oxidative phosphorylation for medical school
TRANSCRIPT


V.S.RAVIKIRAN, MSc., Dept. of Biochemistry, ASRAM Medical college, Eluru-
534005.AP, India

• Food : Food is the source of energy.• Energy : Energy is the capacity to do work.• Bioenergetics : is the quantitative study of
energy transformations that occur within the living cells.
• This follows the laws the thermodynamics.

• Thermodynamics : is a branch of physical sciences that deals with energy changes.
• First Law of Thermodynamics• Stated simply; The total energy of the
universe does not change. This does not mean that
the form of the energy cannot change.
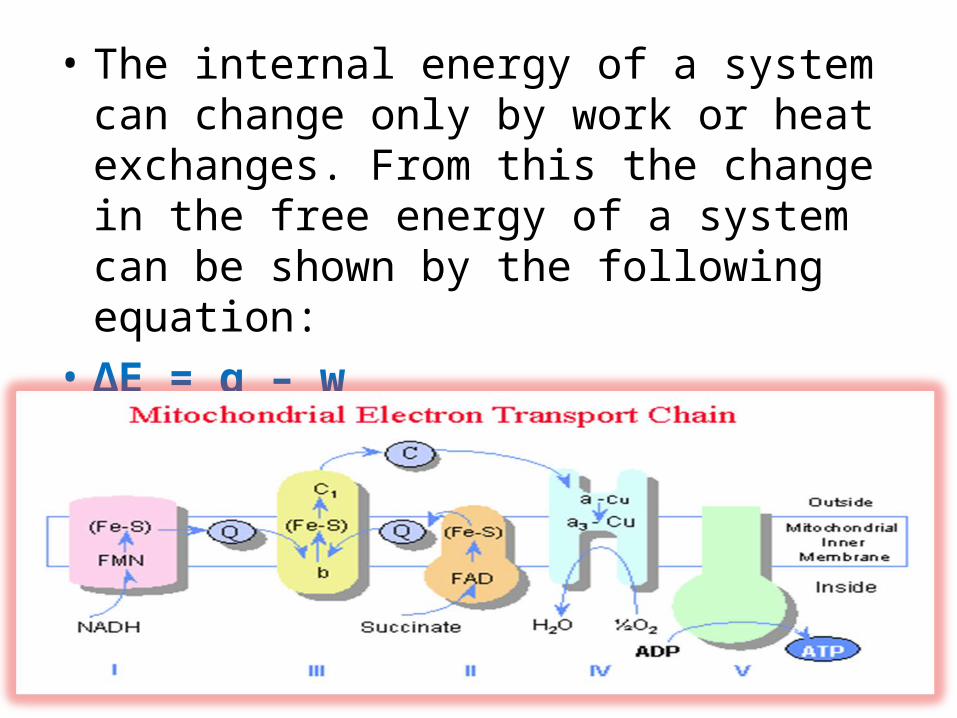
• The internal energy of a system can change only by work or heat exchanges. From this the change in the free energy of a system can be shown by the following equation:
• ΔE = q – w• Where q is the heat absorbed by the system and w is
the work done.

• When q is negative heat has flowed from the system and when q is positive heat has been absorbed by the system.
• Entropy (S): is a measure of the degree of randomness or disorder of a system.
• Enthalpy (H) : is the heat content of a sytem.

• Second Law of Thermodynamics: Entropy• The second law of thermodynamics states that the
universe (i.e. all systems) tend to the greatest degree of randomization.
• For an isothermal reversible reaction the change in entropy can be reduced to the term:
• ΔS = ΔH/T

• ΔS = ΔH/T• In order to effectively evaluate the course
(spontaneity or lack there of) of a reaction and taking into account both the first and second laws of thermodynamics, Josiah Gibbs defined the term, free energy which is defined as:
• ΔG = ΔH – TΔS

• ΔG = ΔH – TΔS• ΔG = change in free energy.• ΔH = change in enthalpy or heat content of
the system.• ΔS = change in entropy.• T = absolute temperature.

• Useful energy is the form of energy that is capable of performing work.- two major types:- (1) Heat energy- (2) Free energy

• (1) Heat energy, which is capable of performing work through change of temperature.
• (2) Free energy which is capable of performing work at constant temperature.
• ΔG = ΔH – TΔS
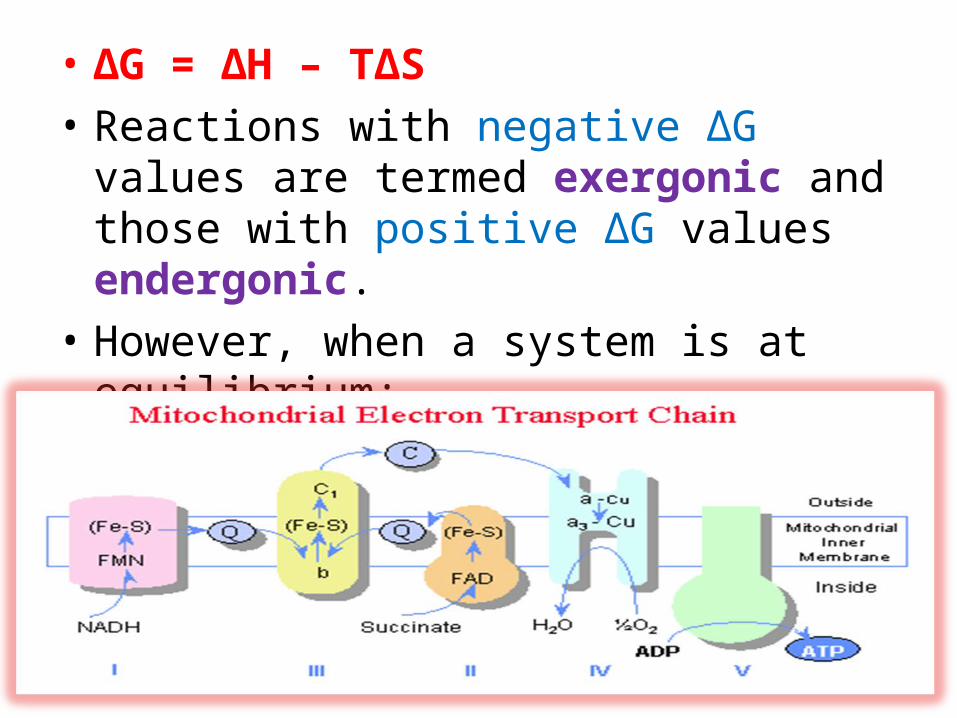
• ΔG = ΔH – TΔS• Reactions with negative ΔG values are termed
exergonic and those with positive ΔG values endergonic.
• However, when a system is at equilibrium:ΔG = 0

• Exergonic reaction :• ATP + H2O → ADP + Pi: ΔGo' = –7.3 kcal/mol
• Endergonic reaction :• ADP + Pi → ATP : ΔGo' = +7.3 kcal/mol

• ΔG = ΔH – TΔS• Exothermic reaction = ΔH negative• Endothermic reaction = ΔH is positive• Iso-thermic reaction = ΔH is zero

• Standard free energy change (ΔGo’)• It is the free energy change under standard
conditions.• The standard conditions are as below:
Temperature of 250C (2980K), Pressure of 1.0 atm (760 mm Hg) pH of 7.0, Reactants and products are all present in their standard concentrations namely 1.0M.

Standard free energies of hydrolysis of some common metabolic intermediates
Metabolite ΔGo’
Phosphoenol pyruvate -14.8 High energy compoundsCarbamoyl phosphate -12.3
1,3-Bisphosphoglycerate -11.8
Acid phosphate -11.2
Creatine phosphate -10.3
Arginine phosphate -7.6
ATP to ADP + Pi -7.3 ΔGo’ = 0ATP to AMP + PPi -7.7 = equilibriumATP to AMP +Pi + Pi -14.6
Glucose 1-phosphate -5.0 Low energy compoundsGlucose 6-phosphate -3.3
Glycerol 1-phosphate -2.2

Phosphoenol pyruvate Pyruvate (ΔGo’ =-14.8 kcal/mol)
Metabolite ΔGo’Phosphoenol pyruvate
-14.8
ΔGo’ = (-) ΔGo’ = (+) ΔGo’ = (0)

ATPATP is the energy carrier for most
biological reactions
ATP + H2O -> ADP + PiATP + H2O -> AMP + PPi

19
Measurement of redox potentialX(oxidized) + X-(reduced) =redox couple
Samplehalf-cell
Standard referencehalf-cell
Electrons, but notX or X-, can flowthrough the agar bridge
If the reaction is,X- + H+ X + 1/2 H2
In half-cells:X- X + e- & H+ + e- 1/2 H2


21
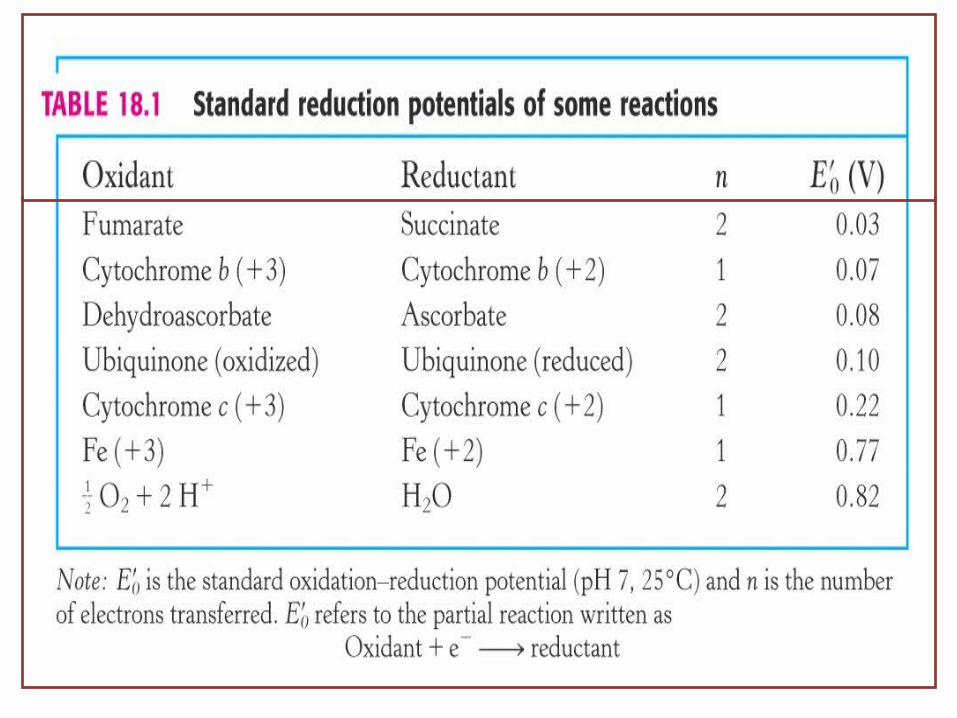
Reduction potentials of biological redox couples
E’o = standard reduction potential
‘prime’ denotes pH 7‘zero’ denotes ‘standard’


Free Energy and Reduction Potential

Number of electrons transferred in the redox reaction Faraday’s constant (96485
J/volt/mole)
Crucial equation: Go' = -nF Eo'
The relative tendency to accept e-s and become reduced.
Reduction Potentials
= Eo'(acceptor) - Eo'(donor)
E0’=standard reduction potential.
If Eo' is positive, an electron transfer reaction is spontaneous (Go' <0)

The free energy of a typical reaction is calculated
directly from its Eo by the Nernst equation as shown ′
below, where n is the number of electrons involved
in the reaction and F is the Faraday constant (23.06
kcal/volt/mol or 94.4 kJ/volt/mol):
ΔG° = –′ nFΔEo′

Order and Reduction Potentials
NAD+
FMN
FeS
ubiquinoneFAD FeS
Cyt b
FeS Cyt c1 Cyt c Cyt a Cyt a3
1/2 O2
ubiquinone
+0.077
+0. 22 +0. 25
+0. 29 +0. 55
-0.32
+0.03
+0.045
+0.82
-0.3

28
ATP from Electron Transport
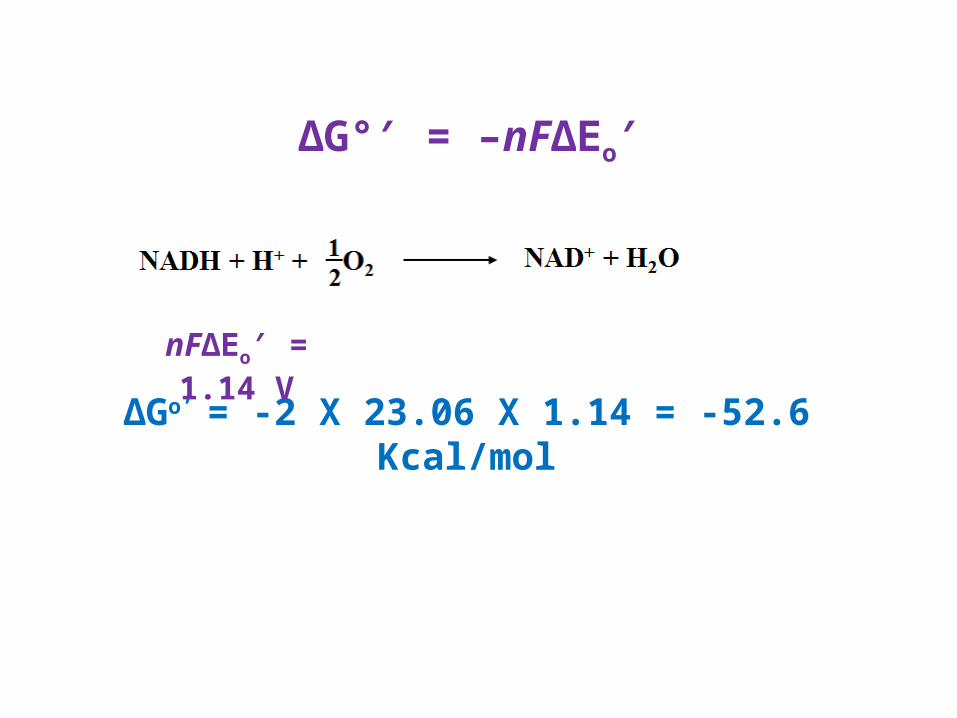
ΔG° = –′ nFΔEo′
∆Go’ = -2 X 23.06 X 1.14 = -52.6 Kcal/mol
nFΔEo = 1.14 V′

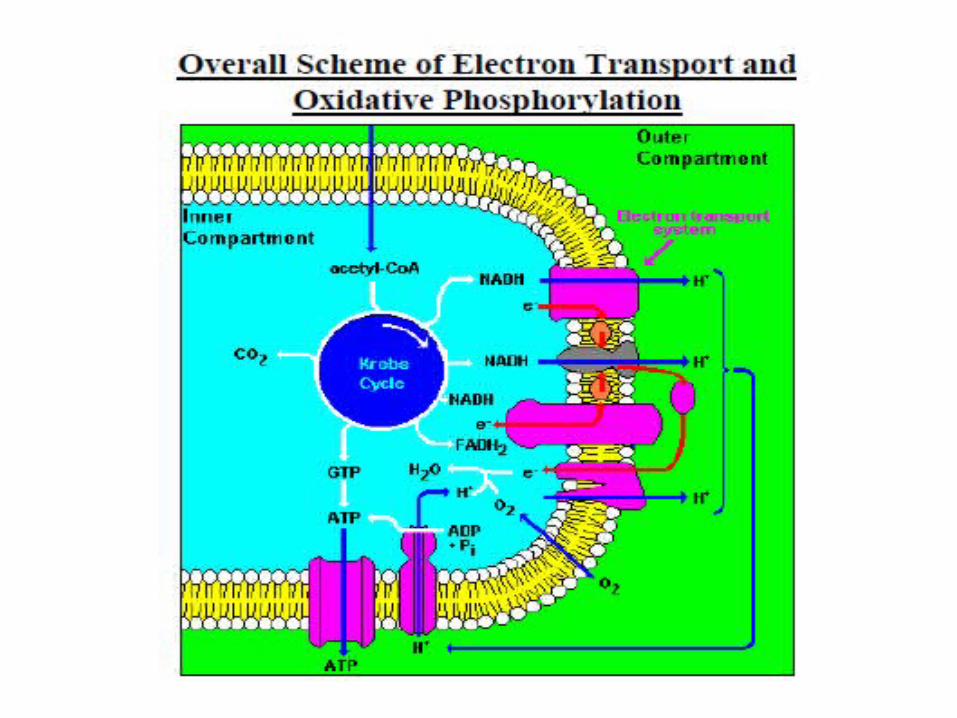
Oxidative Phosphorylation
Oxidative phosphorylation is the process by which the energy stored in NADH and FADH2 is used to produce ATP.
A. Oxidation step: electron transport chain
NADH + H+ + O2NAD+ + H2O
12
FADH2 + O212
FAD + H2O
The free energy available as a consequence of transferring 2 electrons from NADH or succinate to molecular oxygen is –52.6 and –36 kcal/mol, respectively.
–36 kcal/mol
–52.6 kcal/mol

Order and Reduction Potentials
NAD+
FMN
FeS
ubiquinoneFAD FeS
Cyt b
FeS Cyt c1 Cyt c Cyt a Cyt a3
1/2 O2
ubiquinone
+0.077
+0. 22 +0. 25
+0. 29 +0. 55
-0.32
+0.03
+0.045
+0.82
-0.3

Oxidative Phosphorylation
Oxidative phosphorylation is the process by which the energy stored in NADH and FADH2 is used to produce ATP.
A. Oxidation step: electron transport chain
NADH + H+ + O2NAD+ + H2O
12
FADH2 + O212
FAD + H2O
The free energy available as a consequence of transferring 2 electrons from NADH or succinate to molecular oxygen is –52.6 and –36 kcal/mol, respectively.
–36 kcal/mol
–52.6 kcal/mol

Burning glucose in an experiment
Energy released from glucose
(as heat and light)
100%
Energy released from glucose
banked in ATP
“Burning” glucosein cellular respiration
About 40%
Gasoline energy converted to movement
Burning gasolinein an auto engine
25%
How efficient is cell respiration?

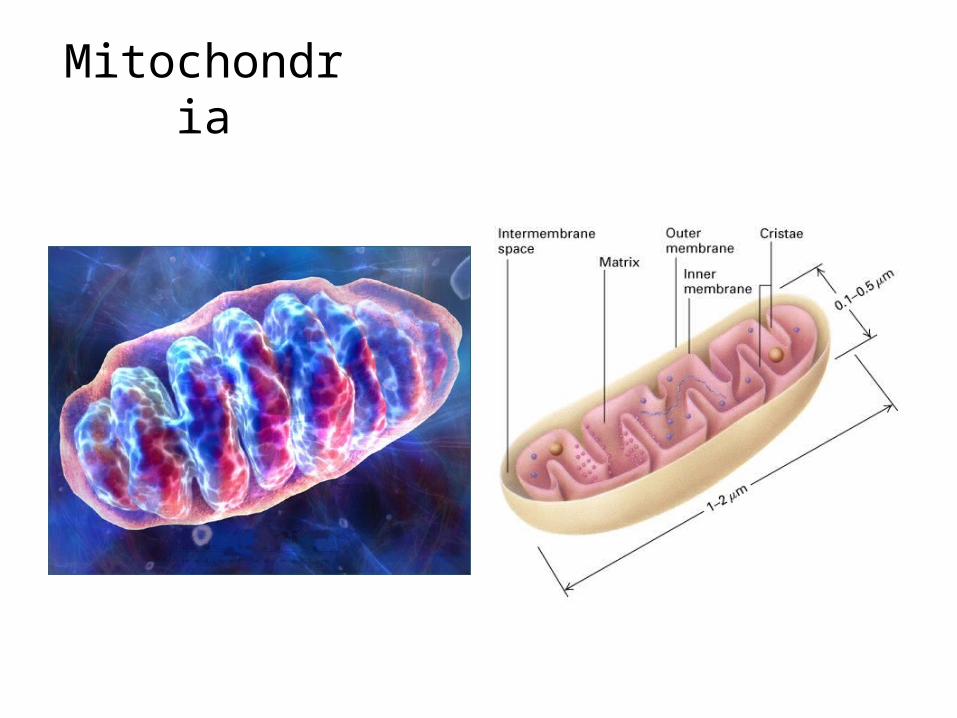
Mitochondria

36

37

Oxidative Phosphorylation
Oxidative phosphorylation is the process by which the energy stored in NADH and FADH2 is used to produce ATP.
A. Oxidation step: electron transport chain
B. Phosphorylation step
NADH + H+ + O2NAD+ + H2O
12
FADH2 + O212
FAD + H2O
ADP + Pi ATP

Electron Carriers
NAD+
FMN
FeS
ubiquinoneFAD FeS
Cyt b
FeS Cyt c1 Cyt c Cyt a Cyt a3
1/2 O2
ubiquinone
NAD+ or FAD
There are 2 sites of entry for electrons into the electron transport chain:
Both are coenzymes for dehydrogenase enzymes
The transfer of electrons is not directly to oxygen but through coenzymes

40






46

Nicotinamide coenzymes: NAD+
N
CONH2
N
CONH2
H HXH2
X
oxidised coenzyme
NAD+ or NADP+reduced coenzymeNADH or NADPH
+ H+
Always a 2-electron reaction transferring 2 e- and 2 H+
48
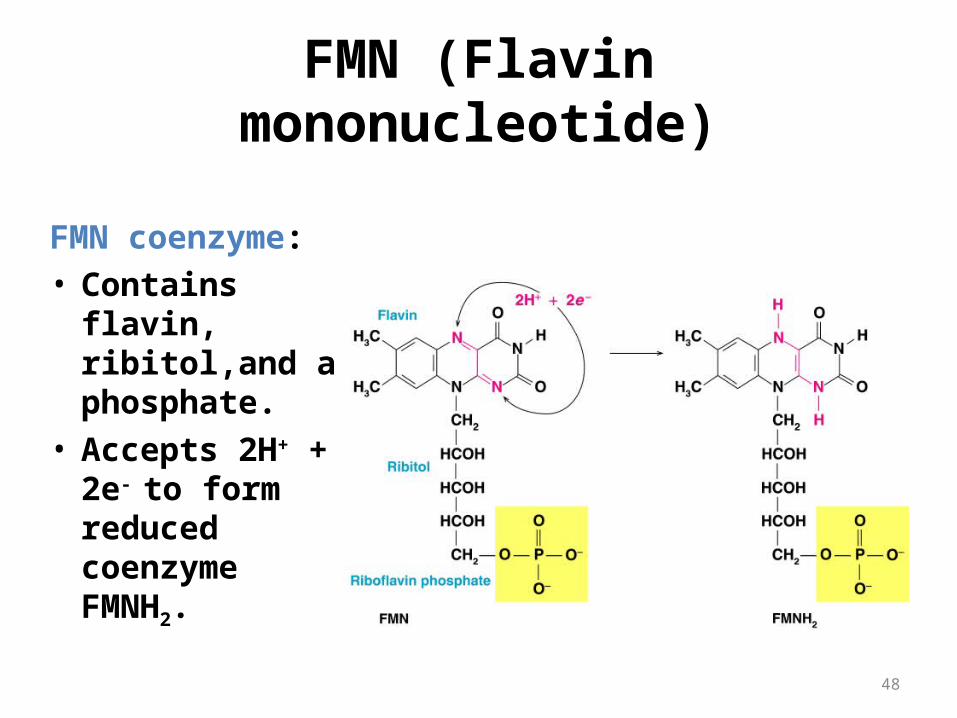
FMN (Flavin mononucleotide)
FMN coenzyme:• Contains flavin,
ribitol,and a phosphate.
• Accepts 2H+ + 2e-
to form reduced coenzyme FMNH2.

The flavin coenzymes / flavoproteins
H3C
H3C NN
N O
O
N
CH2 CH CH CH CH2
OH OH OH
O P OH
OH
O
riboflavin monophosphate(flavin mononucleotide, FMN)
H3C
H3C NN
N O
O
N
CH2 CH CH CH CH2
OH OH OH
O P O P O
OH
O
OH
O
CH2 O
OH OH
N
N
NH2
N
N
flavin adenine dinucleotide (FAD)
FAD Always a 2-electron reaction transferring 2 e- and 2 H+
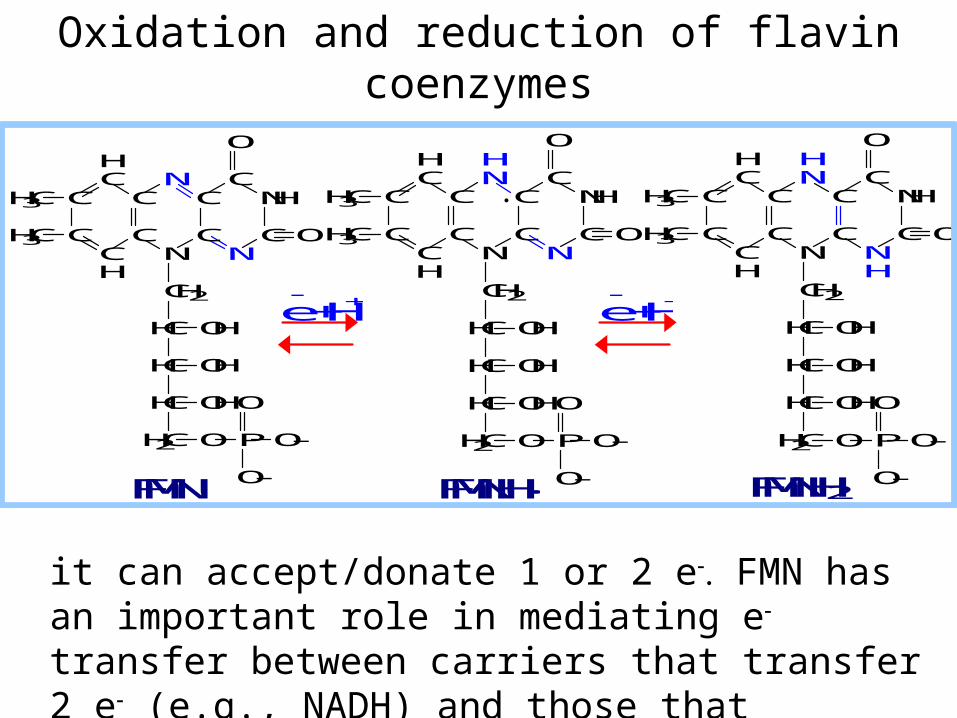
Oxidation and reduction of flavin coenzymes
it can accept/donate 1 or 2 e FMN has an important role in mediating e transfer between carriers that transfer 2 e (e.g., NADH) and those that transfer 1 e (e.g., Fe+++).
C
CCH
C
C
HC
NC
CN
NC
NHC
H3C
H3C
O
O
CH2
HC
HC
HC
H2C
OH
OPO-
O
O-
OH
OH
C
CCH
C
C
HC
NC
C
HN
NC
NHC
H3C
H3C
O
O
CH2
HC
HC
HC
H2C
OH
OPO-
O
O-
OH
OH
C
CCH
C
C
HC
NC
C
HN
NH
C
NHC
H3C
H3C
O
O
CH2
HC
HC
HC
H2C
OH
OPO-
O
O-
OH
OH
e + H+ e
+ H+
FMN FMNH2 FMNH·

Role of FMN mediating between 2e & 1e carriers:
For example, when NADH donates electrons to the respiratory chain, the initial electron transfers are:
NADH + H+ + FMN NAD+ + FMNH2
FMNH2 + Fe+++ FMNH· + Fe++ + H+

52
Iron-Sulfur (Fe-S) Clusters
Fe-S clusters:• Are groups of proteins containing iron ions and
sulfide. • Accept electrons to reduce Fe3+ to Fe2+, and lose
electrons to re-oxidize Fe2+ to Fe3+.

FeFe
S
S
S
Fe
Fe
S
S
S
SS
Cys
Cys
Cys
Cys
S
Fe
S
Fe
S
S
S
S
Cys
CysCys
Cys
Iron-Sulfur Centers

Ubiquinone
QCoenzyme Q CoQ
Other names and abbreviations:
FAD FeS
FeS
FeS
FMN
NAD+
ubiquinone
Cyt b
ubiquinone
O
O
C H 3 O
CH 3C H 3 O
( CH 2 CH C CH 2 ) n H
CH 3
O H
O H
C H 3 O
CH 3C H 3 O
( CH 2 CH C CH 2 ) n H
CH 3
2 e
+ 2 H +
c o e n z y m e Q
c o e n z y m e Q H 2
Most often n = 10Free CoQ can undergo a 2 e oxidation/reduction:Q + 2 e + 2 H+ QH2.

O
O
C H 3 O
CH 3C H 3 O
(CH 2 CH C CH 2 )n H
CH 3
O H
O H
C H 3 O
CH 3C H 3 O
(CH 2 CH C CH 2 )n H
CH 3
e + 2 H +
c o e n zy m e Q
c o e n zy m e Q H 2
O
O
C H 3 O
CH 3C H 3 O
(CH 2 CH C CH 2 )n H
CH 3e
c o e n zy m e Q •
When bound to special sites in respiratory complexes, CoQ can accept 1 e- to form a semiquinone radical (Q•-).

• Coenzyme Q (CoQ, Q or ubiquinone) is lipid-soluble. It dissolves in the hydrocarbon core of a membrane.
• the only electron carrier not bound to a protein.
• it can accept/donate 1 or 2 e-. Q can mediate e- transfer between 2 e- that transfer and 1 e- carriers
57
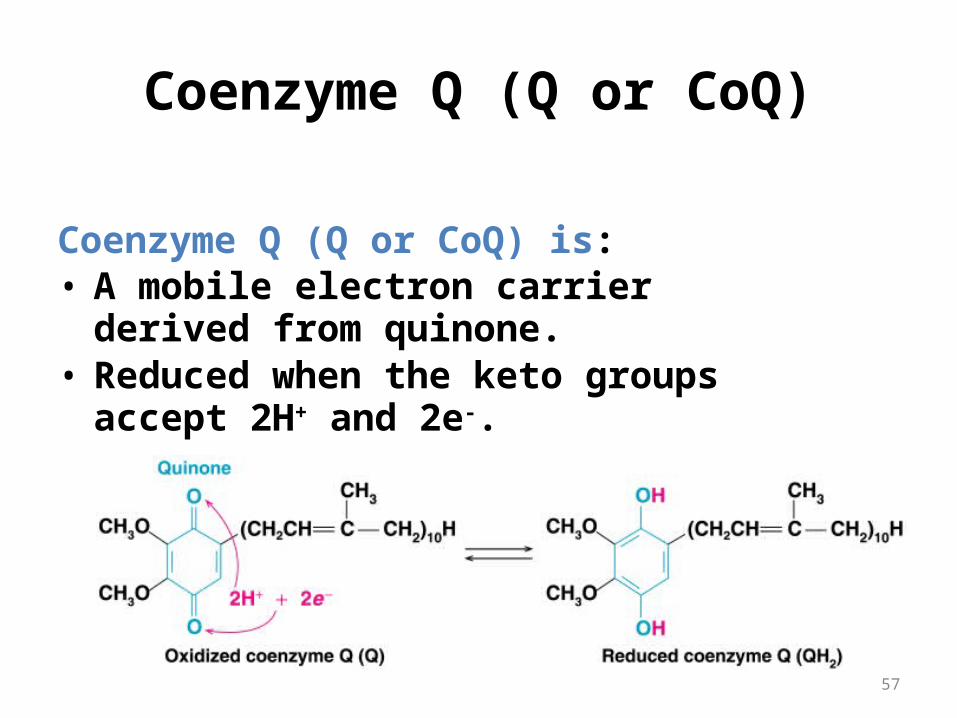
Coenzyme Q (Q or CoQ)
Coenzyme Q (Q or CoQ) is: • A mobile electron carrier derived from
quinone.• Reduced when the keto groups accept 2H+
and 2e-.
58
Cytochromes
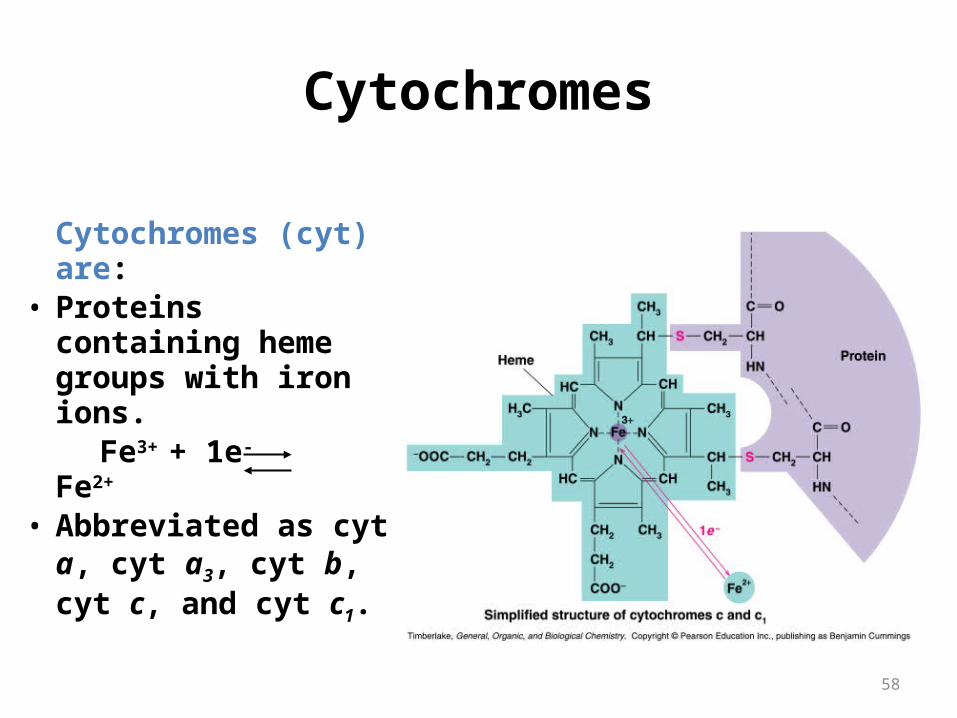
Cytochromes (cyt) are:
• Proteins containing heme groups with iron ions.
Fe3+ + 1e- Fe2+
• Abbreviated as cyt a, cyt a3, cyt b, cyt c, and cyt c1.

Cytochromes
NAD+
FMN
FeS
ubiquinoneFAD FeS
Cyt b
FeS Cyt c1 Cyt c Cyt a Cyt a3
1/2 O2
ubiquinone
proteins that accept electrons from QH2 or FeS
Ultimately transfers the electrons to oxygen

Cytochromes are electron carriers containing hemes . Hemes in the 3 classes of cytochrome (a, b, c) differ in substituents on the porphyrin ring.
Some cytochromes(b,c1,a,a3) are part of large integral membrane protein complexes.
Cytochrome c is a small, water-soluble protein.
Cytochromes
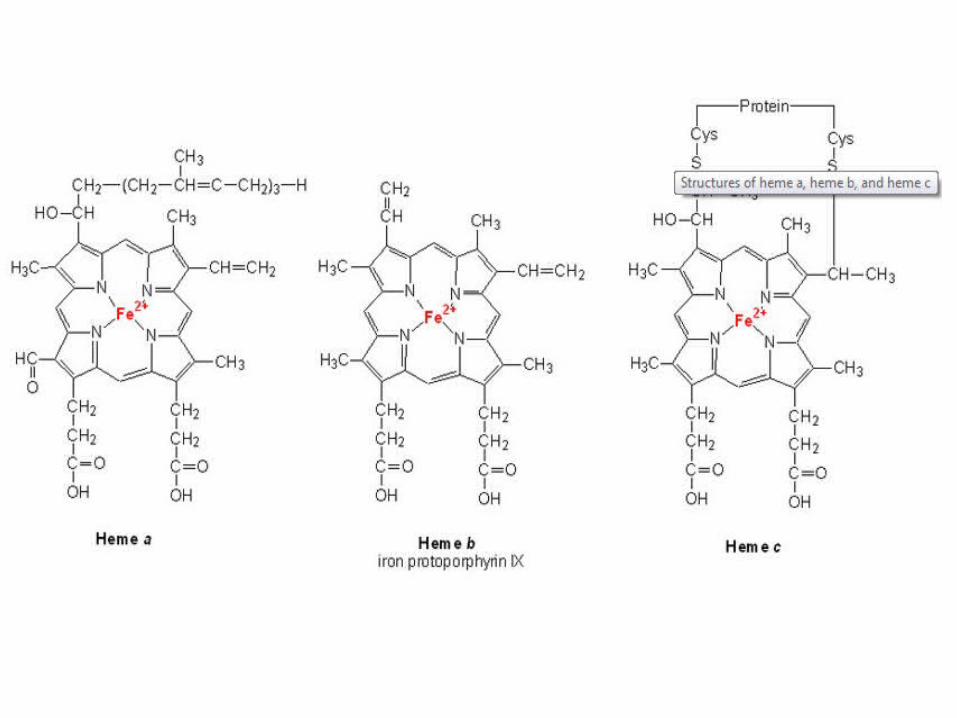


The heme iron can undergo 1 e- transition between ferric and ferrous states: Fe3+ + e- Fe2+
Copper ions besides two heme A groups (a and a3) act as electron carriers in Cyta,a3 Cu2++e- Cu+
Heme is a prosthetic group of cytochromes. Heme contains an iron atom in a porphyrin ring system.

NAD+, flavins and Q carry electrons and H+
Cytochromes and non-haem iron proteins carry only electrons
NAD+ FAD undergoes only a 2 e- reaction; Cytochromes undergo only 1e- reactionsFMN Q undergoes 1e- and 2 e- reaction
Electron carriers

Electron Transport chain(respiratory chain)
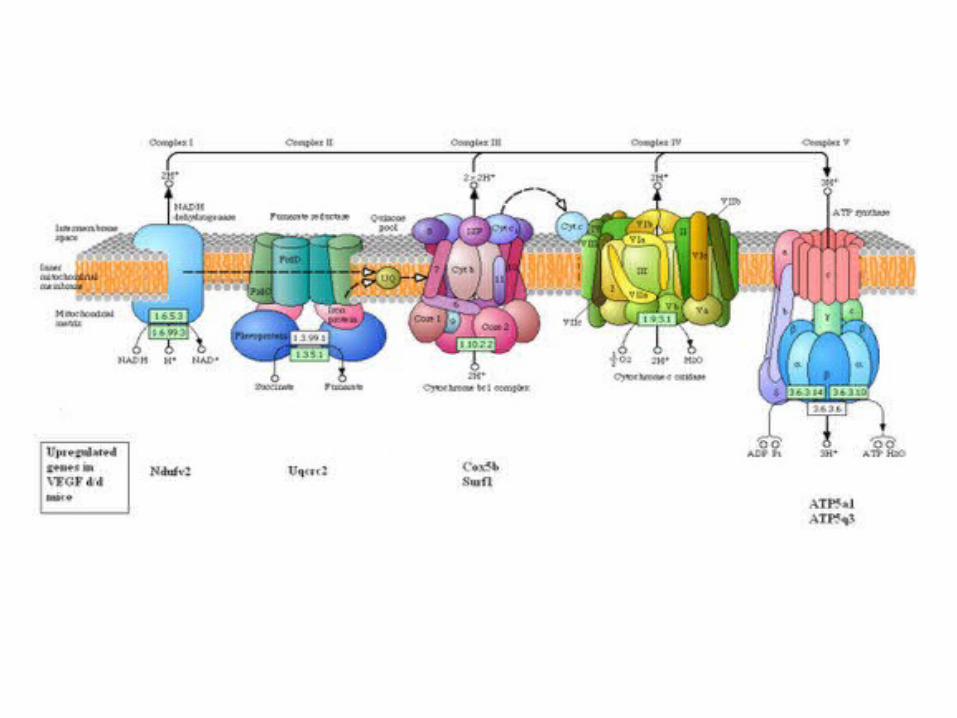
• The electron transport chain in the inner mitochondrial membrane can be isolated in four proteins complexes(I, II, III, IV).
• A lipid soluble coenzyme (Q) and a water soluble protein (cyt c) shuttle between protein complexes
• Electrons transfer through the chain - from complexes I and II to complex IV

The electron transport chain
Mitochondrial Complexes

67
Electron Transport Chain

NAD+
FMN
FeS
ubiquinoneFAD FeS
Cyt b
FeS Cyt c1 Cyt c Cyt a Cyt a3
1/2 O2
ubiquinone
I
II
III IV
Mitochondrial Complexes
NADH Dehydrogenase
Succinate dehydrogenase
CoQ-cyt c Reductase
Cytochrome Oxidase


70
At Complex I, hydrogen and electrons are transferred from:
NADH to FMN.FMN + NADH + H+ FMNH2 + NAD+
FMNH2 to Fe-S clusters and Q, which reduces Q to QH2 and regenerates FMN.
Q + FMNH2 QH2 + FMN
Complex I to Complex III by Q (QH2), a mobile carrier.
Complex I NADH Dehydrogenase



73
At Complex II, hydrogen and electrons are transferred from:
FADH2 to Complex II, which is at a lower energy level than Complex I.
FADH2 to coenzyme Q, which reduces Q and regenerates FAD.Q + FADH2 QH2 + FAD
Complex II to Complex III by Q(QH2), a mobile carrier.
Complex IISuccinate Dehydrogenase



76
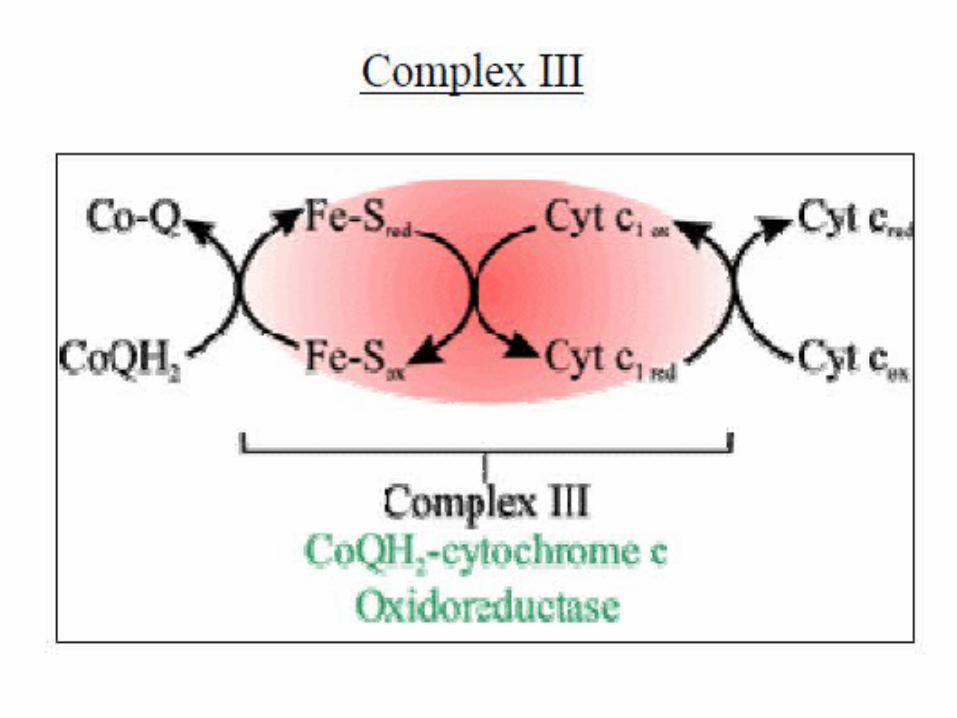
At Complex III, electrons are transferred from: QH2 to two Cyt b, which reduces Cyt b and
regenerates Q.2Cyt b (Fe3+) + QH2 2Cyt b (Fe2+) + Q + 2H+
Cyt b to Fe-S clusters and to Cyt c, the second mobile carrier.
2Cyt c (Fe3+) + 2Cyt b (Fe2+) 2Cyt c (Fe2+) + 2Cyt b (Fe3+)
Complex III CoQ-Cytochrome c reductase


79
Complex IV Cytochrome c Oxidase
At Complex IV, electrons are transferred from:
Cyt c to Cyt a..
2Cyt a (Fe3+) + 2Cyt c (Fe2+) 2Cyt a (Fe2+) + 2Cyt c
(Fe3+) Cyt a to Cyt a3, which provides the
electrons to combine H+ and oxygen to form water. 4H+ + O2 + 4e- (from Cyt a3) 2H2O


Complex Name No. of Proteins
Prosthetic Groups
Complex I NADH Dehydrogenase 46 FMN, 9 Fe-S centers
Complex II Succinate-CoQ Reductase 5 FAD, cyt b560, 3 Fe-S centers
Complex III CoQ-cyt c Reductase 11 cyt bH, cyt bL, cyt c1, Fe-SRieske
Complex IV Cytochrome Oxidase 13 cyt a, cyt a3, CuA, CuB

Support for this order of events
1. Energetically favorable. electrons pass from lower to higher standard reduction potentials.2. Spectra: the absorption spectrum for the reduced carrier differs from that of its oxidized form. carriers closer to oxygen are more oxidized.
3. Specific inhibitors. Those before the blocked step should be reduced and those after be oxidized.
4. Assay of individual complexes.NADH can reduce complex I but not the other complexes.
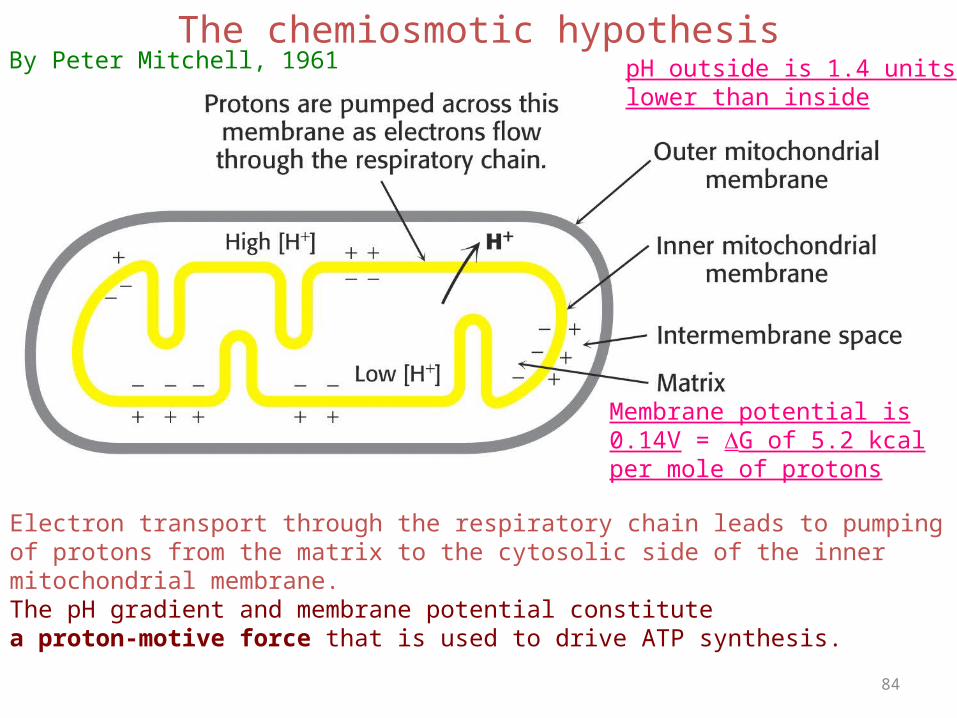
84
The chemiosmotic hypothesis
Electron transport through the respiratory chain leads to pumpingof protons from the matrix to the cytosolic side of the inner mitochondrial membrane.The pH gradient and membrane potential constitute a proton-motive force that is used to drive ATP synthesis.
pH outside is 1.4 unitslower than inside
Membrane potential is0.14V = G of 5.2 kcalper mole of protons
By Peter Mitchell, 1961

Order and Reduction Potentials
NAD+
FMN
FeS
ubiquinoneFAD FeS
Cyt b
FeS Cyt c1 Cyt c Cyt a Cyt a3
1/2 O2
ubiquinone
+0.077
+0. 22 +0. 25
+0. 29 +0. 55
-0.32
+0.03
+0.045
+0.82
-0.3

H+ Transport
Complex I, III, IV drive H+ transport from matrix to the cytosol When e- flow through, which creates proton gradient(electrochemical potential) across the inner membrane
Complex I and Complex IV : The mechanism of H+ transport is still not known.
The mechanism of H+ transport in Complex III is Q cycle.

4H+ are pumped per 2e passing through complex III.
The H+/eratio is less certain for the other complexes: probably 4H+/2e for complex I; 2H+/2e for complex IV.
Matrix
H+ + NADH NAD+
+ 2H+ 2H+ + ½ O2 H2O
2 e – –
I Q III IV
+ +
4H+ 4H+ 2H+ Intermembrane Space
cyt c


89
ATP synthase is composed of 3 fragments: F0, which is localized in the membrane; F1, which protrudes from the inside of the inner membrane into the matrix; and oligomycin sensitivity-conferring protein (OSCP), which connects F0 to F1.
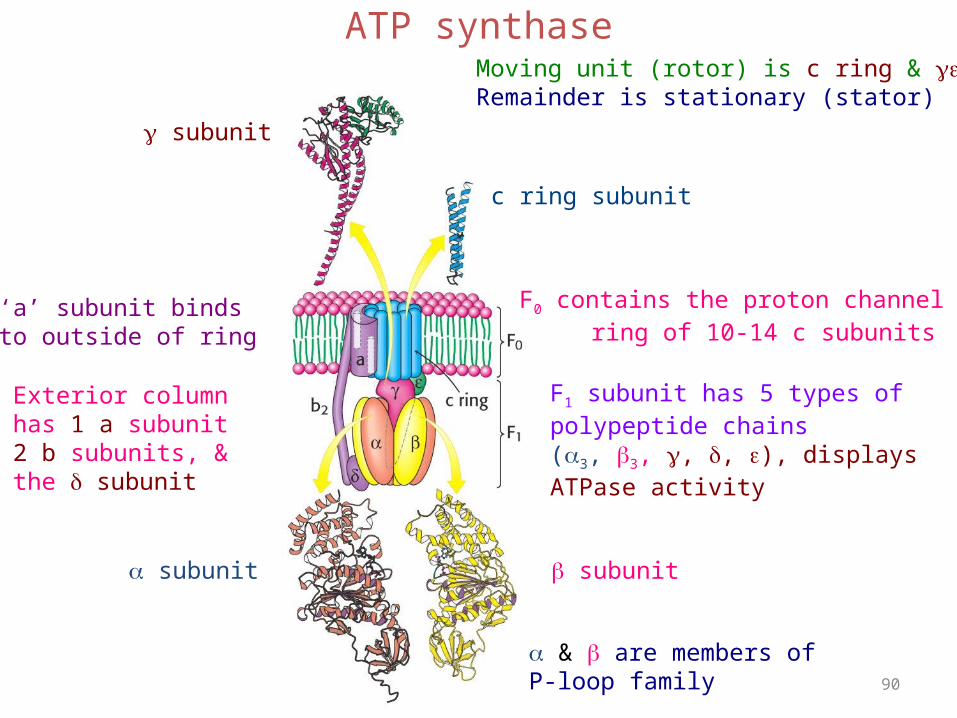
90
ATP synthase
subunit
c ring subunit
subunit subunit
F1 subunit has 5 types ofpolypeptide chains(3, 3, , , ), displaysATPase activity
& are members of P-loop family
F0 contains the proton channel ring of 10-14 c subunits
‘a’ subunit bindsto outside of ring
Exterior columnhas 1 a subunit2 b subunits, &the subunit
Moving unit (rotor) is c ring & Remainder is stationary (stator)

91
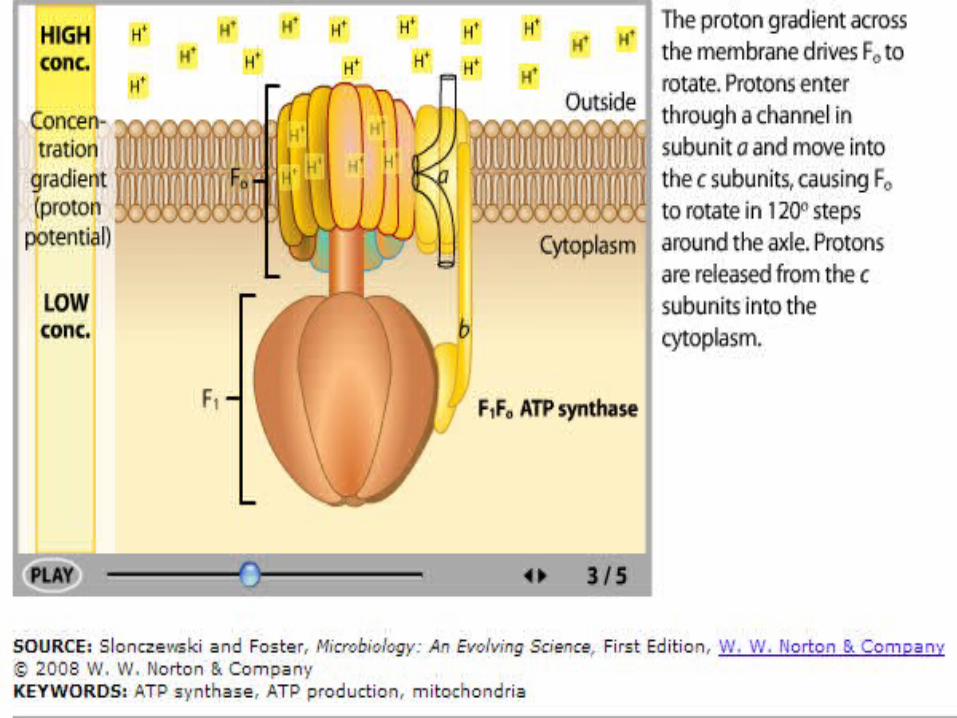
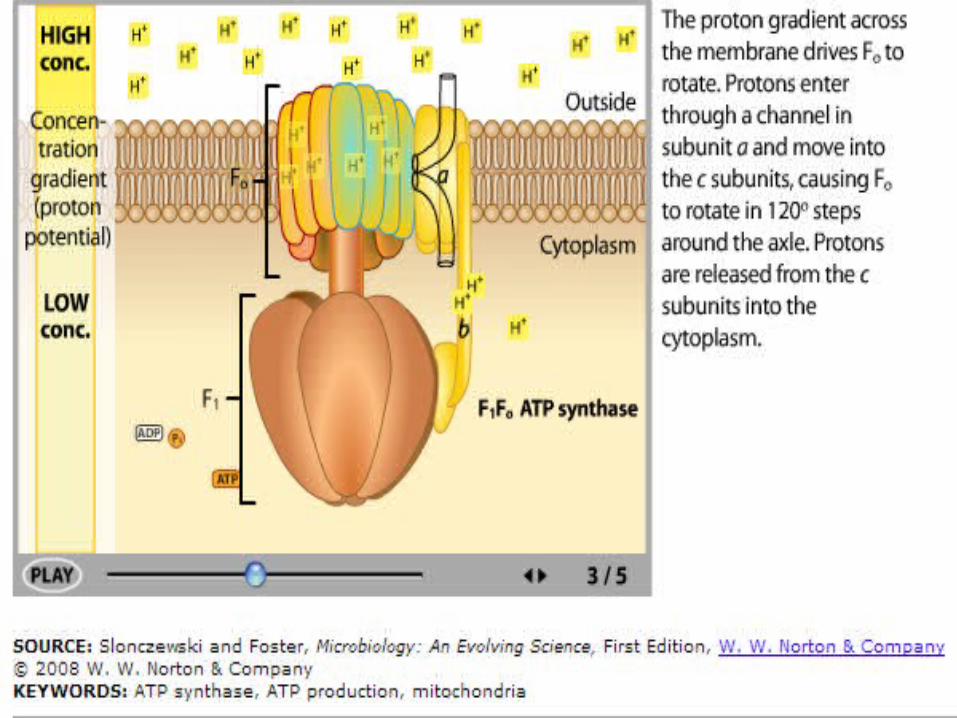
ATP Synthase
In ATP synthase:• Protons flow back to
the matrix through a channel in the F0 complex.
• Proton flow provides the energy that drives ATP synthesis by the F1 complex.
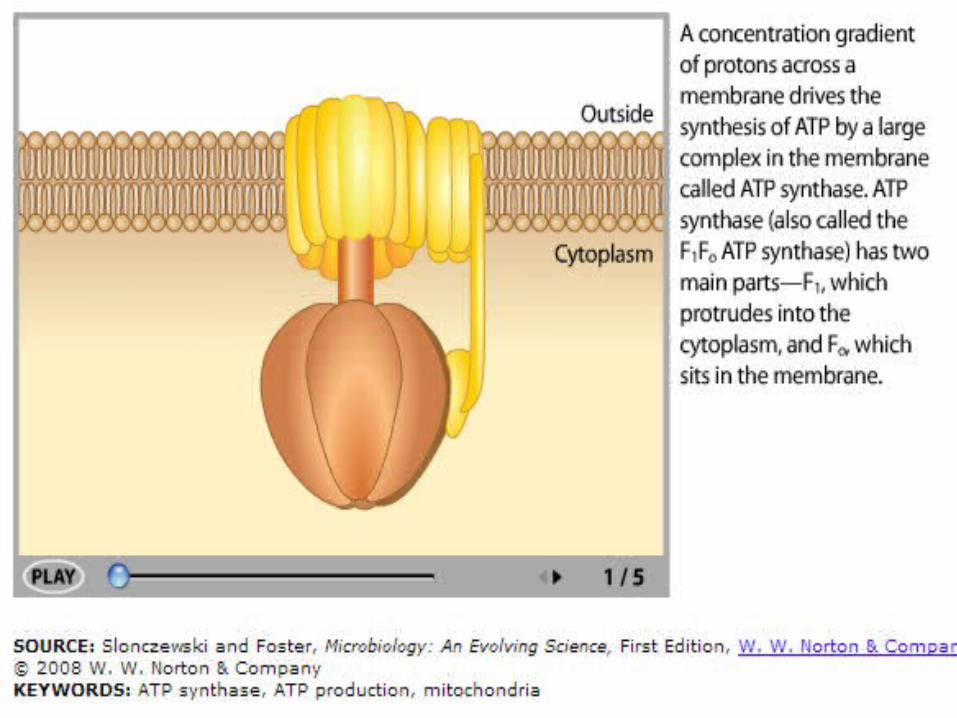






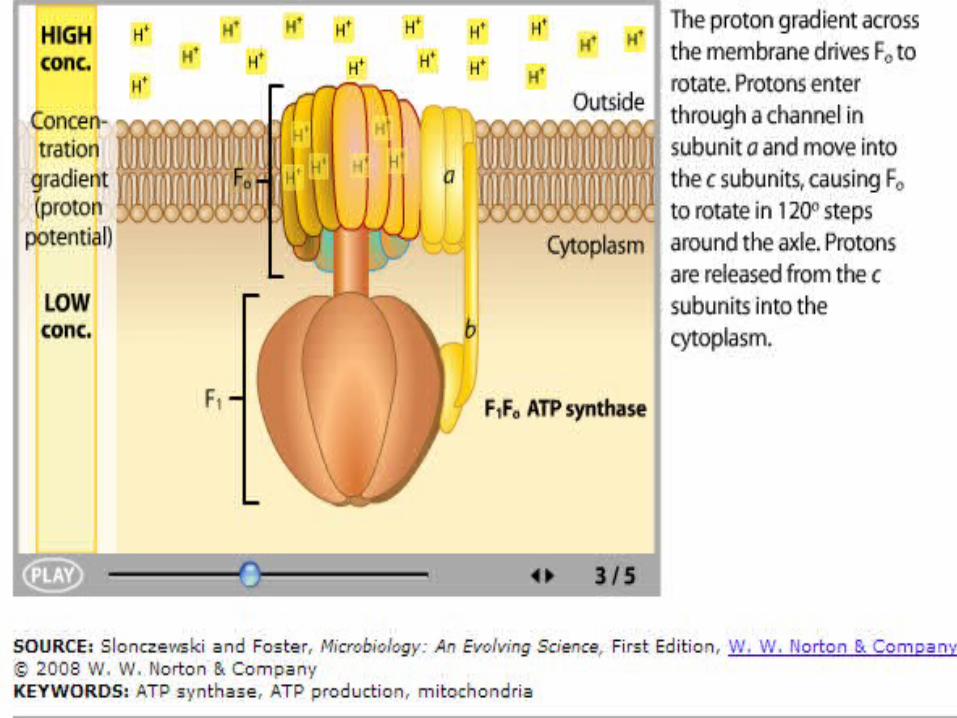







108
ATP Synthase F1 Complex
In the F1 complex of ATP synthase:• A center subunit () is surrounded by three
protein subunits: loose (L), tight (T), and open (O).
• Energy from the proton flow through F0 turns the center subunit (), which changes the shape (conformation) of the three subunits.

109
ATP Synthesis in F1
During ATP synthesis:• ADP and Pi enter the loose L site.• The center subunit turns changing the L site to a
tight T conformation.• ATP is formed in the T site where it remains
strongly bound.• The center subunit turns changing the T site to
an open O site, which releases the ATP.

110
ATP Synthase F1 Diagram

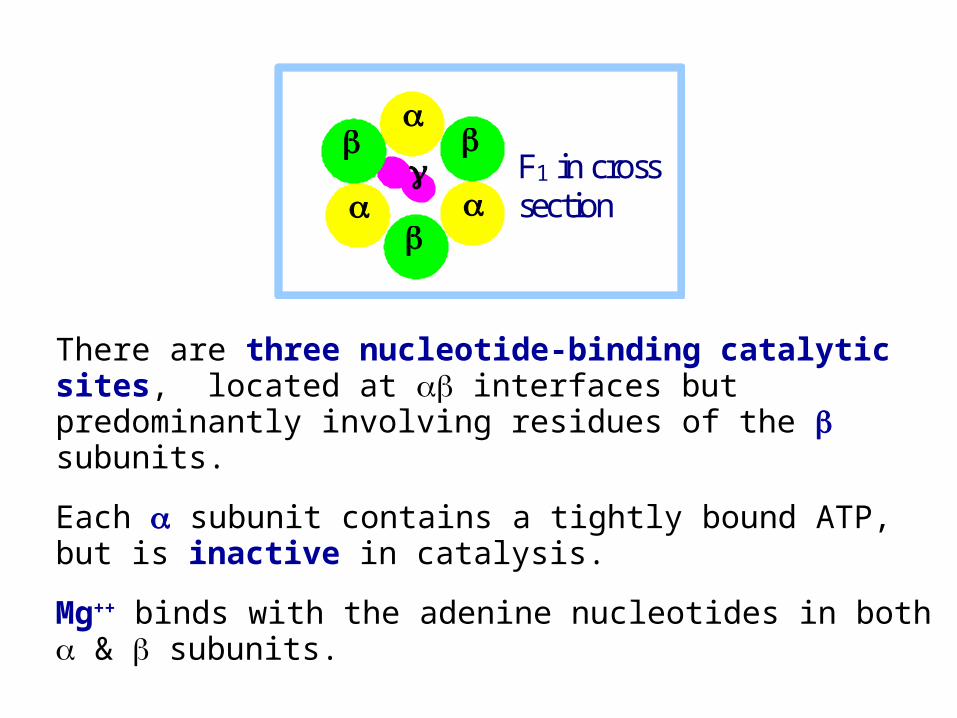
There are three nucleotide-binding catalytic sites, located at interfaces but predominantly involving residues of the subunits.
Each subunit contains a tightly bound ATP, but is inactive in catalysis.
Mg++ binds with the adenine nucleotides in both & subunits.
F1 in cross
section
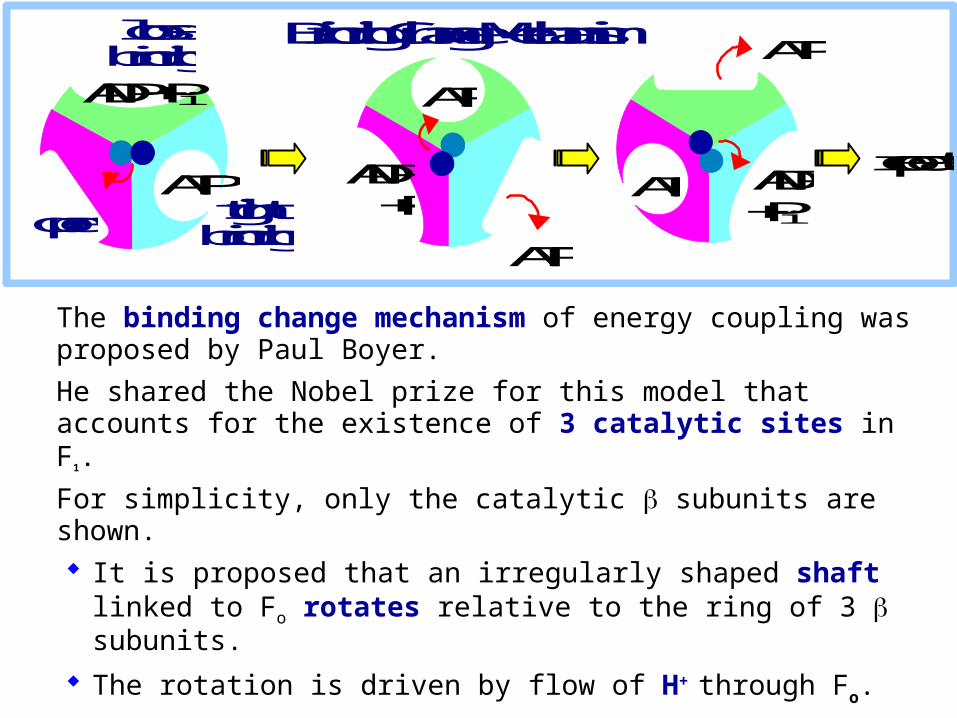
The binding change mechanism of energy coupling was proposed by Paul Boyer.
He shared the Nobel prize for this model that accounts for the existence of 3 catalytic sites in F1.
For simplicity, only the catalytic subunits are shown. It is proposed that an irregularly shaped shaft linked
to Fo rotates relative to the ring of 3 subunits.
The rotation is driven by flow of H+ through Fo.
ADP + Pi ATP
ATP
ATP ADP + Pi
ATP ADP + Pi
ATP open tight
binding
loose binding
repeat
Binding Change Mechanism
ADP + Pi ATP
ATP
ATP ADP + Pi
ATP ADP + Pi
ATP open tight
binding
loose binding
repeat
Binding Change Mechanism
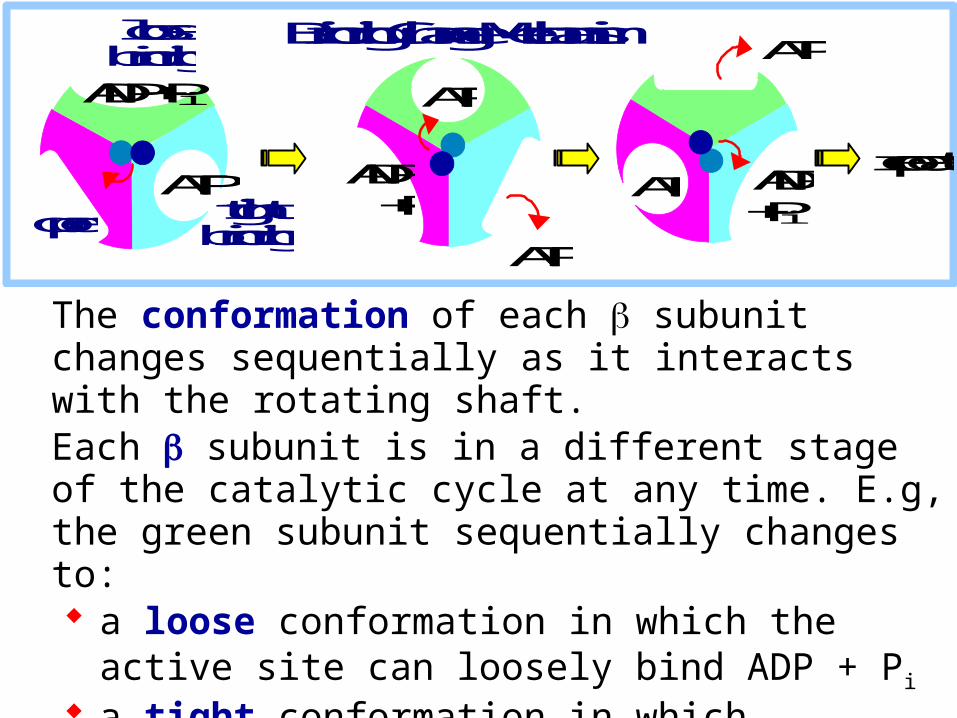
The conformation of each subunit changes sequentially as it interacts with the rotating shaft. Each subunit is in a different stage of the catalytic cycle at any time. E.g, the green subunit sequentially changes to: a loose conformation in which the active site can
loosely bind ADP + Pi
a tight conformation in which substrates are tightly bound and ATP is formed an open conformation that favors ATP release.

Bound to one subunit is a non-hydrolyzable ATP analog (assumed to be the tight conformation).
Bound to another subunit is ADP (loose). The third subunit has an empty active site (open).
This is consistent with the binding change model, whichpredicts that each subunit, being differently affected bythe irregularly shaped rotating shaft, will be in a different stage of the catalytic cycle.
ADP + Pi ATP
ATP
ATP ADP + Pi
ATP ADP + Pi
ATP open tight
binding
loose binding
repeat
Binding Change Mechanism
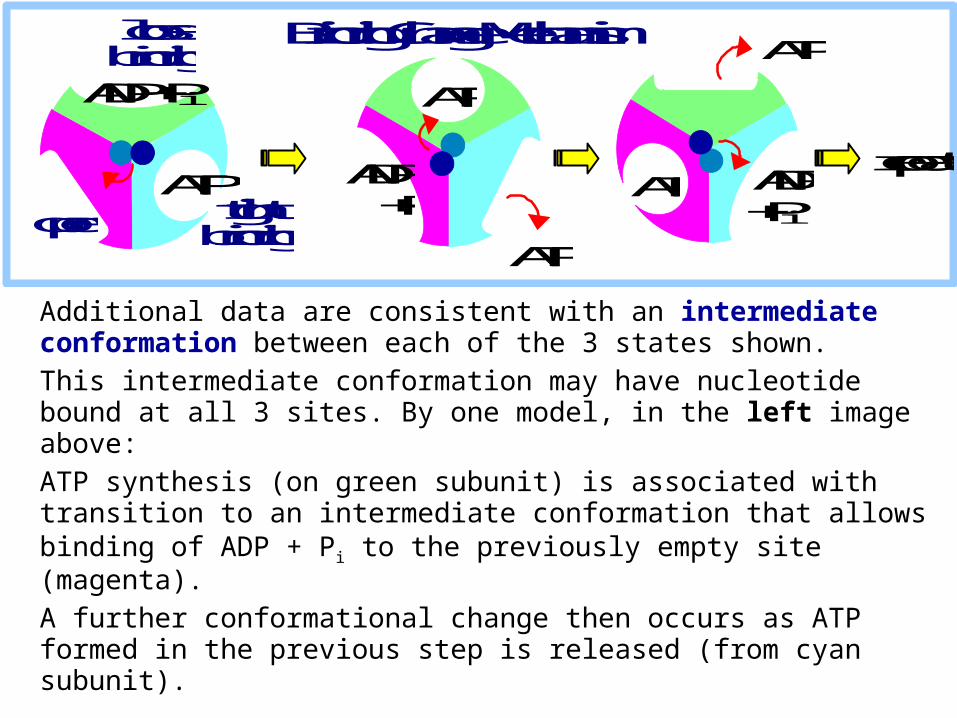
Additional data are consistent with an intermediate conformation between each of the 3 states shown. This intermediate conformation may have nucleotide bound at all 3 sites. By one model, in the left image above:ATP synthesis (on green subunit) is associated with transition to an intermediate conformation that allows binding of ADP + Pi to the previously empty site (magenta).A further conformational change then occurs as ATP formed in the previous step is released (from cyan subunit).
ADP + Pi ATP
ATP
ATP ADP + Pi
ATP ADP + Pi
ATP open tight
binding
loose binding
repeat
Binding Change Mechanism




P :O RATIO
• The P:O ratio is defined as the number of inorganic phosphate molecules incorporated into ATP for every atom of oxygen consumed.
• P:O ratio = NADH = 3• P:O ratio = FADH2 = 2

Transport of ATP, ADP, & Pi
ATP produced in the mitochondrial matrix must exit to the cytosol to be used by transport pumps, kinases, etc.
ADP & Pi arising from ATP hydrolysis in the cytosol must reenter the matrix to be converted again to ATP.
Two carrier proteins in the inner mitochondrial membrane are required.
The outer membrane is considered not a permeability barrier. Large outer membrane VDAC channels are assumed to allow passage of adenine nucleotides and Pi.
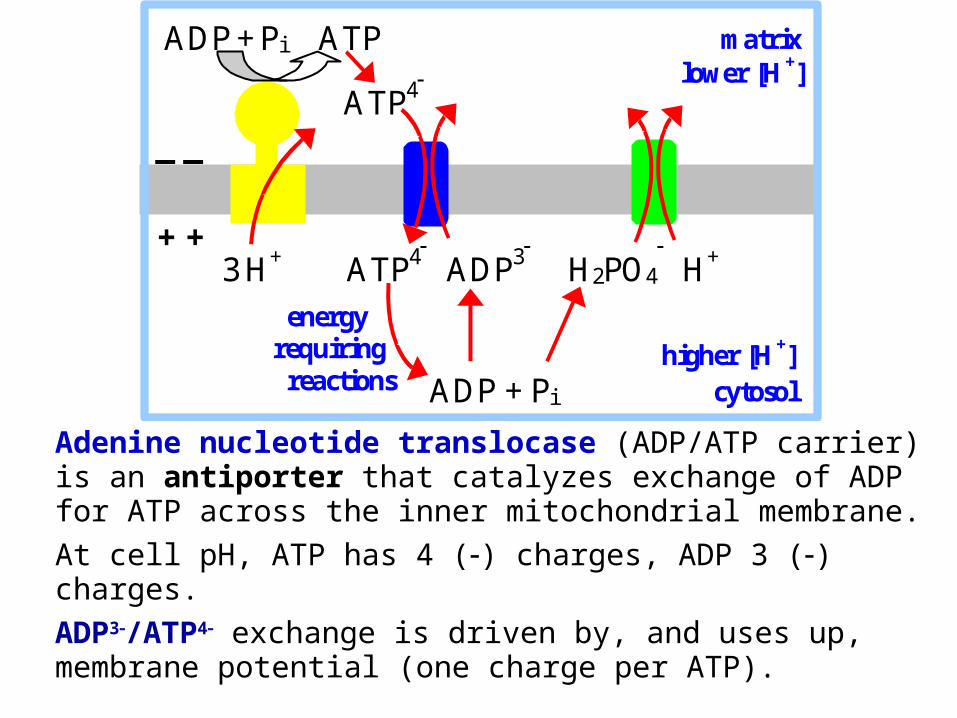
Adenine nucleotide translocase (ADP/ATP carrier) is an antiporter that catalyzes exchange of ADP for ATP across the inner mitochondrial membrane. At cell pH, ATP has 4 () charges, ADP 3 () charges. ADP3/ATP4 exchange is driven by, and uses up, membrane potential (one charge per ATP).
ADP + Pi ATP matrix lower [H+]
_ _
3 H+ ATP4 ADP3 H2PO4
H+
higher [H+] ADP + Pi cytosol
energyrequiring reactions
ATP4
+ +

Phosphate re-enters the matrix with H+ by an electroneutral symport mechanism. Pi entry is driven by, & uses up, the pH gradient (equivalent to one mol H+ per mol ATP).Thus the equivalent of one mol H+ enters the matrix with ADP/ATP exchange & Pi uptake. Assuming 3H+ transported by F1Fo, 4H+ total enter the matrix per ATP synthesized.
ADP + Pi ATP matrix lower [H+]
_ _
3 H+ ATP4 ADP3 H2PO4
H+
higher [H+] ADP + Pi cytosol
energyrequiring reactions
ATP4
+ +
Animation


126
Regulation of Electron Transport
In the electron transport system:• Low levels of ADP, Pi, oxygen, and NADH
decrease electron transport activity.• High levels of ADP activate electron
transport.

Name Function Site of Action
Rotenone e– transport inhibitor Complex I
Amytal e– transport inhibitor Complex I
Antimycin A e– transport inhibitor Complex III
Cyanide e– transport inhibitor Complex IV
Carbon Monoxide e– transport inhibitor Complex IV
Azide e– transport inhibitor Complex IV
2,4,-dinitrophenol Uncoupling agent transmembrane H+ carrier
Pentachlorophenol Uncoupling agent transmembrane H+ carrier
Oligomycin Inhibits ATP synthase OSCP fraction of ATP synthase
Inhibitors of Oxidative Phosphorylation




131
ATP Energy from Glucose
• The complete oxidation of glucose yields 6CO2, 6H2O, and 36 ATP.

132
In glycolysis: • Glucose forms 2 pyruvate, 2 ATP and 2NADH. • NADH produced in the cytoplasm cannot enter the
mitochondria. • A shuttle compound (glycerol-3-phosphate) moves
hydrogen and electrons into the mitochondria to FAD, which forms FADH2.
• Each FADH2 provides 2 ATP.
Glucose 2 pyruvate + 6 ATP
ATP from Glycolysis

133
ATP from Glycolysis
Reaction Pathway ATP for One GlucoseATP from GlycolysisActivation of glucose -2 ATPOxidation of 2 NADH (as FADH2) 4 ATP
Direct ADP phosphorylation (two triose) 4 ATP
Summary: C6H12O6 2 pyruvate + 2H2O + 6 ATP
glucose

134
• One turn of the citric acid cycle provides:3 NADH x 3 ATP = 9 ATP1 FADH2 x 2 ATP = 2 ATP1 GTP x 1 ATP = 1 ATP
Total = 12 ATPAcetyl CoA 2 CO2 + 12 ATP
• Because glucose provides two acetyl CoA , two turns of the citric acid cycle produce 24 ATP.2 Acetyl CoA 4 CO2 + 24 ATP
ATP from Citric Acid Cycle

135
One glucose molecule undergoing complete oxidation provides:From glycolysis 6 ATPFrom 2 Pyruvate 6 ATPFrom 2 Acetyl CoA 24 ATP
Overall ATP Production for One Glucose: C2H12O6 + 6O2 + 36ADP + 36Pi Glucose 6CO2 + 6H2O + 36ATP
ATP from Glucose

Substrate level phosphorylation
• The energy from a high energy compound is directly transferred to nucleotide di-phosphate to form a tri-phosphate without the help of electron transport chain (ETC).
• Example :- Bisphospho gylcerate kinase- Pyruvate kinase- Succinate thiokinase
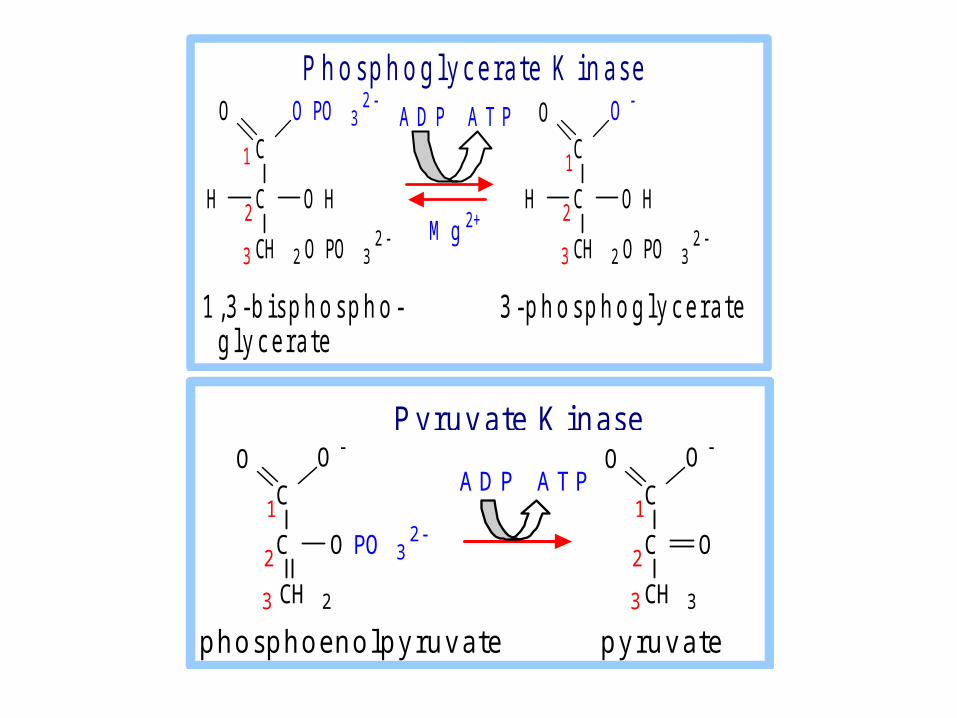
C
C
CH 2 O PO 32
O O PO 32
H O H
C
C
CH 2 O PO 32
O O
H O H
A D P A T P
1
22
3 3
1
M g 2+
1 , 3 - b i s p h o s p h o - 3 - p h o s p h o g l y c e r a t e g l y c e r a t e
P h o s p h o g l y c e r a t e K i n a s e
C
C
CH 3
O O
O2
3
1A D P A T PC
C
CH 2
O O
O PO 32
2
3
1
p h o s p h o e n o l p y r u v a t e p y r u v a t e
P y r u v a t e K i n a s e

5- Succinyl CoA to succinateSuccinyl CoA to succinate Enzyme = succinyl CoA synthetase succinyl-CoA COO-
+ GDP + Pi CH2 + CoA-SH +
CH2 GTP
COO-
succinate
-- substrate level phosphorylation-- GTP is equivalent to ATP; GTP toATP by nucleoside diphosphokinase

THANKS FOR YOUR ATTENTION