growth to adjust
the predictions, and testing the predictions is a powerful tool for making better pre-
dictions. Also, the idea of graphical model output can and has been applied to other
forest growth models.
This dissertation presents an alternative approach to silviculture, explains the
rational behind the approach, presents some tools to implement the approach, and
suggests how the approach could be extended. The practice of viewing silviculture as
an adaptive projection of stand structures not only can be done; it has been and is
done by many silviculturists.
The adaptive stand dynamics model presented here is designed to give a silvicul-
turist the maximum exibility in determining the treatment of a stand. This models
is designed to present its output in a graphic manner. This adaptive management
approach has both advantages and disadvantages. It is exible, intuitive, and ex-
tensible; however, it also gives the user the responsibility for parameterization, for
analysis of the predicted trend, and ethical use of the predictions. The model will
predict any trends given appropriate parameters. Determination of the acceptability
of the predicted trends is the user's responsibility.

BIBLIOGRAPHY
Barnes, G. H. 1949. Site classi�cation for even-aged stands of western hemlock.
Research Note 50, U. S. Dept. Agr. Paci�c Northwest Forest and Range Experiment
Station.
Baskerville, G. 1985. Adaptive management wood availability and habitat avail-
abilty. Forestry Chronicle, 61:171{175.
Baskerville, G. and T. Moore 1988. Forest information systems that really work.
Forestry Chronicle, 64:136{140.
Benjamin, L. R. and R. C. Hardwick 1986. Review Article: Sources of variation and
measures of variabilty in even-aged stands of plants. Annals of Botany, 58:757{778.
Berger, J. O. 1985. Statistical Decision Theory and Bayesian Analysis. Springer-
Verlag, New York, Berlin, Heidelberg, Tokyo, 617, p.
Bickford, C. A. 1957. Stocking, normality, and measurement of stand density.
Journal of Forestry, 55:99{104.
Botkin, D. B., J. F. Janak, and J. R. Wallis 1972b. Rationale, limitations and
assumptions of a Northeastern forest growth simulator. IBM Journal of Research
and Development, 16:101{113.
Botkin, D. B., J. F. Janak, and J. R. Wallis 1972a. Some ecological consquences of
a computer model of forest growth. Journal of Ecology, 60:849{872.

253
Brix, H. 1983. E�ects of thinning and nitrogen fertilization on growth of Douglas-
�r: Relative contribution of foliage quanity and e�ciency. Canadian Journal of
Forest Research, 13:167{175.
Brown, D. and P. Rothery 1978. Randomness and local regularity of points in a
plane. Biometrika, 65:115{122.
Brubaker, L. B. 1986. Responses of tree populations to climate change. Vegetatio,
67:119{130.
Byth, K. and B. D. Ripley 1980. Sampling spatial patterns by distance mehtods.
Biometrics, 36:279{284.
Clark, P. J. and F. C. Evans 1954. Distance to nearest neighbor as a measure of
spatial relationships in populations. Ecology, 35:445{453.
Clatterbuck, W. K., C. D. Oliver, and E. C. Burkhardt 1987. The silvicultural
potential of mixed stands of cherrybark oak and American sycamore: Spacing is
the key. Southern Journal of Applied Forestry, 11:158{161.
Curtis, R. O. 1971. A tree area power function and related stand density measures
for Douglas-�r. Forest Science, 17:146{159.
Dean, T. J. and J. N. Long 1986. Variation in sapwood area-leaf area relastions
within two stands of lodgeploe pine. Forest Science, 32:749{758.
Diggle, P. J. 1983. Statistical Analysis of Spatial Point Patterns. Academic Press,
London, New York.
Dixon, P. M., J. Weiner, T. Michell-Olds, and R. Woodley 1987. Bootstrapping
the Gini coe�cient of inequality. Journal of Ecology, 68 (6):1548{1551.

254
Drew, T. J. and J. W. Flewelling 1979. Stand density management: an alternative
approach and its application to Douglas-�r plantations. Forest Science, 25:518{532.
Espinosa Bancalari, M. A., D. A. Perry, and J. D. Marshall 1987. Leaf area {
sapwood area relationships in adjacent young Douglas-�r stands with di�erent early
growth rates. Canadian Journal of Forest Research, 17:174{180.
Fellows, D. M., G. L. Sprague, and G. L. Baskerville 1983. CROGRO: An interac-
tive forest growth simulator. Simulation, 41:219{228.
Ford, E. D. 1975. Competition and structure in some even-aged plant monocultures.
Journal of Ecology, 63:311{333.
Ford, M. S. 1990. A 10,000-Yr history of natural ecosystem acidi�cation. Ecological
Monographs, 60:57{89.
Goldberg, D. E. 1989. Genetic algorithms in search, optimization, and machine
learning. Addison-Wesley, Reading, MA.
Grier, C. C. and R. H. Waring 1974. Conifer foliage mass related to sapwood area.
Forest Science, 20:205{206.
Henry, J. D. and J. M. A. Swan 1974. Reconstructing forest history from live and
dead plant material { an appraoch to the study of plant sucession in sothewest New
Hampshire. Ecology, 55:772{783.
Holgate, P. 1964. The e�ciency of nearest neighbor estimators. Biometrics, 20:647{
649.
Hopkins, B. 1954. A new method for determining the type of distribution of plant
individuals. Annals of Botany, 18:213{227.

255
Keane, M. G. and G. F. Weetman 1987. Leaf area { sapwood cross-sectional area
relationships in repressed stands of lodgepole pine. Canadian Journal of Forest
Research, 17:205{209.
Knox, R. G., R. K. Peet, and N. L. Christensen 1989. Population dynamics in
loblolly pine stands: Changes in skewness and size inequality. Ecology, 70:1153{
1166.
Krajicek, J. E., K. A. Brinkman, and S. F. Gingrich 1961. Crown competition|a
measure of density. Forest Science, 7:35{42.
Kuiper, L. C. 1988. The structure of natural Douglas-�r forests in Western Wash-
ington and Western Oregon. Agricultural University Wageningen papers 88-5, Wa-
geningen Agricultural University, Agricultural University Wageningen, The Nether-
lands.
Larsen, D. R. 1991. Adaptable stand dynamics model integrating site-speci�c
growth for innovative silvicultural prescriptions. Forest Ecology and Management,
(Submitted).
Lethpere 1957. The in uence of high pruning on the growth of Douglas-�r. Forestry,
30:9{20.
Lexen, B. 1943. Bole area as an expression of growing stock. Journal of Forestry,
41:883{885.
Liu, K. 1990. Holocene paleoecology of the boreal forest and Great Lakes-St.
Lawrence forest in Northern Ontario. Ecological Monographs, 60:179{212.
Lloyd, F. T. and W. R. Harms 1986. An individual stand growth model for mean
plant size based on the rule of self{thinning. Annals of Botany, 57:681{688.

256
Long, J. N., J. B. McCarter, and S. B. Jack 1988. A modi�ed density management
diagram for coastal Douglas-�r. Western Journal of Applied Forestry, 3:88{89.
Long, J. N., F. W. Smith, and D. R. Scott 1981. The role of Douglas-�r stem
sapwood and heartwood in the mechanical and physiological support of crowns
and development of stem form. Canadian Journal of Forest Research, 11:459{464.
Maguire, D. A. and D. W. Hann 1987. Equations for predicting sapwood area at
crown base in southwestern Oregon. Canadian Journal of Forest Research, 17:236{
241.
Maguire, D. A. and D. W. Hann 1989. The relationship between gross crown
dimensions and sapwood area at crown base in Douglas-�r. Canadian Journal of
Forest Research, 19:557{565.
Mar:M�oller, C. 1960. The in uence of pruning on the growth of conifers. Forestry,
33:37{53.
Matthews, D. M. 1935. Management of American forests. McGraw-Hill, New york
and London.
Matthews, J. D. 1989. Silvicultural Systems. Oxford University Press, Oxford 284,
p.
McArdle, R. E., W. H. Meyer, and D. Bruce 1949. The yield of Douglas-�r in the
Paci�c Northwest. Technical Bulletin 201, U. S. Dept. Agr.
McCarter, J. B. and J. N. Long 1986. A lodgepole pine density management
diagram. Western Journal of Applied Forestry, 1:6{11.
McMahon, T. A. and R. E. Kronauer 1976. Tree structure: Deducing the principle
of mechanical design. Journal of Theoretical Biology, 59:443{466.

257
Mitchell, K. J. 1975. Dynamics and simulated yield of Douglas-�r. Forest Science
Monograph, 17:39.
Mitchell, K. J. and I. R. Cameron 1985. Managed Satnd Yield Tables for Coastal
Douglas-�r: Initial Density and Precommerical Thinning. Technical Report 31, B.
C. Ministry of Forests Research Branch, Victoria, B. C. V8W 3E7.
Moore, P. G. 1954. Spacing in plant populations. Ecology, 35:222{227.
Neyman, J. and E. L. Scott 1972. Stochastic Point Processes. P. A. W. Lewis, Ed.
John Wiley and Sons, New York 646{681, p.
O'Hara, K. L. 1989. Stand growth e�cency in a Douglas-�r thinning trial. Forestry,
62:409{418.
Oliver, C. D. 1978. The development of Northern red oak in mixed stands in
central New England. Technical report, Yale University: School of Forestry and
Environmental studies.
Oliver, C. D. 1992. Similarities of stand structure patterns based on uniformities of
stand development processes throughout the world|Some evidence and the appli-
cation to silviculture through adaptive management. In Kelty, M. J., B. C. Larson,
and C. D. Oliver (Eds.), The Ecology and Silviculture of Mixed-Species Forests: A
Festschrift for David M. Smith. Yale University.
Oliver, C. D., A. B. Adams, and R. J. Zasoski 1985. Disturbance patterns and
forest development in a recently deglaciated valley in the northwestern Cascade
range of Washington, U.S.A. Canadian Journal of Forest Research, 15:221{232.

258
Oliver, C. D., W. K. Clatterbuck, and E. C. Burkhardt 1990. Spacing and strati�ca-
tion patterns of cherrybark oak and American sycamore in mixed, even-aged stands
in the southeastern United States. Forest Ecology and Management, 31:67{79.
Oliver, C. D. and B. C. Larson 1990. Forest Stand Dynamics. McGraw-Hill, Inc.,
New York 467, p.
Oliver, C. D. and M. D. Murray 1983. Stand structure, thinning prescriptions,
and density indexes in a Douglas-�r thinning study, Western Washington, U.S.A.
Canadian Journal of Forest Research, 13:126{136.
Oliver, C. D., K. L. O'Hara, G. McFadden, and I. Nagame 1986. Concepts of
thinning regimes. In Douglas-�r Stand Management Symposium, College of Forest
Resources, University of Washington, Seattle Washington.
Oliver, C. D. and E. P. Stephens 1977. Reconstruction of a mixed{species forest in
central New England. Ecology, 58:562{572.
Paine, D. P. and R. J. McCadden 1988. Simpli�ed forest inventory using large-scale
70-mm photography and tarif tables. Photogrammetric Engineering and Remote
Sensing, 54:1423{1427.
Palmer, M. 1988. Fractal geometry: a tool for describing spatial pattern of plant
communities. Vegetatio, 75:91{102.
Perlin, J. 1991. A Forest Journey. Harvard University Press, Cambridge, Mas-
sachusetts.
Pielou, E. C. 1959. The use of point-to-plant distances in the study of the pattern
of plant populations. Journal of Ecology, 47:607{613.

259
Press, W. H., B. P. Flannery, S. A. Teukolsky, and W. T. Vetterling 1988. Numer-
ical Recipes in C; The Art of Scienti�c Computing. Cambridge University Press,
Cambridge.
Pukkala, T. 1987. Simulation model for natural regneration of Pinus sylvestris,
Picea abies, Betula pendula and Betula pubescens.. Silva Fennica, 21:37{53.
Pukkala, T. 1988. E�ect of spatial distribution of trees on the volume increment
of a young Scots pine stand. Silva Fennica, 22:1{17.
Pukkala, T. 1989a. Methods to describe the competition process in a tree stand.
Scandinavian Journal of Forest Research, 4:187{202.
Pukkala, T. 1989b. Predicting diameter growth in even-aged Scots pine stands with
a spatial and non-spatial model. Silva Fennica, 23:101{116.
Pukkala, T. 1990. A method for incorporating the within-stand variation into forest
management planning. Scandinavian Journal of Forest Research, 5:263{275.
Pukkala, T. and T. Kuulivainen 1987. E�ect of conapy structure on the diurnal
interception of direct solar radiation and photosysnthesis in a tree stand. Silva
Fennica, 21:237{250.
Pukkala, T. and T. Kolstr�om 1987. Competition indices and the prediction of radial
groth in Scots pine. Silva Fennica, 21:55{67.
Reineke, L. H. 1933. Perfecting a stand{density index for even{aged forests. Journal
of Agriculture Research, 46:627{638.
Ripley, B. D. 1981. Spatial Statistics. John Wiley and Sons, New York.

260
Rogers, R. and T. Hinckley 1979. Foliar weight and area related to current sapwood
area in oak. Forest Science, 25:298{303.
Rohlf, F. J. and J. W. Archie 1978. Least{squares mapping using interpoint dis-
tances. Ecology, 59:126{132.
Roth, F. 1925. Forest Regulation. George Wahr Publishing Company, Ann Arbor,
Michigan 218, p.
Shachter, R. D. 1986. Evaluating in uence diagrams. Operations Research, 34:871{
882.
Siemon, G. R., W. J. M�uller, G. B. Wood, and W. G. Forrest 1980. E�ect of
thinning on the distribution and biomass of foliage in the crown of radiata pine.
New Zealand Journal of Forestry Science, 10:461{475.
Skellam, J. G. 1952. Studies in statistical ecology, I. Spatial Pattern. Biometrika,
39:346{362.
Smith, N. J. and D. W. Hann 1984. A new analytical model based on the -3/2
power rule of self{thinning. Canadian Journal of Forest Research, 14:605{609.
Staebler, G. R. 1964. Height and diameter growth for four years following pruning
of Douglas-�r. Journal of Forestry, 62:106.
Sugden, M. J. 1962. Tree sway period|A possible new parameter for crown clas-
si�cation and stand competition. Forestry Chronicle, 38:336{344.
Tomppo, E. 1986. Models and methods for analysing spatial patterns of trees.
Communicationes Instituti Forest Alis Fenniae, 138:65.

261
Troup, R. S. 1952. Silvicultural systems (Second ed.). Oxford University Press,
Amen House, London E.C.4 216, p.
Urban, D. L. 1990. A versatile model to simulate forest pattern; A user's guide
to ZELIG version 1.0. Technical report, Environmental Sciences Department, The
University of Virginia, Charlottesville, Virginia 22903.
Valenti, M. A. and Q. V. Cao 1986. Use of crown ratio to improve loblolly pine
taper equations. Canadian Journal of Forest Research, 16:1141{1145.
Von Post, L. 1946. The prospect for pollen analysis in the study of the earth's
climate history. New Phytologist, 45:193{217.
Vose, J. M. 1988. Patterns of leaf area distribution within crowns of nitrogen- and
phosphorous-fertilized loblolly pine trees. Forest Science, 34:564{573.
Vose, J. M. and H. L. Allen 1988. Leaf area, stem wood growth, and nutrition
relationships in loblolly pine. Forest Science, 34:547{563.
Walters, C. 1986. Adaptive Management of Renewable Resources. MacMillan Pub-
lishing Co.
Waring, R. H., P. E. Schroeder, and R. Oren 1982. Application of the pipe model
theory to predict canopy leaf area. Canadian Journal of Forest Research, 12:556{
560.
Weiner, J. 1984. Neighborhood interference amongst Pinus rigida individuals.
Journal of Ecology, 72:183{195.
Weiner, J. and O. Solbrig 1984. The meaning and measurement of size hierarchies
in plant populations. Oecologia (Berlin), 61:334{336.

262
West, M. and J. Harrison 1989. Bayesian Forecasting and Dynamic Models.
Springer-Verlag, New York.
West, P. W. 1983. Comparison of stand density measures in even-aged regrowth
eucalypt forest of southern Tasmania. Canadian Journal of Forest Research, 13:22{
31.
Whitehead, D., W. R. N. Edwards, and P. G. Jarvis 1984. Conducting sapwood
area, foliage area and permeability in mature trees of Picea stichensis and Pinus
contorta. Canadian Journal of Forest Research, 14:940{947.
Whitmore, T. C. 1982. On pattern and process in forests. Blackwell, Oxford 45{59,
p.
Wilson, B. F. and R. R. Archer 1979. Tree design: Some biological solutions to
mechanical problems. Bioscience, 29:293{298.
Wilson, F. G. 1946. Numerical expression of stocking in terms of height. Journal
of Forestry, 44:758{761.
Wilson, F. G. 1951. Control of growing stock in even-aged stands of conifers.
Journal of Forestry, 49:692{695.
Winkler, M. G. 1985. A 12,000-year history of vegetation and climate for Cape
Cod, Massachusetts. Quaternary Research, 23:301{312.

Appendix A
SPATIAL INDICES
� Skellam-Moore
The statistic d� is calculated as:
d� =�n
A
��
mXi=1
d2(t�t)i (A.1)
where n is the number of individuals in area A, m is the number of individuals
in the distance sample and d(t�t)i is the distance from a random plant to the
nearest plant. In this equations the distances are squared and multiplied by �
to obtain area associated with that distance. This is a test that for the Poisson
assumption is �22m. It was noted by Pielou (1959) that estimates of intensity
n=A are poor using distance methods and that independent estimate of intensity
from quadrates should be used .
� Clark and Evans
Clark and Evans (1954) are as follows:
�d(t�t) =
nXi=1
d(t�t)i
n(A.2)
where �d(t�t) is the mean distance to the nearest neighbor plant from a plant.
d(t�t)i is the distance to the nearest neighbor for the ith individual, and n is the

264
number of individuals in the study area. If � is the density of individual and
de�ned as:
� =n
A(A.3)
where n is the number of individuals in area A. Clark and Evans (1954) showed
that the expected distance to the nearest neighbor is:
E( �d(t�t)) =1
2p�
(A.4)
So that an index of aggregation can be shown as:
R =�d(t�t)
E( �d(t�t))(A.5)
where if the spatial pattern is random, R = 1; clustered, R ! 0; and uniform,
R! some arbitrary upper limit. A test of deviation from randomness is given
as:
z =�d(t�t) � E( �d(t�t))
sr(A.6)
where z is a standard normal deviate, sr is the standard error of E( �d(t�t)).
� Brown
The Brown index of aggreation is the geometric mean of squared distances over
the arithmetric mean and is given by:
G =
"nYi=1
d2(t�t)i
#1=n�d2(t�t)
(A.7)

265
where d2(t�t)i is the squared distance from a random individual to it's nearest
neighbor individual and �d2(t�t) is the arithmetic average of the squared distance
from a random individual to it's nearest neighbor individual. G is a index of
aggregation that is independent of area. The index G ranges from 0 to 1 with
G ! 0 for \clustered" pattern and G ! 1 for \uniform" pattern (Brown and
Rothery, 1978).
� Hopkins'
The Hopkins' index of aggregations is the ratio of the distance from a randomly
chosen point and its nearest neighbor tree and the distance from a that nearest
neighbor tree and its nearest neighbor.
HopF =
mXi=1
d2(p�ti)i
mXi=1
d2(t�ti)i
(A.8)
where d(p�ti)i is the distance from a random point to it's nearest neighbor in-
dividual i and d(t�ti)i is the distance from individual i to it's nearest neighbor
individual. This test has an F distribution with F (2m; 2m) (Hopkins, 1954).
Byth and Ripley (1980) presented a standardized index based on this test as:
HopN =1
m
mXi=1
24 d2(p�ti)i(d2(p�ti)i + d2(t�ti)i)
35 (A.9)
This index has a Normal null distribution,N(12; 112m). AsHopN ! 0 it indicates
a more \uniform" pattern and as HopN ! 1 it indicates a more \clustered"
pattern.
� Holgate

266
The Holgate index is based on the ratio of the distance from a random point to
its nearest neighbor and the distance from the same random point and the next
nearest neighbor (Holgate, 1964).
HolN =1
m
mXi=1
24 d2(p�t)id2(p�t2)i
35 (A.10)
where d(p�t)i and d(p�t2)i are the distance from random points to the �rst and
second nearest neighbor individuals.
HolF =
mXi=1
d2(p�t)i
mXi=1
(d2(p�t2)i � d2(p�t)i)
(A.11)
The Holgate indices have the same null distributions as the Hopkins' indices.
� Pielou
Pielou (1954) pointed out the value of the di�erence between the tree to tree
distances, the point to tree distances, and intensity estimates as separate prop-
erties of a spatial pattern.
�d(p�t) =1
n
nXi=1
d(p�t)i (A.12)
where �d(p�t) is the mean distance from a random point to it's nearest neighbor
individual. Independent quadrates are used to estimate � as:
� =1
mA
mXj=1
xj (A.13)

267
where m quadrates are taken from the sample area and xj is the number of
individuals in the jth quadrate and A is the area of each quadrate. Pielou
(1959) de�nes an index of nonrandomness as:
� = �� �d(p�t): (A.14)
Index � is 1 if for a \random" pattern, �! 0 for \uniform" patterns and �!arbitrary upper limit for \clustered" patterns.
� Ripley's K
This second-order test looks at the relationship of each point to all other points
(Ripley, 1981; Diggle, 1983; Tomppo, 1986). If the point pattern is Poisson, the
cumulative distribution of number of individuals within a given distance will
follow �t2, where t is the maximum distance observed.
�K(t) = E[number of events within distance t of an arbitrary event] (A.15)
within a sample A, the area g(A)
�g(A) = E[number of events within sample area] (A.16)
From this the expected number of tree pairs whose distances is less than t, is
�g(A)�K(t) = �2g(A)K(t) (A.17)
where the intensity � is the mean number of events per unit area. This can also
be estimated from the data by counting points less that a give distances within
a sample area. If di;j is the distance from i to j then

268
=Circumference of Circle i=w
=Circumference of Circle jj,i
i,j >1
1
Arc ajb
Circumference of Circle j=w
i,jdi
j
a
b
A
Figure A.1. Edge correction for points in an sample area A
nXi=1
nXj=1
i6=j
I(di;j�t) (A.18)
I(di;j�t) is 1 if di;j is less than t from an arbitrary point in sample area g(A).
This value must be corrected for the amount area outside the sample area which
cannot be observed. This is done by calculating the circumference of a circle of
radius di;j that is within the sample area g(A). The weight factor wi;j is as given
in Figure 3.6. With this weight factor we get an apparently unbiased estimator.
nXi=1
nXj=1
i6=j
wi;jI(di;j�t) (A.19)
If an independent estimate of � is unknown then we can estimate � with n=g(A)
or the number of tree in the sample over the area of the sample. With these
elements we can estimate K(t) as

269
K(t) =g(A)
n2
nXi=1
nXj=1
i 6=j
wi;jI(di;j�t) (A.20)
This statistic provides more information about the underlying process than the
previously mentioned statistics. If K(t) > �t2 the process is \clustered" and if
K(t) < �t2 the process is \uniform." Additionally, the size and range of the
deviations from �t2 can be determined about the process.
� Semi-variance
Semi-variance is the basis for many geostatistic tools and is the concept that a
feature of interest about a spatial pattern is the semi-variance at various spatial
scales (i.e. distance) (Palmer, 1988). This concept may be applied in one, two
or more dimensions, here the two dimensional case is discussed. This statistic
must be applied to a value that can be measured as a continuous variable across
the study area. This technique has potential for resource variables such as
nitrogen N or water content of the soil as well as aggregated stand variables at
landscape scales. The interpretation for discrete variables such as tree size is
unclear.
Given an area mapped for the value Z where Zi is the value of the variable at
location i. The semi-variance d is
d =1
2Nd
NdXi=1
(Zi � Zi+d)2 (A.21)
which is the squared di�erence between the value of Z at location i and the value
of Z at location i + d. This is summed for the number of points N that are
distance d apart Nd. A semivariogram is the plot of d versus d. Palmer (1988)

270
plots several hypothetical process patterns and the resultant semivariogram as
well as several scales of plant data.

Appendix B
RELATED MODELS
The TASS model
The Tree And Stand Simulator (TASS), is a distance dependent, forest growth model
that uses a three-dimensional description of a part of the stand to simulate tree
growth and crown interaction. Th amount of foliage on a given tree determines the
trees height growth and in turn branch growth. The resultant stem increment is
allocated over the stem. A description of the functions in the TASS model are as
follows.
A tree's crown shape is assumed to a function of tree height with layers of foliage
added to the outside of the crown each year. Height growth is the main change
variable in this model and is calculated as,
Hg = Hgo�1� ea0(
FVFVmax
)a1�
(B.1)
where Hg is height growth, Hgo is potential height growth, FV is foliage volume,
and FVmax is the foliage volume maximum.
Branch growth is de�ned as,
Bg = b0 b1ln
"(L1 +Hg + b2)
(L1 + b2)
#(B.2)
where Bg is branch growth, L1 is depth into the crown, b0 is relation of branch
growth to Hg, b1 is correction for crooked branches, and b2 is curvature of crown
pro�le.
This relationship is used to calculate a shell of foliage volume,

272
FVi = �(b0 � b1)2 � [(L1 +Hgi + b2) �
[ln2(L1 +Hgi + b2)� 2(ln(L1 +Hgi + b2)� 1)
�(1 + lnb2) + ln2b2]� (l1 + b2) �
[ln2(L1 + b2)� 2(ln(L1 + b2)� 1) � (1 + lnb2) + ln2b2]]
and then summed for the �ve most recent annual shells,
FV =5X
i=1
wiFVi (B.3)
where wi is a weight based on photosynthetic e�ciency and leaf retention that
assumes the following values: for year 1 = 1.0, year 2 = 0.86, year 3 = 0.75, year 4
= 0.63, and year 5 = 0.40.
The bole increment for the tree is calculated by the equation,
BI = c1FVc2 �
�1� ln
�FV
FVmax
��c3(B.4)
where BI is bole increment. This bole increment is then converted to area in-
crement to calculate diameter inside bark along the stem allowing the stem volumes
to be calculated in a manner similar to that used with taper equations. Maximum
crown length is calculated as a function of height by,
CLmax = d0HT d1: (B.5)
To determine crown interaction and foliage by layers for the area increment, the
plot is divided into 1 foot cubes and each cube is assigned either to open, or to a tree.
The strongest in uence for assignment to a tree is based on the distance to the tree
and the branch length of that tree at crown base.
This model has been calibrated to extensive permanent sample plot data for
Douglas-�r (Pseudotsuga menziesii (Mirb.) Franco ) and versions are being pre-

273
pared for western hemlock (Tsuga heterophylla (Raf.) Sarg.) and sitka spruce (Picea
sitchensis (Bong.) Carr. ). The exibility of the TASS approach is that many treat-
ment e�ects can be simulated for pure stand of Douglas-�r. The TASS models has
been used to generate managed stand yield tables for British Columbia for ease of
use by foresters. The ability to mix species currently has been envisioned but not
implemented.
The CROGRO model
While this growth model had much less e�ort expended in its construction it is no less
interesting in concept. The CROGRO model was built as a tool to teach crown devel-
opment to students. The approach makes a number of assumptions about how crown
interaction, calibrate with regional growth and yield information and was evaluate by
presentation to students, foresters and academics. One of the most interesting fea-
tures of the model is that the model results are displayed in two-dimensional computer
drawing of the trees and their crowns.
Height growth is the driving function of this model. The actual height growth is
determined by reducing the potential height growth by the percentage of the optimal
vertical cross-sectional area of the crown. The size of the vertical cross-section of the
crown is a function of the crown architecture de�ned as
H(t) = hmax ��1� e�bh�t
ch�
(B.6)
where H(t) is height at age t, hmax is the maximum height for the species, bh; ch
are parameters to de�ne the shape of the height age curve. This is the equation form
for a weibull growth curve. The �rst derivative of this function is used to determine
the height growth rate.
dH
dt= h1 � (hmax �H(t)) �
"� log
1� H(t)
ah
!#h2(B.7)

274
h1 = ch � b1=chh (B.8)
h2 =ch � 1
ch(B.9)
A weibull growth curve was also used to describe the crown width growth over
time.
dW
dt= w1 � (wmax �W (t)) �
"� log
1� W (t)
aw
!#w2(B.10)
where W (t) is the crown width at time t, wmax is the maximum crown width
and w1; w2 are constants. Figure 3.8 is a diagram of the crown geometry for the
CROGRO model.
The maximum crown radius wmax is a function of branch angle BA and maximum
branch length BLmax and calculated as
SWMAX = BLmax � sin(BA) (B.11)
above Hmin which can be calculated as
hmin = wmax= tan(BA) (B.12)
and below Hmin for any height H
SWMAX = H � tan(BA) (B.13)
and wmax is determined by minimum of the two SWMAX calculations.
This approach was found acceptable for the design purpose with reservation as to
the extent to which the model is used. The dynamics of open grown crown were not
well represented, however the essential dynamics of stand grown trees are acceptably
represented.

275
Pukkala's Model
Pukkala has build a spatial growth model call MikroMikko for Scot pine (Pinus
sylestris L.) in Finland. Average stand characteristic are used to generate a tree
list for a spatial pattern. In Pukkala (1989b), two methods are presented for gen-
erating diameters from a spatial pattern. The �rst method assumes that trees with
many close neighbors will be small and trees with few close neighbors will be large.
The second method uses the local spatial pattern to predict a diameter distribution
and then this distribution is sampled to determine the diameter for the subject tree.
Then the spatial pattern and the predicted diameter are used to predict tree height.
Pukkala's model uses these relationships in the following sequence.
1. Stand average statistics such as density and total stand basal area are speci�ed
for the stand.
2. The tree coordinates are generated as a realization of a suitable spatial process.
3. The diameters are predicted from the spatial pattern.
4. The heights are then predicted as a function of the spatial pattern and the
diameter.
Pukkala uses a growth model that predicts the diameter increment as
�dbh = b0 + b1 ln
" mXi=1
�i
!+ 0:3
#+ " (B.14)
where �dbh is the future �ve year diameter increment, b0; b1 are equation coe�-
cients, and �i is the angle subtended by all trees larger that the subject tree and no
further that 5 meters away from the subject tree (see Figure 3.9). The heights are
then statically predicted from diameter.

276
h = ch
"1:3 + d2
(a0 + a1d)2
#(B.15)
where h is the new height, ch is a correction factor to obtain a speci�ed height
estimate for the average tree, a0; a1 are equation coe�cients, and d is the newly
predicted diameter.

Appendix C
LATERAL SURFACE AREA EQUATIONS
The formulas for these are; for a paraboloid:
SA =�2� r
12 h
� h(r2 + 4h2)
3
2 � r3i
(C.1)
where SA is the lateral surface area of the solid, h is the height of the solid, and
r is the radius of the base of the solid. For the cone:
SA = �rpr2 + h2 (C.2)
and for a neiloid:
SA =Z h
02�Kx
3
2
s1 +
�K2
3x
1
2
�2dx (C.3)
This is actually the general form where Kx3
2 is the function for the shape of a
neiloid and for any function f(x)
SA =Z h
02�f(x)
q1 + (f 0(x))2 dx (C.4)
where f(x) is any function over the height h of the geometric solid and f 0(x) is
the �rst derivative of f(x), which is used to divide the surface into many small bands
of revolution. In the above case