neelanjan vishnu - uppsala university
TRANSCRIPT

Ribosome assisted protein folding
Neelanjan Vishnu
Degree project in applied biotechnology, Master of Science (2 years), 2010Examensarbete i tillämpad bioteknik 45 hp till masterexamen, 2010Biology Education Centre and Department of Cell and Molecular Biology, Uppsala UniversitySupervisor: Ass. Prof. Suparna Sanyal

Table of Contents Page no. Summary 3 1. Introduction 4 1.1 Protein structure & function 4 1.2 Protein folding 4 1.3 Central dogma, Protein synthesis & Ribosomes 6 1.4 Role of ribosome in Protein folding 7 1.5 Protein misfolding & Prion diseases 8 1.6 Ribosome as a target for Antiprion drugs 9 2. Aims & Objectives 12 2.1 Human carbonic anhydrase I 12 2.2 Ribosomes 12 2.3 Antiprion drugs 11 3. Results 12 3.1 Overexpression & Purification of Human carbonic anhydrase I 12 3.1.1 Overexpression of Human carbonic anhydrase I 12 3.1.2 DEAE Cellulose treatment of Human carbonic anhydrase I 12 3.1.3 Superdex- 75 gel filtration chromatography 13 3.1.4 Determination of Protein concentration 14 3.2 Effect of substrate on Human carbonic anhydrase I activity 15 3.2.1 Kinetics of Human carbonic anhydrase I activity 15 3.2.3 Effect of solvent for paranitrophenyl acetate on Human carbonic anhydrase I activity 16 3.2.4 Effect of Zn in refolding buffer on human carbonic anhydrase I activity 17 3.3 Human carbonic anhydrase I unfolding & refolding 17 3.3.1 Effect of EDTA on Human carbonic anhydrase I unfolding 17 3.3.2 Purification of 70S ribosomes 18 3.3.3 Effect of 70S ribosomes on Human carbonic anhydrase I refolding 19 3.3.4 Effect of varying concentration of 70S, 80S and ribosomal RNA Human carbonic anhydrase I refolding 19 3.3.4.1 Effect of varying concentration of 70S, 80S ribosomes on Human carbonic anhydrase I refolding 19 3.3.4.2 Effect of varying concentration of total ribosomal RNA isolated from 80S ribosomes on Human carbonic anhydrase I refolding 20 3.3.5 Timecourse of refolding of Human carbonic anhydrase I 21 3.4 Drug effects 21 3.4.1 Concentration profile of Psi207 (6AP) & Psi114 (GA) in relation to 70S ribosomes 21 3.4.2.1 Concentration profile of Psi207 (6AP) & Psi114 (GA) in relation to 80S ribosomes 22 3.4.3 Timecourse of refolding Human carbonic anhydrase I in the presence increasing concentration of Psi207 & Psi114 23 4. Discussion 23 5. Materials & Methods 25 5.1 Bacteria, Plasmid & Yeast strain 25
1

5.2 Buffers & Stock solutions 25 5.2.1 For Human carbonic anhydrase I purification 25 5.2.2 Human carbonic anhydrase I unfolding & refolding 26 5.2.3 For 70S ribosome purification 26 5.2.4 For 80S ribosome purification 27 5.2.5 For RNA preparation 28 5.2.6 Drugs 29 5.3 Methods 29 5.3.1 Overexpression & Purification of Human carbonic anhydrase I 29 5.3.2 Kinetics of Human carbonic anhydrase I 30 5.3.3 Human carbonic anhydrase I unfolding & refolding 30 5.3.3.1 Effect of EDTA on Human carbonic anhydrase I unfolding 30 5.3.4 Purification of 70S, 50S ribosomes 30 5.3.5 Purification of 80S ribosomes 31 5.3.6 Isolation of total RNA from ribosomes 31 5.3.7 Ribosome assisted protein folding 32 5.3.8 Total RNA assisted protein folding 32 5.3.9 Antiprion drug activity 32 6. Acknowledgement 33 7. References 33
2

Summary Prion diseases are devastating diseases which afflicts both animals and human equally and are fatal in most cases. The symptoms of this disease vary in different animals. Prion proteins are present in two forms of which PrPsc is the misfolded form of the protein with predisposition of β sheet structure. It has been hypothesized that it is caused by defective protein folding machinery leading to generation of misfolded protein. It has been shown experimentally that ribosome can do protein folding. In my project I have used human carbonic anhydrase I (HCA I) as model system and studied folding and denaturation process with or without ribosomes in relation to antiprion drug activity. Specifically I purified HCA I and standardized the kinetic conditions for its activity wherein I studied the effect of solvent, metal ion and buffers on HCA I activity. Unfolding and refolding conditions for studying ribosome assisted refolding was then characterised. In these assays the self folding level of HCA I was compared in time course experiments with ribosome and its subunit assisted refolding. Finally the inhibitory effect of antiprion drugs on the ribosome assisted refolding was carried out. It was clearly seen from the experiments that antiprion drugs affected the initial fast phase of the ribosome assisted refolding, after which even increase in duration of the incubation with ribosomes did not have significant impact on the inhibition. Further experiments are being performed to determine the kinetic parameters of the inhibition and would be related with other biophysical experiments.
3

1. Introduction 1.1 Protein structure and function Proteins are polymers of amino acids linked together as a linear polypeptide chain. The amino terminal of the polypeptide chain is called N terminus while the carboxy terminus of the protein is called C terminus (Lehninger et al. 2000). Secondary structure of protein consists of local spatial arrangement of polypeptides without regard to conformation of the side chains. Basically there are two forms of secondary structure taken by protein backbones which include α helix and β sheet. In α helix, backbone hydrogen bonds are arranged in such a way so that peptide C=O bond of nth residue points along the helix axis towards the peptide N-H group of the (n+4)th residue (Lehninger et al. 2000). In the β sheet structure hydrogen bonds are between the neighboring peptide chains and are classified into parallel and anti parallel sheets. In tertiary structure the protein is in folded form where the secondary structure elements are arranged so as to give proper 3D orientation to all atoms including the backbone (Lehninger et al. 2000). The major stabilizing forces in this state are noncovalent interactions among the side chain which mainly consists of hydrogen bonds, hydrophobic effects and electrostatic interactions (Stryer, 1995). Electrostatic interactions generally consist of short range forces and do not contribute majorly to the stability of the proteins. Hydrogen bonds on the other hand play important role in maintaining the secondary structure of the proteins. Hydrophobic forces are necessary to maintain the overall 3D structure of the proteins which is based on the fact that hydrophobic amino acids are favoured more towards the interior of the protein core and the hydrophilic amino acids are favoured towards the exterior of the proteins. It is the combination of these forces which give the protein the native active state (Lehninger et al. 2005). 1.2 Protein folding In order for the protein to be in the active state it is important that protein is in 3D folded state otherwise the protein is in inactive state and it can lead to protein aggregation and misfolding diseases. The energy funnel model of the protein folding gives a nice description of protein folding and as can be seen below (Figure1) with local energy and global energy minima in the energy funnel. Misfolded or aggregated proteins are in the local energy minima while in the global energy minima the protein is in the folded or native state (Bartlett and Radford 2009), (Hartl and Hayer-Hartl 2009).
4

Local energy minima
Local energy minima
Global energy minima
Figure1 Energy funnel of protein folding In the energy funnel model protein is in different states when it gets folded to the native form. There are regions of local energy minima and global energy minima. In local energy minima the intermediates in the protein folding pathway and aggregated proteins which eventually would be removed from the cells are present. While in the global energy minima the native folded conformation of the protein is present. The unfolded proteins could be considered to be in the higher energy state and folded protein in the lower energy state. (Adapted from (Bartlett and Radford 2009) with permission from Nature Publishing Group). Various denaturing agents like 8 M urea and 6 M guanidine hydrochloride efficiently disrupt the non covalent bond interactions of proteins thus unfolding and rendering them almost inactive (Stryer, 1995).
5

CN3H6 Guanidinium chloride Generally it has been shown that at a critical concentration of denaturant there occurs sharp transition from folded to unfolded state thus more protein molecules go into denatured state and thus propensity of protein molecules to go into denatured state increase (Figure2). When denaturing agent is removed or is diluted to an extent that it no longer has any denaturing effect the protein refolds back to native state (Stryer, 1995). Although the number of molecules refolded are considerably lower as compared to the total number of protein molecules denatured.
100
piyeiyehglh
Prot
ein
unfo
lded
%
0
Denaturant (conc)
Figure2 Effect of denaturant on percentage of protein denatured At a particular denaturant concentration the number of protein molecules to go into the unfolded state increases dramatically and then they remain at saturation level of unfolded state. Here even with increase in the denaturant concentration the amount of unfolded protein molecules remain the same. (Adapted from (Berg et al. 2002)) Proteins generally follow a pattern of cooperative folding pattern and so some partly folded proteins help in folding of other protein molecules which have not folded yet. 1.3 Central dogma, Protein synthesis and ribosomes The essential tenet of central dogma is the connection between the DNA, RNA and protein (Figure3)
6

Figure3 Information flow in maintenance of life Information from DNA is transcribed into the transient messengers (mRNA) which is in turn translated into protein through Ribosomes & tRNA. Ribosomes are thus central machinery of growth in a cell that converts hereditary information into proteins. Prokaryotic ribosomes are composed of 50S subunit and 30S subunit and the whole complex including 50S and 30S subunits is called 70S (Figure4). These subunits are composed of protein and an RNA part. 30S subunit is composed of 21 different proteins (S1 through S21) and a 16S RNA molecule (Berg et al. 2002). The 50S subunit is composed of 34 different proteins (L1 through L34), and two RNA molecules 23S and 5S RNA. The essential function of the ribosome is to provide a platform on to which protein synthesis occurs. It is composed of A, P and E sites, where A is the acceptor site, P is peptide formation site and E is the exit site. In A site new amino acid loaded on to corresponding tRNA is brought to the ribosome, while at P site dipeptide is formed between new amino acid brought and existing peptide chain and E site is the one where the used tRNA (empty) goes away from the ribosome (Berg et al. 2002).
Figure4 Complex between 50S and 30S ribosome subunits Ribosomes are composed of 50S and 30S subunits (blue is 30S and red is 50S) which join to form 70S ribosome complex in prokaryotic organisms. Adapted from http://en.wikipedia.org/wiki/Ribosome Eukaryotic ribosomes are composed of 60S and 40S subunits and the whole complex is called 80S. Both subunits are composed of protein and RNA parts. 60S subunit consist of 49 proteins and it contains 28S, 5S and 5.8S RNA. While the small subunit 40S is composed of 33 proteins and 18S RNA (Taylor et al. 2009). 1.4 Role of Ribosome in protein folding Originally proteins were believed to be self folding as demonstrated by Christian Anfinsen (Anfinsen, 1973). Subsequently a class of proteins called chaperones or heat shock proteins (HSP) were identified as the post translational regulator of protein folding (Hartl and Hayer-Hartl 2009). Recently ribosomes themselves have been assigned a role in initiating the folding
7

process during synthesis of the nascent polypeptide chain (Das et al. 2008). It has also been experimentally shown that chaperones are also involved in de novo folding, protein transport and proteolytic degradation inside the cellular system. Many of these chaperones are also called heat shock proteins and as the name suggests they are up regulated in the conditions of cellular stress. In prokaryotes this system consists of trigger factor and Hsp70, Hsp40 class of proteins and is involved in folding the nascent chain of protein synthesized on the ribosome. In eukaryotes especially in Saccharomyces cerevisiae this folding is carried out by Hsp70, Hsp40 analogue Ssb/Ssz/Zuotin chaperone triad and Nascent chain associated complex (NAC) (Kramer et al. 2009). It has been recently been suggested that defects in these factors lead to cellular abnormalities including slower growth, increased sensitivity to antibiotics and shock. It is well known that ribosomes play very important role in protein synthesis but that they also have role in protein folding is a recent development. It is also evident from some reports that ribosomes can help in folding of several unrelated proteins (Das, et al. 1996). This folding is not to the same extent in case of all proteins studied but it is significant enough to show that ribosomes do have important role in assisting protein folding (Das, et al. 1996). It has also been shown that not only full 70S ribosomes, but that its component 50S subunit also helps in protein folding ( Das et al. 1996). Domain V in 50S subunit located close to peptide transfer centre itself play significant role in protein folding and many conserved residues have been identified in this region, mutation in them can cause significant loss in assisted folding ( Sanyal et al unpublished), (Samanta et al. 2008). 1.5 Protein misfolding and prion disease A large number of degenerative diseases are now attributed to protein misfolding and points towards defects in the chaperone and protein quality control mechanism inside the cell. These include most noticeably Alzheimers, Parkinsons, Huntingtons and prion diseases (Hartl and Hayer-Hartl 2009). In all these diseases the toxic principle is generally a soluble protein particle, aggregation of which leads to disease phenotype. Prion diseases are really debilitating and fatal diseases resulting in certain death of the animal. Some of the types of prion diseases include bovine spongiform encephalopathy (BSE) in cattle, chronic wasting disease (CWD) in elk, mule and deer, Transmissible mink encephalopathy (TME) in mink. In humans it causes Cruetzfeldt-Jakob disease (CJD), Gerstmann-Sträussler-Scheinker (GSS) syndrome etc (Gilch, et al. 2008). In the case of humans sporadic CJD occurs with pretty much the same with frequency of 1: 1,000,000 per year around the world. Familial forms of the disease are still unknown as the onset and progression of these diseases are difficult to predict. These diseases are known to be originating from prion proteins which are present in two forms PrPc and PrPsc
inside the cell. Misfolding of PrPc to PrPsc leads to prion diseases (Aguzzi and Calella 2009). They were first studied in yeasts which contain an isoform of prion proteins which might be considered complementary to mammalian prions. They are called [URE3] and [PSI+] which have the normal analogue in yeast as Ure2p and Sup35p respectively. Overproduction of these normal analogues in yeast leads to increase in [URE3] and [PSI+] (Wickner et al. 2007). There are different lines of treatment for prion diseases and each is aimed at different stages of the maturation and conversion process of prion proteins. Some of the antiprion drugs and their potential mechanism are given in the table below. Table1 Antiprion drugs being studied Examples of prion drugs which are being currently studied and their possible mechanism of action are given in the table. (Adapted from (Gilch et al. 2008) with permission of informa healthcare)
8

Name of drug Mode of action Reference Congo red Decreases cell surface PrPc,
binds and over stabilizes PrPsc
(Ingrosso et al. 1995)
Dextran sulphate 500 Decrease of cell surface PrPc (Ladogana et al. 1992) Suramin Aggregation and remixing of
PrPc (Ladogana et al. 1992)
Cyclic tetrapyrroles Alteration of PrPsc (Priola et al. 2000) Imitinib Induction of PrPsc clearance,
induction of cellular autophagy
(Ertmer et al. 2004)
si RNA Knock down of PrPc
expression (Pfeifer et al. 2006)
Phosphorothiata oligonucleotides
Inhibition of PrPsc in cell culture, more probability to ProteinaseK digestion
(Karpuj et al. 2007)
Monoclonal Antibody Targeting of PrPc and to PrPsc (Beringue et al. 2004) 1.6 Ribosome as a target for antiprion drugs Most of the antiprion drugs studied act directly on the PrPc to PrPsc conversion or in the stabilization or removal of PrPc or PrPsc. But the lab of Marc Blondel in INCERM France have from the yeast based assay identified many potential drugs acting against mammalian counterparts of prion proteins [URE3] and [PSI+] in yeast (Bach et al. 2006) and deeper analysis of some of the promising drugs have shown that they do not act directly on the prion protein but rather on a internal modulator which is associated with the conversion of Ure2p and Sup35p to [URE3] and [PSI+] in yeast. The work done in Sanyal lab has shown two of these drugs Psi207 (6-aminophenanthridine) and psi114 (Guanabenz) specifically act on the 70S ribosome and prevent ribosome assisted protein folding (Tribouillard-Tanvier et al. 2008). Furthermore it has been shown that Psi207 (6AP) and Psi114 (GA) bind to 23S rRNA in 50S ribosome subunit (Das et al. 1996). Taken together our hypothesis is that after protein synthesis unfolded protein goes through an intermediate state and then it can either get disaggregated or misfolded. The other possibility could be that unfolded protein with the help of chaperones including ribosomes gets folded into native conformation (Figure5).
9

Sanyal lab hypothesis
Ribosome assisted misfolding
Ribosome assisted folding
Figure5 Protein misfolding and folding have similar pathways in different neurodegenerative diseases Protein unfolding and folding generally involve similar pathways in case of different proteins which either remain unfolded or form disorganized aggregates or they form intermediates and then get converted into disorganized aggregates leading to various misfolding diseases including prion. The other possibility is that polypeptide chain once synthesized goes through unfolded then to intermediate and finally form native protein. The Sanyal lab hypothesis is that ribosomes act a chaperone and somehow can assist in folding or misfolding in the case of prion diseases. (Adapted from (Dobson 2003) with permission from Nature Publishing Group) AIMS My goal in this project was to study 70S and 80S ribosome assisted protein folding and later studies the effect of antiprion drugs Psi207 and Psi114 on ribosome assisted protein folding. Specifically I used human carbonic anhydrase I (HCA I) as model protein for studying protein unfolding and refolding. HCA I was used in these experiments because it is an extremely efficient enzyme with rate of reaction for carbon dioxide reaching 106 at pH 9 and 25˚C.The Kcat/Km for HCA I reaches around 5x107 M-1 S-1 (Lehtonen et al. 2004). It is also one of the most stable enzymes and has activity in different conditions of temperature and pH. HCA I is a monomer with 261 amino acids making it ideal for studying enzyme kinetics. The unfolding and refolding experiments were carried out by strategy given below (Figure6).
10

Figure6 Denaturation and renaturation assay strategy Proteins are denatured by chemical denaturant like 6 M guanidine chloride and then they are refolded by dilution to some refolding buffer. When no folding modulators like 50S ribosome subunit etc are added and protein is allowed to fold on its own then it is called self folding. In case of assisted protein folding, modulators like 70S, 80S, 50S ribosome subunit, 23S rRNA or domain V is added to refolding buffer. The figure adapted with permission of Ass. Prof. Suparna Sanyal (Tribouillard-Tanvier, Dos Reis et al. 2008). In my experiments I have tried to study the effects of antiprion drugs on both 70S and 80S assisted protein folding.
11

2. Aims and objectives 2.1 Human carbonic anhydrase
• Purification of Human carbonic anhydrase I (HCA I) • Standardization of conditions for Human carbonic anhydrase I kinetics • Standardization of conditions for Human carbonic anhydrase I unfolding and refolding
assay 2.2 Ribosomes
• Purification of 70S, 50S, 80S ribosomes and total rRNA from 80S ribosomes • Characterization of their activity in refolding assay • Determination of concentration profile of folding modulators
2.3 Antiprion drugs • Effect of antiprion drugs on 70S, 80S assisted folding • Concentration profile of antiprion drugs on 70S and 80S ribosome assisted folding • Time-course of antiprion drugs on 70S and 80S ribosome assisted folding
3. Results 3.1 Overexpression and purification of Human carbonic anhydrase I 3.1.1 Overexpression of Human carbonic anhydrase I Two batches of HCA I (will be called HCA from now) were purified, one was with zinc and the other was without zinc in the bacterial growth media. HCA was cloned in pET3a vector and transferred to BL21 (DE3) E.coli strain. The induction of the HCA (-Zn) (this is HCA which was grown without presence of Zn in the media) was done in the presence of 1mM IPTG (Figure7A). The induction of HCA (+Zn) was done in presence of 0.4 mM IPTG and 0.5 mM zinc sulphate (Figure7B). In both cases protein expression was good. A 1 2 3 4 5 6 7 B 1 2 3 4 5 6 7 8
29K
Figure7 SDS PAGE showing pre and post induction expression of HCA (+Zn) and HCA (-Zn). A: HCA (-Zn) Marker (lane 1), pre induction (lanes 2, 4, 6) and post induction with IPTG (lanes 3, 5, 7). B: HCA (+Zn) Marker (lane 8), pre induction (lanes 2, 3, 5, 6) and post induction with IPTG (lanes 1, 4, 7). 29 K in the figure shows band corresponding to HCA.
12
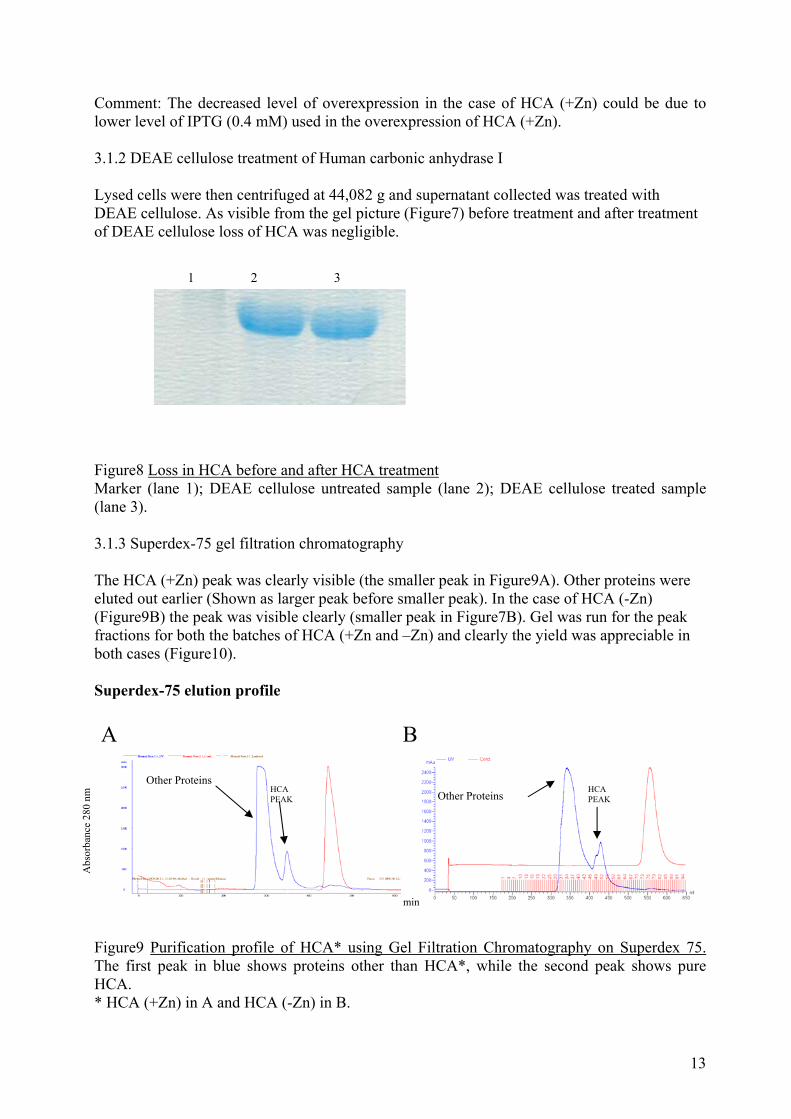
Comment: The decreased level of overexpression in the case of HCA (+Zn) could be due to lower level of IPTG (0.4 mM) used in the overexpression of HCA (+Zn). 3.1.2 DEAE cellulose treatment of Human carbonic anhydrase I Lysed cells were then centrifuged at 44,082 g and supernatant collected was treated with DEAE cellulose. As visible from the gel picture (Figure7) before treatment and after treatment of DEAE cellulose loss of HCA was negligible. 1 2 3 Figure8 Loss in HCA before and after HCA treatment Marker (lane 1); DEAE cellulose untreated sample (lane 2); DEAE cellulose treated sample (lane 3). 3.1.3 Superdex-75 gel filtration chromatography The HCA (+Zn) peak was clearly visible (the smaller peak in Figure9A). Other proteins were eluted out earlier (Shown as larger peak before smaller peak). In the case of HCA (-Zn) (Figure9B) the peak was visible clearly (smaller peak in Figure7B). Gel was run for the peak fractions for both the batches of HCA (+Zn and –Zn) and clearly the yield was appreciable in both cases (Figure10). Superdex-75 elution profile
A B
Other Proteins HCA PEAK
HCA PEAK
Abs
orba
nce
280
nm
Other Proteins
min
Figure9 Purification profile of HCA* using Gel Filtration Chromatography on Superdex 75. The first peak in blue shows proteins other than HCA*, while the second peak shows pure HCA. * HCA (+Zn) in A and HCA (-Zn) in B.
13
SDS-PAGE analysis of purified HCA
B A 1 2 3 4 5 6 7 8 1 2 3 4 5
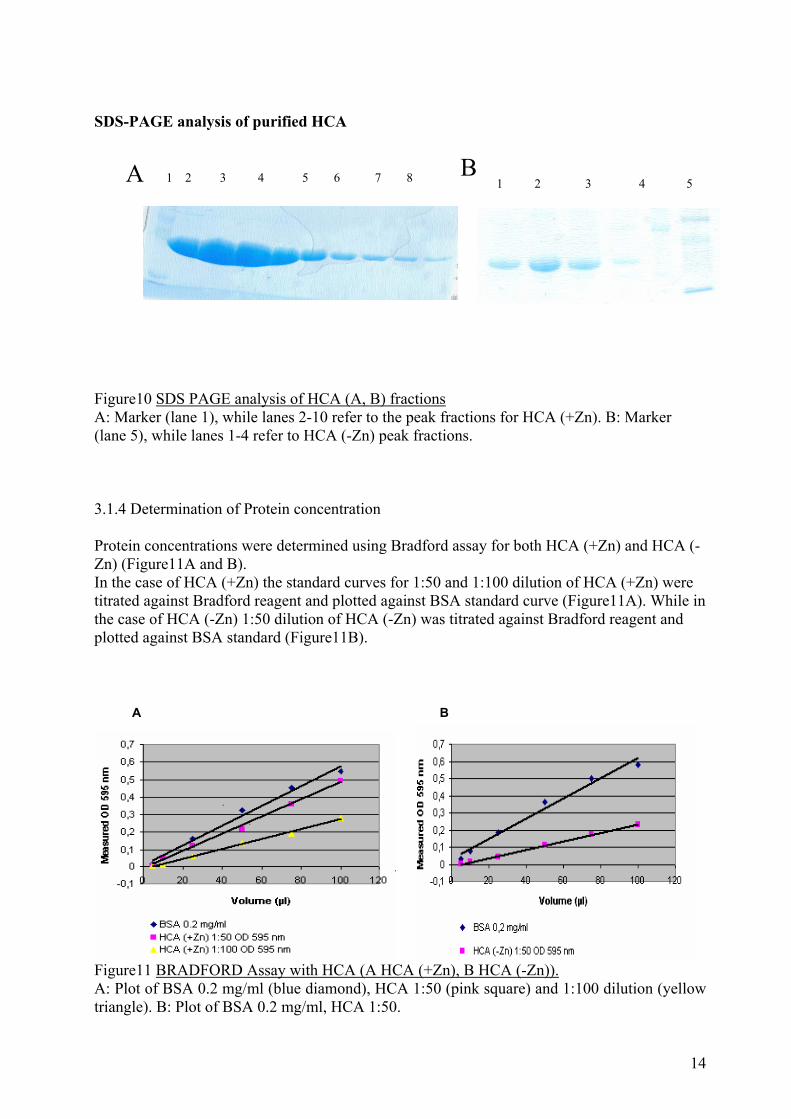
Figure10 SDS PAGE analysis of HCA (A, B) fractions A: Marker (lane 1), while lanes 2-10 refer to the peak fractions for HCA (+Zn). B: Marker (lane 5), while lanes 1-4 refer to HCA (-Zn) peak fractions. 3.1.4 Determination of Protein concentration Protein concentrations were determined using Bradford assay for both HCA (+Zn) and HCA (-Zn) (Figure11A and B). In the case of HCA (+Zn) the standard curves for 1:50 and 1:100 dilution of HCA (+Zn) were titrated against Bradford reagent and plotted against BSA standard curve (Figure11A). While in the case of HCA (-Zn) 1:50 dilution of HCA (-Zn) was titrated against Bradford reagent and plotted against BSA standard (Figure11B).
A B
Figure11 BRADFORD Assay with HCA (A HCA (+Zn), B HCA (-Zn)). A: Plot of BSA 0.2 mg/ml (blue diamond), HCA 1:50 (pink square) and 1:100 dilution (yellow triangle). B: Plot of BSA 0.2 mg/ml, HCA 1:50.
14

Data for Bradford assay is given in table1 below Table2 Data and calculation for Bradford assay Bradford calculation and data generated for HCA (+Zn) and HCA (-Zn) are shown in the table. Protein BSA HCA (+Zn) 1:50 HCA (+Zn) 1:100 HCA (-Zn) 1:50 Mol weight (Da) 29114 29800 29800 29800 Concentration (mg/ml)
0.2
Slope 0.006 0.005 0.003 0.0024 Estimated concentration (mg/ml)
0.17 0.1 0.08
Dilution factor 50 100 50 Concentration of Protein (mg/ml)
8.71 10.12 4.11
Average HCA (+Zn) concentration [µM] = 316.0 Average HCA (-Zn) concentration [µM] =138.0 3.2 Effect of substrate on Human carbonic anhydrase I activity 3.2.1 Kinetics of Human carbonic anhydrase I activity 500 nM of HCA (+Zn), HCA (-Zn) was titrated against increasing concentration of PNPA (from 0 to 10 mM) (Figure12A). From these curves concentration of PNPA to be used for the assay was standardized. In both cases BCA acted as control (Figure12B).
A B
Figure12 Effect of substrate concentration on HCA activity A: Effect of varying substrate concentration (PNPA) on HCA (+Zn) and HCA (-Zn) activity. B: Effect of varying concentration of PNPA on BCA activity. From these graphs kinetic parameters for the enzymes were calculated.
15

From the above graphs (Figure12) the following Km and Vmax values were obtained (Table3). Table3 Km and Vmax for the enzymes Table shows the obtained Km and Vmax values for HCA (+Zn), HCA (-Zn) and BCA.
Km Vmax
HCA (+Zn) 3.8 mM 0.07 mM/min
HCA (-Zn) 2.6 mM 0.04 mM/min
BCA 3.2 mM 0.2 mM/min
3.2.3 Effect of solvent for paranitrophenyl acetate on Human carbonic anhydrase I activity PNPA was dissolved in acetonitrile and 95% ethanol and the activity of HCA with increasing concentration of PNPA was plotted for both HCA (+Zn) and HCA (-Zn) (Figure12B) and was compared to BCA activity profile for both solvents (Figure12A).
B A
Figure13 Effect of solvent on BCA and HCA (-Zn) activity. A: HCA (-Zn) activity, when PNPA was dissolved in acetonitrile and 95% ethanol. B: BCA activity, when PNPA was dissolved in acetonitrile and 95% ethanol. From the above graphs following kinetic parameters were calculated (Table4) Table4 Km and Vmax values for HCA (-Zn) and BCA for dissolving PNPA in different solvents Shown are values for Km and Vmax for HCA (-Zn) and BCA on dissolving PNPA in 95% ethanol and acetonitrile. K m V max HCA (-Zn) acetonitrile 2.8 mM 0.1 mM/min HCA (-Zn) ethanol 2.6 mM 0.04 mM/min BCA acetonitrile 3.2 mM 0.5 mM/min BCA ethanol 3.2 mM 0.2 mM/min
16

3.2.4 Effect of Zn in refolding buffer on Human carbonic anhydrase I activity Effect of Zn on the HCA activity was studied, and as can be seen from Figure14 in the presence of Zn the HCA (+Zn) showed higher activity till 0.1 mM of Zn when PNPA was dissolved either in acetonitrile or in 95% ethanol. While in the case of HCA (-Zn) the optimum activity of HCA (-Zn) was obtained at 0.1 mM Zn in the case when PNPA was dissolved in acetonitrile (Figure14). The optimum activity of HCA (-Zn) was found to be at 0.05 mM Zn when PNPA was dissolved in 95% ethanol. Also as can be seen from the table below that their was significant increase in Vmax for both HCA (+Zn) and HCA (-Zn) when optimum Zn concentration was used for HCA activity as compared to when no Zn was added in the buffer used for dissolving HCA (Figure13).
Figure14 Effect of Zn and solvent in refolding buffer and PNPA respectively on HCA activity. Effect of Zn, and dissolving PNPA in different solvents on HCA (+Zn), HCA (-Zn) activity. From the above curve following kinetic parameters were obtained (Table5) Table5 Zn and solvent effect on HCA (+Zn) activity HCA (+Zn) and HCA (-Zn) activity was assayed in the buffer containing Zn and different solvents for dissolving PNPA. The Km and Vmax values for the same are given in the table. Zn in refolding buffer V max HCA (+Zn),acetonitrile 0.11 mM 0.11 mM/min HCA (+Zn),ethanol 0.1 mM 0.14 mM/min HCA (-Zn),acetonitrile 0.1 mM 0.09 mM/min HCA (-Zn),ethanol 0.05 mM 0.09 mM/min 3.3 Human carbonic anhydrase I unfolding and refolding
17

3.3.1 Effect of EDTA on Human carbonic anhydrase I unfolding EDTA had significant impact on unfolding of HCA (+Zn) (Figure15). As can be seen from the plot (Figure15) without having any Zn in the refolding buffer the HCA (+Zn) refolding activity was around 35% with respect to native enzyme activity. But with increase in the concentration of the Zn concentration till 20 µM, HCA (+Zn) refolding activity fell from 35% to around 15%, after which their was no decrease in the HCA (+Zn) activity even with a Zn concentration of 60 µM.
Figure15 Effect of EDTA on HCA unfolding With the increase in concentration EDTA in the denaturation mix there is significant decrease in HCA refolded after denaturation mix is diluted in the refolding buffer. 3.3.2 Purification of 70S ribosomes 70S ribosomes were purified from JE28 strain of E.coli. Ni Nt column chromatography was used to purify the ribosomes because of the presence of His tag on L12 protein of 50S ribosome subunit (Ederth, Mandava et al. 2009). Using this strategy 70S ribosomes were bound on Ni column and were then eluted using gradient elution using 50% length 500 mM Imidazole (The elution started from 150 mM till 250 mM Imidazole) (Figure16).
Figure16 Ni Nt purification profile for 70S ribosomes 70S ribosomes were purified using 50% length 500 mM Imidazole gradient elution and 70S ribosome peak was observed at 250 mM Imidazole (larger peak). The blue line depicts UV absorbance, green line shows Imidazole concentration and red line in the curve shows conductance.
18
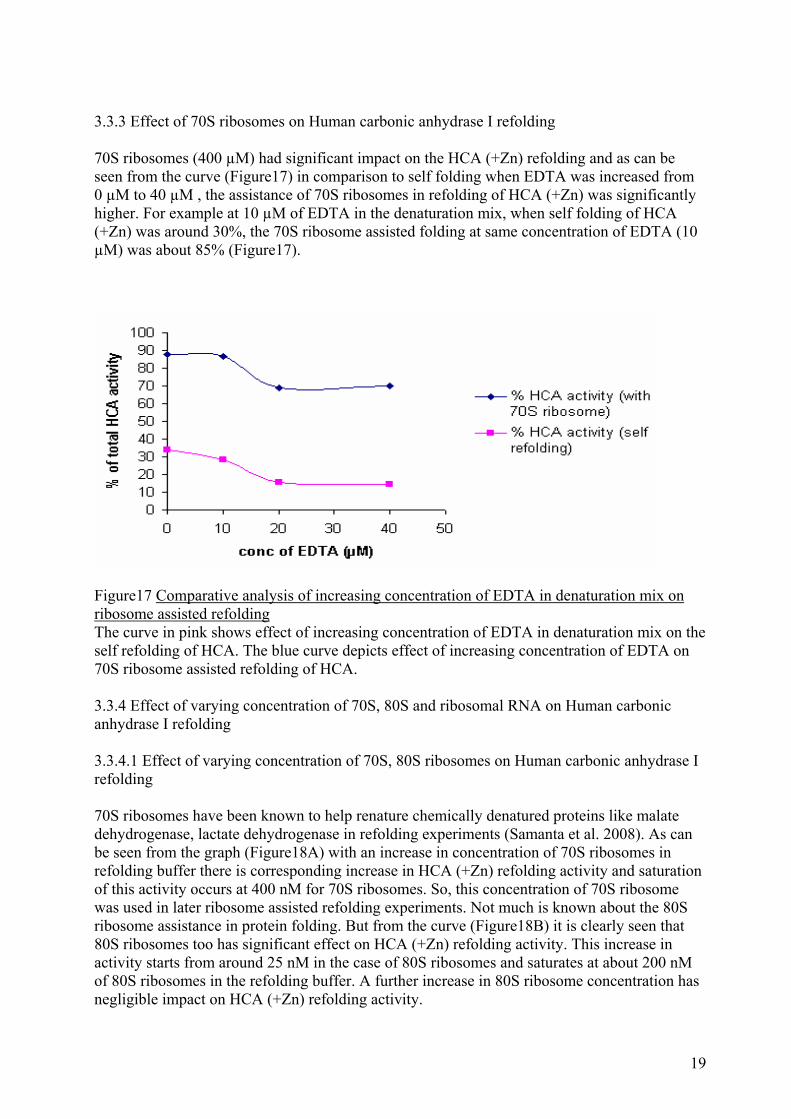
3.3.3 Effect of 70S ribosomes on Human carbonic anhydrase I refolding 70S ribosomes (400 µM) had significant impact on the HCA (+Zn) refolding and as can be seen from the curve (Figure17) in comparison to self folding when EDTA was increased from 0 µM to 40 µM , the assistance of 70S ribosomes in refolding of HCA (+Zn) was significantly higher. For example at 10 µM of EDTA in the denaturation mix, when self folding of HCA (+Zn) was around 30%, the 70S ribosome assisted folding at same concentration of EDTA (10 µM) was about 85% (Figure17).
Figure17 Comparative analysis of increasing concentration of EDTA in denaturation mix on ribosome assisted refolding The curve in pink shows effect of increasing concentration of EDTA in denaturation mix on the self refolding of HCA. The blue curve depicts effect of increasing concentration of EDTA on 70S ribosome assisted refolding of HCA. 3.3.4 Effect of varying concentration of 70S, 80S and ribosomal RNA on Human carbonic anhydrase I refolding 3.3.4.1 Effect of varying concentration of 70S, 80S ribosomes on Human carbonic anhydrase I refolding 70S ribosomes have been known to help renature chemically denatured proteins like malate dehydrogenase, lactate dehydrogenase in refolding experiments (Samanta et al. 2008). As can be seen from the graph (Figure18A) with an increase in concentration of 70S ribosomes in refolding buffer there is corresponding increase in HCA (+Zn) refolding activity and saturation of this activity occurs at 400 nM for 70S ribosomes. So, this concentration of 70S ribosome was used in later ribosome assisted refolding experiments. Not much is known about the 80S ribosome assistance in protein folding. But from the curve (Figure18B) it is clearly seen that 80S ribosomes too has significant effect on HCA (+Zn) refolding activity. This increase in activity starts from around 25 nM in the case of 80S ribosomes and saturates at about 200 nM of 80S ribosomes in the refolding buffer. A further increase in 80S ribosome concentration has negligible impact on HCA (+Zn) refolding activity.
19
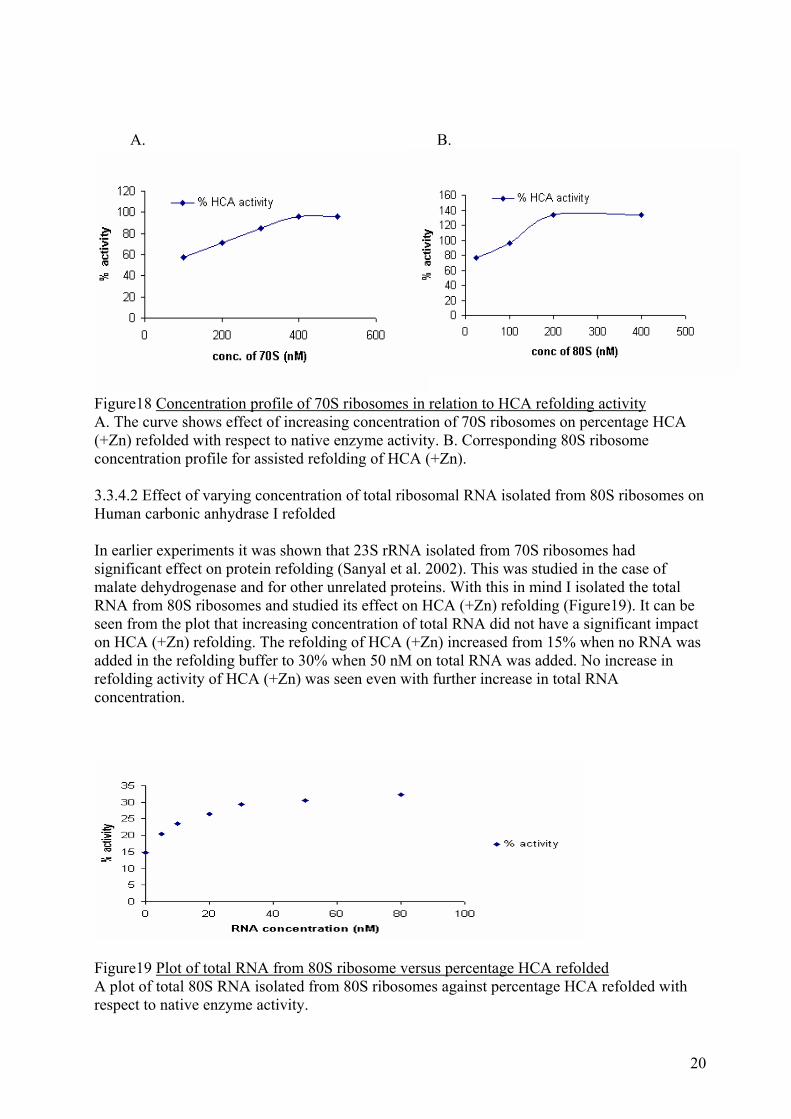
A. B.
Figure18 Concentration profile of 70S ribosomes in relation to HCA refolding activity A. The curve shows effect of increasing concentration of 70S ribosomes on percentage HCA (+Zn) refolded with respect to native enzyme activity. B. Corresponding 80S ribosome concentration profile for assisted refolding of HCA (+Zn). 3.3.4.2 Effect of varying concentration of total ribosomal RNA isolated from 80S ribosomes on Human carbonic anhydrase I refolded In earlier experiments it was shown that 23S rRNA isolated from 70S ribosomes had significant effect on protein refolding (Sanyal et al. 2002). This was studied in the case of malate dehydrogenase and for other unrelated proteins. With this in mind I isolated the total RNA from 80S ribosomes and studied its effect on HCA (+Zn) refolding (Figure19). It can be seen from the plot that increasing concentration of total RNA did not have a significant impact on HCA (+Zn) refolding. The refolding of HCA (+Zn) increased from 15% when no RNA was added in the refolding buffer to 30% when 50 nM on total RNA was added. No increase in refolding activity of HCA (+Zn) was seen even with further increase in total RNA concentration.
Figure19 Plot of total RNA from 80S ribosome versus percentage HCA refolded A plot of total 80S RNA isolated from 80S ribosomes against percentage HCA refolded with respect to native enzyme activity.
20
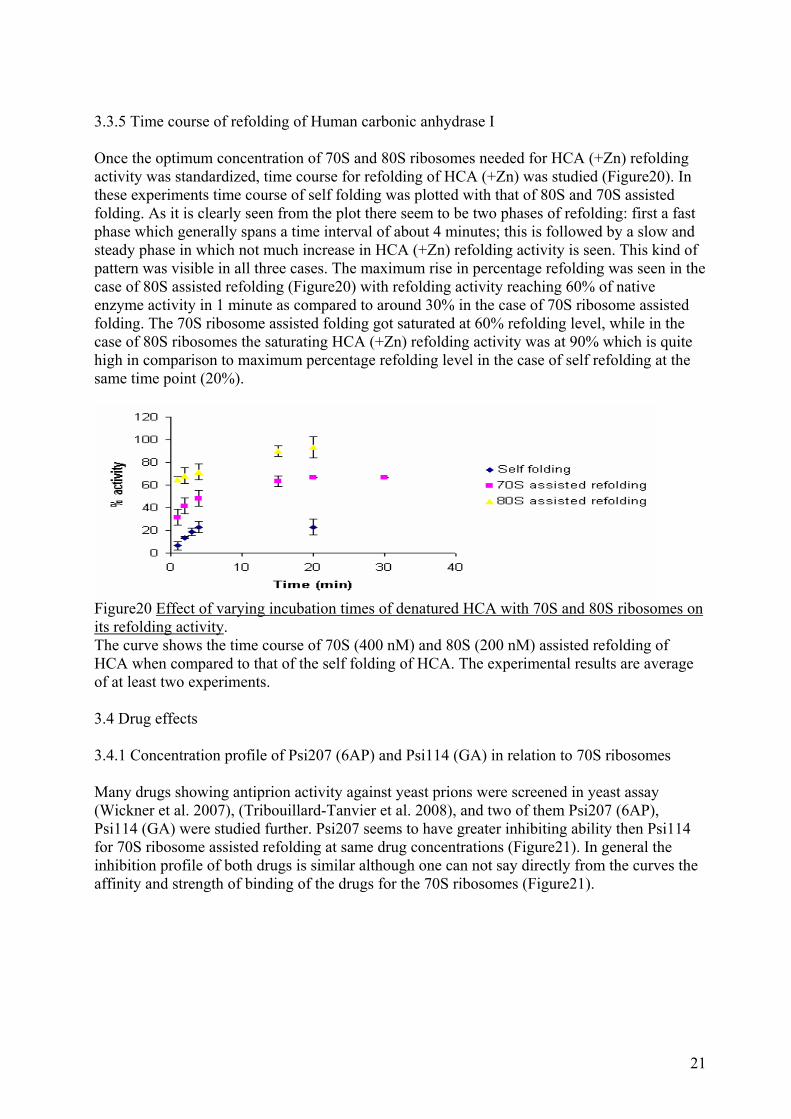
3.3.5 Time course of refolding of Human carbonic anhydrase I Once the optimum concentration of 70S and 80S ribosomes needed for HCA (+Zn) refolding activity was standardized, time course for refolding of HCA (+Zn) was studied (Figure20). In these experiments time course of self folding was plotted with that of 80S and 70S assisted folding. As it is clearly seen from the plot there seem to be two phases of refolding: first a fast phase which generally spans a time interval of about 4 minutes; this is followed by a slow and steady phase in which not much increase in HCA (+Zn) refolding activity is seen. This kind of pattern was visible in all three cases. The maximum rise in percentage refolding was seen in the case of 80S assisted refolding (Figure20) with refolding activity reaching 60% of native enzyme activity in 1 minute as compared to around 30% in the case of 70S ribosome assisted folding. The 70S ribosome assisted folding got saturated at 60% refolding level, while in the case of 80S ribosomes the saturating HCA (+Zn) refolding activity was at 90% which is quite high in comparison to maximum percentage refolding level in the case of self refolding at the same time point (20%).
Figure20 Effect of varying incubation times of denatured HCA with 70S and 80S ribosomes on its refolding activity. The curve shows the time course of 70S (400 nM) and 80S (200 nM) assisted refolding of HCA when compared to that of the self folding of HCA. The experimental results are average of at least two experiments. 3.4 Drug effects 3.4.1 Concentration profile of Psi207 (6AP) and Psi114 (GA) in relation to 70S ribosomes Many drugs showing antiprion activity against yeast prions were screened in yeast assay (Wickner et al. 2007), (Tribouillard-Tanvier et al. 2008), and two of them Psi207 (6AP), Psi114 (GA) were studied further. Psi207 seems to have greater inhibiting ability then Psi114 for 70S ribosome assisted refolding at same drug concentrations (Figure21). In general the inhibition profile of both drugs is similar although one can not say directly from the curves the affinity and strength of binding of the drugs for the 70S ribosomes (Figure21).
21

Figure21 Concentration profile of Psi207 (6AP) and Psi114 (GA) The graph shows the relation between the increasing antiprion drugs concentrations on the percentage fall in 70S ribosome (400 nM) assisted refolding of the HCA. 3.4.2.1 Concentration profile of Psi 207 (6AP) and Psi 114 (GA) in relation to 80S ribosomes Varying the concentration of Psi114 and Psi207 had dramatic effect on the refolding activity of HCA (+Zn). The activity was highest when 80S ribosome was used in the refolding of HCA (+Zn). (50% in the case of Psi114 and 75% in the case of Psi207) (Figure22). A greater decrease in the refolding activity was seen in the case of Psi114 where at 2 µM drug concentration percentage decrease in the refolding activity was around 20% (Figure22A) while in the case of Psi 207 reduction in HCA (+Zn) refolding activity was about 10% (Figure22B). For both drugs the final fall in activity was around 30% (Figure22), but in the case of Psi 207 this fall in HCA (+Zn) refolding activity was more gradual (Figure22B), while in the case of Psi114 after significant fall in HCA (+Zn) refolding activity at 2 µM their was more gradual decrease in refolding activity. HCA (+Zn) refolding activity fall was gradual right from the beginning in the case of Psi 207 (Figure22B).
A B
22

Figure22 Concentration profile of Psi 207 (6AP) A. Percentage fall in 80S ribosome (200 nM) assisted refolding of HCA is plotted with respect to the increasing concentration of Psi 114 (6AP). B. Decrease in percentage refolding activity of HCA (+Zn) is plotted against concentration of Psi 207. In both cases 80S assisted HCA refolding acted as positive control while self refolding acted as negative control. 3.4.3 Time course of refolding of Human carbonic anhydrase I in the presence of increasing concentration of Psi 207 and Psi114. Time course for varying concentration of Psi207 and Psi114 was plotted and as can be seen from the curves there is a definate reduction in HCA (+Zn) refolding activity with increasing concentration of Psi207 (Figure23A) and Psi114 (Figure23B).
A B
Figure23 Overlay of the time courses of varying concentrations of the drug Psi 207 (6AP) and Psi114 (GA) A. An overlay plot of the drug Psi 207 (6AP) showing time course of refolding of HCA (+Zn) with increasing concentration of Psi 207. B. Same plot for the drug Psi114 4. Discussion HCA (+Zn) and HCA (-Zn) activity Protein folding is a fundamental phenomenon since activities of all proteins depend on their correct 3D structure. In many disease processes protein is not folded properly and this leads to misfolded or aggregated forms of protein that is either non functional or toxic to the organism and cause lethality in cells and diseases in animals & humans (Dobson 2003). Prion diseases have been shown to be a protein misfolding problem where the constitutive form of the prion protein PrPc get converted into PrPsc to be scrapie or toxic form (Tribouillard-Tanvier et al. 2008) and the ribosomes are found involved in this process. In my studies I have used human carbonic anhydrase I (HCA I) to show the renaturation of the protein from the denatured state in the presence and absence of ribosomes. Two antiprion drugs (Psi 207 and Psi 114) block this ribosome mediated protein folding assistance. I purified, characterized and quantified the human carbonic anhydrase with and without Zn2+ in the culture media and studied their kinetics of refolding & recovery from denatured state (Figure12, 13, 14). As can be seen from Figure10A curves HCA (+Zn) had much higher activity in the titration assay with PNPA as compared to that of HCA (-Zn); this was also verified from comparative Km and Vmax values for each of the HCA (Table1). During the titration assay the PNPA which was used as the substrate for the HCA activity assay did not dissolve properly in 95% ethanol and used to get
23

precipitated. But from the review of literature I found that PNPA gave very good activity and was much more stable for storage in acetonitrile than in 95% ethanol (Pocker and Stone 1968) and as can be seen from Figure12 the activity of both HCA (+Zn) and HCA (-Zn) improved significantly and also the precipitation problem of PNPA was solved. In different studies it has been shown that metal ions at specific concentrations enhance the activity of enzymes and this is especially true for the metal ions which act as cofactor for the enzyme (Yazgan and Henkens 1972), (Coleman 1984). With this fact in mind I standardized the Zn concentration needed for the activity assay of HCA (Figure14) and curiously the activity of HCA (+Zn) and HCA (-Zn) improved significantly. But concentration higher then 0.1 mM of Zn seemed to have inhibitory effect on the HCA activity (Figure14). This could be due to some kind of oxidizing effect of Zn on the thiol containing groups in the HCA (2 methionine) which might have deleterious effect or toxic effect on the HCA activity as no reducing agent was added in the refolding buffer in which the HCA was dissolved, as some papers had suggested that adding reducing agent in the HCA titration assay might have inhibitory effect on HCA activity (Innocenti et al. 2008).The other reason might be that excess of zinc ions might result in competition for the para nitro phenolate ion thereby preventing the forward reaction of breakdown of para nitro phenyl acetate to para nitro phenol and acetate. This fact was seen in all the cases of using HCA (HCA (+Zn) and HCA (-Zn)) (Figure14). But it is still preliminary to suggest these factors or their might be totally different fact itself to explain these results and since my project was not directly associated with this metal ion effect on HCA activity I did not delve further into it. HCA (+Zn) unfolding and refolding Refolding activity of HCA (+Zn) was pretty high (around 35%) when HCA (+Zn) was unfolded in absence of EDTA, but with the addition of EDTA (to 20 µM) the refolding percentage fell to around 15%. In the case of antiprion inhibitory assays as well which I did later, at 2 µM of Psi114 the fall in 80S ribosome assisted HCA (+Zn) refolding level was around 35%. So, in order to increase the denaturation of HCA (+Zn) EDTA was added in the denaturation mix as EDTA has chelating effect on metal ion (here Zn) which prevents proper folding of HCA (+Zn) and therefore less number of HCA (+Zn) molecules would go into productive folding pathway. Though the 70S ribosome assisted HCA (+Zn) folding was not affected by concentration of EDTA in the denaturation mix and it seemed to get diluted in the refolding buffer and loose its chelating effect which gives reason to believe that EDTA chelation was reversible in protein refolding experiments (Figure15). Effect of Antiprion drugs In the following experiments were the Psi114 and Psi207 activity in inhibiting 70S ribosome assisted HCA (+Zn) refolding activity (Figure20). From the curves it is clear that Psi207 had higher inhibitory effect on 70S ribosome assisted HCA (+Zn) refolding activity at same concentrations of Psi114. Having shown the inhibitory effect of the drugs on the prokaryote 70S ribosomes, we moved in for the inhibition study of the drugs on the eukaryotic 80S ribosomes. These 80S ribosomes were isolated from Saccharomyces cerevisiae and as was done with in the case of the 70S ribosome assisted refolding a concentration profile and time course of 80S ribosomes was generated and drug inhibition study was carried out. In general it seemed that with both drugs the final percentage fall in the refolding activity was pretty much same (Figure22). But their was batch to batch discrepancy in the 80S ribosome assisted HCA (+Zn) refolding which in the Psi207 inhibition studies gave a percentage refolding of around 75% while in the case of the Psi114 it was around 50%. But overall the corresponding fall in the inhibition for both drugs was comparable when compared to the self folding level (Figure22). Finally I performed time course studies with 4 different concentrations of the drugs and in either case their seemed to be increase in inhibition by the drugs with increase in the
24

incubation time. A difficulty encountered in these inhibition studies was that it took time for the spectrophotometer to recognize the sample (around 5 seconds) and then their was time interval of mixing of the samples, so taken together the overall process of sample preparation to that of actual measurement of the sample took some time therefore their was some time variation in the studying the refolding activity at initial time intervals. Also since these experiments were done manually and individually there could be chances of pippeting error in these experiments. More sophisticated kinetic experiments and better machines like quench flow would be needed to study this phenomenon more clearly. Taken together with the data generated by me, and others in our lab and based on the published results it has been shown that both drugs Psi207 and Psi114 bind to the domain V of the ribosomes. Furthermore 70S, 80S ribosomes and their components have important role in protein folding. In order to validate this fact furthermore experiments on other proteins especially for the proteins which are easier to follow in the protein folding experiments like luciferase should be studied. But there are some unanswered questions in this story like if the ribosome helps in protein folding but in other case has role in protein misfolding and prion protein formation. This is quite mysteries to me and to study this fact one need to perform in vivo experiments especially with the prion PrPC isotype. Also related to above question is in which case ribosome assists in protein folding and in which case misfolding is favored. Finally the role of domain V is quite unclear regarding protein misfolding. For example, are their different types of ribosomes in the cell and these antiprion drugs bind specifically to those ribosomes which are associated with the misfolding of the proteins or if they (ribosomes) are the same and drugs bind to all domain V. Answering these questions would require cooperative approach of molecular biology, cell biology and pharmacology and would have great implications on improving lives of people suffering from this debilitating disease. 5. Materials and Methods 5.1 Bacteria, Plasmids & Yeast strains:- pET-3a vector was used for all Hisx6 cloning & Overexpression. All cloning & Overexpression were performed in the Escherichia coli strain BL21 (DE3). Escherichia coli JE 28 (Ederth, Mandava et al. 2009) was used to purify 70S ribosome in single step Ni- column purification, the ribosomes were Hisx6 tagged on L12 of the 50S subunit. Yeast: Saccharomyces cerevisiae MH272 3Fα strain was used for 80S ribosome as described below. 5.2 Buffers and stock solutions 5.2.1 For HCA purification
1. Wash buffer 50 mM Tris HCl (pH7.5) 100 mM Sodium chloride (Merck KGaA, Darmstadt, Germany) Water
2. Lysis buffer 50 mM Tris HCl (pH8.0) (Sigma-Aldrich, GmbH, Germany) 100 mM NaCl (Merck KGaA, Darmstadt, Germany) 0.01 mg/ml DNaseI (Sigma-Aldrich, GmbH, Germany)
25

Water
3. IPTG solution 1 mM IPTG (Apollo Scientific limited, UK)
4. Bradford reagent (1000 ml) 100 mg Coomassie Brilliant Blue G-250 in 50 ml 95% ethanol 100 ml 85% (w/v) phosphoric acid Make up the volume to 1000 ml with water Filter solution and store in dark and cold room
5. Ampicillin stock 50 mg/ml Ampicillin (Sigma-Aldrich, GmbH, Germany) Water 5.2.2 Human carbonic anhydrase unfolding and refolding
6. 8 M Guanidine Hydrochloride (1 ml) 8M Guanidine hydrochloride (Sigma-Aldrich, GmbH, Germany) Water Set the pH to 8.0. Aliquot it and store it at -20˚C.
7. 1M Magnesium Chloride hexahydrate 1M Magnesium chloride (Merck KGaA, Darmstadt, Germany) Water
8. 0.25 M PNPA 0.25 M para nitro phenyl acetate (Sigma-Aldrich, GmbH, Germany) Acetonitrile (Genzyme, Cambridge, USA)
9. Denaturation buffer 6 M Guanidine hydrochloride (Sigma-Aldrich, GmbH, Germany) 20 µM EDTA (Merck KGaA, Darmstadt, Germany) Water
10. Refolding buffer (without Zn) 10 mM Magnesium Chloride (Merck KGaA, Darmstadt, Germany) 20 mM Tris HCl (pH7.5) (Sigma-Aldrich, GmbH, Germany) 100 mM Sodium Chloride (Merck KGaA, Darmstadt, Germany) Water
11. Refolding buffer (with Zn) All chemicals and concentrations are same as 10 but additional 0.1 mM zinc sulphate (Sigma-Aldrich, GmbH, Germany) is added. 5.2.3 For 70S ribosome purification
12. lysis buffer (pH 7.6) 20 mM Tris HCl (pH7.6) (Sigma-Aldrich, GmbH, Germany) 10 mM Magnesium Chloride (Merck KGaA, Darmstadt, Germany)
26

150 mM Potassium Chloride (Merck KGaA, Darmstadt, Germany) 30 mM Ammonium Chloride (Merck KGaA, Darmstadt, Germany) 20 µM PMSF (Sigma-Aldrich, GmbH, Germany) 10 µg/ml DNase I (Sigma-Aldrich, GmbH, Germany) water
13. Wash buffer (pH7.6) 20 mM Tris HCl (pH7.6) (Sigma-Aldrich, GmbH, Germany) 10 mM Magnesium Chloride (Merck KGaA, Darmstadt, Germany) 150 mM Potassium Chloride (Merck KGaA, Darmstadt, Germany) 30 mM Ammonium Chloride (Merck KGaA, Darmstadt, Germany) 5 mM Imidazole (Sigma-Aldrich, GmbH, Germany) Water
14. Elution buffer (pH7.6) 20 mM Tris HCl (pH7.6) (Sigma-Aldrich, GmbH, Germany) 10 mM Magnesium Chloride (Merck KGaA, Darmstadt, Germany) 150 mM Potassium Chloride (Merck KGaA, Darmstadt, Germany) 30 mM Ammonium Chloride (Merck KGaA, Darmstadt, Germany) 150 mM Imidazole (Sigma-Aldrich, GmbH, Germany) Water
15. 1X Polymix 5 mM Ammonium Chloride (Merck KGaA, Darmstadt, Germany) 95 mM Potassium Chloride (Merck KGaA, Darmstadt, Germany) 0.5 mM Calcium Chloride (Merck KGaA, Darmstadt, Germany) 8 mM Putrescine (BIOSYNTH AG, Staad, Switzerland) 1 mM Spermidine (BIOSYNTH AG, Staad, Switzerland) 5 mM Potassium Phosphate (Merck KGaA, Darmstadt, Germany) 1 mM DTT (Sigma-Aldrich, GmbH, Germany)
16. Sucrose cushion 20 mM TrisHCl (pH7.5) (Sigma-Aldrich, GmbH, Germany) 500 mM Ammonium Chloride (Merck KGaA, Darmstadt, Germany) 10.5 mM Magnesium acetate (Merck KGaA, Darmstadt, Germany) 0.5 mM EDTA (Merck KGaA, Darmstadt, Germany) 1.1 M Sucrose (Sigma-Aldrich, GmbH, Germany) 3 mM β mercaptoethanol (Sigma-Aldrich, GmbH, Germany) 5.2.4 For 80S ribosome purification
17. 4X YP media 1% yeast extract 2% peptone water
18. YPD media/plates 1% yeast extract (BactoTM , Becton, Dickinson & company, MD, USA) 2% peptone (BactoTM , Becton, Dickinson & company, MD, USA)
27

Water Autoclave and add 20% glucose (sterile) to final conc. of 2% For plates add 2% agar before autoclaving
19. Lysis buffer 20 mM TrisHCl (pH7.5) 100 mM Potassium Chloride (Merck KGaA, Darmstadt, Germany) 10.5 mM Magnesium Acetate (Merck KGaA, Darmstadt, Germany) 0.5 mM EDTA (Sigma-Aldrich, GmbH, Germany) 3 mM β mercaptoethanol (Sigma-Aldrich, GmbH, Germany) 1 mM PMSF (Sigma-Aldrich, GmbH, Germany)
20. Sucrose cushion 20 mM TrisHCl (pH7.5) (Sigma-Aldrich, GmbH, Germany) 100 mM Potassium Chloride (Merck KGaA, Darmstadt, Germany) 10.5 mM Magnesium Acetate (Merck KGaA, Darmstadt, Germany) 0.5 mM EDTA (Sigma-Aldrich, GmbH, Germany) 3 mM β mercaptoethanol (Merck KGaA, Darmstadt, Germany) 1 mM PMSF 37.5% sucrose (Sigma-Aldrich, GmbH, Germany)
21. High salt wash buffer 20 mM TrisHCl (pH7.5) (Sigma-Aldrich, GmbH, Germany) 10 mM Magnesium Chloride (Merck KGaA, Darmstadt, Germany) 800 mM Potassium Chloride (Merck KGaA, Darmstadt, Germany) Water
22. Low salt wash buffer 20 mM TrisHCl (pH7.5) (Sigma-Aldrich, GmbH, Germany) 10 mM Magnesium Chloride (Merck KGaA, Darmstadt, Germany) 50 mM Sodium Chloride (Merck KGaA, Darmstadt, Germany) Water
23. High salt wash sucrose cushion 20 mM TrisHCl (pH7.5) (Sigma-Aldrich, GmbH, Germany) 10 mM Magnesium Chloride (Merck KGaA, Darmstadt, Germany) 800 mM Potassium Chloride (Merck KGaA, Darmstadt, Germany) 25% sucrose (Sigma-Aldrich, GmbH, Germany)
24. Resuspension buffer 20 mM TrisHCl (pH7.5) (Sigma-Aldrich, GmbH, Germany) 10 mM Magnesium Chloride (Merck KGaA, Darmstadt, Germany) 100 mM Potassium Chloride (Merck KGaA, Darmstadt, Germany) Water 5.2.5 For RNA preparation
28

25. Phenol Phenol (Merck KGaA, Darmstadt, Germany) add Set the pH to 7.9 with TrisHCl (pH7.9) 25 ml of TrisHCl (pH7.9) Store in coloured bottle. 5.2.6 Drugs
26. Psi114 (100 mM) 100 mM Psi114 (Marc Blondel Lab) DMSO (Sigma-Aldrich chemie, GmbH, Riedstr, Steinheim, Germany)
27. Psi207 (100 mM) 100 mM Psi207 (Marc Blondel Lab) DMSO (Sigma-Aldrich chemie, GmbH, Riedstr, Steinheim, Germany) 5.3 Methods 5.3.1 Overexpression and purification of Human carbonic anhydrase I To overexpress HCA, pET3a vector containing HCA gene was transformed into BL21 (DE3) strain of E.coli. Induction of HCA was done in one case using 1 mM IPTG (where Zn was not added in the induction media) (will be called HCA (-Zn)). In other case 0.4 mM IPTG, and 0.5 mM zinc sulphate was used for induction (will be called HCA (+Zn)). For purification of HCA the culture was centrifuged at 4800 g (SorvallR RC3C plus SS-34 rotor, Waltham, MA, USA), for 15 minutes and the cells obtained were used for purification. Cell lysis of both batches of cells (HCA (+Zn) and HCA (-Zn)) was done by lysis buffer for E.coli cells and this was followed by French press (Nike, Eskilstuna, Sweden) to open the cells. The lysed cells were centrifuged at 43920 g (SorvallR RC5C plus, SS-34 rotor, Waltham, MA, USA) for a period of 45 minutes and the supernatant was collected. Supernatant obtained was treated with 0.5 g DEAE cellulose fibre (batch mode of treatment) overnight with gentle rocking, which was followed by centrifugation of the sample at 43920 g and pellet obtained was discarded and the supernatant was collected and stored at 4˚C. Ammonium sulphate based precipitation of the proteins was done and pellet was collected. Finally gel filtration of the ammonium sulphate based precipitated protein was done where pellet was dissolved in the wash buffer and the loaded on to the column at 1ml/min (Superdex 75 (radius 16 cm, height 75 cm) connected to ÄktaprimeTM, GE healthcare, Uppsala, Sweden) and finally elution of the bound proteins was done by excess of wash buffer. The peak fractions were collected and SDS PAGE was done for the peak fractions and the overall yield of the proteins was seen. This was followed by Bradford assay to quantitate the protein yield (Armstrong, Myers et al. 1966), (Kannan, Ramanadham et al. 1984). Briefly BSA standard solution (1:50) was prepared and this was titrated with Bradford reagent which was measured spectrophometrically at 595 nm. Similar dilutions for HCA (+Zn) (1:50, 1:100) and HCA (-Zn) (1:50) was prepared and was titrated with Bradford reagent. The slopes of the curves obtained for each of the proteins was compared and from their concentrations of HCA (+Zn) and HCA (-Zn) was calculated.
29

5.3.2 Kinetics of Human carbonic anhydrase I In order to study the kinetics of HCA, varying concentrations of PNPA was titrated with HCA (500 nM) which was added to the reaction mix and then the colour change was measured spectrophotometrically (Hitachi U-2900 spectrophotometer, Japan) at 400 nm. From these kinetic curves were plotted and Michaelis Menten Constants (Km and Vmax) were calculated for each batch of HCA. In both cases BCA acted as control. The effect the solvent had for dissolving PNPA on HCA activity was then studied. In these studies PNPA was either dissolved in 95% ethanol or in acetonitrile and from their kinetic parameters on using different solvents for PNPA were calculated. Further studies where effect of varying concentrations of metal ion (Zn) on HCA activity was then carried out. Here increasing concentrations of Zn (as zinc sulphate) was added in the refolding buffer wherein HCA was dissolved and titrated against 10 mM PNPA dissolved in either 95% ethanol or acetonitrile and kinetic parameters were calculated. 5.3.3 Human carbonic anhydrase I unfolding and refolding 5.3.3.1 Effect of EDTA on Human carbonic anhydrase I unfolding In order to study the effect of EDTA on HCA unfolding, increasing concentrations of EDTA (0 to 60 µM) was added in the denaturation mix containing 31.6 µM HCA (+Zn) and other components of denaturation buffer was incubated overnight at 25˚C . HCA (+Zn) was then diluted 100 times and incubated for 30 minutes at 25˚C in the refolding buffer giving final HCA concentration of 316 nM and diluting 6 M guanidine HCl to 60 mM. This was followed by titrating refolded HCA activity against 10 mM PNPA which was measured spectrophometrically at 400 nm. A plot was then generated with percentage HCA refolding activity (with respect to native enzyme activity) on Y axis and concentration of EDTA in denaturation mix on the X axis. From this curve the optimum concentration of EDTA needed for maximum HCA unfolding was calculated. 5.3.4 Purification of 70S, 50S ribosomes For purifying 70S ribosomes JE28 strain of E.coli (Ederth, Mandava et al. 2009) was used. 1 colony of JE28 was inoculated in 20 ml of LB with kenamycin (50 µg/ml). The cells were grown overnight. This culture was transferred to 2 L LB and was grown till the absorbance at 595 nm reached 1.3 (Hitachi U-2001 spectrophotometer, Japan). The culture was then kept at 37˚C for 1 hour then it was transferred to 4˚C for half an hour. Cells were then collected by centrifuging the culture at 4800 g (SorvallR RC3C plus SS-34 rotor, Waltham, MA, USA) for 30 minutes. Pellet obtained was weighed and frozen at -80˚C (Liquid nitrogen, LD-35, Liquid Dewar, Taylor- Wharton, Bridgeville, PA, USA) for later lysis of the cells. For lysis the cell pellet was dissolved in sufficient amount of 70S ribosome lysis buffer and then the cells were opened by French press. The lysate collected was centrifuged at 43920 g (SorvallR RC5C plus, Waltham, MA, USA) for 40 minutes. The supernatant was collected and the pellet was discarded. The column (His TrapTM HP, 5 ml) was washed with wash buffer this was followed by loading of the sample on to the column at 2 ml/min. The elution of the sample was done either with 50% length 500 mM imidazole gradient or with elution buffer. The fractions obtained were collected and dialysed overnight with 1X polymix. Then concentration of the pellets obtained was done by loading the sample on sucrose cushion and ultracentrifuging the sample at 200,000 g (Bergman Labora sorvallR RC M150GX S80A3 rotor, Stockholm, Sweden) for 4 hours. The pellet obtained was dissolved in 1X polymix and was later dialysed
30

in refolding buffer. The concentration of ribosome was calculated spectrophotometrically (Hitachi U-2001 spectrophotometer, Japan) at 260 nm. For 50S ribosome subunit separation same protocol as above was followed with exception that no magnesium was added in the wash buffer, and elution was done with elution buffer (with no magnesium). The fractions obtained had in majority 50S ribosome subunits. 5.3.5 Purification of 80S ribosomes For purification of 80S ribosomes single colony of MH272 3Fα stain of Saccharomyces cerevisiae was grown in 20 ml of YPD media overnight and then it was transferred to bigger culture (around 50 ml) to get sufficient starting absorbance at 595 nm. This second preculture was inoculated in 6 L of YPD media and was grown at 30˚C with gentle shaking till the final absorbance reached 0.16. The cells were then pelleted down at 4800 g (SorvallR RC3C plus SS-34 rotor, Waltham, MA, USA) for 42 minutes. The pellet obtained was resuspended in 200 ml of sterile water and were recentrifuged at 2600 g (Eppendorf centrifuge 5810R, A-4-81 rotor, Hamburg, Germany). The pellets were obtained and were stored at -80˚C (Liquid nitrogen, LD-35, Liquid Dewar, Taylor- Wharton, Bridgeville, PA, USA). Lysis of the cells was done by dissolving the cell pellets in 80S ribosome lysis buffer followed by French press 3 times to get complete lysis of the cells. The lysate was collected and centrifuged at 26,900 g (SorvallR
RC5C plus, SS-34 rotor, Waltham, MA, USA) for 30 minutes. The supernatant was carefully collected and was recentrifuged (Beckman ultracentrifuge, Ti50.2 rotor, Krefeld, GmbH, Germany) at 81,000 g for 40 minutes. Supernatant collected was loaded on to the sucrose cushion and was centrifuged at 71,000 g (Beckman ultracentrifuge, Ti50.2 rotor, Krefeld, GmbH, Germany) for 20 hours. The pellet obtained was divided into two parts high salt and low salt wash. The low salt wash pellet was washed with low salt wash buffer and then it was dissolved in low salt wash buffer and its concentration was measured at 260 nm.While high salt wash pellet was dissolved in high salt wash buffer and loaded on to high salt wash sucrose cushion. It was centrifuged at 200,000 g (Bergman Labora SorvallR RC M150GX, S80A3 rotor, Stockholm, Sweden). Finally pellet obtained was dissolved in resuspension buffer and its concentration was measured at 260 nm. 5.3.6 Isolation of total RNA from ribosomes For purification of total RNA in the case of both 70S and 80S ribosomes, 100 µl of the 70S and 80S ribosomes were taken and equal volume of phenol and chloroform were added. The sample was then centrifuged at 21,341 g (Eppendorf centrifuge 5417C, Hamburg, Germany) for 10 minutes and the supernatant was collected carefully. The above step was repeated 9 to 10 times and in every step the supernatant was collected and was resuspended in equal volume of phenol, chloroform mixture. Finally to the supernatant equal volume of chloroform was added and it was centrifuged at 21,341 g (Eppendorf centrifuge 5417C, Hamburg, Germany) and supernatant was collected. To this 0.1X volume 3 M sodium acetate and 2X volume ice cold 95% ethanol was added. This sample was kept overnight at -20˚C and later the supernatant was discarded and pellet was collected and dried using vacuum. Finally the pellet was dissolved in DEPC water and concentration was measured using nano drop (Thermoscientific, Wilmington, DE, USA). RNA was stored in -20˚C.
31

5.3.7 Ribosome assisted protein folding. 70S and 80S ribosomes were used in the protein folding assay. Essentially HCA (+Zn) was denatured in the denaturation mix and then it was refolded in the same fashion in the refolding buffer as explained in section 5.3.4 with the difference that in the refolding buffer also included 70S or 80S ribosomes. First the concentration profile of 70S and 80S ribosomes was generated by increasing the concentrations of 70S (100 nM to 500 nM) and 80S (25 nM to 400 nM) and titrated with 10 mM PNPA, an increase in the refolding activity (spectrophotometrically ) of HCA (+Zn) was plotted against the concentration of 70S or 80S ribosome (Das et al. 1996). Once the concentration of ribosome needed for refolding assay was standardized time course studies were performed with 70S (400 nM) or 80S (200 nM) ribosomes. In the time course studies a denaturation mix containing HCA (+Zn) was prepared and transferred to a refolding buffer from which aliquots were taken at increasing time intervals and to this 10 mM PNPA was added and the change in absorbance was measured spectrophotometrically at 400 nm. In all the experiments self folding activity of HCA (+Zn) (without ribosomes in the refolding buffer) acted as control. 5.3.8 Total RNA assisted protein folding The total RNA isolated from 80S ribosomes was used in protein folding studies. In these experiments HCA (+Zn) was denatured in the denaturation mix as described in section 5.3.4. The refolding of HCA (+Zn) was done by diluting the denaturation mix in the refolding buffer containing all the components of refolding buffer, but also increasing concentration of total RNA. This mix was titrated with 10 mM PNPA and the increase in HCA (+Zn) refolding activity was measured spectrophotometrically at 400 nm. The percentage refolding of HCA (+Zn) was plotted on Y axis while concentration of total RNA was on the X axis. The plot between the two gave concentration profile of total RNA for HCA (+Zn) refolding activity. 5.3.9 Antiprion drug activity In order to study antiprion activity of Psi207 (6AP) and Psi114 (GA), a concentration profile of both drugs in 80S ribosome assisted refolding blocking activity was generated (Tribouillard-Tanvier, Dos Reis et al. 2008). In this assay HCA (+Zn) was denatured as given in section 5.3.4. For refolding in the refolding buffer 80S ribosome (200 nM) and increasing concentrations of Psi207 and Psi114 were added and were incubated together in the refolding buffer for 30 minutes and then to this refolding mix denaturation mix was added and increase in HCA (+Zn) refolding activity was measured spectrophotometrically at 400 nm. From this assay the inhibitory concentrations for the drugs was characterized and was used further for time course studies. For time course studies of antiprion drugs, varying concentrations of Psi207 (10 µM to 400 µM), Psi114 (10 µM to 400 µM) were incubated for 30 minutes with 80S ribosomes (200 nM) and then at increasing time intervals denaturation mix was added in the refolding master mix and aliquots of final mix was titrated against 10 mM PNPA spectrophotometrically at 400 nm.
32

6. Acknowledgement I would like to acknowledge my supervisor Ass. Prof. Suparna Sanyal for her constant encouragement and kind guidance throughout my project. My heartfelt regards goes to Prof. Mans Ehrenberg one of the few people I have seen to have so much vision and knowledge. I am also grateful to Kristine Piesker for her constructive criticism and motivation without which it would have been difficult for me to gain necessary confidence. I would like to thank Chandrashakher Mandava and Ravi Koripella for their help during my experiments. And Finally to Dr Santanu Dasgupta who had inspired me through his great knowledge about not only the topic I worked upon but also from other subjects as well. 7. References Aguzzi, A. and A. M. Calella (2009). "Prions: protein aggregation and infectious diseases."
Physiol Rev 89(4): 1105-1152. Anfinsen, C. B. (1973). "Principles that govern the folding of protein chains." Science 181(96):
223-230. Armstrong, J. M., D. V. Myers, et al. (1966). "Purification and properties of human erythrocyte
carbonic anhydrases." J Biol Chem 241(21): 5137-5149. Bach, S., D. Tribouillard, et al. (2006). "A yeast-based assay to isolate drugs active against
mammalian prions." Methods 39(1): 72-77. Bartlett, A. I. and S. E. Radford (2009). "An expanding arsenal of experimental methods yields
an explosion of insights into protein folding mechanisms." Nat Struct Mol Biol 16(6): 582-588.
Berg, J. M., J. L. Tymoczko, et al. (2002). Biochemistry. New York, W.H. Freeman. Beringue, V., D. Vilette, et al. (2004). "PrPSc binding antibodies are potent inhibitors of prion
replication in cell lines." J Biol Chem 279(38): 39671-39676. Chattopadhyay, S., B. Das, et al. (1996). "Reactivation of denatured proteins by 23S ribosomal
RNA: role of domain V." Proc Natl Acad Sci U S A 93(16): 8284-8287. Coleman, J. E. (1984). "Carbonic anhydrase: zinc and the mechanism of catalysis." Ann N Y
Acad Sci 429: 26-48. Das, B., S. Chattopadhyay, et al. (1996). "In vitro protein folding by ribosomes from
Escherichia coli, wheat germ and rat liver: the role of the 50S particle and its 23S rRNA." Eur J Biochem 235(3): 613-621.
Das, D., A. Das, et al. (2008). "Role of the ribosome in protein folding." Biotechnol J 3(8): 999-1009.
Dobson, C. M. (2003). "Protein folding and misfolding." Nature 426(6968): 884-890. Ederth, J., C. S. Mandava, et al. (2009). "A single-step method for purification of active His-
tagged ribosomes from a genetically engineered Escherichia coli." Nucleic Acids Res 37(2): e15.
Ertmer, A., S. Gilch, et al. (2004). "The tyrosine kinase inhibitor STI571 induces cellular clearance of PrPSc in prion-infected cells." J Biol Chem 279(40): 41918-41927.
Gilch, S., C. Krammer, et al. (2008). "Targeting prion proteins in neurodegenerative disease." Expert Opin Biol Ther 8(7): 923-940.
Hartl, F. U. and M. Hayer-Hartl (2009). "Converging concepts of protein folding in vitro and in vivo." Nat Struct Mol Biol 16(6): 574-581.
Ingrosso, L., A. Ladogana, et al. (1995). "Congo red prolongs the incubation period in scrapie-infected hamsters." Journal of Virology 69(1): 506-508.
33

34
Innocenti, A., M. Hilvo, et al. (2008). "Carbonic anhydrase inhibitors: the very weak inhibitors dithiothreitol, beta-mercaptoethanol, tris(carboxyethyl)phosphine and threitol interfere with the binding of sulfonamides to isozymes II and IX." Bioorg Med Chem Lett 18(6): 1898-1903.
Kannan, K. K., M. Ramanadham, et al. (1984). "Structure, refinement, and function of carbonic anhydrase isozymes: refinement of human carbonic anhydrase I." Ann N Y Acad Sci 429: 49-60.
Karpuj, M. V., K. Giles, et al. (2007). "Phosphorothioate oligonucleotides reduce PrP levels and prion infectivity in cultured cells." Mol Med 13(3-4): 190-198.
Kramer, G., D. Boehringer, et al. (2009). "The ribosome as a platform for co-translational processing, folding and targeting of newly synthesized proteins." Nat Struct Mol Biol 16(6): 589-597.
Ladogana, A., P. Casaccia, et al. (1992). "Sulphate polyanions prolong the incubation period of scrapie-infected hamsters." Journal of General Virology 73 ( Pt 3): 661-665.
Lehninger, A. L., D. L. Nelson, et al. (2000). Lehninger principles of biochemistry. New York, Worth Publishers.
Lehninger, A. L., D. L. Nelson, et al. (2005). Lehninger principles of biochemistry. New York, W.H. Freeman.
Lehtonen, J., B. Shen, et al. (2004). "Characterization of CA XIII, a novel member of the carbonic anhydrase isozyme family." J Biol Chem 279(4): 2719-2727.
Pfeifer, A., S. Eigenbrod, et al. (2006). "Lentivector-mediated RNAi efficiently suppresses prion protein and prolongs survival of scrapie-infected mice." J Clin Invest 116(12): 3204-3210.
Pocker, Y. and J. T. Stone (1968). "The catalytic versatility of erythrocyte carbonic anhydrase. VI. Kinetic studies of noncompetitive inhibition of enzyme-catalyzed hydrolysis of p-nitrophenyl acetate." Biochemistry 7(8): 2936-2945.
Priola, S. A., A. Raines, et al. (2000). "Porphyrin and phthalocyanine antiscrapie compounds." Science 287(5457): 1503-1506.
Samanta, D., D. Mukhopadhyay, et al. (2008). "Protein folding by domain V of Escherichia coli 23S rRNA: specificity of RNA-protein interactions." J Bacteriol 190(9): 3344-3352.
Sanyal, S. C., S. Pal, et al. (2002). "23S rRNA assisted folding of cytoplasmic malate dehydrogenase is distinctly different from its self-folding." Nucleic Acids Res 30(11): 2390-2397.
Stryer, L. (1995). Biochemistry. New York, W.H. Freeman. Taylor, D. J., B. Devkota, et al. (2009). "Comprehensive molecular structure of the eukaryotic
ribosome." Structure 17(12): 1591-1604. Tribouillard-Tanvier, D., S. Dos Reis, et al. (2008). "Protein folding activity of ribosomal RNA
is a selective target of two unrelated antiprion drugs." PLoS One 3(5): e2174. Tribouillard-Tanvier, D., S. Dos Reis, et al. (2008). "Protein Folding Activity of Ribosomal
RNA Is a Selective Target of Two Unrelated Antiprion Drugs." PLoS One 3(5): -. Wickner, R. B., H. K. Edskes, et al. (2007). "Yeast prions: evolution of the prion concept."
Prion 1(2): 94-100. Yazgan, A. and R. W. Henkens (1972). "Role of zinc (II) in the refolding of guanidine
hydrochloride denatured bovine carbonic anhydrase." Biochemistry 11(7): 1314-1318.