plastidial nad-dependent malate dehydrogenase: a … · [email protected] . short title: moonlighting...
TRANSCRIPT

1
RESEARCH ARTICLE
Plastidial NAD-Dependent Malate Dehydrogenase: A Moonlighting Protein Involved in Early Chloroplast Development Through its Interaction with an FtsH12-FtsHi Protease Complex Tina B. Schreier1, Antoine Cléry2, Michael Schläfli1, Florian Galbier1, Martha Stadler1, Emilie Demarsy3,4, Daniele Albertini1, Benjamin A. Maier1, Felix Kessler3, Stefan Hörtensteiner5, Samuel C. Zeeman1* and Oliver Kötting1 1 Institute of Molecular Plant Biology, ETH Zurich, Universitätstrasse 2, CH-8092 Zurich, Switzerland. 2 Institute of Molecular Biology and Biophysics, Department of Biology, ETH Zurich, CH-8093 Zurich, Switzerland. 3 Laboratory of Plant Physiology, University of Neuchâtel, CH-2000 Neuchâtel, Switzerland 4 Department of Botany and Plant Biology, University of Geneva, 30 Quai E. Ansermet, CH-1211 Geneva, Switzerland. 5 Institute of Plant Biology, University of Zürich, Zollikerstrasse 107, CH-8008 Zürich, Switzerland. *Corresponding Author: [email protected] Short title: Moonlighting role of plastidial NAD-MDH One-sentence summary: Plastid NAD-dependent malate dehydrogenase is essential for chloroplast development, not because of its enzymatic activity, but because it interacts with FtsH proteases at the inner envelope membrane. The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy described in the Instructions for Authors (www.plantcell.org) is: Samuel C. Zeeman ([email protected]). ABSTRACT Malate dehydrogenases (MDH) convert malate to oxaloacetate using NAD(H) or NADP(H) as a cofactor. Arabidopsis thaliana mutants lacking plastidial NAD-dependent MDH (pdnad-mdh) are embryo-lethal, and constitutive silencing (miR-mdh-1) causes a pale, dwarfed phenotype. The reason for these severe phenotypes is unknown. Here, we rescued the embryo lethality of pdnad-mdh via embryo-specific expression of pdNAD-MDH. Rescued seedlings developed white leaves with aberrant chloroplasts and failed to reproduce. Inducible silencing of pdNAD-MDH at the rosette stage also resulted in white newly emerging leaves. These data suggest that pdNAD-MDH is important for early plastid development, which is consistent with the reductions in major plastidial galactolipid, carotenoid and protochlorophyllide levels in miR-mdh-1 seedlings. Surprisingly, the targeting of other NAD-dependent MDH isoforms to the plastid did not complement the embryo lethality of pdnad-mdh, while expression of enzymatically inactive pdNAD-MDH did. These complemented plants grew indistinguishably from the wild type. Both active and inactive forms of
Plant Cell Advance Publication. Published on June 22, 2018, doi:10.1105/tpc.18.00121
©2018 American Society of Plant Biologists. All Rights Reserved

2
pdNAD-MDH interact with a heteromeric AAA-ATPase complex at the inner membrane of the chloroplast envelope. Silencing the expression of FtsH12, a key member of this complex, resulted in a phenotype that strongly resembles miR-mdh-1. We propose that pdNAD-MDH is essential for chloroplast development due to its moonlighting role in stabilizing FtsH12, distinct from its enzymatic function. INTRODUCTION 1
2
Malate dehydrogenases (L-malate-NAD-oxidoreductase [MDH]; EC 1.1.1.37) 3
catalyse the reversible interconversion of malate and oxaloacetate, using NAD(H) or 4
NADP(H) as a cofactor. Plant MDHs form a family of enzymes, with isoforms that are 5
present in several compartments within the cell. The Arabidopsis thaliana genome 6
encodes nine isoforms of MDH - two plastidial, two peroxisomal, two mitochondrial, 7
and three that are assumed to be cytosolic, as they have no predicted localisation 8
sequence. All of these isoforms use NAD+ as a cofactor, with the exception of one 9
plastidial isoform that is NADP-dependent (NADP-MDH). Although it is well 10
established that MDHs play an important role in central metabolism in plants, the 11
exact role(s) of many of the individual isoforms in Arabidopsis remains unclear - 12
particularly for the plastidial and cytosolic isoforms. 13
14
MDHs play a critical role in the tricarboxylic acid (TCA) cycle in the mitochondria, 15
where they generate NADH by oxidising malate to oxaloacetate. They can also 16
reduce oxaloacetate back to malate in order to generate NAD+ for the 17
decarboxylation reaction that occurs during the conversion of glycine to serine 18
(Journet et al., 1981). An Arabidopsis double mutant lacking both mitochondrial MDH 19
isoforms is viable but has defects in seed germination, stunted growth, and altered 20
leaf respiration and photorespiration rates (Tomaz et al., 2010; Sew et al., 2016). 21
Mitochondrial MDHs are also involved in the carbon-concentrating mechanism in 22
plants that conduct the NAD-malic enzyme type of C4 photosynthesis (Hatch and 23
Osmond, 1976). The two peroxisomal MDH isoforms generate NAD+, which is mainly 24
required for the β-oxidation of fatty acids. An Arabidopsis mutant deficient in both of 25
these isoforms does not efficiently mobilise triacylglycerols and requires the 26
exogenous supply of sugars for seedling establishment (Pracharoenwattana et al., 27
2007, 2010). 28
29

3
In the plastids, NADP-MDH was proposed to be the key enzyme in the malate valve 30
– a mechanism by which reducing equivalents can be indirectly transported across 31
membranes of organelles (Heber, 1974; Scheibe, 2004; Taniguchi and Miyake, 32
2012). NADP-MDH is redox-regulated by the ferredoxin-thioredoxin system and is 33
therefore thought to be active only in the light (Scheibe, 1987). According to the 34
malate valve hypothesis, NADPH is generated in the chloroplast through the electron 35
transport chain during the day but does not readily diffuse out of the chloroplast. 36
NADP-MDH reduces oxaloacetate to malate by oxidising NADPH to NADP, and the 37
malate is shuttled to the cytosol in exchange for oxaloacetate via the dicarboxylate 38
transporter, AtpOMT1. In the cytosol, malate is oxidised back to oxaloacetate by the 39
cytosolic MDHs, regenerating a reducing equivalent in the form of NADH in that 40
compartment (Kinoshita et al., 2011). However, the importance of the malate valve is 41
unclear. Previous studies of the nadp-mdh knockout mutant have reported either no 42
reduction in the growth of mutants compared to the wild type (Hebbelmann et al., 43
2012) or a slight reduction (Heyno et al., 2014). The latter study also reported that 44
the mutant had higher H2O2 levels under high light conditions, probably because it 45
could not reversibly inactivate catalase activity. 46
47
Unlike NADP-MDH, the activity of the plastidial NAD-dependent malate 48
dehydrogenase (pdNAD-MDH) is not redox sensitive (Berkemeyer et al., 1998). 49
pdNAD-MDH was thus proposed to play an important role in the malate valve in dark 50
and non-green plastids. In contrast to the nadp-mdh mutant, the homozygous pdnad-51
mdh knockout mutant is embryo-lethal (Beeler et al., 2014; Selinski et al., 2014) - a 52
phenotype that was not reported for mutants of any other MDH isoform so far. An 53
artificial microRNA silencing construct was used to generate the miR-mdh-1 line, 54
with constitutively reduced pdNAD-MDH levels using the 35S promoter (Beeler et al., 55
2014). This line was viable but had pale leaves with disordered chloroplast 56
ultrastructure and was severely compromised in growth. Levels of malate, starch, 57
and glutathione during the night were higher in miR-mdh-1 compared to the wild 58
type. The night-time respiration rate was also lower in the silencing line. These 59
findings are consistent with a role for pdNAD-MDH in metabolism in the dark, but it is 60
difficult to pinpoint a specific role for the enzyme due to the highly pleiotropic nature 61
of the miR-mdh-1 phenotype. In addition, Selinski et al. (2014) showed that pollen 62
tube growth is affected in pdnad-mdh in vitro, but not in vivo and proposed that the 63

4
maternal tissue is able to supply substrates for an enzyme generating NAD+, 64
allowing proper tube elongation in vivo. Together, these data suggest that pdNAD-65
MDH is important for embryogenesis and subsequent growth and that its loss cannot 66
be compensated by the presence of the NADP-MDH in the plastid or of NAD-MDH in 67
other compartments. However, the link between the activity of the enzyme and the 68
phenotypes resulting from its loss remain unclear. Here, we aimed to investigate the 69
role of pdNAD-MDH in both embryo development and post-embryonic growth, 70
focusing on chloroplast development, and to test the importance of NAD-MDH 71
activity in these processes. 72
73
74
RESULTS 75
76
Embryo-Specific Expression of pdNAD-MDH Complements the Embryo 77
Lethality of pdnad-mdh 78
79
We previously reported that the Arabidopsis pdnad-mdh knockout mutant has an 80
embryo-lethal phenotype and that the silencing line with reduced pdNAD-MDH 81
expression, miR-mdh-1 (carrying a 35S promoter-driven artificial microRNA 82
construct), has a pale, dwarfed phenotype with aberrant chloroplast ultrastructure 83
(Beeler et al., 2014). To investigate how the complete loss of pdNAD-MDH affects 84
post-embryogenic growth, we expressed pdNAD-MDH specifically in the embryos of 85
pdnad-mdh. For this, we cloned a PABI3:pdNAD-MDH-YFP construct encoding the 86
pdNAD-MDH protein with a C-terminal YFP tag, driven by the embryo-specific ABI3 87
promoter (Figure 1A). Previous studies have effectively used the ABI3 promoter to 88
rescue the embryo lethality of various mutants (Despres et al., 2001; Gómez et al., 89
2010; Candela et al., 2011; Bodi et al., 2012). We transformed heterozygous pdnad-90
mdh plants with the construct and selected transformed T1 plants using a BASTA-91
resistance marker. The genotype of these T1 plants were determined by PCR 92
amplification of the pdnad-mdh T-DNA insertion, as previously described (Beeler et 93
al., 2014). Four independent lines were selected for further analysis. Plants 94
homozygous for the PABI3:pdNAD-MDH-YFP transgene and heterozygous for pdnad-95
mdh were identified in the T3 generation. When we opened the siliques of these 96
plants, we found that all seeds within the silique were green (Figure 1B). All seeds 97
were also green in siliques of wild type Ler plants, and of heterozygous pdnad-mdh 98

5
plants that were complemented with pdNAD-MDH-YFP expressed under its native 99
promoter (homozygous for the PpdNAD-MDH:pdNAD-MDH-YFP construct, described in 100
Beeler et al., 2014). In contrast, approximately one quarter of the seeds contained in 101
siliques of untransformed (UT) heterozygous pdnad-mdh plants were white. 102
Expression of pdNAD-MDH-YFP under the control of the ABI3 promoter was 103
visualised in isolated embryos using fluorescence microscopy, where we observed 104
YFP signal in embryos at various stages of embryogenesis (globular, heart and 105
torpedo stage) (Figure 1C). These findings suggest that the embryo-specific 106
expression of pdNAD-MDH-YFP can overcome the embryo-lethal phenotype of 107
pdnad-mdh. 108
109
We germinated the progeny from plants that were heterozygous for pdnad-mdh and 110
homozygous for the PABI3:pdNAD-MDH-YFP transgene. At the seedling stage, most 111
of these plants resembled the wild-type (Ler) or heterozygous pdnad-mdh plants, but 112
approximately one quarter had pale cotyledons resembling those observed in miR-113
mdh-1 (Figure 2A). These pale seedlings grew very slowly, and once enough 114
material could be harvested for genotyping, they were confirmed to be homozygous 115
for pdnad-mdh. These findings confirmed that the PABI3:pdNAD-MDH-YFP construct 116
could rescue the embryo-lethal phenotype of pdnad-mdh but show that after 117
embryogenesis when the ABI3 promoter is no longer active, these plants become 118
greatly compromised. Aside from the pale cotyledons, plants homozygous for pdnad-119
mdh initiated albino ‘true leaves’ that failed to undergo proper organogenesis, 120
remaining as small white primordia-like stubs on the meristem (Figure 2B). These 121
plants died 2-4 weeks after germination and failed to reach the reproductive stage. 122
The addition of sucrose in the growth medium did not prevent the seedling-lethality 123
of these plants, nor did it improve growth (Supplemental Figure 1). By contrast, 124
homozygous pdnad-mdh plants that were complemented with pdNAD-MDH-YFP 125
expressed under its native promoter were indistinguishable from the wild type 126
(Figure 2A). 127
128
We previously observed that the young leaves of miR-mdh-1 had compromised 129
chloroplast ultrastructure, with fewer thylakoid membranes and starch granules 130
(Beeler et al., 2014). We therefore investigated chloroplast ultrastructure in pdnad-131
mdh PABI3:pdNAD-MDH-YFP seedlings using transmission electron microscopy 132

6
(TEM). We fixed and embedded cotyledon and true leaf samples from 3-4-week-old 133
Ler and pdnad-mdh plants rescued with PABI3:pdNAD-MDH-YFP. The chloroplasts in 134
cotyledons of the rescued pdnad-mdh seedlings contained thylakoid membranes, but 135
they were not as structured as those in wild-type cotyledons (Figure 2C). However, 136
in the white true leaves, no mature chloroplast structures were observed, only 137
proplastid-like structures. We suspect that the chloroplasts in the cotyledons 138
developed further than those in the true leaves due to residual pdNAD-MDH derived 139
from its PABI3-driven expression in cotyledons during embryogenesis (Figure 1C). To 140
confirm that homozygous pdnad-mdh PABI3:pdNAD-MDH-YFP plants do not express 141
pdNAD-MDH protein after embryogenesis, we extracted proteins from these 142
seedlings and performed immunoblots with the pdNAD-MDH antibody, as well as 143
native-PAGE gels followed by MDH activity staining. No bands corresponding to 144
pdNAD-MDH or pdNAD-MDH-YFP were detected in these extracts on the 145
immunoblot (Figure 3A). We observed several unspecific bands on the blot, but our 146
previous cross-reactivity test of this antibody suggests that they are unlikely to be 147
other MDH isoforms (Beeler et al., 2014). These bands were visible in all lines but 148
variable in abundance, possibly due to variation in the amounts of chloroplast 149
proteins such as Rubisco between the lines (gels were loaded on an equal protein 150
basis). On native-PAGE gels, pdNAD-MDH runs as three distinct activity bands – 151
where the lower band corresponds to the free dimer, while the upper bands 152
correspond to pdNAD-MDH in protein-protein interactions. The lower band is difficult 153
to resolve from other NAD-MDH isoforms (Beeler et al., 2014). Thus, it should be 154
noted that only part of the total pdNAD-MDH in the extract can be observed with this 155
technique. The two upper pdNAD-MDH activity bands were clearly observed in 156
extracts of Ler, but not in miR-mdh-1 and pdnad-mdh PABI3:pdNAD-MDH-YFP 157
(Figure 3B). These data confirm that PABI3:pdNAD-MDH-YFP was only active during 158
embryogenesis. Any residual protein from the embryonic tissues was either inactive 159
by this time or below the level of detection. 160
161
Inducible Silencing of pdNAD-MDH at the Rosette Stage Results in the 162
Formation of White Leaves from the Meristem 163
164
To further investigate the post-embryonic role of pdNAD-MDH, we generated 165
Arabidopsis lines where pdNAD-MDH silencing could be induced at later growth 166
stages. The identical microRNA silencing cassette used in miR-mdh-1 was cloned 167

7
downstream of the XVE/OlexA ß–estradiol-inducible promoter system. Wild-type 168
plants were transformed with the construct, and transformed T1 seedlings were 169
selected using the Hygromycin resistance marker. Prior to ß-estradiol treatment, 3-170
week-old resistant T2 plants were phenotypically indistinguishable from the wild type. 171
A ß-estradiol solution was then sprayed onto the entire rosettes, and this treatment 172
was repeated every two days. Six days into the treatment, newly emerging leaves 173
were noticeably pale, whereas the old leaves stayed green (Figure 4A). Paleness 174
was not observed in any leaves of wild-type plants treated with ß-estradiol. 175
Immunoblot analysis of proteins extracted from these rosettes confirmed that 176
pdNAD-MDH protein levels were undetectable in the young white leaves of the 177
transgenic line, and only residual amounts of protein were present in the old green 178
leaves (Figure 4B and C). The phenotype of the newly emerging leaves suggests 179
that pdNAD-MDH deficiency particularly affects tissues in which chloroplasts are 180
developing. The severe reduction in pdNAD-MDH levels did not have a visible effect 181
on the older leaves, which contain mature chloroplasts. 182
183
pdNAD-MDH is Important for Early Etioplast and Chloroplast Development 184
185
To investigate the role of pdNAD-MDH in chloroplast development in more detail, we 186
studied the de-etiolation process in dark-grown miR-mdh-1 seedlings. During de-187
etiolation, plastids undergo a rapid conversion from partially developed chloroplasts 188
(etioplast) to photosynthetic chloroplasts upon illumination (Solymosi and Aronsson, 189
2013). The prolamellar body (PLB), a crystalline lattice structure within etioplasts, 190
contains structural building blocks for the photosynthetic apparatus, including 191
protochlorophyllide, the enzyme protochlorophyllide oxireductase A (PORA), 192
carotenoids, and fragments of membranes that will form the thylakoids 193
(prothylakoids) (Bahl et al., 1976; Ryberg and Sundqvist, 1982, 1988; Park et al., 194
2002). 195
196
Surprisingly, when grown in the dark, the etiolated miR-mdh-1 seedlings were 197
indistinguishable from the wild type, indicating that skotomorphogenic growth is 198
unaffected by pdNAD-MDH deficiency (Figure 5A). Quantification of the hypocotyl 199
length showed no significant difference between miR-mdh-1 and the wild type 200
(Figure 5B). However, growth of the seedlings in the light was affected in the miR-201

8
mdh-1 seedlings. When grown under a 12-h light/12-h dark regime, miR-mdh-1 202
seedlings had cotyledons that were paler and smaller than those of the wild type 203
(Figure 5A). When grown under continuous light, root growth was further 204
compromised in miR-mdh-1. 205
206
We then examined etioplast structure in cotyledons of etiolated wild-type and miR-207
mdh-1 seedlings using TEM. Etioplasts were imaged from sections produced from 208
three different six-day-old seedlings for both miR-mdh-1 and the wild type. The 209
observed etioplasts were categorised according to their PLB structure – either 210
normal, compromised (with a less ordered structure), or absent (no internal structure 211
observed within the etioplast). Examples of wild-type and miR-mdh-1 etioplasts 212
within each category are shown in Supplemental Figure 2. In wild-type cotyledons, 213
we observed that the vast majority of etioplasts (88.5% - example shown in Figure 214
5C) had normal PLB structure, and only 1.4% had compromised structure (Figure 215
5D). PLBs were absent in 10.1% of etioplasts, but this is likely because they were 216
not visible within that thin section of the etioplast. However, in miR-mdh-1, most 217
etioplasts had compromised PLB morphology (59.8% - example shown in Figure 5C) 218
and a substantial proportion had no visible PLB (36.3%). 219
220
We then quantified the abundance of major prolamellar body components in 221
etiolated miR-mdh-1 seedlings. First, we quantified protochlorophyllide and its 222
binding protein, PORA, using a fluorescence-based assay (Cheminant et al., 2011) 223
and immunoblots, respectively. The levels of both components were greatly reduced 224
in 5-day-old etiolated miR-mdh-1 seedlings compared to the wild type (Figure 6A-B). 225
We also quantified major galactolipids and carotenoids in 6-day-old etiolated 226
seedlings and, since these molecules are also major components of mature 227
chloroplasts, we simultaneously analysed extracts of photomorphogenically grown 228
seedlings. Several monogalactosyl- and digalactosyl-diacylglycerol (MGDG, DGDG) 229
species were detected in the extracts. The most abundant MGDG species was 230
MGDG-18:3/16:3, while DGDG-18:3/18:3 was the most abundant DGDG. 231
Interestingly, the levels of MGDG-18:3/16:3 and DGDG-18:3/18:3 were greatly 232
reduced in miR-mdh-1 in both dark-grown and light-grown seedlings, even though 233
there was no difference in the levels of diacylglycerol (DAG) precursor (Figure 6C-E). 234
Similar trends were observed for most other detectable MGDG and DGDG species 235

9
(Supplemental Table 1). Levels of major carotenoids, such as ß-carotene, were also 236
reduced in miR-mdh-1 etiolated seedlings, as well as light-grown seedlings, relative 237
to the wild type (Figure 6F). Similar trends were observed for lutein and 238
viola/zeaxanthin (Supplemental Table 1). In summary, miR-mdh-1 seedlings were 239
deficient in major lipids and carotenoids that are normally present in etioplasts or 240
chloroplasts. Etioplast development is greatly affected by pdNAD-MDH deficiency, 241
even though etiolated growth is not affected. These data further suggest a critical 242
role of pdNAD-MDH in the early stages of chloroplast development. 243
244
The pdNAD-MDH Protein is Required for Proper Embryo and Chloroplast 245
Development, but its Enzymatic Activity is Not 246
247
We tested whether we could complement the pdnad-mdh mutant by introducing 248
other NAD-MDH isoforms into the plastid – thereby restoring NAD-MDH activity. The 249
Arabidopsis NAD-MDH isoforms in the mitochondria, peroxisome and the cytosol 250
have amino acid sequences that are similar to pdNAD-MDH (Supplemental Figure 251
3). We therefore chose one isoform from each compartment (AtcyMDH1, AtmMDH1 252
and AtpMDH1) and a more distantly related mitochondrial MDH from yeast 253
(Saccharomyces cerevisiae; ScmMDH1). The coding sequences of the different 254
MDH isoforms were fused directly downstream of the plastidial transit peptide from 255
the RubisCO small subunit (RbcS; amino acids 1-80). For the mitochondrial 256
isoforms, the mitochondrial transit sequence was first predicted using the TargetP 257
1.1 program (Nielsen and Engelbrecht, 1997; Emanuelsson et al., 2000), and this 258
sequence was replaced with the plastidial one. The constructs were driven by the 259
native pdNAD-MDH promoter and encoded a YFP tag fused to the C-terminal end of 260
each MDH protein (Figure 7A). Constructs were also generated with the smaller 261
Flag-HA peptide tag in place of the YFP. Heterozygous pdnad-mdh plants were 262
transformed with these constructs, and T1 seedlings were selected via BASTA 263
resistance. Expression of the transgene was verified by immunoblotting. All resistant 264
T1 plants were genotyped (5-15 plants per line) for the parental pdnad-mdh 265
mutation, but no homozygous mutant plants were found. We therefore selected T1 266
plants that were both expressing the transgene and heterozygous for pdnad-mdh, 267
and genotyped the BASTA-resistant T2 progeny (120-180 plants per construct from 268
5 independent T1 lines [30-36 T2 plants each]). However, we were not able to isolate 269
any plants that were homozygous for pdnad-mdh (Figure 7B). We verified that all of 270

10
the fusion-proteins were correctly targeted to the chloroplast using confocal 271
microscopy on heterozygous pdnad-mdh plants that were transformed with the YFP-272
tagged constructs. In all lines, YFP-signal was detected exclusively in the 273
chloroplasts (Figure 7C). We also verified that the constructs encoded active MDH 274
isoforms via native-PAGE with activity staining. Additional activity bands were 275
observed in extracts from plants expressing AtmMDH1 or ScmMDH1, suggesting 276
that these proteins were indeed active (Supplemental Figure 4). The other isoforms 277
are also potentially active, but their activity on the native-PAGE gel might be masked 278
by the activity bands of the endogenous MDH isoforms. In summary, restoring 279
plastidial NAD-MDH activity alone in pdnad-mdh could not complement the embryo-280
lethal phenotype. 281
282
Given these findings, we questioned whether the enzymatic activity of pdNAD-MDH 283
is required at all. Therefore, we generated inactive versions of the pdNAD-MDH 284
protein using site-directed mutagenesis to test whether they could complement the 285
pdnad-mdh mutant. The catalytic site of the malate dehydrogenases is highly 286
conserved among all homologs and includes a histidine, two aspartate and three 287
arginine residues (Birktoft and Banaszak, 1983; Musrati et al., 1998; Minárik et al., 288
2002) (Figure 8A). These amino acids are required to co-ordinate both the substrate 289
and nicotinamide ring of the cofactor within the catalytic pocket. The MDH reaction is 290
initiated by cofactor binding, which then facilitates substrate binding (Silverstein and 291
Sulebele, 1969). When the ternary complex is formed, an external loop closes over 292
the substrate and the residues involved in catalysis (Nicholls et al., 1992; Goward 293
and Nicholls, 1994). Additionally, two conserved arginines on a flexible loop are 294
brought into close proximity to the substrate (Clarke et al., 1986; Grau et al., 1981; 295
Wigley et al., 1992). We mutated the arginines at positions 162 (on the flexible loop) 296
and 234 (in the active site) to glutamines, resulting in three different pdNAD-MDH 297
variants - two single amino acid mutations and one containing both mutations. Both 298
Arg162 and Arg234 interact directly with malate, and substitution with glutamine, 299
which has a smaller side chain and lacks a positive charge, should destabilise the 300
substrate-binding site. We tested the effect of these mutations on enzyme activity in 301
vitro. Wild-type pdNAD-MDH protein and proteins containing the mutations were 302
expressed in and purified from Escherichia coli and incubated with oxaloacetate and 303
NADH. The reduction of NADH was monitored spectrophometrically at 340 nm. No 304

11
activity was detected for any of the three mutated proteins (Figure 8B). We then 305
created plant expression constructs encoding these enzymatically-inactive pdNAD-306
MDH proteins fused to a Flag-HA tag at the C-terminal end, and driven by the native 307
pdNAD-MDH promoter. Heterozygous pdnad-mdh plants were transformed with the 308
constructs, and transformed T1 plants were selected via BASTA resistance. As 309
described above for the experiments shown in Figure 7B, we selected T1 individuals 310
that were both expressing the transgene and heterozygous for pdnad-mdh and 311
genotyped their T2 progeny (69-78 plants per construct from 3 independent lines 312
[23-26 plants each]). Surprisingly, we identified individuals that were homozygous for 313
pdnad-mdh in the T2 generation for all catalytically-inactive pdNAD-MDH constructs, 314
indicating that each of them could complement the embryo-lethal phenotype (Figure 315
8C). The complemented pdnad-mdh plants grew like the wild type, showing that the 316
enzymatically-inactive pdNAD-MDH variants also complemented the growth 317
phenotypes of pdnad-mdh (Figure 8D). We confirmed (using native-PAGE) that no 318
activity bands corresponding to pdNAD-MDH were detected in extracts from 319
complemented pdnad-mdh plants (Figure 8E). Thus, the embryo-lethal phenotype of 320
pdnad-mdh, as well as the pale dwarfed phenotype of miR-mdh-1, are caused 321
primarily by the lack of pdNAD-MDH protein itself, rather than the deficiency of NAD-322
MDH activity. 323
324
pdNAD-MDH Interacts with the FtsH12-FtsHi Complex, which is Involved in 325
Chloroplast Development 326
327
Since the pdNAD-MDH protein itself is indispensable for chloroplast and embryo 328
development, we investigated whether it interacts with other plastidial proteins. We 329
generated stable Arabidopsis transgenic lines overexpressing YFP-tagged pdNAD-330
MDH under the control of the constitutive 35S promoter and extracted protein from 331
these lines at three different developmental stages – etiolated and light grown 332
seedlings, as well as rosette leaves. We then conducted immunoprecipitation (IP) 333
experiments with these extracts using beads that specifically bind YFP. Proteins in 334
the IP were digested with trypsin, and the resulting peptides were analysed using 335
LC-MS/MS. The identified peptides (Table 1, Supplemental Data Set 1) were 336
searched against the TAIR10 genome annotation database. As a control, we also 337
analysed IPs conducted on extracts from wild-type plants. 338
339

12
Within the IP, the largest number of peptides, apart from pdNAD-MDH itself, 340
matched hypothetical chloroplast open reading frame 2 (Ycf2), suggesting that it 341
was present in the highest abundance – although it should be noted that our analysis 342
is semi-quantitative, as peptide counts are not strictly correlated with protein 343
abundance. Ycf2 is encoded in the chloroplast genome. It is an essential protein in 344
tobacco (Nicotiana tabacum) and green algae (Chlamydomonas reinhardtii) 345
(Drescher et al., 2000; Nickelsen, 2005). Strikingly, many peptides also matched the 346
FtsH (filamentous temperature sensitive) proteases, chloroplast isoforms of which 347
are also essential (Wagner et al., 2012). The second largest functional group of 348
proteins identified in the IP experiment consists of the proteins involved in starch 349
degradation (including Like SEX4 1 (LSF1), BETA-AMYLASE1 (BAM1) and BETA-350
AMYLASE3 (BAM3)). These proteins were detected particularly at the rosette stage. 351
Although these proteins are involved in starch turnover in the leaf mesophyll and 352
stomatal guard cells, no defects in chloroplast structure were reported for mutants 353
deficient in these proteins (Fulton et al., 2008; Comparot-Moss et al., 2010; Horrer et 354
al., 2016). Given the essential nature of the identified FtsH12-FtsHi complex 355
subunits, we focused on the interaction between pdNAD-MDH and these proteins. 356
The possible role of pdNAD-MDH in starch turnover is being pursued in a separate 357
study. 358
359
FtsH proteins are a family of membrane-bound proteases containing an ATPase 360
associated with various cellular activities (AAA-ATPase) domain and a zinc-binding 361
metalloprotease domain. FtsH proteins are restricted to the mitochondria and 362
chloroplasts in eukaryotes (Wagner et al., 2012). The Arabidopsis genome encodes 363
17 FtsH genes – twelve predicted proteolytic isoforms and five predicted non-364
proteolytic FtsH isoforms that lack the zinc-binding motif for proteolytic activity 365
(FtsHi1-5). FtsH12, along with FtsHi1, FtsHi2, FtsHi3, FtsHi4 and FtsHi5, are located 366
at the inner membrane of the chloroplast envelope, and these subunits are proposed 367
to form a hetero-hexameric complex, as deduced from co-expression analysis and 368
proteomics data (Ferro et al., 2010). FtsH12 and its associated FtsHi subunits (with 369
the exception of FtsHi3) were consistently detected in the IP with pdNAD-MDH at all 370
three of the developmental stages tested. 371
372

13
For further analysis, we focused on FtsH12, as it was the only FtsH protein identified 373
in the IP that has an intact zinc-binding motif, and therefore the potential for 374
proteolytic activity. To confirm the interaction with pdNAD-MDH, we conducted a 375
reciprocal immunoprecipitation with tagged FtsH12 protein. The FtsH12 coding 376
sequence was cloned downstream of the UBIQUITIN10 promoter, and in frame with 377
a YFP-tag on the C-terminal end (UBI10:FtsH12-YFP). Wild-type plants were 378
transformed with the construct, and the IP was performed as described above on T1 379
plants overexpressing FtsH12-YFP. Within the immunoprecipitate, we found 380
peptides matching pdNAD-MDH, as well as those matching all of the FtsHi subunits 381
that were identified in the IP with pdNAD-MDH-YFP and Ycf2 (Table 2). The correct 382
plastidial localisation of the FtsH12-YFP protein was confirmed via confocal 383
microscopy (Supplemental Figure 5A). 384
385
To investigate the function of FtsH12, four independent Arabidopsis mutants 386
harbouring T-DNA insertions in the FtsH12 gene were obtained. We confirmed that 387
ftsh12 mutants are embryo-lethal, as previously described (Patton et al., 1991; 388
Franzmann et al., 1995; Patton et al., 1998). Like pdnad-mdh, the ftsh12 mutants 389
arrested near the globular-to-heart transition stage. However, the exact stage at 390
which the embryo arrested varied between lines, likely due to the position of the T-391
DNA insertions having different effects on transcript and protein accumulation 392
(Supplemental Figure 5B-F). 393
394
To further study the effect of FtsH12 deficiency, we generated lines constitutively 395
expressing artificial microRNAs. We designed microRNA silencing cassettes 396
targeting regions 3259-3279 (amiRNA target B) and 4777-4797 (amiRNA target A) of 397
the FtsH12 coding sequence, respectively (Figure 9A) (Ossowski et al., 2008). Both 398
cassettes were cloned downstream of the constitutive 35S promoter. Wild-type 399
plants were transformed with the constructs, and T1 seedlings were selected via 400
Kanamycin resistance. For both silencing constructs, the T1 plants had varying 401
degrees of paleness - from wild-type-like plants to very pale plants that were stunted 402
in growth (Figure 9B). Notably, the pale plants strongly resembled miR-mdh-1 plants. 403
Immunoblots were performed on protein extracts from the leaves of these silencing 404
lines, using antibodies against FtsH12 and pdNAD-MDH to test the impact of FtsH12 405
silencing on these proteins. The molecular weight of FtsH12 is 115 kDa, including 406

14
the chloroplast transit peptide (49 amino acids), and the mature peptide is predicted 407
to be 110 kDa. No FtsH12 protein was detectable in the T1 plants with the strongest 408
pale phenotype (amiRNA FtsH12 A 3-1, amiRNA FtsH12 B 2-1), and those with 409
intermediate levels of paleness had reduced amounts of FtsH12 protein (Figure 9C). 410
Surprisingly, there was also no detectable FtsH12 protein in the miR-mdh-1 plants. 411
Quantitative immunoblot analysis showed a 7-fold reduction in FtsH12 protein 412
abundance in miR-mdh-1 leaves relative to wild type (Supplemental Figure 6. 413
However, pdNAD-MDH protein levels were unaffected by FtsH12 silencing. 414
415
pdNAD-MDH Activity is not Required for its Interaction with the FtsH12-FtsHi 416
Complex 417
418
We tested whether the FtsH12-FtsHi complex could also interact with the NAD-MDH 419
isoforms from other cellular compartments that could not complement the embryo 420
lethality of pdnad-mdh when targeted to chloroplasts. We extracted proteins from 4-421
week-old rosettes of heterozygous pdnad-mdh plants expressing the YFP-tagged 422
NAD-MDH isoforms (Figure 7A) and performed anti-YFP IPs for analysis by LC-423
MS/MS. IPs were also performed with extracts from pdnad-mdh PpdNAD-MDH:pdNAD-424
MDH-YFP plants (as a positive control) and from wild-type Ler plants (as a negative 425
control). We did not detect peptides matching Ycf2, FtsH12 or the FtsHi proteins in 426
the IPs with most NAD-MDH-YFP isoforms. One exception was the IP with 427
AtmMDH1-YFP, where four peptide hits were found matching Ycf2 and four 428
matching FtsH12, but none matching FtsHi subunits. These peptide counts were 429
very low compared to those found in the pdNAD-MDH-YFP IP, which yielded 193 430
and 155 peptides matching Ycf2 and FtsH12, respectively, and many peptides 431
matching FtsHi subunits (Table 3). Thus, none of these NAD-MDH isoforms readily 432
associated with the FtsH12-FtsHi complex. However, in the IPs of the NAD-MDH 433
isoforms, peptides matching pdNAD-MDH were found. This suggests that the NAD-434
MDH isoforms may dimerise with the endogenous pdNAD-MDH protein, since MDHs 435
are known to form dimers (Minárik et al., 2002). 436
437
We conducted a similar experiment with the enzymatically-inactive pdNAD-MDH-438
FlagHA proteins, which complemented the embryo lethality of homozygous pdnad-439
mdh plants. Instead of beads conjugated to an anti-YFP antibody, we used beads 440

15
conjugated to an anti-HA antibody, since the inactive pdNAD-MDH proteins were 441
tagged with a Flag-HA tag. Again, pdnad-mdh plants complemented with PpdNAD-442
MDH:pdNAD-MDH-Flag-HA and wild-type plants served as positive and negative 443
controls, respectively. Peptides matching the Flag-HA-tagged pdNAD-MDH proteins 444
were present with the highest abundance within each IP, indicating that each of the 445
fusion proteins was effectively enriched. All of the components of the FtsH12-FtsHi 446
complex co-purified with the different pdNAD-MDH catalytic mutants, with peptide 447
counts that were similar to those in the IPs with wild-type protein (Table 4). Thus, the 448
point mutations in the catalytic centre did not affect the protein-protein interaction 449
between pdNAD-MDH with the FtsH12-FtsHi complex. 450
451
Mutated Forms of pdNAD-MDH that Cannot Bind NADH Still Complement the 452
Knockout Phenotype 453
454
Our data show that the enzymatic activity of pdNAD-MDH is dispensable, yet the 455
protein has a critical role within the FtsH12-FtsHi complex. Our strategy to abolish 456
the enzymatic activity of pdNAD-MDH disrupted the malate/oxaloacetate binding 457
site. However, it is still possible that the mutants can bind their cofactor NADH and 458
might function as a redox sensor for the AAA-ATPase complex. To investigate this, 459
we performed Isothermal Titration Calorimetry (ITC) with the enzymatically-inactive 460
pdNAD-MDH proteins to test if they were still capable of binding NADH. ITC 461
measures the heat released or taken up as the titrated cofactor (NADH) binds the 462
protein (pdNAD-MDH), depending on whether it is an exothermic or endothermic 463
binding event. We titrated NADH into the wild-type pdNAD-MDH recombinant protein 464
in four independent experiments, each using freshly purified recombinant protein, 465
and observed binding with a mean dissociation constant (Kd) of 7.54 ± 0.53 µM 466
(Figure 10A). Titration of NADH into the R162Q variant resulted in binding with a 467
similar mean Kd (6.75 ± 0.15 µM, from 2 independent experiments; Figure 10B), 468
suggesting that this mutation does not abolish co-factor-binding. However, no NADH 469
binding could be detected for the R234Q variant or for the R162Q R234Q variant 470
(Figure 10C-D). Since these two variants could also complement pdnad-mdh, we 471
can rule out the possibility that co-factor binding of the pdNAD-MDH protein plays a 472
significant role in the FtsH12-FtsHi complex. 473
474

16
DISCUSSION 475
476
pdNAD-MDH is Required for Chloroplast Development 477
478
Our results demonstrate that the pdNAD-MDH protein plays a vital role in plastid 479
development, both during and after embryogenesis in Arabidopsis. During 480
embryogenesis, plastids start to differentiate into chloroplasts in cotyledons at the 481
late globular stage. They expand during the early heart stage, as thylakoids and 482
grana stacks begin to develop (Mansfield and Briarty, 1991). Embryos of the pdnad-483
mdh knockout mutant arrest in the globular-to-heart transition stage (Beeler et al., 484
2014), like many other mutants that are defective in genes essential for chloroplast 485
biogenesis (Yu et al., 2004; Ruppel and Hangarter, 2007; Feng et al., 2014; Lu et al., 486
2014). 487
488
Previously, we studied the viable constitutive silencing line, miR-mdh-1, where 489
pdNAD-MDH levels were reduced, but not abolished, at most or all stages of plant 490
growth. Using the ABI3 promoter to drive embryo-specific expression of pdNAD-491
MDH enabled us to complement the embryo lethality of homozygous pdnad-mdh 492
plants and determine the role of pdNAD-MDH in vegetative growth (Figure 1). These 493
plants were seedling-lethal, with albino leaves containing highly aberrant plastids 494
(Figure 2, 3). This confirms the notion that pdNAD-MDH plays a vital role in post-495
embryogenesis chloroplast development. The phenotype of seedlings resulting from 496
the embryo-specific complementation of pdnad-mdh is more severe than – but 497
consistent with - those observed in miR-mdh-1 plants, which also have pale leaves 498
with aberrant chloroplast ultrastructure. This pale phenotype of miR-mdh-1 results 499
from MDH-deficiency during leaf development rather than from defective 500
embryogenesis. This is further supported by the finding that inducible silencing of 501
pdNAD-MDH at the rosette stage resulted in white/pale newly emerging leaves 502
(Figure 4). 503
504
We propose that pdNAD-MDH is required for the early stages of plastid 505
differentiation. This is most obviously reflected by the absence of internal membrane 506
structure in leaf plastids of embryo-complemented pdnad-mdh and by the reduced 507
thylakoid structure in miR-mdh-1 chloroplasts. However, the aberrant formation of 508

17
internal structure was even observed in etioplasts of miR-mdh-1, the majority of 509
which lacked normal prolamellar bodies (PLBs; Figure 5) and were deficient in many 510
of the precursors required for forming photosynthetic chloroplasts 511
(protochlorophyllide, PORA, galactolipids, and carotenoids; Figure 6). Thus, the 512
presence of pdNAD-MDH is required – either directly or indirectly - for the proper 513
synthesis of several component classes, which serve as the building blocks of 514
thylakoid membranes. Deficiency of these components, caused by other mutations, 515
is known to lead to aberrant PLBs. For example, etioplasts of Arabidopsis carotenoid 516
and chloroplast regulation (ccr) mutants have reduced lutein levels and lack PLBs, 517
suggesting that specific carotenoids are essential for PLB formation (Park et al., 518
2002). PLB formation is absent or aberrant in mutants that cannot make 519
protochlorophyllide (Mascia, 1978; Solymosi and Aronsson, 2013) and in the 520
constitutive photomorphogenic 1 (cop1) mutant (Deng et al., 1991; Lebedev et al., 521
1995), where the lack of PLBs is attributed to a deficiency in PORA or PORB 522
(Sperling et al., 1998). 523
524
Monogalactosyl diacylglycerol (MGDG) and diagalactosyl diacylglycerol (DGDG) are 525
the most abundant lipids in thylakoid membranes in chloroplasts, and MGDG is the 526
most dominant lipid in PLBs. The interaction between PORA and MGDG is thought 527
to stabilize the formation of PLBs (Klement et al., 1999; Engdahl et al., 2001; 528
Selstam et al., 2002). There are conflicting data regarding the effects of suppressing 529
MGDG synthase 1 (MGD1) involved in MGDG synthesis (Jarvis et al., 2000; 530
Kobayashi et al., 2007), possibly resulting from differences between the T-DNA 531
alleles. However, recent findings suggest that MGD1 is involved in the initial step of 532
etioplast development by providing a lipid matrix for protochlorophyllide biosynthesis 533
(Fujii et al., 2017). The silencing of MGD1 decreased MGDG levels in etiolated 534
seedlings, as well as total protochlorophyllide levels. Given that these reports show 535
that the deficiency of one component can cause a pleiotropic decrease in the 536
accumulation of another, the finding that the levels of all major PLB components are 537
reduced in miR-mdh-1 does not allow us to pin down a specific metabolic pathway 538
where pdNAD-MDH is required. 539
540
In seedlings undergoing photomorphogenesis, proplastids develop into chloroplasts, 541
bypassing the etioplast stage, and the extent to which the etioplast is a good model 542

18
for this direct route of chloroplast development is debatable (Solymosi and Schoefs, 543
2010). However, it is important to note that all major PLB components, with the 544
exception of protochlorophyllide and PORA, are also major components of 545
developed chloroplasts, and we observed similar reductions in the levels of these 546
compounds in miR-mdh-1 relative to the wild type in photomorphogenic seedlings. 547
The reduced amounts of these compounds suggest that pdNAD-MDH is required for 548
their proper synthesis in both etioplast-dependent and independent routes of 549
chloroplast development. 550
551
552
Enzymatically-Inactive pdNAD-MDH Complements the pdnad-mdh Mutant 553
554
Unexpectedly, we discovered that the embryo-lethal phenotype of the pdnad-mdh 555
mutant is not caused by the loss of plastidial NAD-MDH activity. On one hand, three 556
different mutated forms of the enzyme that were enzymatically-inactive and, in two 557
cases, that additionally could not bind NADH, could complement the embryo-lethal 558
phenotype when expressed in pdnad-mdh (Figure 8, 10). The complemented plants 559
grew normally and were not pale or albino, as were miR-mdh-1 or embryo-560
complemented pdnad-mdh plants. On the other hand, expressing other NAD-MDH 561
isoforms in chloroplasts failed to complement the pdnad-mdh phenotype (Figure 7). 562
These observations suggest that the phenotypes of pdnad-mdh plants are 563
specifically caused by the absence of the pdNAD-MDH protein itself (Figure 8). 564
565
Our data do not rule out the previously proposed role of pdNAD-MDH in balancing 566
redox equivalents via its enzymatic interconversion of malate and oxaloacetate (i.e. 567
the malate valve model; Scheibe, 2004), but we argue that this is not the essential 568
function of the protein. Nevertheless, it is important to note that the pdNAD-MDH 569
protein has MDH activity and that the catalytic residues are conserved among 570
orthologous proteins in other plants (Supplemental Figure 7). Thus, it is likely that the 571
enzymatic activity is important under specific conditions or tissues, such as in pollen 572
tubes during expansion (Selinski et al., 2014). Indeed, in a recent genome-wide 573
association study, pdNAD-MDH was mapped as a quantitative trait locus for malate 574
levels in Arabidopsis (Fusari et al., 2017). Furthermore, mutations in pdNAD-MDH 575
rescued the phenotype of the mosaic death 1 (mod1) mutant, which accumulates 576

19
reactive oxygen species (ROS) and shows abnormal patterns of programmed cell 577
death (PCD) (Zhao et al., 2018). Interestingly, the mod1 mutant is proposed to 578
generate ROS in the mitochondria in response to a signal from the chloroplast. The 579
fact that mMDH1 mutations could also suppress the mod1 phenotype suggests that 580
a malate valve may facilitate the communication between mitochondria and 581
chloroplasts (Zhao et al., 2018). These findings are consistent with our results, as 582
they suggest that the malate valve is not essential. 583
584
Interestingly, we found slightly fewer than the expected 25% of pdnad-mdh plants in 585
the T2 generation (Figure 8C), particularly for the construct that encoded proteins 586
with two amino acid substitutions. This may reflect incomplete complementation by 587
these non-enzymatic proteoforms at certain developmental stage, either because 588
MDH activity is beneficial or because the introduction of amino-acid substitutions in 589
the catalytic centre may affect the integrity or half-life of the protein itself. Further 590
work will be required to assess the importance of NAD-MDH activity in the 591
chloroplast and the extent to which its role can be compensated by the presence of 592
NADP-MDH. As the loss of NADP-MDH activity alone also has a relatively mild 593
impact on plant growth (Hebbelmann et al., 2012), generating lines expressing the 594
inactive pdNAD-MDH constructs in the pdnad-mdh nadp-mdh double mutant 595
background would be highly valuable for reassessing the importance of MDH activity 596
and the malate valve in the chloroplast. 597
598
pdNAD-MDH Functions in Complex with AAA-Proteases at the Chloroplast 599
Inner Envelope 600
601
We demonstrate that pdNAD-MDH interacts with members of a proposed large AAA 602
protease complex localised to the chloroplast inner envelope composed of FtsH12 603
and FtsHi subunits, as well as with Ycf2 (Table 1, 2; Figure 11). Proteomics data 604
indicate that most of the pdNAD-MDH protein is localized to the stroma and 605
chloroplast envelope, and only a minority of the protein associated to the thylakoid 606
membranes (Ferro et al., 2010), suggesting that a fraction of total pdNAD-MDH is 607
stably involved in this interaction. Further, Cvetić et al. (2008) identified pdNAD-MDH 608
in both the stromal and chloroplast envelope fractions of spinach leaf chloroplasts. 609
Plants lacking FtsH12 (emb1047, emb156), FtsHi2 (emb2083), FtsHi4 (emb3144) or 610

20
FtsHi5 (emb2458) are all embryo-lethal (Patton et al., 1991; Franzmann et al., 1995; 611
Patton et al., 1998; Sokolenko et al., 2002; Wagner et al., 2012; Lu et al., 2014). 612
Although no Arabidopsis mutant for the chloroplast genome-encoded Ycf2 is 613
currently available, a knockout mutant of the gene could not be generated using 614
plastome transformation in tobacco, suggesting that it is also essential (Drescher et 615
al., 2000). We verified that FtsH12 knockout plants were embryo-lethal and arrested 616
near the globular-to-heart transition stage, similar to pdnad-mdh. Furthermore, 617
Arabidopsis lines with constitutive silencing of FtsH12 expression had a striking 618
resemblance to the miR-mdh-1 line. Thus, the phenotypes observed from the loss of 619
pdNAD-MDH could be explained by the loss of FtsH12 function. Interestingly, in miR-620
mdh-1, FtsH12 protein levels were strongly reduced compared to wild type, whereas 621
the converse was not true; in the amiRNA FtsH12 lines, pdNAD-MDH protein levels 622
were similar to the wild type (Figure 9C). Taken together, these data support a 623
hypothesis where pdNAD-MDH plays a role in stabilising FtsH12, and possibly the 624
entire complex. The exact mechanism by which pdNAD-MDH stabilises FtsH12 is 625
still unknown, but it does not require pdNAD-MDH catalytic activity or NADH-binding, 626
as the inactive pdNAD-MDH proteins could still interact with FtsH12-FtsHi complex 627
members in plants (Table 4). However, other MDH isoforms from different cell 628
compartments could not interact with FtsH12, and no other MDH isoform was 629
identified in the immunoprecipitation experiment with FtsH12, suggesting that the 630
interaction is specific to pdNAD-MDH in chloroplasts (Table 2, 3). 631
632
Possible Role of the FtsH12-FtsHi Complex 633
634
AAA-type proteins generally form hexamers, where each subunit has an N-terminal 635
transmembrane segment and a C-terminal AAA-ATPase domain expanded in the 636
stroma. The FtsH12-FtsHi complex contains several members of the FtsHi family 637
(FtsHi1, FtsHi2, FtsHi4 and FtsHi5), which lack the zinc-binding motif that is 638
considered essential for its metalloprotease activity. In a co-expression network, all 639
members of the FtsH12-FtsHi complex clustered together with genes involved in 640
plastid translation, division and positioning, as well as amino acid metabolism 641
(Majsec et al., 2017). Non-proteolytic FtsHi proteins were reported to be absent from 642
the cyanobacterium Synechocystis and are thought to have evolved at a later stage 643
of evolution through gene duplication (Sokolenko et al., 2002). However, the activity 644

21
of the AAA protease complex does not require all six subunits to be active (Martin et 645
al., 2005). AAA-type proteases are also known to generate a pulling force. The 646
mechanism of target protein unfolding by AAA-type proteases includes a 647
conformational change in the AAA domain, which moves conserved, substrate-648
binding hydrophobic residues towards the inner pore of the hexameric complex. This 649
draws the substrate proteins inside the pore and unfolds them (Lee et al., 2001; 650
Langklotz et al., 2012). In mitochondria, the m-AAA protease, described as an ATP-651
dependent protease that degrades misfolded proteins and mediates protein 652
processing, is proposed to be further involved in the dislocation of imported 653
preproteins from the inner membrane by functioning as an ATP-driven molecular 654
motor (Tatsuta et al., 2007; Botelho Calado et al., 2013). In yeast mitochondria, 655
many nucleus-encoded preproteins are imported into the mitochondrial matrix via the 656
TIM23 translocon (Demishtein-Zohary and Azem, 2017). The FtsH12-FtsHi complex 657
could potentially play a similar role in chloroplasts. 658
659
Nakai (2018) recently proposed a novel ATP-driven import motor associated with the 660
TIC (translocon on the inner chloroplast membrane) complex at the inner chloroplast 661
envelope membrane. It is likely that the FtsH12-FtsHi complex, together with 662
pdNAD-MDH, is the proposed ATP-driven motor that imports pre-proteins across the 663
chloroplast envelopes. Consistent with this hypothesis, we also found large numbers 664
of peptides matching components of the chloroplast protein import machinery (e.g. 665
Ycf1 and Toc159) in our IP with FtsH12 (Table 2). However, further investigations 666
are needed to determine whether the function of FtsH12 in chloroplast development 667
is dependent on its proteolytic activity or solely on its ATPase activity. Further 668
studies will provide exciting new insights into the role of the FtsH12-FtsHi complex in 669
chloroplast development and function. 670
671
In conclusion, our data define pdNAD-MDH as a moonlighting protein essential for 672
chloroplast development. Moonlighting enzymes perform more than one function, 673
often serving a structural or regulatory function in addition to their known catalytic 674
function (Jeffrey, 1999, 2003; Copley, 2003; Moore, 2004). Moonlighting functions 675
occur frequently in highly conserved proteins and are thought to evolve more 676
commonly for soluble, highly abundant proteins that are constitutively expressed 677
(Huberts and van der Klei, 2010; Copley, 2015). Soluble abundant proteins are likely 678

22
to encounter many more biomolecular interactions within their environment, and 679
advantageous interactions can evolve over time (Copley, 2015). It has also been 680
proposed that acquiring a moonlighting function might be an easier way to expand 681
the functional tool box of an organism without the drawbacks resulting from an 682
expanding genome (Jeffery, 1999). Interestingly, lactate dehydrogenase, a homolog 683
of the MDH family, also plays a moonlighting role as a structural protein in the lenses 684
of bird eyes, and was one of the first examples of moonlighting proteins described 685
(Wistow et al., 1987; Hendriks et al., 1988; Huberts and van der Klei, 2010). A well-686
known example of a moonlighting enzyme in plant metabolism is hexokinase, which 687
is a key enzyme in central metabolism but also acts as a sugar sensor (Jang, 1997; 688
Moore, 2003). Also, Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) 689
isoforms, aside from their roles in glycolysis and the Calvin-Benson-Bassham cycle, 690
can regulate DNA stability, control gene expression, function in apoptosis, and act as 691
redox sensors (Zaffagnini et al., 2013; He et al., 2013; Yang and Zhai, 2017). 692
Recently, AROGENATE DEHYDRATASE2 (ADT2) was shown to localise to the 693
chloroplast division machinery in Arabidopsis, suggesting an additional non-694
enzymatic function besides its enzymatic role in phenylalanine biosynthesis (Bross et 695
al., 2017). However, pdNAD-MDH appears thus far to be a unique example in which 696
the moonlighting function is essential for plant survival. 697
698
METHODS 699
700
Plant Growth 701
Arabidopsis thaliana plants were grown in soil in growth cabinets (Percival AR-95 702
[CLF Plant Climatics]; OR Kälte 3000) fitted with fluorescent lamps and 703
supplemented with red LED panels. Unless stated otherwise, the chambers provided 704
a 12-h light/12-h dark cycle, with light intensity of 150 μmol photons m−2 s−1, 705
temperature of 20°C, and relative humidity of 65%. 706
707
For experiments with plate-grown seedlings, seeds were surface sterilized and 708
placed onto ½-strength Murashige and Skoog (MS) medium with vitamins and MES 709
(Duchefa Biochemie BV) at pH 5.8, solidified with 0.8 % (w/v) agar. The seeds were 710
stratified by incubating the plates in the dark at 4°C for two days. For etiolated 711
seedlings, germination was stimulated by placing the seeds on plates under white 712

23
light (150 μmol photons m−2 s−1) for 4-6 h, followed by growth for 5-6 days in the 713
dark. Light-grown seedlings were placed in either standard growth conditions (as 714
above for plants grown in soil) or in continuous light. For seedlings for lipid, 715
carotenoid and protochlorophyllide/PORA analysis, seeds were germinated on a 100 716
μm nylon mesh placed on solid medium to aid harvest. Dark-grown seedlings were 717
harvested under low green light illumination. 718
719
To select transformants on plates, the growth medium contained either 15 mg/L 720
BASTA, 15 mg/L Hygromycin or 50 mg/L Kanamycin, depending on the resistance 721
marker on the transgene. pdnad-mdh plants transformed with the constructs 722
encoding inactive pdNAD-MDH (R162Q, R234Q and R162Q R234Q) were selected 723
on soil by spraying them with BASTA (final concentration of 0.018% (w/v) glufosinate 724
(Omya). 725
726
The pdnad-mdh T-DNA insertion mutant (ET8629) and constitutive pdNAD-MDH 727
silencing line, miR-mdh-1, were characterized in (Beeler et al., 2014). The ftsh12 T-728
DNA lines (emb1047-1, emb1047-2, ftsh12 1-1, emb156-1) were ordered from the 729
Nottingham Arabidopsis Stock Centre (NASC). The heterozygous pdnad-mdh 730
mutant is in the Landsberg erecta (Ler) background, while miR-mdh-1 and ftsh12 T-731
DNA insertion lines are in the Columbia (Col-0) background. 732
733
Recombinant Protein Expression in Escherichia coli 734
735
All sequences of oligonucleotide primers used for cloning the following constructs 736
are listed in Supplemental Table 2. For the recombinant expression of pdNAD-MDH-737
His and its catalytic inactive variants in E. coli, pdNAD-MDH was cloned into the 738
pET21a+ expression vector. First, the length of the chloroplast transit peptide (cTP) 739
was predicted using the Target P server (http://www.cbs.dtu.dk/services/TargetP/). 740
The full-length coding sequence of pdNAD-MDH without the cTP was amplified with 741
NdeI and NotI restriction sites, using the pdNAD-MDH:pDONR221 vector as a 742
template (Beeler et al., 2014). The PCR product was cloned into the pET21a+ vector 743
(Novagen) using the restriction sites. The point mutations in the catalytic centre of 744
pdNAD-MDH were generated in the pdNAD-MDH:pDONR221 vector using a 745
QuikChange Site-Directed Mutagenesis Kit (Agilent Technologies) according to the 746

24
manufacturer’s instructions. The mutated coding sequences were then cloned into 747
pET21a+ as described for the wild-type sequence. 748
749
For protein expression, the vectors were transformed into E. coli BL21 (DE3) 750
CodonPlus cells (Agilent Technologies). The cells were cultured in LB medium at 751
37°C until OD600 of 0.5-0.7. Protein expression was induced by adding 1 mM 752
Isopropyl β-D-1-thiogalactopyranoside (IPTG), and the cultures were incubated 753
overnight at 20°C. Cell lysis and purification of the His-tagged proteins were carried 754
out as described in (Seung et al., 2013). 755
756
NAD-MDH Enzyme Activity Measurements 757
758
Enzyme activity of the pdNAD-MDH recombinant proteins was measured 759
spectrophotometrically in an infinite® M1000 Pro plate reader (Tecan). Each reaction 760
contained 0.092 M Tris-HCl, pH 7.9, 0.01 M MgCl2, 0.2 mM NADH, and 0.01 μg 761
recombinant protein. The baseline rate at 340 nm was acquired for 5 min at 20°C, 762
measuring absorbance every 20 s. The reaction was started by adding 0.09 mM 763
oxaloacetate. The linear decline in absorbance at 340 nm was monitored and used 764
to calculate the rate of NADH consumption. Activity was determined three times on 765
the same protein preparation to calculate the mean ± SE. 766
767
Immunoblotting and Native PAGE 768
769
To extract soluble proteins, young leaves were homogenised in protein extraction 770
medium (100 mM 3-[N-Morpholino]-2-hydroxypropanesulfonic acid [MOPS, pH 7.2], 771
1 mM EDTA, 10% (v/v) ethylene glycol, 2 mM dithiothreitol, 1 x Complete Protease 772
Inhibitor cocktail [Roche]). Insoluble material was pelleted at 20,000 g. Protein 773
content in the supernatant was determined using the Bradford assay. To extract total 774
protein from leaves (Figure 4), frozen 7-mm leaf disc were ground with 2-3 glass 775
beads in a mixer mill (Retsch MM 200). The powder was suspended in SDS-PAGE 776
loading buffer (50mM Tris-HCl, pH 6.8, 100 mM DTT, 2% [w/v] SDS, 30% [v/v] 777
glycerol, 0.005% [w/v] bromophenol blue) and heated to 95°C for 5 min. Insoluble 778
material was removed via centrifugation, and 5 µl samples were loaded onto SDS-779
PAGE gels. To extract total protein from seedlings (Figure 6), frozen seedling 780

25
samples were extracted with SDS-PAGE loading buffer (at 100 mg/mL) and 10 µl 781
samples were loaded onto SDS-PAGE gels. 782
783
For immunodetection of pdNAD-MDH, we used rabbit antisera raised against the 784
Arabidopsis pdNAD-MDH protein (Beeler et al., 2014). For FtsH12, we used rabbit 785
antisera raised against the Arabidopsis FtsH12 protein, which was a gift from Dr. 786
Masato Nakai (Osaka University). For the detection of epitope-tagged proteins, we 787
used α-GFP/YFP (ab290, Abcam), α-HA (ab9110, Abcam), α-Flag M2 (F1804, 788
Sigma), and α-PORA (Agrisera). Proteins were detected based on infrared 789
fluorescence using IR800-conjugated secondary antibodies and an Odyssey CLx 790
detection system (Li-cor). Where actin was detected as a loading control, the rabbit 791
primary antibody was co-incubated with a mouse monoclonal antibody against plant 792
actin (A0480, Sigma), which was detected in the same blot using a 680RD-793
conjugated anti-mouse secondary antibody. Primary antibody dilutions are as 794
follows: α-pdNAD-MDH - 1:2,000, α-FtsH12 - 1:1000, α-GFP/YFP - 1:10,000, α-HA - 795
1:7,000, α-Flag M2 - 1:5,000, α-PORA – 1:2,000, α-actin - 1:10,000. 796
797
Native PAGE observation of NAD-MDH activity was performed as described (Beeler 798
et al., 2014). 799
800
Bioinformatic Analyses 801
802
For amino acid sequence alignments, all Arabidopsis MDH isoform sequences were 803
retrieved from TAIR, while pdNAD-MDH sequences from different plant species were 804
retrieved from the Phytozome v12 database 805
(https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3245001/). Sequences were aligned 806
using Clustal Omega. 807
808
For homology-based modelling of the pdNAD-MDH protein structure, the pdNAD-809
MDH protein sequence without the cTP was modelled using SWISS-MODEL 810
(https://www.ncbi.nlm.nih.gov/pubmed/24782522) onto the human MMDH2 structure 811
as a template (PDB: 2DFD), which was co-crystallized with malate and NAD. 812
813

26
Cloning of Expression Vectors for Plant Transformation 814
815
Constructs for Arabidopsis transformation were assembled using Gateway 816
technology (Invitrogen). For multi-site Gateway assembly, we prepared the promoter, 817
coding sequence or tag in appropriate pENTR or pDONR vectors. The ABI3 818
promoter (2176 bp) was amplified from Arabidopsis genomic DNA, flanked with 819
HindIII restriction sites, and ligated into the pENTR vector between the attL4 and 820
attR1 recombination sites at the 5’ and 3’ ends, respectively. The pdNAD-MDH 821
promoter in pDONR P4-P1r and the pdNAD-MDH coding sequence in pDONR221 822
were described previously. The coding sequences of the various NAD-MDH isoforms 823
fused to the chloroplast transit peptide of the Rubisco small subunit were 824
synthesized by Biomatik, flanked by attB1 and atttB2 recombination sites. These 825
sequences were recombined directly into the pDONR221 vector. The eYFP and 826
Flag-HA tags were cloned into pDONR P2R-P3 (Beeler et al., 2014; Tschopp et al., 827
2017). The appropriate promoter, coding sequence, and C-terminal tag were 828
recombined via an LR Clonase reaction into the multisite Gateway binary vector 829
pB7m34GW,0. 830
831
To generate overexpressor lines of pdNAD-MDH and FtsH12, the pDONR221 vector 832
containing the corresponding coding sequence was recombined in a LR Clonase 833
reaction into the single Gateway binary vector pB7YWG2,0 (with a 35S promoter and 834
C-terminal YFP tag, Karimi et al., 2002) and pUBC-YFP (Grefen et al., 2010), 835
respectively. 836
837
To generate artificial microRNA silencing constructs, we recombined the amiRNA 838
miR-mdh-1 cassette in pDONR221 (described previously; Beeler et al., 2014) into a 839
vector containing an estradiol inducible cassette (XVE) driven by the UBIQUITIN10 840
promoter, via the LR Clonase reaction (Lee et al., 2013; Gujas et al., 2017). Two 841
artificial microRNA silencing cassettes for FtsH12 were designed and cloned using 842
the Web MicroRNA Designer tool (WMD3, wmd3.weigelworld.org; Ossowski et al., 843
(2008) according to the provided instructions. The primers used are provided in 844
Supplemental Table 2., The assembled amiRNAs were flanked with attB1 and attB2 845
recombination sites using the ‘attB1-amiRNA FW’ and ‘attB2-amiRNA REV’ primers 846
(Supplemental Table 2) and recombined in pDONR221 vectors using the BP 847

27
Clonase reaction. Instead of the pART27 vector, we further recombined the insert 848
into the binary pJCV52 vector via an LR Clonase reaction. 849
850
To generate stable Arabidopsis lines, the appropriate constructs were transformed 851
into Agrobacterium tumefaciens (strain GV3101). Wild-type Columbia and 852
heterozygous pdnad-mdh plants were transformed using the floral dip method as 853
described previously (Zhang et al., 2006). The T1 generation of transformants were 854
selected using the BASTA or Hygromycin selection marker (as described above for 855
plant growth). Up to 10 seedlings per line were screened for transgene expression 856
using immunoblotting (using the antibody against the epitope tag) and genotyped for 857
the pdnad-mdh T-DNA insertion in the case of transformed pdnad-mdh plants. For 858
plants expressing the transgene, the number of T-DNA insertion loci was determined 859
by segregation analysis of the resistance marker gene in the T2 generation. Two 860
independent transformants with single insertions, originating from different T0 861
parents, were selected for further analysis - either in the T2 generation (BASTA 862
resistant plants that are heterozygous or homozygous for the transgene) or T3 863
generation (plants that are homozygous for the transgene), as specified. 864
865
Transient expression in Nicotiana benthamiana leaves was performed according to 866
Seung et al. (2015). 867
868
Immunoprecipitation Experiments 869
870
Proteins were extracted from leaf tissue of Arabidopsis plants expressing the epitope 871
tagged proteins by homogenising in immunoprecipitation medium (50 mM Tris-HCl, 872
pH 8.0, 150 mM NaCl, 1% [v/v] Triton X-100, 1 mM DTT and Complete Protease 873
Inhibitor cocktail [Roche]). Insoluble material was removed by centrifugation. The 874
supernatant was incubated for 1 h at 4°C with µMACS magnetic beads conjugated to 875
α-YFP or α-HA (Miltenyi Biotec). After incubation, the beads were recovered using a 876
µColumn (Miltenyi Biotec) on a magnetic stand. The beads were washed five times 877
with immunoprecipitation medium before eluting the bound proteins with elution 878
buffer (50 mM Tris-HCl, pH 7.5, 2% [w/v] SDS). The proteins were precipitated by 879
the addition of 10% (w/v) TCA. After two washes with cold acetone, the protein pellet 880
was dissolved in 10 mM Tris, pH 8.2, 2 mM CaCl2 and digested with trypsin for 30 881

28
min at 60°C. The resulting peptides were dried, dissolved in 0.1% (v/v) formic acid, 882
and analysed by LC-MS/MS on a nanoAcquity UPLC (Waters Inc.) connected to a Q 883
Exactive mass spectrometer (Thermo Scientific) equipped with a Digital PicoView 884
source for electrospray ionisation (New Objective). Peptides were trapped on a 885
Symmetry C18 trap column (5 µm, 180 µm x 20 mm, Waters Inc.) and separated on 886
a BEH300 C18 column (1.7 µm, 75 µm x 150 m, Waters Inc.) at a flow rate of 250 887
nl/min using a gradient from 1% solvent B (0.1% formic acid in acetonitrile, 888
Romil)/99% solvent A (0.1% formic acid in water, Romil) to 40% solvent B/60% 889
solvent A within 90 min. The mass spectrometer settings were as follows: Data 890
dependent analysis; Precursor scan range 350 – 1500 m/z, resolution 70,000, 891
maximum injection time 100 ms, threshold 3e6; Fragment ion scan range 200 – 2000 892
m/z, Resolution 35,000, maximum injection time 120 ms, threshold 1e5. Proteins 893
were identified using the Mascot search engine (Matrix Science, version 2.4.1) 894
against the TAIR10 Arabidopsis proteome database, with fragment ion mass 895
tolerance of 0.030 Da, parent ion tolerance of 10.0 PPM, and oxidation of methionine 896
was specified in Mascot as a variable modification. Scaffold (Proteome Software 897
Inc.) was used to validate MS/MS based peptide and protein identifications. Peptide 898
identifications were accepted if they achieved a false discovery rate (FDR) of less 899
than 0.1% by the Scaffold Local FDR algorithm. Protein identifications were 900
accepted if they achieved an FDR of less than 1.0% and contained at least 2 901
identified peptides. 902
903
904
Light and Electron Microscopy 905
906
Transmission electron microscopy analyses were conducted as previously described 907
(Beeler et al., 2014). For DIC images of embryos, seeds were cleared with a 908
glycerin:chloral hydrate:water (1:8:3, w/w/w) solution for 2-6 h, depending on the 909
developmental stage of the seeds (Breuninger et al., 2008). Embryos were imaged 910
under an Axio Imager 2 microscope with ZEN pro 2011 software (Zeiss). For imaging 911
of YFP in leaf tissue, confocal laser scanning microscopy was conducted as 912
previously described (Beeler et al., 2014). 913
914
915
Isothermal Titration Calorimetry 916
917

29
Binding of NADH to recombinant pdNAD-MDH proteins (WT, R162Q, R234Q, 918
R162Q R234Q) was measured with a VP-ITC instrument at 25°C. 40 injections of 919
cofactor solution (400 µM NADH), each with a volume of 6 µL, were made into a 920
recombinant protein solution (40 µM). An injection spacing of 300 s was selected, 921
and duration was automatically calculated. An initial injection of 2 µL was excluded 922
from data analysis. Default sample cell volume was 1.4644 mL. Stirring speed and 923
reference power were set to 307 RPM and 10 µCal.sec-1, respectively. Before 924
measurement, samples were degassed for 5 min at 23°C. The proteins were 925
desalted into ITC buffer (20 mM Tris-HCl, pH 7.5, 10 mM NaCl) with and without 2.5 926
mM 2-Mercaptoethanol (Sigma) using Illustra NAP-5 (Fisher Scientific AG) prior to 927
the experiment. The ligand tested was NADH (Grade I, Roche) dissolved directly into 928
the ITC buffer. Concentrations of the proteins and NADH were determined 929
spectrophotometrically using following extinction coefficients (Ɛ340nm [NADH] = 930
6220 M-1 cm-1; Ɛ280nm [pdNAD-MDH] = 8940 M-1 cm-1). 931
932
933
Lipid and Carotenoid Measurements 934
935
For targeted lipid profiling, 6-day-old light or dark-grown seedlings were ground into 936
a fine powder in liquid nitrogen using a mortar and pestle. Ground plant material (30-937
60 mg, precisely weighed for each sample) was extracted with 0.4 mL 938
tetrahydrofuran/methanol (50:50, v/v) including 5 µg/mL hydrogenated MDGD 939
(Matreya) as an internal standard. 10-15 glass beads were added and the samples 940
were homogenized for 3 min at 30 Hz in a tissue lyser (Qiagen). Insoluble material 941
was removed by two-rounds of centrifugation at 16,000 g for 3 min. 200 µL of 942
supernatant was transferred to an appropriate glass vial for immediate UHPLC-943
QTOF-MS analysis (Waters) as previously described (Martinis et al., 2011; Kessler 944
and Glauser, 2014). Raw data were processed using MassLynx version 4.1 (waters) 945
for automatic peak analysis. Identification and relative quantification were performed 946
as described in Spicher et al., 2017. 947
Protochlorophyllide Measurements 948
Six-day-old etiolated seedlings were harvested and ground into fine powder in liquid 949
nitrogen using a mortar and pestle. Protochlorophyllide was extracted twice in 9:1 950

30
mix (v/v) of acetone:0.1 M NH4OH, and insoluble material was removed by pelleting 951
at 13,000 g for 10 min after each extraction (Cheminant et al., 2011). The two 952
extractions were combined, and emission spectra were recorded from 500-750 nm 953
following excitation at 440 nm using a fluorescence spectrophotometer (Tecan 954
infinite M1000). Protochlorophyllide was quantified at the emission peak of 632 nm, 955
and expressed as fluorescence per mg of fresh weight. 956
Accession Numbers 957
Sequence data from this article can be found in TAIR (www.arabidopsis.org) under 958
the following accession numbers: pdNAD-MDH (At3g47520), FtsH12 (At1g79560), 959
FtsHi1 (At4g23940), FtsHi2 (At3g16290), FtsHi4 (At5g64580), FtsHi5 (At3g04340), 960
and YCF2 (AtCg00860). TAIR accession numbers of potential interaction partners 961
are provided in Tables 1–4. 962
Supplemental Data 963
Supplemental Figure 1 Exogenous supply of sucrose did not rescue pdnad-mdh 964
seedlings. 965
Supplemental Figure 2 Examples of normal, compromised, and absent prolamellar 966
body structures in Col and miR-mdh-1, observed via transmission electron 967
microscopy. 968
Supplemental Figure 3 Multiple protein sequence alignment of all nine Arabidopsis 969
thaliana malate dehydrogenase isoforms. 970
Supplemental Figure 4 Native-PAGE gel analysis of heterozygous pdnad-mdh 971
plants expressing various NAD-MDH isoforms. 972
Supplemental Figure 5 FtsH12 localisation and embryo lethality of the ftsh12 973
mutant. 974
Supplemental Figure 6 Quantification of FtsH12 in miR-mdh-1. 975
976
Supplemental Figure 7 Sequence alignment of pdNAD-MDH isoforms in various 977
plant species, algae and E. coli. 978
Supplemental Table 1 Total set of lipids measured in 6-day-old wild-type (Col) and 979
miR-mdh-1 seedlings. 980
Supplemental Table 2 Oligonucleotide primers used in this study. 981

31
Supplemental Data Set 1 Complete list of proteins identified in immunoprecipitates 982
by MS/MS. 983
984
ACKNOWLEDGMENTS 985
986
This work was funded by the Swiss National Foundation (SNF) (Grant Number 987
31003A_156987 to O.K. and S.C.Z.) and by ETH Zurich. We thank Claudia Di 988
Césaré from the Neuchâtel Platform of Analytical Chemistry for technical assistance, 989
as well as Peter Hunziker, Yolanda Joho-Auchli and Simone Wüthrich from the 990
Functional Genomic Centre Zurich (FGCZ) for mass spectrometry analysis. We 991
thank Masato Nakai for sharing his unpublished work and his thoughts with us, 992
Enrico Martinoia and Alisdair Fernie for fruitful discussions during the course of this 993
work and Andrea Ruckle for help in plant culture. 994
995
AUTHOR CONTRIBUTIONS 996
997
T.B.S., S.C.Z., and O.K. conceived and directed the research; T.B.S., A.C., S.C.Z., 998
and O.K. designed the experiments; T.B.S., A.C., E.D., M.S., F.G., M.S., F.K., S.H., 999
D.A., and B.A.M. performed research and analysed data; T.B.S. and S.C.Z. wrote1000
the manuscript with input from all of the authors. 1001
1002
REFERENCES 1003 1004
Bahl, J., Francke, B., and Monéger, R. (1976). Lipid composition of envelopes, prolamellar bodies 1005 and other plastid membranes in etiolated, green and greening wheat leaves. Planta 129: 193–1006 201. 1007
Beeler, S., Liu, H.-C., Stadler, M., Schreier, T., Eicke, S., Lue, W.-L., Truernit, E., Zeeman, S.C., 1008 Chen, J., and Kotting, O. (2014). Plastidial NAD-Dependent Malate Dehydrogenase Is Critical 1009 for Embryo Development and Heterotrophic Metabolism in Arabidopsis. Plant Physiol. 164: 1010 1175–1190. 1011
Berkemeyer, M., Scheibe, R., and Ocheretina, O. (1998). A novel, non-redox-regulated NAD-1012 dependent malate dehydrogenase from chloroplasts of Arabidopsis thaliana L. J. Biol. Chem. 1013 273: 27927–27933. 1014
Birktoft, J.J. and Banaszak, L.J. (1983). The presence of a histidine-aspartic acid pair in the active 1015 site of 2-hydroxyacid dehydrogenases. X-ray refinement of cytoplasmic malate dehydrogenase. 1016 J. Biol. Chem. 258: 472–482.1017
Bodi, Z., Zhong, S., Mehra, S., Song, J., Graham, N., Li, H., May, S., and Fray, R.G. (2012). 1018 Adenosine Methylation in Arabidopsis mRNA is Associated with the 3′ End and Reduced Levels 1019 Cause Developmental Defects. Front. Plant Sci. 3: 1–10. 1020
Botelho Calado, S., Tatsuta, T., Von Heijne, G., and Kim, H. (2013). Dislocation by the m-AAA 1021 protease increases the threshold hydrophobicity for retention of transmembrane helices in the 1022 inner membrane of yeast mitochondria. J. Biol. Chem. 288: 4792–4798. 1023
Breuninger, H., Rikirsch, E., Hermann, M., Ueda, M., and Laux, T. (2008). Differential Expression 1024 of WOX Genes Mediates Apical-Basal Axis Formation in the Arabidopsis Embryo. Dev. Cell 14: 1025

32
867–876. 1026 Bross, C.D., Howes, T.R., Rad, S.A., Kljakic, O., and Kohalmi, S.E. (2017). Subcellular localization 1027
of Arabidopsis arogenate dehydratases suggests novel and non-enzymatic roles. J. Exp. Bot. 1028 68: 1425–1440. 1029
Candela, H., Pérez-Pérez, J.M., and Micol, J.L. (2011). Uncovering the post-embryonic functions of 1030 gametophytic- and embryonic-lethal genes. Trends Plant Sci. 16: 336–345. 1031
Cheminant, S., Wild, M., Bouvier, F., Pelletier, S., Renou, J.-P., Erhardt, M., Hayes, S., Terry, 1032 M.J., Genschik, P., and Achard, P. (2011). DELLAs Regulate Chlorophyll and Carotenoid 1033 Biosynthesis to Prevent Photooxidative Damage during Seedling Deetiolation in Arabidopsis. 1034 Plant Cell 23: 1849–1860. 1035
Clarke, A.R., Wigley, D.B., Chia, W.N., Barstow, D., Atkinson, T., and Holbrook, J.J. (1986). Site-1036 directed mutagenesis reveals role of mobile arginine residue in lactate dehydrogenase catalysis. 1037 Nature 324: 699–702. 1038
Comparot-Moss, S., Kötting, O., Stettler, M., Edner, C., Graf, A., Weise, S.E., Streb, S., Lue, W.-1039 L., MacLean, D., Mahlow, S., Ritte, G., Steup, M., Chen, J., Zeeman, S.C., and Smith A.M. 1040 (2010). A Putative Phosphatase, LSF1, Is Required for Normal Starch Turnover in Arabidopsis 1041 Leaves. Plant Physiol. 152: 685–697. 1042
Copley, S.D. (2015). An Evolutionary Perspective on Protein Moonlighting. Biochem Soc Trans. 42: 1043 1684–1691. 1044
Copley, S.D. (2003). Enzymes with extra talents: Moonlighting functions and catalytic promiscuity. 1045 Curr. Opin. Chem. Biol. 7: 265–272. 1046
Cvetić, T., Veljović-Jovanović, S., and Vučinić, Ž (2008) Characterization of NAD-dependent 1047 malate dehydrogenases from spinach leaves, Protoplasma 232:247-253. 1048
Demishtein-Zohary, K. and Azem, A. (2017). The TIM23 mitochondrial protein import complex: 1049 function and dysfunction. Cell Tissue Res. 367: 33–41. 1050
Deng, X.W., Caspar, T., and Quail, P.H. (1991). Cop1 : a regulatory locus involved m hght-controlled 1051 development and gene expression in Arabidopsis. Genes Dev. 5: 1172–1182. 1052
Despres, B., Delseny, M., and Devic, M. (2001). Partial complementation of embryo defective 1053 mutations: A general strategy to elucidate gene function. Plant J. 27: 149–159. 1054
Drescher, A., Stephanie, R., Calsa, T., Carrer, H., and Bock, R. (2000). The two largest chloroplast 1055 genome-encoded open reading frames of higher plants are essential genes. Plant J. 22: 97–1056 104. 1057
Engdahl, S., Aronsson, H., Sundqvist, C., Timko, M.P., and Dahlin, C. (2001). Association of the 1058 NADPH:protochlorophyllide oxidoreductase (POR) with isolated etioplast inner membranes from 1059 wheat. Plant J. 27: 297–304. 1060
Emanuelsson, O., Nielsen, H., Brunak, S., and Von Heijne, G. (2000). Predicting subcellular 1061 localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 300: 1005–1062 1016. 1063
Feng, J., Fan, P., Jiang, P., Lv, S., Chen, X., and Li, Y. (2014). Chloroplast-targeted Hsp90 plays 1064 essential roles in plastid development and embryogenesis in Arabidopsis possibly linking with 1065 VIPP1. Physiol. Plant. 150: 292–307. 1066
Ferro, M. et al. (2010). AT_CHLORO, a Comprehensive Chloroplast Proteome Database with 1067 Subplastidial Localization and Curated Information on Envelope Proteins. Mol. Cell. Proteomics 1068 9: 1063–1084. 1069
Franzmann, L.H., Yoon, E.S., and Meinke, D.W. (1995). Saturating the Genetic-Map of Arabidopsis 1070 thaliana with Embryonic Mutations. Plant J. 7: 341–350. 1071
Fujii, S., Kobayashi, K., Nagata, N., Masuda, T., and Wada, H. (2017). 1072 Monogalactosyldiacylglycerol Facilitates Synthesis of Photoactive Protochlorophyllide in 1073 Etioplasts. Plant Physiol. 174: 2183–2198. 1074
Fulton, D.C., Stettler, M., Mettler, T., Vauhjan, C.K., Li, J., Francisco, P., Gil, M., Reinhold, H., 1075 Eicke, S., Messerli, G., Dorken, G., Halliday, K., Smith, A.M., Smith, S.M., and Zeeman, S.C. 1076 (2008). ß-AMYLASE4, a Noncatalytic Protein Required for Starch Breakdown, Acts Upstream of 1077 Three Active ß-Amylases in Arabidopsis Chloroplasts. Plant Cell Online 20: 1040–1058. 1078
Fusari, C.M., Kooke, R., Lauxmann, M.A., Annunziata, M.G., Encke, B., Hoehne, M., Krohn, N., 1079 Becker, F.F.M., Schlereth, A., Sulpice, R., Stitt, M., and Keurentjes, J.J.B. (2017). Genome-1080 wide Association Mapping Reveals that Specific and Pleiotropic Regulatory Mechanisms Fine-1081 tune Central Metabolism and Growth in Arabidopsis. Plant Cell: tpc.00232.2017. 1082
Gómez, L.D., Gilday, A., Feil, R., Lunn, J.E., and Graham, I.A. (2010). AtTPS1-mediated trehalose 1083 6-phosphate synthesis is essential for embryogenic and vegetative growth and responsiveness 1084 to ABA in germinating seeds and stomatal guard cells. Plant J. 64: 1–13. 1085

33
Goward, C.R. and Nicholls, D.J. (1994). Malate dehydrogenase: A model for structure, evolution, 1086 and catalysis. Protein Sci. 3: 1883–1888. 1087
Grau, U.M., Trommer, W.E., and Rossmann, M.G. (1981). Structure of the active ternary complex of 1088 pig heart lactate dehydrogenase with S-lac-NAD at 2.7 Å resolution. J. Mol. Biol. 151: 289–307. 1089
Grefen, C., Donald, N., Hashimoto, K., Kudla, J., Schumacher, K., and Blatt, M.R. (2010). A 1090 ubiquitin-10 promoter-based vector set for fluorescent protein tagging facilitates temporal 1091 stability and native protein distribution in transient and stable expression studies. Plant J. 64: 1092 355–365. 1093
Gujas, B., Cruz, T.M.D., Kastanaki, E., Vermeer, J.E.M., Munnik, T., and Rodriguez-Villalon, A. 1094 (2017). Perturbing phosphoinositide homeostasis oppositely affects vascular differentiation in 1095 Arabidopsis thaliana roots. Development: dev.155788. 1096
Hatch, M.D. and Osmond, C.B. (1976). Compartmentation and Transport in C4 Photosynthesis. In 1097 Stocking CR, Heber U, eds. Transport in plants III, Vol. 3. Berlin:Springer, pp. 144-184. 1098
He, H., Lee, M.-C., Zheng, L.-L., Zheng, L., and Luo, Y. (2013). Integration of the metabolic/redox 1099 state, histone gene switching, DNA replication and S-phase progression by moonlighting 1100 metabolic enzymes. Biosci. Rep. 33: e00018. 1101
Hebbelmann, I., Selinski, J., Wehmeyer, C., Goss, T., Voss, I., Mulo, P., Kangasjärvi, S., Aro, E.-1102 M., Oelze, M.-L., Dietz, K.-J., Nunes-Nesi, A., Do, P.T., Fernie, A.R., Talla, S.K., 1103 Raghavendra, A.S., Linke, V., and Scheibe, R. (2012). Multiple strategies to prevent oxidative 1104 stress in Arabidopsis plants lacking the malate valve enzyme NADP-malate dehydrogenase. J. 1105 Exp. Bot. 63: 1445–1459. 1106
Heber, U. (1974). Metabolite exchange between chloroplasts and cytoplasm. Annu. Rev. Plant 1107 Physiol. 25: 393–421. 1108
Hendriks, W., Mulders, J.W., Bibby, M. a, Slingsby, C., Bloemendal, H., and de Jong, W.W. 1109 (1988). Duck lens epsilon-crystallin and lactate dehydrogenase B4 are identical: a single-copy 1110 gene product with two distinct functions. Proc. Natl. Acad. Sci. U. S. A. 85: 7114–8. 1111
Heyno, E., Innocenti, G., Lemaire, S.D., Issakidis-Bourguet, E., and Krieger-Liszkay, A. (2014). 1112 Putative role of the malate valve enzyme NADP-malate dehydrogenase in H2O2 signalling in 1113 Arabidopsis. Philos. Trans. R. Soc. B Biol. Sci. 369: 20130228–20130228. 1114
Horrer, D., Flütsch, S., Pazmino, D., Matthews, J.S.A., Thalmann, M., Nigro, A., Leonhardt, N., 1115 Lawson, T., and Santelia, D. (2016). Blue light induces a distinct starch degradation pathway in 1116 guard cells for stomatal opening. Curr. Biol. 26: 362–370. 1117
Huberts, D.H.E.W. and van der Klei, I.J. (2010). Moonlighting proteins: An intriguing mode of 1118 multitasking. Biochim. Biophys. Acta - Mol. Cell Res. 1803: 520–525. 1119
Jang, J.C. (1997). Hexokinase as a Sugar Sensor in Higher Plants. Plant Cell Online 9: 5–19. 1120 Jarvis, P., Dormann, P., Peto, C.A., Lutes, J., Benning, C., and Chory, J. (2000). Galactolipid 1121
deficiency and abnormal chloroplast development in the Arabidopsis MGD synthase 1 mutant. 1122 Proc. Natl. Acad. Sci. 97: 8175–8179. 1123
Jeffery, C.J. (1999). Moonlighting proteins. Trends Biochem. Sci. 24: 8–11. 1124 Jeffery, C.J. (2003). Moonlighting proteins: Old proteins learning new tricks. Trends Genet. 19: 415–1125
417. 1126 Journet, E.P., Neuburger, M., and Douce, R. (1981). Role of glutamate-oxaloacetate transaminase 1127
and malate dehydrogenase in the regeneration of NAD for glycine oxidation by spinach leaf 1128 mitochondria. Plant Physiol. 67: 467–469. 1129
Karimi, M., Inze, D., and Depicker, A. (2002). GATEWAY™ vectors for Agrobacterium-mediated 1130 plant transformation. Trends Plant Sci. 7: 193–195. 1131
Kinoshita, H., Nagasaki, J., Yoshikawa, N., Yamamoto, A., Takito, S., Kawasaki, M., Sugiyama, 1132 T., Miyake, H., Weber, A.P.M., and Taniguchi, M. (2011). The chloroplastic 2-1133 oxoglutarate/malate transporter has dual function as the malate valve and in carbon/nitrogen 1134 metabolism. Plant J. 65: 15–26. 1135
Klement, H., Helfrich, M., Oster, U., Schoch, S., and Rüdiger, W. (1999). Pigment-free 1136 NADPH:protochlorophyllide oxidoreductase from Avena sativa L. Purification and substrate 1137 specificity. Eur. J. Biochem. 265: 862–874. 1138
Kobayashi, K., Kondo, M., Fukuda, H., Nishimura, M., and Ohta, H. (2007). Galactolipid synthesis 1139 in chloroplast inner envelope is essential for proper thylakoid biogenesis, photosynthesis, and 1140 embryogenesis. Proc. Natl. Acad. Sci. 104: 17216–17221. 1141
Langklotz, S., Baumann, U., and Narberhaus, F. (2012). Structure and function of the bacterial AAA 1142 protease FtsH. Biochim. Biophys. Acta - Mol. Cell Res. 1823: 40–48. 1143
Lebedev, N., Van Cleve, B., Armstrong, G., and Apel, K. (1995). Chlorophyll Synthesis in a 1144 Deetiolated (det340) Mutant of Arabidopsis without NADPH-Protochlorophyllide (PChlide) 1145

34
Oxidoreductase (POR) A and Photoactive PChlide-F655. Plant Cell 7: 2081–2090. 1146 Lee, C., Schwartz, M.P., Prakash, S., Iwakura, M., and Matouschek, A. (2001). ATP-dependent 1147
proteases degrade their substrates by processively unraveling them from the degradation signal. 1148 Mol. Cell 7: 627–637. 1149
Lee, Y., Rubio, M.C., Alassimone, J., and Geldner, N. (2013). A mechanism for localized lignin 1150 deposition in the endodermis. Cell 153: 402–412. 1151
Lu, X., Zhang, D., Li, S., Su, Y., Liang, Q., Meng, H., Shen, S., Fan, Y., Liu, C., and Zhang, C. 1152 (2014). FtsHi4 is essential for embryogenesis due to its influence on chloroplast development in 1153 Arabidopsis. PLoS One 9. 1154
Majsec, K., Bhuiyan, N.H., Sun, Q., Kumari, S., Kumar, V., Ware, D., and van Wijk, K. (2017). The 1155 Plastid and Mitochondrial Peptidase Network in Arabidopsis: A Foundation for Testing Genetic 1156 Interactions and Functions in Organellar Proteostasis. Plant Cell 29: 2687-2710. 1157
Mansfield, S.G. and Briarty, L.G. (1991). Early Embryogenesis in Arabidopsis thaliana .II. the 1158 Developing Embryo. Can. J. Bot. Can. Bot. 69: 461–476. 1159
Martin, A., Baker, T.A., and Sauer, R.T. (2005). Rebuilt AAA + motors reveal operating principles for 1160 ATP-fuelled machines. Nature 437: 1115–1120. 1161
Mascia, P.N. (1978). Studies of Chloroplast Development in Four Maize Mutants Defective in 1162 Chlorophyll Biosynthesis. Planta. 143: 207–211. 1163
Minárik, P., Tomaásková, N., Kollárová, M., and Antalík, M. (2002). Malate Dehydrogenases - 1164 Structure and function. Gen. Physiol. Biophys. 21: 257–265. 1165
Moore, B. (2003). Role of the Arabidopsis Glucose Sensor HXK1 in Nutrient, Light, and Hormonal 1166 Signaling. Science 300: 332–336. 1167
Moore, B.D. (2004). Bifunctional and moonlighting enzymes: Lighting the way to regulatory control. 1168 Trends Plant Sci. 9: 221–228. 1169
Musrati, R.A., Kollárová, M., Mernik, N., and Mikulášová, D. (1998). Malate Dehydrogenase: 1170 Distribution, Function and Properties. Gen. Physiol. Biophys. 17: 193–210. 1171
Nakai, M. (2018). New perspectives on chloroplast protein import. Plant and Cell Physiology, pcy083, 1172 doi.org/10.1093/pcp/pcy083. 1173
Nicholls, D.J., Miller, J., Scawen, M.D., Clarke, A.R., John Holbrook, J., Atkinson, T., and 1174 Goward, C.R. (1992). The importance of arginine 102 for the substrate specificity of Escherichia 1175 coli malate dehydrogenase. Biochem. Biophys. Res. Commun. 189: 1057–1062. 1176
Nickelsen, J. (2005). The Green Alga Chlamydomonas reinhardtii – A Genetic Model Organism. In 1177 Progress in Botany (Heidelberg, Springer-Verlag Berlin), pp. 68–89. 1178
Nielsen, H. and Engelbrecht, J. (1997). Identification of prokaryotic and eukaryotic signal peptides 1179 and prediction of their cleavage sites Artificial neural networks have been used for many 1180 biological. Protein Eng. 10: 1–6. 1181
Ossowski, S., Schwab, R., and Weigel, D. (2008). Gene silencing in plants using artificial 1182 microRNAs and other small RNAs. Plant J. 53: 674–690. 1183
Park, H., Kreunen, S.S., Cuttriss, A.J., DellaPenna, D., and Pogson, B.J. (2002). Identification of 1184 the Carotenoid Isomerase Provides Insight into Carotenoid Biosynthesis, Prolamellar Body 1185 Formation, and Photomorphogenesis. Plant Cell 14: 321–332. 1186
Patton, D.A., Franzmann, L.H., and Meinke, D.W. (1991). Mapping genes essential for embryo 1187 development in Arabidopsis thaliana. Mol. Gen. Genet. 227: 337–347. 1188
Patton, D.A., Schetter, A.L., Franzmann, L.H., Nelson, K., Ward, E.R., and Meinke, D.W. (1998). 1189 An Embryo-Defective Mutant of Arabidopsis Disrupted in the Final Step of Biotin Synthesis. 1190 Plant Physiol. 116: 935–946. 1191
Pracharoenwattana, I., Cornah, J.E., and Smith, S.M. (2007). Arabidopsis peroxisomal malate 1192 dehydrogenase functions in beta-oxidation but not in the glyoxylate cycle. Plant J. 50: 381–90. 1193
Pracharoenwattana, I., Zhou, W., and Smith, S.M. (2010). Fatty acid beta-oxidation in germinating 1194 Arabidopsis seeds is supported by peroxisomal hydroxypyruvate reductase when malate 1195 dehydrogenase is absent. Plant Mol. Biol. 72: 101–109. 1196
Ruppel, N.J. and Hangarter, R.P. (2007). Mutations in a plastid-localized elongation factor G alter 1197 early stages of plastid development in Arabidopsis thaliana. BMC Plant Biol. 7: 37. 1198
Ryberg, M. and Sundqvist, C. (1982). Characterization of prolamellar bodies and prothylakoids 1199 fractionated from wheat etioplasts. Physiol. Plant. 56: 125–132. 1200
Ryberg, M. and Sundqvist, C. (1988). The regular ultrastructure of isolated prolamellar bodies 1201 depends on the presence of membrane‐bound NADPH‐protochlorophyllide oxidoreductase. 1202 Physiol. Plant. 73: 218–226. 1203
Scheibe, R. (2004). Malate valves to balance cellular energy supply. Physiol. Plant. 120: 21–26. 1204 Scheibe, R. (1987). NADP+‐malate dehydrogenase in C3‐plants: Regulation and role of a light‐1205

35
activated enzyme. Physiol. Plant. 71: 393–400. 1206 Selinski, J., König, N., Wellmeyer, B., Hanke, G.T., Linke, V., Ekkehard Neuhaus, H., and 1207
Scheibe, R. (2014). The plastid-localized NAD-dependent malate dehydrogenase is crucial for 1208 energy homeostasis in developing Arabidopsis thaliana seeds. Mol. Plant 7: 170–186. 1209
Selstam, E., Schelin, J., Brain, T., and Williams, W.P. (2002). The effects of low pH on the 1210 properties of protochlorophyllide oxidoreductase and the organization of prolamellar bodies of 1211 maize (Zea mays). Eur. J. Biochem. 269: 2336–2346. 1212
Seung, D., Thalmann, M., Sparla, F., Abou Hachem, M., Lee, S.K., Issakidis-Bourguet, E., 1213 Svensson, B., Zeeman, S.C., and Santelia, D. (2013). Arabidopsis thaliana AMY3 is a unique 1214 redox-regulated chloroplastic α-amylase. J. Biol. Chem. 288: 33620–33633. 1215
Seung D., Soyk, S., Coiro, M., Maier, B.A., Eicke, S., and Zeeman, S.C. (2015). PROTEIN 1216 TARGETING TO STARCH is required for localising GRANULE-BOUND STARCH SYNTHASE 1217 to starch granules and for normal amylose synthesis in Arabidopsis. PLoS Biol. 13:e1002080. 1218
Sew, Y.S., Ströher, E., Fenske, R., and Millar, A.H. (2016). Loss of Mitochondrial Malate 1219 Dehydrogenase Activity Alters Seed Metabolism Impairing Seed Maturation and Post-1220 Germination Growth in Arabidopsis. Plant Physiol. 171: 849–63. 1221
Silverstein, E. and Sulebele, G. (1969). Catalytic mechanism of pig heart mitochondrial malate 1222 dehydrogenase studied by kinetics at equilibrium. Biochemistry 8: 2543–50. 1223
Sokolenko, A., Pojidaeva, E., Zinchenko, V., Panichkin, V., Glaser, V.M., Herrmann, R.G., and 1224 Shestakov, S. V (2002). The gene complement for proteolysis in the cyanobacterium 1225 Synechocystis sp. PCC 6803 and Arabidopsis thaliana chloroplasts. Curr. Genet. 41: 291–310. 1226
Solymosi, K. and Aronsson, H. (2013). Etioplasts and their significance in chloroplast biogensis 36: 1227 155-167. 1228
Solymosi, K. and Schoefs, B. (2010). Etioplast and etio-chloroplast formation under natural 1229 conditions: The dark side of chlorophyll biosynthesis in angiosperms. Photosynth. Res. 105: 1230 143–166. 1231
Sperling, U., Franck, F., van Cleve, B., Frick, G., Apel, K., and Armstrong, G.A. (1998). Etioplast 1232 differentiation in arabidopsis: both PORA and PORB restore the prolamellar body and 1233 photoactive protochlorophyllide-F655 to the cop1 photomorphogenic mutant. Plant Cell 10: 283–1234 296. 1235
Spicher, L., Almeida, J., Gutbrod, K., Pipitone, R., Dörmann, P., Glauser, G., Rossi, M., and 1236 Kessler, F. (2017). Essential role for phytol kinase and tocopherol in tolerance to combined light 1237 and temperature stress in tomato. J. Exp. Bot. 68: 5845–5856. 1238
Taniguchi, M. and Miyake, H. (2012). Redox-shuttling between chloroplast and cytosol: Integration 1239 of intra-chloroplast and extra-chloroplast metabolism. Curr. Opin. Plant Biol. 15: 252–260. 1240
Tatsuta, T., Augustin, S., Nolden, M., Friedrichs, B., and Langer, T. (2007). m-AAA protease-1241 driven membrane dislocation allows intramembrane cleavage by rhomboid in mitochondria. 1242 EMBO J. 26: 325–335. 1243
Tomaz, T., Bagard, M., Pracharoenwattana, I., Linden, P., Lee, C.P., Carroll, A.J., Stroher, E., 1244 Smith, S.M., Gardestrom, P., and Millar, A.H. (2010). Mitochondrial Malate Dehydrogenase 1245 Lowers Leaf Respiration and Alters Photorespiration and Plant Growth in Arabidopsis. Plant 1246 Physiol. 154: 1143–1157. 1247
Tschopp, M.-A., Iki, T., Brosnan, C.A., Jullien, P.E., and Pumplin, N. (2017). A complex of 1248 Arabidopsis DRB proteins can impair dsRNA processing. RNA 23: 782–797. 1249
Wagner, R., Aigner, H., and Funk, C. (2012). FtsH proteases located in the plant chloroplast. 1250 Physiol. Plant. 145: 203–214. 1251
Wigley, D.B., Gamblin, S.J., Turkenburg, J.P., Dodson, E.J., Piontek, K., Muirhead, H., and 1252 Holbrook, J.J. (1992). Structure of a ternary complex of an allosteric lactate dehydrogenase 1253 from Bacillus stearothermophilus at 2.5 A resolution. J. Mol. Biol. 223: 317–35. 1254
Wistow, G.J., Mulders, J.W., and de Jong, W.W. (1987). The enzyme lactate dehydrogenase as a 1255 structural protein in avian and crocodilian lenses. Nature 326: 622–4. 1256
Yang, S.S. and Zhai, Q.H. (2017). Cytosolic GAPDH: a key mediator in redox signal transduction in 1257 plants. Biol. Plant. 61: 417–426. 1258
Yu, B., Wakao, S., Fan, J., and Benning, C. (2004). Loss of plastidic lysophosphatidic acid 1259 acyltransferase causes embryo-lethality in Arabidopsis. Plant Cell Physiol. 45: 503–510. 1260
Zaffagnini, M., Fermani, S., Costa, A., Lemaire, S.D., and Trost, P. (2013). Plant cytoplasmic 1261 GAPDH: redox post-translational modifications and moonlighting properties. Front. Plant Sci. 4: 1262 1–18. 1263
Zhang, X., Henriques, R., Lin, S.-S., Niu, Q.-W., and Chua, N.-H. (2006). Agrobacterium-mediated 1264 transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 1: 641–6. 1265

36
Zhao, Y., Luo, L., Xu, J., Xin, P., Guo, H., Wu, J., Bai, L., Wang, G., Chu, J., Zuo, J., Hong, Y., 1266 Huang, X., and Li, J. (2018). Malate transported from chloroplast to mitochondrion triggers 1267 production of ROS and PCD in Arabidopsis thaliana. Cell Research 28 (4): 448-461. 1268
1269 1270
FIGURE LEGENDS 1271
Figure 1 Embryo-specific expression of pdNAD-MDH to rescue the embryo lethality 1272
of pdnad-mdh plants. 1273
(A) Schematic diagram of the PABI3:pdNAD-MDH-YFP and PpdNAD-MDH:pdNAD-MDH-1274
YFP constructs. For the former construct, the ABI3 promoter (PABI3) was placed 1275
upstream of the pdNAD-MDH coding sequence fused at its C-terminus to YFP. The 1276
latter construct was described by Beeler et al. (2014). 1277
(B) Opened siliques of pdNAD-MDH+/− plants transformed with the PABI3:pdNAD-1278
MDH-YFP construct. Siliques from the wild type (Ler), untransformed (UT) pdNAD-1279
MDH+/− plants, and pdNAD-MDH+/− transformed with the PpdNAD-MDH:pdNAD-MDH-1280
YFP construct are shown for comparison. White seeds are indicated with an arrow. 1281
Bar = 1 mm. 1282
(C) Expression of the PABI3:pdNAD-MDH-YFP construct in Arabidopsis embryos after 1283
transformation of pdNAD-MDH+/− plants. Expression was detected by fluorescence 1284
microscopy on embryos at three different developmental stages (globular, heart and 1285
torpedo). Bar = 50 µM. 1286
1287
1288
Figure 2 Phenotype of pdnad-mdh mutants rescued by the embryo-specific 1289
expression of pdNAD-MDH. 1290
(A) Photographs of 14-day-old seedlings of pdnad-mdh plants expressing the 1291
PABI3:pdNAD-MDH-YFP construct, grown on ½-strength MS agar plates under a 12-h 1292
light/12-h dark regime. The PABI3:pdNAD-MDH-YFP construct was transformed into 1293
pdNAD-MDH+/− plants, and a T3 population that was segregating for the pdnad-mdh 1294
T-DNA insertion but not for the PABI3:pdNAD-MDH-YFP construct was obtained. Two 1295
examples of pale plants that resembled the constitutive silencing line, miR-mdh-1, 1296
are shown, while the other plants resembled the wild-type Ler, or pdnad-mdh+/−. A 1297
pdnad-mdh −/− plant complemented with the PpdNAD-MDH:pdNAD-MDH-YFP construct 1298
is also shown. Bars = 500 µm. 1299
(B) Photographs of 4-week-old pdnad-mdh−/− seedlings rescued with the 1300
PABI3:pdNAD-MDH-YFP construct. Close-up images of their meristematic zones and 1301

37
their cotyledons are shown. Bars in the left-hand panels = 500 µm, Bars in the 1302
middle and right-hand panels = 100 µm. 1303
(C) Transmission electron micrographs of plastids in pdnad-mdh−/− seedlings 1304
rescued with the PABI3:pdNAD-MDH-YFP construct. Plastids from cotyledons are 1305
shown for the wild type (Ler) and pdnad-mdh −/−, and a plastid in a true leaf of pdnad-1306
mdh −/− is shown on the right. Bars = 500 nm. 1307
1308
Figure 3 Immunoblot and native-PAGE detection of pdNAD-MDH in Ler, miR-mdh-1 1309
and pdnad-mdh−/− seedlings rescued with the PABI3:pdNAD-MDH-YFP construct. 1310
(A) Immunoblots were conducted with the pdNAD-MDH antibody. Equal amounts of 1311
soluble protein (5 µg) were loaded. The migration of molecular weight markers is 1312
indicated (left). 1313
(B) In-gel activity assay of the NAD-MDH. The two bands corresponding to pdNAD-1314
MDH activity are indicated (arrows, right). Equal amounts of protein (15 µg) were 1315
loaded. 1316
1317
Figure 4 β-estradiol-inducible silencing of pdNAD-MDH in mature plant rosettes. 1318
XVE OLEXA:miR-mdh-1 plants photographed before and after a 6 day treatment 1319
with β-estradiol or a mock treatment. 1320
(A) Photographs in the top row were taken before β-estradiol treatment (D0), and 1321
those in the bottom row after 6 days (D6). β-estradiol solution (20 µM) was sprayed 1322
every second day onto the entire rosette. β-estradiol was applied to wild-type (Col) 1323
plants as a control. Mock-treated samples were sprayed with water containing the 1324
same amount of DMSO as the treatment solution (used to dissolve the β-estradiol). 1325
Red arrows indicate examples of white, newly emerging leaves. 1326
(B) Immunoblot detection of pdNAD-MDH in total protein extracts of wild-type and 1327
XVE OLEXA:miR-mdh-1 plants before (D0) and 6 days into β-estradiol treatment 1328
(D6). For the treated XVE OLEXA:miR-mdh-1 plants, proteins were extracted from 1329
the old, green leaves (D6o) and young, white leaves (D6y) separately. Gels were 1330
loaded on an equal leaf area basis. The migration of molecular weight markers is 1331
indicated on the left. pdNAD-MDH and actin (as a loading control) were detected 1332
concurrently on the same membrane using secondary antibodies conjugated to 1333
different infrared fluorescence dyes (800CW for pdNAD-MDH and 680RD for actin). 1334
(C) As for (B), but with old leaves from mock-treated samples. 1335

38
1336
Figure 5 Etiolated growth of miR-mdh-1 and etioplast structure. 1337
(A) Wild-type (Col) and miR-mdh-1 seedlings were grown on ½-strength MS agar 1338
plates in different diel light conditions (24 h dark, 12 h light/12 h dark, 24 h light). Bar 1339
= 1 cm. 1340
(B) Quantification of the hypocotyl length of etiolated wild-type and miR-mdh-1 1341
seedlings. Values represent the mean ± SE of measurements conducted on n=89 1342
and n=86 seedlings for the wild type and miR-mdh-1, respectively. 1343
(C) Etioplast ultrastructure in cotyledons of etiolated wild-type and miR-mdh-1 1344
seedlings observed by transmission electron microscopy. Bars = 500 nm 1345
(D) Proportion of etioplasts showing normal, compromised, or absent prolamellar 1346
body structures in cotyledons of the wild type and miR-mdh-1. The first 139 and 256 1347
etioplasts, observed in wild-type and miR-mdh-1 cotyledons, respectively, were 1348
categorized based on their prolamellar body structure. 1349
1350
Figure 6 Protochlorophylide, galactolipid and β-carotene contents of miR-mdh-1 1351
seedlings. 1352
(A) Protochlorophyllide levels in 5-day-old wild-type (Col) and miR-mdh-1 etiolated 1353
seedlings, measured by fluorescence after excitation at 440 nm. Values are the 1354
means ± SE of three biological replicates, each measured with four technical 1355
replicates. Each biological replicate is a pool of 100-200 individual seedlings. 1356
(B) Immunoblot detection of protochlorophyllide oxidoreductase A (PORA) in total 1357
protein extracts from 6-day-old etiolated wild-type and miR-mdh-1 seedlings (upper 1358
panel). Immunoblots for actin was performed as a loading control (lower panel). Gels 1359
were loaded on an equal fresh weight basis. Three biological replicates are shown. 1360
Each biological replicate is a pool of 50-100 individual seedlings. 1361
(C-F) Lipids and carotenoids were extracted from 6-day-old wild-type and miR-mdh-1 1362
seedlings, either etiolated (e) or light grown (l; grown under a 12-h light/12-h dark 1363
regime). Levels of Diacylglycerol (DAG) (C), MGDG 18:3/16:3 (D) DGDG 18:3/18:3 1364
(E) and β-carotene (F) are shown. For the lipids, only the most common species (in 1365
terms of fatty acid chain composition) is shown; similar trends were seen in other 1366
detected species (Supplemental Table 1). All lipids and carotenoids were quantified 1367
relative to an internal standard, and values were corrected for differences in fresh 1368
weight (see Methods for details). Values are the mean ± SE from four biological 1369

39
replicates. Each biological replicate is a pool of 50-100 individual seedlings. 1370
Significant differences (p < 0.05) within the respective light-grown and etiolated 1371
samples of miR-mdh-1 and the wild type, determined using a two-tailed t-test, are 1372
indicated with an asterisk. 1373
1374
Figure 7 Complementation test with various NAD-MDH isoforms expressed in 1375
pdnad-mdh. 1376
(A) Constructs encoding NAD-MDH isoforms under the control of the pdNAD-MDH 1377
promoter. The plastidial transit peptide from RubisCO small subunit was fused to the 1378
N-terminus of each isoform. The constructs shown are tagged at the C-terminus with 1379
YFP. Similar constructs were cloned with the Flag-HA tag in place of YFP. 1380
(B) Genotyping results of the T2 progeny from T1 plants heterozygous for the pdnad-1381
mdh mutation and expressing the different NAD-MDH isoforms. Numbers above the 1382
bars indicate the number of BASTA-resistant T2 plants that were genotyped. 1383
(C) Chloroplast localization of the NAD-MDH isoforms. Similar constructs to those 1384
shown in A, except with the 35S promoter in place of the native promoter, were 1385
transiently expressed in Nicotiana benthamiana leaves and imaged using confocal 1386
microscopy. Bar = 5 µm. 1387
1388
Figure 8 Enzymatically-inactive pdNAD-MDH proteins can complement the embryo 1389
lethality and growth defects of the pdnad-mdh mutant. 1390
(A) Catalytic centre of the Arabidopsis pdNAD-MDH structure. The pdNAD-MDH 1391
protein sequence (without cTP) was modelled using the human malate- and NADH-1392
bound MMDH2 crystal structure (PDB: 2DFD) as a template. Left panel shows 1393
NADH (red) and malate (yellow), and the relative position of the conserved catalytic 1394
amino acid residues of pdNAD-MDH. A detailed view of the malate-binding site is 1395
shown in the upper right panel. The surface of the catalytic pocket is shown in the 1396
lower right panel. 1397
(B) An in-vitro activity assay was performed using purified recombinant proteins. The 1398
proteins (10 µg) were incubated with an excess of cofactor (NADH) at 22°C. The 1399
reaction was started by the addition of excess substrate (oxaloacetate). The velocity 1400
was determined by measuring the decrease in absorbance at 340 nm resulting from 1401
the conversion of NADH to NAD+. Error bars indicate mean ± SE (n=3). 1402
(C) Genotyping results of T2 progeny from T1 plants heterozygous for pdnad-mdh 1403

40
and expressing the enzymatically-inactive pdNAD-MDH proteins. Numbers above 1404
the bars indicate the number of BASTA-resistant T2 plants that were genotyped. 1405
(D) Photographs showing 3-week-old rosettes of homozygous pdnad-mdh plants 1406
complemented with enzymatically-inactive pdNAD-MDH mutants. For comparison, 1407
wild-type (Ler) plants are shown. Plants were grown under long days (16 h light/8 h 1408
dark). 1409
(E) NAD-MDH activity observed by native-PAGE. Equal amounts of protein (15 µg) 1410
were loaded per lane, and all lanes were run on the same gel. pdNAD-MDH runs in 1411
distinct activity bands (a,b,c). While the fastest migrating band (c) corresponding to 1412
the free dimer is masked by other NAD-MDH activity, the two slower migrating bands 1413
that correspond to NAD-MDH in protein complexes can be easily observed (a, b). 1414
Additional activity bands (a’, b’) are observed for plants heterozygous for the pdnad-1415
mdh T-DNA insertion expressing the catalytic-inactive pdNAD-MDH variants, 1416
possibly due to dimer formation between an inactive pdNAD-MDH with an 1417
endogenous form of pdNAD-MDH protein. 1418
1419
Figure 9 Constitutive and inducible silencing of the FtsH12 protein. 1420
(A) In Arabidopsis, the FtsH12 gene consists of 19 exons and 18 introns. The 1421
sequence and position of the target for artificial microRNA silencing are indicated. 1422
Numbers represent nucleotide positions relative to the translational start site +1. 1423
(B) Constitutive silencing of FtsH12 resulted in plants with varying degrees of 1424
paleness and delayed growth phenotype in the T1 generation. Plants were grown 1425
under a 12-h light/12-h dark regime for 6 weeks. Both amiRNA constructs produced 1426
plants with comparable phenotypes. The identical wild type (Col) plant was used for 1427
both the left and right panels. Bar = 2 cm. 1428
(C) Immunoblot analysis of total protein extracts from the amiRNA FtsH12 lines, 1429
using the FtsH12 antibody (upper panel), pdNAD-MDH antibody (lower panel) and 1430
actin antibody as a loading control (both panels). FtsH12 and actin, as well as 1431
pdNAD-MDH and actin, were detected concurrently on the same membrane using 1432
secondary antibodies conjugated to different infrared fluorescence dyes (800CW for 1433
FtsH12 and pdNAD-MDH, and 680RD for actin). The migration of molecular weight 1434
markers is indicated (left). The gel was loaded on an equal protein (15 µg) basis. 1435
1436
Figure 10 Isothermal titration calorimetry (ITC) thermograms of NADH titrated into 1437

41
the pdNAD-MDH WT and inactive proteins. 1438
(A) ITC profile of NADH injected into a solution of recombinant His-pdNAD-MDH WT 1439
protein at 25°C. The upper panel shows the raw calorimetric data. The plot below 1440
shows the integrated enthalpy as a function of the NADH/pdNAD-MDH molar ratio. 1441
Reactions are exothermic. 1442
(B-D) Experiments were conducted as described for (A) with the His-pdNAD-MDH 1443
R162Q protein (B), the His-pdNAD-MDH R234Q protein (C), and the His-pdNAD-1444
MDH R162Q R234Q protein (D). 1445
1446
Figure 11 A model for the interaction of pdNAD-MDH with the heteromeric FtsH12-1447
FtsHi AAA-ATPase complex at the chloroplast inner envelope membrane, which 1448
plays an essential role in chloroplast development. 1449
1450
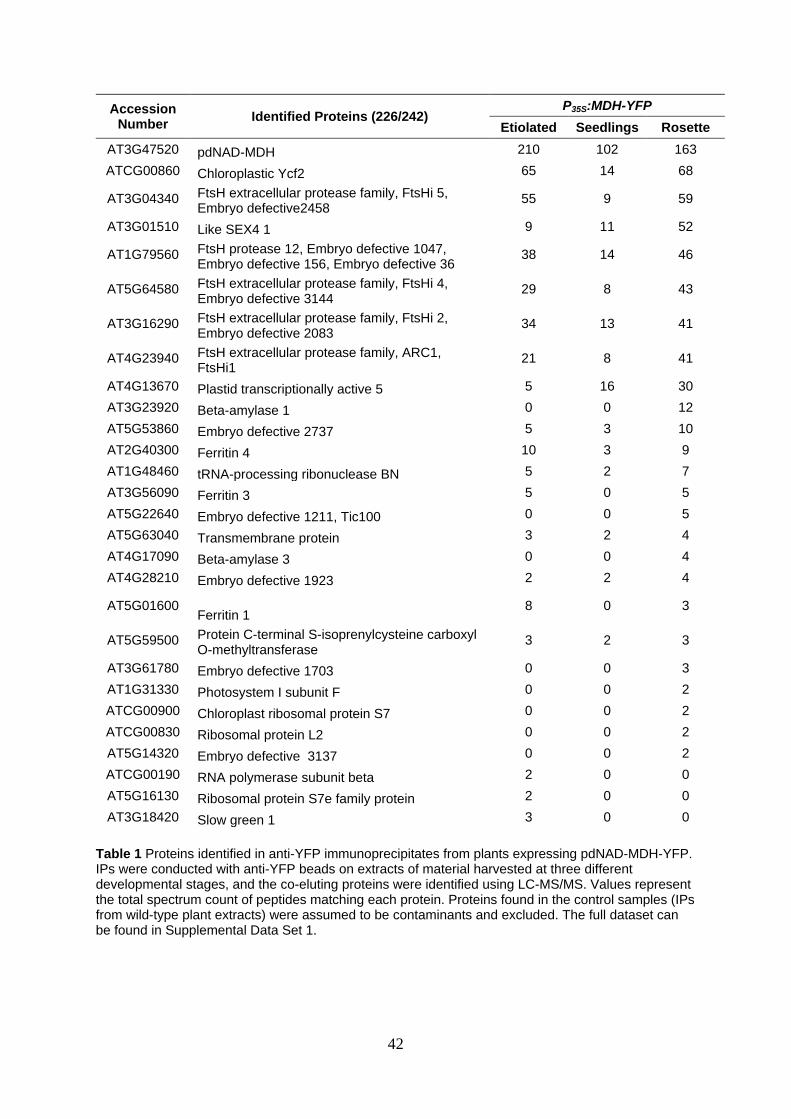
42
Accession Number
Identified Proteins (226/242) P35S:MDH-YFP
Etiolated Seedlings Rosette
AT3G47520 pdNAD-MDH 210 102 163
ATCG00860 Chloroplastic Ycf2 65 14 68
AT3G04340 FtsH extracellular protease family, FtsHi 5, Embryo defective2458
55 9 59
AT3G01510 Like SEX4 1 9 11 52
AT1G79560 FtsH protease 12, Embryo defective 1047, Embryo defective 156, Embryo defective 36
38 14 46
AT5G64580 FtsH extracellular protease family, FtsHi 4, Embryo defective 3144
29 8 43
AT3G16290 FtsH extracellular protease family, FtsHi 2, Embryo defective 2083
34 13 41
AT4G23940 FtsH extracellular protease family, ARC1, FtsHi1
21 8 41
AT4G13670 Plastid transcriptionally active 5 5 16 30
AT3G23920 Beta-amylase 1 0 0 12
AT5G53860 Embryo defective 2737 5 3 10
AT2G40300 Ferritin 4 10 3 9
AT1G48460 tRNA-processing ribonuclease BN 5 2 7
AT3G56090 Ferritin 3 5 0 5
AT5G22640 Embryo defective 1211, Tic100 0 0 5
AT5G63040 Transmembrane protein 3 2 4
AT4G17090 Beta-amylase 3 0 0 4
AT4G28210 Embryo defective 1923 2 2 4
AT5G01600 Ferritin 1
8 0 3
AT5G59500 Protein C-terminal S-isoprenylcysteine carboxyl O-methyltransferase
3 2 3
AT3G61780 Embryo defective 1703 0 0 3
AT1G31330 Photosystem I subunit F 0 0 2
ATCG00900 Chloroplast ribosomal protein S7 0 0 2
ATCG00830 Ribosomal protein L2 0 0 2
AT5G14320 Embryo defective 3137 0 0 2
ATCG00190 RNA polymerase subunit beta 2 0 0
AT5G16130 Ribosomal protein S7e family protein 2 0 0
AT3G18420 Slow green 1 3 0 0
Table 1 Proteins identified in anti-YFP immunoprecipitates from plants expressing pdNAD-MDH-YFP. IPs were conducted with anti-YFP beads on extracts of material harvested at three different developmental stages, and the co-eluting proteins were identified using LC-MS/MS. Values represent the total spectrum count of peptides matching each protein. Proteins found in the control samples (IPs from wild-type plant extracts) were assumed to be contaminants and excluded. The full dataset can be found in Supplemental Data Set 1.

43
Accession Number
Identified Proteins (236/247) PUBQ10:FtsH12-YFP
1 2
AT1G79560 FtsH protease 12, Embryo defective 1047, Embryo defective 156, Embryo defective 36
386 371
ATCG00860 Chloroplast Ycf2 181 237
AT3G04340 FtsH extracellular protease family, FtsHi 5, Embryo defective2458
247 179
ATCG01130 Chloroplast Ycf1, TIC214 70 111
AT3G16290 FtsH extracellular protease family, FtsHi 2, Embryo defective 2083
46 86
AT5G64580 FtsH extracellular protease family, FtsHi 4, Embryo defective 3144
66 83
AT4G23940 FtsH extracellular protease family, ARC1, FtsHi1 69 52
AT1G01320 Tetratricopeptide repeat (TPR)-like superfamily protein
6 32
AT4G02510 Translocon at the outer envelope membrane of chloroplasts 159
37 31
AT5G53860 Embryo defective 2737 13 26
ATCG00480 ATP synthase subunit beta 8 23
AT3G47520 pdNAD-MDH 5 22
AT1G07920 GTP binding Elongation factor Tu family protein 15 20
AT1G43170 Ribosomal protein 1 5 20
ATCG00120 ATP synthase subunit alpha 42 19
ATCG00490 Ribulose-bisphosphate carboxylase 48 16
AT5G01590 Histone-lysine N-methyltransferase ATXR3-like protein
14 16
AT2G41840 Ribosomal protein S5 family protein 6 16
Table 2 Proteins identified in anti-YFP immunoprecipitates from two independent lines overexpressing FtsH12-YFP. The IPs were conducted with anti-YFP beads on extracts of material harvested from rosette leaves, and the co-eluting proteins were identified using LC-MS/MS. Values represent the total spectrum count of peptides matching each protein. Proteins found in the control sample (IPs from wild-type plants) were assumed to be contaminants and excluded. The full dataset can be found in Supplemental Data Set 1.

44
Accession Number
Identified Proteins (102/115)
pdnad-mdh+/-
At pdNAD-
MDH
At cyMDH1
Sc mMDH1
At mMDH1
At pMDH1
GFP_AEQVI YFP-tag 108 7 3 66 162
AT3G47520 pdNAD-MDH 142 3 0 18 6
AT1G04410 cyMDH1 0 8 5 0 0
AT1G53240 mMDH1 4 0 0 96 0
AT2G22780 pMDH1 0 2 2 0 312
ATCG00860 Chloroplast Ycf2 193 0 0 4 0
AT3G04340 FtsH extracellular protease family, FtsHi 5, Embryo defective2458
192 0 0 0 0
AT1G79560 FtsH protease 12, Embryo defective 1047, Embryo defective 156, Embryo defective 36
155 0 0 4 0
AT5G64580 FtsH extracellular protease family, FtsHi 4, Embryo defective 3144
102 0 0 0 0
AT3G01510 Like SEX4 1 82 0 0 0 0
AT4G23940 FtsH extracellular protease family, ARC1, FtsHi1
69 0 0 0 0
ATCG01130 Chloroplast Ycf1 protein 59 0 0 0 0
AT3G16290 FtsH extracellular protease family, FtsHi 2, Embryo defective 2083
54 0 0 0 0
AT5G53860 Embryo defective 2737 29 0 0 0 0
AT4G02510 Translocon at the outer envelope membrane of chloroplasts 159
29 0 0 0 0
Table 3 Proteins identified in IPs of YFP-tagged NAD-MDH proteins (AtpdNAD-MDH, AtcyMDH1, ScmMDH1, AtmMDH1 and AtpMDH) expressed in heterozygous pdnad-mdh plants. IPs were conducted with anti-YFP beads on extracts of rosette leaves, and the co-eluting proteins were identified using LC-MS/MS. Values represent the total spectrum count of peptides matching each protein. Proteins also identified in the control sample (Ler) were assumed to be contaminants and excluded. The full dataset can be found in Supplemental Data Set 1.

45
Accession Number
Identified Proteins (917/929)
pdnad-mdh-/-
pdNAD-MDH WT
pdNAD-MDH
R162Q
pdNAD-MDH
R234Q
pdNAD-MDH
R163Q R234Q
AT3G47520 pdNAD-MDH 96 49 31 22
ATCG00860 Chloroplast Ycf2 283 457 332 405
AT3G04340 FtsH extracellular protease family, FtsHi 5, Embryo defective2458
408 330 210 362
AT1G79560 FtsH protease 12, Embryo defective 1047, Embryo defective 156, Embryo defective 36
373 309 212 317
AT5G64580 FtsH extracellular protease family, FtsHi 4, Embryo defective 3144
221 207 164 159
AT3G16290 FtsH extracellular protease family, FtsHi 2, Embryo defective 2083
125 228 158 179
ATCG01130 Chloroplast Ycf1 70 166 174 223
AT4G23940 FtsH extracellular protease family, ARC1, FtsHi1
153 141 153 109
AT4G02510 Translocon at the outer envelope membrane of chloroplasts 159
25 95 113 133
AT3G01510 like SEX4 1 159 58 105 23
AT5G53860 Embryo defective 2737 30 54 55 54
Table 4 Proteins identified in anti-HA immunoprecipitates from homozygous pdnad-mdh plants expressing non-catalytic versions of pdNAD-MDH. The IPs were conducted with anti-HA beads on extracts of rosette leaves, and the co-eluting proteins were identified using LC-MS/MS. Values represent the total spectrum count of peptides matching each protein. Proteins found in the control sample (IPs from wild-type [Ler] plant extracts) were assumed to be contaminants and excluded. The full dataset can be found in Supplemental Data Set 1.

46
1451

Figure 1 Embryo-specific expression of pdNAD-MDH to rescue the embryo lethality of pdnad-mdh plants.
(A) Schematic diagram of the PABI3:pdNAD-MDH-YFP and PpdNAD-MDH:pdNAD-MDH-YFP constructs. For the former construct,
the ABI3 promoter (PABI3) was placed upstream of the pdNAD-MDH coding sequence fused at its C-terminus to YFP. The
latter construct was described by Beeler et al. (2014).
(B) Opened siliques of pdNAD-MDH+/− plants transformed with the PABI3:pdNAD-MDH-YFP construct. Siliques from the wild
type (Ler), untransformed (UT) pdNAD-MDH+/− plants, and pdNAD-MDH+/− transformed with the PpdNAD-MDH:pdNAD-MDH-
YFP construct are shown for comparison. White seeds are indicated with an arrow. Bar = 1 mm.
(C) Expression of the PABI3:pdNAD-MDH-YFP construct in Arabidopsis embryos after transformation of pdNAD-MDH+/−
plants. Expression was detected by fluorescence microscopy on embryos at three different developmental stages (globular,
heart and torpedo). Bar = 50 µM.
A
1000 bp
Scale
C
Ler UT
PABI3:
pdNAD-MDH-
YFP
PpdNAD-MDH:
pdNAD-MDH-
YFP
pdnad-mdh+/- B
YFP pdNAD-MDH PABI3
YFP pdNAD-MDH PpdNAD-MDH

Figure 2 Phenotype of pdnad-mdh mutants rescued by the embryo-specific expression of pdNAD-MDH.
(A) Photographs of 14-day-old seedlings of pdnad-mdh plants expressing the PABI3:pdNAD-MDH-YFP construct, grown on ½-
strength MS agar plates under a 12-h light/12-h dark regime. The PABI3:pdNAD-MDH-YFP construct was transformed into
pdNAD-MDH+/− plants, and a T3 population that was segregating for the pdnad-mdh T-DNA insertion but not for the
PABI3:pdNAD-MDH-YFP construct was obtained. Two examples of pale plants that resembled the constitutive silencing line,
miR-mdh-1, are shown, while the other plants resembled the wild-type Ler, or pdnad-mdh+/−. A pdnad-mdh −/− plant
complemented with the PpdNAD-MDH:pdNAD-MDH-YFP construct is also shown. Bars = 500 µm.
(B) Photographs of 4-week-old pdnad-mdh−/− seedlings rescued with the PABI3:pdNAD-MDH-YFP construct. Close-up images
of their meristematic zones and their cotyledons are shown. Bars in the left-hand panels = 500 µm, Bars in the middle and
right-hand panels = 100 µm.
(C) Transmission electron micrographs of plastids in pdnad-mdh−/− seedlings rescued with the PABI3:pdNAD-MDH-YFP
construct. Plastids from cotyledons are shown for the wild type (Ler) and pdnad-mdh −/−, and a plastid in a true leaf of pdnad-
mdh −/− is shown on the right. Bars = 500 nm.
C
Ler (cotyledon)
pdnad-mdh−/− PABI3:pdNAD-
MDH-YFP #1 (cotyledon)
pdnad-mdh−/− PABI3:pdNAD-
MDH-YFP #1 (true leaf)
Ler pdnad-mdh+/− pdnad-mdh
PABI3:pdNAD-MDH-YFP (T3 gen.)
A miR-mdh-1
pdnad-mdh −/− PpdNAD-MDH:
pdNAD-MDH-
YFP
B pdnad-mdh −/− PABI3:pdNAD-MDH-YFP #1
pdnad-mdh −/− PABI3: pdNAD-MDH-YFP #2

Figure 3 Immunoblot and native-PAGE detection of pdNAD-MDH in Ler, miR-mdh-1 and pdnad-mdh−/− seedlings rescued
with the PABI3:pdNAD-MDH-YFP construct.
(A) Immunoblots were conducted with the pdNAD-MDH antibody. Equal amounts of soluble protein (5 µg) were loaded. The
migration of molecular weight markers is indicated (left).
(B) In-gel activity assay of the NAD-MDH. The two bands corresponding to pdNAD-MDH activity are indicated (arrows, right).
Equal amounts of protein (15 µg) were loaded.
A
Ler
pdnad-mdh−/− PABI3:pdNAD-
MDH-YFP #2
miR-
mdh-1 kDa
35 pdNAD-
MDH
B
55

Figure 4 β-estradiol-inducible silencing of pdNAD-MDH in mature plant rosettes. XVE OLEXA:miR-mdh-1 plants photographed
before and after a 6 day treatment with β-estradiol or a mock treatment.
(A) Photographs in the top row were taken before β-estradiol treatment (D0), and those in the bottom row after 6 days (D6).
β-estradiol solution (20 µM) was sprayed every second day onto the entire rosette. β-estradiol was applied to wild-type (Col)
plants as a control. Mock-treated samples were sprayed with water containing the same amount of DMSO as the treatment
solution (used to dissolve the β-estradiol). Red arrows indicate examples of white, newly emerging leaves.
(B) Immunoblot detection of pdNAD-MDH in total protein extracts of wild-type and XVE OLEXA:miR-mdh-1 plants before (D0)
and 6 days into β-estradiol treatment (D6). For the treated XVE OLEXA:miR-mdh-1 plants, proteins were extracted from the
old, green leaves (D6o) and young, white leaves (D6y) separately. Gels were loaded on an equal leaf area basis. The
migration of molecular weight markers is indicated on the left. pdNAD-MDH and actin (as a loading control) were detected
concurrently on the same membrane using secondary antibodies conjugated to different infrared fluorescence dyes (800CW
for pdNAD-MDH and 680RD for actin).
(C) As for (B), but with old leaves from mock-treated samples.
A
XVE OLEXA:miR-mdh-1
Col #1-2 #2-3
XVE OLEXA:miR-mdh-1
Col #1-2 #2-3
β-estradiol treatment mock treatment
kDa
55
35 pdNAD-MDH
actin
Col
XVE OLEXA:miR-mdh-1
D0 D6
1 2 1 2 1 2 1 2 1 2 1 2
D0 D6 D0
#1-2 #2-3
D6
C
B
pdNAD-
MDH
kDa
55
35
Col
XVE OLEXA:miR-mdh-1
D0 D6
1 2 1 2 1 2 1o 1y 2o 2y 1 2
D0 D6 D0 D6
#1-2
1o 1y 2o 2y
#2-3
actin

miR-mdh-1
miR-mdh-1
miR-mdh-1
Col
Col
miR-mdh-1
Col D
88.5
3.9
1.4
59.8
10.1
36.3
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Col miR-mdh-1
normal compromised absent
Prolamellar body (PLB) structure
0
2
4
6
8
10
12
14
16
18
20
Col-0 miR-mdh-1
hypocoty
l le
ngth
[m
m]
Col miR-mdh-1
Figure 5 Etiolated growth of miR-mdh-1 and etioplast structure.
(A) Wild-type (Col) and miR-mdh-1 seedlings were grown on ½-strength MS agar plates in different diel light conditions (24 h
dark, 12 h light/12 h dark, 24 h light). Bar = 1 cm.
(B) Quantification of the hypocotyl length of etiolated wild-type and miR-mdh-1 seedlings. Values represent the mean ± SE of
measurements conducted on n=89 and n=86 seedlings for the wild type and miR-mdh-1, respectively.
(C) Etioplast ultrastructure in cotyledons of etiolated wild-type and miR-mdh-1 seedlings observed by transmission electron
microscopy. Bars = 500 nm
(D) Proportion of etioplasts showing normal, compromised, or absent prolamellar body structures in cotyledons of the wild
type and miR-mdh-1. The first 139 and 256 etioplasts, observed in wild-type and miR-mdh-1 cotyledons respectively, were
categorized based on their prolamellar body structure.
A B
C
Col miR-mdh-1
Col miR-mdh-1
24 h dark
12 h light/12 h dark
24 h light
100
90
80
70
60
50
40
30
20
10
Perc
enta
ge o
f etiopla
sts
[%
]

A B
0
0.5
1
1.5
2
2.5
Col-0 miR-mdh-1
Flu
ore
sce
nce
/mg
FW
Col miR-mdh-1
E F
0
50
100
150
200
250
300
350
Col-0, lightgrown
Col-0, darkgrown
miR-mdh-1,light grown
miR-mdh-1,dark grown
β-carotene
Re
l. q
uantifica
tion
Col miR-mdh-1
e e l l
* *
C
Re
l. q
uantifica
tion
*
α-PORA
Col
miR-
mdh-1 Col
miR-
mdh-1 Col
miR-
mdh-1
α-actin
Protochlorophyllide
Figure 6 Protochlorophylide, galactolipid and β-carotene content of miR-mdh-1 seedlings.
(A) Protochlorophyllide levels in 5-day-old wild-type (Col) and miR-mdh-1 etiolated seedlings, measured by fluorescence after
excitation at 440 nm. Values are the means ± SE of three biological replicates, each measured with four technical replicates.
Each biological replicate is a pool of 50-100 individual seedlings.
(B) Immunoblot detection of protochlorophyllide oxidoreductase A (PORA) in total protein extracts from 6-day-old etiolated
wild-type and miR-mdh-1 seedlings (upper panel). Immunoblots for actin was performed as a loading control (lower panel).
Gels were loaded on an equal fresh weight basis. Three biological replicates are shown. Each biological replicate is a pool of
50-100 individual seedlings.
(C-F) Lipids and carotenoids were extracted from 6-day-old wild-type and miR-mdh-1 seedlings, either etiolated (e) or light
grown (l; grown under a 12-h light/12-h dark regime). Levels of Diacylglycerol (DAG) (C), MGDG 18:3/16:3 (D) DGDG
18:3/18:3 (E) and β-carotene (F) are shown. For the lipids, only the most common species (in terms of fatty acid chain
composition) is shown; similar trends were seen in other detected species (Supplemental Table 1). All lipids and carotenoids
were quantified relative to an internal standard, and values were corrected for differences in fresh weight (see Methods for
details). Values are the mean ± SE from four biological replicates. Each biological replicate is a pool of 50-100 individual
seedlings. Significant differences (p < 0.05) within the respective light-grown and etiolated samples of miR-mdh-1 and the
wild type, determined using a two-tailed t-test, are indicated with an asterisk.
Re
l. q
uantifica
tion
D
Re
l. q
uantifica
tion
0
500
1000
1500
2000
2500
Col-0, lightgrown
Col-0, darkgrown
miR-mdh-1,light grown
miR-mdh-1,dark grown
MGDG-18:3/16:3
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
Col-0, lightgrown
Col-0, darkgrown
miR-mdh-1,light grown
miR-mdh-1,dark grown
DAG-18:0/18:0
e e l l
Col miR-mdh-1
e e l l
*
*
Col miR-mdh-1
0
200
400
600
800
1000
1200
Col-0, lightgrown
Col-0, darkgrown
miR-mdh-1,light grown
miR-mdh-1,dark grown
DGDG-18:3/18:3
Col miR-mdh-1
e e l l
*
*
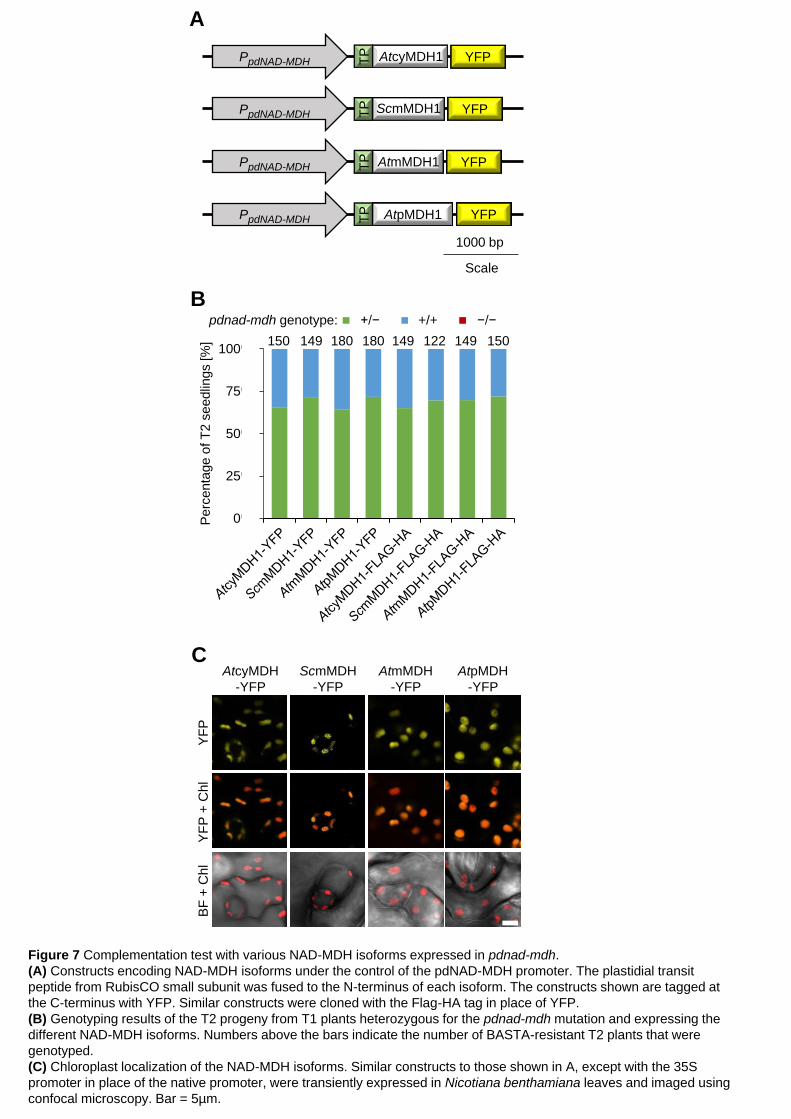
Figure 7 Complementation test with various NAD-MDH isoforms expressed in pdnad-mdh.
(A) Constructs encoding NAD-MDH isoforms under the control of the pdNAD-MDH promoter. The plastidial transit
peptide from RubisCO small subunit was fused to the N-terminus of each isoform. The constructs shown are tagged at
the C-terminus with YFP. Similar constructs were cloned with the Flag-HA tag in place of YFP.
(B) Genotyping results of the T2 progeny from T1 plants heterozygous for the pdnad-mdh mutation and expressing the
different NAD-MDH isoforms. Numbers above the bars indicate the number of BASTA-resistant T2 plants that were
genotyped.
(C) Chloroplast localization of the NAD-MDH isoforms. Similar constructs to those shown in A, except with the 35S
promoter in place of the native promoter, were transiently expressed in Nicotiana benthamiana leaves and imaged using
confocal microscopy. Bar = 5µm.
1000 bp
Scale
A
PpdNAD-MDH TP
YFP
PpdNAD-MDH TP
PpdNAD-MDH TP
YFP
PpdNAD-MDH AtpMDH1 TP
YFP
YFP
AtmMDH1
ScmMDH1
AtcyMDH1
0%
25%
50%
75%
100%
At
CY
MD
H-Y
FP
Sc M
MD
H-Y
FP
At
MM
DH
-YF
P
At
PM
DH
-YF
P
At
CY
MD
H-F
lag
HA
Sc M
MD
H-F
lag
HA
At
MM
DH
-Fla
gH
A
At
PM
DH
-Fla
gH
A
pdnad-mdh+/-
He Wt Ho
Pe
rcenta
ge o
f T
2 s
eedlin
gs [%
]
+/− +/+ −/− pdnad-mdh genotype:
B
150 149 180 180 149 122 149 150
AtcyMDH
-YFP
ScmMDH
-YFP
AtmMDH
-YFP
AtpMDH
-YFP
YF
P
YF
P +
Ch
l B
F +
Ch
l
C

Figure 8 Enzymatically-inactive pdNAD-MDH proteins can complement the embryo lethality and growth defects of the pdnad-mdh
mutant.
(A) Catalytic centre of the Arabidopsis pdNAD-MDH structure. The pdNAD-MDH protein sequence (without cTP) was modelled
using the human malate- and NADH-bound MMDH2 crystal structure (PDB: 2DFD) as a template. Left panel shows NADH (red)
and malate (yellow), and the relative position of the conserved catalytic amino acid residues of pdNAD-MDH. A detailed view of the
malate-binding site is shown in the upper right panel. The surface of the catalytic pocket is shown in the lower right panel.
(B) An in-vitro activity assay was performed using purified recombinant proteins. The proteins (10 µg) were incubated with an
excess of cofactor (NADH) at 22°C. The reaction was started by the addition of excess substrate (oxaloacetate). The velocity was
determined by measuring the decrease in absorbance at 340 nm resulting from the conversion of NADH to NAD+. Error bars
indicate mean ± SE (n=3).
(C) Genotyping results of T2 progeny from T1 plants heterozygous for pdnad-mdh and expressing the enzymatically-inactive
pdNAD-MDH proteins. Numbers above the bars indicate the number of BASTA-resistant T2 plants that were genotyped.
(D) Photographs showing 3-week-old rosettes of homozygous pdnad-mdh plants complemented with enzymatically-inactive pdNAD-
MDH mutants. For comparison, wild-type (Ler) plants are shown. Plants were grown under long days (16 h light/8 h dark).
(E) NAD-MDH activity observed by native-PAGE. Equal amounts of protein (15 µg) were loaded per lane, and all lanes were run on
the same gel. pdNAD-MDH runs in distinct activity bands (a,b,c). While the fastest migrating band (c) corresponding to the free
dimer is masked by other NAD-MDH activity, the two slower migrating bands that correspond to NAD-MDH in protein complexes
can be easily observed (a,b). Additional activity bands (a’, b’) are observed for plants heterozygous for the pdnad-mdh T-DNA
insertion expressing the catalytic-inactive pdNAD-MDH variants, possibly due to dimer formation between an inactive pdNAD-MDH
with an endogenous form of pdNAD-MDH protein.
H258
D231
R162
R168
R234
D231
R234
H258
R168
R162
D115
0.00
0.02
0.04
0.06
0.08
0.10
0.12
ve
loctiy [m
in-1
]
pdNAD-MDH
WT R162Q R234Q R162Q
R234Q
blank
A
C
E
0%
25%
50%
75%
100%
pdNAD-MDHR162Q
pdNAD-MDHR234Q
pdNAD-MDHR162QR234Q
pdnad-mdh+/-
He Wt Ho
Pe
rcenta
ge o
f T
2 s
eedlin
gs [%
]
R162Q R234Q R162Q
R234Q
pdNAD-MDH
construct:
+/− +/+ −/− pdnad-mdh genotype:
pdnad-mdh background
Le
r
#2
-3-1
#3
-1-1
7
#2
-3-2
1
+/− −/−
R162Q pdNAD-MDH construct:
pdnad-mdh genotype:
#2
-4-1
#2
-4-1
3
#3
-6-2
0
#3
-4-1
#3
-4-3
#4
-5-2
8
+/− −/−
R234Q
+/− −/−
R162Q R234Q
B
D
Ler pdnad-mdh-/-
R162Q R234Q
R162Q
R234Q
pdNAD-MDH construct: 73 69 78
Co
l
miR
-
md
h-1
a
b
c
a’
b’

+3259 +3279
5’UTR 3’UTR
+1 +5333bp
At1G79560
GAGACAAGCTGTGATATTTAT
CCGTGAGTTGACAAGGTATAT
+4777 +4797 amiRNA target A
amiRNA target B
A
B
C
Figure 9 Constitutive and inducible silencing of the FtsH12 protein.
(A) In Arabidopsis, the FtsH12 gene consists of 19 exons and 18 introns. The sequence and position of the target for
artificial microRNA silencing are indicated. Numbers represent nucleotide positions relative to the translational start site
+1.
(B) Constitutive silencing of FtsH12 resulted in plants with varying degrees of paleness and delayed growth phenotype
in the T1 generation. Plants were grown under a 12-h light/12-h dark regime for 6 weeks. Both amiRNA constructs
produced plants with comparable phenotypes. The identical wild type (Col) plant was used for both the left and right
panels. Bar = 2 cm.
(C) Immunoblot analysis of total protein extracts from the amiRNA FtsH12 lines, using the FtsH12 antibody (upper
panel), pdNAD-MDH antibody (lower panel) and actin antibody as a loading control (both panels). The migration of
molecular weight markers is indicated (left). The gel was loaded on an equal protein (15 µg) basis.
miR-mdh-1
amiRNA
FtsH12 A 3-1
amiRNA
FtsH12 B 2-1
Col
Col amiRNA FtsH12 A
14-1 11-1
Col amiRNA FtsH12 B
4-1 1-1
Col amiRNA FtsH12 A
actin
FtsH12
actin
14-1 11-1 3-1 1 2
70
100
130
55
55
35
kDa
pdNAD-MDH
miR-
mdh-1
amiRNA FtsH12 B
4-1 1-1 2-1

Figure 10 Isothermal titration calorimetry (ITC) thermograms of NADH titrated into the pdNAD-MDH WT
and inactive proteins.
(A) ITC profile of NADH injected into a solution of recombinant His-pdNAD-MDH WT protein at 25°C. The
upper panel shows the raw calorimetric data. The plot below shows the integrated enthalpy as a function of
the NADH/pdNAD-MDH molar ratio. Reactions are exothermic.
(B-D) Experiments were conducted as described for (A) with the His-pdNAD-MDH R162Q protein (B), the
His-pdNAD-MDH R234Q protein (C), and the His-pdNAD-MDH R162Q R234Q protein (D).
pdNAD-MDH R234Q pdNAD-MDH R162Q R234Q
A B
C D
pdNAD-MDH WT pdNAD-MDH R162Q
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5
-6
-4
-2
0
-0.4
-0.3
-0.2
-0.1
0.0
0.10 25 50 75 100 125 150 175 200 225
Time (min)
µca
l/sec
Molar ratio
kcal/m
ole
of in
ject
ant
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5-6
-4
-2
0
-0.4
-0.3
-0.2
-0.1
0.0
0.10 25 50 75 100 125 150 175 200 225
Time (min)
µca
l/sec
Molar ratio
kcal/m
ole
of in
ject
ant
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0-6
-4
-2
0
2
-0.6
-0.5
-0.4
-0.3
-0.2
-0.1
0.0
0.10 25 50 75 100 125 150 175 200 225
Time (min)
µca
l/sec
Molar ratio
kcal/m
ole
of in
ject
ant
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0-6
-4
-2
0
-0.7
-0.6
-0.5
-0.4
-0.3
-0.2
-0.1
0.0
0.10 25 50 75 100 125 150 175 200 225
Time (min)
µca
l/sec
Molar ratio
kcal/m
ole
of in
ject
ant
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5
-6
-4
-2
0
-0.4
-0.3
-0.2
-0.1
0.0
0.10 25 50 75 100 125 150 175 200 225
Time (min)
µca
l/sec
Molar ratio
kcal/m
ole
of in
ject
ant
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5-6
-4
-2
0
-0.4
-0.3
-0.2
-0.1
0.0
0.10 25 50 75 100 125 150 175 200 225
Time (min)
µca
l/sec
Molar ratio
kcal/m
ole
of in
ject
ant
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0-6
-4
-2
0
2
-0.6
-0.5
-0.4
-0.3
-0.2
-0.1
0.0
0.10 25 50 75 100 125 150 175 200 225
Time (min)
µca
l/sec
Molar ratio
kcal/m
ole
of in
ject
ant
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0-6
-4
-2
0
-0.7
-0.6
-0.5
-0.4
-0.3
-0.2
-0.1
0.0
0.10 25 50 75 100 125 150 175 200 225
Time (min)
µca
l/sec
Molar ratio
kcal/m
ole
of in
ject
ant

Figure 11 A model for the interaction of pdNAD-MDH with the heteromeric FtsH12-FtsHi AAA-
ATPase complex at the chloroplast inner envelope membrane, which plays an essential role in
chloroplast development.
Ycf2
pdNAD-
MDH
stroma
putative Zinc
binding domain
ATPase
domain
inner chloroplast membrane
outer chloroplast membrane
pdNAD-
MDH
Chloroplast
development
Prolamellar body
formation
FtsH12/FtsHi1/FtsHi2/
FtsHi4/FtsHi5 complex

Parsed CitationsBahl, J., Francke, B., and Monéger, R. (1976). Lipid composition of envelopes, prolamellar bodies and other plastid membranes inetiolated, green and greening wheat leaves. Planta 129: 193–201.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Beeler, S., Liu, H.-C., Stadler, M., Schreier, T., Eicke, S., Lue, W.-L., Truernit, E., Zeeman, S.C., Chen, J., and Kotting, O. (2014).Plastidial NAD-Dependent Malate Dehydrogenase Is Critical for Embryo Development and Heterotrophic Metabolism in Arabidopsis.Plant Physiol. 164: 1175–1190.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Berkemeyer, M., Scheibe, R., and Ocheretina, O. (1998). A novel, non-redox-regulated NAD-dependent malate dehydrogenase fromchloroplasts of Arabidopsis thaliana L. J. Biol. Chem. 273: 27927–27933.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Birktoft, J.J. and Banaszak, L.J. (1983). The presence of a histidine-aspartic acid pair in the active site of 2-hydroxyaciddehydrogenases. X-ray refinement of cytoplasmic malate dehydrogenase. J. Biol. Chem. 258: 472–482.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bodi, Z., Zhong, S., Mehra, S., Song, J., Graham, N., Li, H., May, S., and Fray, R.G. (2012). Adenosine Methylation in Arabidopsis mRNA isAssociated with the 3′ End and Reduced Levels Cause Developmental Defects. Front. Plant Sci. 3: 1–10.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Botelho Calado, S., Tatsuta, T., Von Heijne, G., and Kim, H. (2013). Dislocation by the m-AAA protease increases the thresholdhydrophobicity for retention of transmembrane helices in the inner membrane of yeast mitochondria. J. Biol. Chem. 288: 4792–4798.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Breuninger, H., Rikirsch, E., Hermann, M., Ueda, M., and Laux, T. (2008). Differential Expression of WOX Genes Mediates Apical-BasalAxis Formation in the Arabidopsis Embryo. Dev. Cell 14: 867–876.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bross, C.D., Howes, T.R., Rad, S.A., Kljakic, O., and Kohalmi, S.E. (2017). Subcellular localization of Arabidopsis arogenatedehydratases suggests novel and non-enzymatic roles. J. Exp. Bot. 68: 1425–1440.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Candela, H., Pérez-Pérez, J.M., and Micol, J.L. (2011). Uncovering the post-embryonic functions of gametophytic- and embryonic-lethalgenes. Trends Plant Sci. 16: 336–345.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Cheminant, S., Wild, M., Bouvier, F., Pelletier, S., Renou, J.-P., Erhardt, M., Hayes, S., Terry, M.J., Genschik, P., and Achard, P. (2011).DELLAs Regulate Chlorophyll and Carotenoid Biosynthesis to Prevent Photooxidative Damage during Seedling Deetiolation inArabidopsis. Plant Cell 23: 1849–1860.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Clarke, A.R., Wigley, D.B., Chia, W.N., Barstow, D., Atkinson, T., and Holbrook, J.J. (1986). Site-directed mutagenesis reveals role ofmobile arginine residue in lactate dehydrogenase catalysis. Nature 324: 699–702.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Comparot-Moss, S., Kötting, O., Stettler, M., Edner, C., Graf, A., Weise, S.E., Streb, S., Lue, W.-L., MacLean, D., Mahlow, S., Ritte, G.,Steup, M., Chen, J., Zeeman, S.C., and Smith A.M. (2010). A Putative Phosphatase, LSF1, Is Required for Normal Starch Turnover inArabidopsis Leaves. Plant Physiol. 152: 685–697.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Copley, S.D. (2015). An Evolutionary Perspective on Protein Moonlighting. Biochem Soc Trans. 42: 1684–1691.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Copley, S.D. (2003). Enzymes with extra talents: Moonlighting functions and catalytic promiscuity. Curr. Opin. Chem. Biol. 7: 265–272.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Cvetić, T., Veljović-Jovanović, S., and Vučinić, Ž (2008) Characterization of NAD-dependent malate dehydrogenases from spinach

leaves, Protoplasma 232:247-253.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Demishtein-Zohary, K. and Azem, A. (2017). The TIM23 mitochondrial protein import complex: function and dysfunction. Cell TissueRes. 367: 33–41.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Deng, X.W., Caspar, T., and Quail, P.H. (1991). Cop1 : a regulatory locus involved m hght-controlled development and gene expressionin Arabidopsis. Genes Dev. 5: 1172–1182.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Despres, B., Delseny, M., and Devic, M. (2001). Partial complementation of embryo defective mutations: A general strategy to elucidategene function. Plant J. 27: 149–159.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Drescher, A., Stephanie, R., Calsa, T., Carrer, H., and Bock, R. (2000). The two largest chloroplast genome-encoded open readingframes of higher plants are essential genes. Plant J. 22: 97–104.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Engdahl, S., Aronsson, H., Sundqvist, C., Timko, M.P., and Dahlin, C. (2001). Association of the NADPH:protochlorophyllideoxidoreductase (POR) with isolated etioplast inner membranes from wheat. Plant J. 27: 297–304.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Emanuelsson, O., Nielsen, H., Brunak, S., and Von Heijne, G. (2000). Predicting subcellular localization of proteins based on their N-terminal amino acid sequence. J. Mol. Biol. 300: 1005–1016.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Feng, J., Fan, P., Jiang, P., Lv, S., Chen, X., and Li, Y. (2014). Chloroplast-targeted Hsp90 plays essential roles in plastid developmentand embryogenesis in Arabidopsis possibly linking with VIPP1. Physiol. Plant. 150: 292–307.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ferro, M. et al. (2010). AT_CHLORO, a Comprehensive Chloroplast Proteome Database with Subplastidial Localization and CuratedInformation on Envelope Proteins. Mol. Cell. Proteomics 9: 1063–1084.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Franzmann, L.H., Yoon, E.S., and Meinke, D.W. (1995). Saturating the Genetic-Map of Arabidopsis thaliana with Embryonic Mutations.Plant J. 7: 341–350.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fujii, S., Kobayashi, K., Nagata, N., Masuda, T., and Wada, H. (2017). Monogalactosyldiacylglycerol Facilitates Synthesis of PhotoactiveProtochlorophyllide in Etioplasts. Plant Physiol. 174: 2183–2198.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fulton, D.C., Stettler, M., Mettler, T., Vauhjan, C.K., Li, J., Francisco, P., Gil, M., Reinhold, H., Eicke, S., Messerli, G., Dorken, G.,Halliday, K., Smith, A.M., Smith, S.M., and Zeeman, S.C. (2008). ß-AMYLASE4, a Noncatalytic Protein Required for Starch Breakdown,Acts Upstream of Three Active ß-Amylases in Arabidopsis Chloroplasts. Plant Cell Online 20: 1040–1058.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fusari, C.M., Kooke, R., Lauxmann, M.A., Annunziata, M.G., Encke, B., Hoehne, M., Krohn, N., Becker, F.F.M., Schlereth, A., Sulpice, R.,Stitt, M., and Keurentjes, J.J.B. (2017). Genome-wide Association Mapping Reveals that Specific and Pleiotropic RegulatoryMechanisms Fine-tune Central Metabolism and Growth in Arabidopsis. Plant Cell: tpc.00232.2017.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gómez, L.D., Gilday, A., Feil, R., Lunn, J.E., and Graham, I.A. (2010). AtTPS1-mediated trehalose 6-phosphate synthesis is essential forembryogenic and vegetative growth and responsiveness to ABA in germinating seeds and stomatal guard cells. Plant J. 64: 1–13.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Goward, C.R. and Nicholls, D.J. (1994). Malate dehydrogenase: A model for structure, evolution, and catalysis. Protein Sci. 3: 1883–1888.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

Google Scholar: Author Only Title Only Author and Title
Grau, U.M., Trommer, W.E., and Rossmann, M.G. (1981). Structure of the active ternary complex of pig heart lactate dehydrogenasewith S-lac-NAD at 2.7 Å resolution. J. Mol. Biol. 151: 289–307.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Grefen, C., Donald, N., Hashimoto, K., Kudla, J., Schumacher, K., and Blatt, M.R. (2010). A ubiquitin-10 promoter-based vector set forfluorescent protein tagging facilitates temporal stability and native protein distribution in transient and stable expression studies.Plant J. 64: 355–365.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gujas, B., Cruz, T.M.D., Kastanaki, E., Vermeer, J.E.M., Munnik, T., and Rodriguez-Villalon, A. (2017). Perturbing phosphoinositidehomeostasis oppositely affects vascular differentiation in Arabidopsis thaliana roots. Development: dev.155788.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hatch, M.D. and Osmond, C.B. (1976). Compartmentation and Transport in C4 Photosynthesis. In Stocking CR, Heber U, eds. Transportin plants III, Vol. 3. Berlin:Springer, pp. 144-184.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
He, H., Lee, M.-C., Zheng, L.-L., Zheng, L., and Luo, Y. (2013). Integration of the metabolic/redox state, histone gene switching, DNAreplication and S-phase progression by moonlighting metabolic enzymes. Biosci. Rep. 33: e00018.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hebbelmann, I., Selinski, J., Wehmeyer, C., Goss, T., Voss, I., Mulo, P., Kangasjärvi, S., Aro, E.-M., Oelze, M.-L., Dietz, K.-J., Nunes-Nesi,A., Do, P.T., Fernie, A.R., Talla, S.K., Raghavendra, A.S., Linke, V., and Scheibe, R. (2012). Multiple strategies to prevent oxidativestress in Arabidopsis plants lacking the malate valve enzyme NADP-malate dehydrogenase. J. Exp. Bot. 63: 1445–1459.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Heber, U. (1974). Metabolite exchange between chloroplasts and cytoplasm. Annu. Rev. Plant Physiol. 25: 393–421.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hendriks, W., Mulders, J.W., Bibby, M. a, Slingsby, C., Bloemendal, H., and de Jong, W.W. (1988). Duck lens epsilon-crystallin andlactate dehydrogenase B4 are identical: a single-copy gene product with two distinct functions. Proc. Natl. Acad. Sci. U. S. A. 85: 7114–8.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Heyno, E., Innocenti, G., Lemaire, S.D., Issakidis-Bourguet, E., and Krieger-Liszkay, A. (2014). Putative role of the malate valve enzymeNADP-malate dehydrogenase in H2O2 signalling in Arabidopsis. Philos. Trans. R. Soc. B Biol. Sci. 369: 20130228–20130228.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Horrer, D., Flütsch, S., Pazmino, D., Matthews, J.S.A., Thalmann, M., Nigro, A., Leonhardt, N., Lawson, T., and Santelia, D. (2016). Bluelight induces a distinct starch degradation pathway in guard cells for stomatal opening. Curr. Biol. 26: 362–370.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Huberts, D.H.E.W. and van der Klei, I.J. (2010). Moonlighting proteins: An intriguing mode of multitasking. Biochim. Biophys. Acta - Mol.Cell Res. 1803: 520–525.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jang, J.C. (1997). Hexokinase as a Sugar Sensor in Higher Plants. Plant Cell Online 9: 5–19.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jarvis, P., Dormann, P., Peto, C.A., Lutes, J., Benning, C., and Chory, J. (2000). Galactolipid deficiency and abnormal chloroplastdevelopment in the Arabidopsis MGD synthase 1 mutant. Proc. Natl. Acad. Sci. 97: 8175–8179.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jeffery, C.J. (1999). Moonlighting proteins. Trends Biochem. Sci. 24: 8–11.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jeffery, C.J. (2003). Moonlighting proteins: Old proteins learning new tricks. Trends Genet. 19: 415–417.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

Journet, E.P., Neuburger, M., and Douce, R. (1981). Role of glutamate-oxaloacetate transaminase and malate dehydrogenase in theregeneration of NAD for glycine oxidation by spinach leaf mitochondria. Plant Physiol. 67: 467–469.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Karimi, M., Inze, D., and Depicker, A. (2002). GATEWAY™ vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 7:193–195.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kinoshita, H., Nagasaki, J., Yoshikawa, N., Yamamoto, A., Takito, S., Kawasaki, M., Sugiyama, T., Miyake, H., Weber, A.P.M., andTaniguchi, M. (2011). The chloroplastic 2-oxoglutarate/malate transporter has dual function as the malate valve and in carbon/nitrogenmetabolism. Plant J. 65: 15–26.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Klement, H., Helfrich, M., Oster, U., Schoch, S., and Rüdiger, W. (1999). Pigment-free NADPH:protochlorophyllide oxidoreductase fromAvena sativa L. Purification and substrate specificity. Eur. J. Biochem. 265: 862–874.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kobayashi, K., Kondo, M., Fukuda, H., Nishimura, M., and Ohta, H. (2007). Galactolipid synthesis in chloroplast inner envelope isessential for proper thylakoid biogenesis, photosynthesis, and embryogenesis. Proc. Natl. Acad. Sci. 104: 17216–17221.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Langklotz, S., Baumann, U., and Narberhaus, F. (2012). Structure and function of the bacterial AAA protease FtsH. Biochim. Biophys.Acta - Mol. Cell Res. 1823: 40–48.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lebedev, N., Van Cleve, B., Armstrong, G., and Apel, K. (1995). Chlorophyll Synthesis in a Deetiolated (det340) Mutant of Arabidopsiswithout NADPH-Protochlorophyllide (PChlide) Oxidoreductase (POR) A and Photoactive PChlide-F655. Plant Cell 7: 2081–2090.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lee, C., Schwartz, M.P., Prakash, S., Iwakura, M., and Matouschek, A. (2001). ATP-dependent proteases degrade their substrates byprocessively unraveling them from the degradation signal. Mol. Cell 7: 627–637.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lee, Y., Rubio, M.C., Alassimone, J., and Geldner, N. (2013). A mechanism for localized lignin deposition in the endodermis. Cell 153:402–412.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lu, X., Zhang, D., Li, S., Su, Y., Liang, Q., Meng, H., Shen, S., Fan, Y., Liu, C., and Zhang, C. (2014). FtsHi4 is essential for embryogenesisdue to its influence on chloroplast development in Arabidopsis. PLoS One 9.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Majsec, K., Bhuiyan, N.H., Sun, Q., Kumari, S., Kumar, V., Ware, D., and van Wijk, K. (2017). The Plastid and Mitochondrial PeptidaseNetwork in Arabidopsis: A Foundation for Testing Genetic Interactions and Functions in Organellar Proteostasis. Plant Cell 29: 2687-2710.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mansfield, S.G. and Briarty, L.G. (1991). Early Embryogenesis in Arabidopsis thaliana .II. the Developing Embryo. Can. J. Bot. Can. Bot.69: 461–476.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Martin, A., Baker, T.A., and Sauer, R.T. (2005). Rebuilt AAA + motors reveal operating principles for ATP-fuelled machines. Nature 437:1115–1120.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mascia, P.N. (1978). Studies of Chloroplast Development in Four Maize Mutants Defective in Chlorophyll Biosynthesis. Planta. 143:207–211.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Minárik, P., Tomaásková, N., Kollárová, M., and Antalík, M. (2002). Malate Dehydrogenases - Structure and function. Gen. Physiol.Biophys. 21: 257–265.

Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Moore, B. (2003). Role of the Arabidopsis Glucose Sensor HXK1 in Nutrient, Light, and Hormonal Signaling. Science 300: 332–336.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Moore, B.D. (2004). Bifunctional and moonlighting enzymes: Lighting the way to regulatory control. Trends Plant Sci. 9: 221–228.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Musrati, R.A., Kollárová, M., Mernik, N., and Mikulášová, D. (1998). Malate Dehydrogenase: Distribution, Function and Properties. Gen.Physiol. Biophys. 17: 193–210.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nakai, M. (2018). New perspectives on chloroplast protein import. Plant and Cell Physiology, pcy083, doi.org/10.1093/pcp/pcy083.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nicholls, D.J., Miller, J., Scawen, M.D., Clarke, A.R., John Holbrook, J., Atkinson, T., and Goward, C.R. (1992). The importance ofarginine 102 for the substrate specificity of Escherichia coli malate dehydrogenase. Biochem. Biophys. Res. Commun. 189: 1057–1062.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nickelsen, J. (2005). The Green Alga Chlamydomonas reinhardtii – A Genetic Model Organism. In Progress in Botany (Heidelberg,Springer-Verlag Berlin), pp. 68–89.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nielsen, H. and Engelbrecht, J. (1997). Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavagesites Artificial neural networks have been used for many biological. Protein Eng. 10: 1–6.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ossowski, S., Schwab, R., and Weigel, D. (2008). Gene silencing in plants using artificial microRNAs and other small RNAs. Plant J. 53:674–690.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Park, H., Kreunen, S.S., Cuttriss, A.J., DellaPenna, D., and Pogson, B.J. (2002). Identification of the Carotenoid Isomerase ProvidesInsight into Carotenoid Biosynthesis, Prolamellar Body Formation, and Photomorphogenesis. Plant Cell 14: 321–332.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Patton, D.A., Franzmann, L.H., and Meinke, D.W. (1991). Mapping genes essential for embryo development in Arabidopsis thaliana. Mol.Gen. Genet. 227: 337–347.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Patton, D.A., Schetter, A.L., Franzmann, L.H., Nelson, K., Ward, E.R., and Meinke, D.W. (1998). An Embryo-Defective Mutant ofArabidopsis Disrupted in the Final Step of Biotin Synthesis. Plant Physiol. 116: 935–946.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pracharoenwattana, I., Cornah, J.E., and Smith, S.M. (2007). Arabidopsis peroxisomal malate dehydrogenase functions in beta-oxidationbut not in the glyoxylate cycle. Plant J. 50: 381–90.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pracharoenwattana, I., Zhou, W., and Smith, S.M. (2010). Fatty acid beta-oxidation in germinating Arabidopsis seeds is supported byperoxisomal hydroxypyruvate reductase when malate dehydrogenase is absent. Plant Mol. Biol. 72: 101–109.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ruppel, N.J. and Hangarter, R.P. (2007). Mutations in a plastid-localized elongation factor G alter early stages of plastid development inArabidopsis thaliana. BMC Plant Biol. 7: 37.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ryberg, M. and Sundqvist, C. (1982). Characterization of prolamellar bodies and prothylakoids fractionated from wheat etioplasts.Physiol. Plant. 56: 125–132.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

Ryberg, M. and Sundqvist, C. (1988). The regular ultrastructure of isolated prolamellar bodies depends on the presence of membrane‐bound NADPH‐protochlorophyllide oxidoreductase. Physiol. Plant. 73: 218–226.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Scheibe, R. (2004). Malate valves to balance cellular energy supply. Physiol. Plant. 120: 21–26.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Scheibe, R. (1987). NADP+‐malate dehydrogenase in C3‐plants: Regulation and role of a light‐activated enzyme. Physiol. Plant. 71: 393–400.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Selinski, J., König, N., Wellmeyer, B., Hanke, G.T., Linke, V., Ekkehard Neuhaus, H., and Scheibe, R. (2014). The plastid-localized NAD-dependent malate dehydrogenase is crucial for energy homeostasis in developing Arabidopsis thaliana seeds. Mol. Plant 7: 170–186.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Selstam, E., Schelin, J., Brain, T., and Williams, W.P. (2002). The effects of low pH on the properties of protochlorophyllideoxidoreductase and the organization of prolamellar bodies of maize (Zea mays). Eur. J. Biochem. 269: 2336–2346.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Seung, D., Thalmann, M., Sparla, F., Abou Hachem, M., Lee, S.K., Issakidis-Bourguet, E., Svensson, B., Zeeman, S.C., and Santelia, D.(2013). Arabidopsis thaliana AMY3 is a unique redox-regulated chloroplastic α-amylase. J. Biol. Chem. 288: 33620–33633.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Seung D., Soyk, S., Coiro, M., Maier, B.A., Eicke, S., and Zeeman, S.C. (2015). PROTEIN TARGETING TO STARCH is required forlocalising GRANULE-BOUND STARCH SYNTHASE to starch granules and for normal amylose synthesis in Arabidopsis. PLoS Biol.13:e1002080.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sew, Y.S., Ströher, E., Fenske, R., and Millar, A.H. (2016). Loss of Mitochondrial Malate Dehydrogenase Activity Alters Seed MetabolismImpairing Seed Maturation and Post-Germination Growth in Arabidopsis. Plant Physiol. 171: 849–63.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Silverstein, E. and Sulebele, G. (1969). Catalytic mechanism of pig heart mitochondrial malate dehydrogenase studied by kinetics atequilibrium. Biochemistry 8: 2543–50.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sokolenko, A., Pojidaeva, E., Zinchenko, V., Panichkin, V., Glaser, V.M., Herrmann, R.G., and Shestakov, S. V (2002). The genecomplement for proteolysis in the cyanobacterium Synechocystis sp. PCC 6803 and Arabidopsis thaliana chloroplasts. Curr. Genet. 41:291–310.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Solymosi, K. and Aronsson, H. (2013). Etioplasts and their significance in chloroplast biogensis 36: 155-167.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Solymosi, K. and Schoefs, B. (2010). Etioplast and etio-chloroplast formation under natural conditions: The dark side of chlorophyllbiosynthesis in angiosperms. Photosynth. Res. 105: 143–166.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sperling, U., Franck, F., van Cleve, B., Frick, G., Apel, K., and Armstrong, G.A. (1998). Etioplast differentiation in arabidopsis: bothPORA and PORB restore the prolamellar body and photoactive protochlorophyllide-F655 to the cop1 photomorphogenic mutant. PlantCell 10: 283–296.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Spicher, L., Almeida, J., Gutbrod, K., Pipitone, R., Dörmann, P., Glauser, G., Rossi, M., and Kessler, F. (2017). Essential role for phytolkinase and tocopherol in tolerance to combined light and temperature stress in tomato. J. Exp. Bot. 68: 5845–5856.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Taniguchi, M. and Miyake, H. (2012). Redox-shuttling between chloroplast and cytosol: Integration of intra-chloroplast and extra-chloroplast metabolism. Curr. Opin. Plant Biol. 15: 252–260.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

Google Scholar: Author Only Title Only Author and Title
Tatsuta, T., Augustin, S., Nolden, M., Friedrichs, B., and Langer, T. (2007). m-AAA protease-driven membrane dislocation allowsintramembrane cleavage by rhomboid in mitochondria. EMBO J. 26: 325–335.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tomaz, T., Bagard, M., Pracharoenwattana, I., Linden, P., Lee, C.P., Carroll, A.J., Stroher, E., Smith, S.M., Gardestrom, P., and Millar,A.H. (2010). Mitochondrial Malate Dehydrogenase Lowers Leaf Respiration and Alters Photorespiration and Plant Growth inArabidopsis. Plant Physiol. 154: 1143–1157.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tschopp, M.-A., Iki, T., Brosnan, C.A., Jullien, P.E., and Pumplin, N. (2017). A complex of Arabidopsis DRB proteins can impair dsRNAprocessing. RNA 23: 782–797.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wagner, R., Aigner, H., and Funk, C. (2012). FtsH proteases located in the plant chloroplast. Physiol. Plant. 145: 203–214.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wigley, D.B., Gamblin, S.J., Turkenburg, J.P., Dodson, E.J., Piontek, K., Muirhead, H., and Holbrook, J.J. (1992). Structure of a ternarycomplex of an allosteric lactate dehydrogenase from Bacillus stearothermophilus at 2.5 A resolution. J. Mol. Biol. 223: 317–35.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wistow, G.J., Mulders, J.W., and de Jong, W.W. (1987). The enzyme lactate dehydrogenase as a structural protein in avian andcrocodilian lenses. Nature 326: 622–4.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yang, S.S. and Zhai, Q.H. (2017). Cytosolic GAPDH: a key mediator in redox signal transduction in plants. Biol. Plant. 61: 417–426.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yu, B., Wakao, S., Fan, J., and Benning, C. (2004). Loss of plastidic lysophosphatidic acid acyltransferase causes embryo-lethality inArabidopsis. Plant Cell Physiol. 45: 503–510.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zaffagnini, M., Fermani, S., Costa, A., Lemaire, S.D., and Trost, P. (2013). Plant cytoplasmic GAPDH: redox post-translationalmodifications and moonlighting properties. Front. Plant Sci. 4: 1–18.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang, X., Henriques, R., Lin, S.-S., Niu, Q.-W., and Chua, N.-H. (2006). Agrobacterium-mediated transformation of Arabidopsis thalianausing the floral dip method. Nat. Protoc. 1: 641–6.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhao, Y., Luo, L., Xu, J., Xin, P., Guo, H., Wu, J., Bai, L., Wang, G., Chu, J., Zuo, J., Hong, Y., Huang, X., and Li, J. (2018). Malatetransported from chloroplast to mitochondrion triggers production of ROS and PCD in Arabidopsis thaliana. Cell Research 28 (4): 448-461.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

DOI 10.1105/tpc.18.00121; originally published online June 22, 2018;Plant Cell
Oliver KöttingDaniele Albertini, Benjamin A Maier, Felix Kessler, Stefan Hörtensteiner, Samuel C. Zeeman and
Tina B Schreier, Cléry Antoine, Michael Schläfli, Florian Galbier, Martha Stadler, Emilie Demarsy,Chloroplast Development Through its Interaction with an FtsH12-FtsHi Protease Complex
Plastidial NAD-Dependent Malate Dehydrogenase: A Moonlighting Protein Involved in Early
This information is current as of September 17, 2018
Supplemental Data /content/suppl/2018/06/27/tpc.18.00121.DC2.html /content/suppl/2018/06/21/tpc.18.00121.DC1.html
Permissions X
https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists