prime-ome: "a molecular approach towards defense priming"
TRANSCRIPT

Department Of Agricultural Biotechnology
(Centre Of Excellence In Biotechnology)
Anand Agricultural University
Anand-388110.
‘Prime-ome’ : A Molecular Approach Towards Defense Modulation
Speaker : Dhanya A J
Degree : M. Sc. (Agri.) Plant Molecular
Biology and Biotechnology
Major Guide : Dr. G. B. Patil
Minor Guide : Dr. Sasidharan N
Course No : MBB 591
Reg. No. : 04-2348-2014
Date : 17/10/2015
Time : 16:00

Contents
Introduction
SAR and ISR
Priming
Prime-omics
Prime-omics in defense against pathogens and pest
Conclusion
Future thrust
1
5
6
7
2
3
4
2
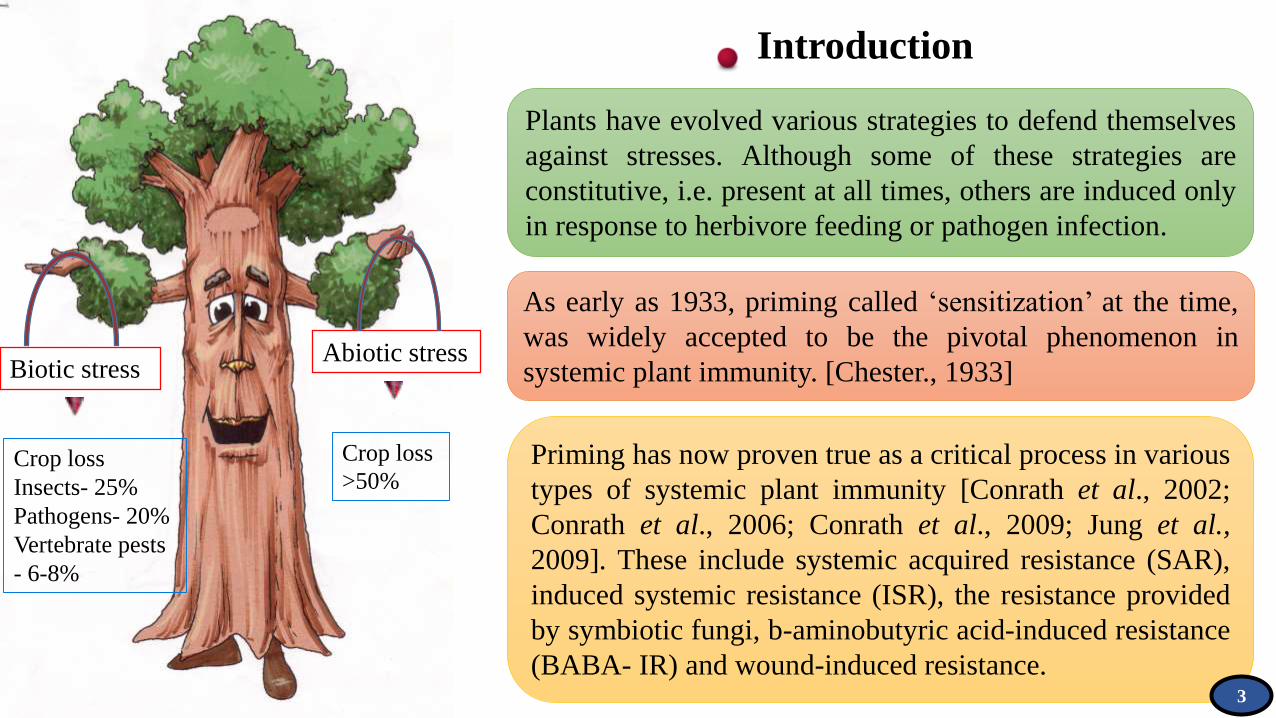
Biotic stressAbiotic stress
Crop loss
>50% Crop loss
Insects- 25%
Pathogens- 20%
Vertebrate pests
- 6-8%
Plants have evolved various strategies to defend themselves
against stresses. Although some of these strategies are
constitutive, i.e. present at all times, others are induced only
in response to herbivore feeding or pathogen infection.
As early as 1933, priming called ‘sensitization’ at the time,
was widely accepted to be the pivotal phenomenon in
systemic plant immunity. [Chester., 1933]
Priming has now proven true as a critical process in various
types of systemic plant immunity [Conrath et al., 2002;
Conrath et al., 2006; Conrath et al., 2009; Jung et al.,
2009]. These include systemic acquired resistance (SAR),
induced systemic resistance (ISR), the resistance provided
by symbiotic fungi, b-aminobutyric acid-induced resistance
(BABA- IR) and wound-induced resistance.
Introduction
3

Induced systemic resistance (ISR): type of systemic,
broad spectrum immunity in plants. Induced systemic
resistance is elicited by colonization with selected
strains of non-pathogenic, plant growth-promoting
rhizobacteria and depends on the plant hormones
ethylene and jasmonic acid.
Systemic acquired resistance (SAR): type of
systemic, broad-spectrum immunity in plants. Systemic
acquired resistance is induced by local contact with a
MAMP*, PAMP*, or effector and depends on the plant
hormone salicylic acid.
* Microbe-associated molecular pattern (MAMP): molecular signatures typical of whole classes of microbes.
Their recognition plays a key role in innate immunity in plants and animals.* Pathogen-associated molecular pattern (PAMP): molecular signatures typical for potential microbial
pathogens of a given host organism.
Fig. 1
4

* Microbe-associated molecular pattern (MAMP): molecular signatures typical of whole classes of microbes.
Their recognition plays a key role in innate immunity in plants and animals.* Damage-associated molecular pattern (DAMP): signals arising from plants because of damage caused by
microbes; originally referred to as ‘endogenous elicitors’.
It is the induction of a
physiological state that
enables cells to respond to
very low levels of a
stimulus in a more rapid and
robust manner than non-
primed cells. In plants,
priming plays a role in
defense (‘defense priming’)
and seed germination (‘seed
priming’).
PRIMING
Fig. 2
5

Primed state: It is the physiological state of a plant that has been subjected to
priming. Usually starts on exposure of such a plant to a stress.
Fig. 36

Priming events can occur as a result of inter-individual or inter-species communication,
such as induced resistance mediated by rhizobacteria, mycorrhizal fungi, or virulent or a
virulent pathogens or by natural or axenic compounds. Plants ‘remember’ such events
(Priming memory) and, depending on the type of primary stimulus or priming
stimulus(initial trigger for priming) and the pathosystem involved (target of priming),
primed plants can deploy a diverse set of defense mechanisms.
Types based on the stimuli which
induces priming
Natural priming (by microorganisms)
Chemical priming (treatment with
chemicals)
Constitutive priming (alterations of specific
defense-repressive constitutive genes)
7
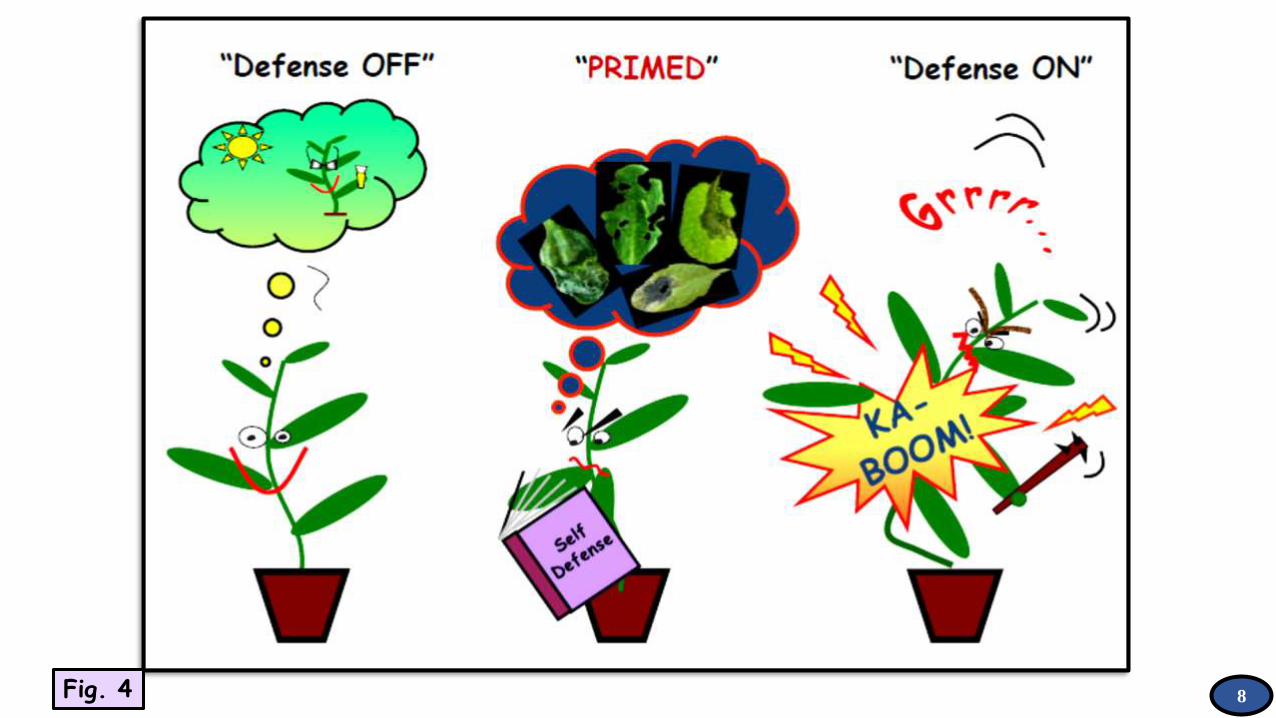
Fig. 4 8
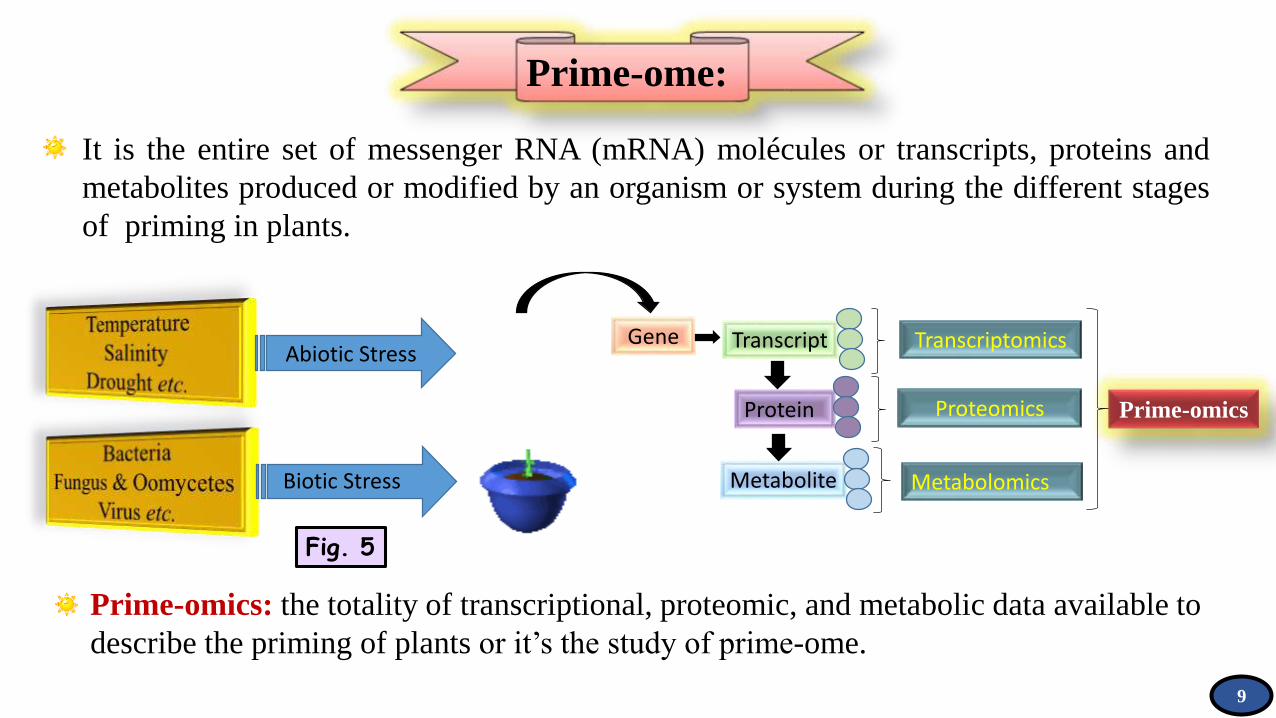
Prime-ome:
Biotic Stress
Gene Transcript
Protein
Metabolite
Transcriptomics
Proteomics
Metabolomics
Prime-omics
Abiotic Stress
Prime-omics: the totality of transcriptional, proteomic, and metabolic data available to
describe the priming of plants or it’s the study of prime-ome.
It is the entire set of messenger RNA (mRNA) molécules or transcripts, proteins and
metabolites produced or modified by an organism or system during the different stages
of priming in plants.
Fig. 5
9
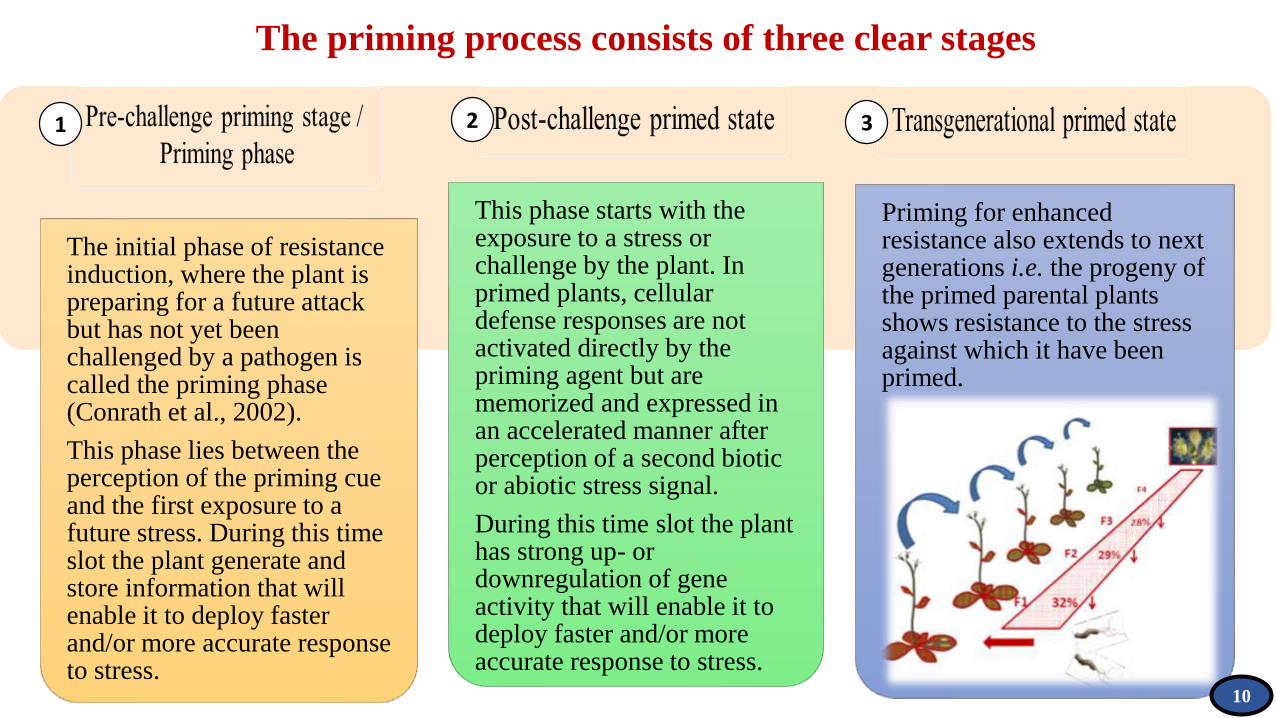
The priming process consists of three clear stages
The initial phase of resistance induction, where the plant is preparing for a future attack but has not yet been challenged by a pathogen is called the priming phase (Conrath et al., 2002).
This phase lies between the perception of the priming cue and the first exposure to a future stress. During this time slot the plant generate and store information that will enable it to deploy faster and/or more accurate response to stress.
This phase starts with the exposure to a stress or challenge by the plant. In primed plants, cellular defense responses are not activated directly by the priming agent but are memorized and expressed in an accelerated manner after perception of a second biotic or abiotic stress signal.
During this time slot the plant has strong up- or downregulation of gene activity that will enable it to deploy faster and/or more accurate response to stress.
Priming for enhanced resistance also extends to next generations i.e. the progeny of the primed parental plants shows resistance to the stress against which it have been primed.
21 3
10

Pla
nt
react
ion
Priming stimulus Challenge Challenge
Priming phase Post – challenge primed state
Time
Transgenerational primed state
Stages of primingFig. 611

Seed collection & next generationInoculation
(12-oxo-phytodienoic acid)
12Gamir et al., 2014Fig. 7
(Indole-3-carboxylic acid)

Prime-omics in defenses against pathogens and pest
13

Plant Priming Tools Suggested mechanisms Refs
By Against
Priming phase Nicotiana
tabacum
Agrobacterium
tumefaciens
GV3101
– qRT-PCR Protein
immunoblotting
detection
SA, ROS, MAPK Sheikh et al., 2014
Solanum
tuberosum
Phosphite
BABA
GABA
Laminarin
INA
– Microarray
LC-MS/MS
SA, PR proteins, PTI, HR,
wall-associated kinase, primary
metabolism, TCA, ROS, Ca2+-
dependent
pathway redox- regulating
enzymes, sesquiterpene
phytoalexin biosynthesis
Bengtsson et al., 2014
Massoud et al., 2014
Lim et al., 2013
Jelonek et al., 2013
Hordeum
vulgare
Pseudomonas
fluorescens
– Microarray
RT-PCR
Detoxification, lipid transfer,
cell wall biosynthesis, JA
Petti et al., 2010
Zea mays Synthetic
indole
dispensers
– GC-flame
ionisation detector
(FID)
HIPVs, ABA, JA, JA-Ile Erb et al., 2015
Post-
challenge
primed state
Oomycetes
Vitis spp. Methionine Plasmopara viticola qRT-PCR H2O2
measurement
(FOX1 method)
ROS Boubakri et al., 2013
S. Tuberosum Phosphite Phytophthora
infestans
LC-MS/MS Callose deposition, HR, TCA Lim et al., 2013
Overview of the omics involved in various stages of priming in plantsTable 1
Balmer et al., 201514
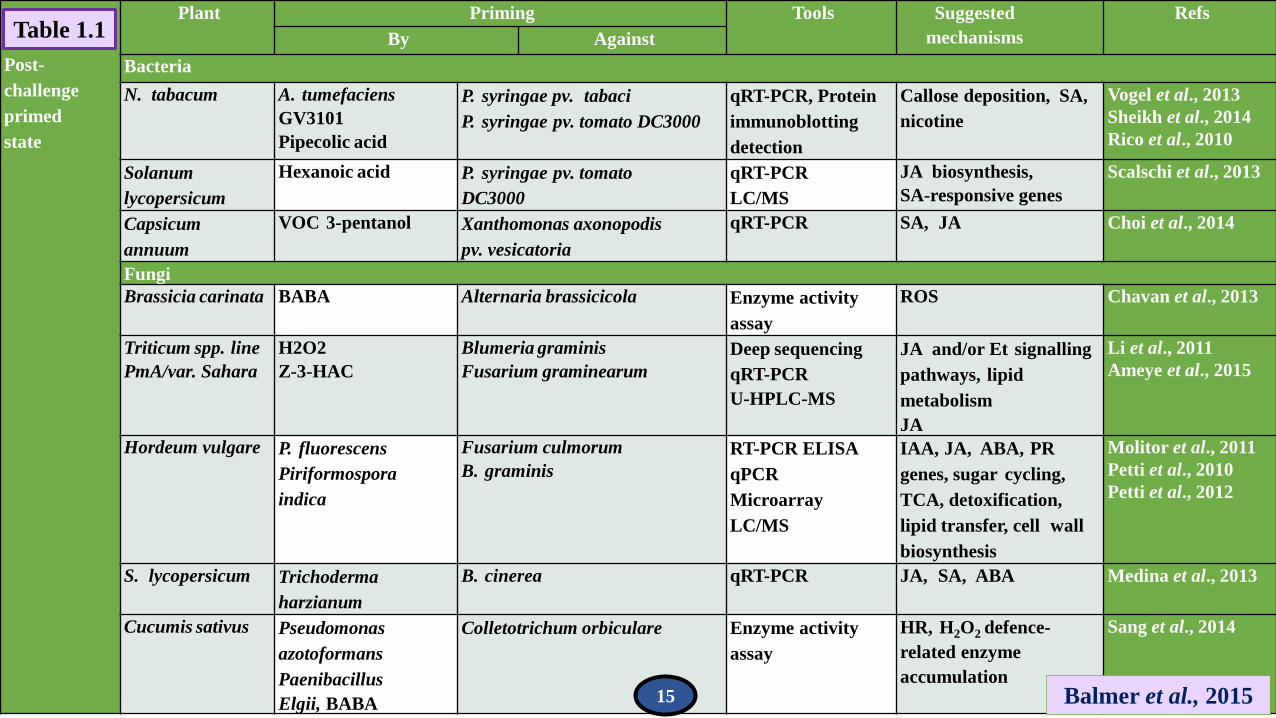
Post-
challenge
primed
state
Plant Priming Tools Suggested
mechanisms
Refs
By Against
Bacteria
N. tabacum A. tumefaciens
GV3101
Pipecolic acid
P. syringae pv. tabaci
P. syringae pv. tomato DC3000
qRT-PCR, Protein
immunoblotting
detection
Callose deposition, SA,
nicotine
Vogel et al., 2013
Sheikh et al., 2014
Rico et al., 2010
Solanum
lycopersicum
Hexanoic acid P. syringae pv. tomato
DC3000
qRT-PCR
LC/MS
JA biosynthesis,
SA-responsive genes
Scalschi et al., 2013
Capsicum
annuum
VOC 3-pentanol Xanthomonas axonopodis
pv. vesicatoria
qRT-PCR SA, JA Choi et al., 2014
Fungi
Brassicia carinata BABA Alternaria brassicicola Enzyme activity
assay
ROS Chavan et al., 2013
Triticum spp. line
PmA/var. Sahara
H2O2
Z-3-HAC
Blumeria graminis
Fusarium graminearum
Deep sequencing
qRT-PCR
U-HPLC-MS
JA and/or Et signalling
pathways, lipid
metabolism
JA
Li et al., 2011
Ameye et al., 2015
Hordeum vulgare P. fluorescens
Piriformospora
indica
Fusarium culmorum
B. graminis
RT-PCR ELISA
qPCR
Microarray
LC/MS
IAA, JA, ABA, PR
genes, sugar cycling,
TCA, detoxification,
lipid transfer, cell wall
biosynthesis
Molitor et al., 2011
Petti et al., 2010
Petti et al., 2012
S. lycopersicum Trichoderma
harzianum
B. cinerea qRT-PCR JA, SA, ABA Medina et al., 2013
Cucumis sativus Pseudomonas
azotoformans
Paenibacillus
Elgii, BABA
Colletotrichum orbiculare Enzyme activity
assay
HR, H2O2 defence-
related enzyme
accumulation
Sang et al., 2014
Balmer et al., 2015
Table 1.1
15

Plant Priming Tools Suggested mechanisms Refs
By Against
Post-
challenge
primed state
Nematodes
S. lycopersicum Arbuscular
mycorrhizal fungi
(AMF)
Meloidogyne
incognitaRT-PCR Suppression
subtractive
hybridisation
Phenylpropanoid pathway,
ROS metabolism
Vos et al., 2013
Vitis spp. Arbuscular
mycorrhizal fungi
Xiphinema index RT-PCR Suppression
subtractive
hybridisation
Chitinase, PR genes,
shikimate enzyme pathway
Hao et al., 2012
Arthropods
Phaseolus
lunatus
JA
(E)-b-Ocimene
Tetranychus urticae RT-PCR GC/MS
Olfactory choices
PIOS, volatile emission, JA,
predator attraction
Muroi et al., 2011
Gols et al., 2003
A. thaliana MeSA + feeding
larvae
Caterpillar feeding
Pieris brassicae RT-PCR
Choice assays
Northern blot
ABA, JA, oviposition
deterrence
Groux et al., 2014
Vos et al., 2013
S. lycopersicum AMF
Aphid feeding
Helicoverpa
arimigera
Bemisia tabaci
RT-PCR
Olfactory choices
Larval deterrence, JA
signalling, systemin signalling,
HIPVs – indirect defence
Song et al., 2013
Ammopiptanthus
mongolicus
Conspecifics
HIPVs
Orgyia ericae NMR- metabolomics TCA, amino acids, lipids,
glycolate, sugars
Sun et al., 2014
Oryza sativa Silicon Cnaphalocrocis
medinalis
RT-PCR JA signalling Ye et al., 2013
Z. mays (E)-b-Ocimene Mythimna separata GC/MS
Olfactory choices
Parasitoid attraction Muroi et al., 2011
Transgenera-
tional primed
state
S. lycopersicum MeJA/herbivory Pieris raphae qPCR JA signalling Rasmann et al., 2012
N. tabacum Tobacco mosaic
virus
Tobacco mosaic virus NMR GC/MS Sugars, amino acids Mandal et al., 2012
Table 1.2
Balmer et al., 201516

Prime-omics in defense against bacteria

Vogel et al., 2013Germany
Also proved that exogenous application of Pipecolic acid to tobacco plants provides significant
protection to infection by Pstb and hypersensitive cell death-inducing P. syringae pv maculicola
(Psm).
Pipecolic acid thereby primes tobacco for rapid and strong accumulation of SA and nicotine
following bacterial infection.
L-Pipecolic acid is a Lys-derived non-protein amino acid of plants whose levels increase upon
plant treatment with growth-affecting chemicals, in response to osmotic stress, and in the course
of pathogen infections.
Tobacco plants respond to leaf infection by the compatible bacterial pathogen Pseudomonas
syringae pv tabaci (Pstb) with a significant accumulation of several amino acids, including Lys,
branched-chain, aromatic, and amide group amino acids. Moreover, Pstb strongly triggers,
alongside the biosynthesis of SA and increases in the defensive alkaloid nicotine, the production
of the Lys catabolites Pipecolic acid (Pip) and α-aminoadipic acid (Aad).
Pipecolic acid enhances resistance to bacterial infection and primes salicylic acid and nicotine accumulation in tobacco
18

Mean values of 3 to 5 replicate samples are given in μg g-1 fresh weight (FW) ± SD. Mock-treatments were performed by infiltration of
leaves with a 10 mM MgCl2 solution. Asterisks denote statistically significant differences between Pstb- and MgCl2- samples (2-tailed t-
test; ***: p < 0.001; **: p < 0.01; *: p < 0.05). Ratios of the values of the Pstb (P) and the MgCl2 (M)-samples (P/M) are also given.
Changes in the levels of free amino acids and amines in N. tabacum cv Xanthi leaves(4 week old) upon
inoculation with compatible P. syringae pv tabaci (Pstb) 2 d post inoculation (dpi). Table 2
19

Time course of (A) pipecolic acid (Pip) and (B) α-aminoadipic acid (Aad) accumulation in tobacco leaves inoculated with
compatible Pstb at indicated times after inoculation.
Time course of (A) salicylic acid (SA ) accumulation and (B) nicotine production in tobacco leaves inoculated with compatible
Pstb at indicated times after inoculation.
Fig. 8
Fig. 9
20

Exogenous Pip primes tobacco plants for effective SA and nicotine production
upon Pstb inoculation. H2O or 10 μmol Pip were applied to plants through the soil. Leaves were infiltrated 1 d later with Pstb (dark bars) or MgCl2 (light bars), and leaf
metabolite levels were scored 8 h later. (A) SA contents in leaves. (B) Leaf nicotine levels. Bars represent the mean ± SD of 3 replicate samples. Different
letters above the bars denote statistically significant differences between pairwise compared samples (p < 0.05, 2-tailed t-test).
Fig. 10
21

Exogenous Pip enhances disease resistance of tobacco plants to Pstb and Psm.
Plant pots were supplied with 10 ml of H2O or 10 ml of 1 mM (10 μmol) Pip 1 d prior to bacterial inoculation. (A) Bacterial numbers of Pstb (applied in titers
of OD 0.001) in leaves at 0 dpi and 5 dpi. The y-axis is depicted in a logarithmic scale. Bars represent the mean ± SD of at least 7 replicate samples. Asterisks
denote statistically significant differences between leaf samples of control- and Pip-treated plants (***: p < 0.001; 2-tailed t-test). (C) Bacterial numbers of
Psm (applied in titers of OD 0.005) in leaves at 0 dpi and 5 dpi.
Fig. 11
22

Representative disease symptoms of Pstb-infected tobacco leaves from H2O and
Pip pre-treated plants.
Pstb Psm
Fig. 12
0 dpi 0 dpi5 dpi 5 dpi
23

Prime-omics in defense against oomycetes

Burra et al., 2014Sweden
.
Phosphite-induced changes of the transcriptome and secretome in Solanum tuberosum leading to resistance against Phytophthora infestans
Investigated the transcriptome of Solanum tubersoum (cv. Desiree) and characterized the
secretome by quantitative proteomics after foliar application of the protective agent
phosphite.
It seems that multiple defense pathways are rapidly induced by phosphite treatment that
causes heightened defense leading to enhanced resistance after pathogen infection in local
tissue.
Phosphite had a rapid and transient effect on the transcriptome, with a clear response after
3 h of treatment. This effect lasted less than 24 h, whereas protection was observed
throughout all time points tested.
It activates the genes associated to both biotic and abiotic stress response.
In field applications, the dual nature of the phosphite molecule both being an inducer of
plant resistance and having a direct toxic effect on oomycetes might explain the high
efficacy.
25

Detached leaflet assay of potato plants
Potatoes (cv. Desiree) were foliar sprayed either with 36 mM proalexin (Potassium phosphite; phosphite treated) or tap water
(Water treated). “Covered leaves” leaflets were obtained by covering two leaves per plant during phosphite spray. Washed
leaflets were obtained by spraying leaves with 36 mM proalexin, washing and drying away the phosphite present on the
leaves. Infection was measured as lesion size 7 days after inoculation with P. infestans. Data corresponds to mean ± SD
obtained from 12 biological replicates.
Fig. 13
26

Gene ontology (GO) analysis: representation of processes and associated
example transcripts (in brackets) significantly regulated at each time point.Fig. 14
27

Time(h)
No
: of
sign
ific
ant
tran
scri
pts
Differentially expressed genes: A
comparison of number of transcripts
induced and repressed at each time
point.
Area proportional Venn
diagram depicting overlap of
transcripts significantly altered
at all the time points .
Fig. 15Fig. 16
28


Molitor et al., 2011Germany
• Colonization of barley roots with the basidiomycete fungus Piriformospora indica
(Sebacinales) induces systemic resistance against the biotrophic leaf pathogen Blumeria
graminis f. sp. hordei (B. graminis).
• In plants that were more B. graminis resistant due to P. indica root colonization, 22
transcripts, including those of pathogenesis related genes and genes encoding heat-
shock proteins were differentially expressed.
• Detailed expression analysis revealed a faster induction after B. graminis inoculation
between 8 and 16 hpi, suggesting that priming of these genes is an important
mechanism of P. indica-induced systemic disease resistance.
Barley leaf transcriptome and metabolite analysis reveals new aspects of compatibility and Piriformospora indica–mediated systemic induced
resistance to powdery mildew
30

Upregulated
Downregulated
Powdery mildew-regulated
transcripts in barley leaves. Venn diagram showing numbers of Blumeria
graminis f. sp. hordei-responsive transcripts (false
discovery rate < 0.05) in 3-week-old P. indica
colonized barley plants 12, 24, and 96 h post
inoculation (hpi) with the pathogen. Mean fold-inductions of transcript levels were calculated from
levels detected in three independent experiments by
hybridization to the Affymetrix Barley1 Gene Chip.
Fig. 17
31

Systemic effect of
Piriformospora indica on
expression of Blumeria
graminis f. sp. hordei (B.
graminis)–induced genes in
barley. • Transcript levels determined by quantitative real-
time polymerase chain reaction were calculated
relative to the mean of three constitutively expressed
genes.
• The gray frame highlights the 12-hpi time point for
which differential induction was originally detected
in the Gene Chip experiment.
Fig. 18
32

Systemic effect of Piriformospora indica on expression of defense-associated BCI-7
(barley chemically induced 7 ). • Determined by quantitative real-time polymerase chain reaction, were calculated relative to the mean of three constitutively expressed genes.
Fig. 19
33

Lower
Higher
(compared with mock control)
• Arrow thickness correlates with the proposed metabolic
flux relative to the other depicted metabolic pathways.
Only significantly altered metabolites and transcripts
with a fold change >2 relative to mock control are
included in the model.
• Abbreviations• Metabolites: ADPglc (ADP-glucose), aro-aa
(aromatic amino acids), aKG ( ketoglutarate), bc-aa
(branched-chain amino acid),Cit (citrate), Hex
(hexoses), HexP (hexose phosphates), Icit (isocitrate),
PEP (phosphoenol pyruvate), 3PGA (3
phosphoglycerate), Suc (sucrose), TP (triose
phosphates), UDPglc (UDP-glucose).
• Transcripts: AsnS (asparagine synthetase), CS
(chorismate synthase), cwINV (cell wall invertase),
GDH (glutamate dehydrogenase),
GS (glutamine synthetase), IPMS
(isopropylmalate synthase), P5CDH ( ’1-
pyrroline 5-carboxylate dehydrogenase),
PGM (phosphoglycerate mutase), PPT
(phosphoenolpyruvate / phosphate translocator),
SuSy (sucrose synthase).
Based on the results of the combined metabolome and transcriptome analysis, a model illustrating the major
redirection of metabolism in B. graminis–infected barley leaves was developed that integrates the dynamics in
central metabolism observed at 24 and 96 h post inoculation (hpi) (changes observed only at 96 hpi are depicted
in gray).
Fig. 20
34

Prime-omics in defense against nematode

Arbuscular mycorrhizal fungi (AMF) have great potential as biocontrol organisms against
the root-knot nematode Meloidogyne incognita which causes severe gall formation in plants.
Suppression subtractive hybridization (SSH) was used to investigate plant genes that are
specifically up-regulated in tomato roots (Solanum lycopersicum cv. Marmande) pre-colonized
by the AMF Glomus mosseae (BEG 12) after 12 days of soil inoculation with M. incognita
juveniles.
The higher expression of a selection of defense-related plant genes specifically in the
biocontrol interaction compared to in plants that were only mycorrhizal or only nematode
infected was confirmed, which pleads for the existence of mycorrhiza-induced priming of
plant defense responses.
In particular, the involvement of the phenylpropanoid pathway and reactive oxygen species
(ROS) metabolism could explain the reduced root-knot nematode infection in mycorrhizal
tomato roots, processes that have also been reported to play a pivotal role in plant resistance to
nematodes.
Vos et al., 2013Belgium
Mycorrhiza-induced resistance against the root-knot nematode Meloidogyne incognita involves priming of defense gene responses in tomato.
36
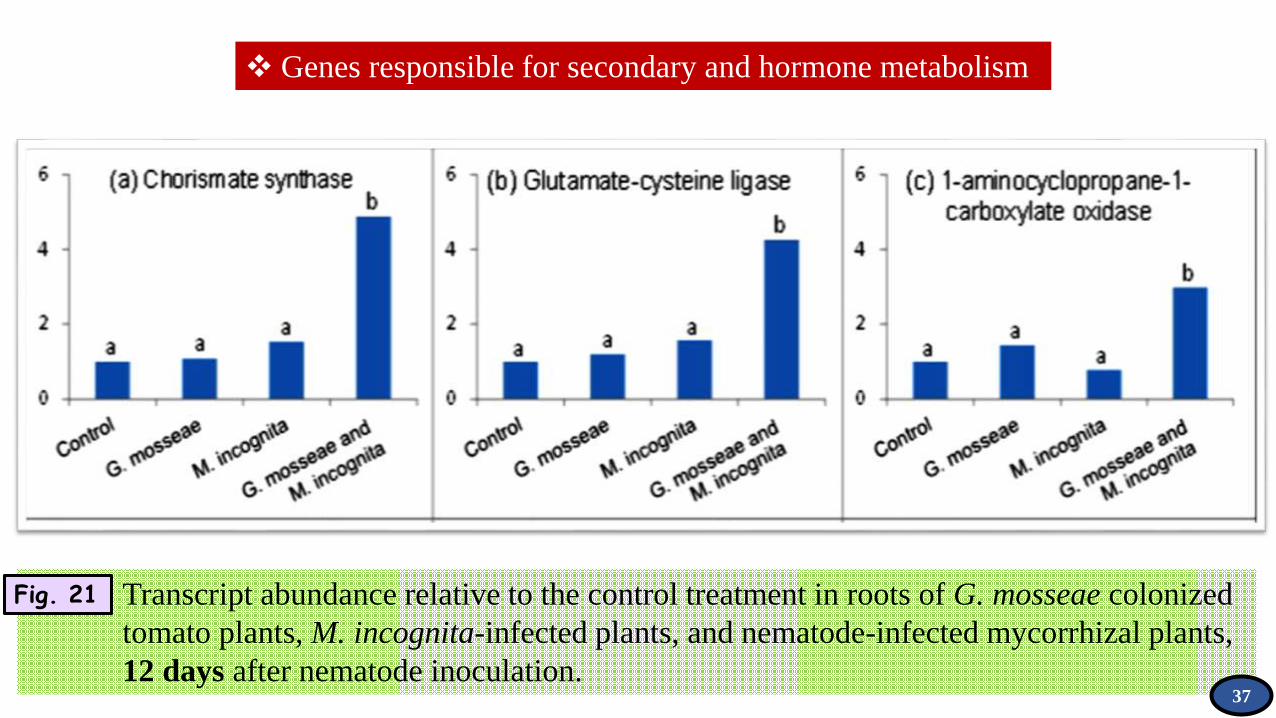
Genes responsible for secondary and hormone metabolism
Transcript abundance relative to the control treatment in roots of G. mosseae colonized
tomato plants, M. incognita-infected plants, and nematode-infected mycorrhizal plants,
12 days after nematode inoculation.
Fig. 21
37
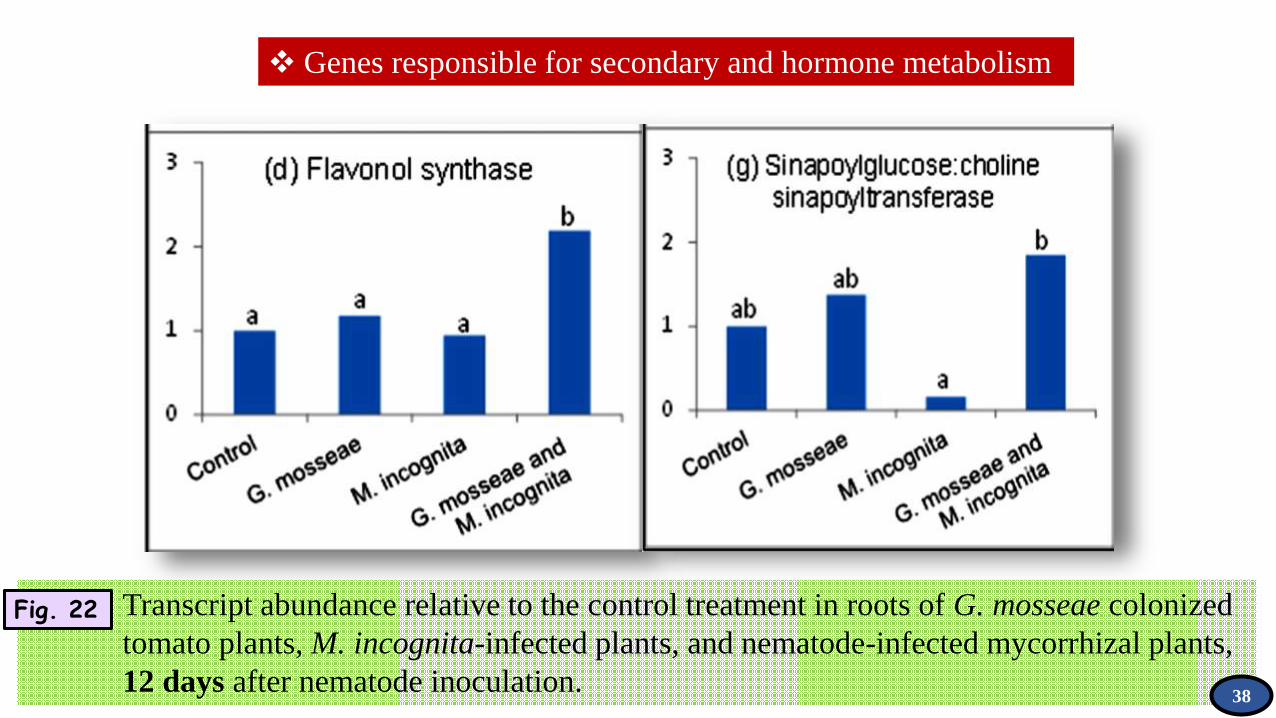
Genes responsible for secondary and hormone metabolism
Transcript abundance relative to the control treatment in roots of G. mosseae colonized
tomato plants, M. incognita-infected plants, and nematode-infected mycorrhizal plants,
12 days after nematode inoculation.
Fig. 22
38

Fig. 2339

Nematode infection parameters in tomato plants colonized or not by the AMF G.
mosseae, 12 days after M. incognita inoculation (Pi ¼ 1000).
Treatment Number
of J2 & J3
Number
of J4
Total number
of nematodes
per root system
Gall indexa
- G. mosseae 118 ± 24 b 34 ± 9 b 151 ± 27 b 2.3 ± 0.2 b
+ G. mosseae 48 ± 13 a 9 ± 2 a 58 ± 14 a 1.7 ± 0.2 a
P (treatment) 0.024 0.013 0.008 0.048
Data represent mean standard error (n ¼ 8). Within each column, values followed by different letters are significantly different (P 0.05) according to one-
way ANOVA and Tukey’s HSD test. Statistical analysis was performed on log(x þ 1) transformed nematode infection data: n.s. ¼ not significant; J2, 2nd
stage juveniles; J3, 3rd stage juveniles; J4, 4th stage juveniles. a Gall index was rated on a scale of 1e10, according to Bridge and Page (1980).
Table 3
40

Prime-omics in defense against arthropods

Stress
severity
Time
Defense against arthropods
Fig. 24Niinemets et al., 201342

Defense against arthropods
Direct defense priming can be achieved through either chemical or natural stimuli. Natural stimuli can stem from arbuscular mycorrhiza, nonpathogenic rhizobacteria, or various arthropod cues such as oviposition or insect wounding. In priming against arthropods, abscisic acid (ABA) is a key regulator that activates defenses coordinately with jasmonic acid (JA)- and systemin-dependent signaling.
Indirect defense priming is more diverse. It involves the enhancement of defense responses in neighbouring plants and also more efficient attraction of predators. Primed defenses in surrounding plants activate ocymene synthase (OS), which catalyzes the accumulation of b-ocimene.
43
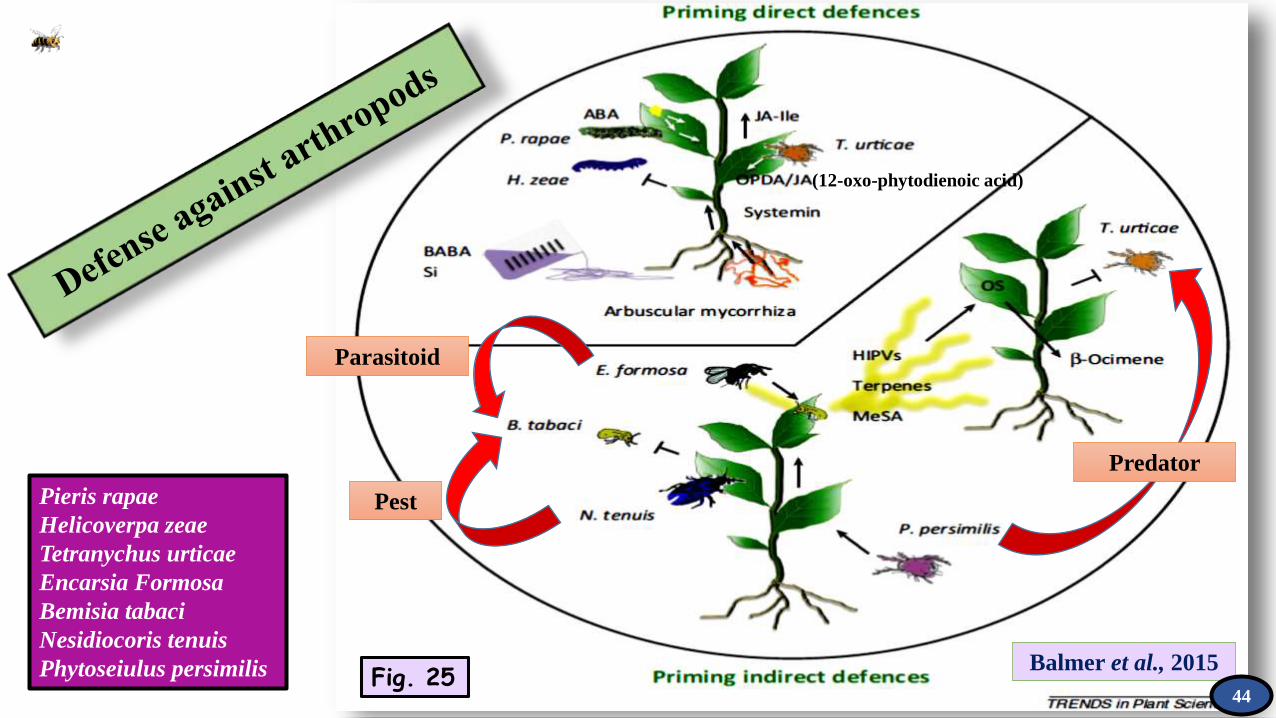
Pest
Parasitoid
Predator
(12-oxo-phytodienoic acid)
Balmer et al., 2015Fig. 25
Pieris rapae
Helicoverpa zeae
Tetranychus urticae
Encarsia Formosa
Bemisia tabaci
Nesidiocoris tenuis
Phytoseiulus persimilis44

Prime-omics of direct defense against arthropods

Mao et al., 2013China
• These results suggest a strong interaction between Si and JA in defense against insect herbivores
involving priming of JA-mediated defense responses by Si and the promotion of Si accumulation by JA.
Priming of jasmonate-mediated anti-herbivore defense responses in rice by silicon
• To explore the role of JA in Si-enhanced resistance, the expression of allene oxide synthase (OsAOS;
active in JA biosynthesis) and CORONATINE INSENSITIVE1 (OsCOI1; active in JA perception)
genes are silenced in transgenic rice plants via RNAi and examined resulting changes in Si accumulation
and defense responses against caterpillar Cnaphalocrocis medinalis (rice leaffolder, LF) infestation.
• Reduced Si deposition and Si cell expansion were observed in leaves of OsAOS and OsCOI1 RNAi
plants in comparison with wild-type (WT) plants, and reduced steady-state transcript levels of the Si
transporters OsLsi1, OsLsi2, and OsLsi6 were observed in Si-pretreated plants after LF attack.
• Upon LF attack, wild-type plants subjected to Si pretreatment exhibited enhanced defense responses relative to
untreated controls, including higher levels of JA accumulation; increased levels of transcripts encoding defense
marker genes; and elevated activities of peroxidase, polyphenol oxidase, and trypsin protease inhibitor.
46

Gain in mass of LF larvae fed on WT rice plants and the OsCOI1 RNAi and OsAOS RNAi lines
treated with Si and MeJA. • Seven-day-old seedlings were transplanted to nutrient solution containing 2 mM K2SiO3. In the Si-deficient treatment, potassium chloride was used to
replenish potassium.
• OsCOI1 and OsAOS RNAi lines and WT rice plants were sprayed with 1 mM MeJA or buffer (control) 20 d after transplanting.
• Two days later all plants were infected by third-instar LF larvae at leaf node 3.
• The individual larvae were measured 3 d later, and the mean percentage of gain in mass was calculated. Values are mean ± SE (n ( 20). Letters above
bars indicate significant differences among treatments (P < 0.05 according to Tukey’s multiple range test).
Fig. 26
47

Steady-state levels of OsAOS (A and C) and OsCOI1 (B and D) transcripts in the leaves of WT rice plants
not treated with Si or treated with Si with or without LF infestation (A and B) or treatment with MeJA (C
and D). For gene-expression experiments the treatments included no Si added (Si−); no Si added followed by MeJA treatment (Si−MeJA+) or LF infestation
(Si−LF+); 2 mM potassium silicate added (Si+); and 2 mM potassium silicate added followed by treatment with 1 mM MeJA (Si+MeJA+) or LF
infestation (Si+LF+). Real-time qRT-PCR analysis was used to determine the relative steady-state transcript levels shown. Values shown are mean ±
SE (n ( 3).
Fig. 27
Time after treatment (h)
Rel
ati
ve g
ene
exp
ress
ion
48

Steady-state levels of JA levels (E) in the leaves of WT rice plantsFor JA analysis the six treatments were Si−, Si+, Si−MeJA+, Si−LF+, Si+MeJA+, and Si+LF+; values are mean ± SE (n ( 6). For each time point,
letters above bars indicate significant difference among treatments (P < 0.05 according to Tukey’s multiple range test).
Fig. 28
49

Scanning electron micrographs of rice
leaf cross-sections. Bilobed (“dumbbell”-
shaped) Si cells from node 3 leaves of
WT, OsAOS RNAi, and OsCOI1 RNAi
plants are shown.(Scale bar: 20 μm.) Silica cells were quantified at a magnification of 2,000×. The
length and width of the Si cell were measured; then the Si area was calculated.
SC, silica cell.
Fig. 29
50
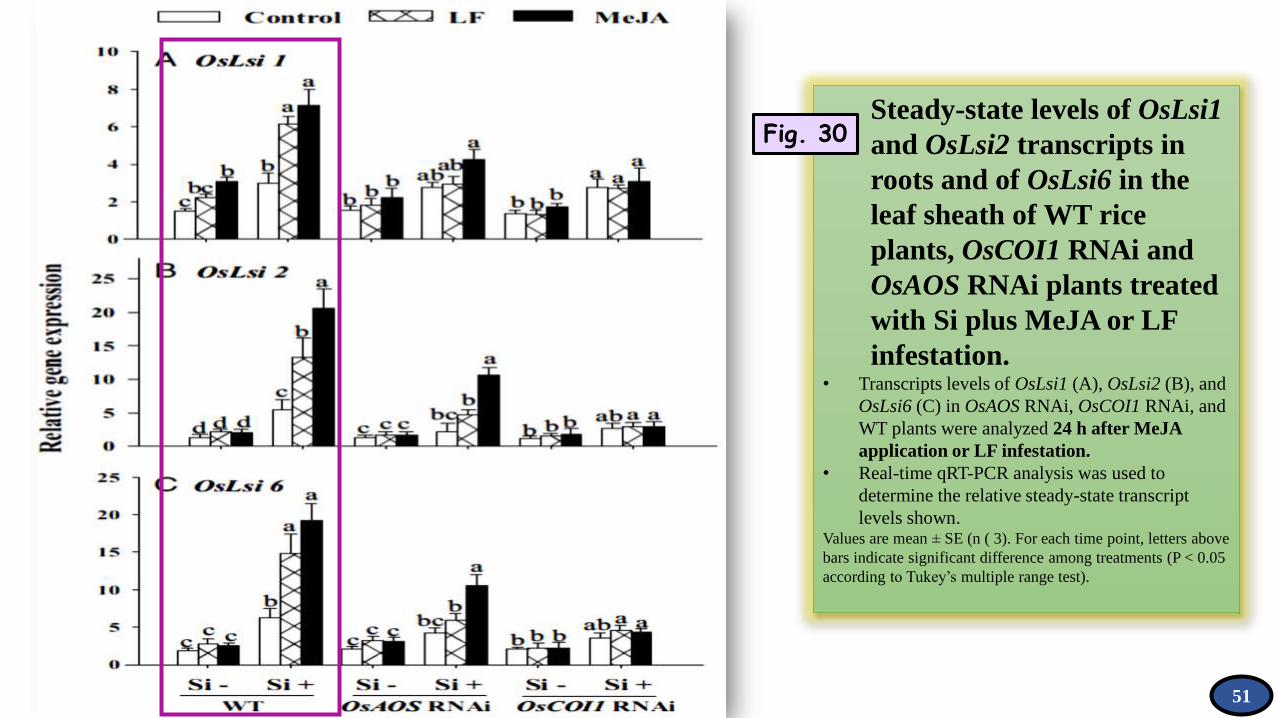
Steady-state levels of OsLsi1
and OsLsi2 transcripts in
roots and of OsLsi6 in the
leaf sheath of WT rice
plants, OsCOI1 RNAi and
OsAOS RNAi plants treated
with Si plus MeJA or LF
infestation. • Transcripts levels of OsLsi1 (A), OsLsi2 (B), and
OsLsi6 (C) in OsAOS RNAi, OsCOI1 RNAi, and
WT plants were analyzed 24 h after MeJA
application or LF infestation.
• Real-time qRT-PCR analysis was used to
determine the relative steady-state transcript
levels shown. Values are mean ± SE (n ( 3). For each time point, letters above
bars indicate significant difference among treatments (P < 0.05
according to Tukey’s multiple range test).
Fig. 30
51

Prime-omics of indirect defense by and against arthropods

Predatory mites
(Dicke & Sabelis, 1988)
Parasitoid wasps
(Turlings et al., 1990)
Predatory bugs
(Drukker et al., 1995)
Predatory lady beetles
(Ninkovic et al., 2001) Herbivorous moths, which are repelled
(de Moraes et al., 2001)Nematodes
(Rasmann et al., 2005)
Parasitic plants
(Runyon et al., 2006)
Predatory birds
(Mantyla et al., 2008)
Resistance to pathogens
(Yi et al., 2009)
Other plants
(Baldwin & Schultz, 1983;
Rhoades,1983)
Depolarization events and Ca2+ influxes in
membranes exposed to HI-VOCs (Zebelo et
al., 2012) it is likely that the next discovery
will concern their perception by local cells
that are surrounding the injured tissues.
Milestones in the research of herbivore induced-volatile organic
compounds(HI-VOCs)
Fig. 31 Martin., 2014
53

Reports on herbivore-induced volatile organic compounds (HI-VOCs) published
between 1983-2013.
Martin., 2014Fig. 32
54

Vieira et al., 2012Brazil
• It induce indirect defenses in soybean plants
against stink bugs by egg parasitoid Telenomus
podisi.
• Spraying cis-jasmone increased number of
Scelionidae egg parasitoids in soybean plots.
• Their results suggest that treatment with cis-
jasmone effectively attracted and enhanced the
population of Scelionid parasitoids, but had no
effect on the occurrence and intensity of parasitism
and in the number of stink bugs.
• The secondary metabolite cis-Jasmone activates the metabolic pathway that produces
volatile organic compounds attractive to natural enemies and, sometimes, repellent to
herbivores. cis-Jasmone is an herbivore induced plant volatile.
cis-Jasmone indirect action on egg parasitoids (Hymenoptera: Scelionidae) and its application in biological control of soybean stink bugs (Hemiptera:
Pentatomidae)
Constitutive
volatiles
Selective attraction
HIPVs
Spraying
cis-jasmone
Soybean field plotsFig. 33 55

Abundance of adult parasitoids sampled in soybean plots, with yellow sticky
traps for nine weeks in control and cis-jasmone treated plots in two areas. Legend: Aph ( Aphelinidae), Bet ( Bethylidae), Bra ( Braconidae), Cer ( Ceraphronidae), Chal ( Chalcididae), Chalni ( Chalcidoidea unidentified), Chr (
Chrysididae), Dia ( Diapriidae), Dry ( Dryinidae), Euc ( Eucoilidae), Eul ( Eulophidae), Eup ( Eupelmidae), Eur ( Eurytomidae), Eva ( Evaniidae), Fig (
Figitidae), Ich (Ichneumonidae), Meg ( Megaspilidae), Mma ( Mymarommatidae), Mym ( Mymaridae), Per ( Perilampidae), Pla ( Platygastridae), Pro (
Proctrupidae), Sce ( Scelionidae), Sig ( Signiphoridae), Tet ( Tetracampidae), Tor ( Torimidae), Tri ( Trichogrammatidae).Indicate significant difference (P
< 0,05).
Fig. 34
Ab
un
dan
ce
Family
d
56

The costs and benefits of priming
Fig. 3557

Conclusion
Prime-omics is an integrated approach for defense priming which involves different techniques spanning the fields of transcriptomics, proteomics, and metabolomics together with adequate bioinformatics tools.
Primed plants show faster and stronger defense responses when subsequently challenged by microbes, insects, or abiotic stress, and this is frequently linked to development of local and systemic immunity and stress tolerance.
The availability of rapidly growing transcriptomic, proteomic, and metabolomicdata sets – the prime-ome – describes the state of primed plants.
The knowledge of prime-omics can be exploited for a better defence modulation in plants.
58

Future thrust
Studies on adaptation occurring in the attacker stresses in response to priming
and priming response at different developmental stages of plants will suffice the
strategies to manage any deviations in response.
Research on the priming response in combination with two or more stress
should be investigated.
The work on development of metadata will speed up further research in this
area.59
