oxidative phosphorylation
DESCRIPTION
This complex mechanical art form can be viewed as a metaphor for the molecular apparatus underlying electron transport and ATP synthesis by oxidative phosphorylation. OXIDATIVE PHOSPHORYLATION. A PROTON GRADIENT POWERS THE SYNTHESIS OF ATP. - PowerPoint PPT PresentationTRANSCRIPT

OXIDATIVE PHOSPHORYLATIONOXIDATIVE PHOSPHORYLATION
This complex mechanical art form can be viewed as a
metaphor for the molecular apparatus underlying electron
transport and ATP synthesis by oxidative phosphorylation.

A PROTON GRADIENT POWERS THE SYNTHESIS OF ATP
The transport of electrons from NADH or FADH2 to O2 via the electron-transport chain is exergonic process:
NADH + ½O2 + H+ H2O + NAD+
FADH2 + ½O2 H2O + FAD+
Go’ = -52.6 kcal/mol for NADH
-36.3 kcal/mol for FADH2
How this process is coupled to the synthesis of ATP (endergonic process)? ADP + Pi ATP + H2O Go’=+7.3 kcal/mol

•Proposed by Peter Mitchell in the 1960’s (Nobel Prize, 1978)
•Chemiosmotic theory: electron transport and ATP synthesis are coupled by a proton gradient across the inner mitochondrial membrane Mitchell’s postulates for chemiosmotic
theory1. Intact inner mitochondrial membrane is required
2. Electron transport through the ETC generates a proton gradient
3. ATP synthase catalyzes the phosphorylation of ADP in a reaction driven by movement of H+ across the inner membrane into the matrix
The Chemiosmotic Theory
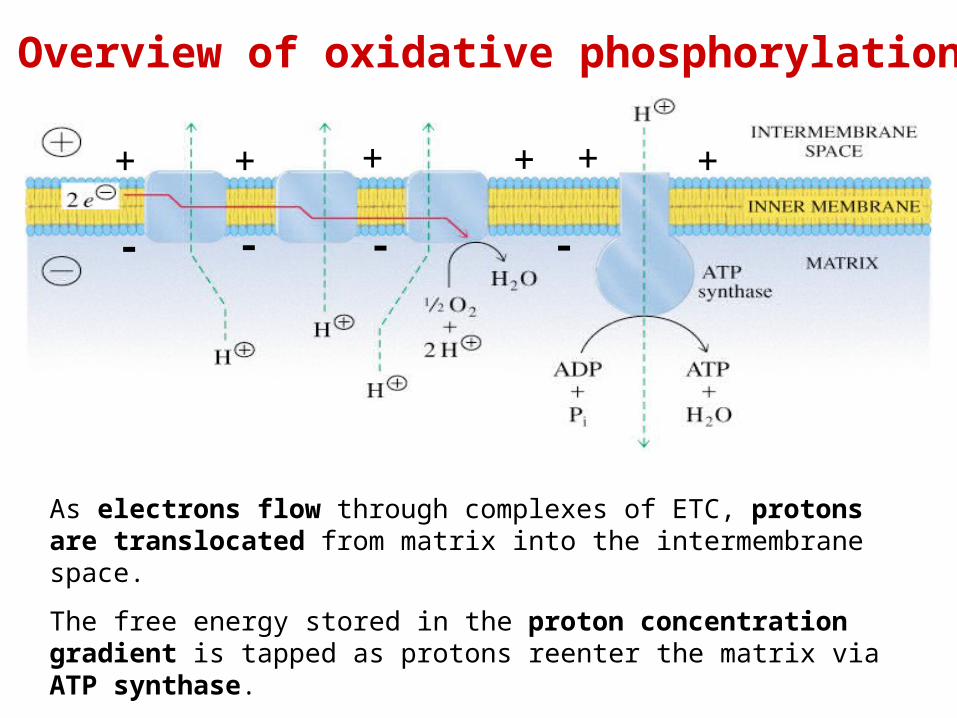
As electrons flow through complexes of ETC, protons are translocated from matrix into the intermembrane space.
The free energy stored in the proton concentration gradient is tapped as protons reenter the matrix via ATP synthase.
As result ATP is formed from ADP and Pi.
++ + ++ +
----
Overview of oxidative phosphorylation
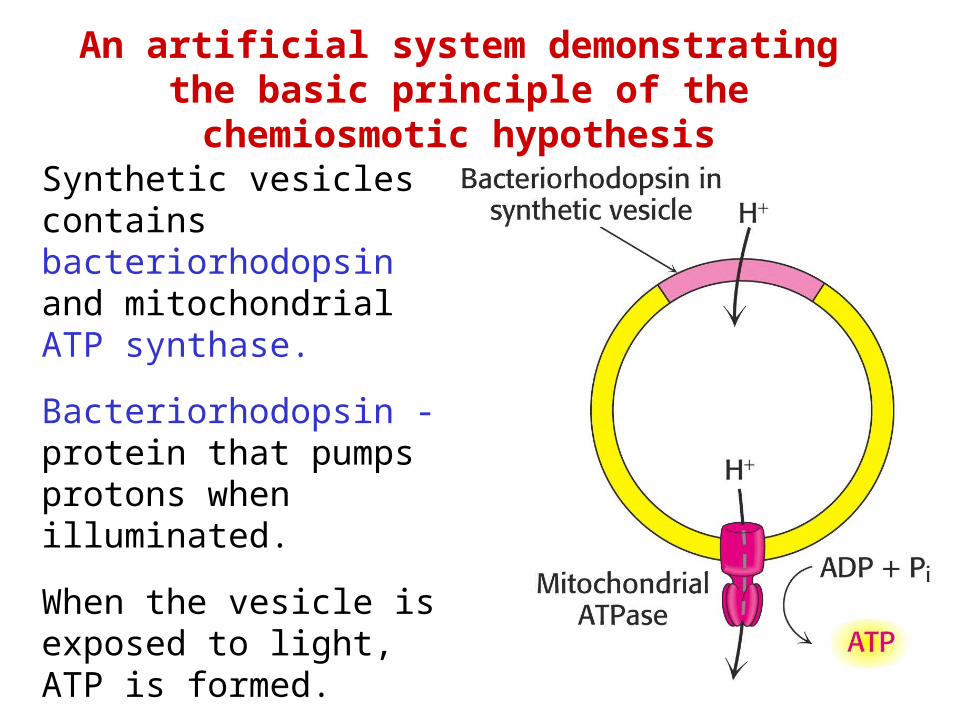
An artificial system demonstrating the basic principle of the chemiosmotic
hypothesisSynthetic vesicles contains bacteriorhodopsin and mitochondrial ATP synthase. Bacteriorhodopsin - protein that pumps protons when illuminated. When the vesicle is exposed to light, ATP is formed.
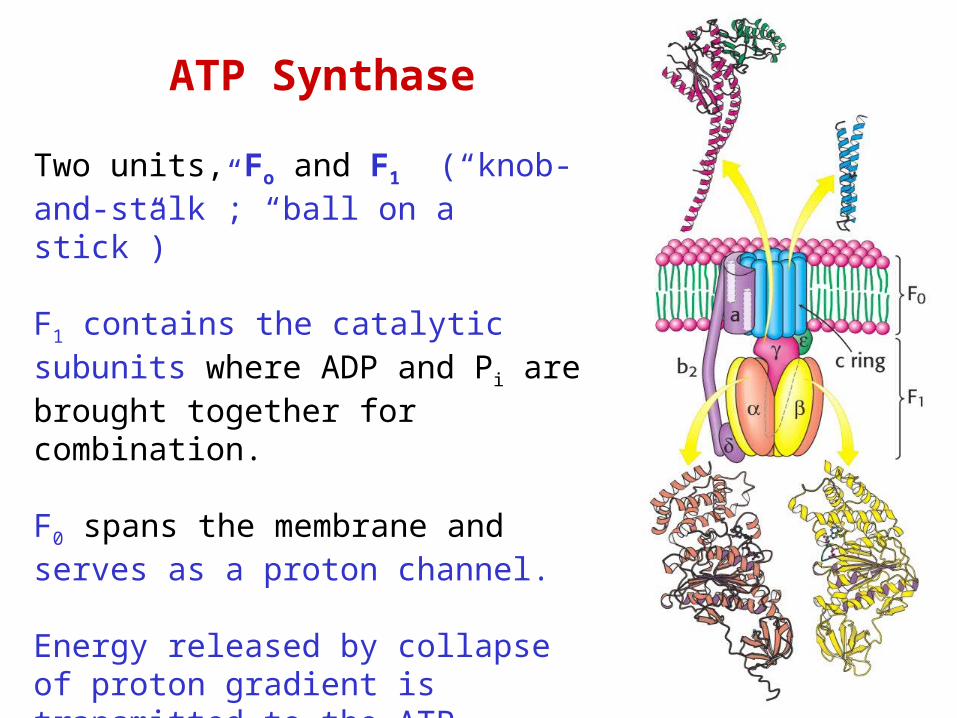
ATP Synthase
Two units, Fo and F1 (“knob-and-stalk”; “ball on a stick”)
F1 contains the catalytic subunits where ADP and Pi are brought together for combination.
F0 spans the membrane and serves as a proton channel.
Energy released by collapse of proton gradient is transmitted to the ATP synthesis.

•F1 contains 5 types of polypeptide chains - 33
•Fo - a1b2c10-14
(c subunits form cylindrical, membrane-bound base)
•Fo and F1 are connected by a stalk and by exterior column (a1b2 and
•The proton channel – between c ring and a subunit.

• there are 3 active sites, one in each subunit
• c- unit forms a “rotor”
• a-b--33 unit is the “stator”
• passage of protons through the Fo channel causes the rotor to spin
• rotation of the subunit inside the 33 hexamer causes domain movements in the -subunits, opening and closing the active sites
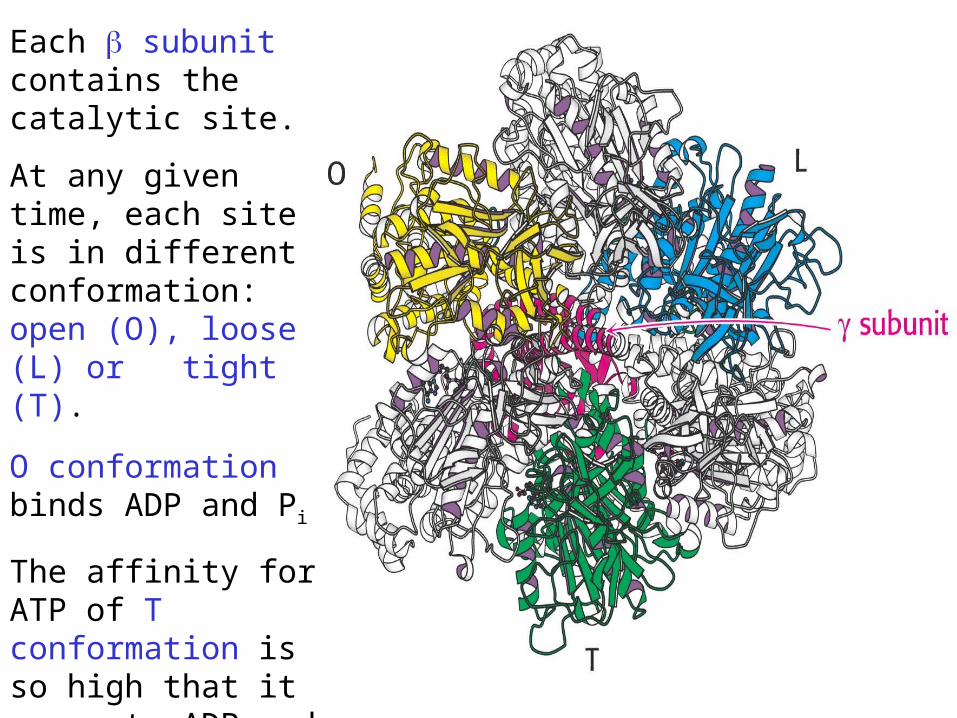
Each subunit contains the catalytic site.
At any given time, each site is in different conformation: open (O), loose (L) or tight (T).
O conformation binds ADP and Pi
The affinity for ATP of T conformation is so high that it converts ADP and Pi into ATP.

1. ADP and Pi bind to an open site
2. Passage of protons causes each of three sites to change conformation.
3. The open conformation (containing the newly bound ADP and Pi) becomes a loose site. The loose site filled with ADP and Pi becomes a tight site. The ATP containing tight site becomes an open site.
4. ATP released from open site, ADP and Pi form ATP in the tight site
Binding-Change Mechanism of ATP Synthase

Experimental observation of ATP synthase rotation
•Fluorescent protein arm (actin) attached to subunits
• subunits bound to a glass plate
•Arm seen rotating when ATP added (observed by microscopy)