nouvelles stratégies thérapeutiques dans le syndrome de barth
TRANSCRIPT

Nouvelles stratégies thérapeutiques dans le syndrome de Barth
Chris Ottolenghi Biochimie Métabolique et Centre de Référence des
Maladies Métaboliques, Hôpital Necker, Paris
• Plasma amino acid profiling in BTHS • Transcriptional profiling in a mouse model • Metabolic flux analysis

INTRODUCTION
Nouvelles stratégies thérapeutiques dans le syndrome de Barth – SFEIM Nantes 2014

Syndrome de Barth (BTHS) • myocardiopathie, myopathie, neutropénie, retard de croissance • acidurie 3-méthylglutaconique modérée • mortalité importante avant 5 ans • maladie génétique très rare liée à l'X • Tafazzine (TAZ) :
• acyltransférase • remodelage des cardiolipines (phospholipides mitochondriaux)
Thérapie • « standard » : insuffisance cardiaque, neutropénie • métabolique débattue :
Coenzyme Q, ac.linoléique, ac.pantothénique, leucine, arginine • Essai clinique Bézafibrate en cours

Proposed mechanisms of pathology for BTHS TAZ deficiency : differential mitochondrial protein transport/assembly This may account for several mechanisms of pathology, including :
1) Respiratory chain deficiency-like features Mitochondrial structure abnormalities Intermittently high lactate and often 3-methylglutaconic aciduria Occasional lactic acidosis, hyperNH3, hypoglycaemia but : ATP production not impaired under basal metabolism 2) Krebs cycle dysfunction, possibly critical

- Krebs cycle dysfunction may preferentially involve isocitrate dehydrogenase
- Arginine could by-pass this block
Source: Richard Kelley (Kennedy Krieger Institute at John Hopkins)

1. PLASMA AMINO ACID PROFILES
IN BARTH SYNDROME PATIENTS (unpublished)
Nouvelles stratégies thérapeutiques dans le syndrome de Barth – SFEIM Nantes 2014

Plasma amino acids : 15 samples from 8 BTHS patients (France) vs single-hospital cohort (20408 samples, 5 yrs)

Plasma amino acids : age-adjusted standard deviations

Age-adjusted standard deviations, ranked by median deviation from the mean
Heatmap and clustering
Plasma amino acids from 15 BTHS samples

Three multivariate parameters (unsupervised and supervised) provide a strong plasma amino acid predictor of BTHS status
ROC AUC=98%

Principal component analysis on BTHS patient samples Correlation of single amino acids to PC axes :
PC3 = blood sampling technique ? PC4 = ??
PC1 = meal PC2 = nutritional status
ASN,ARG,SER,THR,LYS (r-squared > 0.15)
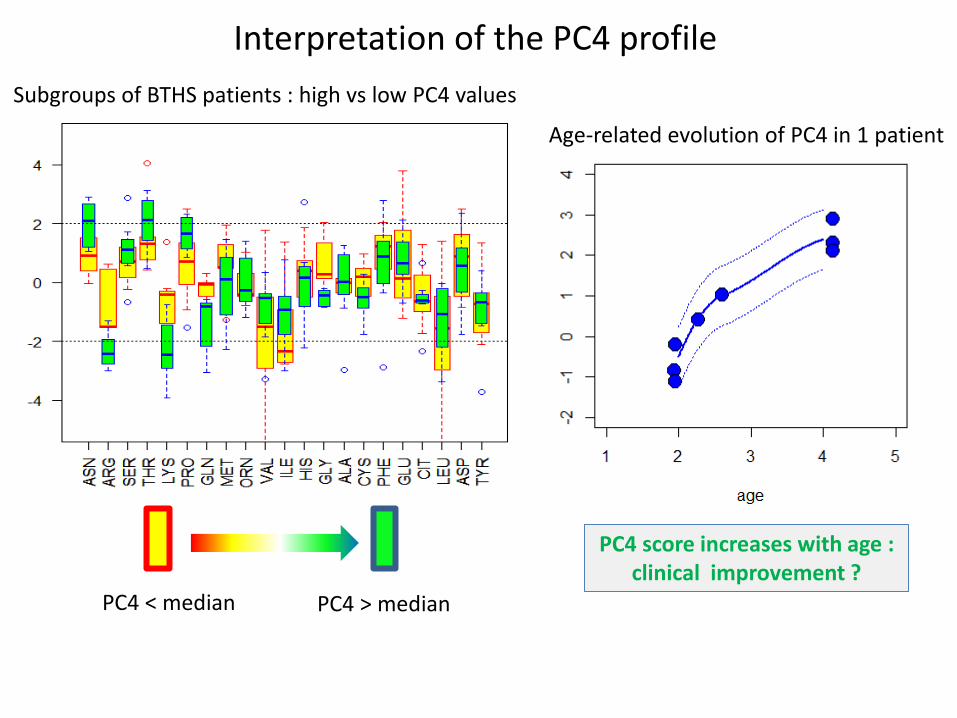
PC4 score increases with age : clinical improvement ?
Interpretation of the PC4 profile
PC4 < median PC4 > median
Subgroups of BTHS patients : high vs low PC4 values
Age-related evolution of PC4 in 1 patient

CONCLUSIONS 1 : - BTHS plasma amino acid profile (ASN, ARG, SER, THR, LYS)
• strong predictor by multivariate analysis (but test performance with additional BTHS patients)
• deviates with age and/or clinical improvement - Are amino acids reflecting a compensatory mechanism ? Are they markers or causal effectors ?

2. MOUSE MODEL OF BARTH SYNDROME
Nouvelles stratégies thérapeutiques dans le syndrome de Barth – SFEIM Nantes 2014
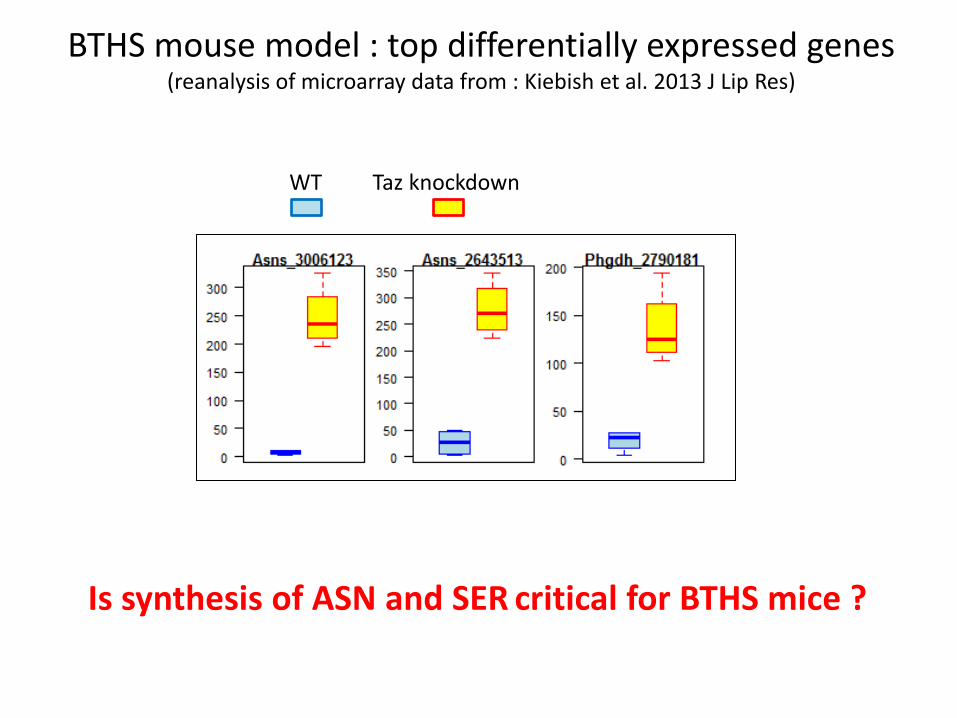
BTHS mouse model : top differentially expressed genes (reanalysis of microarray data from : Kiebish et al. 2013 J Lip Res)
WT Taz knockdown
Is synthesis of ASN and SER critical for BTHS mice ?

Barth Sd heart model :
WT TazKO
Fetal heart (dpc) :
16-18 12-14
Unpublished from Cardiogenomics Lab., Harvard Univ.
ASN SER : both fetal and BTHS (mouse heart)

ASN SER : top scoring hits genomewide Global correlation young fetus - BTHS
Early fetus
WT TazKD
ASN synthesis
SER synthesis
Myocardial transcriptome in BTHS mouse model (25697 genes) vs mouse fetal heart development (30506 genes) (unpublished)
Late fetus

CONCLUSIONS 2 :
- High plasma ASN and SER may reflect
- transcriptional upregulation in humans as in mice
- global reexpression of « fetal heart » transcriptome

3. METABOLIC FLUX ANALYSIS
Nouvelles stratégies thérapeutiques dans le syndrome de Barth – SFEIM Nantes 2014

Respirometry : amino acid relevance in BTHS mice. Kiebish M A et al. J. Lipid Res. 2013
Palmitoylcarnitine Glutamate
Overall respiratory chain activity : increased by amino acid (glutamate) in BTHS model vs WT. decreased by fatty acid (b-oxidation) in BTHS model vs WT.
WT
Taz knock-down
Oxy
gen
co
nsu
mp
tio
n
(mo
use
he
art
mit
och
on
dri
a)

m+5 / m ornithine
m+5 / m glutamate
ARG
GLU Krebs GLN
LEU,LYS
Metabolic flux analysis in patient fibroblasts by mass spectrometry (unpublished)
ISOTOPE RATIOS (reference = 1)
ORN
P5C
0
10
20
30
40
50
60
70
ref. R* R* K* L* Q*
BTHS
control
0
100
200
300
400
ref. R* R* K* L* Q* X100
BTHS
control
BTHS Increased ‘cataplerotic’ flux from glutamate to ornithine Different response of arginine catabolism to Leu + Lys supplements

ALDH18A1 ALDH4A1
Mouse BTHS model : more about the GLU – ARG connection WT TazKO
ARG
GLU Krebs GLN
ORN
P5C
BTHS (mouse model transcriptome) : Increased ‘cataplerotic’ flux from glutamate to ornithine may occur in part by regulation at the RNA level

CONCLUSIONS 3 : - BTHS Krebs cycle needs more, but gets less glutamate
- Arginine may not be the best anaplerotic substrate
- Role of amino acid context - Combined amino acids could improve therapy

SUMMARY
Nouvelles stratégies thérapeutiques dans le syndrome de Barth – SFEIM Nantes 2014

SUMMARY - BTHS is an interesting case of secondary aminoacidopathy = therapy !
- Asparagine, Serine, Threonine and Arginine, Lysine in plasma
= candidate markers for BTHS and/or cardiomyocyte differentiation
- Amino acid coktails may improve Barth syndrome therapy - Caveats :
- Developmental/aging factors - Accurate cell / mouse models of pathology - Other dysfunctions
- Perspectives - High resolution MS (GC qTOF) - Integrated multivariate analyses on open cohorts

REMERCIEMENTS Biochimie métabolomique et protéomique, Necker
Florence Habarou, Maxime Janin Bernadette Chadefaux, Robert Barouki Centre de Référence des Maladies Métaboliques, Necker Pascale de Lonlay and collaborators Autres collaborations : A.S. Lebre, M. Rio, A. Rötig, A. Munnich (Génétique, Necker) D. Bonnet (Cardiopédiatrie, Necker) J. Donadieu (Hémato-oncologie pédiatrique, Trousseau) Association Barth France


CKL Phoon 2012, Barth Sd Foundation conference
Several possible sources of heart pathology in BTHS
Amino acid metabolism


Pei W et al. Development 2010;137:2587-2596

Characterization of lymphoblast mitochondria from patients with Barth syndrome Yang Xu, John J Sutachan, Heide Plesken, Richard I Kelley and Michael Schlame
Lab Invest. 2007 Jan;87(1):40-8. Epub 2006 Oct 16. Comparison of lymphoblast mitochondria from normal subjects and patients with Barth syndrome using electron microscopic tomography. Acehan D1, Xu Y, Stokes DL, Schlame M.
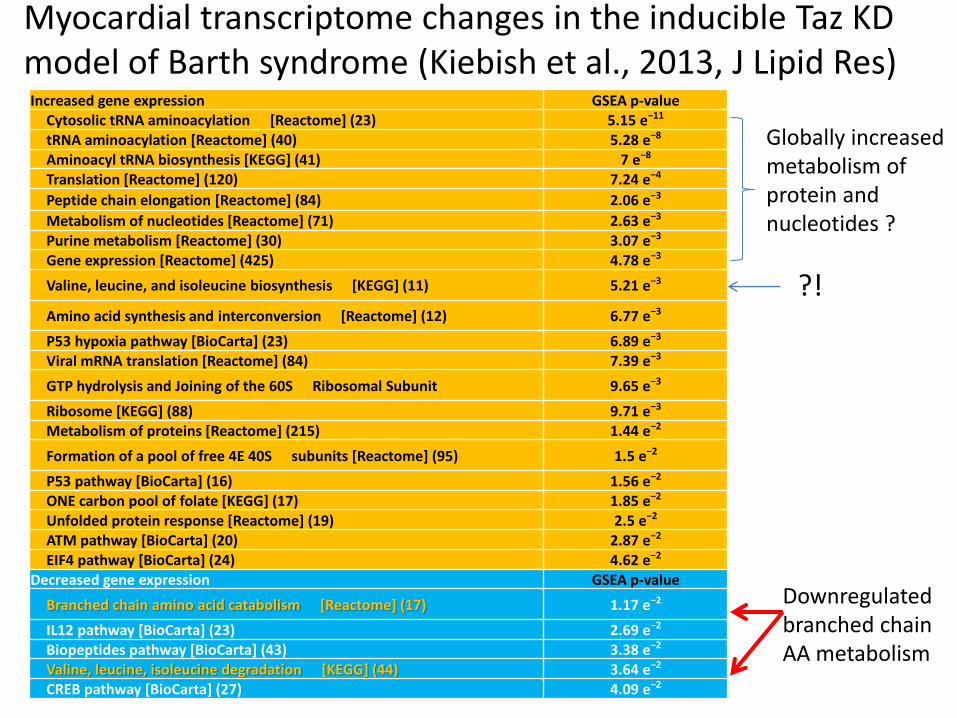
Increased gene expression GSEA p-value Cytosolic tRNA aminoacylation [Reactome] (23) 5.15 e−11 tRNA aminoacylation [Reactome] (40) 5.28 e−8 Aminoacyl tRNA biosynthesis [KEGG] (41) 7 e−8 Translation [Reactome] (120) 7.24 e−4
Peptide chain elongation [Reactome] (84) 2.06 e−3
Metabolism of nucleotides [Reactome] (71) 2.63 e−3 Purine metabolism [Reactome] (30) 3.07 e−3 Gene expression [Reactome] (425) 4.78 e−3
Valine, leucine, and isoleucine biosynthesis [KEGG] (11) 5.21 e−3
Amino acid synthesis and interconversion [Reactome] (12) 6.77 e−3
P53 hypoxia pathway [BioCarta] (23) 6.89 e−3 Viral mRNA translation [Reactome] (84) 7.39 e−3
GTP hydrolysis and Joining of the 60S Ribosomal Subunit 9.65 e−3
Ribosome [KEGG] (88) 9.71 e−3 Metabolism of proteins [Reactome] (215) 1.44 e−2
Formation of a pool of free 4E 40S subunits [Reactome] (95) 1.5 e−2
P53 pathway [BioCarta] (16) 1.56 e−2 ONE carbon pool of folate [KEGG] (17) 1.85 e−2 Unfolded protein response [Reactome] (19) 2.5 e−2 ATM pathway [BioCarta] (20) 2.87 e−2 EIF4 pathway [BioCarta] (24) 4.62 e−2 Decreased gene expression GSEA p-value
Branched chain amino acid catabolism [Reactome] (17) 1.17 e−2
IL12 pathway [BioCarta] (23) 2.69 e−2 Biopeptides pathway [BioCarta] (43) 3.38 e−2 Valine, leucine, isoleucine degradation [KEGG] (44) 3.64 e−2 CREB pathway [BioCarta] (27) 4.09 e−2
?!
Downregulated branched chain AA metabolism
Globally increased metabolism of protein and nucleotides ?
Myocardial transcriptome changes in the inducible Taz KD model of Barth syndrome (Kiebish et al., 2013, J Lipid Res)

- Gcn2, +igf1/GH