osteoporosis of the vertebra and osteochondral remodeling ... · osteoporosis of the vertebra and...
TRANSCRIPT

RESEARCH ARTICLE Open Access
Osteoporosis of the vertebra andosteochondral remodeling of the endplatecauses intervertebral disc degeneration inovariectomized miceZhi-feng Xiao1,2,3†, Jian-bo He1,2,3†, Guo-yi Su1,3, Mei-hui Chen1,2,3, Yu Hou1,3, Shu-dong Chen1,3
and Ding-kun Lin1,2,3*
Abstract
Background: Studies on the relationship between osteoporosis and intervertebral disc degeneration (IVDD) areinconsistent. Therefore, we assessed whether IVDD is affected by vertebral osteoporosis in ovariectomized mice andinvestigated the underlying pathogenesis of IVDD related to osteoporosis.
Methods: Thirty healthy female C57BL/6 J mice aged 8 weeks were randomly divided into two groups: a controlgroup (sham operation, n = 15) and an ovariectomy group (OVX; bilateral ovariectomy, n = 15). At 12 weeks aftersurgery, the bone quantity and microstructure in the lumbar vertebra and endplate as well as the volume of the L4/5disc space were evaluated by microcomputed tomography (micro-CT). The occurrence and characteristic alterations ofIVDD were identified via histopathological staining. The osteoclasts were detected using tartrate-resistant acidphosphatase (TRAP) staining. Type II collagen (Col II), osterix (OSX), osteopontin (OPN), and vascular endothelial growthfactor (VEGF) expression in the intervertebral disc were detected by immunohistochemical analysis.
Results: OVX significantly increased the body weight and decreased the uterus weight. Micro-CT analysis showed thatosteoporosis of the vertebra and osteochondral remodeling of the endplate were accompanied by an increase in theendplate porosity and a decrease in the disc volume in the OVX group. Likewise, histological evaluation revealed thatIVDD occurred at 12 weeks after ovariectomy, with features of endochondral ossification of the endplate, loose andbroken annulus fibrosus, and degeneration of nucleus pulposus. TRAP staining showed that numerous activeosteoclasts appeared in the subchondral bone and cartilaginous endplate of OVX mice, whereas osteoclasts were rarelydetected in control mice. Immunohistochemical analysis demonstrated that the expression of osterix was significantlyincreased, notably in the endplate of OVX mice. In addition, Col II was decreased in the ossification endplate and thedegenerative annulus fibrosus, where OPN and VEGF expressions were elevated in OVX mice.
(Continued on next page)
* Correspondence: [email protected]†Zhi-feng Xiao and Jian-bo He contributed equally to this work.1The Department of Spinal Surgery, The Second Affiliated Hospital ofGuangzhou University of Chinese Medicine, No. 111, Dade Road, YuexiuDistrict, Guangzhou 510120, China2The Laboratory Affiliated to Orthopaedics and Traumatology of ChineseMedicine of Linnan Medical Research Center of Guangzhou University ofChinese Medicine, No. 12, Jichang Road, Baiyun District, Guangzhou 510405,ChinaFull list of author information is available at the end of the article
© The Author(s). 2018 Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, andreproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link tothe Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver(http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
Xiao et al. Arthritis Research & Therapy (2018) 20:207 https://doi.org/10.1186/s13075-018-1701-1

(Continued from previous page)
Conclusions: OVX induced vertebral osteoporosis and osteochondral remodeling of the cartilaginous endplatecontributing to the angiogenesis and an increase in porosity of the bone-cartilage surface, and also affected the matrixmetabolism which consequently had detrimental effects on the intervertebral disc. Our study suggests that preservingthe structural integrity and the function of the adjacent structures, including the vertebrae and endplates, may protectthe disc against degeneration.
Keywords: Intervertebral disc degeneration, Osteoporosis, Ovariectomy, Endplate, Osteochondral remodeling,Microcomputed tomography,
BackgroundLower back pain and spinal compression nerve pain arethe major symptoms caused by intervertebral disc de-generation (IVDD) in the clinic [1, 2]. Spinal instabilityor disability is common in serious IVDD which leads toenormous human suffering and significant socioeco-nomic losses [3]. Unfortunately, there are no currentlyeffective methods to repair IVDD [4] and disc resectionwith interbody fusion are often the final choice. There-fore there is an urgent need to explore the key mechan-ism of IVDD and to develop drugs for its treatment.It has long been recognized that musculoskeletal de-
generation disorders such as osteoporosis, IVDD, andosteoarthritis are a difficult focus in locomotor diseaseresearch. An intimate relationship between cartilage andsubchondral bone has been proven in recent years. Ana-tomically, the vertebrae and the intervertebral discs arecombined in bundles to form the motion segments ofthe spine. From the mechanical and biological points ofview they are closely linked and are considered as afunctional unit [5–7]. Although it is not fully clearwhether it precedes or follows nucleus pulposus degen-eration, the modic change in the endplate is an import-ant feature of IVDD physiopathology [8]. Therefore, thehealth of the bone and its attached nonosseous tissuessuch as cartilage and disc are tightly associated. Researchhas shown that crosstalk between bone and cartilage iselevated in osteoarthritis where the coupling of boneand cartilage turnover is even aggravated [9–11]. More-over, remodeling of the subchondral bone microstruc-ture due to osteoporosis could further exacerbateexperimental osteoarthritis [12]. All of this suggests thatthe subchondral bone is an indispensable factor in theprocess of osteoarthritis. In terms of IVDD, however,there is a lack of research on the pathological mecha-nisms of subchondral bone in IVDD, and it is also un-clear how osteoporosis affects the nonosseous tissuessuch as the intervertebral disc.Because of the structural similarities between joint and
intervertebral discs [13, 14], it is well known that theintervertebral disc is a nonvascular structure and the ex-change of substances between the intervertebral disc andthe vertebra depends on the cranial and caudal endplates
[15–17]. The endplate contains marrow contact chan-nels which provide nutrients for the intervertebral discand discharge metabolic waste through diffusion andliquid flow under cyclic loading [4, 18, 19]. Hence,changes in the vertebra microenvironment resultingfrom rapid bone turnover during vertebral osteoporosisand obstruction of marrow contact channels induced bycalcification of the cartilaginous endplate may accelerateIVDD.Recent studies have provided increasing evidence that
osteoporosis is associated with the evolution of IVDD.The osteoporosis of vertebrae in postmenopausalwomen was correlated with IVDD [20, 21], and sex hor-mones can affect the severity of IVDD [21–23]. Further-more, in osteoprotegerin (OPG) knockout mice,ossification occurred in the cartilage endplate and re-sulted in IVDD [24]. IVDD often occurs with osteopor-osis of the vertebrae, indicating that the development ofosteoporosis and IVDD might be a coupling processwhich could explain why postmenopausal womenhave more lower back pain than men. Some studieshave reported the prospect of delaying the course ofdisc degeneration through improving bone metabol-ism and vertebral osteoporosis. For example, it wasfound that alendronate could retard the progressionof lumbar IVDD in ovariectomized rats by improvingthe bone quality [25, 26]. Calcitonin could also sup-press intervertebral disk degeneration and preservelumbar vertebral bone mineral density and bonestrength [27]. However, it is still puzzling that someclinical and epidemiological studies have shown thatosteoporosis is inversely related to spinal degenerativediseases and IVDD [19, 28, 29]. These studies supportosteoporosis in increasing endplate permeability anddelaying IVDD [19]. In addition, radiographic featuresof lumbar disc degeneration were associated with anincreased bone mineral density (BMD) in the spine[28, 30]. Thus, the relationship between osteoporosisand IVDD is still controversial and confusing. Conse-quently, whether a positive correlation exists betweenosteoporosis and IVDD and how the vertebral bodyaffects the intervertebral disc are still undefined andremain to be further clarified.
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 2 of 15

Mice are commonly used as an animal model forosteoporosis and intervertebral disc degeneration [31–33]. Many studies have demonstrated that the ovariecto-mized mouse is a good model for postmenopausal osteo-porosis [34]. Thus, the purpose of this study was todetermine the roles of postmenopausal osteoporosis inIVDD and to further clarify its underlying mechanismby assessing the detailed pathological changes in L4–L5spine motion segments including vertebrae, the endplate,and the intervertebral disc in ovariectomized mice.
MethodsAnimals and designsFemale C57BL/6 J mice (8 weeks old) were purchasedfrom the Animal Center of Guangzhou University ofChinese Medicine, Guangzhou, China (license number:SCXK (YUE) 2013–0034). The mice were housed inconditions of controlled temperature (22–25 °C) at 40–60% relative humidity with alternate day and night andwere allowed food and water freely. The animals wererandomly divided into two groups. The mice underwenteither sham operation (control group, n = 15) or bilateralovariectomy (OVX group, n = 15) while under anesthesiawith 4% chloral hydrate at a dose of 400 mg/kg weightvia intraperitoneal injection. The body weight of themice was recorded weekly. The mice were sacrificed at12 weeks after surgery, and L3–L6 spinal motion seg-ments and hind limbs were harvested for subsequent ex-periments. All experimental protocols were approved bythe Ethics Committee of Guangzhou University of Chin-ese Medicine and implemented according to the Guidefor Use and Care of Animals.
Microcomputed tomography analysesL3–L6 spinal motion segments were harvested and imme-diately fixed in 4% neutral paraformaldehyde for 72 h at4 °C. The samples were washed three times withphosphate-buffered saline (PBS) for 15 min each time.The L4–L5 segment of samples (n = 6) was measured byhigh resolution microcomputed tomography (micro-CT;Skyscan1172) according to a previously described protocol[31]. Briefly, the scanner was set at a voltage of 59 kV, acurrent of 100uA, and a resolution of 9 μm per pixel tomeasure the spinal segment. Images were reconstructedand analyzed using NRecon v1.6 and CTAn v1.9 software,respectively. Three-dimensional (3D) reconstruction im-ages were obtained using CTvox v3.0. The coronal imagesof the L4/5 segment were used to perform 3D histomor-phometric analyses of the intervertebral disc and cartilageendplate, while transverse images of L5 vertebrae wereused to measure the vertebral body. The region of interest(ROI) of vertebrae, endplates, and intervertebral discswere depicted using CTAn v1.9. A total of 200 or 20 con-secutive images of the ROI were respectively used to show
the 3D reconstruction of the microarchitecture in the ver-tebra and endplate. The ROI of the intervertebral disc wasshown as the mid-plane coronal images of the L4/5 seg-ment. The disc volume was defined as the ROI covered bythe entire invisible space between the L4/5 vertebrae. Thecartilage endplate volume was defined as the visible boneplate volume that covers the vertebrae. Three dimensionalstructural parameters of vertebrae included the total vol-ume of bone mineral density (BMDtv; reflecting bonemass per unit volume), percentage bone volume (bonevolume (BV)/total volume (TV)), trabecular number(Tb.N; the inverse of the mean distance between themid-axes of the structure), trabecular thickness (Tb.Th),trabecular separation (Tb.Sp; the average separation be-tween the mid-axes), trabecular pattern factor (Tb.Pf;measuring the degree of convex surfaces and concave sur-faces of the trabecular, where having many concave sur-faces represents a well-connected spongy lattice, whilemore convex surfaces indicates a bad connectedness; therising value represents the decrease in trabecular connect-edness); connectivity density (CONN.D; reflecting theconnection in the trabecular, with a lower value indicatingincreased interruption of the trabecular), and structuralmodel index (SMI; reflecting the proportion of the plateand rod structure of the trabecular with a value range of0–3 where a higher value indicates an increase inrod-shaped trabecular volume). The parameters of theendplate included the percentage bone volume (BV/TV),number of closed pores (Po.N(cl); representing the num-ber of pores with a closed cavity in the endplate structure),open porosity (PO(op); open pore volume over total porevolume), and total volume of pore space (Po.V(tot)).
Histology and immunohistochemistry examinationsAfter fixation in 4% neutral paraformaldehyde for 72 h,the dorsal attachment including the vertebral arch/lam-ina and facet joint were removed from lumbar L4/5spinal segments (n = 9) and were decalcified in 10%EDTA (pH 7.4) for 14 days at room temperature. Thetibia received the same process. The samples were thendehydrated and embedded in paraffin. The L4/5 seg-ments were sliced into 4-μm thick sections to performhematoxylin and eosin (H&E) staining, safranin O andfast green staining, tartrate-resistant acid phosphatase(TRAP) staining, and immunohistochemistry.H&E and safranin O staining were guided by the in-
structions of the reagent kits (Servicebio BiologicalTechnology Co. Ltd., China). TRAP staining was han-dled according to the protocol of the staining kit (Solar-bio Science & Technology Co. Ltd., China). Briefly, afterdewaxing and hydration, slices were soaked in TRAPstaining solution and incubated at 37 °C for 50 min.After washing in tap water, specimens were counter-stained by methyl green.
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 3 of 15
Immunostaining was performed using standard protocolsaccording to the Abcam/Santa Cruz official website (https://www.abcam.cn/protocols/ihc-tissue-processing-protocol andhttps://www.scbt.com/scbt/zh/resources/protocols/immuno-fluorescence-cell-staining). Briefly, sections were dewaxedand hydrated and then boiled in EDTA antigen repair solu-tion (pH 8.0) for 20 min for antigen retrieval. Tissues weretreated with 3% H2O2 to block endogenous peroxidase. Thesections were then incubated with normal goat serum forblocking and subsequently with primary antibodies includ-ing mouse collagen II (Col II; Abcam, 1:200, ab34712),osterix (OSX; Santa Cruz, 1:200, sc-393,060), osteopontin(OPN; Santa Cruz, 1:200, sc-73,631), and vascular endothe-lial growth factor (VEGF; Santa Cruz, 1:200, sc-7269) at 4 °Covernight. Negative control slices were incubated in PBSwithout antibodies. For immunohistochemical staining,the rest of the procedures were manipulated accordingto the PV-6001 Two-Step IHC Detection Reagent in-structions. After coloration with 3,3’-diaminobenzidine(DAB) solution (ZSGB-BIO Corporation, China), thesections were counterstained with hematoxylin and theyellow or brown color was considered as positive stain-ing. For the immunofluorescent assay, the slides wereincubated with the secondary antibody conjugated withfluorescence-Alexa Fluor® 555 (CST Corporation, USA,#4409) for 1 h avoiding light, followed by counterstain-ing with 4’, 6-diamidino-2-phenylindole (DAPI), andthen detected under fluorescence microscopy (Olympus
DP80, Japan). Positive staining in the disc was quanti-fied using ImageJ Pro Plus software (Media Cybernet-ics, Baltimore, MD, USA).
StatisticsAll data were analyzed using the paired t test of SPSS20.0 software (SPSS Inc., Chicago, IL). The comparisonsof means were performed between the control and OVXgroups. All data were checked for normality and homo-geneity of variance and represent the mean ± standarddeviation (SD). In all analyses, p < 0.05 was consideredstatistically significant.
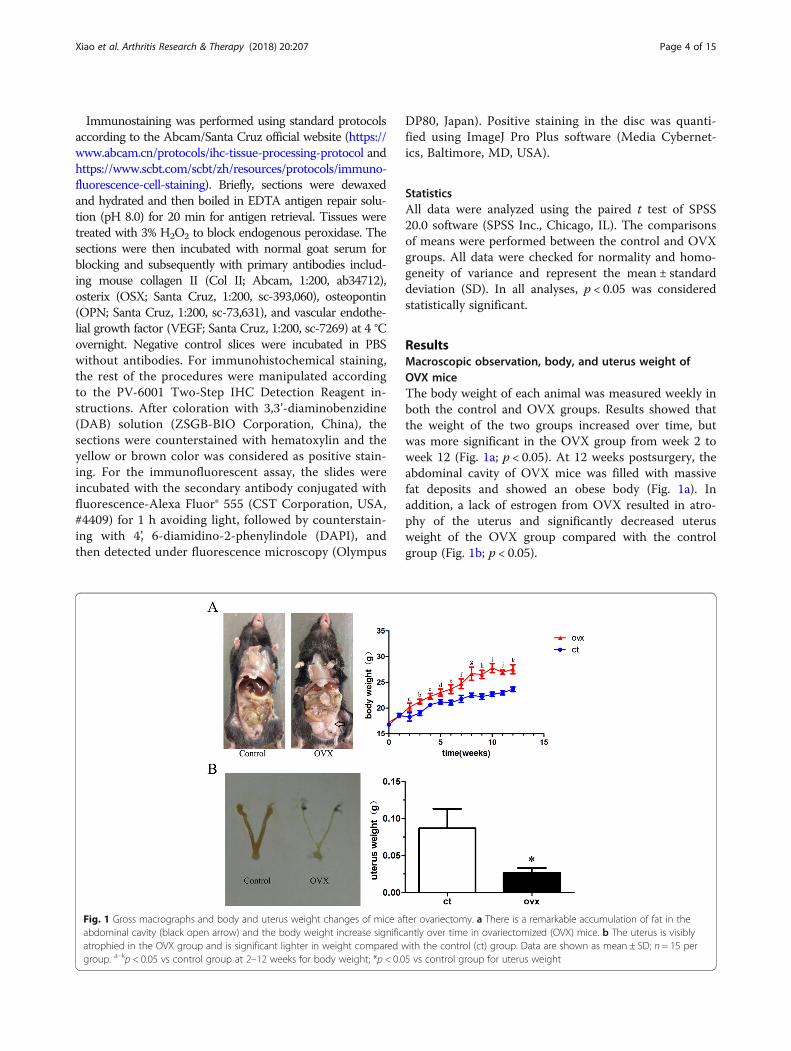
ResultsMacroscopic observation, body, and uterus weight ofOVX miceThe body weight of each animal was measured weekly inboth the control and OVX groups. Results showed thatthe weight of the two groups increased over time, butwas more significant in the OVX group from week 2 toweek 12 (Fig. 1a; p < 0.05). At 12 weeks postsurgery, theabdominal cavity of OVX mice was filled with massivefat deposits and showed an obese body (Fig. 1a). Inaddition, a lack of estrogen from OVX resulted in atro-phy of the uterus and significantly decreased uterusweight of the OVX group compared with the controlgroup (Fig. 1b; p < 0.05).
Fig. 1 Gross macrographs and body and uterus weight changes of mice after ovariectomy. a There is a remarkable accumulation of fat in theabdominal cavity (black open arrow) and the body weight increase significantly over time in ovariectomized (OVX) mice. b The uterus is visiblyatrophied in the OVX group and is significant lighter in weight compared with the control (ct) group. Data are shown as mean ± SD; n = 15 pergroup. a–kp < 0.05 vs control group at 2–12 weeks for body weight; *p < 0.05 vs control group for uterus weight
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 4 of 15
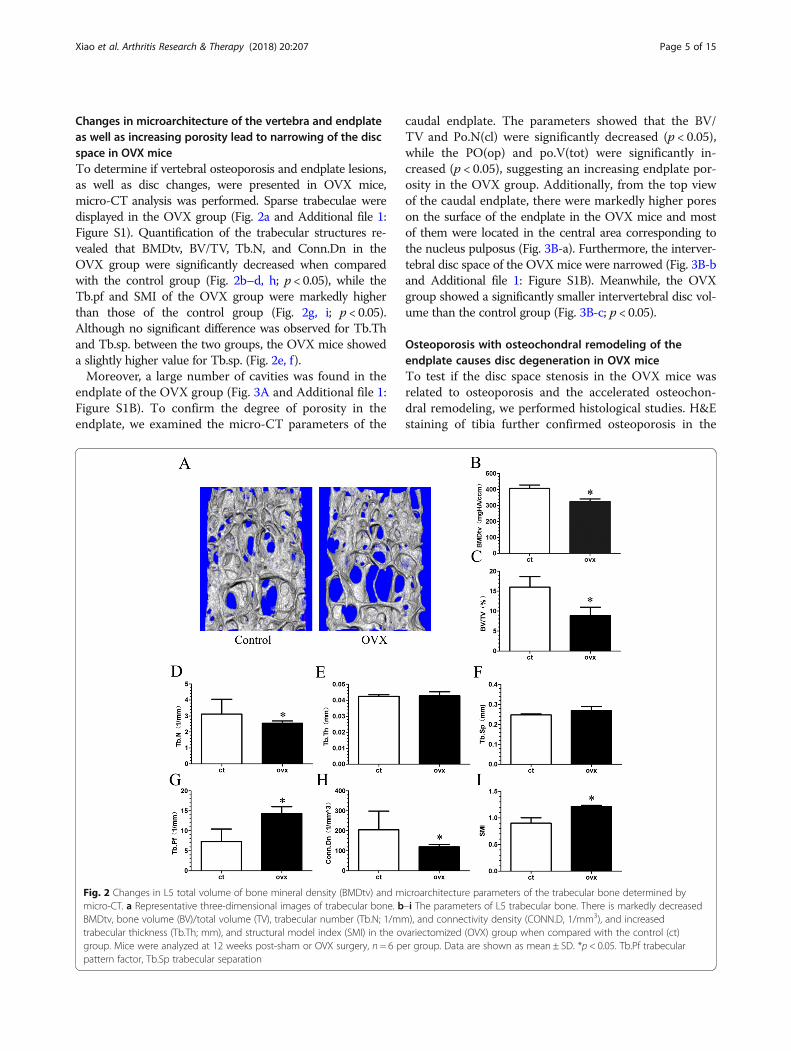
Changes in microarchitecture of the vertebra and endplateas well as increasing porosity lead to narrowing of the discspace in OVX miceTo determine if vertebral osteoporosis and endplate lesions,as well as disc changes, were presented in OVX mice,micro-CT analysis was performed. Sparse trabeculae weredisplayed in the OVX group (Fig. 2a and Additional file 1:Figure S1). Quantification of the trabecular structures re-vealed that BMDtv, BV/TV, Tb.N, and Conn.Dn in theOVX group were significantly decreased when comparedwith the control group (Fig. 2b–d, h; p < 0.05), while theTb.pf and SMI of the OVX group were markedly higherthan those of the control group (Fig. 2g, i; p < 0.05).Although no significant difference was observed for Tb.Thand Tb.sp. between the two groups, the OVX mice showeda slightly higher value for Tb.sp. (Fig. 2e, f).Moreover, a large number of cavities was found in the
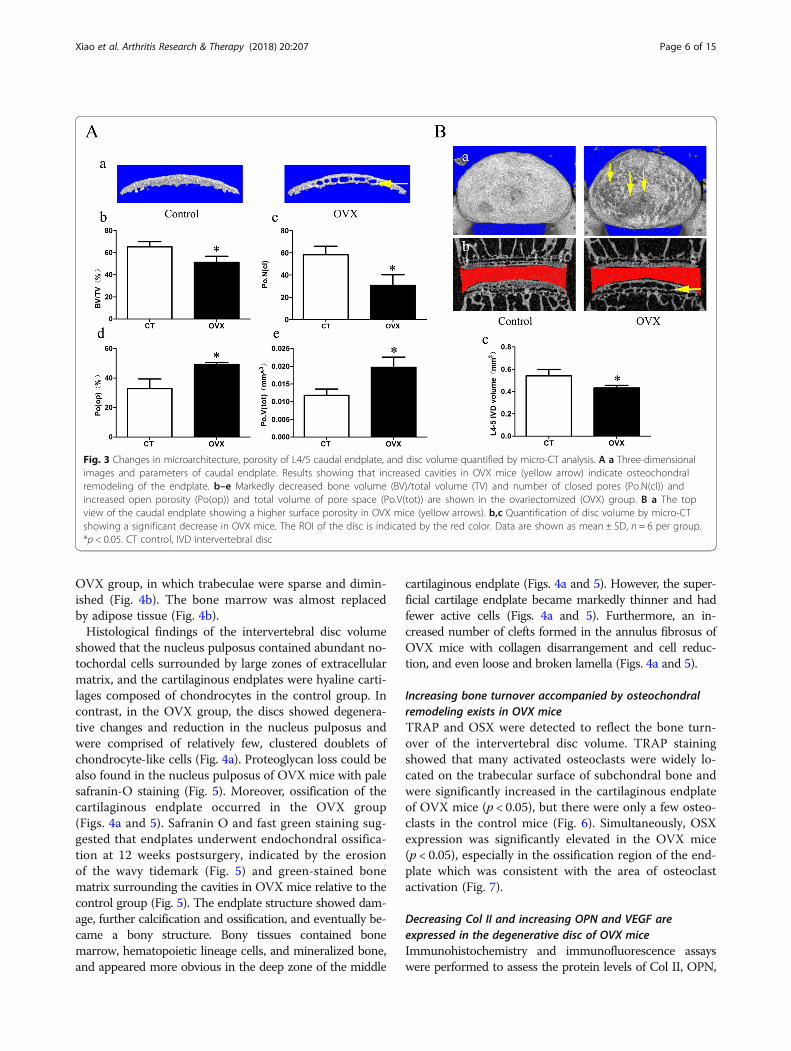
endplate of the OVX group (Fig. 3A and Additional file 1:Figure S1B). To confirm the degree of porosity in theendplate, we examined the micro-CT parameters of the
caudal endplate. The parameters showed that the BV/TV and Po.N(cl) were significantly decreased (p < 0.05),while the PO(op) and po.V(tot) were significantly in-creased (p < 0.05), suggesting an increasing endplate por-osity in the OVX group. Additionally, from the top viewof the caudal endplate, there were markedly higher poreson the surface of the endplate in the OVX mice and mostof them were located in the central area corresponding tothe nucleus pulposus (Fig. 3B-a). Furthermore, the interver-tebral disc space of the OVXmice were narrowed (Fig. 3B-band Additional file 1: Figure S1B). Meanwhile, the OVXgroup showed a significantly smaller intervertebral disc vol-ume than the control group (Fig. 3B-c; p < 0.05).
Osteoporosis with osteochondral remodeling of theendplate causes disc degeneration in OVX miceTo test if the disc space stenosis in the OVX mice wasrelated to osteoporosis and the accelerated osteochon-dral remodeling, we performed histological studies. H&Estaining of tibia further confirmed osteoporosis in the
Fig. 2 Changes in L5 total volume of bone mineral density (BMDtv) and microarchitecture parameters of the trabecular bone determined bymicro-CT. a Representative three-dimensional images of trabecular bone. b–i The parameters of L5 trabecular bone. There is markedly decreasedBMDtv, bone volume (BV)/total volume (TV), trabecular number (Tb.N; 1/mm), and connectivity density (CONN.D, 1/mm3), and increasedtrabecular thickness (Tb.Th; mm), and structural model index (SMI) in the ovariectomized (OVX) group when compared with the control (ct)group. Mice were analyzed at 12 weeks post-sham or OVX surgery, n = 6 per group. Data are shown as mean ± SD. *p < 0.05. Tb.Pf trabecularpattern factor, Tb.Sp trabecular separation
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 5 of 15
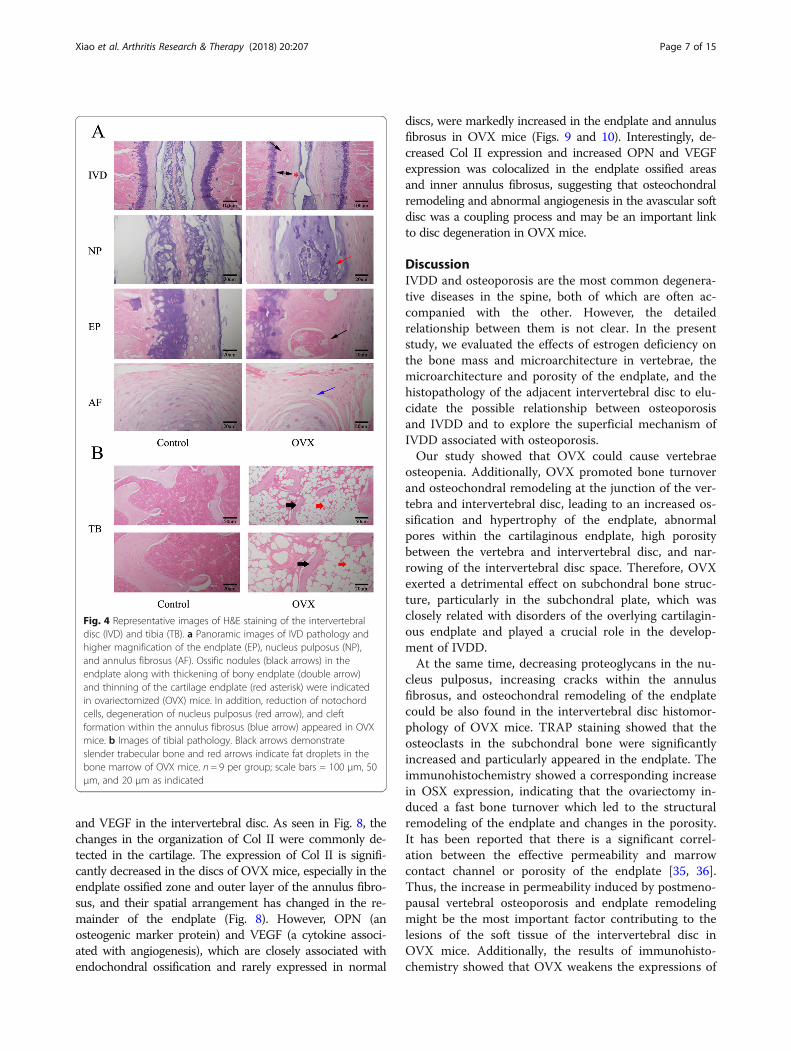
OVX group, in which trabeculae were sparse and dimin-ished (Fig. 4b). The bone marrow was almost replacedby adipose tissue (Fig. 4b).Histological findings of the intervertebral disc volume
showed that the nucleus pulposus contained abundant no-tochordal cells surrounded by large zones of extracellularmatrix, and the cartilaginous endplates were hyaline carti-lages composed of chondrocytes in the control group. Incontrast, in the OVX group, the discs showed degenera-tive changes and reduction in the nucleus pulposus andwere comprised of relatively few, clustered doublets ofchondrocyte-like cells (Fig. 4a). Proteoglycan loss could bealso found in the nucleus pulposus of OVX mice with palesafranin-O staining (Fig. 5). Moreover, ossification of thecartilaginous endplate occurred in the OVX group(Figs. 4a and 5). Safranin O and fast green staining sug-gested that endplates underwent endochondral ossifica-tion at 12 weeks postsurgery, indicated by the erosionof the wavy tidemark (Fig. 5) and green-stained bonematrix surrounding the cavities in OVX mice relative to thecontrol group (Fig. 5). The endplate structure showed dam-age, further calcification and ossification, and eventually be-came a bony structure. Bony tissues contained bonemarrow, hematopoietic lineage cells, and mineralized bone,and appeared more obvious in the deep zone of the middle
cartilaginous endplate (Figs. 4a and 5). However, the super-ficial cartilage endplate became markedly thinner and hadfewer active cells (Figs. 4a and 5). Furthermore, an in-creased number of clefts formed in the annulus fibrosus ofOVX mice with collagen disarrangement and cell reduc-tion, and even loose and broken lamella (Figs. 4a and 5).
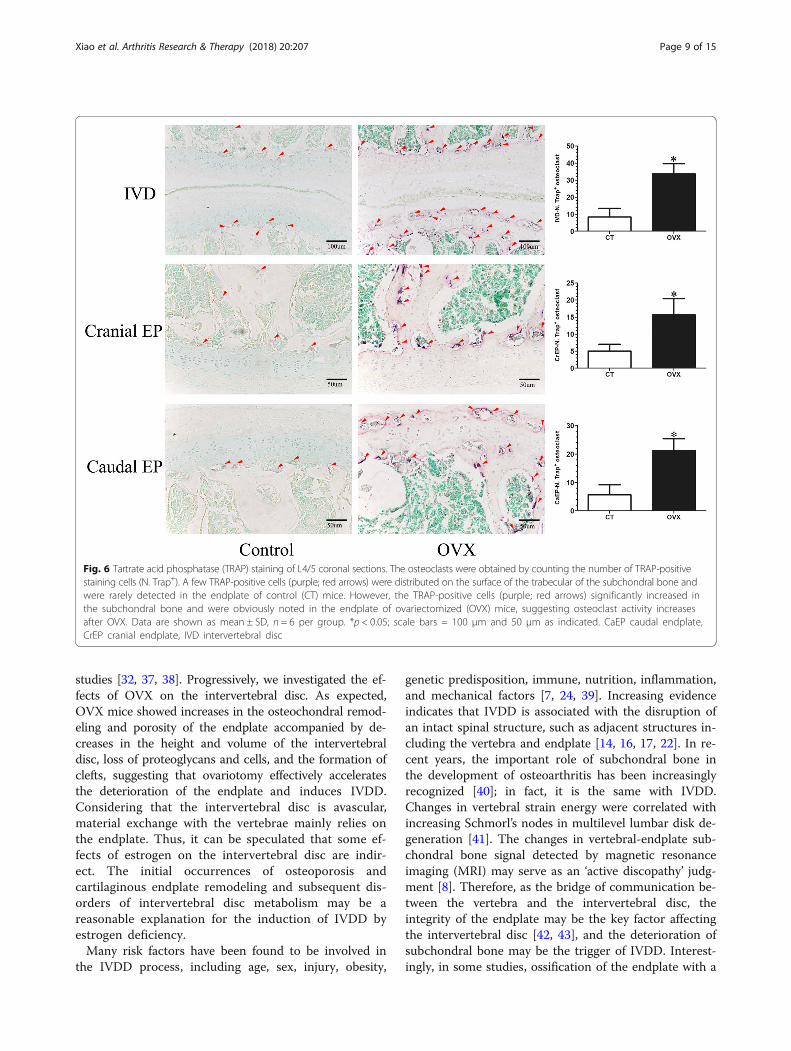
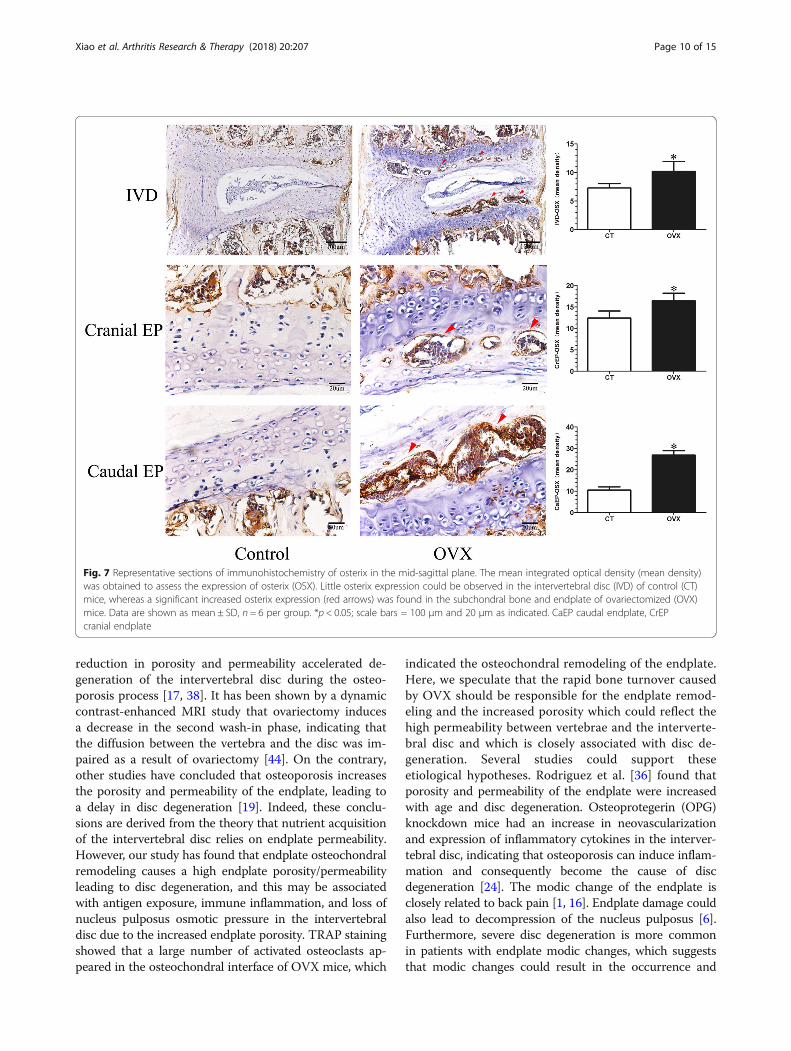
Increasing bone turnover accompanied by osteochondralremodeling exists in OVX miceTRAP and OSX were detected to reflect the bone turn-over of the intervertebral disc volume. TRAP stainingshowed that many activated osteoclasts were widely lo-cated on the trabecular surface of subchondral bone andwere significantly increased in the cartilaginous endplateof OVX mice (p < 0.05), but there were only a few osteo-clasts in the control mice (Fig. 6). Simultaneously, OSXexpression was significantly elevated in the OVX mice(p < 0.05), especially in the ossification region of the end-plate which was consistent with the area of osteoclastactivation (Fig. 7).
Decreasing Col II and increasing OPN and VEGF areexpressed in the degenerative disc of OVX miceImmunohistochemistry and immunofluorescence assayswere performed to assess the protein levels of Col II, OPN,
Fig. 3 Changes in microarchitecture, porosity of L4/5 caudal endplate, and disc volume quantified by micro-CT analysis. A a Three-dimensionalimages and parameters of caudal endplate. Results showing that increased cavities in OVX mice (yellow arrow) indicate osteochondralremodeling of the endplate. b–e Markedly decreased bone volume (BV)/total volume (TV) and number of closed pores (Po.N(cl)) andincreased open porosity (Po(op)) and total volume of pore space (Po.V(tot)) are shown in the ovariectomized (OVX) group. B a The topview of the caudal endplate showing a higher surface porosity in OVX mice (yellow arrows). b,c Quantification of disc volume by micro-CTshowing a significant decrease in OVX mice. The ROI of the disc is indicated by the red color. Data are shown as mean ± SD, n = 6 per group.*p < 0.05. CT control, IVD intervertebral disc
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 6 of 15
and VEGF in the intervertebral disc. As seen in Fig. 8, thechanges in the organization of Col II were commonly de-tected in the cartilage. The expression of Col II is signifi-cantly decreased in the discs of OVX mice, especially in theendplate ossified zone and outer layer of the annulus fibro-sus, and their spatial arrangement has changed in the re-mainder of the endplate (Fig. 8). However, OPN (anosteogenic marker protein) and VEGF (a cytokine associ-ated with angiogenesis), which are closely associated withendochondral ossification and rarely expressed in normal
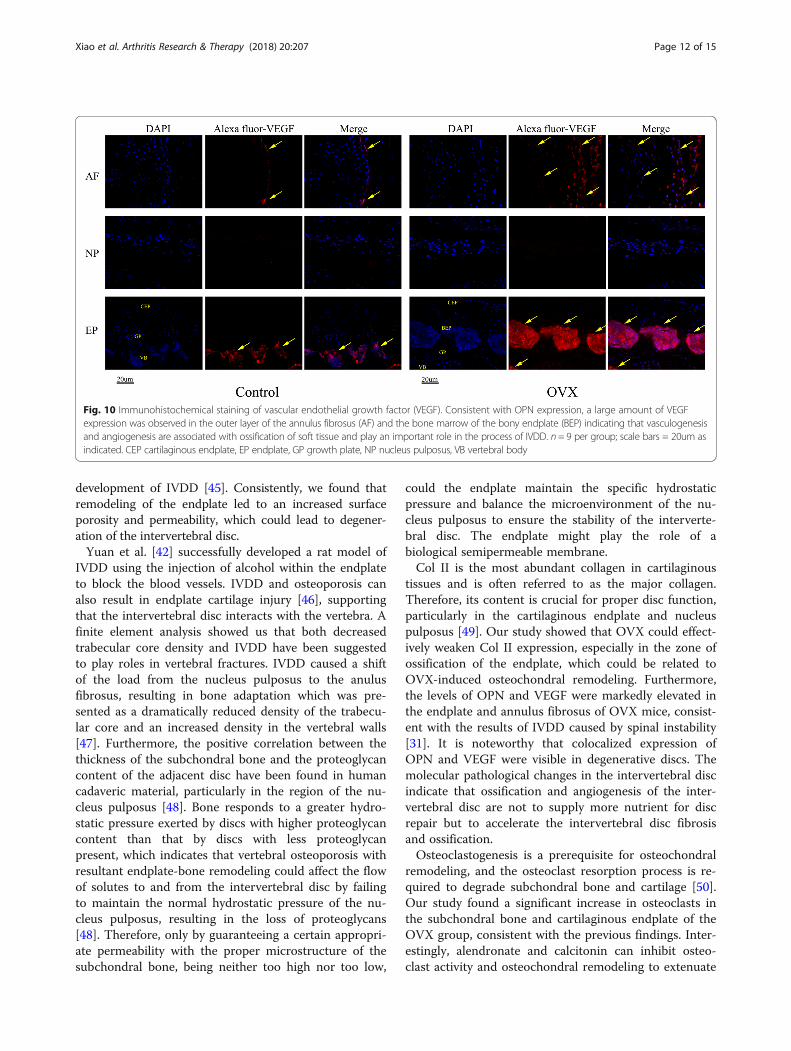
discs, were markedly increased in the endplate and annulusfibrosus in OVX mice (Figs. 9 and 10). Interestingly, de-creased Col II expression and increased OPN and VEGFexpression was colocalized in the endplate ossified areasand inner annulus fibrosus, suggesting that osteochondralremodeling and abnormal angiogenesis in the avascular softdisc was a coupling process and may be an important linkto disc degeneration in OVX mice.
DiscussionIVDD and osteoporosis are the most common degenera-tive diseases in the spine, both of which are often ac-companied with the other. However, the detailedrelationship between them is not clear. In the presentstudy, we evaluated the effects of estrogen deficiency onthe bone mass and microarchitecture in vertebrae, themicroarchitecture and porosity of the endplate, and thehistopathology of the adjacent intervertebral disc to elu-cidate the possible relationship between osteoporosisand IVDD and to explore the superficial mechanism ofIVDD associated with osteoporosis.Our study showed that OVX could cause vertebrae
osteopenia. Additionally, OVX promoted bone turnoverand osteochondral remodeling at the junction of the ver-tebra and intervertebral disc, leading to an increased os-sification and hypertrophy of the endplate, abnormalpores within the cartilaginous endplate, high porositybetween the vertebra and intervertebral disc, and nar-rowing of the intervertebral disc space. Therefore, OVXexerted a detrimental effect on subchondral bone struc-ture, particularly in the subchondral plate, which wasclosely related with disorders of the overlying cartilagin-ous endplate and played a crucial role in the develop-ment of IVDD.At the same time, decreasing proteoglycans in the nu-
cleus pulposus, increasing cracks within the annulusfibrosus, and osteochondral remodeling of the endplatecould be also found in the intervertebral disc histomor-phology of OVX mice. TRAP staining showed that theosteoclasts in the subchondral bone were significantlyincreased and particularly appeared in the endplate. Theimmunohistochemistry showed a corresponding increasein OSX expression, indicating that the ovariectomy in-duced a fast bone turnover which led to the structuralremodeling of the endplate and changes in the porosity.It has been reported that there is a significant correl-ation between the effective permeability and marrowcontact channel or porosity of the endplate [35, 36].Thus, the increase in permeability induced by postmeno-pausal vertebral osteoporosis and endplate remodelingmight be the most important factor contributing to thelesions of the soft tissue of the intervertebral disc inOVX mice. Additionally, the results of immunohisto-chemistry showed that OVX weakens the expressions of
Fig. 4 Representative images of H&E staining of the intervertebraldisc (IVD) and tibia (TB). a Panoramic images of IVD pathology andhigher magnification of the endplate (EP), nucleus pulposus (NP),and annulus fibrosus (AF). Ossific nodules (black arrows) in theendplate along with thickening of bony endplate (double arrow)and thinning of the cartilage endplate (red asterisk) were indicatedin ovariectomized (OVX) mice. In addition, reduction of notochordcells, degeneration of nucleus pulposus (red arrow), and cleftformation within the annulus fibrosus (blue arrow) appeared in OVXmice. b Images of tibial pathology. Black arrows demonstrateslender trabecular bone and red arrows indicate fat droplets in thebone marrow of OVX mice. n = 9 per group; scale bars = 100 μm, 50μm, and 20 μm as indicated
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 7 of 15

Col II and upregulates OPN and VEGF expressions inthe endplate and annulus fibrosus, suggesting that ab-normal ossification and angiogenesis are involved in theprocess of IVDD related to osteoporosis.Estrogen as an endocrine hormone influences the me-
tabolism of various tissues and organs in the body, suchas the rich collagenous tissues of bone, cartilage, disc, ar-tery, and skin, etc. [23]. It has been clearly shown that arapid decline of estrogen levels is an important factor forosteoporosis in postmenopausal women. Insufficient es-trogen caused by OVX is the common method to mimicpostmenopausal status and can effectively induce osteo-porosis. Many studies have confirmed that ovariectomycould cause osteoporosis in various animals, such as rats,
mice, monkeys, and so forth [25, 32, 37]. In the currentstudy, our results show that ovariotomy caused an in-creased body weight with abdominal fat accumulation buta decreased uterine weight with a severe atrophy inmorphology. Furthermore, a mass of fat in the bone mar-row of the tibia and thin trabeculae were observed inOVX mice, which supports that estrogen deficiency mayinduce adipogenic differentiation of bone marrow stemcells, but not osteogenesis. Moreover, the poorer resultsfor BMDtv and the bone parameters including BV/TV,Tb.N, Tb.pf, CONN.D, and SMI were seen in the L5 verte-bra of the OVX group when compared with the controlgroup suggesting a deterioration in bone quality andquantity due to ovariectomy, consistent with previous
Fig. 5 Representative images of safranin O and fast green staining of the intervertebral disc (IVD). This showed consistent results with H&Estaining. Specially, thickening of the bony endplate (EP; double arrow) accompanied by accelerated osteochondral remodeling (black arrows) andduplication of tidemarks (yellow arrow) were displayed more clearly in the EP of ovariectomized (OVX) mice by safranin O fast green staining.Moreover, reduction of aggrecan in the nucleus pulposus (NP), as indicated by a paucity of safranin O staining (blue thick arrow), cleft/crackformation in the annulus fibrosus (AF) (blue arrow), and loss of cells indicate intervertebral disc degeneration in OVX mice. n = 9 per group; scalebars = 100 μm and 20 μm as indicated
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 8 of 15
studies [32, 37, 38]. Progressively, we investigated the ef-fects of OVX on the intervertebral disc. As expected,OVX mice showed increases in the osteochondral remod-eling and porosity of the endplate accompanied by de-creases in the height and volume of the intervertebraldisc, loss of proteoglycans and cells, and the formation ofclefts, suggesting that ovariotomy effectively acceleratesthe deterioration of the endplate and induces IVDD.Considering that the intervertebral disc is avascular,material exchange with the vertebrae mainly relies onthe endplate. Thus, it can be speculated that some ef-fects of estrogen on the intervertebral disc are indir-ect. The initial occurrences of osteoporosis andcartilaginous endplate remodeling and subsequent dis-orders of intervertebral disc metabolism may be areasonable explanation for the induction of IVDD byestrogen deficiency.Many risk factors have been found to be involved in
the IVDD process, including age, sex, injury, obesity,
genetic predisposition, immune, nutrition, inflammation,and mechanical factors [7, 24, 39]. Increasing evidenceindicates that IVDD is associated with the disruption ofan intact spinal structure, such as adjacent structures in-cluding the vertebra and endplate [14, 16, 17, 22]. In re-cent years, the important role of subchondral bone inthe development of osteoarthritis has been increasinglyrecognized [40]; in fact, it is the same with IVDD.Changes in vertebral strain energy were correlated withincreasing Schmorl’s nodes in multilevel lumbar disk de-generation [41]. The changes in vertebral-endplate sub-chondral bone signal detected by magnetic resonanceimaging (MRI) may serve as an ‘active discopathy’ judg-ment [8]. Therefore, as the bridge of communication be-tween the vertebra and the intervertebral disc, theintegrity of the endplate may be the key factor affectingthe intervertebral disc [42, 43], and the deterioration ofsubchondral bone may be the trigger of IVDD. Interest-ingly, in some studies, ossification of the endplate with a
Fig. 6 Tartrate acid phosphatase (TRAP) staining of L4/5 coronal sections. The osteoclasts were obtained by counting the number of TRAP-positivestaining cells (N. Trap+). A few TRAP-positive cells (purple; red arrows) were distributed on the surface of the trabecular of the subchondral bone andwere rarely detected in the endplate of control (CT) mice. However, the TRAP-positive cells (purple; red arrows) significantly increased inthe subchondral bone and were obviously noted in the endplate of ovariectomized (OVX) mice, suggesting osteoclast activity increasesafter OVX. Data are shown as mean ± SD, n = 6 per group. *p < 0.05; scale bars = 100 μm and 50 μm as indicated. CaEP caudal endplate,CrEP cranial endplate, IVD intervertebral disc
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 9 of 15
reduction in porosity and permeability accelerated de-generation of the intervertebral disc during the osteo-porosis process [17, 38]. It has been shown by a dynamiccontrast-enhanced MRI study that ovariectomy inducesa decrease in the second wash-in phase, indicating thatthe diffusion between the vertebra and the disc was im-paired as a result of ovariectomy [44]. On the contrary,other studies have concluded that osteoporosis increasesthe porosity and permeability of the endplate, leading toa delay in disc degeneration [19]. Indeed, these conclu-sions are derived from the theory that nutrient acquisitionof the intervertebral disc relies on endplate permeability.However, our study has found that endplate osteochondralremodeling causes a high endplate porosity/permeabilityleading to disc degeneration, and this may be associatedwith antigen exposure, immune inflammation, and loss ofnucleus pulposus osmotic pressure in the intervertebraldisc due to the increased endplate porosity. TRAP stainingshowed that a large number of activated osteoclasts ap-peared in the osteochondral interface of OVX mice, which
indicated the osteochondral remodeling of the endplate.Here, we speculate that the rapid bone turnover causedby OVX should be responsible for the endplate remod-eling and the increased porosity which could reflect thehigh permeability between vertebrae and the interverte-bral disc and which is closely associated with disc de-generation. Several studies could support theseetiological hypotheses. Rodriguez et al. [36] found thatporosity and permeability of the endplate were increasedwith age and disc degeneration. Osteoprotegerin (OPG)knockdown mice had an increase in neovascularizationand expression of inflammatory cytokines in the interver-tebral disc, indicating that osteoporosis can induce inflam-mation and consequently become the cause of discdegeneration [24]. The modic change of the endplate isclosely related to back pain [1, 16]. Endplate damage couldalso lead to decompression of the nucleus pulposus [6].Furthermore, severe disc degeneration is more commonin patients with endplate modic changes, which suggeststhat modic changes could result in the occurrence and
Fig. 7 Representative sections of immunohistochemistry of osterix in the mid-sagittal plane. The mean integrated optical density (mean density)was obtained to assess the expression of osterix (OSX). Little osterix expression could be observed in the intervertebral disc (IVD) of control (CT)mice, whereas a significant increased osterix expression (red arrows) was found in the subchondral bone and endplate of ovariectomized (OVX)mice. Data are shown as mean ± SD, n = 6 per group. *p < 0.05; scale bars = 100 μm and 20 μm as indicated. CaEP caudal endplate, CrEPcranial endplate
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 10 of 15

Fig. 8 Representative sections of immunohistochemistry of Col II. Positive immunostaining was noted as brown staining. Black arrows indicateloss of Col II, especially in the ossification area of the cartilaginous endplate (EP) and the outer layer of the annulus fibrosus (AF) in ovariectomized(OVX) mice. n = 9 per group; scale bars = 20 μm as indicated. NP nucleus pulposus
Fig. 9 Immunohistochemical staining of osteopontin (OPN). DAPI stains nuclei blue and OPN expression was detected as red. Both control andovariectomized (OVX) mice have OPN expression in the vertebral body (VB) and outer annulus fibrosus (AF), but it was almost undetectable inthe nucleus. However, remarkable expression of OPN was found in the bony endplate (BEP) and outer AF in OVX mice. Meanwhile, the expression ofOPN was also detected in the inner AF. n = 9 per group; scale bar = 20 μm as indicated. CEP cartilaginous endplate, EP endplate, GP growth plate, NPnucleus pulposus
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 11 of 15
development of IVDD [45]. Consistently, we found thatremodeling of the endplate led to an increased surfaceporosity and permeability, which could lead to degener-ation of the intervertebral disc.Yuan et al. [42] successfully developed a rat model of
IVDD using the injection of alcohol within the endplateto block the blood vessels. IVDD and osteoporosis canalso result in endplate cartilage injury [46], supportingthat the intervertebral disc interacts with the vertebra. Afinite element analysis showed us that both decreasedtrabecular core density and IVDD have been suggestedto play roles in vertebral fractures. IVDD caused a shiftof the load from the nucleus pulposus to the anulusfibrosus, resulting in bone adaptation which was pre-sented as a dramatically reduced density of the trabecu-lar core and an increased density in the vertebral walls[47]. Furthermore, the positive correlation between thethickness of the subchondral bone and the proteoglycancontent of the adjacent disc have been found in humancadaveric material, particularly in the region of the nu-cleus pulposus [48]. Bone responds to a greater hydro-static pressure exerted by discs with higher proteoglycancontent than that by discs with less proteoglycanpresent, which indicates that vertebral osteoporosis withresultant endplate-bone remodeling could affect the flowof solutes to and from the intervertebral disc by failingto maintain the normal hydrostatic pressure of the nu-cleus pulposus, resulting in the loss of proteoglycans[48]. Therefore, only by guaranteeing a certain appropri-ate permeability with the proper microstructure of thesubchondral bone, being neither too high nor too low,
could the endplate maintain the specific hydrostaticpressure and balance the microenvironment of the nu-cleus pulposus to ensure the stability of the interverte-bral disc. The endplate might play the role of abiological semipermeable membrane.Col II is the most abundant collagen in cartilaginous
tissues and is often referred to as the major collagen.Therefore, its content is crucial for proper disc function,particularly in the cartilaginous endplate and nucleuspulposus [49]. Our study showed that OVX could effect-ively weaken Col II expression, especially in the zone ofossification of the endplate, which could be related toOVX-induced osteochondral remodeling. Furthermore,the levels of OPN and VEGF were markedly elevated inthe endplate and annulus fibrosus of OVX mice, consist-ent with the results of IVDD caused by spinal instability[31]. It is noteworthy that colocalized expression ofOPN and VEGF were visible in degenerative discs. Themolecular pathological changes in the intervertebral discindicate that ossification and angiogenesis of the inter-vertebral disc are not to supply more nutrient for discrepair but to accelerate the intervertebral disc fibrosisand ossification.Osteoclastogenesis is a prerequisite for osteochondral
remodeling, and the osteoclast resorption process is re-quired to degrade subchondral bone and cartilage [50].Our study found a significant increase in osteoclasts inthe subchondral bone and cartilaginous endplate of theOVX group, consistent with the previous findings. Inter-estingly, alendronate and calcitonin can inhibit osteo-clast activity and osteochondral remodeling to extenuate
Fig. 10 Immunohistochemical staining of vascular endothelial growth factor (VEGF). Consistent with OPN expression, a large amount of VEGFexpression was observed in the outer layer of the annulus fibrosus (AF) and the bone marrow of the bony endplate (BEP) indicating that vasculogenesisand angiogenesis are associated with ossification of soft tissue and play an important role in the process of IVDD. n= 9 per group; scale bars = 20um asindicated. CEP cartilaginous endplate, EP endplate, GP growth plate, NP nucleus pulposus, VB vertebral body
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 12 of 15

IVDD [26, 27]. Although parathyroid hormone 1–34 hassubstantial anabolic effects on bone mass and trabecularmicroarchitecture, nonsignificant effects have not yetbeen found on disc degeneration [51]. This leads us tosuggest that preventing the remodeling of the osteo-chondral structure caused by the initial osteoclastogene-sis after the menopause could be an important aspect inthe fight against the occurrence of intervertebral disc de-generation. Furthermore, some reports have confirmedthat suppressing osteoclastogenesis and aberrant angio-genesis could blunt IVDD and osteoarthritis [52, 53],which provides further promise for the treatment ofIVDD in the future. It may be beneficial to renew theintervertebral disc by balancing bone metabolism andregulating permeability of the endplate.
ConclusionsIn summary, this study uncovered the mystery of the de-teriorative effects of OVX on IVDD, clearly showing thatOVX can induce and accelerate the progression of discdegeneration. The underlying mechanisms could be re-lated to the destruction of the structural integrity andthe function of the vertebra and endplate induced byOVX, both of which are essential structures for main-taining disc function. OVX modulates bone turnoverand results in osteochondral interface remodeling whichcould also influences the expressions of Col II, OSX,OPN, and VEGF to stimulate the disc degeneration. It isconceivable that osteoporosis of vertebrae and endplateremodeling resulting from estrogen depletion may affectthe bone marrow microenvironment and endplate per-meability which subsequently could alter the metabolismand biomechanics of the intervertebral disc. Simultan-eously, the reduction in estrogen caused by OVX couldalso directly affect the metabolism of the intervertebraldisc because of the presence of estrogen receptors in thenucleus pulposus. These changes may be a reasonableexplanation for disc degeneration related to osteopor-osis. However, our observations are preliminary andneed to be further confirmed through clinical trials oradditional animal experiments. Based on the above re-sults, intervention for osteoporosis and improvement inthe vertebral body and endplate structure may be an ef-fective way to retard IVDD; the mechanisms of IVDDrelated to osteoporosis need to be further studied.
Shortcomings and limitationsThere are several limitations for this study. First, thefindings from the current study are preliminary becauseof the limited sample sizes and insufficient evidence.The penetration of the endplate lacks intuitive measure-ment methods and the porosity of the endplate was de-fined by an indirect index for assessing permeability.Second, our study lacks investigation at different time
points and thus we cannot observe progressive changesof vertebrae and discs. Third, although some differen-tially expressed proteins were found using immunohisto-chemistry, other methods of detection were not useddue to research grant limitations and insufficient micedisc tissue.
Additional file
Additional file 1: Figure S1. Representative mid-coronal images of L4/5segment and transverse images of L5. The results demonstrate that the tra-becular bone structure of L4 and L5 vertebrae are poorer and significantlythinner in OVX mice (blue arrows) suggesting osteoporosis in OVX mice. Thecaudal endplate shows obviously increased cavities (red arrow) with a morenarrowed disc (red asterisk) which may indicate osteochondral remodeling ofthe endplate and intervertebral disc degeneration in OVX mice. (TIF 592 kb)
AbbreviationsBMD: Bone mineral density; BV: Bone volume; Col: Collagen;CONN.D: Connectivity density; CT: Computed tomography; H&E: Hematoxylinand eosin; IVDD: Intervertebral disc degeneration; OPN: Osteopontin;OSX: Osterix; OVX: Ovariectomy; PO(op): Open porosity; Po.N(cl): Number ofclosed pores; Po.V(tot): Total volume of pore space; ROI: Region of interest;SMI: Structural model index; Tb.N: Trabecular number; Tb.Pf: Trabecularpattern factor; Tb.Sp: Trabecular separation; Tb.Th: Trabecular thickness;TRAP: Tartrate-resistant acid phosphatase; TV: Total volume; VEGF: Vascularendothelial growth factor
AcknowledgmentsThe authors would like to thank Mr. Fang Yang for helping with micro-CTscanning and analyses, and we thank the Laboratory of Orthopedics andTraumatology of Lingnan Medical Research Center of Guangzhou Universityof Chinese Medicine for providing associated facilities.
FundingThis study is supported by a National Natural Science Foundation of China(NSFC; grant number 81673992) and the Open Foundation of The NationalKey Discipline and The Laboratory of Orthopedics and Traumatology ofGuangzhou University of Chinese Medicine.
Availability of data and materialsThe authors are committed to sharing their data, publishing the data, andmaking available the molecular resources described in this publication to thescientific community.
Authors’ contributionsAll authors made substantial contributions to the data analysis andinterpretation, drafting of the manuscript and critical revision of the manuscriptfor important intellectual content. All authors gave final approval of the versionto be published. ZX and DL had full access to all the data in the study and takeresponsibility for the integrity of the data and the accuracy of the data analysis.Conception and design: DL, ZX, MC, and YH. Collection and assembly of data:ZX, JH, MC, GS, and SC. Analysis and interpretation of the data: GS, YH, MC, ZX,and SC. Manuscript drafting: ZX, JH, DL, MC, and YH. Manuscript revision andsupplementary experiments: JH and GS.
Ethics approvalAnimal studies were performed under institutional guidelines and inaccordance with protocols approved by the Ethics Committee, GuangzhouUniversity of Chinese Medicine.
Consent for publicationNot applicable.
Competing interestsThe authors declare that they have no competing interests.
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 13 of 15

Publisher’s NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations.
Author details1The Department of Spinal Surgery, The Second Affiliated Hospital ofGuangzhou University of Chinese Medicine, No. 111, Dade Road, YuexiuDistrict, Guangzhou 510120, China. 2The Laboratory Affiliated toOrthopaedics and Traumatology of Chinese Medicine of Linnan MedicalResearch Center of Guangzhou University of Chinese Medicine, No. 12,Jichang Road, Baiyun District, Guangzhou 510405, China. 3GuangzhouUniversity of Chinese Medicine, No. 12, Jichang Road, Baiyun District,Guangzhou 510405, China.
Received: 16 January 2018 Accepted: 15 August 2018
References1. Luoma K, Vehmas T, Kerttula L, Grönblad M, Rinne E. Chronic low back pain in
relation to modic changes, bony endplate lesions, and disc degeneration in aprospective MRI study. Eur Spine J. 2016;25(9):2873–81.
2. Izzo R, Popolizio T, D'Aprile P, Muto M. Spinal pain. Eur J Radiol. 2015;84(5):746–56.
3. Wenig CM, Schmidt CO, Kohlmann T, Schweikert B. Costs of back pain inGermany. Eur J Pain. 2009;13(3):280–6.
4. Paesold G, Nerlich AG, Boos N. Biological treatment strategies for discdegeneration: potentials and shortcomings. Eur Spine J. 2007;16(4):447–68.
5. Humzah MD, Soames RW. Human intervertebral disc: structure and function.Anat Rec. 1988;220(4):337–56.
6. Dolan P, Luo J, Pollintine P, Landham PR, Stefanakis M, Adams MA.Intervertebral disc decompression following endplate damage: implicationsfor disc degeneration depend on spinal level and age. Spine (Phila Pa 1976).2013;38(17):1473–81.
7. Vergroesen PPA, Kingma I, Emanuel KS, Hoogendoorn RJW, Welting TJ, vanRoyen BJ, van Dieen JH, Smit TH. Mechanics and biology in intervertebraldisc degeneration: a vicious circle. Osteoarthr Cartilage. 2015;23(7):1057–70.
8. Nguyen C, Poiraudeau S, Rannou F. From Modic 1 vertebral-endplatesubchondral bone signal changes detected by MRI to the concept of ‘activediscopathy’. Ann Rheum Dis. 2015;74(8):1488–94.
9. Karsdal MA, Bay-Jensen AC, Lories RJ, Abramson S, Spector T, Pastoureau P,Christiansen C, Attur M, Henriksen K, Goldring SR, et al. The coupling ofbone and cartilage turnover in osteoarthritis: opportunities for boneantiresorptives and anabolics as potential treatments? Ann Rheum Dis.2014;73(2):336–48.
10. Yuan XL, Meng HY, Wang YC, Peng J, Guo QY, Wang AY, Lu SB. Bone-cartilage interface crosstalk in osteoarthritis: potential pathways and futuretherapeutic strategies. Osteoarthr Cartil. 2014;22(8):1077–89.
11. Funck-Brentano T, Cohen-Solal M. Crosstalk between cartilage and bone:when bone cytokines matter. Cytokine Growth Factor Rev. 2011;22(2):91–7.
12. Bellido M, Lugo L, Roman-Blas JA, Castaneda S, Caeiro JR, Dapia S, Calvo E,Largo R, Herrero-Beaumont G. Subchondral bone microstructural damageby increased remodelling aggravates experimental osteoarthritis precededby osteoporosis. Arthritis Res Ther. 2010;12(4):R152.
13. Shapiro IM, Vresilovic EJ, Risbud MV. Is the spinal motion segment adiarthrodial polyaxial joint: what a nice nucleus like you doing in a joint likethis? Bone. 2012;50(3):771–6.
14. Chen S, Fu P, Wu H, Pei M. Meniscus, articular cartilage and nucleuspulposus: a comparative review of cartilage-like tissues in anatomy,development and function. Cell Tissue Res. 2017;370(1):53–70.
15. Kang R, Li H, Ringgaard S, Rickers K, Sun H, Chen M, Xie L, Bunger C.Interference in the endplate nutritional pathway causes intervertebral discdegeneration in an immature porcine model. Int Orthop. 2014;38(5):1011–7.
16. Moore RJ. The vertebral endplate: disc degeneration. Eur Spine J. 2006;153:S333–7.17. Tomaszewski KA, Adamek D, Konopka T, Tomaszewska R, Walocha JA.
Endplate calcification and cervical intervertebral disc degeneration: the roleof endplate marrow contact channel occlusion. Folia Morphol (Warsz). 2015;74(1):84–92.
18. Holm S, Maroudas A, Urban JP, Selstam G, Nachemson A. Nutrition of theintervertebral disc: solute transport and metabolism. Connect Tissue Res.1981;8(2):101–19.
19. Mattei TA. Osteoporosis delays intervertebral disc degeneration byincreasing intradiscal diffusive transport of nutrients through bothmechanical and vascular pathophysiological pathways. Med Hypotheses.2013;80(5):582–6.
20. Lou C, Chen H, Feng X, Xiang G, Zhu S, Tian N, Jin Y, Fang M, Wang C, XuH. Menopause is associated with lumbar disc degeneration: a review of4230 intervertebral discs. Climacteric. 2014;17(6):700–4.
21. Wang YJ, Griffith JF. Effect of menopause on lumbar disk degeneration.Radiology. 2010;257(2):319–20.
22. Wang YJ, Griffith JF. Menopause causes vertebral endplate degenerationand decrease in nutrient diffusion to the intervertebral discs. MedHypotheses. 2011;77(1):18–20.
23. Calleja-Agius J, Muscat-Baron Y, Brincat MP. Estrogens and the intervertebraldisc. Menopause Int. 2009;15(3):127–30.
24. Li X, Xue C, Zhao Y, Cheng S, Zhao D, Liang Q, Chen L, Wang Q, Lu S, Shi Q,et al. Deletion of OPG leads to increased neovascularization and expressionof inflammatory cytokines in the lumbar intervertebral disc of mice. Spine.2017;42(1):E8–E14.
25. Luo Y, Zhang L, Wang W, Hu Q, Song H, Su Y, Zhang Y. Alendronate retardsthe progression of lumbar intervertebral disc degeneration inovariectomized rats. Bone. 2013;55(2):439–48.
26. Song H, Luo Y, Wang W, Li S, Yang K, Dai M, Shen Y, Zhang Y, Zhang L.Effects of alendronate on lumbar intervertebral disc degeneration withbone loss in ovariectomized rats. Spine J. 2017;17(7):995–1003.
27. Tian FM, Yang K, Wang WY, Luo Y, Li SY, Song HP, Zhang YZ, Shen Y, ZhangL. Calcitonin suppresses intervertebral disk degeneration and preserveslumbar vertebral bone mineral density and bone strength inovariectomized rats. Osteoporos Int. 2015;26(12):2853–61.
28. Nanjo Y, Morio Y, Nagashima H, Hagino H, Teshima R. Correlation betweenbone mineral density and intervertebral disk degeneration in pre- andpostmenopausal women. J Bone Miner Metab. 2003;21(1):22–7.
29. Miyakoshi N, Itoi E, Murai H, Wakabayashi I, Ito H, Minato T. Inverse relationbetween osteoporosis and spondylosis in postmenopausal women asevaluated by bone mineral density and semiquantitative scoring of spinaldegeneration. Spine. 2003;28(5):492–5.
30. Pye SR, Reid DM, Adams JE, Silman AJ, O'Neill TW. Radiographic features oflumbar disc degeneration and bone mineral density in men and women.Ann Rheum Dis. 2006;65(2):234–8.
31. Bian Q, Jain A, Xu X, Kebaish K, Crane JL, Zhang Z, Wan M, Ma L, Riley LH,Sponseller PD, et al. Excessive activation of TGFbeta by spinal instabilitycauses vertebral endplate sclerosis. Sci Rep. 2016;6:27093.
32. Bonucci E, Ballanti P. Osteoporosis-bone remodeling and animal models.Toxicol Pathol. 2014;42(6):957–69.
33. Martin JT, Gorth DJ, Beattie EE, Harfe BD, Smith LJ, Elliott DM. Needle punctureinjury causes acute and long-term mechanical deficiency in a mouse model ofintervertebral disc degeneration. J Orthop Res. 2013;31(8):1276–82.
34. Naito Y, Wakabayashi H, Kato S, Nakagawa T, Iino T, Sudo A. Alendronateinhibits hyperalgesia and suppresses neuropeptide markers of pain in amouse model of osteoporosis. J Orthop Sci. 2017;22(4):771–7.
35. Laffosse JM, Accadbled F, Molinier F, Bonnevialle N, de Gauzy JS, Swider P.Correlations between effective permeability and marrow contact channelssurface of vertebral endplates. J Orthop Res. 2010;28(9):1229–34.
36. Rodriguez AG, Slichter CK, Acosta FL, Rodriguez-Soto AE, Burghardt AJ,Majumdar S, Lotz JC. Human disc nucleus properties and vertebral endplatepermeability. Spine (Phila Pa 1976). 2011;36(7):512–20.
37. Ren H, Liang D, Shen G, Yao Z, Jiang X, Tang J, Cui J, Lin S. Effects ofcombined ovariectomy with dexamethasone on rat lumbar vertebrae.Menopause. 2016;23(4):441–50.
38. Ding Y, Jiang J, Zhou J, Wu X, Huang Z, Chen J, Zhu Q. The effects ofosteoporosis and disc degeneration on vertebral cartilage endplate lesionsin rats. Eur Spine J. 2014;23(9):1848–55.
39. Dario AB, Ferreira ML, Refshauge KM, Lima TS, Ordonana JR, Ferreira PH. Therelationship between obesity, low back pain, and lumbar disc degenerationwhen genetics and the environment are considered: a systematic review oftwin studies. Spine J. 2015;15(5):1106–17.
40. Zhen G, Wen C, Jia X, Li Y, Crane JL, Mears SC, Askin FB, Frassica FJ, ChangW, Yao J, et al. Inhibition of TGF-beta signaling in mesenchymal stem cellsof subchondral bone attenuates osteoarthritis. Nat Med. 2013;19(6):704.
41. Von Forell GA, Nelson TG, Samartzis D, Bowden AE. Changes in vertebralstrain energy correlate with increased presence of Schmorl's nodes in multi-level lumbar disk degeneration. J Biomech Eng-T ASME. 2014;136:0610026.
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 14 of 15

42. Yuan W, Che W, Jiang Y, Yuan F, Wang H, Zheng G, Li X, Dong J.Establishment of intervertebral disc degeneration model induced byischemic sub-endplate in rat tail. Spine J. 2015;15(5):1050–9.
43. Maatta JH, Kraatari M, Wolber L, Niinimaki J, Wadge S, Karppinen J, WilliamsFMK. Vertebral endplate change as a feature of intervertebral discdegeneration: a heritability study. Eur Spine J. 2014;23(9):1856–62.
44. Deng M, Griffith JF, Zhu X, Poon WS, Ahuja AT, Wang YJ. Effect ofovariectomy on contrast agent diffusion into lumbar intervertebral disc: adynamic contrast-enhanced MRI study in female rats. Magn Reson Imaging.2012;30(5):683–8.
45. Hu Z, Zhao F, Fang X, Fan S. Modic changes, possible causes andpromotion to lumbar intervertebral disc degeneration. Med Hypotheses.2009;73(6):930–2.
46. Wang L, Cui W, Kalala JP, Van Hoof T, Liu B. Effect of osteoporosis andintervertebral disc degeneration on endplate cartilage injury in rats. AsianPac J Trop Med. 2014;7(10):796–800.
47. Homminga J, Aquarius R, Bulsink VE, Jansen CTJ, Verdonschot N. Canvertebral density changes be explained by intervertebral disc degeneration?Med Eng Phys. 2012;34(4):453–8.
48. Roberts S, McCall IW, Menage J, Haddaway MJ, Eisenstein SM. Does thethickness of the vertebral subchondral bone reflect the composition of theintervertebral disc? Eur Spine J. 1997;6(6):385–9.
49. Kim KW, Ha KY, Park JB, Woo YK, Chung HN, An HS. Expressions ofmembrane-type I matrix metalloproteinase, Ki-67 protein, and type IIcollagen by chondrocytes migrating from cartilage endplate into nucleuspulposus in rat intervertebral discs—a cartilage endplate-fracture modelusing an intervertebral disc organ culture. SPINE. 2005;30(12):1373–8.
50. Lofvall H, Newbould H, Karsdal MA, Dziegiel MH, Richter J, Henriksen K,Thudium CS. Osteoclasts degrade bone and cartilage knee jointcompartments through different resorption processes. Arthritis Res Ther.2018;20(1):67.
51. Luo Y, Li SY, Tian FM, Song HP, Zhang YZ, Zhang L. Effects of humanparathyroid hormone 1–34 on bone loss and lumbar intervertebral discdegeneration in ovariectomized rats. Int Orthop. 2018;42(5):1183–90.
52. Kwon WK, Moon HJ, Kwon TH, Park YK, Kim JH. Influence of rabbitnotochordal cells on symptomatic intervertebral disc degeneration: anti-angiogenic capacity on human endothelial cell proliferation under hypoxia.Osteoarthr Cartil. 2017;25(10):1738–46.
53. Ji B, Zhang Z, Guo W, Ma H, Xu B, Mu W, Amat A, Cao L. Isoliquiritigeninblunts osteoarthritis by inhibition of bone resorption and angiogenesis insubchondral bone. Sci Rep. 2018;8(1):1721.
Xiao et al. Arthritis Research & Therapy (2018) 20:207 Page 15 of 15