new species of troglobitic shrimps from · pdf filejose' luis villalobos, fernando...
TRANSCRIPT

JOURNAL OF CRUSTACEAN BIOLOGY, 19(1): 111-122, 1999
NEW SPECIES OF TROGLOBITIC SHRIMPS FROM MEXICO, WITH THE DESCRIPTION OF TROGLOMEXICANUS, NEW GENUS
(DECAPODA: PALAEMONIDAE)
Jose' Luis Villalobos, Fernando Alvarez, and Thomas M. Ilifle
A B S T R A C T
A new genus of cavernicolous fresh-water palaemonid shrimp, Troglomexicanus, and two new species, 7: tarnaulipasensis and 7: huastecae, are described from El Abra limestone caves in the states of San Luis Potosi and Tamaulipas, northeastern Mexico. Troglomexicanus includes 7:perez-farfantar, formerly in the genus Troglocubanus. distributed also in El Abra limestone caves. The following characters shared by the 3 species support the creation of the new genus: rostrum with- out dorsal or ventral teeth, maxilla with single-lobed endite, bilobed epipodite of first maxilliped, second maxilliped without podobranch, and absence of antennal, hepatic, or branchiostegal spines of carapace. The maxilla with single-lobed endite and a bilobed epipodite of first maxilliped are two characters not shared with any other genus in the subfamily Palaemoninae.
AND METHODS Troglocubanus Holthuis, 1949, included six
four distributed in Cuba [ T . eigen- Individuals of the new species described herein were
1903), collected during the Blindcat Collecting Expedition to the manni ( H ~ ~ , ':ibarensis (chase, Sierra de El Abra, San Luis Potosi-Tamaulipas. Mexico,
1943)3 T. inermis (Chase, 1943)9 and ' in March 1997. Descriptions of the two caves where the cis (Rathbun, 1912)1, One in Jamaica (T ':a- new s ~ e c i e s were ca~tured. ~hvs ica l and chemical con-
Previous to this study, the palaemonid genus MATERIALS
.- , L , maicensis Holthuis, 1963), and one in M ~ X -ditiong encountered, and collecting details are as follows.
ico (T . perezfarfantae Villalobos, 1971). Troglocubanus as proposed by Holthuis Cueva del Nacimiento del Rio Frb , Ejido El Nacimiento,
Municipio de Gdmez Farias, Tamaulipas. Mbxico, 9 and (1949), included troglobitic species, unpig- 10 March 1997 (Fig. l).-The cave entrance, situated at mented, with reduced eyes lacking a cornea, 155 m elevation, is located in a dry channel on the moun- a carapace without hepatic or branchiostegal tain side immediately above the springs that give rise to
spines, and a mandible without a palp. Vari- Rio Frio. The small cave entrance leads about 10 m down over breakdown to a dirt-floored room. From this point,
able characters within the genus were the ros- a flowstone slope extends up through a well-decorated trum, which could be short and smooth or passage back to the level of the entrance. About 50 m well developed with many dorsal teeth, the from the entrance, a large dome pit is encountered. This
presence or absence of an antennal spine, the 12-m diameter pit drops 40 m to a deep lake. Underwa- ter, the cave descends over breakdown to a smooth-walled endite of the second maxilla which is bilobed phreatic passage at a depth of 50 m. This passage ex-
in the Antillean species and single-lobed in tends horizontally for about 100 m to a vertical shaft that the Mexican species, and the epipodite of the goes up to an isolated lake with a 25-m high dome above first maxilliped which is bilobed in the Mex- it. No water currents were noticed.
ican species. These important differences, Two ho~nogeneous but discrete water masses were found in the cave, based on in situ measurements of wa-
arising between the Mexican T. pereifarfan- ter quality taken with a Hydrolab Data Sonde 3 Multi- tae and the Antillean species, were recog- probe Logger (Hydrolab Corporation, P.O. Box 50116, nized and discussed by Villalobos (1971). Austin, Texas 78763). Measurements of depth, tempera-
However, Troglocubanus was preserved as a ture, specific conductance, pH, dissolved oxygen, and re- dox potential were taken at 2-s intervals over the course valid genus for all six species. of the dive. The upper 40 rn of the column in the cave
Based on the finding of two new species water were uniform at 21.52"C, 0.43 milliSiernens/cm from Mexico, collected in caves that are part (pS1cm) specific conductance, 3.4 mgll dissolved oxygen, of the same formation as the type locality for pH 7.76, and 361 mV redox potential. Below 41 m, tem-
perature, specific conductance, dissolved oxygen, and re- T. peredarfantae, in which these important dox potential increased slightly to 21.63"C, 0.52 pSIcm, morphological differences between the An- 4.4 mgll, and 363 mV, respectively, while pH decreased tillean and Mexican species are preserved, a slightly to 7.72. Both sets of values were markedly dif- new genus, Troglomexicanus, is proposed. ferent from the Rio Frio springs located below the cave

112 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 19, NO. 1 . 1999
Fig. 1. Map showing the location of the type localities of the species of Troglornexicanus, new genus, in the Sierra de El Abra, within the states of San Luis Potosi and Tamaulipas, MCxico.
entrance. Spring water measured at 12 separate sites had markedly increased temperature, specific conductance, dissolved oxygen, and redox potential at 23.05"C. 0.73 pSlcm, 5.4 mgll, and 380 mV, respectively, while pH de- creased to 7.70. These data suggest that the spring and cave waters are not directly connected and, considering that the spring-water parameters significantly extend the depth trends observed in the cave, come from consider- able depths.
Shrimps were collected by hand in individual vials. The shrimps were observed swimming in the water col- umn primarily at depths below the 40-m chemocline. Troglobitic catfish (Ictaluridae, Prietella),mysids, isopods (Cirolanidae and Trichoniscidae), gastropods, and bran- chiobdelid worms were also collected from the same lo- cations within the cave.
Manantial de Sun Rafael de Los Castro, Municinio de Ciudad Mante, ~ a m a > i ~ a s , Mexico, 13 March 1997 (Fig. l).-The s~ r ingentrance is located at the southern edge of the village :f San Rafael de Los Castros at an eleva- tion of 98 m. The water surface is accessible at the bot- tom of a 4-m deep pit, but flows under a short natural bridge before emerging permanently on the surface. From this pit, the underwater cave continues as an irregular series of small to medium-sized chambers before adeep fissure extends to depths in excess of 60 m. The flow rate from the spring was very low at the time of visitation.
Water quality measurements were made at 2-s inter- vals with a Hydrolab Data Sonde 3. The water mass
within the cave was homogeneous at 27.3"C, 0.76 pS1cm specific conductance, 0.9 mgll dissolved oxygen, pH 7.47, and 335 mV redox potential. These values show markedly higher temperatures and lower dissolved oxygen than those at Cueva del Nacimiento del Rio Frio.
Shrimps were collected by hand primarily from sec- tions of the cave near the entrance at 0-15-m water depths. Troglobitic catfish (Ictaluridae, Prietella), isopods (Stenasellidae and Cirolanidae), and amphipods were also collected.
Material of the new species treated herein is deposited in the Colecci6n Nacional de Crustaceos, Instituto de Bio- logia, Universidad Nacional Aut6noma de Mexico (CNCR). The abbreviations used are: cl = carapace length and tl = total length.
RESULTS
Troglomexicanus, new genus Troglocuhanus Villalobos, 1971: 1; Hobbs et al., 1977:
64; Reddell, 1981: 16; Holthuis, 1986: 607.
Diagnosis.--Carapace smooth without an-tennal. he~atic. or branchiostegal s~ ines . An-terOlaierafmargin with roundeisudorbital an- gle. Rostrum short, devoid of dorsal or ven- tral spines. Eyes reduced, cornea absent, unpigmented. Mandible without palp, maxilla

113 VILLALOBOS ETAL.: NEW SPECIES OF TROGLOBITIC SHRIMPS
with single-lobed endite, first maxilliped with bilobed epipodite, and second maxilliped without podobranch. First pair of pereiopods slender and chelate; second pair longer than first, with palm and fingers of chela elon- gated; remaining pereiopods not chelate, with simple dactyl. Propodus of fifth pair of pe- reiopods with sparse setae along posterior margin and with dense distal tuft.
Distribution.-The distribution range of the genus is confined to limestone caves in the El Abra Formation, part of the Sierra de El Abra, a small mountain range of 125 km, that runs north-south 150 km west from the coast of the Gulf of Mexico. Sierra de El Abra is located in the northeastern portion of the state of San Luis Potosi and the adjacent southwestern sec- tion of the state of Tamaulipas (Fig. 1).
Type Species.-Troglomexicanus perezfar-fantae (Villalobos, 197 1).
Etymology.-The genus name is derived from Greek troglo (dwelling in a cave) and Mex- ican, since the new genus is distributed only in Mexican caves.
Troglomexicanus perezfarfantae (Villalobos, 197 1)
Troglocubanus perez&arfantae Villalobos, 197 1 : 1; Strenth, 1976: 13; Hobbs et al., 1977: 64; Holthuis, 1977: 181; Mitchell et al., 1977: 49; Reddell, 1981: 16; Villalo- bos, 1982: 218; Holthuis, 1986: 607; Villalobos et al., 1993: 281; Alvarez et a[., 1996: 110.
Diagnosis.-Carapace, eyes, mandible, max- illa, and first maxilliped same as in descrip- tion of genus. Rostrum short, not reaching distal end of first antennular segment, curv- ing upward. Hepatic and branchial grooves of carapace visible.
Abdomen smooth, sornites 1-5 with rounded pleura, pleura of sixth somite with posterior portion falcate. Telson shorter than external branch of uropods, longer than internal branch, posterior margin rounded, bearing 3 or 4 spines laterally and 2 pairs on distolateral angles, ex- ternal pair longer.
Antennular peduncle shorter than antenna1 scale, stylocerite short and blunt with deep dorsolateral groove on first article. External flagellum divided into 2 rami from fourth or fifth basal segment onward; internal ramus short with 10-12 segments. Antenna with an- tennal flagellum 3 times as long as total length of shrimp. Antenna1 scaphocerite reaching be- yond antennular peduncle.
First pair of pereiopods shorter than second pair. Ischium short, distal portion wider, bear- ing setae on internal margin. Merus longer than carpus. Second pair of pereiopods sub- equal, slender, longer than total length of shrimp. Surface of chela smooth with scat- tered setae, movable finger two-thirds length of cheliped, cutting edges of fingers smooth, without teeth. Carpus 2.5 times length of palm and longer than merus. Remaining 3 pairs of pereiopods increasing in length pos- teriorly.
Second pair of pleopods with an appendix interna more than half length of endopod. Uropods with external ramus longer than in- ternal one. Exopod bearing 2 spines on lat- erodistal angle. Endopod as long as telson.
Distribution.-Known only from the type lo- cality, S6tano de Tinaja (22"04'N, 89"56'W), 11.7 km east northeast from Ciudad Valles, San Luis Potosi, Mexico.
Remarks.-The single known specimen of this species, a female (USNM 1391 36) was collected by J. Fish and D. McKenzie in a large lake, 107 m long and 9 m wide, with a temperature of 22S°C, near the deepest sec- tion of the cave (Mitchell et al., 1977). Al- though Sotano de Tinaja has been explored Since 1946, and several expeditions have en-tered to collect blind fish, the holotype remains the only known specimen of the species,
Troglomexicanus tamaulipasensis, new species
Figs. 2, 3 Ho1otype.-5, cl 5.6 mm, tl 17.0 mm; 10 March 1997; Cueva del Nacimiento del Rio Frio (23"01fN, 99"04'W), Municipio de G6mez Farias, Tamaulipas; collected by T. M. Iliffe and D. A. Hendrickson; CNCR 17028.
A1lotype.-c, cl 8.1 mm, tl 22.0 mm; same date, locality, and collectors as holotype; CNCR 17029.
Paratypes.--4 x i , cl 5.0, 7.1, 7.5, 7.7 mm, tl 15.5, 21.0, 21.0, 21.5 mm; 3 I?, cl 5.5, 7.5, 8.2 mm, tl 16.8, 21.5, 23.0 mm; same date, locality, and collectors as holotype; CNCR 17030.
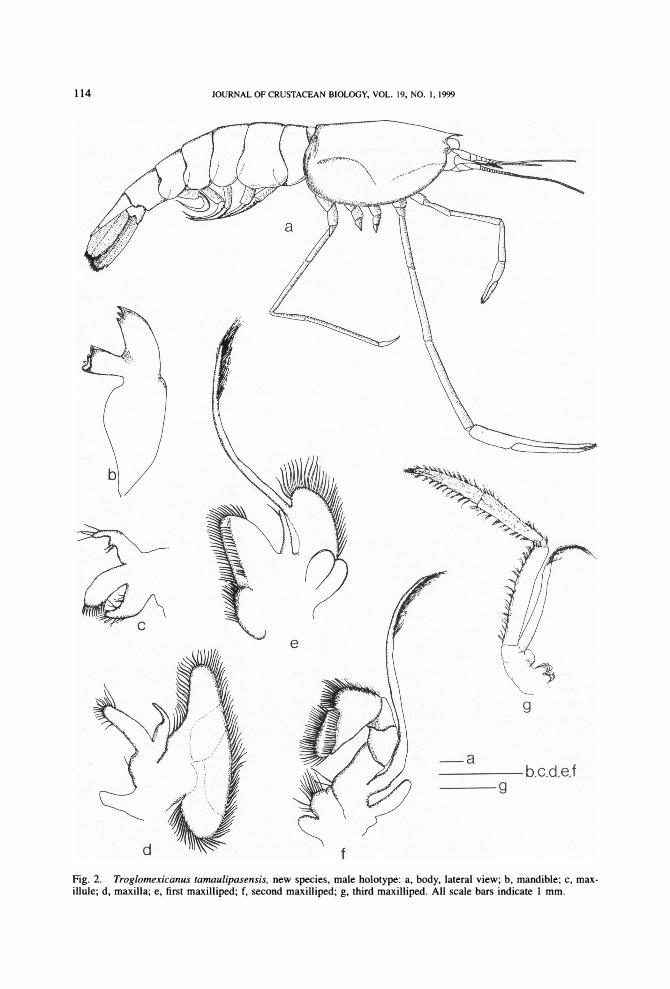
Description.-Rostrum slender, short, un-armed dorsally or ventrally, proximal half straight, distal half curving downward, end- ing in sharp tip, reaching slightly beyond oc- ular peduncles and proximal third of first an- tennular segment. Carapace smooth, with shallow hepatic depression and without an- tennal branchiostegal, or hepatic spines (Fig. 2a). Carapace with anterior margin projected next to rostrum, partially covering ocular pe-
JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 19, NO. 1,1999
Fig. 2. Troglomexicanus tamaulipasensis, new species, male holotype: a, body, lateral view; b, mandible; c, max- illule; d, maxilla; e, first maxilliped; f, second maxilliped; g, third maxilliped. All scale bars indicate 1 mm.

VILLALOBOS E T A L : NEW SPECIES OF TROGLOBITIC SHRIMPS
Fig. 3. Troglomexicanus tamaulipasensis, new species, male holotype (except g): a, dorsal view of anterior portion of carapace; b, antennule; c, antenna and antenna1 scale; d, first pereiopod; e, second pereiopod; f, dorsal view of telson and uropods; g, second pleopod of female; h, second pleopod of male; i, appendix masculina. All scale bars indicate 1 mm.
duncles; anterolateral margin rounded at in- ferior orbital angle; posterior margin formed laterally by broadly rounded expansion over- lapping with first abdominal somite. Eyes re- duced, without cornea.
Abdomen smooth. Anterior margin of pleura of first somite overlapping carapace. Second somite with pleura broadly rounded and with shallow depression along inferior border. Somites 3-5 with posterolateral mar-
gin of pleura rounded (Fig. 2a). Sixth somite slightly longer than second, dorsally project- ing on top of anterior portion of telson, with rounded notch on posterolateral margin of pleura at articulation of uropod. Telson shorter than internal pair of uropods; with 2 pairs of small, articulated spines on dorsal surface next to lateral margin. Posterior pair next to posterior margin, anterior pair next to posterior one on distal fifth of length of

116 JOURNAL OF CRLISTACE.AN BIOLOGY. V O I - . 19. NO. I. 1999
telson. Posterior margin rounded, bearing 14 long, plumose setae, with 2 spines on each posterolateral angle, external one shortest (Fig. 3f).
Antennular peduncle shorter than antenna1 scale (Fig. 3a). Stylocerite strong, with sub- acute apex reaching distal fourth of first an- tennular segment. First antennular segment flattened and twice as long as second. Second segment with rounded, deep notch on ante- rior margin; external flagellum divided into 2 rami from fourth basal segment onward, short ramus with 16 segments (Fig. 3b).
Antenna with scaphocerite reaching beyond antennular peduncle, twice as long as broad, with lateral margin ending distally in small acute spine, not reaching distal margin.
Mandible without palp, elongate molar pro- cess, incisor process with 3 conical teeth (Fig. 2b).
Maxillule with bilobed palp: distal lobe slender with 4 long setae, 2 anterior, 2 on apex; proximal lobe strong, with cylindrical projection on posterior margin; anterior lacinia with mesial margin armed with sharp setae; posterior lacinia arched with fine setae along anterior and posterior margins (Fig. 2c).
Maxilla with scaphognathite bearing plu- mose setae, anterior lobe slender, thinner than posterior one; palp strong, decreasing in thick- ness distally, ending in tip devoid of setae; en- dite single-lobed and slim, with distal end rounded and bearing plumose setae (Fig. 2d).
First maxilliped with 3 endites bearing marginal and submarginal setae (Fig. 2e). Ex- opod slender, 4 times length of palp, distal third bearing long setae; caridean lobe obvi- ous, joined to basal portion of exopod, with long setae along margins; palp strong proxi- mally, decreasing in thickness distally, with long seta on mesial margin. Epipodite com- posed of 2 lobes, approximately rounded, sep- arated by deep notch as long as epipodite; an- terior lobe wider than posterior one.
Second maxilliped subpediform and with- out podobranch, endopod 4-segmented, 2 dis- tal articles with gnathal border armed with spines and setae; exopod slender, almost twice as long as endopod; epipodite single- lobed, slender, and simple (Fig. 2f).
Third maxilliped pediform, thin, with dis- tal segment reaching beyond scaphocerite. Well-developed arthrobranch present, coxa with flattened, chitinous, rounded protuber- ance. Basis short. endopod and exopod joined
to basis at distal margin. Endopod 3-seg- mented, bearing tufts of setae, becoming denser distally; first segment thin, 1.3 times length of second segment; third segment end- ing in rounded tip, shorter than second seg- ment. Exopod slender, slightly longer than first segment of endopod; distal third bear- ing long setae (Fig. 2g).
First pair of pereiopods slender, shorter than second pair; carpus reaching beyond dis- tal end of scaphocerite, longer in males than in females. Ischium short, less than half of merus length, about half length of carpus and chela; proximal half of ventral border bear- ing wide notch; tufts of setae on distal half. Merus longer than carpus and chela, dorsal and ventral margins parallel, with tuft of se- tae ventrally on proximal half. Proximal two- thirds of carpus slender, becoming wider dis- tally, of about same length as chela, with scat- tered setae dorsally and distal tuft of setae ventrally. Chela slightly thicker than distal portion of carpus; palm three-fourths length of fingers, with ventral tuft of setae on prox- imal portion. Fingers slender, straight, leav- ing small gap when closed, with tufts of se- tae becoming denser distally. Cutting edges of fingers chitinized, devoid of teeth. Tips of fingers sharp, corneous, with tuft of setae (Fig. 3d).
Second pair of pereiopods subequal, elon- gate, devoid of setae except for 1 tuft on tip of each finger; distal third of merus reaching beyond scaphocerite. Length ratios of seg- ments as follows. Ischium shortest, 0.6-0.8 times length of merus, 0.5-0.7 times length of carpus, and 0.6-0.8 times length of dactyl. Merus shorter than carpus and of same length as dactyl. Carpus of same length as dactyl or slightly longer. Chela longer than carpus, 1.28-1.46 times its length; palm slightly thicker than carpus, about half length of dactyl. Fingers thin, elongate, with sharp cor- neous tips; cutting edges devoid of teeth, closing completely (Fig. 3e).
Remaining pereiopods thin, increasing in length from third to fifth pair; smooth with sparse short setae near articulations of seg- ments, along posterior margin of propodus, and on middorsal portion of dactyl. In all pairs, dactyl being shortest segment, simple, ending in sharp tip.
Third pair of pereiopods with distal margin of carpus reaching beyond scaphocerite. Is- chium slightly more than half length of merus

117 VILLALOBOS ETAL.: NEW SPECIES OF TROGLOBITIC SHRIMPS
and propodus, shorter than carpus, and longer than dactyl. Merus 1.5 times length of carpus, 3 times length of dactyl, and about same length as propodus. Carpus shorter than prop- odus and twice length of dactyl. Propodus 3 times length of dactyl. Fourth pair of pereio- pods with distal third of carpus reaching be- yond scaphocerite. Ischium about half length of merus and propodus, shorter than carpus, and longer than dactyl. Merus 1.5 times length of carpus and 3 times that of dactyl, about same length as propodus. Carpus shorter than propodus and twice length of dactyl. Propo- dus 3 or more times length of dactyl.
Fifth pair of pereiopods with distal third of carpus reaching beyond scaphocerite. Is- chium slightly less than half length of merus and propodus, shorter than carpus, and twice length of dactyl. Merus 1.3 times length of carpus, 3 times length of dactyl, and slightly shorter than propodus. Carpus shorter than propodus, more than twice length of dactyl. Propodus 4 times length of dactyl.
Pleopods with well-developed rami, except for first pair in both sexes where endopod re- duced, without appendix interna (Fig. 3g). In male, second pair with appendix masculina slender, long, reaching distal third of endo- pod; bearing setae distally, 4 located on apex; appendix interna, slim, shorter than appen- dix masculina (Fig. 3h, i).
Uropods with both rami of subequal length, reaching beyond tip of telson with distal fourth. Protopod with external lobe ending in acute spine and medial lobe rounded. En- dopod bearing setae on posterior margin and on distal portion of lateral margins, posterior margin rounded. Exopod with straight exter- nal lateral margin ending in unmovable spine, with small, movable, sharp spine next to it on mesial margin. Diaeresis incomplete, appear- ing as border reaching middle of exopod sur- face; posterior margin broadly rounded, bear- ing setae (Fig. 3f).
Etymology.-The specific name is derived from Tamaulipas, the State where the species occurs.
Distribution.-Known only from the type lo- cality.
Remarks.-Troglomexicanus tamaulipasensis seems to be more closely related to 7: huaste-cue, new species, described below, than to 7: perez$arfantae. From the former, it differs in
having a slimmer body and rostrum, a rounded posterior margin of the telson, and the rami of the uropods of subequal length. The new species can be readily separated from 7: peredarfantae by its shorter rostrum, the tel- son shorter than uropods, with two pairs of lateral spines and rounded posterior margin.
Troglomexicanus huastecae, new species
Figs. 4, 5
Holot)?pe.--. cl 8.0 mm, tl 21.0 mm: 13 March 1997: Manantial de San Rafael de 10s Castro (22"46'N, 99"03'W), Municipio de Ciudad Mante, San Luis Potosi; collected by T. M. lliffe and D. A. Hendrickson; CNCR 1703 I .
Description.-Rostrum short, sharp, slightly oriented upward, reaching beyond distal mar- gin of ocular peduncles and proximal third of first segment of antennular peduncles; dor- sal and ventral margins devoid of teeth. Cara- pace smooth, without antennal, branchioste- gal, or hepatic spines; with shallow bran- chiostegal depression (Fig. 4a). Anterior portion of carapace produced next to rostrum, partially covering ocular peduncles; antero- lateral margin with inferior orbital angle slightly marked by smooth, rounded projec- tion. Posterolateral margin of carapace end- ing in broadly rounded section overlapping first abdominal somite. Eyes reduced, ocular peduncles partially exposed, cornea absent.
Surface of abdomen smooth. Anterolateral margin of pleura of first somite overlapping posterior margin of carapace. Second somite with pleura broadly rounded, with shallow depression on middle section of inferior mar- gin. Third to fifth somite with posterolateral angle of pleura rounded and projected poste- riorly (Fig. 4a). Sixth somite as long as second somite, with rounded notch on pos- terolateral margin of pleura at articulation of uropod, angles limiting rounded notch suba- cute; dorsal posterior margin slightly pro- jected on top of anterior portion of telson. Telson shorter than internal rami of uropods, 2 small, movable pairs of sharp spines dor- sally along lateral margin; first pair at ap- proximately four-fifths length of telson, sec- ond pair smaller, next to posterior margin. Posterior margin of telson almost straight, with 14 long, plumose setae, and with pair of spines on each posterolateral angle, exter- nal one smaller.
Antennular peduncle shorter than antennal scale; stylocerite strong, apex subacute, reach-

JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 19, NO. 1,1999
Fig. 4. Troglornexicanus huastecae, new species, female holotype: a, body, lateral view; b, mandible; c, maxillule; d, maxilla; e, first maxilliped; f, second maxilliped; g, third maxilliped. All scale bars indicate 1 mm.
VILLALOBOS ETAL: NEW SPECIES OF TROGLOBITIC SHRIMPS 119
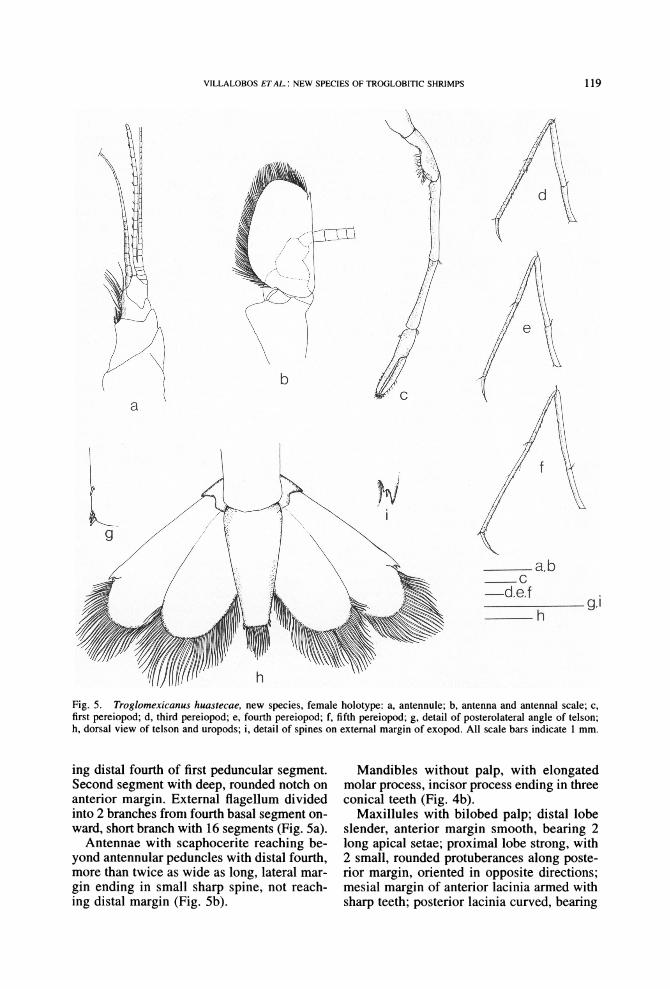
Fig. 5. Troglomexicanus huastecae, new species, female holotype: a, antennule; b, antenna and antenna1 scale; c, first pereiopod; d, third pereiopod; e, fourth pereiopod; f, fifth pereiopod; g, detail of posterolateral angle of telson; h, dorsal view of telson and uropods; i, detail of spines on external margin of exopod. All scale bars indicate 1 mm.
ing distal fourth of first peduncular segment. Second segment with deep, rounded notch on anterior margin. External flagellum divided into 2 branches from fourth basal segment on- ward, short branch with 16 segments (Fig. 5a).
Antennae with scaphocerite reaching be- yond antennular peduncles with distal fourth, more than twice as wide as long, lateral mar- gin ending in small sharp spine, not reach- ing distal margin (Fig. 5b).
Mandibles without palp, with elongated molar process, incisor process ending in three conical teeth (Fig. 4b).
Maxillules with bilobed palp; distal lobe slender, anterior margin smooth, bearing 2 long apical setae; proximal lobe strong, with 2 small, rounded protuberances along poste- rior margin, oriented in opposite directions; mesial margin of anterior lacinia armed with sharp teeth; posterior lacinia curved, bearing

120 JOURNAL OF CRGSTACLAN BIOLOGY. VOL. 19, NO. I . 1'199
thin setae along tip and anterior and poste- rior margins (Fig. 4c).
Scaphognathite of maxillae with sinuous margin bearing plumose setae. anterior lobe thinner than posterior lobe; palp devoid of se- tae, proximal third strong, tapering distally, ending in rounded apex; endite single-lobed, apex rounded, bearing thin, plumose setae (Fig. 4d).
First maxilliped with 3 endites bearing marginal and submarginal setae. Exopod slen- der, bearing long setae on distal third, 4 times as long as palp; caridean lobe well formed bearing long setae marginally, separated from basal portion of exopod by V-shaped cleft. Palp moderately flattened, strong, mesial mar- gin straight with long, thin subapical seta, lat- eral margin curving toward mesial margin distally, forming sharp tip. Epipodite formed by 2 strong, broadly rounded lobes, separated by deep cleft as long as half length of epipodite; anterior lobe wider than posterior one (Fig. 4e).
Second maxilliped subpediform and with- out podobranch, endopod 4-segmented, 2 dis-tal articles oriented mesially with gnathal bor- der bearing setae and spines. Exopod slender, distal half reaching beyond margin of endo- pod, bearing long setae distally. Epipodite single-lobed, with borders smooth (Fig. 4f).
Third maxilliped pediform, strong, distal segment reaching beyond scaphocerite; well- developed arthrobranch basally. Coxa simple, bearing flattened, chitinous, rounded protu- berance. Basis short, endopod and exopod joined to basis at distal margin. Endopod with 3 segments bearing tufts of setae, becoming denser distally; first segment 1.4 times length of second segment, third segment shorter than second segment, ending in rounded tip. Exo- pod slender, longer than first segment of en- dopod, distal third bearing long setae (Fig. 4g).
First pair of pereiopods slender and shorter than second pair. Ischium short, slightly more than half length of merus, carpus, and chela; ventral border with wide proximal notch and tufts of setae along distal half. Merus slightly longer than carpus and chela, dorsal and ven- tral margins parallel, tuft of setae on proxi- moventral margin. Proximal two-thirds of carpus slender, becoming wider distally, as long as chela, scattered setae on dorsal mar- gin, and tuft of setae on distoventral margin. Chela thicker than distal portion of carpus, palm shorter, about half length of fingers,
with proximoventral tuft of setae. Fingers slender, straight, with scattered setae becom- ing denser distally; leaving narrow space when closed, cutting borders smooth without teeth, tips corneous, with tuft of setae (Fig. 5c).
Second pair of pereiopods subequal, long, slender, with scattered setae; reaching beyond distal margin of scaphocerite with distal third of merus. Length ratios of segments as fol- lows. Ischium shortest, 0.76 times length of merus, 0.63 times length of carpus, and 0.72 times length of dactyl. Merus shorter than carpus (0.8) and slightly shorter than dactyl (0.95). Carpus longer than dactyl (1.14). Chela 1.4 times length of carpus; palm slightly thicker than carpus, more than half length of dactyl. Fingers thin, elongate, with sharp, cor- neous tips; cutting edges devoid of teeth, closing completely (Fig. 4a).
Remaining pereiopods slender, increasing in length from third to fifth pair; smooth with sparse short setae near articulations of seg- ments, along posterior margin of propodus, and on mid-dorsal portion of dactyl. In all pairs, dactyl shortest segment, simple, ending in sharp tip.
Third pair of pereiopods with ischium more than half length of merus and propodus, 0.9 times length of carpus, 1.75 times length of dactyl. Merus 1.5 times length of carpus, 3 times as long as dactyl, and of about same size as propodus. Carpus shorter than propo- dus and twice length of dactyl. Propodus 3 times length of dactyl (Fig. 5d).
Fourth pair of pereiopods with ischium more than half length of merus, shorter than carpus, half length of propodus, and 1.6 times length of dactyl. Merus 1.5 times length of carpus, 2.7 times length of dactyl, and slightly shorter than propodus. Carpus shorter than propodus and about twice length of dactyl. Propodus 3.2 times length of dactyl (Fig. 5e).
Fifth pair of pereiopods with ischium more than half length of merus, shorter than car- pus and propodus, and 1.6 times length of dactyl. Merus 1.3 times length of carpus and 2.8 times length of dactyl, shorter than prop- odus. Carpus more than half length of prop- odus and more than twice length of dactyl. Propodus 3.8 times length of dactyl (Fig. 5f).
Pleopods with well-developed rami, except for first pair where endopod reduced without appendix interna.
Uropods with internal branch slightly shorter than external one, reaching beyond

121 VILLALOBOS ETAL.: NEW SPECIES OF TROGLOBITIC SHRIMPS
distal margin of telson with distal fourth. Pro- topod with external lobe forming sharp spine and internal lobe rounded. Endopod with se- tae along posterior margin and distal portion of lateral margins, posterior margin rounded. Exopod with lateral margin straight, ending distally in unmovable spine, with internally small, sharp, movable spine; diaeresis incom- plete, appearing as border reaching middle of dorsal surface of exopod; posterior margin broadly rounded, bearing setae (Fig. 5h).
Etymology.-The specific name is derived from Huasteca, the name used for this region of Mexico.
Remarks.-Troglomexicanus huastecae seems to be more closely related to T. tamaulipasen-sis than to 7: perezlfalfantae, as discussed above. However, the three species are very similar. The caves in which 7: huastecae and T. tamaulipasensis were collected are closer to each other (27 km) than to S6tano de Tinaja, the type locality of i? perezlfalfantae, 75 and 102 km to the south.
KEY TO THE SPECIESOF TROGLOMEXICANUS
coastal plain of the Gulf of Mexico, through the border of the states of Tamaulipas and San Luis Potosi (Fig. 1). In the area, more than 40 caves have been registered so far, of which 27 are inhabited by cavernicolous fishes of the genus Astyanax (see Mitchell et al., 1977). The presence of blind fishes, the recording of permanent bodies of water, and the availability of large quantities of organic matter inside the caves, suggest that it is likely that more new species of decapod crus- taceans will be found. In the case of Troglo-mexicanus, S6tano de Tinaja (22"05'N, 89"56'W), the type locality of I: perezfalfan-tae, is located 75 and 102 km south from the type localities of i? huastecae and T. tamauli-pasensis, respectively, on the western slope in the southern tip of El Abra limestone (Fig. l), leaving an extensive area unexplored for crustaceans.
Troglomexicanus is clearly related to Trog-locubanus. However, following the ideas of Villalobos (1971) and Hobbs et al. (1977), the separation of the ancestral marine stock that invaded fresh water to become the Antillean and Mexican branches must have taken place
1. Telson longer than internal branch of uropods . . . . . . . . . . . . . . Troglomexicanus perezfa$antae
- Telson shorter than internal branch of uropods 2 2. Posterior margin of telson rounded . . . . . . . . . . . . .
. . . . . . . . . . . . . Troglomexicanus tamaulipensis - Posterior margin of telson straight . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . Troglomexicanus huastecae
D ~ s c u s s ~ o ~
Villalobos (1971), in the description of Troglocubanus perezlfalfantae, stated that the Mexican species of Troglocubanus differed from the Antillean species in three characters: the absence of spines on the carapace, the endite of the maxilla single-lobed, and the epipodite of the first maxilliped bilobed. Hobbs et al. (1 977) figured and presented di- agnosis of the six species that belonged to Troglocubanus, recording again the notice- able differences between the Antillean and the Mexican species. In this study, based on these three characters. not shared by any other genus of fresh-water shrimp of the subfam- ily Palaemoninae, the genus Troglomexicanus is proposed.
The species of Troglomexicanuscome from caves that have formed in the limestone of Sierra de El Abra, a small mountain range that runs along a north-south axis in the
very early in the evolution of the group. Chace and Hobbs (1969) postulated a Mio- cene origin for the Antillean species through separate invasions of Cuba and Jamaica. Vil- lalobos (1971) proposed a Cretaceous origin for the Mexican stock, coinciding with the probable time of uplifting of the coastal area from which the El Abra limestone originates (Mitchell et al., 1977). The different postu- lated times at which invasions of the fresh- water habitat may have taken place would suggest an older age for the Mexican stock. Under this scheme, the very specialized mouthparts of Troglomexicanus would be a consequence of a longer period of isolation. As pointed out by Chace and Hobbs (1969) and recognized by other authors, no epigean species closely related to Troglocubanus, and now to Troglomexicanus, can be identified, making the establishment of relationships of these two genera a difficult task.
Collection of the shrimp described here was part of the March 1997 Blindcat Collecting Expedition led by Dr. Dean A. Hendrickson. Major funding for the expedition was supplied by U.S.-Mexico Foundation for Science (Project 54-E-94). Specimens were collected under SE-MARNAP permit 3120 (29 August 1996) issued to Dr. Hendrickson. We thank DiveRite Manufacturing, Inc., and

122 JOURNAL OF CRUSTACEAN BIOLOGY. VOL. 19, NO. 1. 1999
U.S. Divers Co., lnc., for supplying diving equipment used during the expedition and the Texas Memorial Mu- seum for general support. We are especially grateful to Rolando Mendoza for the drawings. Members of the ex- pedition included Dean A. Hendrickson, James C. Brown, Thomas M. Iliffe, Thomas L. Morris, Jean Krejca, Doug Feakes, Hector Espinosa Perez, Francisco Javier Garcia de Leon, Tania Flores Martinez, Jost Manuel Padilla Gutitrrez, Juan Manuel Rodriguez Martinez, and Harris W. Martin. This is contribution number 82 of the Texas Memorial Museum, University of Texas at Austin, 2400 Trinity, Austin, Texas 78705.
Alvarez, F., J. L. Villalobos, and E. Lira. 1996. Decipo- dos.-In: J. Llorente, A. Garcia-Aldrete, and E. Gon-zilez, eds., Biodiversidad, taxonomia y biogeografia de artr6podos de MCxico: hacia una sintesis de su conocimiento. Instituto de Biologia, Universidad Na- cional Aut6noma de Mtxico, pp. 103-129.
Chace, Jr., F. A. 1943. Two new blind prawns from Cuba with a synopsis of the subterranean Caridea of Amer- ica.-Proceedings of the New England Zoological Club 22: 25-40.
,and H. H. Hobbs, Jr. 1969. The freshwater and terrestrial decapod crustaceans of the West Indies with special reference to Dominica.-United States National Museum Bulletin 292: 1-258.
Hay, W. P. 1903. On a small collection of crustaceans from the island of Cuba.-Proceedings of the United States National Museum 26: 429-435.
Hobbs, Jr., H. H., H. H. Hobbs 111, and M. A. Daniel. 1977. A review of the troglobitic decapod crustaceans of the Americas,-Smithsonian Contributions to Zo- ology 244: 1-183.
Holthuis, L. B. 1949. Note on the species of Palaemo- netes (Crustacea: Decapoda) found in the United States of America.-Proceedings, Koninklijke Nederlandse Akademie van Wetenschappen 52: 87-95.
. 1963. Two new species of fresh-water shrimp (Crustacea: Decapoda) from the West Indies.-Pro- ceedings, Koninklijke Nederlandse Akademie van Wetenschappen 66: 61-69.
. 1977. Subterranean fauna of Mexico.-Acca- demia Nazionale dei Lincei Quaderno 171 : 172-195.
. 1986. Decapoda.-In: L. Botosaneanu, ed.. Sty- gofauna mundi. Pp. 589-615. E. J. Brill, Leiden, The Netherlands.
Mitchell, R. W., W. H. Russell, and W. R. Elliot. 1977. Mexican eyeless characin fishes, genus Astyanax: en- vironment, distribution, and evolution.-The Museum, Special Publications No. 12, pp. 1-89. Texas Tech Uni- versity, Lubbock, Texas.
Rathbun, M. J. 1912. Some Cuban Crustacea, with notes on the Astacidae, by Walter Faxon, and a list of the Isopoda, by Harriet Richardson.-Bulletin of the Mu- seum of Comparative Zoology 54: 449-460.
Reddell. J . R. 1981. A review of the cavernicole fauna of Mexico, Guatemala, and Belize.-Texas Memorial Museum Bulletin 27: 1-327.
Strenth, E. N. 1976. A review of the systematics and zoogeography of the freshwater species of Palaenzo- netes Heller (Crustacea: Decapoda) of North Amer- ica.-Smithsonian Contributions to Zoology 228: 1-27.
Villalobos, A, 1971. Una nueva especie de Troglocu- banus (Crustacea: Decapoda: Palaemonidae), de San Luis Potosi, MCxico.-Anales del Instituto de Biologia, Universidad Nacional Aut6noma de MCxico, Serie Ciencias del Mar y Limnologia 42: 1-6.
. 1982. Decapoda.-In: S. H. Hurlbert and A. Villalobos, eds., Aquatic biota of Mexico, Central America, and the West Indies. Pp. 215-248. San Diego State University. San Diego, California.
Villalobos, J. L.. A. Cantti. and E. Lira. 1993. Los crustaceos de agua dulce de Mtxico.-Revista de la Sociedad Mexicana de Historia Natural, Volumen Es- pecial (XLIV): 267-290.
RECEIVED:10 December 1997 ACCEFTED: 19 May 1998.
Addresses: (JLV, FA) Colecci6n Nacional de Crusti- ceos, Instituto de Biologia, Universidad Nacional Aut6- noma de MCxico, Apartado Postal 70-153, MCxico 04510 D.F., MCxico. (e-mail: [email protected]); (TI) Department of Marine Biology, Texas A&M University at Galveston, Galveston, Texas 77553-1675, U.S.A.