molecular characterization and expression analysis of nac family transcription factors in tomato
TRANSCRIPT

ORIGINAL PAPER
Molecular Characterization and Expression Analysis of NACFamily Transcription Factors in Tomato
Xiaohong Kou & Shuang Wang & Mengshi Wu &
Runzi Guo & Zhaohui Xue & Nan Meng & Xiaomin Tao &
Mimi Chen & Yifei Zhang
Published online: 13 October 2013# Springer Science+Business Media New York 2013
Abstract The NAC family is considered one of the largestplant-specific transcription factors families, and functions indiverse and vital physiological processes during development.In the present study, we performed a complete bioinformaticsanalysis of the NAC family transcription factors in tomato, andannotated the non-redundant SlNAC1-74 proteins into 12 sub-groups. Six NAC genes from tomato, which were designatedSNAC4–SNAC9 , were studied instensively. The expression anal-ysis indicated that each SNAC gene exhibited a specific expres-sion pattern in the tissues examined. SNAC4 and SNAC6 weremost highly expressed in the stems and leaves, whereas theexpression levels of SNAC5 , SNAC8 and SNAC9 were higherin young leaves and old leaves, respectively. In addition, theexpression patterns of SNAC genes were characterized duringthe development of tomato fruits. All of the genes were furtherinvestigated to determine their responsiveness to hormones, anda coordinated expression was observed. The expression of theSNAC gene transcripts was induced by ABA, SA and short-timeethylene treatment, whereas their transcription was inhibited byGA, 6-BA and IAA. Our present study provides a useful refer-ence for future investigations of NAC genes in tomato and otherfleshy fruits.
Keywords NAC . Transcription factor . Tomato .
Development . Hormones
Introduction
Transcription factors (TFs) are proteins that interact withDNA promoters and are involved in gene expression andregulation. Various plant TFs, which contain various function-al domains, such as MYB, DREB and NAC, have been wellstudied. The NAC gene family name is an acronym derivedfrom the first three identified genes containing this domain:NAM from Petunia and, ATAF1/2 and CUC2 fromArabidopsis (Souer et al. 1996; Aida et al. 1997). The NACfamily members are characterized with a highly conservedNAC domain region at the N-terminal as a functional domain,which can be further divided into five subdomains (A-E). TheC-terminal region is highly variable in sequence compositionand length and, plays a vital role as a transcriptional activatoror repressor (Xie et al. 2000; Ooka et al. 2003; Tran et al.2004; Kim et al. 2007a).
The plant-specific NAC family refers to proteins contain-ing the NAC domain, and is one of the largest transcriptionfactor families in plants. A continuously increasing number ofNAC genes have been identified and studied in differentplants. There are 105 and 140 NAC or NAC-like genes inArabidopsis and rice, respectively, and these can be furtherclassified into 18 subfamilies (Ooka et al. 2003; Fang et al.2008). The specific subfamilies are composed of multi-functional proteins involved in various plant biological pro-cesses related to growth and development, including celldivision (Kim et al. 2006), seed and embryo development(Duval et al. 2002; Sperotto 2009), the formation of lateralroots and cotyledons (Aida et al. 1997; Weir et al. 2004; He
Xiaohong Kou and Shuang Wang made equal contributions to this work.
Electronic supplementary material The online version of this article(doi:10.1007/s11105-013-0655-3) contains supplementary material,which is available to authorized users.
X. Kou : S. Wang (*) :M. Wu :R. Guo : Z. Xue (*) :N. Meng :X. Tao :M. Chen :Y. ZhangSchool of Chemical Engineering and Technology, Tianjin University,Tianjin 300072, People’s Republic of Chinae-mail: [email protected]: [email protected]
Plant Mol Biol Rep (2014) 32:501–516DOI 10.1007/s11105-013-0655-3

et al. 2005), the maintenance of shoot apical meristems (Soueret al. 1996; Aida et al. 1997; Kim et al. 2007b; Mao et al.2007), organ growth and senescence (Sablowski andMeyerowitz 1998; Guo and Gan 2006; Liu et al. 2009; Kouet al. 2012a, b), the synthesis of secondary walls (Zhong et al.2007b; Ko et al. 2007) and plant nutrition regulation (Uauyet al. 2006; Ogo et al. 2008).
Transcriptome studies have shown that many NAC tran-scription factors are involved in the response to abiotic stressand hormonal treatments. Furthermore, some of the NACtranscription factors have been reported to be potential genesfor the engineering of stress resistance during the plant life-cycle. Interestingly, phylogenetic analysis shows that most ofthe stress-responsive NACs belong to the ATAF and AtNAC3subfamilies. The SNAC1 and SNAC2 genes in rice are in-duced by cold, salinity, drought, and ABA (abscisic acid)treatment, and the transgenic plants that overexpress thesegenes demonstrate increased tolerance against cold, salt, anddrought (Hu et al. 2006; Hu et al. 2008). SNAC2 andOsNAC10 can improve the expression of some related genesunder stress conditions (Jeong et al. 2010). The ANAC19 , 55and 72 genes in Arabidopsis are inducible by drought, saltand ABA, and have been confirmed to be positive regulatorsin the ABA signaling pathway and stress tolerance (Tran et al.2004; Fujita et al. 2004). NAC genes are also involved in thedefense against biotic stress and are up-regulated under con-ditions of pathogen infection and attack. It has been shownthat OsNAC19 in rice is elevated by Magnaporthe griseainfection (Lin et al. 2007).
Tomato is one of the most popular fleshy fruits in the worldand exhibits a wealth of nutrition. The abundant genetic re-sources available make tomato an essential model system forthe study of the development, ripening and senescence of fleshyfruits (Alexander and Grierson 2002; Giovannoni 2004; Kleeand Giovannoni 2011). Most available research focuses mainlyon model species, such as Arabidopsis and rice, and the char-acterization of NAC genes in tomato is still rudimentary. AnNAC gene, SlNAC3 , isolated from a tomato flower cDNAlibrary was recently reported (Han et al. 2012). A digital anal-ysis revealed that SlNAC3 demonstrates a tissue-specific ex-pression pattern and is inhibited by drought, salt stress andABA treatment. The completion of the high-quality sequencingof the tomato genome has provided an excellent opportunity forthe genome-wide analysis of NAC family genes (InternationalTomato Genome Sequencing Project 2012).
In the present study, 74 SlNAC genes were identified andclassified according to their phylogeny. The molecular fea-tures of the genes and their encoded proteins were also ana-lyzed. We also studied the developmental expression patternsof six genes, namely SNAC4-9 , which belong to the NAP andAtNAC3 subgroups in tomato. Through molecular cloningand sequence characterization, we found that all six genesshare a similar genomic organization and high sequence
similarities, especially within the NAC domains. A tissue-specific expression analysis showed that these genes exhibitdifferent expression patterns, particularly during fruit devel-opment. We further investigated the responses of the genes tohormone treatments. The present study constitutes the firstsystematic study ofNAC genes in tomato, and the informationgenerated suggests that the NAC genes may regulate tomatogrowth and fruit development.
Materials and Methods
Plant Growth and Treatments
Plants of tomato cultivar AC were used throughout the exper-iments described in this manuscript. The plants were field-grown under normal conditions at Tianjin University.
The vegetable tissues, such as leaves, stems and roots, werecollected from seedlings starting 30 days after flowering(DAF), and the open flowers were sampled and prepared fortissue-specific expression assays. The developing fruits from35 to 55 DAF, at the green mature, breaker, pink, and red ripestages, were collected, divided into the pericarp, radial peri-carp, and columella, frozen immediately in liquid nitrogen andstored at −80°C until further use.
For exogenou hormones treatments, 10 g tomato fruittissue discs (diameter of 10 mm) at the green mature stagewere immersed in 50 ml vials containing 0.1 mM ABA(abscisic Acid), 0.1 mM IAA (indole Acetic Acid), 0.1 mMGA (gibberellic Acid), 0.1 mM 6-BA (6-benzylaminopurine)and 1mMSA (salicylic Acid), respectively. The control groupdiscs were treated with the buffer solution (50 mmol/L citricacid, 100 mmol/L sorbitol and 0.33 mmol/L chlorampheni-col). To evaluate the effects of ethylene treatment, tomatofruits were placed in vials, which were injected with 50 μLof 2 % (v/v) ethylene (air as a control) to obtain a finalconcentration of 100 μL/L. The hormone treatments lasted0, 2, 4, 6, 8, 10, 12, 24, or 48 h at 23°C. All of the sampleswere rinsed, frozen in liquid nitrogen and stored at −80°C forfurther use. Three biological replications were performed foreach treatment of sampling time point.
Collection of SlNAC Gene Family Members
We selected 10 reported NAC protein sequences from fivesubgroups of the NAC family as queries for a BLASTP searchin the tomato SOL database (http://solgenomics.net/), using an e-value of 1e-10. The GenBank accession numbers of the repre-sentative genes are the followings: ATAF1/2 (X74755/X74766),AtNAM/NAP/AtNAC2 (AF123311/AJ222713/AB049071),CUC1/2/3 (AB049069/AB002560/AF543 194), TIP(AF281062), and AtNAC072 (At4g27410). ConservedDomain Database (CDD) search in NCBI (http://www.ncbi.
502 Plant Mol Biol Rep (2014) 32:501–516

nlm.nih.gov/ Structure/cdd/wrpsb.cgi) was performed to confirmthe NAC domain of each sequence.
Phylogenetic Analysis and Sequence Alignment
Previously published NAC sequences from Arabidopsis andother species, including Oryza sativa, Brassica napus, Triticumsativa , Antirrhinum majus, were retrieved from the GenBankdatabase. For the phylogenetic analysis, an unrooted tree wasconstructed with the MEGA5 software (Tamura et al. 2007)using the Neighbor-Joining method with 1000 bootstrapreplicates.
A multiple sequence alignment of the representative pro-tein sequences of SNAC4-9 was performed using theClustalX1.83 program (Thompson et al. 1997). TheGenBank accession numbers of the reported proteins werethe following: CitNAC (EF185419)/AtNAM (AF123311)/NAP (AJ222713)/ AtNAC2 (AB049071)/GmNAC1(AY974349) for NAP and, AtNAC3 (AB049070)/AtNAC019 (At1g52890) /AtNAC072 (At4g27410)/GmNAC3 (AY974351)/GmNAC4 (AY974352) for AtNAC3.
Sequence Analysis
Information regarding the SlNAC gene structures, chromosomallocalizations, and transcripts was procured from the SOL data-base. The positions of the SlNAC genes on the tomato chromo-somemaps were drawn andmodified manually, with annotation.The protein-encoding characteristics were procured fromExPASy ProtParam (http://web.expasy.org/protparam/), and thehydrophobicmapswere further verified by ProtScale (http://web.expasy.org/protscale). A protein motif analysis wasaccomplished using the SMART database (http://smart.embl-heidelberg.de/), and the transmembrane proteins were furtherconfirmed by TMHMM (http://www.cbs.dtu.dk/services/TMHMM-2.0/). The subcellular localizations were predictedby WoLF PSORT (http://wolfpsort.org/). A transcriptomeanalysis of various tissues in the tomato cultivar Heinz and thewild relative Solanum pimpinellifolium was used to predict theSlNAC gene expression patterns. The data corresponding to theSlNAC genes were downloaded from the Tomato Expressiondatabase (http://ted.bti.cornell.edu/). Based on the data, the heatmap was generated using theMeV4.8 software (http://www.tm4.org/mev.html) following the instruction of the software. Byexpressing the signal strength and expression directly reflectsthe gap, screening candidate genes. The digital expressionanalysis of tomato SlNAC genes was performed by the visualreflection of signal strength and expression disparity.
RNA Extraction and RT-qPCR Analysis
Isolating RNA from samples ready for the reverse transcriptasequantitative PCR (RT-qPCR) applications was performed using
the Column Plant RNAOUT kit (Tiandz#71203, China). Theconcentration and quality of the RNA samples were examinedusing spectrophotometer (Nanodrop ND-1000). Samples cDNAwas synthesized using the Transcript One-Step gDNA removaland cDNA synthesis supermix (TransGen#AH11-03, China).qPCR analyses of individual genes were designed usingPrimer5. Briefly, for each qPCR reaction, 1 μL each dilutedsample was used as a template in a 25 μL reaction containing12.5 μL 2× SYBR green supermix (TransGen#AQ101), 8.5 μLddH2O and 0.5 μL of each primer. All qPCR reactions wereperformed on Light Cycler 480 thermocycler with 45 cycles.Cycle threshold (Ct) values were determined by the Light Cyclersoftware assuming 100 % primer efficiency. The β-tublin genewas used as an internal control. The Primer pairs used in thisresearch are shown in Table 1. For the development analyses, theexpression levels of individual genes in the root (R) or pericarp atgreen stage (G1) were set to 1. For the hormone treatmentanalyses, the expression levels of individual genes under 0 htreatment were set to 1.
Statistical Methods
The RT-qPCR gene expression was quantified using the 2-△△Ct
comparative methods. Results are presented as the means ±standard deviation from three biological replicates of eachexperiment. The significant differences (p =0.05) betweenmean values were determined by analysis of variance(ANOVA) using IBM SPSS Statistics 20 (SPSS commercialsoftware, SPSS Inc., Chicago, IL, USA) software.
Results
Identification of SlNAC Genes
After the sequencing of the tomato genome was completed,we used 10 reported NACs as sequence entries for an exhaus-tive search. By removing any redundant sequences and dif-ferent transcripts of the same gene, we identified 74 putativeSlNAC genes preliminarily. The protein-encoding sequenceswere confirmed by CDD for the presence of the NAM do-main. For convenience, we designated the genes SlNAC1 toSlNAC74 according to the chromosome distribution.
Phylogenetic Analysis and Sequence Alignment
The clarification of the phylogenetic relationships and classi-fications is important for the functional prediction of this genefamily. A combined phylogenetic tree was constructed withthe aligned SlNAC and published NAC sequences using theMEGA5 software (Fig. 1).
Of the 74 total SlNACs, 54 SlNACs were divided into 12subgroups with high bootstrap support. The subgroups were
Plant Mol Biol Rep (2014) 32:501–516 503

named by the homologous NACs, according to Ooka’s classifi-cation (Ooka et al. 2003). Eleven, nine, seven, and five SlNACsbelonged to the NAM,VND, TERN, andANAC063 subgroups,respectively, whereas the NAC2 and TIP subgroups containedfour SlNACs each, the NAP, ATAF and ANAC011 subgroupscontained three SlNACs each, the AtNAC3 and SENU5 sub-groups contained two SlNACs each, and the NAC1 subgroupcontained one SlNAC. The remaining 20 sequences had lesshomology with the known NACs and were clustered into differ-ent unknown subgroups. SlNAC20 was clustered as a memberof the ATAF subgroup and was most closely related to StNAC.SlNAC46 belonged to the NAM subgroup, and SlNAC49 wasclustered into the NAC1 subgroup. SlNAC69 was a member ofthe TERN subgroup. The identities of SlNAC70 and NTL, werehigh, i.e., approximately 99 %. Our data showed that SlNAC1was homologous to SENU5 with high bootstraps support.SlNAC48 and SlNAC59 were highly related, shared 99 % se-quence similarity and were classified as members of the NAPsubgroup. The deduced proteins SlNAC47 and SlNAC71 wereclassified as members of the AtNAC3 subgroup. For furtherresearch, we selected six genes belonged to the NAP andAtNAC3 subgroups, and conveniently renamed SlNAC48, 71,59, 24, 47 and 19 as SNAC4, 5, 6, 7, 8 and 9, respectively, basedon a previous report regarding SlNAC3 (Han et al. 2012) .
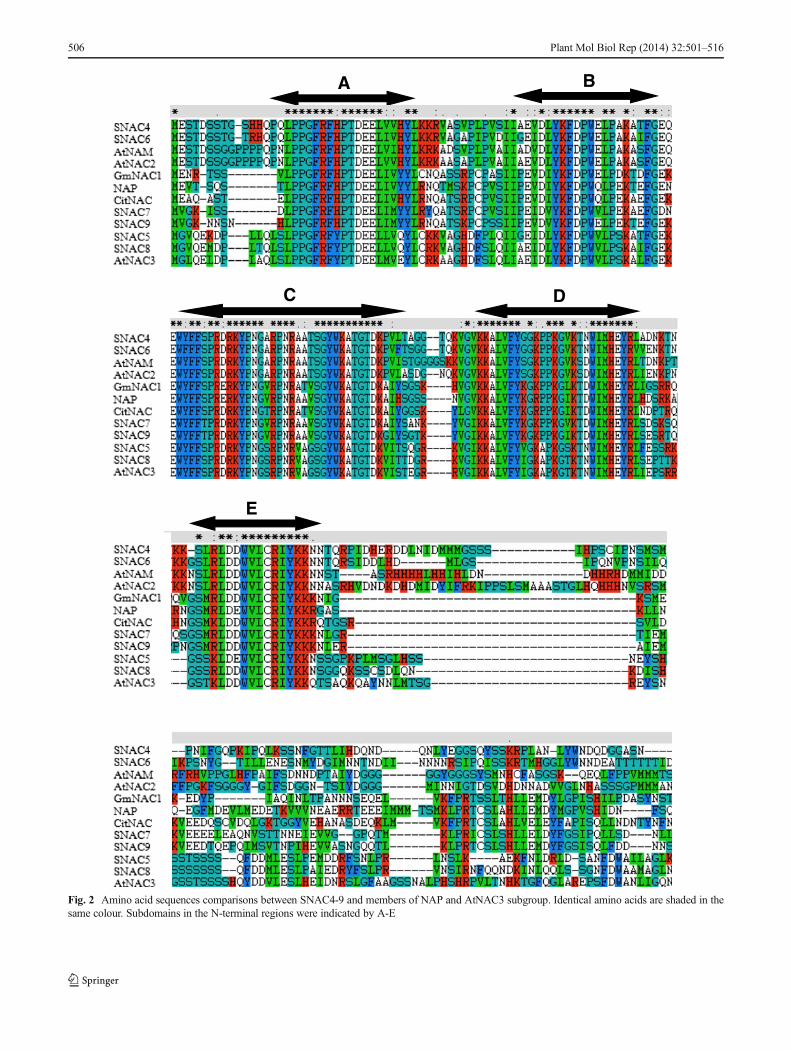
The amino acid alignments of SNAC4, 5, 6, 7, 8 and 9 andother members of the NAP and AtNAC3 subgroups are shownin Fig. 2. Higher sequence similarities were found in the N-terminus regions. The N-terminnus was composed of approxi-mately 150 amino acids and contained a conserved NAM do-main, which had five distinguishable subdomains (A–E). The C-terminal regions were divergent in amino acid composition, andcontained a number of simple amino acids, such as Ser, Thr, Proand Glu.
Features of SlNAC Genes
To examine the specific properties of SlNAC genes, we identifiedtheir genomic distributions and the structural features of eachgene using the SOL database (Supplementary Table 1). In total,74 genes were localized on 12 chromosomes with an unevendistribution (Fig. 3). The genes were present in different regions
of the chromosomes, including at the telomeric ends, around thecentromere, and in between the telomere and the centromere.Chromosome 6 had the largest number (13) of SlNAC genes,followed by eight genes on chromosomes 7 and 11. In contrast,only two genes were found on chromosome 9. Most of theSlNAC genes on chromosomes 2, 3, 6 and 7 were found onthe long arms of the chromosomes, whereas the SlNAC genes onchromosomes 4, 5, 8 and 7 were located at both ends.
The SlNAC genes could be classified into three typesaccording to their number indexes of exons/introns. The firsttype contained 48 genes with the number index 3/2. Thesecond type had six genes that had one or no intron, such asSlNAC12, 33 and 45 in the AtNAC063 subgroup. The thirdtype could be further divided into smaller subtypes with thenumber indexes 4/3, 5/4 and 6/5. The highest numbers of theexons and introns were found in SlNAC8 , which contained 17exons and 16 introns.
Features of SlNAC Proteins
To analyze the subcellular localizations, conserved domainsand motifs of SlNAC proteins, we used website-accessiblesoftware as described in materials and methods. As shown inSupplementary Table 1, most of the proteins were acidic, withPI values ranging from 4.58 to 7.0, and the highest observedPI value was 9.47. The instability indexes ranged from 24.28to 61.3, and most SlNAC proteins had an index higher than40. According to the amino acid indexes, the proteins wererich in Ser, Lys, and Asn.
The subcellular localization predictions suggested that ap-proximately half of the SlNACs were localized in the nucleus,whereas five, four, and three were localized in the peroxisome,cytosol, and chloroplast, respectively. In addition, subcellularlocalization predictions suggested the cytoskeletal localizationof SlNAC5, the mitochondrial localization of SlNAC29, andthat SlNAC11 and 37 could be located in either the nucleus orthe cytosol.
SlNAC proteins were divergent in their motif compositions.The SlNAC members all had one NAM domain as the charac-teristic conserved domain. Forty-four SlNAC proteins containedone or more low complexity region in the downstream region of
Table 1 Primer pairs used in thisresearch Gene Forward-Primer ( 5′-3′) Reverse-Primer (5′-3′) Tm (°C)
SNAC4 TGCCTCTGTTCCTCTTCCTG TCTTGTTCTCCAAATGTCGC 53
SNAC5 ATTCTCGCTGGGCTCAAAC GGAGGATGGGCGTAAACAT 53
SNAC6 TGTTGAGAACAACGAGGACG AGGAAATTGGCAATGGAGC 53
SNAC7 CTCTGATCTTCCTCCTGGATTT CAGGGATCGAACTTGTAGACAT 53
SNAC8 CTGGGAACTTCGATTGGGCT GTTTGATTTCCCGGCGTTGG 53
SNAC9 CCCTCCTGGATTTAGGTTTC CCAGGGATCGAACTTATAGACA 53
β-tublin CACGTGGGTCCAGCAATAC GGTCAGCAGCACACATCATGT 60
504 Plant Mol Biol Rep (2014) 32:501–516

the sequence. Some proteins in the TIP and NAC2 subgroupshad coiled coil regions. SlNAC21, 27, 64 and 65 in the ATAFand NAC2 subgroups included a transmembrane region. Inaddition, SlNAC68 had two internal repeat regions in sequence,and SlNAC37 had a VQ conserved domain.
Digital Expression Patterns of SlNAC Genes
To forecast the expression profiles of the SlNAC genes duringthe tomato development process, we acquired the digital ex-pression normalized (RPKM) data from a transcriptome anal-ysis of various tissues in the tomato cultivar Heinz and thewild relative Solanum pimpinellifolium from the TED data-base. As shown in Fig. 4, some members of the NAP, ATAF,AtNAC3, NAC2, TIP and TERN families, such as SlNAC14,20, 24, 28 and 61 were constitutively expressed in all of thetested organs. However, members of the VND, NAM andTERN subgroups were weakly expressed. SlNAC9, 30, 34,
35, 52 and 72 , which belong to subgroupVND, were stronglyexpressed in the roots, and SlNAC18 and 41 were stronglyexpressed in the flower buds and petals. Notably, there weretwo opposite expression patterns observed during fruit devel-opment. The first, which was observed for SlNAC3 , exhibitedan increase in expression during maturation. And the second,which was found for SlNAC20 , exhibited a decrease in ex-pression in the transition from the immature to mature stages.
Tissue-Specific Expression of SNAC Genes
Although the phylogenetic analysis provided important bio-informatics support for candidate genes selection, expressionanalysis is better for the further investigation of SNAC tran-scription factors. To elucidate the expression patterns ofSNAC genes, RT-qPCR analysis was performed (Fig. 5).The β-tubulin gene, which was used as an internal controlfor constitutive expression, was uniformly expressed in all of
Fig. 1 Evolutionary relationship of SlNAC and some homologous pro-teins in other plants. The multiple alignment was made using ClustalXand N-J tree was constructed with a 1000-bootstrap replication support.The subfamilies within the NAC family were grouped as indicated.Accession numbers of the reported NACs are as follows: ATAF1(X74755); ATAF2 (X74755); AtNAM (AF123311); NAP (AJ222713);AtNAC2 (AB049071); TIP (AF281062); CUC1 (AB049069); CUC2(AB002560); CUC3 (AF543194); AtNAC072 (At4g27410); AtNAC3(AB049070); NAC2 (AF201456); ANAC055 (At3g15500); ANAC019(At1g52890); ANAC011(EFH67253); BnNAC1-1 (AY245879);
BnNAC3 (AY245880); OsNAC1 (AB028180); OsNAC2 (AB028181);OsNAC3 (AB028182); OsNAC4 (AB028183); OsNAC5 (AB028184);OsNAC6 (AB028185); OsNAC7 (AB028186); OsNAC8 (AB028187);GRAB1 (AB028187); GRAB2 (AJ010830); NAM (X92205); ZmNAC1(ABY67929); SENU5 (Z75524); StNAC (AJ401151); GmNAC1(AY974349); GmNAC2 (AY974350); GmNAC3 (AY974351);GmNAC4 (AY974352); GmNAC5 (AY974353); GmNAC6(AY974354); CitNAC (EF185419); TERN (AB021178); CarNAC5(ACS94038); ANAC063 (AEE79353); VND1 (AEC06722); SND1(ABL67723); NTL8 (EFH57163)
Plant Mol Biol Rep (2014) 32:501–516 505
E
DC
BA
Fig. 2 Amino acid sequences comparisons between SNAC4-9 and members of NAP and AtNAC3 subgroup. Identical amino acids are shaded in thesame colour. Subdomains in the N-terminal regions were indicated by A-E
506 Plant Mol Biol Rep (2014) 32:501–516

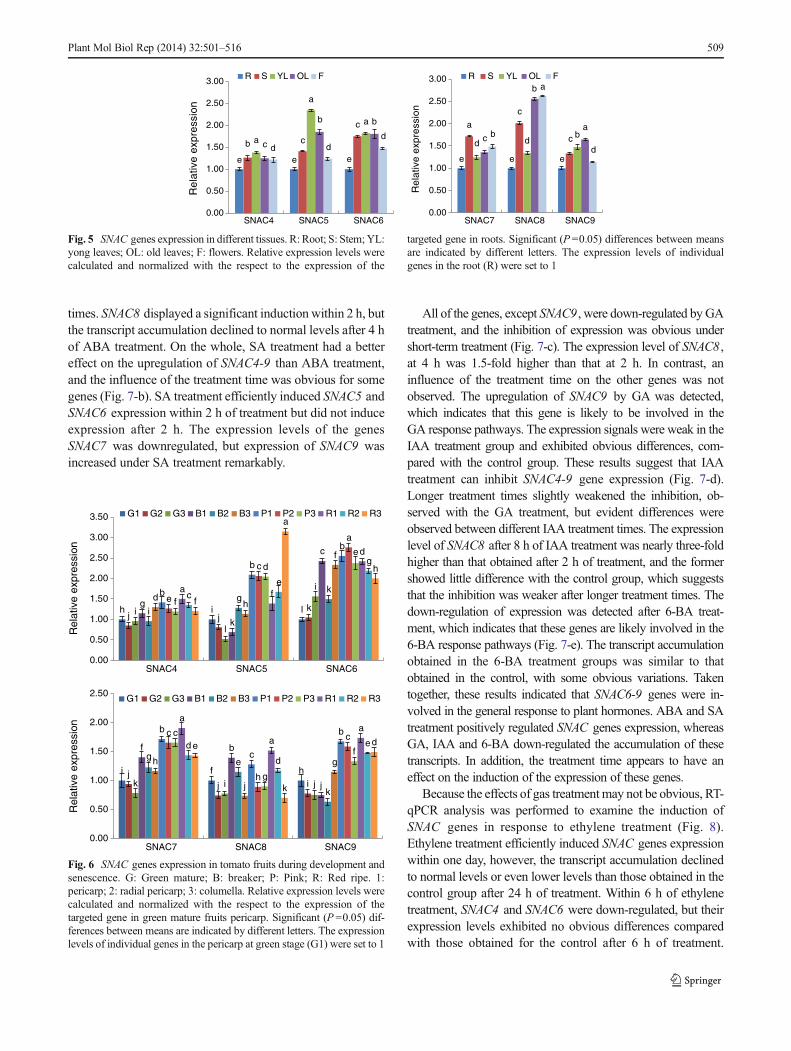
the tissues examined. Each SNAC gene showed a uniqueexpression pattern. SNAC4 mRNA was accumulated in allof the tissues at a similar expression level. SNAC5 wasexpressed strongly in the leaves but weakly expressed in theroots and flowers. Moreover, the expression level of SNAC5in young leaves was obviously higher than that obtained in oldleaves. The analysis of SNAC6 revealed a higher level ofexpression in the stems and leaves and, a relatively lowerexpression level in the roots. SNAC7 was predominantlyexpressed in the stems. The expression of SNAC8 was mostlyobserved in old leaves and flowers, but expression was alsoobserved at very low levels in the roots and young leaves,which is different from the expression pattern of SNAC5 ,although the two genes had a high sequence similarity. Theexpression pattern was consistent with the development.SNAC9 was constitutively expressed in all of the tissues,although a relatively higher level of expression was observedin old leaves.
The expression patterns of SNAC genes during fruit devel-opment were also studied (Fig. 6). For the expression analysis,the β-tubulin gene was used as an internal control. In tomatofruits, the expression level of SNAC4 increased with thematuration process and reached its highest expression levelsat the red ripe stage, which is consisten with the maturationprocess. The SNAC4 expression level was high in the colu-mella at the green mature and breaker stages, but was signif-icantly higher in the pericarp compared with the columella andthe radial pericarp late in the maturation process. The analysisof the expression of SNAC5 revealed that it is the expressed athigher levels at the pink stage in different parts of the fruits,with no obvious difference. Throughout development, theexpression of SNAC5 first decreased in the pericarp and thenincreased, and the lowest level was observed at the breakerstage. The SNAC5 expression in the columella presented aclear upward trend, and the highest levels were observed at the
red ripe stage. However, the obvious expression differencesbetween the columella and other regions during the sameperiod still require further investigation. On the whole, thehighest expression levels of SNAC6 were observed during thepink period, followed by the breaker stage. At the red ripestage, the tomato fruits progress from late-maturation to thesenescence process, and the expression signals for SNAC6slowly weakened. SNAC6 expression in the pericarp andradial pericarp was weak in the early stage and graduallyincreased with the development process. There was a promi-nent increase in the expression of SNAC7 in different regionsthroughout the during the entire development process. TheSNAC7 expression in the pericarp reached a maximum levelat the red ripe stage, whereas its expression in the radialpericarp and columella was low at the pink stage. Comparedwith the other genes, SNAC8 was expressed in the earliestdevelopmental stage. Fruits at the breaker stage and the redripe stage exhibited a higher SNAC8 expression level. Theexpression profile of SNAC9 was similar to that of SNAC7 ,and the levels in the pericarp were higher than those obtainedin other regions at similar stages.
Expression Profiles with Hormone Treatments
Plant hormones play an important role in the regulation of thedevelopment and maturation of fruits. To detect the impact ofhormones on the expression of the target genes (SNAC4-9 ),green mature fruit discs were treated with ABA, IAA, GA, 6-BA, SA, and ethylene.
Compared with the control group, SNAC4 was observed tobe down-regulated following ABA treatment (Fig. 7-a).SNAC5 were up-regulation during short-term treatment withABA for 2–6 h. ABA treatment promoted SNAC6 , SNAC7and SNAC9 genes expression during the 6–8 h, and the up-regulation of this gene increased with extended treatment
Fig. 3 Distributions of SlNAC genes on the 12 chromosomes in tomato. Chromosome numbers were indicated at the top of each bar. The scale on theleft was in megabases (Mb)
Plant Mol Biol Rep (2014) 32:501–516 507

Fig. 4 Heat map representation of tissue-specific expression of 74SlNAC genes. Color bar at top shows level of expression. Red indicatedexpressed genes and green indicated unexpressed ones. R=root; L=leaf;FB, F=flower buds and opened flowers; Fr (1)/ (2)/ (3)=1/2/3 cm fruit,respectively; MG=mature green fruit; Br and Br+10=fruits at breaker
and breaker+10 days stage. The above were samples of tomato cultivarHeinz . l=leaf; img=immature green fruit; brand br+5=fruit at breakerand breaker+5 days stage. The last four ones were samples of Solanumpimpinellifolium
508 Plant Mol Biol Rep (2014) 32:501–516
times. SNAC8 displayed a significant induction within 2 h, butthe transcript accumulation declined to normal levels after 4 hof ABA treatment. On the whole, SA treatment had a bettereffect on the upregulation of SNAC4-9 than ABA treatment,and the influence of the treatment time was obvious for somegenes (Fig. 7-b). SA treatment efficiently induced SNAC5 andSNAC6 expression within 2 h of treatment but did not induceexpression after 2 h. The expression levels of the genesSNAC7 was downregulated, but expression of SNAC9 wasincreased under SA treatment remarkably.
All of the genes, except SNAC9 , were down-regulated byGAtreatment, and the inhibition of expression was obvious undershort-term treatment (Fig. 7-c). The expression level of SNAC8 ,at 4 h was 1.5-fold higher than that at 2 h. In contrast, aninfluence of the treatment time on the other genes was notobserved. The upregulation of SNAC9 by GA was detected,which indicates that this gene is likely to be involved in theGA response pathways. The expression signals were weak in theIAA treatment group and exhibited obvious differences, com-pared with the control group. These results suggest that IAAtreatment can inhibit SNAC4-9 gene expression (Fig. 7-d).Longer treatment times slightly weakened the inhibition, ob-served with the GA treatment, but evident differences wereobserved between different IAA treatment times. The expressionlevel of SNAC8 after 8 h of IAA treatment was nearly three-foldhigher than that obtained after 2 h of treatment, and the formershowed little difference with the control group, which suggeststhat the inhibition was weaker after longer treatment times. Thedown-regulation of expression was detected after 6-BA treat-ment, which indicates that these genes are likely involved in the6-BA response pathways (Fig. 7-e). The transcript accumulationobtained in the 6-BA treatment groups was similar to thatobtained in the control, with some obvious variations. Takentogether, these results indicated that SNAC6-9 genes were in-volved in the general response to plant hormones. ABA and SAtreatment positively regulated SNAC genes expression, whereasGA, IAA and 6-BA down-regulated the accumulation of thesetranscripts. In addition, the treatment time appears to have aneffect on the induction of the expression of these genes.
Because the effects of gas treatment may not be obvious, RT-qPCR analysis was performed to examine the induction ofSNAC genes in response to ethylene treatment (Fig. 8).Ethylene treatment efficiently induced SNAC genes expressionwithin one day, however, the transcript accumulation declinedto normal levels or even lower levels than those obtained in thecontrol group after 24 h of treatment. Within 6 h of ethylenetreatment, SNAC4 and SNAC6 were down-regulated, but theirexpression levels exhibited no obvious differences comparedwith those obtained for the control after 6 h of treatment.
e e e
b c
c
a
a
a
c
b b
d d d
0.00
0.50
1.00
1.50
2.00
2.50
3.00
SNAC4 SNAC5 SNAC6
Rel
ativ
e ex
pres
sion
R S YL OL F
e e e
a
c
c d db c
b
a b
a
d
0.00
0.50
1.00
1.50
2.00
2.50
3.00
SNAC7 SNAC8 SNAC9
Rel
ativ
e ex
pres
sion
R S YL OL F
Fig. 5 SNAC genes expression in different tissues. R: Root; S: Stem;YL:yong leaves; OL: old leaves; F: flowers. Relative expression levels werecalculated and normalized with the respect to the expression of the
targeted gene in roots. Significant (P=0.05) differences between meansare indicated by different letters. The expression levels of individualgenes in the root (R) were set to 1
h i l j j
k i
l
i
g
k
c
i g
k d
h
f
b
b
b
e
c
a
f
d
e
a f
d
c e
g
f
a
h
0.00
0.50
1.00
1.50
2.00
2.50
3.00
3.50
SNAC4 SNAC5 SNAC6
Rel
ativ
e ex
pres
sion
G1 G2 G3 B1 B2 B3 P1 P2 P3 R1 R2 R3
i f h j j i k i j
f b
j
g e
k
h
j
g
b
c
b c
h
c c
g
f
a
a a
d
d
e e
k
d
0.00
0.50
1.00
1.50
2.00
2.50
SNAC7 SNAC8 SNAC9
Rel
ativ
e ex
pres
sion
G1 G2 G3 B1 B2 B3 P1 P2 P3 R1 R2 R3
Fig. 6 SNAC genes expression in tomato fruits during development andsenescence. G: Green mature; B: breaker; P: Pink; R: Red ripe. 1:pericarp; 2: radial pericarp; 3: columella. Relative expression levels werecalculated and normalized with the respect to the expression of thetargeted gene in green mature fruits pericarp. Significant (P =0.05) dif-ferences between means are indicated by different letters. The expressionlevels of individual genes in the pericarp at green stage (G1) were set to 1
Plant Mol Biol Rep (2014) 32:501–516 509

0.00
0.50
1.00
1.50
2.00
0h 2h 4h 6h 8h 10h 12h
Rel
ativ
e ex
pres
sion
SNAC4 SNAC5 SNAC6
0.00
0.50
1.00
1.50
2.00
0h 2h 4h 6h 8h 10h 12h
Rel
ativ
e ex
pres
sion
SNAC7 SNAC8 SNAC9
(a) ABA treatment
0.00
0.50
1.00
1.50
2.00
0h 2h 4h 6h 8h 10h 12h
Rel
ativ
e ex
pres
sion
SNAC4 SNAC5 SNAC6
0.00
0.50
1.00
1.50
2.00
0h 2h 4h 6h 8h 10h 12h
Rel
ativ
e ex
pres
sion
SNAC7 SNAC8 SNAC9
(b) SA treatment
0.00
0.50
1.00
1.50
0h 2h 4h 6h 8h 10h 12h
Rel
ativ
e ex
pres
sion
SNAC4 SNAC5 SNAC6
0.00
0.50
1.00
1.50
0h 2h 4h 6h 8h 10h 12h
Rel
ativ
e ex
pres
sion
SNAC7 SNAC8 SNAC9
(c) GA treatment
(e) 6-BA treatment
0.00
0.50
1.00
1.50
0h 2h 4h 6h 8h 10h 12h
Rel
ativ
e ex
pres
sion
SNAC4 SNAC5 SNAC6
0.00
0.50
1.00
1.50
0h 2h 4h 6h 8h 10h 12h
Rel
ativ
e ex
pres
sion
SNAC7 SNAC8 SNAC9
(d) IAA treatment
0.00
0.50
1.00
1.50
0h 2h 4h 6h 8h 10h 12h
Rel
ativ
e ex
pres
sion
SNAC4 SNAC5 SNAC6
0.00
0.50
1.00
1.50
0h 2h 4h 6h 8h 10h 12h
Rel
ativ
e ex
pres
sion
SNAC7 SNAC8 SNAC9
Fig. 7 Effects of exogenous planthormone (a) ABA, (b) SA, (c)GA, (d) IAA and (e) 6-BA onSNAC genes expression. Theexpression levels of individualgenes under 0 h treatment wereset to 1
510 Plant Mol Biol Rep (2014) 32:501–516

Remarkably, the expression peaks for SNAC4 and SNAC6were both detected 8 h after treatment, but the transcript accu-mulations decreased with longer treatment times. The SNAC5and SNAC7-9 genes presented a similar expression pattern,which showed that the ethylene regulation of SNAC genespresented fluctuations with the extension of the treatment time,as shown in Fig. 8. SNAC5 and SNAC7-9 were expressed athigh levels after 2 h, 6 h, and 12 h of treatment, and at normal orlower levels after 4 h, 8 h and 10 h of treatment. Taken together,these results suggest that short-term treatment with ethylene canpromote gene expression, but this effect was suppressed after24 h of treatment.
Discussion
SlNAC Genes May Be Involved in Diverse PhysiologicalProcesses in Tomato
The NAC family is a newly discovered transcription factorfamilly, with a variety of biological functions. NAC genes
have been found in approximately 20 species such asArabidopsis thaliana, Oryza sativa , Triticum aestivum , Zeamays , Solanum tuberosum , Cucurbita moschata , Glycine max ,andCitrus sinensis. It has been speculated that there may be over100 NAC genes in tomato (Han et al. 2012). Through bioinfor-matics methods, we conducted a genome-wide inventory of theNAC family of transcription factors in tomato. Similar workshave been performed by other groups, and the numbers of NACgenes identified differ (http://planttfdb_v1.cbi.pku.edu.cn:9010/web/family_view.php?sp=le&fn=NAC&listall=0; http://bioinfo.bti.cornell.edu/cgi-bin/itak/family_gene_list.cgi?acc=NAC&org=Tomato). We identified 74 SlNAC genes from thetomato genome using an e-value of 1e-10 and classfied them into12 subgroups in accordance with Ooka’s nomenclature (Ookaet al. 2003).
As shown in Fig. 1, SlNAC20 belonged to the ATAFsubgroup, which participates in the response to biotic andabiotic stress (Ohnishi et al. 2005). The NAM subgroupincludes the known NAM and CUC3 proteins and otherproteins with high homology. SlNAC46 was clustered as amember of the NAM subgroup, whose members are involved
0
0.5
1
1.5
2
2.5
0 2 4 6 8 10 12 24 48
Rel
ativ
e ex
pres
sion
Hours after treatment (h)
SNAC4
C2H4 Control
0
0.2
0.4
0.6
0.8
1
0 2 4 6 8 10 12 24 48
Rel
ativ
e ex
pres
sion
Hours after treatment (h)
SNAC6
C2H4 Control
0
1
2
3
4
5
0 2 4 6 8 10 12 24 48
Rel
ativ
e ex
pres
sion
Hours after treatment (h)
SNAC8C2H4 Control
0
3
6
9
12
0 2 4 6 8 10 12 24 48
Rel
ativ
e ex
pres
sion
Hours after treatment (h)
SNAC9C2H4 Control
0
5
10
15
20
25
0 2 4 6 8 10 12 24 48
Rel
ativ
e ex
pres
sion
Hours after treatment (h)
SNAC7
C2H4 Control
0
4
8
12
16
20
24
0 2 4 6 8 10 12 24 48
Rel
ativ
e ex
pres
sion
Hours after treatment (h)
SNAC5
C2H4 ControlFig. 8 Effects of exogenousethylene on SNAC genesexpression. X-ais represents thehours after 100 μL/L C2H4 andcontrol treatment on AC tomatofruits at greent mature stage. Theexpression levels of individualgenes under 0 h treatment wereset to 1
Plant Mol Biol Rep (2014) 32:501–516 511

in maintenance of shoot meristems and the separation ofcotyledons during the developmental process (Olsen et al.2005; Guilherme et al. 2009). SlNAC49 fell into the NAC1subgroup, which has been shown to be related to the develop-ment of lateral root (Meng et al. 2007). SlNAC69 was a memberof the TERN subgroup, and themembers of the TERN group areinduced by elicitors of the pathogen response (Ooka et al. 2003).SND1 inArabidopsis is themain switch for fiber secondary walldevelopment, and the VND1-7 genes are related to vascularbundle development. The SlNACs related to SND1 and VND1were predicted to be involved in plant secondary metabolismprocesses (Kubo et al. 2005; Zhong and Ye 2007b; Yamaguchiet al. 2008). The TIP subgroup contains the known membersNTL and TIP. TIP acts as a positive regulator of defense re-sponses. The TIP subgroup members can influence seed germi-nation during the response to external stimuli (Kim et al. 2008;Yoshii et al. 2010). SENU5 was a subgroup of unknown NACproteins from tomato and is related to leaf senescence. In addi-tion, the SlNACs in other subgroups have different functions,and some have not yet been functionally characterized. TheNAPsubgroup includes AtNAP from Arabidopsis , and has beenshown to be involved in the development of leaf, siliques, andother organs and senescence (He and Gan 2002; Guo and Gan2006; Uauy et al. 2006; Kou et al. 2012a, b). The AtNAC3subgroup includes AtNAC019, 055 and 072, which are inducedby ABA and drought stress and potentially enhance plant toler-ance and development (Tran et al. 2004; Bu et al. 2008; Causieret al. 2012). As a class of important transcriptional factors, theNAC proteins in the NAP and AtNAC3 subgroups have beenproposed to play vital roles during tomato development.According to the analysis conducted in the present study, wetargeted SNAC4-9 as candidate genes for further research. Insummary, the NAC transcription factors in tomato may playdivergent roles in plant development, signal transduction andresponses to stress.
The Features of Gene and Protein Sequences May Relateto Gene Functions
The features of the SlNAC genes were systematically analyzed.The alignment analysis results were consistent with previousreports (Ooka et al. 2003; Olsen et al. 2005). In the present report(Fig. 2), the N-terminals were conserved with five subdomains.Subdomains A, C and D were more conserved, and might beresponsible for binding DNA and other proteins (Ernst et al.2004). In contrast, the C-terminals were divergent in compositionand length and act as a transcription activators or repressorsduring transcriptional regulation (Kim et al. 2007a).
The genomic distribution data suggested that more SlNACgenes were localized on chromosomes 6, 7 and 11, particular-ly at both ends (Fig. 3). These results indicate that thesechromosomes are more related to the NAC family, but therewas no clear association between the gene classification and
chromosome location. According to a previous report, theNAC genes of Arabidopsis mostly have three exons and twointrons. The first two exons encode the conserved NAC do-main and the third one is responsible for the transcriptionactivation domain (Duval et al. 2002). Based on the exonand intron number (Supplementary Table 1), the SlNAC genescan be divided into several subfamilies, and most genescontained three exons and two introns, which is in accordancewith the features reported for NAC genes. However, therewere several genes with number indexes of 4/3, 5/4 and 6/5,and the highest numbers of exons and introns found inSlNAC8 , which contained 17 exons and 16 introns. There isevidence showing that the rate of intron gain is lower than theloss rate after segmental duplication, but the mechanisms arenot yet clear (Lin et al. 2006; Nuruzzaman et al. 2008;Nuruzzaman et al. 2010). Based on this evidence, the genesin each subgroup containing more introns may be the originalgenes of the group.
The protein features were analyzed as shown inSupplementary Table 1. The instability index is a measure ofthe protein stability. Most of the SlNAC proteins had instabil-ity index values higher than 40, which suggests that theseproteins are unstable. The amino acid index refers to theamino acid contents. Most of the SlNAC proteins were richin Ser, Lys and Asn, and these amino acids were mostlydistributed in the middle and lower reaches of the proteinsequences. This distribution of amino acids may be an impor-tant component for the constitution of a specific domainstructure in the C-terminals of the proteins. The averagehydrophobic values of the SlNAC proteins were negative,which indicates that these proteins are hydrophilic (data notshown). The subcellular localization prediction wasperformed using the WoLF PSORT software. The subcellularlocalization demonstrated that the subcellular localizations ofthe SlNAC proteins are divergent and complex, but mostSlNAC proteins were found to be nuclear proteins. The dif-ferences in the subcellular localizationmay playmultiple rolesin tomato cells (Li et al. 2010).
All of the SlNAC members had one NAM domain as thecharacteristic conserved domain but were divergent in thecompositions of their motifs. Forty-four SlNAC proteins hadone or more low complexity regions in the downstream regionof the sequence, which also had a higher instability index thanthe other regions. These regions were abundant in Ser, Asnand other simple amino acids and are thought to play roles astranscriptional activation regions (TARs). The coiled coilregions observed for some of the proteins may be relevant tothe higher structures of those proteins. The proteins belongingto the ATAF and NAC2 subgroups contained a transmem-brane region. To validate these results, the TMHMM softwarewas used. The exp number of AAs in TMHs (En) is animportant parameter and refers to the expected number ofamino acids number in the transmembrane helical structure.
512 Plant Mol Biol Rep (2014) 32:501–516

In general, if the index is higher than 18, the protein is likely atransmembrane protein or contain signal peptide structures.Through certification, the indexes of SlNAC21, /27, /64, and /65 were found to be 19.770, 21.377, 20.407 and 22.830,respectively. The first three were confirmed to have a trans-membrane helical structure in the C-terminus, but that ofSlNAC65 was in the N-terminus. The exp number, first 60AAs (Ef) is another important parameter. The index found forSlNAC65 was greater than 10 (22.767), which indicate thatthe so-called transmembrane helical structure in the N-terminus is a signal peptide.
Taken together, the various subcellular localizations andmultiple protein motifs may exert assignable impacts on theadvanced structures and functions of SlNAC proteins.
SlNAC Genes are Differentially Expressed During Plantand Fruit Development
Increasing evidence has shown that NAC transcription factorsare involved in plant development and senescence throughtissue-specific expression (Guo and Gan 2006; Liu et al.2009; Huang et al. 2012). To forecast the expression profilesof SlNAC genes, we acquired digital expression normalized(RPKM) data (Fig. 4). RPKM (Reads per Kilo bases perMillion reads) is the most commonly used method for theestimation of the gene expression level and consider the influ-ence of both the sequencing depth and the gene length on theread count. The experiment was performed by the USDARobert W Holley Center and included Illumina RNA-Seqanalyses of the leaves, roots, flower buds, fully opened flowers,and 1-cm, 2-cm, 3-cm sized, mature green, breaker, and break-er+10 fruits of the tomato cultivar Heinz , and of the leaves,immature green, breaker, and breaker+5 fruits of Solanumpimpinellifolium . Due to variety differences and incompletemeasured data, the digital expression can only be used as areference, and tissues-specific analysis is indispensable.
SNAC4-9 genes were moderately expressed in differenttissues (Fig. 5). The RT-qPCR analysis revealed that SNAC4and SNAC6 may be involved in the development of stems andleaves. SNAC5 may play roles in the early development ofleaves, because it exhibits a higher expression level in youngleaves and may be restrained by the senescence process.SNAC8 and SNAC9 both regulated the senescence of leaves,and SNAC8 was found to be highly expressed in flowers, andthusmay be related to flower formation and development. TheSNAC8 expression pattern was different from that of SNAC5 ,although the two genes had a high sequence similarity. Thedifferent expression patterns of the SNAC genes suggest thattheir products likely played functionally diverse roles duringthe processes of tomato growth and development.
The ripening and development of tomato fruits refers to theperiod from 35 to 55 DAF, which includes the green mature,breaker, pink and red ripe stages. A coordinated expression
was observed among the six SNAC genes (Fig. 6).Interestingly, the expression levels were divergent in the peri-carp, radial pericarp and columella of fruits at similar stages.Brecht reported that the maturity of the different parts of thetomato fruit is also inconsistent (Brecht 1987). There was aprominent increase in the expression of SNAC7 throughoutthe development in different regions. The expressions in peri-carp reached a maximum level at the red ripe stage, whereasthose in the radial pericarp and columella ware low at the pinkstage. These results show that the regulation of SNAC7 in thepericarp was later than in the other tissues during develop-ment. In general, SNAC4-9 , which belong to the NAP andAtNAC3 subgroups, were found to be involved in the regu-lation of tomato fruit development. These results indicate thatthe phylogenetic analysis is reliable to a certain degree.SNAC4, SNAC6 and SNAC7 showed similar expression pat-terns during the greenmature and breaker stages, but exhibitedsignificant differences at the pink and red ripe stages. Basedon the overall similarity of the expression patterns of theSlNAC genes, it is tempting to speculate that these genesparticipate in the same or similar regulatory networks.Currently, we do not know the exact roles of SlNAC genesin fruit ripening. However, it is possible that the proteins areinvolved in ethylene-stimulated or hormone signalingpathways.
SlNAC Genes Are Involved in the Response to Hormone
The hormone levels in vivo and hormone treatment in vitroplay crucial roles during fruit and vegetable maturation pro-cesses. Hormones also affect the ripening-related gene expres-sion, ethylene production, and the effects of ethylene. Theregulation of plant hormones during plant growth, fruit ripen-ing and senescence is a complex process that depends not onlyon hormone concentrations, but also on the balance and inter-action between different hormones. Previous studies havemostly focused on the effect of hormones on the regulatingof the stress response by NAC genes (Shinozaki andYamaguchi-Shinozaki 2000; 2007; Peng et al. 2009; Zhanget al. 2012; Han et al. 2012), and few studies have examinedthe effect of hormones on the fruit ripening mechanismsmediated by NAC genes. The present experiment examinedthe effects of 0.1 mM ABA, 0.l mM GA, 0.l mM IAA,0.1 mM 6-BA and 1 mM SA treatment on AC green maturetomato fruit tissue discs to analyze the roles of SNAC4-9 geneexpression on the regulation of maturity.
Abscisic acid (ABA) is an important factor in the regula-tion of fruit ripening and senescence as well as ethylene,which is effective for fruit ripening, abiotic stresses and otheraspects. ABA has close ties to ethylene generation (Josphet al. 1990), and may be an endogenous regulatory factorlocated upstream of ethylene during the apple ripening processto promote ethylene production. It has been reported that the
Plant Mol Biol Rep (2014) 32:501–516 513

ABA levels increase during fruit ripening, and that ABA canstimulate endogenous ethylene synthesis, which indicates thatthere is cross-talk between ABA and ethylene (Beaudoin et al.2000; Carbonell-Bejerano et al. 2010; Zhang et al. 2012).Recently, it was shown that ABA treatment can promoterespiration and ethylene production in WT siliques, but hasno effect on the atnap mutant siliques, which indicates thatABA may regulate ethylene through NAC genes (Kou et al.2012a, b). The results of the present study showed that theSNAC5-9 genes can promote the mature development of fruitthrough the ABA pathway (Fig. 7-a).
Salicylic acid (SA) is a plant growth regulator involved ingrowth, development, maturation, senescence, stress resis-tance induction and other metabolic processes (Manoj andSrivastava 2000). At present, the mechanisms of SA in fruitripening, softening and ripening-related gene expression reg-ulation have not been well investigated. In the present study,treatment with 1 mM SA significantly promoted NAC geneexpression in AC tomato fruit tissue discs, and the effect wasstronger than that of ABA (Fig. 7-b).
Gibberellic acid (GA) can play an important role in seedgermination, stem elongation, flower induction and seed for-mation throughout the entire life-cycle of higher plants.Studies have shown that GA can inhibit fruit chlorophylland peroxidase activity, and thereby delays strawberry fruitcoloring (Martinez et al. 1996). Treatment with 0.1 mM GAwas shown to inhibit SNAC gene expression in the presentexperiment, and this inhibition was weakened with the exten-sion of the treatment time (Fig. 7-c).
Similarly, indole acetic acid (IAA) can also play a regula-tory role in the plant growth and development stages. It hasbeen reported that the majority of gene expression is negativeregulated by IAA during the strawberry growth and matureaging processes (Civello et al. 1999). Bouzayen (2002) sug-gested that there is a signaling conversation between IAA andethylene in tomato ripening-related transcription regulation.IAA regulates plants metabolism, because exogenous auxincan promote ethylene production and after-ripening and reg-ulates both the tissue reaction to ethylene and the inhibitoryeffects of ripening (Kende and Zeevaart 1997). The roles ofIAA at high levels (100 to 1000 μM) and low levels (1 to10 μM) are different within the threshold value. A highconcentration of IAA can stimulate ethylene production, andmay promote after-ripening. In contrast, a low concentrationof IAA can inhibit ethylene production to delay ripening. Inthis experiment, treatment with IAA inhibited the gene ex-pression of the NAC genes, which can delay fruit maturation(Fig. 7-d).
6-Benzylaminopurine (6-BA) was the first artificially syn-thesized cytokinin, and can inhibit the decomposition of plantleaf chlorophyll, nucleic acids and protein to keep green fruitsimmature. However, the test crop types, usage, concentration,treatment time and location can affect the reactions of a plant
to 6-BA treatment. The 6-BA treatment in the present exper-iment showed an inhibitory effect on SNAC gene expression(Fig. 7-e).
It is widely accepted that ethylene is an important factorthat promote fruit and leave senescence (Oeller et al. 1991;Watkins 2002; Giovannoni 2004; Jing et al. 2005; Kou et al.2012a, b), but the link between ethylene and NAC transcrip-tion factors is barely understood. It has been reported thatexogenous ethylene treatment promotes the earlier occurrenceof the surge in respiration inWTsiliques, but the treatment didnot alter the respiration pattern in the atnap mutant siliques,which suggests that ethylene may regulate fruit developmentthrough NAC genes (Kou et al. 2012a, b). In this study, SNACgenes were up-regulated in response to short-time ethylenetreatment within 24 hours. In contrast, the induction declinedto normal or lower levels after 24 h of treatment (Fig. 8).These data indicate that SlNAC genes are involved in ethylenepathways during fruit senescence, but further analysis isrequired.
Acknowledgements This work was supported by the National NaturalScience Foundation of China (project no. 31171769) and PostdoctoralScience Founding Special Foundation Project of China (project no.201003300)
Conflict of Interest The authors have declared no conflict of interest.
Reference
Aida M, Ishida T, Fukaki H, Fujisawa H, Tasaka M (1997) Genesinvolved in organ separation in Arabidopsis : an analysis of thecup-shaped cotyledon mutant. Plant Cell 9:841–857
Alexander L, Grierson D (2002) Ethylene biosynthesis and actionin tomato: a model for climacteric fruit ripening. J Exp Bot53:2039–2055
Beaudoin N, Serizet C, Gosti (2000) Interactions between abscisicacid and ethylene signaling acscades. The Plant Cell 12:1103–1115
BouzayenM (2002) Ripening- associated transcriptional regulation in thetomato. A case of cross- talk between ethylene and auxin.Comparative Biochemistry and Physiology, Part A: Molecular andIntegrative. Physiology 132:S97
Brecht JK (1987) Locular gel formation in developing tomato fruit andinitiation of ethylene production. Hortscience 22(3):476–479
Bu Q, Jiang H, Li CB, Zhai Q, Zhang J, Wu X, Sun J, Xie Q, Li C (2008)Role of the Arabidopsis thaliana NAC transcription factorsANAC019 and ANAC055 in regulating jasmonic acid-signaleddefense responses. Cell Res 18:756–767
Carbonell-Bejerano P, Urbez C, Carbonell J, Granell A, Perez-Amador MA (2010) A fertilization-independent develop-mental program triggers partial fruit development and se-nescence processes in pistills of Arabidopsis . Plant Physiol154:163–172
Causier B, Ashworth M, Guo W, Davies B (2012) The TOPLESSinteractome: a framework for gene repression in Arabidopsis .Plant Physiol 158:423–438
Civello PM, Powell AL, Sabehat A (1999) An expansion gene expressedin ripening strawberry fruit. Plant Physiol 121:1273–1279
514 Plant Mol Biol Rep (2014) 32:501–516

Duval M, Hsieh TF, Kim SY, Thomas TL (2002) Molecular characteri-zation of AtNAM: a member of the Arabidopsis NACdomainsuperfamily. Plant Mol Biol 50:237–248
Ernst HA, Olsen AN, Skriver K, Larsen S, Lo Leggio L (2004) Structureof the conserved domain of ANAC, a member of the NAC family oftranscription factors. EMBO Rep 5:297–303
Fang Y, You J, Xie K, Xie W, Xiong L (2008) Systematic sequenceanalysis and identification of tissue-specificor stress-responsivegenes of NAC transcription factor family in rice. Mol GenGenomics 280:547–563
Fujita M, Fujita Y, Maruyama K, Seki M, Hiratsu K, Ohme-Takagi M,Tran LS, Yamaguchi-Shinozaki K, Shinozaki K (2004) Adehydration-induced NAC protein, RD26, is involved in a novelABA-dependent stress- signaling pathway. Plant J 39:863–876
Giovannoni JJ (2004) Genetic regulation of fruit development and ripen-ing. Plant Cell Online 16(suppl 1):S170–S180
Guilherme LP, Carolina SM, Maximiller DC, Pedro AR, Murilo SA,ClaudineMC, LucianoGF, Elizabeth PF (2009) Complete inventoryof soybean NAC transcription factors: sequence conservation andexpression analysis uncover their distinct roles in stress response.Gene 444:10–23
Guo YF, Gan SS (2006) AtNAP, a NAC family transcription factor, hasan important role in leaf senescence. Plant J 46:601–612
Han QQ, Zhang JH, Li HX, Luo ZD, KhurramZiaf WTT, Ye ZB (2012)Identification and expression pattern of one stress-responsive NACgene from Solanum lycopersicum . Mol Biol Rep 39:1713–1720
He XJ, Mu RL, Cao WH, Zhang ZG, Zhang JS, Chen SY (2005)AtNAC2, a transcription factor downstream of ethylene and auxinsignaling pathways, is involved in salt stress response and lateralroot development. Plant J 44:903–916
He YH, Gan SS (2002) A gene encoding an acyl hydrolase is involved inleaf senescence in Arabidopsis . Plant Cell 14:805–815
Hu HH, Dai MQ, Yao JL, Xiao BZ, Li XH, Zhang QF, Xiong LZ (2006)Overexpressing a NAM, ATAF, and CUC (NAC) transcriptionfactor enhances drought resistance and salt tolerance in rice. ProcNatl Acad Sci 103:12987–12992
Hu HH, You J, Fang YJ, Zhu XY, Qi ZY, Xiong LZ (2008)Characterization of transcription factor gene SNAC2 conferring coldand salt tolerance in rice. Plant Mol Biol 67:169–181
Huang H, Wang Y, Wang S, Wu X, Yang K, Niu Y, Dai SL (2012)Transcriptome-wide survey and expression analysis of stress-responsive NAC genes in Chrysanthemum lavandulifolium . PlantSci 193:18–27
International Tomato Genome Sequencing Project (2012) The tomatogenome sequence provides insights into fleshy fruit evolution.Nature 485:635–641
Jeong JS, Kim YS, Baek KH, Hairn J, Ha SH, Choi YD, Minkyun K,Reuzeau C, Kim JK (2010) Root-specific expression of OsNAC10improves drought tolerance and grain yield in rice under fielddrought conditions. Plant Physiologyl 153:185–197
Jing HC, Schippers JHM, Hille J, Dijkwel PP (2005) Ethylene-inducedleaf senescence depends on age-related changes and OLD genes inArabidopsis. J Exp Bot 56:2915–2923
Josph R, Eliahu G, Yang SF (1990) Characterization of abscisic acid-induced ethylene production in citrus leaf and tomato fruit tissues.Plant Physiol 92:48–53
Kende H, Zeevaart JAD (1997) The five classical plant hormones. PlantCell 9:1197–1210
Kim SG, Kim SY, Park CM (2007a) A membrane-associated NACtranscription factor regulates salt-responsive flowering viaFLOWERING LOCUS T in Arabidopsis . Planta 226:647–654
Kim SG, Lee AK, Yoon HK, Park CM (2008) A membrane bound NACtranscription factor NTL8 regulates gibberellic acid mediated salt sig-naling in Arabidopsis seed germination. Plant Journal 55(1):77–88
Kim SY, Kim SG, Kim YS, Seo PJ, Bae M, Yoon HK, Park CM (2007b)Exploring membrane-associated NAC transcription factors in
Arabidopsis: implications for membrane biology in genome regu-lation. Nucleic Acids Res 35:203–213
KimYS, Kim SG, Park JE (2006) Amembrane-bound NAC transcriptionfactor regulates cell division in Arabidopsis . Plant Cell 18:3132–3144
Klee HJ, Giovannoni JJ (2011) Genetics and control of tomato fruitripening and quality attributes. Annu Rev Genet 45:41–59
Ko JH, Yang SH, Park AH, Lerouxel O, Han KH (2007) ANAC012, amember of the plant-specific NAC transcription factor family, neg-atively regulates xylary fiber development in Arabidopsis thaliana .Plant J 50:1035–1048
Kou XH,Watkins CB, Gan SS (2012)Arabidopsis AtNAP regulates fruitsenescence. J Exp Bot 63:6139–6147
Kubo M, Udagawa M, Nishikubo N, Horiguchi G, Yamaguchi M, Ito J,Mimura T, Fukuda H, Demura T (2005) Transcription switches forprotoxylem and metaxylem vessel formation. Genes Dev 19:1855–1860
Li LB, Zhang YR, Liu KC, Ni ZF, Fang ZJ, Sun QX, Gao JW (2010)Identification and bioinformatics analysis of SnRK2 and CIPKfamily genes in sorghum. Agric Sci China 9:19–30
Lin HN, ZhuW, Silva CJ, Gu X, Buell CR (2006) Intron gain and loss insegmentally duplicated genes in rice. Genome Biol 7:R41
Lin R, Zhao W, Meng X, Wang M, Peng Y (2007) Rice gene OsNAC19encodes a novel NAC-domain transcription factor and responds toinfection byMagnaporthe grisea. Plant Sci 172:120–130
Liu YZ, BaigMN, Fan R, Ye JL, Cao YC, DengXX (2009) Identificationand expression pattern of a novel NAM, ATAF, and CUC-like genefrom Citrus sinensis Osbeck . Plant Mol Biol Rep 27:292–297
Manoj K, Srivastava UN (2000) Delayed ripening of banana fruit bysalicylic acid. Plant Sci 158:87–96
Mao C,DingW,WuY, Yu J, He X, ShouH,Wu P (2007) Overexpressionof a NAC-domain protein promotes shoot branching in rice. NewPhytol 176:288–298
Martinez GA, Chaves AR, Anon MC (1996) Effect of exogenous appli-cation of gibberellic acid on color change and phenylalanineammonia-lyase, chlorophyllase, and peroxidase activities duringripening of strawberry fruit (Fragaria×ananassa Duch.). J PlantGrowth Regul 15:139–146
Meng Q, Zhang C, Gai J, Yu D (2007) Molecular cloning, sequencecharacterization and tissue-specific expression of six NAC-likegenes in soybean (Glycine max (L.) Merr.). J Plant Physiol 164:1002–1012
Nuruzzaman M, Gupta M, Zhang CJ, Wang L, Xie WB, Xiong LZ,Zhang QF, Lian XM (2008) Sequence and expression analysis ofthe thioredoxin protein gene family in rice. Mol Gen Genomics 280:139–151
Nuruzzaman M, Manimekalai R, Sharoni AM, Satoh K, Kondoh H,Ooka H, Kikuchi S (2010) Genome-wide analysis of NAC tran-scription factor family in rice. Gene 465:30–44
Oeller PW, Lu MW, Taylor LP, Pike DA, Theologis A (1991) Reversibleinhibition of tomato fruit senescence by antisense RNA. Science254:437–439
Ogo Y, Kobayashi T, Nakanishi-Itai R, Nakanishi H, Kakei Y, TakahashiM, Toki S, Mori S, Nishizawa NK (2008) A novel NAC transcrip-tion factor, IDEF2, that recognizes the iron deficiency-responsiveelement 2 regulates the genes involved in iron homeostasis in plants.J Biol Chem 283:13407–13417
Ohnishi T, Sugahara S, Yamada T, Kikuchi K, Yoshiba Y, Hirano HY,Tsutsumi N (2005) OsNAC6, a member of the NAC gene family, isinduced by various stresses in rice. Gene Genet Syst 80:135–139
Olsen AN, Ernst HA, Leggio LL (2005) NAC transcription factors:structurally distinct, functionally diverse. Trends Plant Sci 10:79–87
OokaH, SatohK,Doi K, Nagata T, OtomoY,MurakamiK,Matsubara K,Osato N, Kawai J, Carninci P (2003) Comprehensive analysis ofNAC family genes inOryza sativa and Arabidopsis thaliana. DNARes 10:239–247
Plant Mol Biol Rep (2014) 32:501–516 515

Peng H, Cheng HY, Yu XW, Shi QH, Zhang H, Li JG, Ma H (2009)Characterization of a chickpea (Cicerarietinum L.) NACfamilygene, CarNAC5, which is both developmentally- and stress-regulated. Plant Physiol Biochem 47:1037–1045
Sablowski RW, Meyerowitz EM (1998) A homolog of NO APICALMERISTEM is an immediate target of the floral homeotic genesAPETALA3/PISTILLATA. Cell 92:93–103
Shinozaki K, Yamaguchi-Shinozaki K (2000) Molecular responses to dehy-dration and low temperature, differences and cross-talk between twostress signaling pathways. Curr Opin Plant Biol l3:217–223
Shinozaki K, Yamaguchi-Shinozaki K (2007) Gene networks in-volved in drought stress response and tolerance. J ExprimentalBot 58:221–227
Souer E, van Houwelingen A, Kloos D, Mol J, Koes R (1996) The NoApical Meristem gene of Petunia is required for pattern formation inembryos and flowers and is expressed at meristem and primordiaboundaries. Cell 85:159–170
Sperotto RA (2009) Identification of up-regulated genes in flag leavesduring rice grain filling and characterization of OsNAC5, a newABA-dependent transcription factor. Planta 230:985–1002
Tamura K, Dudley J, Nei M (2007) MEGA4: molecular evolutionarygenetics analysis (MEGA) software version4.0. Mol Biol Evol 24:1596–1599
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG(1997) The CLUSTAL-X windows interface: flexible strategies formultiple sequence alignment aided by quality analysis tools. NucleicAcids Res 25:4876–4882
Tran LS, Nakashima K, Sakuma Y, Simpson SD, Fujita Y, Maruyama K,Fujita M, Seki M, Shinozaki K, Yamaguchi-Shinozaki K (2004)Isolation and functional analysis of Arabidopsis stress-inducible
NAC transcription factors that bind to a drought-responsive cis-element in the earlyresponsive to dehydration stress 1 promoter.Plant Cell 16:2481–2498
Uauy C, Distelfeld A, Fahima T, Blechl A, Dubcovsky J (2006) A NACgene regulating senescence improves grain protein, zinc, and ironcontent in wheat. Science 314:1298–1301
Watkins CB (2002) Ethylene synthesis, mode of action, consequencesand control. In: Knee M (ed) Fruit quality and its biological basis.Sheffield Academic Press, Sheffield, pp 180–224
Weir I, Lu J, Cook H, Causier B, Schwarz SZ, Davies B (2004)CUPULIFORMIS establishes lateral organ boundaries inAntirrhinum . Development 131:915–922
Xie Q, Frugis G, Colgan D, Chua NH (2000) Arabidopsis NAC1 trans-duces auxin signal downstream of TIR1 to promote lateral rootdevelopment. Gene Dev 14:3024–3036
Yamaguchi M, Kubo M, Fukuda H, Demura T (2008) Vascular relatedNAC-domain7 is involved in the differentiation of all types of xylemvessels in Arabidopsis roots and shoots. Plant J 55:652–664
Yoshii M, Yamazaki M, Rakwal R, Kishi-Kaboshi M, Miyao A,Hirochika H (2010) The NAC transcription factor RIM1 of rice isa new regulator of jasmonate signaling. Plant J 61:804–815
Zhang K, Xia X, Zhang Y, Gan SS (2012) An ABA-regulated and Golgi-localized protein phosphatase controls water loss during leaf senes-cence in Arabidopsis. Plant J 69:667–678
Zhong RQ, Richardson EA, Ye ZH (2007b) Two NAC domain transcrip-tion factors, SND1 and NST1, function redundantly in regulation ofsecondary wall synthesis in fibers of Arabidopsis. Planta 225:1603–1611
Zhong RQ, Ye ZH (2007) Regulation of cell wall biosynthesis. Curr OpinPlant Biol 10:564–572
516 Plant Mol Biol Rep (2014) 32:501–516