nonhuman transferrin receptor 1 is an efficient cell entry receptor...
TRANSCRIPT

Nonhuman Transferrin Receptor 1 Is an Efficient Cell Entry Receptorfor Ocozocoautla de Espinosa Virus
Yíngyún Caì,a Shuıqìng Yú,a Steven Mazur,a Lián Dong,b Krisztina Janosko,a Téngfe� i Zha�ng,a Marcel A. Müller,c Lisa E. Hensley,a
Sina Bavari,b Peter B. Jahrling,a Sheli R. Radoshitzky,b Jens H. Kuhna
Integrated Research Facility at Fort Detrick, National Institute of Allergy and Infectious Diseases, National Institutes of Health, Fort Detrick, Frederick, Maryland, USAa;United States Army Medical Research Institute of Infectious Diseases, Fort Detrick, Frederick, Maryland, USAb; Institute of Virology, University of Bonn Medical Centre,Bonn, Germanyc
Ocozocoautla de Espinosa virus (OCEV) is a novel, uncultured arenavirus. We found that the OCEV glycoprotein mediates entryinto grivet and bat cells through transferrin receptor 1 (TfR1) binding but that OCEV glycoprotein precursor (GPC)-pseu-dotyped retroviruses poorly entered 53 human cancer cell lines. Interestingly, OCEV and Tacaribe virus could use bat, but nothuman, TfR1. Replacing three human TfR1 amino acids with their bat ortholog counterparts transformed human TfR1 into anefficient OCEV and Tacaribe virus receptor.
Classified arenaviruses have been assigned to four phylogeneticgroups, the Old World (OW) viruses and clades A to C New
World (NW) viruses (1). Each arenavirus is adapted to a distinctvertebrate host, which maintains the virus chronically withoutdeveloping disease (2). The arenavirus glycoprotein (GP1 subunitof glycoprotein precursor [GPC]) primarily determines host tro-pism (3–5) by binding to one of several cell surface receptors. Theprimary receptor for two pathogenic OW arenaviruses, Lassa virus(LASV) and lymphocytic choriomeningitis virus (LCMV), is�-dystroglycan (6). All classified pathogenic clade B arenaviruses(i.e., Chapare, Guanarito [GTOV], Junín [JUNV], Machupo[MACV], and Sabiá) and at least two nonpathogenic clade B are-naviruses (i.e., Amaparí [AMAV] and Tacaribe [TCRV]) can usetransferrin receptor 1 (TfR1) to enter host cells. TfR1 seems to bethe primary cell entry receptor for these viruses (7), but TfR1-independent replication has been observed in vitro, as well as incertain laboratory-mouse models (8–12). Importantly, humanTfR1 is utilized only by the pathogenic clade B viruses but not bynonpathogenic AMAV or TCRV (13, 14). The GP1s of JUNV andMACV, but not of GTOV, efficiently bind the TfR1 ortholog of theMACV host, the big laucha (Calomys callosus). All three GP1s ofGTOV, JUNV, and MACV can utilize the TfR1 ortholog of theGTOV host, the short-tailed zygodont (Zygodontomys brevi-cauda). JUNV GP1 efficiently binds to the TfR1 ortholog of itshost, the drylands laucha (Calomys musculinus), whereas GTOVand MACV GP1 do not (15). TfR1 orthologs from related rodents,the house mouse (Mus musculus) and brown rat (Rattus norvegi-cus), do not support infection by GTOV, JUNV, or MACV (15).Finally, the nonpathogenic AMAV and TCRV efficiently use theTfR1 orthologs of their primary hosts, the common neacomys(Neacomys spinosus) and the Jamaican fruit-eating bat (Artibeusjamaicensis), respectively. Whereas TCRV can bind to both TfR1s,AMAV cannot use the bat receptor, but both viruses can use short-tailed zygodont and big laucha TfR1 (16). These observations in-dicate that TfR1, and especially TfR1 orthologs, determine therelative (but not necessarily absolute) permissibility of individualmammal hosts to NW clade B arenaviruses.
Between 1545 and 1815, viral hemorrhagic fever (VHF) epi-demics that may have been caused by arenaviruses were recordedin Mexico (17, 18). In 1967, a VHF epidemic occurred in Chiapas
State among several hundred Maya people. Low titers of comple-ment-fixing antibodies against arenaviruses found in survivorssuggested an arenavirus etiology (19). In 2012, Cajimat et al. se-quenced the genome of a novel arenavirus, Ocozocoautla de Es-pinosa virus (OCEV), found in kidneys of Mexican deer mice(Peromyscus mexicanus) captured in Chiapas State (20). OCEVcould not be isolated, but phylogenetic studies placed the OCEVgenome firmly in NW clade B. Consequently, Cajimat et al. spec-ulated that OCEV could have been the etiological agent of the 1967Mexican epidemic.
To identify OCEV’s cell entry determinants, we first evaluatedwhether the OCEV genome encodes a functional GPC. Retrovi-ruses pseudotyped with arenavirus glycoproteins have been usedsuccessfully to characterize arenavirus cell entry determinants (10,13, 15, 16, 21–24). We therefore produced enhanced green fluo-rescent protein (eGFP)-expressing Moloney murine leukemia vi-rus (MoMLV) pseudotyped with control (TCRV, MACV, orLASV) GPC (15, 16, 25) or OCEV GPC (synthesized by DNA2.0based on GenBank accession number JN897398) and evaluatedtransduction efficiencies in cells obtained from ATCC. Grivet(Chlorocebus aethiops) Vero E6 cells (Fig. 1A) and human HeLacells (Fig. 1B) were exposed to pseudotypes for 5 h and imaged foreGFP 48 h later. As published previously, MACV and LASV pseu-dotypes transduced Vero E6 (Fig. 1A) and HeLa (Fig. 1B) cellsefficiently (10, 16, 21, 22). Although not as efficiently as MACVand LASV pseudotypes, OCEV pseudotypes entered Vero E6 cellsat levels comparable to those of TCRV, indicating that the OCEVGPC is functional.
Since OCEV pseudotypes entered HeLa cells poorly (Fig. 1B),we expanded our screening of the four pseudotypes (MACV,LASV, OCEV, and TCRV) to 53 human NCI-60 panel cancer cell
Received 17 September 2013 Accepted 3 October 2013
Published ahead of print 9 October 2013
Address correspondence to Jens H. Kuhn, [email protected], or Sheli R.Radoshitzky, [email protected].
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.02701-13
13930 jvi.asm.org Journal of Virology p. 13930 –13935 December 2013 Volume 87 Number 24
on Novem
ber 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

lines (26) and measured transduction efficiencies by fluorescence-activated cell sorting (FACS) (Fig. 1C). As expected, LASV andMACV pseudotypes efficiently transduced most human cancercells (data not shown). OCEV pseudotype transduction was ex-tremely low in 52 of 53 tested cell lines, with some lines reachingthe low level of OCEV susceptibility of Vero E6 cells (Fig. 1C, redline) and one cell line (SNB19) being somewhat more susceptiblethan Vero E6 cells (Fig. 1C).
To identify cells that are highly permissive to OCEV, wescreened human 293T/17, Egyptian rousette (Rousettus aegyptia-cus) RoNi/7.2 (27), Büttikofer’s epauletted fruit bat (Epomopsbuettikoferi) EpoNi/22.1 (27), grivet BS-C-1, Chinese hamster(Cricetulus griseus) CHO-K1, Syrian hamster (Mesocricetus aura-tus) BHK-21, house mouse (Mus musculus) NIH/3T3 and M2-10B4, hispid cotton rat (Sigmodon hispidus) CRL, and domesticdog (Canis familiaris) MDCK cells (Fig. 2). Results obtained withgrivet BS-C-1 cells were similar to those obtained with Vero E6cells. As expected (10, 16, 21), hamster and house mouse cells were
largely refractory to transduction with MACV, OCEV, and TCRVpseudotypes but were susceptible to LASV pseudotypes. Hispidcotton rat cells proved to be excellent targets for LASV and MACVpseudotypes but were largely resistant to TCRV and OCEV pseu-dotype transduction. Dog cells were somewhat susceptible toLASV pseudotype transduction but not permissive to other pseu-dotypes. Unexpectedly, OCEV pseudotypes transduced RoNi/7.2and EpoNi/22.1 cells with high efficiency and TCRV, MACV, andLASV pseudotypes also entered both bat cell lines at lower effi-ciencies (Fig. 2). These results identified the two bat cell lines assuitable tools for further OCEV studies.
Since OCEV clusters with clade B NW arenaviruses, we hy-pothesized that TfR1 is an OCEV receptor. We cloned Egyptianrousette TfR1 (reTfR1) by extracting cellular RNA from RoNi/7.2cells using an RNeasy minikit (Qiagen) followed by SuperScriptIII one-step reverse transcription (RT)-PCR (Life Technologies)using primers 5=-ATGATGGATCAAGCCAGATCAGCAWTCTCT-3= and 5=-AAAYTCATTGTCAATGTCCCAAAYGTCACCA-
FIG 1 OCEV glycoprotein GPC mediates cell entry. eGFP-encoding MoMLV was pseudotyped with OCEV, TCRV, MACV, or LASV GPC and used to transducegrivet Vero E6 (A), human HeLa (B), or human NCI-60 panel (C) cells. (A and B) Images of transduced cells (blue, DAPI stained). (C) Quantification oftransduction of OCEV pseudotype using FACS. Shown are the results of one of two independent, but highly similar, NCI-60 screens. Error bars indicate standarddeviations for duplicate samples.
Nonhuman TfR1 Is an OCEV Entry Receptor
December 2013 Volume 87 Number 24 jvi.asm.org 13931
on Novem
ber 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

3=. Amplified PCR products were cloned by bidirectional TOPO TAcloning (Life Technologies), and five clones were analyzed. A newset of primers, 5=-CACCATGATGGATCAAGCCAGATCAGCA-3= and 5=-TTACTTATCGTCATCGTCCTTGTAGTCAAATTCATTGTCAATGTCCCA-3=, was used to clone reTfRI with aC-terminal FLAG tag using a pcDNA3.1 directional TOPO ex-pression kit (Life Technologies). Following protocols and reagentspreviously described (15, 16), we transiently overexpressed FLAG-
tagged control human (hTfR1), house mouse (mTfR1), Jamaicanfruit-eating bat (ajTfR1), big laucha (ccTfR1), common neacomys(nsTfR1), cat (fTfR1), or reTfR1 or transfected empty plasmidinto naturally hTfR1-expressing HEK 293T/17 cells and trans-ducedthese cells with OCEV, TCRV, MACV, or LASV pseu-dotypes (Fig. 3). Anti-FLAG antibody (Sigma-Aldrich) detectedsurface TfR1s (Fig. 3A). Unsurprisingly, LASV pseudotypes,which are dependent on �-dystroglycan for cell entry, entered
FIG 2 OCEV pseudotypes transduce bat cells with high efficiency. MoMLV pseudotyped with the indicated arenavirus GPCs were used to transduce cells fromthe indicated mammals, followed by imaging.
Caì et al.
13932 jvi.asm.org Journal of Virology
on Novem
ber 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
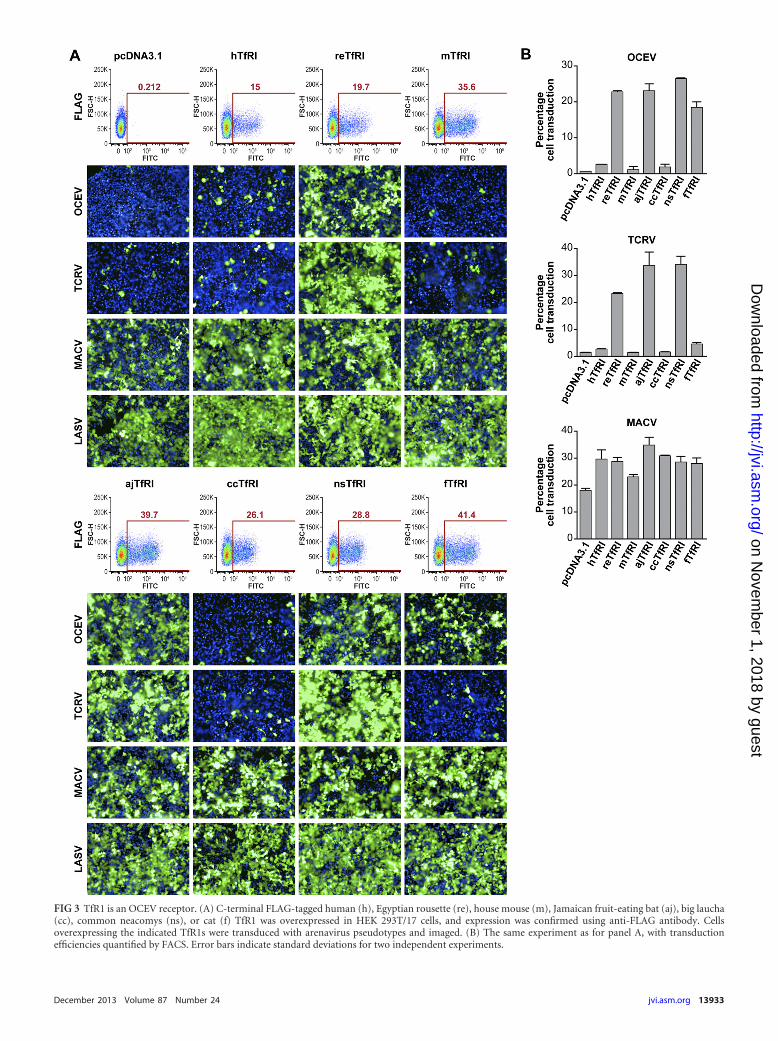
FIG 3 TfR1 is an OCEV receptor. (A) C-terminal FLAG-tagged human (h), Egyptian rousette (re), house mouse (m), Jamaican fruit-eating bat (aj), big laucha(cc), common neacomys (ns), or cat (f) TfR1 was overexpressed in HEK 293T/17 cells, and expression was confirmed using anti-FLAG antibody. Cellsoverexpressing the indicated TfR1s were transduced with arenavirus pseudotypes and imaged. (B) The same experiment as for panel A, with transductionefficiencies quantified by FACS. Error bars indicate standard deviations for two independent experiments.
December 2013 Volume 87 Number 24 jvi.asm.org 13933
on Novem
ber 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from
HEK 293T/17 cells with roughly equivalent efficiencies indepen-dently of TfR1 expression. As expected, hTfR1 and ccTfR1 over-expression increased susceptibility to MACV pseudotypes com-pared to that for empty plasmid (Fig. 3B). However, these TfR1orthologs as well as mTfR1 did not appreciably affect cell permis-siveness to OCEV or TCRV pseudotypes. Bat (reTfR1 and ajTfR1)and nsTfR1 conferred susceptibility to OCEV and TCRV pseu-dotypes and increased cell susceptibility to MACV pseudotypescompared to that of the empty-plasmid controls. Interestingly,OCEV could utilize fTfR1, which was also efficiently used by NWclade B hemorrhagic fever arenaviruses (15) and the nonpatho-genic AMAV, but not the nonpathogenic TCRV (16). Together,these results indicate that TfR1 is a receptor for OCEV.
A local region in the apical domain of TfR1 (residues 208 to212), and in particular residue Y211, determines GP1 bindingspecificity (15, 16, 23). Following published strategies (15, 16, 25),we aligned hTfR1 residues 201 to 216 with those of other orthologs(Fig. 4) and then created three FLAG-tagged h/reTfR1 chimeras.These ORFs were stably overexpressed in CHO-K1 cells using theFlp-In system (Life Technologies) (Fig. 5). Cells were exposed topseudotypes, imaged (Fig. 5A), and analyzed by FACS (Fig. 5B).
FIG 4 Alignment of TfR1 ortholog residues 201 to 216. Alignment of TfR1apical domain residues 201 to 216 of the indicated mammals and generatedhuman/Egyptian rousette TfR1 (h/re) chimeras. TfR1 apical domain residuescritical for NW clade B hemorrhagic fever arenavirus GP1 binding (residues208 to 212) are underlined, and critical residue Y211 is shown in green. Resi-dues deviating from the hTfR1 sequence are shown in red.
FIG 5 Three amino acid changes turn human TfR1 into a functional OCEV receptor. Effect of TfR1 chimera expression on arenavirus pseudotype transduction.(A and B) Stable expression in CHO-K1 cells. (A) Confirmation of expression of FLAG-tagged chimera TfR1 using anti-FLAG antibody (FACS) and images oftransduced cells. (B) FACS analysis of transduction efficiency. Error bars indicate standard deviations for two independent experiments.
Caì et al.
13934 jvi.asm.org Journal of Virology
on Novem
ber 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from

As little as three amino acid changes (h/re2TfR1, GRL206 to209¡SPT) transformed hTfR1 into a functional OCEV andTCRV receptor.
The results of our study indicate that OCEV, as it currentlycirculates in Mexico, cannot use human TfR1 efficiently to enterhuman cells. The amino acid sequence of OCEV GP1 supports thisconclusion. In the case of MACV GP1, residues R111, Y122, andD123 were found to be essential and residues D155 and P160 to beimportant for direct engagement of human TfR1 (28). Of theseresidues, only D123 and P160 are conserved in OCEV GP1. Fur-thermore, the three bat ortholog residues that transformed hu-man TfR1 into an efficient OCEV receptor are located within TfR1motif 5. These human TfR1 residues contact MACV GP1 residuesS97 and F98 (23). Interestingly, S97 is conserved among GTOV,JUNV, MACV, and SABV (23) but T occupies the same positionin OCEV and TCRV. Mutational studies with OCEV GP1 shouldbe performed to elucidate which, and how many, mutationswould be necessary for adaptation of OCEV to human TfR1.
ACKNOWLEDGMENTS
We thank Maria N. B. Cajimat and Charles F. Fulhorst (University ofTexas Medical Branch, Galveston, TX, USA) for providing the OCEVGPC amino acid sequence before it was deposited in GenBank. We alsothank Michael Farzan (The Scripps Research Institute, Jupiter, FL) forproviding plasmids expressing TCRV GPC and FLAG-tagged ajTfR1 andnsTfR1. We are grateful to our colleagues Jiro Wada and Laura Bollingerfor assisting us with the preparation of figures and critically editing themanuscript.
Y.C. and J.H.K. performed this work as employees of Tunnell Con-sulting, Inc., S.M. as an employee of MRI Global, T.Z. as an employee ofLovelace, and S.Y. and K.J. as employees of Battelle Memorial Institute, allunder Battelle’s prime contract with NIAID under contract numberHHSN272200700016I. This work was in part funded by the Joint Scienceand Technology Office for Chemical and Biological Defense (proposalnumber TMTI0048_09_RD_T to S.B.).
The content of this publication does not necessarily reflect the views orpolicies of the U.S. Department of the Army, the U.S. Department ofDefense, or the U.S. Department of Health and Human Services or of theinstitutions and companies affiliated with the authors.
REFERENCES1. Salvato MS, Clegg JCS, Buchmeier MJ, Charrel RN, Gonzalez JP,
Lukashevich IS, Peters CJ, Romanowski V. 2011. Family Arenaviridae, p715–723. In King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ (ed), Virustaxonomy. Ninth report of the International Committee on Taxonomy ofViruses. Elsevier/Academic Press, London, United Kingdom.
2. Charrel RN, de Lamballerie X. 2010. Zoonotic aspects of arenavirusinfections. Vet. Microbiol. 140:213–220.
3. Buchmeier MJ, Southern PJ, Parekh BS, Oldstone MB. 1986. Molecularand topographic analysis of the glycoproteins of lymphocytic choriomen-ingitis virus. Med. Microbiol. Immunol. 175:71–72.
4. Parekh BS, Buchmeier MJ. 1986. Proteins of lymphocytic choriomenin-gitis virus: antigenic topography of the viral glycoproteins. Virology 153:168 –178.
5. Rojek JM, Spiropoulou CF, Kunz S. 2006. Characterization of the cellu-lar receptors for the South American hemorrhagic fever viruses Junin,Guanarito, and Machupo. Virology 349:476 – 491.
6. Cao W, Henry MD, Borrow P, Yamada H, Elder JH, Ravkov EV, NicholST, Compans RW, Campbell KP, Oldstone MB. 1998. Identification ofalpha-dystroglycan as a receptor for lymphocytic choriomeningitis virusand Lassa fever virus. Science 282:2079 –2081.
7. Demogines A, Abraham J, Choe H, Farzan M, Sawyer SL. 2013. Dualhost-virus arms races shape an essential housekeeping protein. PLoS Biol.11:e1001571. doi:10.1371/journal.pbio.1001571.
8. Campetella OE, Galassi NV, Sanjuan N, Barrios HA. 1988. Susceptibleadult murine model for Junin virus. J. Med. Virol. 26:443– 451.
9. Cuevas CD, Lavanya M, Wang E, Ross SR. 2011. Junin virus infectsmouse cells and induces innate immune responses. J. Virol. 85:11058 –11068.
10. Flanagan ML, Oldenburg J, Reignier T, Holt N, Hamilton GA, MartinVK, Cannon PM. 2008. New World clade B arenaviruses can use trans-ferrin receptor 1 (TfR1)-dependent and -independent entry pathways,and glycoproteins from human pathogenic strains are associated with theuse of TfR1. J. Virol. 82:938 –948.
11. Kolokoltsova OA, Yun NE, Poussard AL, Smith JK, Smith JN, SalazarM, Walker A, Tseng CT, Aronson JF, Paessler S. 2010. Mice lackingalpha/beta and gamma interferon receptors are susceptible to Junin virusinfection. J. Virol. 84:13063–13067.
12. Weissenbacher MC, Calello MA, Quintans CJ, Panisse H, WoyskowskyNM, Zannoli VH. 1983. Junin virus infection in genetically athymic mice.Intervirology 19:1–5.
13. Radoshitzky SR, Abraham J, Spiropoulou CF, Kuhn JH, Nguyen D, LiW, Nagel J, Schmidt PJ, Nunberg JH, Andrews NC, Farzan M, Choe H.2007. Transferrin receptor 1 is a cellular receptor for New World haem-orrhagic fever arenaviruses. Nature 446:92–96.
14. Helguera G, Jemielity S, Abraham J, Cordo SM, Martinez MG, Rodrí-guez JA, Bregni C, Wang JJ, Farzan M, Penichet ML, Candurra NA,Choe H. 2012. An antibody recognizing the apical domain of humantransferrin receptor 1 efficiently inhibits the entry of all New World hem-orrhagic fever arenaviruses. J. Virol. 86:4024 – 4028.
15. Radoshitzky SR, Kuhn JH, Spiropoulou CF, Albariño CG, Nguyen DP,Salazar-Bravo J, Dorfman T, Lee AS, Wang E, Ross SR, Choe H, FarzanM. 2008. Receptor determinants of zoonotic transmission of New Worldhemorrhagic fever arenaviruses. Proc. Natl. Acad. Sci. U. S. A. 105:2664 –2669.
16. Abraham J, Kwong JA, Albarino CG, Lu JG, Radoshitzky SR, Salazar-Bravo J, Farzan M, Spiropoulou CF, Choe H. 2009. Host-species trans-ferrin receptor 1 orthologs are cellular receptors for nonpathogenic NewWorld clade B arenaviruses. PLoS Pathog. 5:e1000358. doi:10.1371/journal.ppat.1000358.
17. Marr JS, Kiracofe JB. 2000. Was the huey cocoliztli a haemorrhagic fever?Med. Hist. 44:341–362.
18. Acuna-Soto R, Romero LC, Maguire JH. 2000. Large epidemics of hem-orrhagic fevers in Mexico 1545–1815. Am. J. Trop. Med. Hyg. 62:733–739.
19. Goldsmith RS, Shields KP. 1971. Epidemic in southern Mexico of diseaseresembling virus haemorrhagic fevers. Lancet 2:151–154.
20. Cajimat MN, Milazzo ML, Bradley RD, Fulhorst CF. 2012. Ocozo-coautla de Espinosa virus and hemorrhagic fever, Mexico. Emerg. Infect.Dis. 18:401– 405.
21. Reignier T, Oldenburg J, Noble B, Lamb E, Romanowski V, BuchmeierMJ, Cannon PM. 2006. Receptor use by pathogenic arenaviruses. Virol-ogy 353:111–120.
22. Oldenburg J, Reignier T, Flanagan ML, Hamilton GA, Cannon PM.2007. Differences in tropism and pH dependence for glycoproteins fromthe clade B1 arenaviruses: implications for receptor usage and pathoge-nicity. Virology 364:132–139.
23. Abraham J, Corbett KD, Farzan M, Choe H, Harrison SC. 2010.Structural basis for receptor recognition by New World hemorrhagic feverarenaviruses. Nat. Struct. Mol. Biol. 17:438 – 444.
24. Reignier T, Oldenburg J, Flanagan ML, Hamilton GA, Martin VK,Cannon PM. 2008. Receptor use by the Whitewater Arroyo virus glyco-protein. Virology 371:439 – 446.
25. Martin VK, Droniou-Bonzom ME, Reignier T, Oldenburg JE, Cox AU,Cannon PM. 2010. Investigation of clade B New World arenavirus tro-pism by using chimeric GP1 proteins. J. Virol. 84:1176 –1182.
26. Weinstein JN. 2006. Spotlight on molecular profiling: “integromic” anal-ysis of the NCI-60 cancer cell lines. Mol. Cancer Ther. 5:2601–2605.
27. Kühl A, Hoffmann M, Müller MA, Munster VJ, Gnirss K, Kiene M,Tsegaye TS, Behrens G, Herrler G, Feldmann H, Drosten C, PöhlmannS. 2011. Comparative analysis of Ebola virus glycoprotein interactionswith human and bat cells. J. Infect. Dis. 204(Suppl. 3):S840 –S849.
28. Radoshitzky SR, Longobardi LE, Kuhn JH, Retterer C, Dong L, ClesterJC, Kota K, Carra J, Bavari S. 2011. Machupo virus glycoprotein deter-minants for human transferrin receptor 1 binding and cell entry. PLoSOne 6:e21398. doi:10.1371/journal.pone.0021398.
Nonhuman TfR1 Is an OCEV Entry Receptor
December 2013 Volume 87 Number 24 jvi.asm.org 13935
on Novem
ber 1, 2018 by guesthttp://jvi.asm
.org/D
ownloaded from