opean rev iewfor med icaland pharmacol sci 2016;20:3327 ... · 2d ep artm n ofb ich s ydm lu g, lp...
TRANSCRIPT

Abstract. – OBJECTIVE: To investigate thecorrelation between LeY glycan expression andembryo implantation.
MATERIALS AND METHODS: Uterine epithe-lial cells before implantation were transfectedwith FUT1siRNA to inhibit FUT1 (the gene encod-ing the key enzyme of LeY synthesis) expressionand treated with 10 ng/ml leukemia inhibitoryfactor (LIF). Murine embryo implantation modelin vitro was prepared by late blastocysts withidentical morphology and treated uterine epithe-lial cells co-culture. Using RT-PCR, dot blot andobservation of embryo attachment to analyzeFUT1 gene expression and LeY synthesis ofuterine epithelial cells and studied further thecorrelation of LeY expression level and embryoimplantation.
RESULTS: FUT1 gene expression and LeYsynthesis declined after cells were transfectedwith FUT1siRNA, and LIF promoted FUT1 expres-sion and LeY synthesis. After expression ofFUT1 gene was inhibited, attachment rate ofembryos lowered, but LIF up-regulated FUT1 ex-pression and increased the attachment rate ofembryos.
CONCLUSIONS: These results indicated reg-ulating FUT1 expression affected LeY synthe-sis, and then LeY regulated the recognition andattachment of uterus-embryo and participatesin embryo implantation further.
Key Words:LeY glycan, LIF, Uterine epithelial cell, Embryo im-
plantation, RNA interference.
Introduction
Ovum would gradually develop into “invasiveblastocyst” in the process of migrating into theuterus through oviduct after fertilization andwould interact with the endometria that has al-
European Review for Medical and Pharmacological Sciences
Effects of LeY glycan expression on embryoimplantation
J. GU1, L.-L. SUI2, D. CUI2, Y.-N. MA2, C.-Y. ZHU2, Y. KONG2
1Xuzhou Central Hospital, The Affiliated XuZhou Hospital of Medical College of Southeast University,Xuzhou Clinical School of Xuzhou Medical College, Xuzhou Clinical Medical College of NanjingUniversity of Chinese Medicine, Southeast University Reproductive Medicine Institute, Xuzhou,Jiangsu, China2Department of Biochemistry and Molecular Biology, Liaoning Provincial Key Lab of Glycobiologyand Glycoengineering, Dalian Medical University, Dalian, Liaoning, China
Corresponding Author: Ying Kong, MD; e-mail: [email protected] 3327
ready developed into “receptive” state to start upembryo implantation. Embryo implantation is di-vided into three processes: location, adhesionand invasion. The specific expression of cytokine(CK), matrix metalloproteinases (MMPs) and itsinhibition factor (TIMPs), cell adhesion mole-cules (CAMS), extracellular matrix (ECM) andother implantation related factors in peri-implan-tation would interact with each other under theregulation of hormone and form a “regulationnetwork” to regulate the microenvironment of theembryo and the uterus, and create conditions forsuccessful implantation1.
Blastocyst would firstly contact mutually withthe glycan chain on the surface of cells while ap-proaching endometrium. The LeY glycan on thecell surface is involved in the recognition and ad-hesion between the embryo and the uterine ep-ithelium. To obstruct the expression of LeY gly-can before implantation by LeY monoclonal anti-body could significantly inhibit embryo implanta-tion2,3, and also inhibit the secretion and gene ex-pression of leukemia inhibitory factor (LIF),MMPs, epidermal growth factor (EGF), trans-forming growth factor (TGF), and vascular en-dothelial growth factor (VEGF) in embryo andendometrial cells before implantation, and mean-while promote the secretion of fibronectin (FN)and laminin (LN). The FN and LN increase as theexpression of proteolytic enzymes decreased4-6 in-dicating that LeY glycan was involved in the im-plantation regulation as an information molecule.The expression of fucosyltransferase (FUTI) thatcatalyzed the synthesis of LeY glycan in embryoand endometrium during peri-implantation wasconsistent with LeY glycan7, which indicated thatthe expression level of glycan was regulated by itssynthesis key enzyme gene.
2016; 20: 3327-3335

J. Gu, L.-L. Sui, D. Cui, Y.-N. Ma, C.-Y. Zhu, Y. Kong
MaterialsFUT1 interference reagent kit was from Qia-
gen, Hilden, Germany. LIF was introduced fromAmerica Chemical Company, AH6 monoclonalantibody (anti-LeY) was given away by Profes-sor Hakomori from University of Washington,USA, RT-PCR kit was from Dalian TakaraCompany, β-actin antibody from Wuhan BosterCompany.
Preparation and Treatment ofMonolayered Endometrial Epithelial Cells
The uterus of mice was dissected in the thirdday of pregnancy (D3). The uterus tissue was di-gested with trypsin solution (Sigma-Aldrich, St.Louis, MO, USA), followed by centrifugation atlow speed (500 rpm). The endometrial epithelialcells were collected, and allowed to grow in 24-well culture plate (Sigma-Aldrich, St. Louis,MO, USA). Subsequently, 0.5 ml Ham’s F-10(Gibco, Rockville, MD, USA)+10% FBS (fetalbovine serum, Institute of Hematology, ChineseAcademy of Medical Sciences) was added to theeach well. The culture medium was composed of1×105 cells, and cultured in incubator of 37°Cand 5% CO2
13. The well on culture plate were di-
vided into control and treatment groups, andwere transfected with HiPerFect transfectionreagent (Qiagen, Hilden, Germany). The cells ininterference group were transfected with 20µmol/L FUT1 siRNA 0.15 µl, cells in interferencecontrol group were transfected with 20 µmol/Lcontrol siRNA 0.15 µl, cells in LIF group weretreated with 10 ng/ml LIF, and cells cultured inpure culture medium were grouped into a blankgroup. Subsequent experiments were conducted24 h after transfection and 12 h after LIF.
Detection of FUT1 Gene Expressionin Endometrial Cells
The cells from all groups were collected andwashed with 75% ethanol after Trizol split (Gib-co, Rockville, MD, USA), chloroform extractionand isopropanol precipitation. Finally, the precip-itants were completely dissolved in 10 µl RNaseFree ddH2O. The absorbance was measured atOD260, OD280 (UV754) (TianMei BiotechnologyCompany, Beijing, China) and RNA purity andquality was analyzed after 1% formaldehyde de-naturing gel electrophoresis, estimation of RNAcontent. Reverse transcription was carried out in20 µl reaction system composed of 2 µg RNA, 4µl Mg2+ (25 mmol/L), 2 µl 10× reverse transcrip-tion buffer, 2 µl dNTP (10 mmol/L), 0.5 µl
LIF is a kind of secreted glycoprotein withextensive biological function with a regular ex-pression in the embryo and endometrium of hu-man, mouse, and other mammal and high ex-pression in “implantation window phase”,which could affect the blastocyst developmentin morula and blastula stage, and start up blasto-cyst implantation8-10. LIF gene knockout micecould fertilize normally and produce blasto-cysts, but the blastocysts could not support im-plantation11. Therefore, the present view holdsthat LIF is the most important cytokine thatwould mediate blastocyst implantation12 andthat the uterus lacking of LIF is not responsiveto the implantation stimuli.
To investigate the correlation between LeYglycan expression and embryo implantation,we have prepared endometrial epithelial cellson the 3rd day of pregnancy (D3) and groupedthem, meanwhile transfected the uterine ep-i thelial cel ls before implantat ion withFUT1siRNA to inhibit FUT1 expression. Then,we treated them with 10 ng/ml of LIF to up-regulate the FUT1 expression. Murine embryoimplantation model in vitro was prepared bylate blastocysts with identical morphology andtreated uterine epithelial cells co-culture. Weused RT-PCR, Dot-blot and observation of em-bryo attachment to analyze FUT1 gene expres-sion and LeY synthesis of uterine epithelialcells, to study the correlation of LeY expres-sion level and embryo implantation, and to un-derstand further the effect of LeY in embryoimplantation.
Materials and Methods
Experimental AnimalsAdult female Kunming mice of 6-8 weeks age
(22-24 g) were provided by the experimental ani-mal center of Dalian Medical University andwere fed for 12 h in the light cycle and 12 h indark cycle. 5-10 IU pregnant mare serum go-nadotropin (PMSG, Tianjin animal experimentcenter) were injected into the mice for superovu-lation. 48 h later, we injected 5-10 IU of humanchorionic gonadotropin (hCG, Shandong PenglaiHuatai Pharmaceutical co. Ltd), after which wemated them with adult male mice in the ratio of1:1. Examined their vagina the next day. Theones with vaginal plug were confirmed as thefirst day of pregnancy (D1).
3328

Effects of LeY glycan expression on embryo implantation
3329
min. The image was developed in a dark roomafter exposing membrane to NBT/BCIP (Gibco,Rockville, MD, USA). The image was processedby Lab works 4.60 software and the relative in-tensity of each sample was calculated on the ba-sis of β-actin, then protein content of LeY werecompared and analyzed.
Blastocyst Collection and Co-Culturewith Endometrial Epithelial Cells
Female mice were decapitated in the fourthday of pregnancy (D4) to rapidly collect theiruterus to remove blastocysts from the uterusbefore implantation. The well-developed anduniform Level A advanced blastula was select-ed14 and allowed to migrate into 35 mm culturedish. Afterwards, it was placed into 50 µl ofpre-warmed Ham’s F-10 culture mediumdroplet at 37°C in 5% CO2 incubator for cul-ture. After 2 h, blastula was transferred into 24-well culture plate and co-cultured with en-dometrial epithelial cells from all groups. Eachwell with 10 blastocysts was cultured for 12 h,24 h, and 36 h individually in 5% CO2 incuba-tor at 37°C and then the attachment of embryowas observed and the ratio was calculated. Theattachment rate of each group was presented bypercentage between the number of attachmentembryos and that of total embryos. The identi-fication of attachment of blastocyst on the sur-face of uterine epithelial cell monolayer wassubject to the methods of Masahide et al15.
Statistical AnalysisSPSS 11.5 software (SPSS Inc., Chicago, IL,
USA) was used for data analysis. One-WayANOVA was applied to make comparisonsamong the three groups and independent-sam-ple t-test was applied to make a comparison be-tween the groups. Repeated Measure ANOVAwas applied to evaluate the outcomes of co-cul-ture and p<0.05 was considered to be statisti-cally significant.
RNase inhibitor (40 u/µl), 1 µl AMV reversetranscriptase (5 ug/µl), 1 µl random primer (Oli-go dT), and complemented the system by DEPCwater. The PCR conditions were annealing at50°C for 50 min, extension 99°C for 5 min, final-ly at 4°C for 5 min. 2 µl of cDNA was taken forFUT1 and β-actin gene amplification, 2 µl Mg2+
(25 mmol/L), 2 µl dNTP (2.5 mmol/L), 2.5 µl10×Ex Taq Buffer, 0.5 µl Taq DNA polymerase,0.5 µl (30 pmol/L) forward primer and 0.5 µl (30pmol/L) reverse primer, complemented withddH2O till the volume of 25 µl was used. PCR re-action condition was pre-denaturation at 94°C for5 min, denaturation at 94°C for 50 s, annealing at56°C for 50 s, extension at 72°C for 1 min, then,a final extension at 72°C for 5 min after 35 cy-cles. The primers sequence is shown in Table I.
The PCR products were loaded on to 1%agarose gel for electrophoresis and were subse-quently processed by Lab works 4.60 software.The relative intensity of each band was normal-ized with β-actin.
Protein Extraction and Dot-blot Analysisof Endometrial Cells
100 µl protein extract (1000 µl PBS contained10 µl NP-40 and 10 µl PMSF) was added to eachwell and was transferred into Eppendorf tube af-ter blowing, beating and binding at 4°C for 2 h.Subsequently, the tube contents were centrifugedat 4°C for 15 min at 10,000 rpm and the super-natant was transferred into a new tube and ex-tracted proteins were quantified by Kaunas Brad-ford method. 20 µg of the extracted proteins ineach group was attached to NC membrane byDot blot instrument (MBI Company), incubatedwith 5% BSA at 37oC for 2 h. The membranewas washed with TBS and allowed to incubatewith LeY primary antibody at 37°C for 2 h. Themembrane was washed again with TBS and thenplaced into alkaline phosphatase labeled anti-IgM second antibody (Santa Cruz Biotechnology,Santa Cruz, CA, USA) to incubate at 37°C for 40
Genes Sequences (5’to3’) Size (bp)
FUT1 (F)5’-CCTGGCATTTCTGTTGGTCTGT-3’ 559 bp(R)5’-ACCTTCCCGTAGATGGTTATGC-3’
β-actin (F)5’-GTGGGCCGCCCTAGGCACCAA-3’ 540 bp(R)5’-CTCTTTGATGTCACGCACGATTTC-3’
Table I. Primers used for this study.
(F) Stands for forward primer, (R) reverse primer, primers designed by primer 5.0.
J. Gu, L.-L. Sui, D. Cui, Y.-N. Ma, C.-Y. Zhu, Y. Kong
Results
RT-PCR detection on Gene Expression ofEndometrial cell FUT1 After Transfection
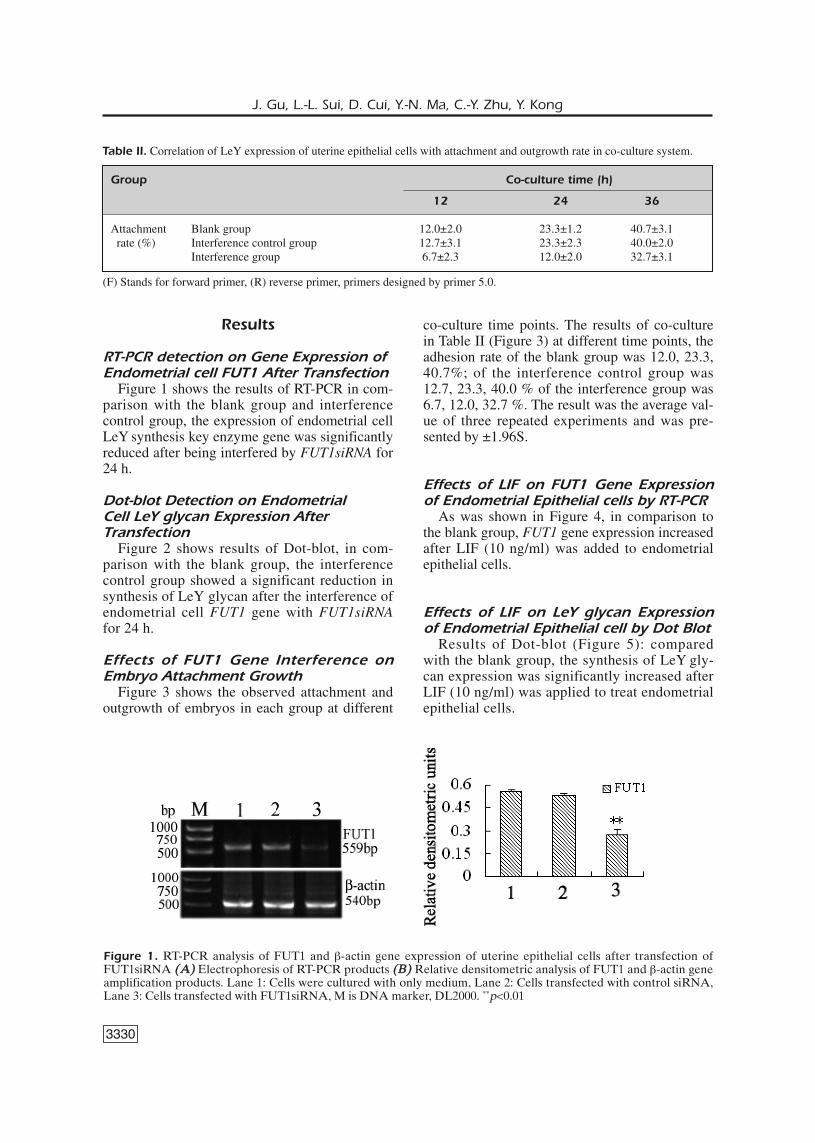
Figure 1 shows the results of RT-PCR in com-parison with the blank group and interferencecontrol group, the expression of endometrial cellLeY synthesis key enzyme gene was significantlyreduced after being interfered by FUT1siRNA for24 h.
Dot-blot Detection on EndometrialCell LeY glycan Expression AfterTransfection
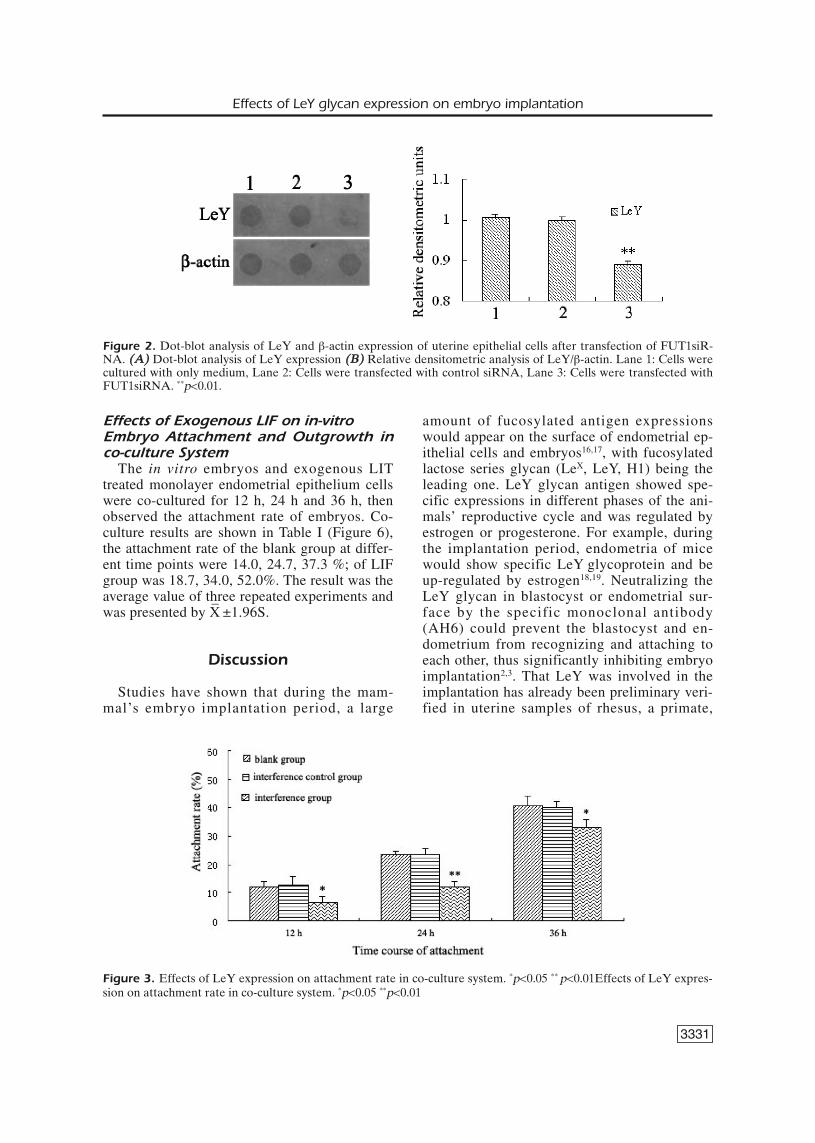
Figure 2 shows results of Dot-blot, in com-parison with the blank group, the interferencecontrol group showed a significant reduction insynthesis of LeY glycan after the interference ofendometrial cell FUT1 gene with FUT1siRNAfor 24 h.
Effects of FUT1 Gene Interference onEmbryo Attachment Growth
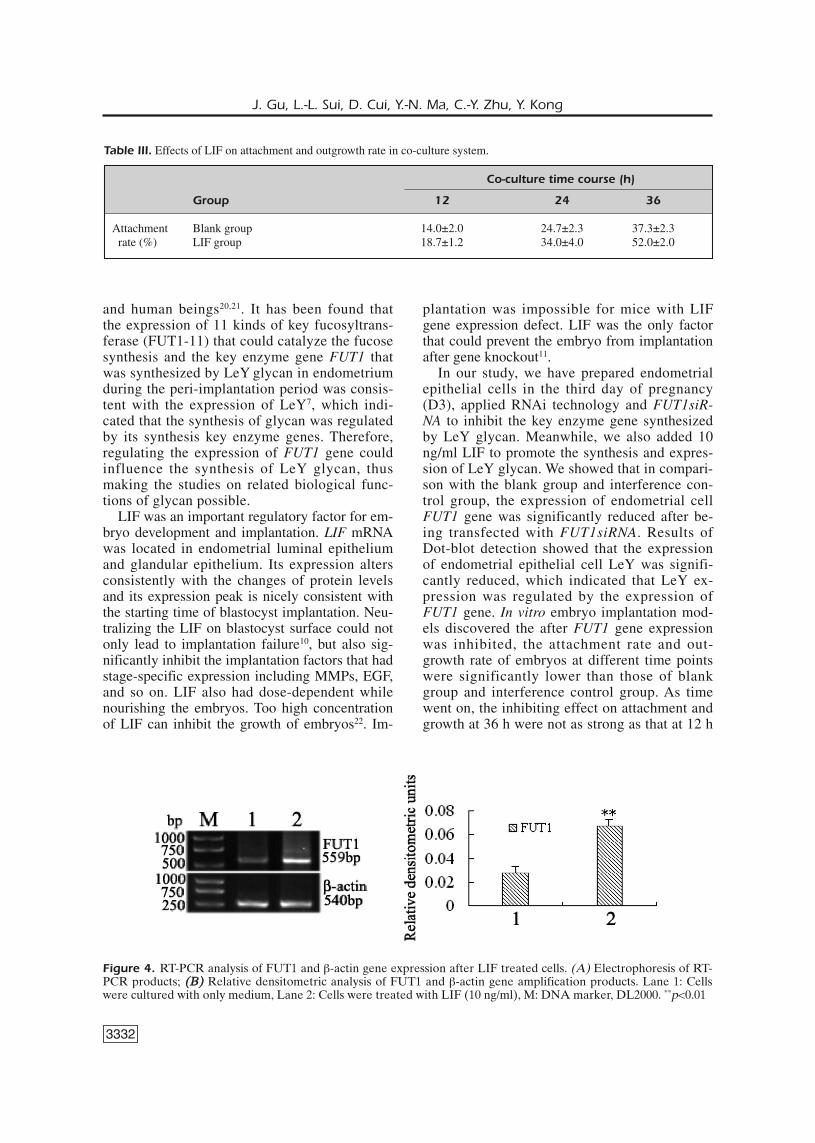
Figure 3 shows the observed attachment andoutgrowth of embryos in each group at different
3330
co-culture time points. The results of co-culturein Table II (Figure 3) at different time points, theadhesion rate of the blank group was 12.0, 23.3,40.7%; of the interference control group was12.7, 23.3, 40.0 % of the interference group was6.7, 12.0, 32.7 %. The result was the average val-ue of three repeated experiments and was pre-sented by ±1.96S.
Effects of LIF on FUT1 Gene Expressionof Endometrial Epithelial cells by RT-PCR
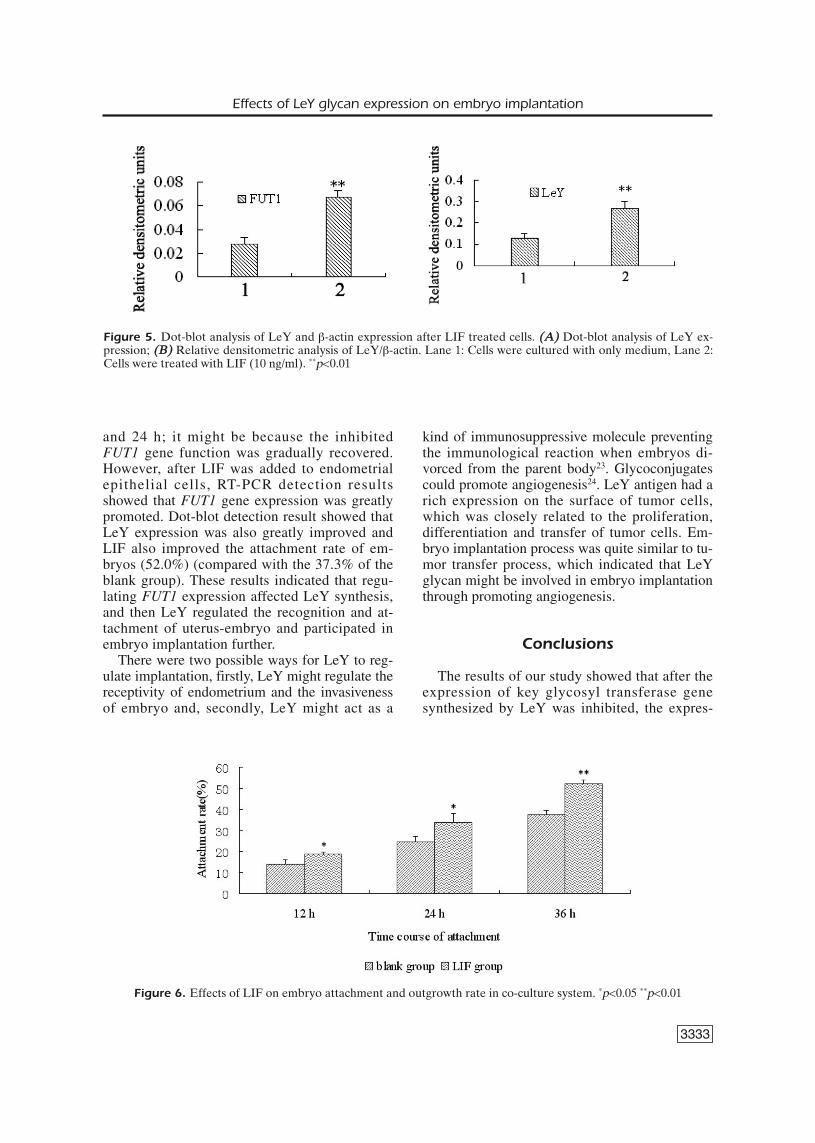
As was shown in Figure 4, in comparison tothe blank group, FUT1 gene expression increasedafter LIF (10 ng/ml) was added to endometrialepithelial cells.
Effects of LIF on LeY glycan Expressionof Endometrial Epithelial cell by Dot Blot
Results of Dot-blot (Figure 5): comparedwith the blank group, the synthesis of LeY gly-can expression was significantly increased afterLIF (10 ng/ml) was applied to treat endometrialepithelial cells.
Group Co-culture time (h)
12 24 36
Attachment Blank group 12.0±2.0 23.3±1.2 40.7±3.1rate (%) Interference control group 12.7±3.1 23.3±2.3 40.0±2.0
Interference group 6.7±2.3 12.0±2.0 32.7±3.1
Table II. Correlation of LeY expression of uterine epithelial cells with attachment and outgrowth rate in co-culture system.
(F) Stands for forward primer, (R) reverse primer, primers designed by primer 5.0.
Figure 1. RT-PCR analysis of FUT1 and β-actin gene expression of uterine epithelial cells after transfection ofFUT1siRNA ((AA)) Electrophoresis of RT-PCR products ((BB)) Relative densitometric analysis of FUT1 and β-actin geneamplification products. Lane 1: Cells were cultured with only medium, Lane 2: Cells transfected with control siRNA,Lane 3: Cells transfected with FUT1siRNA, M is DNA marker, DL2000. **p<0.01
Effects of LeY glycan expression on embryo implantation
3331
amount of fucosylated antigen expressionswould appear on the surface of endometrial ep-ithelial cells and embryos16,17, with fucosylatedlactose series glycan (LeX, LeY, H1) being theleading one. LeY glycan antigen showed spe-cific expressions in different phases of the ani-mals’ reproductive cycle and was regulated byestrogen or progesterone. For example, duringthe implantation period, endometria of micewould show specific LeY glycoprotein and beup-regulated by estrogen18,19. Neutralizing theLeY glycan in blastocyst or endometrial sur-face by the specific monoclonal antibody(AH6) could prevent the blastocyst and en-dometrium from recognizing and attaching toeach other, thus significantly inhibiting embryoimplantation2,3. That LeY was involved in theimplantation has already been preliminary veri-fied in uterine samples of rhesus, a primate,
Effects of Exogenous LIF on in-vitro Embryo Attachment and Outgrowth inco-culture System
The in vitro embryos and exogenous LITtreated monolayer endometrial epithelium cellswere co-cultured for 12 h, 24 h and 36 h, thenobserved the attachment rate of embryos. Co-culture results are shown in Table I (Figure 6),the attachment rate of the blank group at differ-ent time points were 14.0, 24.7, 37.3 %; of LIFgroup was 18.7, 34.0, 52.0%. The result was theaverage value of three repeated experiments andwas presented by X
–±1.96S.
Discussion
Studies have shown that during the mam-mal’s embryo implantation period, a large
Figure 2. Dot-blot analysis of LeY and β-actin expression of uterine epithelial cells after transfection of FUT1siR-NA. ((AA)) Dot-blot analysis of LeY expression ((BB)) Relative densitometric analysis of LeY/β-actin. Lane 1: Cells werecultured with only medium, Lane 2: Cells were transfected with control siRNA, Lane 3: Cells were transfected withFUT1siRNA. **p<0.01.
Figure 3. Effects of LeY expression on attachment rate in co-culture system. *p<0.05 ** p<0.01Effects of LeY expres-sion on attachment rate in co-culture system. *p<0.05 **p<0.01
3332
and human beings20,21. It has been found thatthe expression of 11 kinds of key fucosyltrans-ferase (FUT1-11) that could catalyze the fucosesynthesis and the key enzyme gene FUT1 thatwas synthesized by LeY glycan in endometriumduring the peri-implantation period was consis-tent with the expression of LeY7, which indi-cated that the synthesis of glycan was regulatedby its synthesis key enzyme genes. Therefore,regulating the expression of FUT1 gene couldinfluence the synthesis of LeY glycan, thusmaking the studies on related biological func-tions of glycan possible.
LIF was an important regulatory factor for em-bryo development and implantation. LIF mRNAwas located in endometrial luminal epitheliumand glandular epithelium. Its expression altersconsistently with the changes of protein levelsand its expression peak is nicely consistent withthe starting time of blastocyst implantation. Neu-tralizing the LIF on blastocyst surface could notonly lead to implantation failure10, but also sig-nificantly inhibit the implantation factors that hadstage-specific expression including MMPs, EGF,and so on. LIF also had dose-dependent whilenourishing the embryos. Too high concentrationof LIF can inhibit the growth of embryos22. Im-
plantation was impossible for mice with LIFgene expression defect. LIF was the only factorthat could prevent the embryo from implantationafter gene knockout11.
In our study, we have prepared endometrialepithelial cells in the third day of pregnancy(D3), applied RNAi technology and FUT1siR-NA to inhibit the key enzyme gene synthesizedby LeY glycan. Meanwhile, we also added 10ng/ml LIF to promote the synthesis and expres-sion of LeY glycan. We showed that in compari-son with the blank group and interference con-trol group, the expression of endometrial cellFUT1 gene was significantly reduced after be-ing transfected with FUT1siRNA. Results ofDot-blot detection showed that the expressionof endometrial epithelial cell LeY was signifi-cantly reduced, which indicated that LeY ex-pression was regulated by the expression ofFUT1 gene. In vitro embryo implantation mod-els discovered the after FUT1 gene expressionwas inhibited, the attachment rate and out-growth rate of embryos at different time pointswere significantly lower than those of blankgroup and interference control group. As timewent on, the inhibiting effect on attachment andgrowth at 36 h were not as strong as that at 12 h
J. Gu, L.-L. Sui, D. Cui, Y.-N. Ma, C.-Y. Zhu, Y. Kong
Co-culture time course (h)
Group 12 24 36
Attachment Blank group 14.0±2.0 24.7±2.3 37.3±2.3rate (%) LIF group 18.7±1.2 34.0±4.0 52.0±2.0
Table III. Effects of LIF on attachment and outgrowth rate in co-culture system.
Figure 4. RT-PCR analysis of FUT1 and β-actin gene expression after LIF treated cells. (A) Electrophoresis of RT-PCR products; ((BB)) Relative densitometric analysis of FUT1 and β-actin gene amplification products. Lane 1: Cellswere cultured with only medium, Lane 2: Cells were treated with LIF (10 ng/ml), M: DNA marker, DL2000. **p<0.01
and 24 h; it might be because the inhibitedFUT1 gene function was gradually recovered.However, after LIF was added to endometrialepithelial cells, RT-PCR detection resultsshowed that FUT1 gene expression was greatlypromoted. Dot-blot detection result showed thatLeY expression was also greatly improved andLIF also improved the attachment rate of em-bryos (52.0%) (compared with the 37.3% of theblank group). These results indicated that regu-lating FUT1 expression affected LeY synthesis,and then LeY regulated the recognition and at-tachment of uterus-embryo and participated inembryo implantation further.
There were two possible ways for LeY to reg-ulate implantation, firstly, LeY might regulate thereceptivity of endometrium and the invasivenessof embryo and, secondly, LeY might act as a
kind of immunosuppressive molecule preventingthe immunological reaction when embryos di-vorced from the parent body23. Glycoconjugatescould promote angiogenesis24. LeY antigen had arich expression on the surface of tumor cells,which was closely related to the proliferation,differentiation and transfer of tumor cells. Em-bryo implantation process was quite similar to tu-mor transfer process, which indicated that LeYglycan might be involved in embryo implantationthrough promoting angiogenesis.
Conclusions
The results of our study showed that after theexpression of key glycosyl transferase genesynthesized by LeY was inhibited, the expres-
3333
Effects of LeY glycan expression on embryo implantation
Figure 5. Dot-blot analysis of LeY and β-actin expression after LIF treated cells. ((AA)) Dot-blot analysis of LeY ex-pression; ((BB)) Relative densitometric analysis of LeY/β-actin. Lane 1: Cells were cultured with only medium, Lane 2:Cells were treated with LIF (10 ng/ml). **p<0.01
Figure 6. Effects of LIF on embryo attachment and outgrowth rate in co-culture system. *p<0.05 **p<0.01

3334
sion of LeY was lowered, and the attachmentrate as well as the outgrowth rate of embryos,were also lowered. The acknowledged implan-tation essential factor, LIF played quite an im-portant role in promoting the LeY expressionon the surface of the endometrium during peri-implantation period while facilitating embryodevelopment and implantation process. Al-though some studies have reported that em-bryos, after gene knockout, could still achieveimplantation25. It might be because the com-plexity and precision of blastocyst implanta-tion required multiple influencing factors tocoordinate expression and co-regulate toachieve implantation. After an in-depth studyon the LeY glycan on the surface of endome-trial cells, we have come to that, to develop abetter understanding of the intrauterine envi-ronment during implantation period and themolecular markers of endometrium in differentstates are quite significant in reproductivemedicine, including the etiologic study andtreatment of infertility, and the improvement ofpregnancy rate of assisted reproductive tech-nique. Also, it also provides a new clue andmethod for clinical contraception.
–––––––––––––––––-––––Conflict of InterestThe Authors declare that they have no conflict of interests.
References
1) SHARKEY A. Cytokines and implantation. Rev Re-prod 1998; 3: 52-61.
2) ZHU ZM, KOJIMA N, STROUD MR, HAKOMORI S, FEND-ERSON BA. Monoclonal antibody directed to Le(y)oligosaccharide inhibits implantation in themouse. Biol Reprod 1995; 52: 903-912.
3) WANG XQ, ZHU ZM, FENDERSON BA, ZENG GQ, CAOYJ, JIANG GT. Effects of monoclonal antibody di-rected to Le(y) on implantation in the mouse. MolHum Reprod 1998; 4: 295-305.
4) GE CH, KONG Y, WANG H, ZHU ZM. Effect of Le(y)oligosaccharide on the expression and secretionof MMPs mice blastocyst. J Biochem Biophys2002; 34: 45-49.
5) KONG Y, GE CH, LI H. Effect of Le(y) oligosaccha-ride on surface of blastocyst on the expressionand secretion of blastocyst epidermal growth fac-tor and its receptor. J Biochem Biophys 2002; 34:116-120.
6) HU Y, KONG Y, FENG B. Effects of monoclonal anti-body blocked Le(y) oligosaccharides on extracel-
lular matrix of blastocyst and matrix metallopro-teinases. Reproduction and Contraception 2004;24: 204-209.
7) LIU N, JIN C, ZHU ZM, ZHANG J, TAO H, GE C, YANGS, ZHANG S. Stage-specific expression of α1,2-fu-cosyltransferase and α1,3-fucosyltransferase (FT)during mouse embryogenesis. Eur J Biochem1999; 265: 258-263.
8) BHATT H, BRUNET L J, STEWARD C L. Uterine expres-sion of luekaemia inhibitory factor coincides withonset of blastocyst implantation. Proc Natl AcadSci USA 1991; 88: 11408-11412.
9) MEZHEVIKINA LM, KAPRALOVA IV, FESENKO EE. Stimu-lating effect of recombinant cytokine LIF on themouse blastocysts during implantation. BiomedKhim 2006; 52: 620-626.
10) SENGUPTA J, LALITKUMAR PG, NAJWA AR, GHOSH D.Monoclonal anti-leukemia inhibitory factor an-tibody inhibits blastocyst implantation in therhesus monkey. Contraception. 2006; 74: 419-425.
11) STEWARD CL, KASPAR P, BRUNET LJ, BHATT H, GADI I,KÖNTGEN F, ABBONDANZO SJ. Blastocyst implanta-tion depends on maternal expression ofleukeamia inhibitory factor. Nature 1992; 359: 76-79.
12) SENTURK LM, ARICI A. Leukemia inhibitory factor inhuman reproduction. Am J Reprod Immunol 1998;39: 144-151.
13) ZENG G, CAO Y. Study on co-culture of embryo anduterine epithelial cell monolayer. Current Zoology1996; 42: 212-217.
14) STEER CV, MILLS CL, TAN SL. The cumulative embryoscore: a predictive embryo scoring technique toselect the optimal number of embryos to transferin an in-vitro fertilization and embryo transfer pro-gramme. Hum Reprod 1992; 7: 117-119.
15) MASAHIDE S, YOICHI N, TAKAHIDE M. Embryo-depen-dent induction of uterine receptivity assessed byan in vitro model of implantation in mice. Bio Re-prod 1993; 49: 794-801.
16) FENDERSON BA. Saccharides involved in implanta-tion. Trends Glycosci Glycotechnol 1993; 5: 271-285.
17) ZHU ZM, CHENG LM, HAKOMORI S. Glycosphingolipids of rabbit endometrium and their changesduring pregnancy. J Reprod Fertil 1992; 95: 813-823.
18) ILLINGWORTH IM, KIMBER SJ. Demonstration of oe-strogenic control of H-type-1 carboh- ydrate anti-gen in the murine endometrial epithelium by useof ICI 182, 780. J Reprod Fertil 1999; 117: 89-95.
19) WANG H, GE CH, KONG Y, XIN Y, ZHU ZM. Regula-tion of ovarian hormone on the expression of Leyoligosaccharide in peri-implantation period. JBiochem Biophys 2001; 33: 542-546.
20) GHOSH D, LIU N, ZHU ZM, SENGUPTA J. Immunolo-calization of LeY oligosaccharide in endometrium
J. Gu, L.-L. Sui, D. Cui, Y.-N. Ma, C.-Y. Zhu, Y. Kong

during menstrual cycle and effect of early lutealphase mifepristone administration on its expres-sion in implantation stage endometrium of therhesus monkey. Hum Reprod 1998; 13: 1374-1379.
21) ZHANG X, CHEN J, ZHU Z. Expression of LeYoligose in endometrium of different states. Chi-na Journal of Modern Medicine, 2001; 1:36-37.
22) MITCHELL MH, SWANSON RJ, OEHNINGER S. In vivo ef-fect of leukemia inhibitory factor (LIF) and an an-tiLIF polyclonal antibody on murine embryo andfetal development following exposure at the timeof transcervical blastocyst transfer. Biol Reprod2002; 67: 460-464.
23) ZHU ZM, WANG XQ. Role for cell surface oligosac-charide in cell-cell recognition during implanta-tion. Mol Hum Reprod 1998; 4: 735-738.
24) HALLORAN MM, CARLEY WW, POLVERINI PJ, HASKELL CJ,PHAN S, ANDERSON BJ, WOODS JM, CAMPBELL PL,VOLIN MV, BÄCKER AE, KOCH AE. Ley/H: an endothe-lial-selective, cytokine-inducible, angiogenic medi-ator. J Immunol 2000; 164: 4868-4877.
25) DOMINO S E, ZHANG L, GILLESPIE PJ, SAUNDERS TL,LOWE JB. Deficiency of reproductive tract α (1, 2)fucosylated glycans and normal fertility in micewith targeted deletions of the FUT1 or FUTα (1,2) fucosyltransferase locus. Mol Cell Biol 2001;21: 8336-8345.
3335
Effects of LeY glycan expression on embryo implantation