ophiostomatoid fungiassociated with ipscembrae injapan
TRANSCRIPT

Mycoscience 39: 367-378, 1998
367
Ophiostomatoid fungi associated with Ips cembrae in Japanand their pathogenicity to Japanese larchYuichi Yamaoka1), Michael J. Wingfield2!, Masashi Ohsawa31 and Yoshio Kuroda1!
11Institute of Agriculture and Forestry, University of Tsukuba, Tsukuba, Ibaraki 305-8572, Japan2)Forestry and Agricultural Biotechnology Institute (FAB!), University of Pretoria, Pretoria 0002, South Africa3)Yamanashi Forestry and Forest Products Research Institute, Masuhomachi, Minamikomagun, Yamanashi 400-0502,
Japan
Accepted for publication 29 July 1998
Six ophiostomatoid fungi, Le" Ceratocystiopsis minuta, Ceratocystis laricicola, Ophiostoma brunneo~ciliatum, O. laricis,O. piceae and Ophiostoma sp., were isolated from the galleries of Ips cembrae on Japanese larch (Larix kaempfef/llogs incentral Honshu, Japan. Japanese larch trees approximately 30 yr old were inoculated with all six fungi. Ceratocystislaricicola produced the largest lesions on the inner bark around the inoculation point and the largest dry zone in the sap~wood. Furthermore, C. laricicola was the only fungus associated with I. cembrae that had the ability to kill Japaneselarch, death occurring in 30wyr-old trees within 3.5 mo after inoculation.
Key Words-Ceratoeystiopsis minuta; Ceratocystis laricieola; inoculation experiment; Ips cembrae; Ophiostoma spp.
Ips cembrae (Heer) infests trees and recently cut logs oflarch (Larix spp.) in Europe and Asia (Crooke and Bevan,1957; Nobuchi, 1974; Redfern et al" 1987; Koizum;,1990, 1994). The insect also damages plantations ofEuropean larch (L. decidua Miller) in the United KingdomICrooke and Bevan, 1957; Redfern, 19891 and Japaneselarch IL. kaempferi ILamb.1 Carr. I in Japan IKoizumi,1990). Redfern et al. 119871 described Ceratocystislarieieola Redfern & Minter from I. eembrae in Scotlandand showed that the fungus has a high degree of viru~lence.
In Japan, Aoshima (1965) isolated six species ofophiostomatoid fungi, some of them undescribed, from I.eembrae infesting Japanese larch: Ceratocystiseoeruleseens (Munch) Bakshi, C. jezoensis Aoshimanom. nud., C. macrospara Aoshima nom. nud., Ophjosto~ma brunneo-eiliatum MathiesenwKaarik, O. olivaceumMathiesen, and O. pieeae Munch. Amongst them, O.pieeae was shown to be pathogenic {Maeto et aI., 1991;Yamaguchi, 1993, 19951 and was considered to beresponsible for the mortality of larch trees infested by I.cembrae IMaeto et al" 1991; Yamaguchi, 19951. OnlyO. piceae was used in the inoculation study, and it wasnot compared with the other ophiostomatoid fungiassociated with I. eembrae.
Although C. laricicalaRedfern u Z. Minter is knownas the dominant fungus associated with I. cembrae on in~troduced larch plantations in Scotland (Redfern et aI.,1987), its occurrence on native stands of larch, in eitherEurope or Japan, has not been reported. Ips eembrae inScotland is thought to have been introduced from Con-tinental Europe and it is logically assumed that C. lariei~cola was introduced with this insect. Aoshima (1965)invalidly described C.jezoensis from larch in Japan and it
is probable that this fungus is the same as C. ladcieo/a.One of the aims of this study was to characterize the
ophiostomatoid fungi from larch in Japan and to compareand contrast these findings with previous investigations.Based on a previous study of fungi associated with I.typographus L. f.japonicus Niijima in Japan (Yamaoka et
al" 1997) and the study of Redfern et al. (1987), wehave hypothesized that C. larieicala would be the mostvirulent and important fungal associate of I. cembrae inJapan. A further aim of this study was to prove thishypothesis. If correct, this would be in contrast to thefindings of Maeto et al. 11991) and Yamaguchi (1995)who did not isolate C. larieieola and who suggested thatO. piceae is an important and virulent associate of I.cembrae.
Materials and Methods
Isolation of fungi Disks approximately 5 cm thick andstrips of bark (about 10 x 20 cm) including galleries of I.cembrae were cut on 20 July 1989 from Japanese larchlogs left in plantations labout 1,500 m a.s.1.1after thin-ning in Experimental Forests in Yatsugatake (EFY),Agricultural and Forestry Research Center, University ofTsukuba, Kawakami-mura, Nagano Prefecture. On 10July 1992, similar samples were collected from aJapanese larch plantation 11.420 m a.s.l.) at the foot ofMt. Fuji, Fujiyoshida~shi, Yamanashi Prefecture. In bothareas, the populations of the beetles seemed to be en-demic. Samples were placed in plastic bags and transwferred to the laboratory for further study. With the aid ofa dissection microscope, masses of ascospores accumuwlating at the tips of perithecia, or masses of conidia ac-cumulating on conidiophores were carefully lifted from

368 Y. Yamaoka et a1.
these structures. This was accomplished by using a finetungsten needle, with which the spore masses weretransferred onto 1% ma!t extract agar (1% MA; 10 9malt extract, 15 g agar/1,OOO ml distilled waterl in 9-cmPetri dishes. The dishes were incubated at 20°C in thedark and the fungi were allowed to sporulate. Theprocess of purifying and identifying cultures was similarto that described by Yamaoka et al. 119971. Culturesused for identification were grown on 2% malt extractagar, 236 malt extract Ebios agar {2% MEA; 20 9 malt ex~tract, 1 9 Ebios (Brewer's yeast preparation, Tanabe),15 g agar/1 ,000 ml distilled water). and 1%Pablum agarIPA; 10 g Pablum mixed cereal, 15 g agar/1 ,000 ml dis-tilled water), with or without the addition of small (about2 em x 5 mm x 3 mm) pieces of autoclaved Japaneselarch bark.
Cultures representing each distinct taxon ofophiostomatoid fungus have been deposited in the cul-ture collection of the Laboratory of Plant Pathology andMycology, Institute of Agriculture and Forestry, Universi-ty of Tsukuba, Tsukuba, Japan and in the Japan Collec-tion of Microorganisms (JCM). Dried specimens ofthese cultures have also been deposited with the Herbari-um of the Institute of Agriculture and Forestry, Universityof Tsukuba ITSHI.Inoculations To assess the relative virulence of theophiostomatoid fungi associated with I. cembrae onlarch, eight isolates representing the six taxa (Table 1)were selected. These included two isolates each of C.larieieola and O. pieeae and one each of the other fourtaxa. Isolates YCC-273 (C. /aric/cola) and YCC-299 10.pieeae) were acquired from Japanese larch logs collectedin Experimental Forests in Yatsugatake, and all other iso-lates were from Mt. Fuji.Experiment I. In this experiment, all six fungi were test-ed for their relative virulence. Each fungus was grownon 23~ MA in 9-cm Petri dishes at 20°C for 2 wk. Disksof bark (1 cm in diam and 2-3 mm in thickness), whichwere punched out with a cork borer from healthyJapanese larch bark after removing the scaly outermostlayer of bark, were autoclaved for 20 min at 121°C andadded to the plates. The plates were incubated underthe same conditions for an additional 3 wk. Peritheciaand/or conidiophores of the test fungi were produced onthe bark discs, which were used as inoculum discs. Forcontrols, sterilized bark discs were placed on 2% MA andthese were incubated at 20°C until use.
Four Japanese larch trees approximately 30 yr old115.5 to 18 em D8HI were selected in a plantation labout1,500 m a.s.\.) in the EFY. Each tree was inoculatedwith all test fungi and controls on 30 May 1995. Barkdiscs (1 cm in diam) were removed using a cork borer atintervals of about 3 cm in a horizontal ring at a height of1.5 m (Fig. 1a). Inoculum discs or control discs wereplaced into the wounds and covered with vaseline fol-lowed by parafilm and adhesive tape. The trunks of thetrees were then wrapped with nylon sheet from groundlevel to a height of 2 m to prevent invasion by bark bee-tles.
Two of the inoculated trees were felled on 24 July
1995 155 d after inoculation). Three 1-m logs were cutfrom the bottom of each tree and to the laboratory for ex-amination. Bark was peeled from the bolts and the sizeof lesions formed in the inner bark around the inoculationpoints was measured. The logs were then cut into disksat about 1O-cm intervals above and below the inoculationpoints. Staining and dried zones of sapwood were ob-served on the surface of each disc. Small pieces ofwood were cut from the stained sapwood and trans-ferred onto 2% MA plates to confirm the presence of theinoculated fungi. The remaining two trees were felledon 11 September 1995 (104 d after inoculation). Le-sions on the inner bark, as well as staining and thepresence of dry zones of sapwood, were observed usingthe technique described above.Experiment II. Inthis experiment, six trees approximate-ly 30 yr old (13-15 em dbhl in the same stand of the ex-perimental forest as in the first inoculation experimentwere subjected to three different treatments on 30 May1995. The aim here was to compare the relative viru-lence of C. /aricico/aand O. piceae. Two trees eachwere inoculated with inoculum discs of C. /aricicolaYCC-285 or O. p/ceae YCC-295, or with sterile bark discs ascontrols. Inoculation was achieved by removing discs of
<a)
.. ..,.
.. .................. .., '. ....
". . . .
.'
(b)
.. ... ......"". ,1 em,~Z em'
....... .. .
... . . . . . .','
','.' . ... .. ..... ....... ......
nOU'.
.......
., ..,'"'.','.
',',
'
'.','.. .... .... .
'.'.'.' .'.. ...
"..'.
, . . . ,. ,. . . .. ... , . .... ....
\/ 0\\!! !! i
; :: : : : : : . : : ;i : i i ;! ~
:
'...
.......
'". ,..
,"'..'. . .. .,,.. ., ,.,. .,,..'. .........'::::::::::::::::""';.::-::
.n'..~...~~:cm,
....
. ...... ',','
..
. . .. ......
'". ... .'"
dCo'..
, .
...."
... ..'.
..., ..,...,... ............ ,:::::::::; ;'<: :::'. .' -:::)H~Un nn~ :~:.
.. , .... ,. .... ' .... ... ...,, . , . . . . ........., '..
.. ... ..'.'
...
Fig. 1. Patterns of inoculation in Experiment! (a) and n {bl.

Ips cembrae-associated ophiostomatoid fungi
bark with a cork borer (1 cm in diam), at intervals of 2 cmin three horizontal rings, which were approximately1.5 m above ground and separated by 4 cm verticallyIFig. 1bl. Thus a total of 45 bark discs were removedfrom each tree with 15 cm dbh. The inoculations werecovered as described above.
Development of the external symptoms of inoculat-ed trees was monitored at intervals of 2-3 wk. Twotrees inoculated with C. laricicola, and one of each of thetrees inoculated with O. piceae or sterile bark discs werefelled on 11 September 1995 1104 d after inoculation).One-meter bolts were cut from the boles from the groundlevel up to 3 m above ground and transferred to thelaboratory for further study. Lesions formed in the innerbark around the inoculated area as well as stained anddry zones of sapwood were measured. Symptoms ofthe remaining two trees inoculated with O. piceae orsterile bark discs were observed until early November,when needles of uninoculated normal larch trees in thesame stand turned yellow. They were felled on 7November 1995 (161 d after inoculation) and treated asdescribed above.
Results and Discussion
Isolation of fungiSix distinct species of ophiostomatoid fungi were isolat-ed from Japanese larch infested with I. cembrae. Theseare as follows.
Ceratocystiopsis minuta (Siem.) Upadhyay & Kendrick,Mycologia 67: 800. 1975. Figs. 2-5
=Ophiostoma minutum Siem.. Planta Pol. 7: 23. 1939.=Ceratocystis minuta ISiem.} Hunt, lIoydia 19: 49.
1956.Morphological characteristics of the fungus isolated
in the present study were almost the same as those of C.minuta associated with Yezo spruce (Picea jezoensis(Sieb. & Zucc.) Carr. I infested by I. typographus f.japonicus in Japan (Yamaoka et aI., 1997). One excep-tion was that the perithecia of the present fungus wereuniformly brown to dark brown (Fig. 2), while those ofthe fungus associated with Yezo spruce had lighter baseswhen produced on the bark placed on the surface of agar(Yamaoka et aI., 1997). They also fitted well with thedescription for C. minuta (Hunt, 1956; Upadhyay, 1981).
Ceratocystiopsis minuta is known to be associatedwith Norway spruce infested by I. typographus in EuropeISiemaszko, 1939; Mathiesen, 1950; Kotynkova-Sychrova, 1966; Kiiiirik, 1975; Solheim, 1986, 1993;Harding, 1989), and other spruce and pine trees associat-ed with various bark beetles including Dendroctonus spp.and Ips spp. in North America (Davidson, 1942; Wrightand Cain, 1961; Robinson, 1962; Griffin, 1968; Olcho-weeki and Reid, 1974; Upadhyay, 1981). In Japan,Yamaoka et a!. (1997) provided the first record of aCeratocystiopsis sp.. and the present study is the secondreport of C. minute associated with a bark beetle inJapan.
Living cultures deposited: YCC-293, isolate from a
369
Figs. 2-5. Ceratocystiopsis minuta.2. Ascocarp (bar, 20 pm). 3. Top of neck (bar, 15 pm). 4.Ascospores (bar, 5 (1m). 5. Hyalorhinocladiella anamorph(bar, 15flm).
gallery wall of I. cembrae in L. kaempferi at the foot
of Mt. Fuji, Fujiyoshida-shi, Yamanashi, Japan (FMF).Collected on 10 July 1992 by Y. Yamaoka (YYI, M. J.Wingfield (MJWL and M. Ohsawa (MOl; YCC-294, iso-late from a gallery wall of I. cembrae in L. kaempferi atFMF. Collected on 10 July 1992 by yy, MJW, and MO.
Dried specimens: TSH-C127, dried culture YCC-293grown on 196 Pablum agar with pieces of autoclaved
bark of L. kaempferi (PASI at 20°C; TSH-C128, dried cul-ture YCC-294 grown on PAS at 20°C.
Ceratocystis laricicola Redfern & Minter, in Redfern et al.,Plant Pathol. 36: 468.1987. Figs. 6-10
Perithecia of this fungus were produced superficiallyboth on the surface of the agar medium and on barkplaced on the surface of the agar. The bases of perithe~cia (Fig. 6) were black, globose to subglobose,145-300 pm in diam, ornamented by dark brown septatehyphae. Perithecia! necks were slender, black,

370 Y. Yamaoka et al.
Figs. 6-10. Ceratacystis laricicola.6. Ascocarp (bar, 100,urn). 7. Top of neck (bar, 20 11m).8. Ascospores (bar, 10 pm), 9. Chalara anamorph (bar,15f1m). 10. Conidia (bar, 15pm).
540-1,150 I'm long; terminating ostiolar hyphae (Fig. 7)aseptate, hyaline, straight, 14-22, slightly divergent,32-67 I'm long. Ascospores (Fig. 8) ellipsoid,4.8-6.0x 2.4-2.8 I'm, surrounded by hyaline sheaths,oval in face view, 4.8-6.4 x 3.6-4.4 pm, and orange-sec-tion shaped in side view. Conidia ph ores (Fig. 9)mononematous, cylindrical or slightly tapered. palebrown to brown, composed of 2-8 cells, 94-274 pmlong, including termina! conidiogenous cell, and 4-8 pmwide at the broadest part. Conidiogenous cells phialidic,cylindrical or lageniform, and 56-100 x 4-8 pm. Conidia(Fig. 10) hyaline, oblong, barrel-shaped or ellipsoidal,
6-15 x 3.5-6 [.m.This fungus was first described by Redfern et al.
(19B7) as being associated with I. cembrae attacking in-troduced European larch in Scotland. It has recentlybeen suggested that C. laricicola might be con specificwith C. polonica (Siem.) C. Moreau (Visser et al" 1995;Harrington et aI., 1996; Witthuhn et al" 1998). whichis a common associate of I. typographus in Europe(Siemaszko, 1939; Mathiesen, 1950, 1951; Mathiesen-Kaarik, 1953, 1960; Kaarik, 1975; Solheim, 1986,1993; Harding, 1989; Furniss et aI., 1990; Vilrl and vanWeissenberg, 1995). The present fungus was virtuallyidentical in morphological characteristics to C. laricicoladescribed by Redfern et al. (1 987). except for the size ofascospores, which were about 4 x 2 pm excluding thesheath. It was also morphologically indistinguishablefrom C. polonica associated with I. typographus f. japoni-cus (Yamaoka et aI., 1997) except that diameters of theperithecial bases of the Japanese isolates (145-300 I'm)were slightly smaller than those of C. p%nica (200-390 I'm). and the length of conidiophores (94-274[lmincluding terminal conidiogenous cells) were longer thanthose of C. p%nica (74-160 11m).
Aoshima (1965) described a Ceratocystis species as-sociated with I. typographus f. japonicus which appearsto be the same as C. polonica (Yamaoka et al., 1997).He named it C. jezoensis, but this description was notvalid, since it was not formally published. He also notedthat this fungus was associated with I. cembrae in Japan.It is apparent that he considered the fungus associatedwith I. cembrae to be conspecific with that associatedwith I. typographus f. japonicus. Ceratocystis laricicolaand C. polonica are very similar and morphologically in-distinguishable. Visser et a!. (1995) were unable to dis-tinguish these species by comparison of sequence datafrom the ITS region of the ribosomal DNA. These datahave also recently been confirmed in a more comprehen-sive study, at the molecular level, of Ceratocystis spp.from conifers (Witthuhn et al., 1998). Similarly, isolatesof C.laricicola grouped closely with those of C. polonicain a comparison of Ceratocystis spp. based on isozymes(Harrington et al" 1996). However, in the latter study,isolates of C. laricicola could be distinguished from C.polonica, and the fungi have very different hosts and dis-tinct vectors. The fungi might thus represent distincttaxa that have recently undergone speciation and, thus,have not developed distinct morphological characteris-tics. For the present, we believe that they are best treat-ed as distinct.
Living cultures deposited: YCC-273, isolate from agallery wall of I. cembrae in L. kaempferi at ExperimentalForests in Yatsugatake, Agricultural and ForestryResearch Center, University of Tsukuba, Kawakami-mura, Nagano, Japan (EFY). Collected on 20 July 1989by YY;YCC-285" (JCM9810). isolate from a gallery wallof I. cembrae in L. kaempferi at FMF. Collected on 10July 1992 by yy, MJW, and MO.
Dried specimens: TSH-C106, dried culture YCC-285
,i'
HThe same culture as C745 used in Harrington et a!. (1996).
Ips cembrae-associated ophiostomatoid fungi
grown on PAB at 20°C; TSH-C132, dried culture YCC-273 grown on 2.% malt extract Ebios agar with pieces ofautoclaved bark of L. kaempferi IMEABI at 20°C.
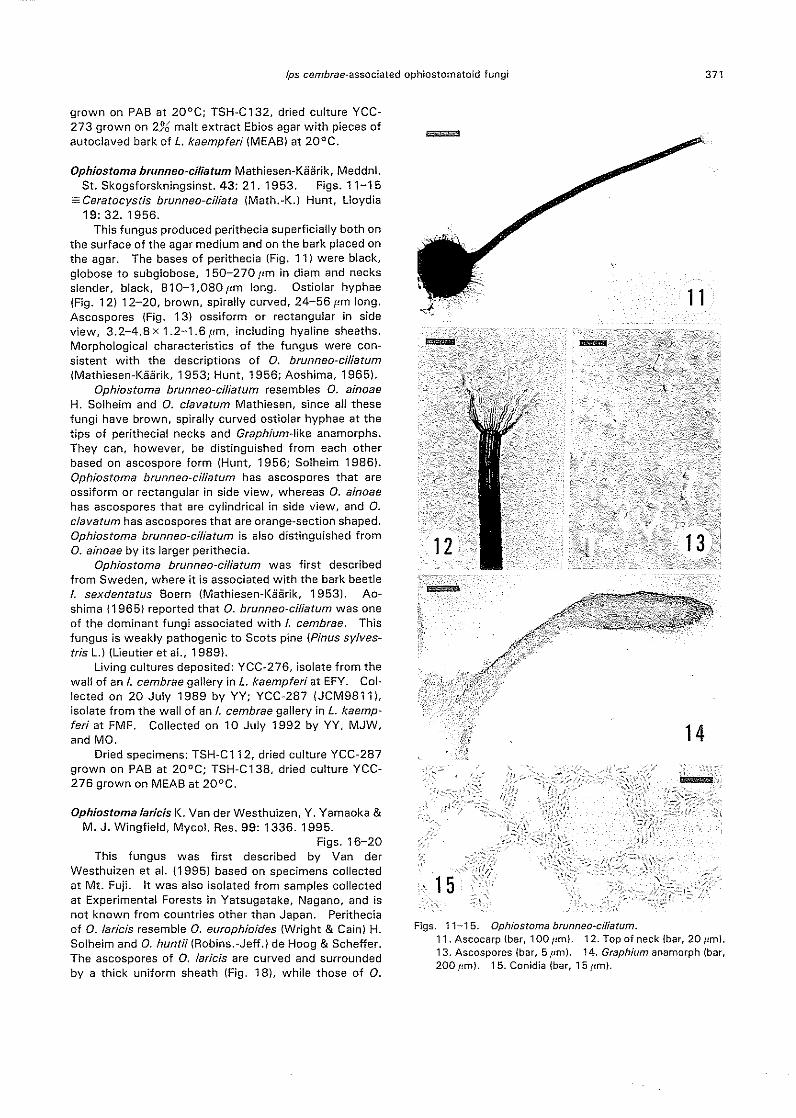
Ophiostoma brunneo-ciliatum Mathiesen-Kaarik, Meddnl.St. Skogsforskningsinst. 43: 21. 1953. Figs. 11-15
=Ceratocystis brunneo-ciliata (Math.~K.) Hunt, Lloydia19: 32. 1956.
This fungus produced perithecia superficially both onthe surface of the agar medium and on the bark placed onthe agar. The bases of perithecia (Fig. 11) were black,globose to subg!obose, 150-270 pm in diam and necksslender, black, 810-1,080 IJm long. Ostiolar hyphaeIFig. 12) 12-20, brown, spirally curved, 24-56 I'm long.Ascospores (Fig. 13) ossiform or rectangular in sideview, 3.2-4.8 x 1.2-1.6 11m, including hyaline sheaths.Morphological characteristics of the fungus were con-sistent with the descriptions of O. brunneo~ciliatumIMathiesen-Kiiiirik, 1953; Hunt, 1956; Aoshima, 1965).
Ophiostoma brunneo-ciliatum resembles O. ainoaeH. Solheim and O. clavatum Mathiesen, since all thesefungi have brown, spirally curved ostiolar hyphae at thetips of perithecial necks and Graphium-like anamorphs.They can, however, be distinguished from each otherbased on ascospore form IHunt, 1956; Solheim 19861.Ophiostoma brunneo~ciliatum has as co spores that areossiform or rectangular in side view, whereas O. ainoaehas ascospores that are cylindrical in side view, and O.clavatum has ascospores that are orange~section shaped.Ophiostoma brunneo~ciliatum is also distinguished fromO. ainoae by its larger perithecia.
Ophiostoma brunneo-ciliatum was first describedfrom Sweden, where it is associated with the bark beetleI. sexdentatus 80em (Mathiesen-Kaarik, 1953). Ao-shima {1965} reported that O. brunneo~ciliatum was oneof the dominant fungi associated with I. cembrae. Thisfungus is weakly pathogenic to Scots pine (Pinus sylves-tris L.IILieutier et aI., 1989).
Living cultures deposited: YCC-276, isolate from thewall of an I. cembrae gallery in L. kaempferi at EFY. Col-lected on 20 July 1989 by YY; YCC-287 IJCM9811).isolate from the wall of an I. cembrae gallery in L. kaemp-feri at FMF. Collected on 10 July 1992 by yy, MJW,and MO.
Dried specimens: TSH-C 112, dried culture YCC-287grown on PAB at 20°C; TSH-C138, dried culture YCC-276 grown on MEAB at 20°C.
Ophiostoma laricis K. Van der Westhuizen, Y. Yamaoka &M. J. Wingfield, Mycol. Res. 99: 1336. 1995.
Figs. 16-20This fungus was first described by Van der
Westhuizen et a!. (1995) based on specimens collectedat Mt. Fuji. It was also isolated from samples collectedat Experimental Forests in Yatsugatake, Nagano, and isnot known from countries other than Japan. Peritheciaof O. laricis resemble O. europhioides (Wright & Cain) H.Solheim and O. huntii IRobins.-Jeff.1 de Hoog & Scheffer.The ascospores of O. lariel's are curved and surroundedby a thick uniform sheath (Fig. 18). while those of O.
371
Figs. 11-15. Ophiostoma brunneo-cil/atum.11. Ascocarp (bar, 100 pm). 12. Top of neck (bar, 20 pm).13. Ascospores (bar, 5 pm). 14. Graph/um anamorph {bar,200 pm}. 15. Conidia (bar, 15 I,m).

372 Y. Yamaoka et al.
--
Figs. 16-20. Ophiostoma ladcis.16. Ascocarp (bar, 5Crim). 17. Top of neck (bar, 20 pm).18. Ascospores (bar, 10 pm). 19. Leptographium ana-morph (bar, 15 pm). 20, Conidia (bar, 15 pm).
europhioides and O. huntii have cucullate ascospores.Van der Westhuizen et al. (1995) described the lengthsof perithecial necks of O. laricis as 400-1,320 pm whenthe perithecia were produced on host tissue or in thepresence of host tissue and on autoclaved pieces of Pinuspatula Schl. & Cham. on 2% MEA plates. The presentfungus, however, produced perithecia with shorter necks(200-370 ~m long) when the cultures were grown on
1% Pablum agar amended with pieces of autoclaved barkof Japanese larch (PAB) at 20°C (Fig. 16).
Aoshima (1965) described a Ceratocystis species (C.macrospora nom. nud.) associated with I. cembrae thatwas morphologically very similar to O. larids. Cerato~
cystis macrospora was not valid, since it was not formal-ly published.
Living cultures deposited: YCC-277, isolate from thewall of I. cembrae gallery in L. kaempferi at EFY. Col-lected on 20 July 1989 by YY; YCC-289 (JCM9812),isolate from the wall of I. cembrae gallery in L. kaempferiat FMF. Collected on 10 July 1992 by yy, MJW, andMO.
Dried specimens: TSH-C 117, dried culture YCC-289grown on PAB at 20°C; TSH-C140, dried culture YCC-277 grown on MEAB at 20°C.
Ophiastama piceae (Munch) H. & P. Sydow, Ann. Mycol.17: 43. 1919.
=Ceratostomella piceae Munch, Naturw. Z. Land~ u.Forstw. 5: 547.1907. .
=Ceratocystis piceae (Munch) Bakshi, Trans. Br. Mycol.Soc. 33: 113. 1950.
Ophiostoma piceae isolated in this study had thesame morphological characteristics as the fungus isolat-ed from I. typographus f. japanicus in Hakkaido (Yama-
oka et aI., 1997). It was also very similar to the descrip-tion of O. piceae by Hunt (19561 and Upadhyay (1981),except that the bases of perithecia were larger (up to280 pm in diam). Detailed descriptions of Japanese iso~lates of O. piceae have been presented previously(Nishikada and Yamauti, 1935; Aoshima, 1965),
Aoshima (1965) reported that O. piceae was one ofthe most common fungi associated with I. cembrae.This fungus has been isolated from Yezo spruce (Tochinaiand Sakamoto, 1934; Aoshima, 1965; Yamaoka et aI.,1997), Japanese red pine (Pinus densiflora Sieb. & Zucc.)(Nishikado and Yamauti, 1935; Aoshima, 1965), beech(Fagus crenata Blume) (Aoshima and Hayashi, 1953;Aoshima, 1965), and various other conifers and hard-woods (Otani, 1988).
Brasier and Kirk (1993) separated O. piceae into twointersterile mating groups, one from hardwood sources(OPH) and one from conifers (OPC). They further sug-gested that OPH probably represents O. querci (Geor-gevitch) Nannf.=Ceratostomella quercus Georgevitch(1927) and OPC the original O. piceae=Ceratostomellapiceae sensu Munch (1907). Halmschlager et al. (1994)supported their findings by morphological investigationof synnematal size and RAPD analysis of DNA. The fun-gus associated with I. cembrae in the present study isthus considered to be O. piceae. However, Japaneseisolates of O. piceae from conifer and hardwood sourcesmust stilt be examined using mating tests and molecularcomparisons to clarify their taxonomic position.
Living cultures deposited: YCC-299, isolated fromthe walt of an I. cembrae galleryin L. kaempferi at EFY.Collected on 20 July 1989 by YY; YCC-301, isolate fromthe wall of an I. cembrae gallery in L. kaempferi at EFY.Collected on 20 July 19B9 by YY.
Dried specimens: TSH-C156, dried culture YCC-301grown on PAB at 20°C; TSH-C157, dried culture YCC-299 grown on PAB at 20°C.

Ips cembrae-associated ophiostomatoid fungi
Ophiostoma sp. Figs. 21-25Perithecia (Fig. 21) on agar medium and on bark
placed on the surface of the agar, superficial or semi-im-mersed. Perithecial bases black, globose to subglobose,210-270 pm in diam, and the necks black, cylindricaiwith an apical taper, 300-540 pm long, 56-72 pm wide
Figs. 21-25. Ophiastama sp.21. Ascocarp (bar, 100 pm). 22. Top of neck (bar, 20 pm).23. Ascospores (bar, 10 pm). 24. Leptagraphium ana-morph (bar, 25 pm). 25. Conidia (bar, 15 pm).
373
above the globose bases, 12-20.um wide at the apex.Ostiolar hyphae absent (Fig. 221. Ascospores (Fig. 231hyaline, 1-celled surrounded by hyaline sheaths, appear-ing cucullate in side view, 5.6-6.4x 2.8-3.2 pm, quad-rangular in face view, triradiate in end view. Conidialstate a Leptographium sp. Conidiophores (Fig. 24) erect,macronematous, mononematous, brown to dark brown,200-410 pm long including a penicillately branchedconidiogenous apparatus. Conidia (Fig. 25) hyaline,obovoid to ellipsoid, 4-9 x 2-4 pm.
This fungus is morphologically very similar to O.europhioides. In particular, its perithecial characteristicsfitted well with descriptions of O. europhioides (Wrightand Cain, 1961; Davidson et aI., 19671. Both fungi haveascospores surrounded by hyaline walls, and appearingcucullate in side view, quadrangular in face view, andtriangular in end view. However, ascospores of thepresent fungus were narrower and had more distinct,sharp brims in side view than those reported for O. euro-phioides. The fungus also produces longer conidio-phares than those typical of O. europhioides. Only oneisolate of this Ophiostoma sp. was obtained in thepresent study and we have, therefore, selected not toname it yet.
living cultures deposited: YCC-300 (JCM9B141. iso-late from the wall of an I. cembrae gallery in L. kaempferiat FMF. Collected on 10 July 1992 by yy, MJW, andMO.
Dried specimens: TSH-C121, dried culture YCC-300grown on MEAB at 20°C.
Of the six species of the fungi isolated from I.cembrae by Aoshima (1965), C. coerulescens and O.olivaceum were not isolated in the present study.
Inoculation experiments
Relative virulence of the ophiostomatoid fungi Necroticlesions were formed around all the inoculation points onthe inner bark when ophiostomatoid fungi were inoculat-ed (Table 1, Fig. 261. lesions mainly spread iongitudi-nally, and only slightly in the tangential direction. Thelengths of lesions differed depending on the fungi inocu-lated. When sterile bark was used as an inoculum, no le-sion or very small necrotic lesions (less than 5 mm inlength) developed around the inoculation points (Table1).
Lesion lengths could not be statistically analyzed dueto the low number of trees available for inoculation.However, the relative lengths of the lesions associatedwith the test fungi on each tree showed the same tend-ency in all four trees treated. Among the fungi tested,C. laricicola produced larger lesions than the others(Table 1, Fig. 261. Isolate YCC-2B5 of C. !addco!aproduced longer lesions than did isolate YCC-273.Ophiostoma laricis and the undescribed Ophiostoma sp.produced relatively large lesions, but these were alwaysconsiderably smaller than those associated with C. larici-cola. Ophiostoma brunneo-ciliatum and O. piceaeproduced smaJJer lesions than O. laricis and Ophiostoma
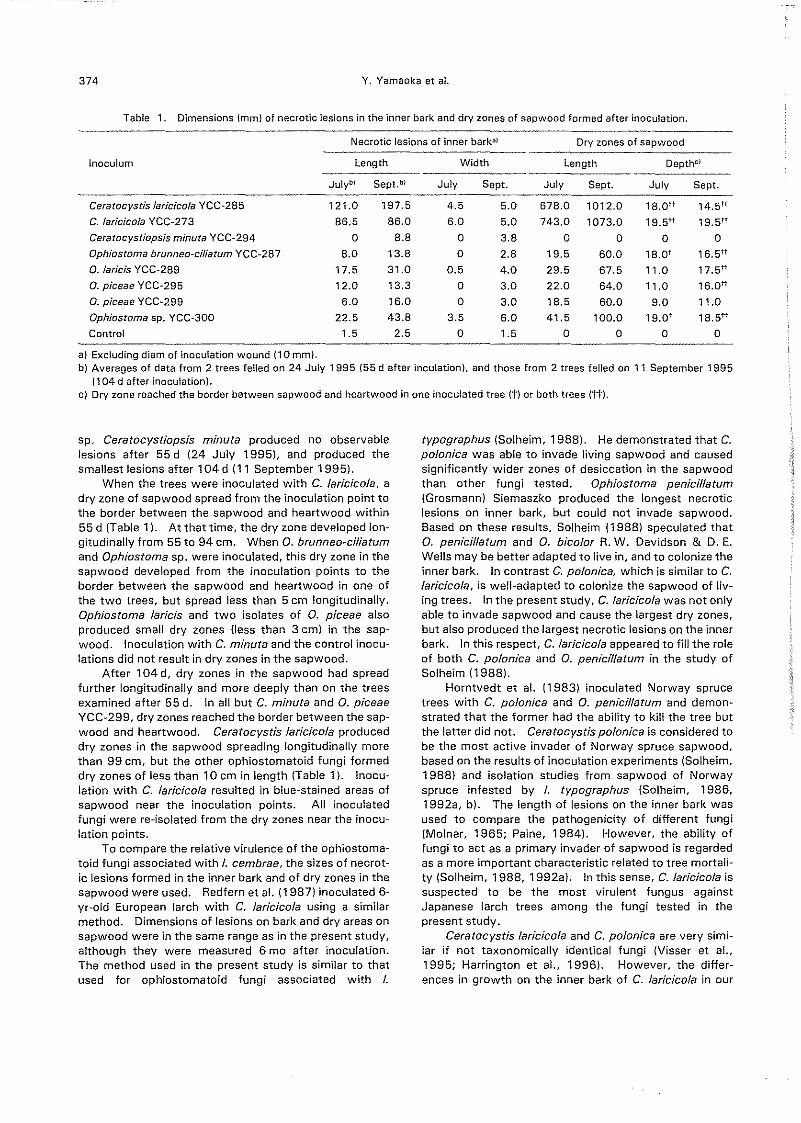
Necrotic lesions of inner bark:!1 Dry zones of sapwood
Inoculum Length Width Length DepthC)
JUlybl Sept.b! July Sept. July Sept. July Sept.
Ceratocystis laricicola YCC-285 121.0 197.5 4.5 5.0 678.0 1012.0 18.0tt 14,5tt
C. laricicola YCC-273 86.5 86.0 6.0 5.0 743.0 1073.0 19.5tt 19.5tt
Ceratocystiopsis minuta YCC-294 0 8.8 0 3.8 0 0 0 0
Ophiostoma brunneo-ciliatum YCC~287 8.0 13.8 0 2.8 19.5 60.0 18.0t 16.5tt
O. laricis YCC-289 17.5 31.0 0.5 4.0 29.5 67.5 11.0 17,5tt
0, piceae YCC-295 12.0 13.3 0 3.0 22.0 64.0 11.0 16.att
O. piceae YCC-299 6.0 16.0 0 3.0 18.5 60.0 9.0 11.0
Ophiostoma sp. YCC-300 22.5 43.8 3.5 6.0 41.5 100.0 19.0' 18.5tt
Control 1.5 2.5 0 1.5 0 0 0 0
374 Y. Yamaoka et a!.
Table 1. Dimensions (mm) of necrotic lesions in the inner bark and dry zones of sapwood formed after inoculation.
a) Excluding diam of inoculation wound (10 mm).b) Averages of data from 2 trees felled on 24 July 1995 (55 dafter inculation), and those from 2 trees felled on 11 September 1995
(104 d after inoculation).c) Dry zone reached the border between sapwood and heartwood in one inoculated tree (t) or both trees (tt).
sp. Ceratoeystiopsis minuta produced no observablelesions after 55 d (24 July 19951, and produced thesmallest lesions after 104d (11 September 19951.
When the trees were inoculated with C. larieieola, adry zone of sapwood spread from the inoculation point tothe border between the sapwood and heartwood within55 d (Table 11. At that time, the dry zone developed lon-gitudinally from 55 to 94 em, When 0, brunneo-ciliatumand Ophiostoma sp. were inoculated, this dry zone in thesapwood developed from the inoculation points to theborder between the sapwood and heartwood in one ofthe two trees, but spread less than 5 cm longitudinally.Ophiostoma laricis and two isolates of 0, piceae alsoproduced small dry zones (less than 3 cm) in the sap-wood. Inoculation with C. minuta and the control inocu-lations did not result in dry zones in the sapwood.
After 104 d, dry zones in the sapwood had spreadfurther longitudinally and more deeply than on the treesexamined after 55 d. In all but C. minuta and O. piceaeYCC-299, dry zones reached the border between the sap-wood and heartwood, Ceratocystis laricicola produceddry zones in the sapwood spreading longitudinally morethan 99 em, but the other ophiostomatoid fungi formeddry zones of less than 10 em in length (Table 11. Inocu-lation with C. laricicola resulted in blue-stained areas ofsapwood near the inoculation points. All inoculatedfungi were re-isolated from the dry zones near the inocu-lation points.
To compare the relative virulence of the ophiostoma-toid fungi associated with I. eembrae, the sizes of necrot-ic lesions formed in the inner bark and of dry zones in thesapwood were used, Redfern et a!. (1987) inoculated 6~yr-old European larch with C. laricicola using a similarmethod. Dimensions of lesions on bark and dry areas onsapwood were in the same range as in the present study,although they were measured 6 mo after inoculation.The method used in the present study is similar to thatused for ophiostomatoid fungi associated with I.
tvpographus (Solheim, 19881. Hedemonstrated that C.polonica was able to invade living sapwood and causedsignificantly wider zones of desiccation in the sapwoodthan other fungi tested, Ophiostoma penicillatum(Grosmann) Siemaszko produced the longest necroticlesions on inner bark, but could not invade sapwood.8ased on these results, Solheim 119881 speculated thatO. penieillaturn and O. bieolor R. W. Davidson & D. E.Wells may be better adapted to live in, and to colonize theinner bark. In contrast C. poloniea, which is similar to C.laricicola, is well-adapted to colonize the sapwood of liv-ing trees. In the present study, C. laricicola was not onlyable to invade sapwood and cause the largest dry zones,but also produced the largest necrotic lesions on the innerbark. In this respect, C. laricicola appeared to fill the roleof both C. polonica and O. penici'l/atum in the study ofSolheim (1988).
Horntvedt et al. (1983) inoculated Norway sprucetrees with C. polonica and O. penicillatum and demon~strated that the former had the ability to kill the tree butthe latter did not. Ceratocystis polonica is considered tobe the most active invader of Norway spruce sapwood,based on the results of inoculation experiments (Solheim,1988) and isolation studies from sapwood of Norwayspruce infested by I. tvpographus (Solheim, 1986,1992a, b). The length of lesions on the inner bark wasused to compare the pathogenicity of different fungi(Molnar, 1965; Paine, 1984). However, the ability offungi to act as a primary invader of sapwood is regardedas a more important characteristic related to tree mortali-ty (Solheim, 1988, 1992a). In this sense, C. larieieola issuspected to be the most virulent fungus againstJapanese larch trees among the fungi tested in thepresent study.
Ceratocystis laricieola and C. polonica are very simi-lar if not taxonomically identical fungi (Visser et aL,1995; Harrington et aI., 1996). However, the differ-ences in growth on the inner bark of C. laricicola in our

Inoculum Tree No.32 55 66 SS 104 161
Ceratocystfs larfcfcola No.1 -", , , , D
YCC-285 NO.2 ++ 0Ophfostoma pfceae No.3
YCC-295 No.4
Control No. 5
NO.6
Ips cembrae-associated ophiostomatoid fungi
study and C. p%nica in the study of Solheim (19881.suggest that they might differ physiologically. Thisdifference might also correspond with the fact that thevectors and hosts of the fungi are different.Inoculation with C. laricicola and O. piceae No symp-toms were observed on trees mass-inoculated with C./aricico/a, O. piceae, or controls after 32 d 11 July 19951.However, 55 d after inoculation (24 July). the crown ofone tree inoculated with C. laricieola had discolored, andit turned brown 66 d after inoculation (4 August) (Table2). The second tree inoculated with C. laricieola showeddiscoloration of needles after 66 d, and most of the nee-dles had fallen 88 d after inoculation (26 Augustl IFig.271. These trees died by 104d after inoculation (11September) (Table 2). when they were cut to observesymptoms on the inner bark and in the sapwood. One ofeach of the trees inoculated with O. pieeae and a controlwere felled together with the two C. larieicola-treatedtrees for comparison. At that stage, they showed no ex-ternal symptoms. The remaining two trees (one inocu-lated with O. pieeae and the other control) were observeduntil 7 November 1995 1161 dafter inoculation I, whenall trees in the stand began to loose needles due to theonset of winter. These trees were also felled to observesymptoms formed on the inner bark and in the sapwood.
Substantially larger necrotic lesions were formed onthe inner bark of trees inoculated with C. laricieola (Fig.28, Table 3) than those inoculated with O. pieeae or thecontrols. They had merged with each other IFig. 281
375
and completely girdled the tree trunks around the inocula-tion zone. The inner bark of the trees inoculated with C.larieicola was completely dead above the inoculationzone. The entire sapwood of the trees near the inocula-tion zone was blue~stained (Fig. 31). Blue-stained areasof the sapwood spread more than 1 m above and belowthe inoculation zone. Ceratocystis laricicola was almostalways re~isolated from the blue-stained sapwood.
In the case of O. pieeae (Fig. 29) and the controls(Fig. 30). small necrotic lesions were formed around theinoculation points (Table 3), but these never merged witheach other. In the trees inoculated with O. piceae, thesapwood near the inoculation zone was stained palebrown (Fig. 32). The stained areas spread 5 to 8 cmabove and below the inoculation points. No stained sap-wood or dry zone of sapwood was observed in the con-trol trees IFig. 331.
The results of the present study showed that C.laricieola has the ability to kill Japanese larch trees andthat it is more virulent to Japanese larch trees than O.pieeae. Redfern et al. (1987) first described C. laricieolaand demonstrated its pathogenicity to European larch.These authors did not, however, consider the ability ofthe fungus to kill trees. Maeto et al. (19911 describedthe association between O. pieeae and I. cembrae inJapan and demonstrated that the fungus is pathogenic toJapanese larch. However, the inoculated trees werekilled only when they were inoculated more densely withthe fungus IMaeto et aI., 1991; Yamaguchi et aI., 19911
Table 2. Leaf symptom development in inoculated Japanese larch trees.
Days after inoculation
a) -; no symptom, +; slight discoloration of leaves, +f-; browning of leaves and serious defoliation, 0;
death.
Table 3. Comparison of lesion length on the inner bark of trees inoculated withC. larfcfcola and O. pfceae.
Inoculum
Ceratocystis larfcfcolaYCC-285
Ophfostoma piceaeYCC.295
Control
Tree No.D\ Length of lesionl!\ (mm)
NO.1
No.2
No.3
No.4
NO.5
No.6
115.4:t40.6a
96.4z 27.4b
8.9:t 1.8c
11.4:t2.2c
4.3:t0.9c
3.6:t0.9c
a) Tree Nos. 4 and 6 were felled on 7 November 1995; all other trees were felled
on 11 September 1995.b) Values (mean:tSD, n=25) followed by the same letter are not significantly
different (P=O.05) by Duncan's new multiple range test.

376 Y. Yamaoka et al.

Ips cembrae-associated ophiostomatoid fungi
than the inoculum density used in the present study.Neither the isolates of O. piceae used by Maeto et al-119911 and Yamaguchi et ai. 119911 nor the ones isolatedin Hokkaido were used in the present study. Thus, therelative virulence of these isolates can not be clarified,since they were not inoculated under the same inocula-tion conditions at the same time. However, the sizes oflesions on inner bark resulting from the inoculation withO. piceae in the earlier studies were almost the same asthose observed in the present study and considerablysmaller than the lesions formed by C. laricicola. Maetoet ai. 119911 and Yamaguchi 119951 considered O. piceaeto be the causal agent of larch mortality after the massattack by I. cembrae. In contrast, we believe that C.laricicola is the more important fungal associate of I.cembrae and that it plays a significant role in tree death.
Acknowledgements-Wethank the Experimental Forests inYatsugatake, Agricultural and Forestry Research Center, Univer-sity of Tsukuba, for providing Japanese larch trees for inocula-tion. Part of this study was supported by a Grant-in-Aid for En-couragement of Young Scientists, The Ministry of Education,Science, and Culture, Japan to Y. Yamaoka. Part of this studywas presented at the 40th Annual Meeting of the MycologicalSociety of Japan, May 25-26, 1996, University of Tsukuba,Tsukuba, Ibaraki, Japan, and at the Asian International Mycolog-ical Congress '96 (AIMC'96), July 3-5, 1996, Chiba University,Chiba, Japan. Contribution No. 136, laboratories of PlantPathology and Mycology, Institute of Agriculture and Forestry,University of Tsukuba.
literature cited
Aoshima, K. 1965. Studies on wood-staining fungi of Japan.PhD thesis, University of Tokyo, Tokyo. (In Japanese.)
Aoshima, K. and Hayashi, Y. 1953. Blue stain of beech log(Fagus crenata) and its causal fungus. J. Jpn. For. Soc.35: 26B-269. ([n Japanese.)
Bakshi, S. K. 1950. Fungi associated with ambrosia beetles inGreat Britain. Trans. Sr. Mycol. Soc, 33: 111-120.
Brasier, C. M. and Kirk, S. A. 1993. Sibling species withinOphiostoma piceae. Mycol. Res. 97: 811-816.
Crooke, M, and Bevan, D, 1957. Note on the first British oc-currence of Ips cembrae Heer (CoL Scolytidae). Forestry
377
30: 21-28.Davidson, R. W. 1942. Some additional species of Cerato-
stomella in the United States. Mycologia 34: 650-662.Davidson, R. W., Francke-Grosmann, H, and Kaarik, A, 1967.
A restudy of Ceratocystis penicillata and report of twoAmerican species of this genus from Europe. Mycologia59: 928-932.
Furniss, M. M., Solheim, H, and Christiansen, E. 1990, Trans-mission of blue-stain fungi by Ips typographus (Coleoptera:Scolytidae) in Norway spruce. Ann. Entomol. Soc. Am.83: 712-716.
Georgevitch, P. 1927. Ceratostomella quercus n, sp. EinParasit der slawonischen Eichen. 8iologia Generalis 3:245-252.
Griffin, H, D. 1968. The genus Ceratocystis in Ontario, Can.J. 80t. 46: 689-718.
Harding,S, 1989. The influence of mutualistic blue stain fungion bark beetle population dynamics. PhD thesis, RoyalVeterinary and Agricultural University, Copenhagen.
Harrington, T. C., Steimel, J. P., Wingfield, M. J, and Kile, G, A.1996. Isozyme variation and species delimitation in theCeratocystis coeruleseens complex. Mycologia 88: 104-113.
Halmschlager, E., Messner, R., Kowalski, T, and Prillinger, H.1994. Differentiation of Ophiostoma pieeae and Ophio-stoma quercus by morphology and RAPD analysis. Sys-tem. Appl. MicrobioL 17: 554-562.
Horntvedt, Ro, Christiansen, E" Soheim, H. and Wang, S. 1983,Artificial inoculation with Ips typographus-associated blue-stain fungi can kill healthy Norway spruce trees, Medd.Nor, Inst. Skogforsk. 38: 1-20.
Hunt, J, 1956. Taxonomy of the genus Ceratoeystis, Uoydia19: 1-59.
Kaarik, A, 1975. Succession of microorganisms during wooddecay. In: Biological transformation of wood by microor~ganisms. Proceedings of the Sessions on Wood ProductsPathology at the 2nd Intern. Congr, Plant Pathology. Sept.10-12, 1973., (ed. by Liese, W.), pp, 39-51. Springer~Verlag, Berlin.
Koizumi, C. 1990. Ips cembrae. Ringyo-to-Yakuzai 111:1-10. (In Japanese,)
Koizum!, C, 1994. Larch Ips, Ips cembrae (Heer). In: ShinrinKonchu (Forest Insects), (ed. by Kobayashi, F. and Taketa-ni, A.J, pp. 183-184. Yokendo, Tokyo. (In Japanese,)
Kotynkova-Sychrova, E. 1966. Mykofl6ra chodeb kurovcu vCeskoslovensku. (The mycoflora of bark-beetle galleries inCzechoslovakia.) Ceska Myko1. 20: 45-53.
Fig. 26, Necrotic lesions formed around the inoculation points on the inner bark 55 d after inoculation with ophiostomatoid fungiassociated with Ips cembrae.a) Control, b) Ceratocystis larieicola YCC-285, c) Ophiostoma piceae YCC-295, d} O. brunneo-ciliatum YCC-287, e} O. larids YCC-289, f} Ophiostoma sp, YCC-300, g) Ceratocystiopsis minuta YCC-294, h} C. laricicola YCC-273, i} O. pieeae YCC-299.
Fig. 27, A dying larch tree (No.2) inoculated with C.laricicola YCC-285, 88 d after inoculation.Note that most of the leaves have fallen.
Figs. 28-30, Lesions formed around the inoculation points on the inner bark 104 d after inoculation.I, inoculation point; C, callus tissue (bars. 2 em). Fig. 28. Tree No.1 inoculated with C. laricicola. Note that large necroticlesions merged with each other and completely encircled trunks of the trees around the inoculation zone. Fig. 29. Tree No.3inoculated with O. piceae. Note that small necrotic lesions were formed around the inoculation zone, but they never merged.Callus tissues developed around some of the lesions, Fig. 30, Tree No, 5 inoculated with control inoculum, Note that the lesionsformed around the inoculation zone were very small. Some of the holes were plugged by callus tissue.
Figs. 31-33, Cross sections of the inoculated trees 2 em above the upper inoculation points, 104 d after inoculation.H, heartwood; S, sapwood; Arrowheads, border between heartwood and sapwood. Fig.31. Tree No, 1 inoculated with C. lariei-
cola. Note that the entire sapwood of the tree was blue-stained. Fig. 32, Tree No, 3 inoculated with O. piceae, Note that thesapwood was partiaJly stained a pale brown color. Fig, 33. Tree No.5 inoculated with control inoculum. Note the absence ofstained sapwood or dry zone of sapwood.

378 Y. Yamaoka et al.
Lieutier, F., Vart, A., Garcia, J., Ham, M. C.. Morele!, M. andLevieux, J. 1989. Champignons phytopathogenes as-Baciesa deux col!~opteres scolytidae du pin sylvestre (Pinussylvestris L), et etude preliminaire de leur aggressivite eo-vers !'hote. Ann. Sci. For. {Paris} 46: 201-216. (InFrench.)
Maeto, 1<', Ozaki, K., Hayashi, Y. and Koizum!, C. 1991. Wiltof Japanese larch inoculated with Ips cembrae-associatedblue-stain fungi CeratDcystis piceae. Trans. 39th Ann.Mtg. Hokkaido Branch Jpn. For. Soc. 39: 79-82. {InJapanese. }
Mathiesen, A. 1950. Uber elolge mit Borkenkafern assoziierteBlauepiJze in Schweden. Dikas 2: 275-308,
Mathiesen, A. 1951. Einige neue Ophiostoma-arten inSchweden. $v. Bot. Tidskr. 45: 203-232.
Mathiesen~IC3arik, A. 1953. Eine Obersicht uber die gew6hn-Hchsten mit Borkenkafern assoziierten Blauepilze inSchweden. Meddn. St. Skogsforsklnst. 43: 1-74.
Mathiesen-Kaarik, A. 1960. Studies on the ecology, taxonomyand physiology of Swedish insect-associated blue stainfungi, especially the genus Ceratocysas. Oikos 11: 1-25.
Molnar, A. C. 1965. Pathogenic fungi associated with a barkbeetle on alpine fir. Can. J. Bot. 43: 563-570.
Munch, E. 1907. Die Blaufaule des Nadelholzes. Naturw. Z.Land- u. Forstw. 5: 531-573.
Nisikado, Y. and Yamauti, K. 1935. Contributions to theknowledge of the sap stains of wood in Japan. III. Studieson Ceratostomella piceae Munch, the cause of a blue stainof pine trees. Ohara Inst. Landw. Forsch. Ber. 6: 539-560.
Nobuchi, A. 1974. Studies on Scolytidae XII. The bark beetlesof the tribe Ipini in Japan (Coleoptera). Bull. Gov. For. Exp.Sta. 266: 33-60.
Olchowecki. A. and Reid, J. 1974. Taxonomy of the genusCeratocystis in Manitoba. Can. J. Bot. 52: 1675-1711.
Otani, Y. 1988. Ophiostomatales Benny et Kimbrough. In:Seiya Ito's Mycological flora of Japan, led. by Otani, Y.j,pp.134-147. Yokendo, Tokyo. (In Japanese.)
Paine, T. D. 1984. Seasonal response of ponderosa pine to in~oculation of the mycangial fungi from the western pine bee-tle. Can. J. Bot. 62: 551-555.
Redfern, D. B. 1989. The roles of the bark beetle Ips cembrae,the wood wasp Urocerus gigas and associated fungi in die-back and death of larches. In: Insect~fungus interactions,(ed. by Wilding, N., Collins, N. M., Hammond, P. M. andWebber, J. F.Lpp. 195-204. Academic Press, London.
Redfern, D. B., Stoakley, J. T., Steele, H. and Minter, D. W.1987. Dieback and death of larch caused by Ceratocystislaricicola sp. novo following attack by Ips cembrae. PlantPathol. 36: 467-480.
Robinson, R. C. 1962. Blue stain fungi in lodgepole pine (Pinuscontorta Dougl. var./atifo/ia Engelm.) infested by the moun-tain pine beetle (Dendroctonus monticolae Hopk.). Can. J.80t. 40: 609-614.
Siemaszko, W. 1939. Zespoly grzybow fowarzyszacych kor-nikom polskim. (Fungi associated with bark~beetles inPoland.) Planta Pol. 7: 1-54.
Solheim, H. 1986. Species of Ophiostomataceae isolated fromPicea abies infested by the bark beetle Ips typographus.Nord. J. Bot. 6: 199-207.
Solheim, H. 1988. Pathogenicity of some Ips typographus-
associated blue-stain fungi to Norway spruce. Medd. Nor.Inst. Skogforsk. 40: 1-11.
Solheim, H. 1992a. The early stages of fungal invasion in Nor~way spruce infested by the bark beetle Ips typographus.Can. J. Bot. 70: 1-5.
Solheim, H. 1992b. Fungal succession in sapwood of Norwayspruce infested by the bark beetle Ips typographus. Eur. J.For. Pathol. 22: 136-148
Solheim, H. 1993. Fungi associated with the spruce bark bee~tie Ips typographus in an endemic area in Norway. Scand.J. For. Res. 8: 118-122.
Sydow, H. and Sydow, P. 1919. Mykologische Mitteilungen.Ann. Mycol. 17: 33-47.
Tochinai, Y. and Sakamoto, M. 1934. Studieson blue-stain ofwood of Picea jezoensis. Hokkaido Ringyo Kaiho 32:334-342. (In Japanese.)
Upadhyay, H. P. 1981. A Monograph of Ceratocystis andCeratocystiopsis. Univ. Georgia Press, Athens, Georgia.
Upadhyay, H. P. and Kendrick, W. B. 1975. Prodromus for arevision of Ceratocystis (Microascales, Ascomycetes) andits conidial states. Mycologia 67: 798-805.
Viiri, H. and Weissenberg, 1<.v. 1995, Ophiostoma blue-stain-ing fungi associated with Ips typographus in Finland. In:Forest pathology research in the Nordic countries 1994,Proceedings from the SNS-meeting in Forest Pathology atSkogbrukets Kurssenter, Biri, Norway, 9-12 August 1994.Aktuelt fra Skogforsk 4/95, (ed. by Aamlid, D.), pp. 58-60.Nor.lnst. Skogforsk., Norway.
Visser, C., Wingfield, M. J., Wingfield, B. D. and Yamaoka, Y.1995. Ophiostoma polonicum is a species of Ceratocystissensu stricto. System. Appl. Microbiol. 18: 403-409.
Westhuizen, 1<.van der, Wingfield, M. J., Yamaoka, Y., Kemp,G. H. J. and Crous, P. W. 1995. A new species ofOphiostoma with a Leptographium anamorph from larch inJapan. MycoL Res. 99: 1334-1338.
Witthuhn, R. C., Wingfield, B. D., Wingfield, M. J., Wolfaardt,M, and Harrington,T. C. 1998. Monophylyof the coniferspecies in the Ceratocystis coerulescens complex based onDNA sequence data. Mycologia 90: 96-101.
Wright, E, F. and Cain, R. F. 1961. New species of the genusCeratocystis. Can. J. Bot. 39: 1215-1230.
Yamaguchi, T. 1993. Inoculation tests with Ceratocystispiceae to Japanese larch seedlings (1) Water potentialchanges associated with girdling and inoculation. Trans.104th Ann. Mtg. Jpn. For. Soc. 104: 615-616. OnJapanese.)
Yamaguchi, T, 1995. Anatomical and water potential changesin Japanese larch inoculated with Ophiostoma piceae. In:Bark beetles, blue-stain fungi, and conifer defence systems,(ed. by Christiasen, E.!, pp. 24-25. NISK, Norw. For. Res.Inst. (NISK) in As, Norway.
Yamaguchi, T., Sasaki, 1<'and Matsuzaki. S. 1991. Inoculationtests of Ceratocystis piceae to Japanese larch !!II) Wilt ofinoculated trees. Trans. 39th Ann. Mtg. Hokkaido BranchJpn. For. Soc. 39: 76-78. (InJapanese.)
Yamaoka, Y., Wingfield, M. J., Takahashi, L and Solheim, H.1997. Ophiostomatoid fungi associated with the sprucebark beetle Ips typographus f. japonicus in Japan. Mycol.Res. 101: 1215-1227.