migration neuronale et développement du système...
TRANSCRIPT

Migration neuronale et développement
du système nerveux
UE21/Master Sante/2010-2011Pr.A.AUTILLO-TOUATI
I Introduction : la migration neuronale dans le développement.
II De nouvelles techniques ont permis une meilleure compréhension de
la complexité de la migration neuronale.
III Migration radiale et neurones pyramidaux du cortex.
IV Migration tangentielle et inter neurones du cortex.
V Le mutant Reeler
VI Conclusion

I Introduction:la migration neuronale dans le développement.

Rappel des principales étapes du développement du SN
-Origine embryonnaire du SN
-origine commune: l’ectoderme
- trois sources distinctes de neurones: neuro-épithélium, la
crête neurale et les placodes (épaississements de l'ectoderme
rostral)
-Plusieurs stades
-Induction et Régionalisation axiale de la plaque neurale
-Neurulation / Tube neural : processus morphogénétique qui
donne le cerveau , la moelle épinière.
-Le SNP dérive de la crête neurale
-Les épithéliums olfactif et auditif dérivent des placodes
-Polarité dorso-ventrale et rostro caudale du tube neural

Plaque neurale Cerveau adulteAu niveau cellulaire: la neurogénèse
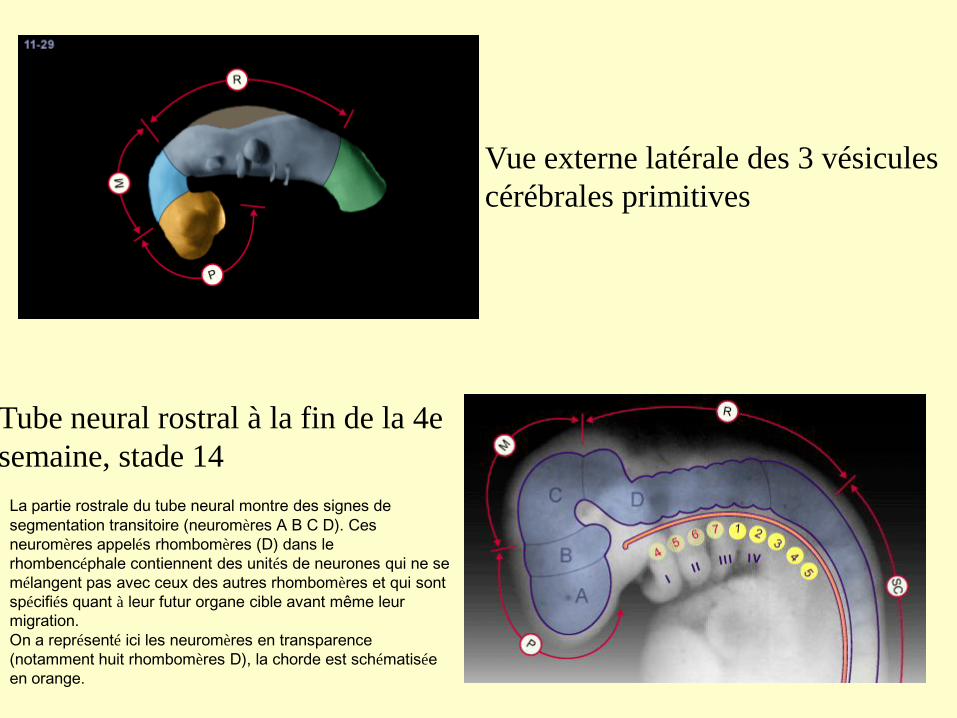
Vue externe latérale des 3 vésicules
cérébrales primitives
Tube neural rostral à la fin de la 4e
semaine, stade 14
La partie rostrale du tube neural montre des signes de
segmentation transitoire (neuromères A B C D). Ces
neuromères appelés rhombomères (D) dans le
rhombencéphale contiennent des unités de neurones qui ne se
mélangent pas avec ceux des autres rhombomères et qui sont
spécifiés quant à leur futur organe cible avant même leur
migration.
On a représenté ici les neuromères en transparence
(notamment huit rhombomères D), la chorde est schématisée
en orange.

Forebrain
(Prosencephalon)
Midbrain
(Mesencephalon)
Hindbrain
(Rhombencephalon)
Telencephalon
Diencephalon
Mesencephalon
Metencephalon
Myelencephalon
Olfactory lobes
Hippocampus
Cerebrum
Retina
Epithalamus
Thalamus
Hypothalamus
Midbrain
Cerebellum
Pons
Medulla
Neurogenin
bHLH
NeuroD
bHLH
Proneural genes Neural precursor genes Neurons
Determination Differentiation
Formation of the Brain
Neuron
‘Birthday’ –
final division
Hindbrain segmented
(rhombomeres) – isolated
‘territories’ of neurons
Inhibitory HLHs may
limit effects of proneural
genes to specific tissues

Eight Phases in Embryonic and Fetal
Development at a Cellular Level
1. Mitosis 2. Migration 3. Aggregation and
4. Differentiation
5. Synaptogenesis 6. Death 7. Rearrangement
8. Myelination

1. Mitosis/Proliferation
•Occurs in ventricular zone
•Rate can be 250,000/min
•After mitosis “daughter”
cells become fixed post
mitotic
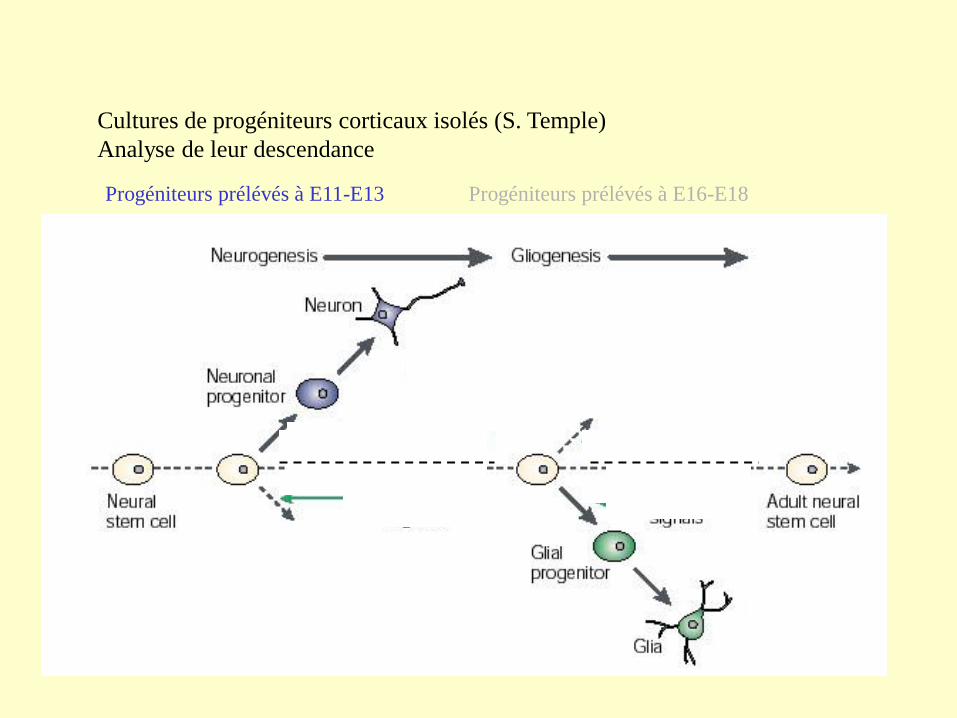
Cultures de progéniteurs corticaux isolés (S. Temple)
Analyse de leur descendance
Progéniteurs prélévés à E11-E13 Progéniteurs prélévés à E16-E18

Neurons Arise before Glia in E10 Cortical Stem Cell Clones
E10 stem cells were cultured in serum-free basal medium
supplemented with 10 ng/ml FGF2, and astrocyte-meningeal
cell conditioned medium. Under these culture conditions,
cortical stem cells generate neurons, astrocytes, and
oligodendrocytes

In Vivo, Cortical Neurons Differentiate during the Embryonic
Period and Glia Largely during the Postnatal Period
Neuron. 2000 Oct;28(1):1-3. Timing of CNS cell generation: a
programmed sequence of neuron and glial cell production from isolated
murine cortical stem cells. Qian X, Shen Q, Goderie SK, He W, Capela
A, Davis AA, Temple S.


2. Migration
Note that
differentiation is
going on as neurons
migrate.

2. Migration
Radial glial cells
act as guide
wires for the
migration of
neurons
Radial Glia

2. Migration
Growth Cones
Growth cones crawl forward as they
elaborate the axons training behind them.
Their extension is controlled by cues in
their outside environment that ultimately
direct them toward their appropriate
targets.
The fine threadlike
extensions shown in red
and green are filopodia,
which find adhesive
surfaces and pull the
growth cone and
therefore the growing
axon to the right.
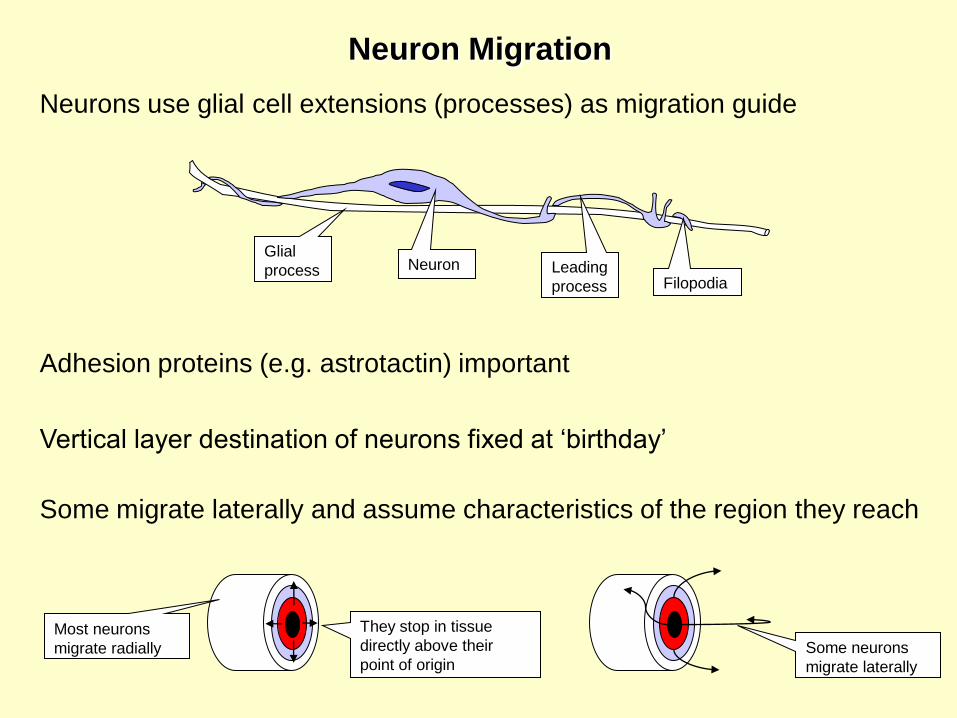
NeuronGlial
process Leading
process Filopodia
Neurons use glial cell extensions (processes) as migration guide
Adhesion proteins (e.g. astrotactin) important
Vertical layer destination of neurons fixed at ‘birthday’
Some migrate laterally and assume characteristics of the region they reach
Neuron Migration
Most neurons
migrate radially Some neurons
migrate laterally
They stop in tissue
directly above their
point of origin

II De nouvelles approches ont permis une meilleure
compréhension de la complexité de la migration neuronale.
-Vidéomicroscopie.
-Analyse de mutants murins.
-Identification de facteurs et de voies de signalisation
impliquées dans la migration.
-Electroporation et siRNA.
-Pathologies humaines de la migration.
-3 modèles: cortex cérébral, cortex cérébelleux, système visuel.

Comment s’organise le cortex cérébral?

Le développement du cortex cérébral des mammifères est une cascade d’événements
complexes comprenant deux neurogenèses successives.
La première survient précocement chez les foetus et permet la mise en place d’une pré
plaque corticale composée des premiers neurones post mitotiques, formant une couche
située entre la matrice proliférative ventriculaire et la surface méningée.
La plupart des cellules de la pré plaque disparaîtront lorsque l’ensemble des neurones
de la plaque corticale seront mis en place durant la seconde neurogenèse; la plaque
corticale est destinée à se transformer en substance grise du cortex définitif.
Lors du développement normal, la plaque corticale s’insère dans la pré plaque et isole
les cellules de Cajal-Retzius en une couche de neurones tangentiels exclusivement
situés dans la zone marginale (ZM).
Deux neurogenèses successives dans le cortex cérébral

-Migration neuronale au cours de la gestation entre E days 11-18
chez la souris et E weeks 10-20 chez l’homme.
-3 étapes:
-stade de la pré plaque (PP), migration de neurones
post mitotiques dans la PP à partir de la ZV
proliférative et des cellules de Cajal-Retzius dans la
zone marginale.
-stade de la plaque corticale (PC), migration radiale et
par vagues de neurones qui vont constituer les couches
2-6 du cortex .
-stade de maturation

III Migration radiale et neurones pyramidaux du cortex.
Les neurones pyramidaux dérivent du télencéphale dorsal

A: 4 étapes dans la migration des neurones pyramidaux du
cortex associées à des modifications morphologiques: migration
radiale initiale, arrêt dans la zone sub ventriculaire, migration
rétrograde, migration radiale secondaire.

Les 4 phases de migration
2 types de mouvement des
neurones corticaux:
-dans la corticogenèse précoce
-dans la corticogenèse tardive




Les Glies Radiaires sont des précurseurs astrocytaires?



Glial cells make intimate contact with synaptic terminals
Reconstruction 3D d’une cellule de la glie de Bergmann
Haydon Ph., Nature 2001, 186-196.

B:Les neurones les plus jeunes forment les couches les plus
superficielles.

Phénotypes des anomalies de
migration dans le cortex.

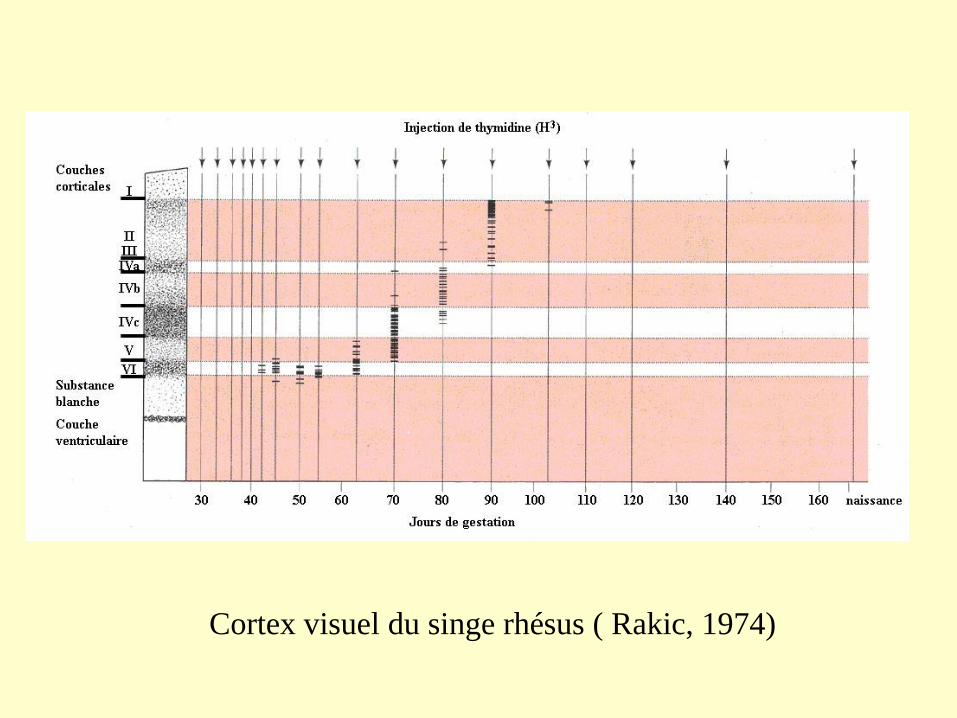
Cortex visuel du singe rhésus ( Rakic, 1974)

Détermination de l’identité laminaire dans le cerveau du furet
Précurseurs neuronaux jeunes
migrent dans la couche 6
Précurseurs neuronaux tardifs
migrent dans les couches 2/3

Expériences de transplantation de précurseurs neuronaux jeunes dans des
zones ventriculaires plus agées

Dans la plaque corticale la séquence de mise en place des neurones se réalise selon un
gradient intéro-externe de migration amenant les neurones derniers nés à occuper
toujours l’interface entre la couche marginale et la plaque corticale. Le cortex s’accroît
donc par sa périphérie mais est toujours contenu par les éléments de la couche
marginale qui forment un réseau neuronal perpendiculaire au vecteur d’épaississement
du cortex.
CONCLUSION

IV Migration tangentielle et inter neurones du cortex.
Kriegstein and Noctor,TINS, 2004, 27, 7,392-398
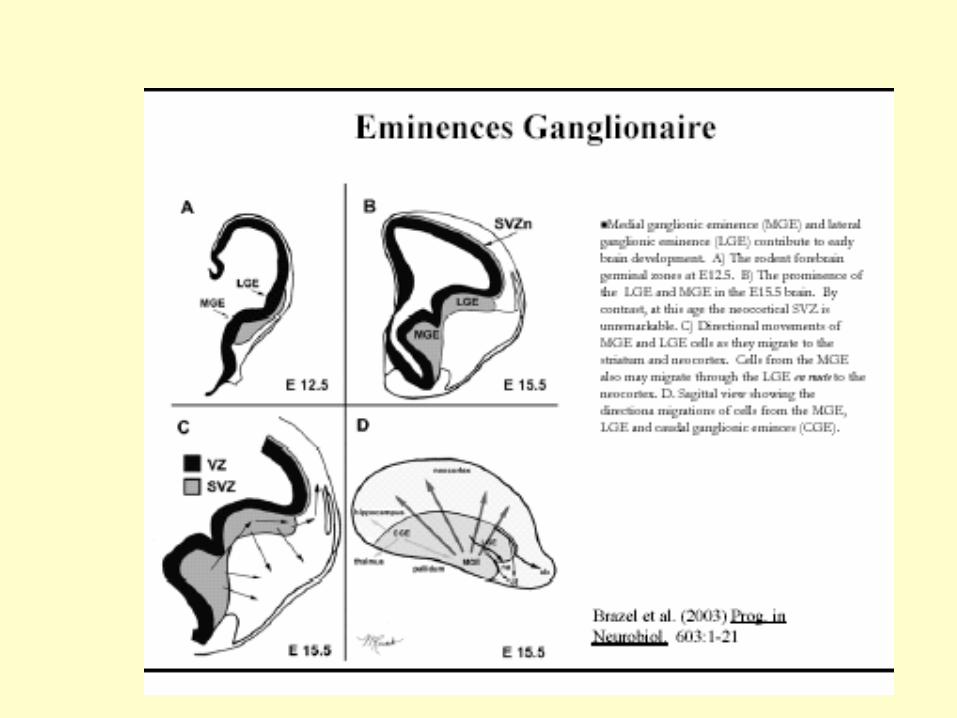

Les inter neurones corticaux dérivent du télencephale ventral et
migrent vers le télencéphale dorsal de façon tangentielle et
peuvent ensuite changer de direction pour rejoindre la PC
MGE: eminence ganglionnaire mediane
LGE: eminence ganglionnaire laterale


Des schémas différents de migration pour les inter neurones
et les neurones pyramidaux du cortex



En 1995, plusieurs groupes ont réussi à cloner le gène reelin (rln)
codant pour une protéine sécrétée de 400 kDa exclusivement
synthétisée dans le cortex par les cellules de Cajal-Retzius (cCR) .
Chez la souris normale, le développement en présence de reeline
aboutit à un cortex ordonné où la disposition des neurones
pyramidaux est essentiellement radiaire.
Chez la souris mutante homozygote reeler, en l’absence de reeline, le
cortex est désorganisé et l’orientation des neurones pyramidaux
devient aléatoire. La plaque corticale ne s’intercale plus dans la pré
plaque et le gradient de mise en place des neurones pyramidaux
apparaît relativement inversé (extéro-interne). De plus les cCR sont
situées en position sub normale et souvent collées contre la lame
basale méningée car la limitante gliale apparaît souvent perforée.
Effet répulsif de la reeline sur la migration neuronale
V Le mutant Reeler

Chez l'embryon de la souris homozygote reeler, la migration
des neurones se déroule normalement jusqu'au moment où
ceux-ci arrivent près de leur destination. La reeline n'étant pas
sécrétée dans la matrice extracellulaire par les cellules de Cajal-
Retzius, le gradient répulsif vis-à-vis des neurones de la plaque
corticale n'a pas lieu si bien que les cellules de la sous-plaque
sont comme repoussées vers l'extérieur en dehors de la plaque
corticale sous-jacent très désorganisée. Cela se traduit chez
l'animal très précocement par un cortex où les couches sont peu
apparentes : la plaque corticale ne s'intercale plus dans la
préplaque et le gradient de mise en place des neurones
pyramidaux se fait de manière quasiment inversée c'est-à-dire
selon un gradient extéro-interne.


Le cervelet du mutant homozygote est constitué
par l'emboîtement de deux structures : un cortex
cérébelleux atrophié et une masse cellulaire
centrale comportant la plupart des cellules de
Purkinje et des cellules des noyaux profonds. On
ne distingue aucune différence entre le témoin et
le mutant hétérozygote.
Technique utilisée : coloration à la thionine
phéniquée
Mutant Reeler

1 - Chez la souris normale
Dans le cortex cérébelleux, le rôle joué par les cellules de Cajal-Retzius dans le cortex cérébral en formation, serait joué par
les cellules granulaires externes. En effet, ces cellules sécréteraient la reeline dans la matrice extracellulaire qui aurait un rôle
répulsif dans la zone marginale repoussant la plaque des cellules de Purkinje à l'interface de la couche moléculaire et de la
couche granulaire.
2 - Chez le mutant homozygote reeler
Dans le cortex cérébelleux où la reeline est pourtant peu exprimée, la modification de la séquence nucléotidique de cette
protéine entraîne de grandes perturbation dans l'organisation du cortex cérébelleux chez le mutant homozygote. Les cellules
de Purkinje sont disposées de manière aléatoire vraisemblablement à cause d'un arrêt de leur migration si bien que le cervelet
paraît constitué de l'emboîtement de deux structures : à l'extérieur, un cortex cérébelleux dont l'architecture est semblable à
celle que l'on trouve chez l'animal normal mais très mince et à l'intérieur une masse cellulaire comportant la plupart des
cellules de Purkinje mélangées aux cellules des noyaux profonds.

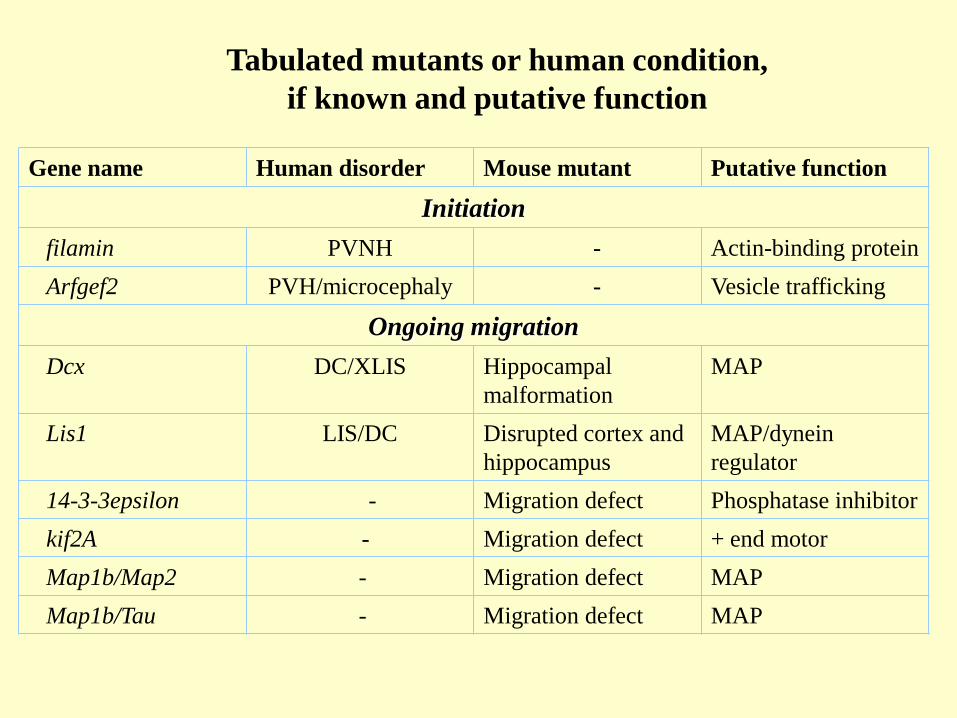
Gene name Human disorder Mouse mutant Putative function
Initiation
filamin PVNH - Actin-binding protein
Arfgef2 PVH/microcephaly - Vesicle trafficking
Ongoing migration
Dcx DC/XLIS Hippocampal
malformation
MAP
Lis1 LIS/DC Disrupted cortex and
hippocampus
MAP/dynein
regulator
14-3-3epsilon - Migration defect Phosphatase inhibitor
kif2A - Migration defect + end motor
Map1b/Map2 - Migration defect MAP
Map1b/Tau - Migration defect MAP
Tabulated mutants or human condition,
if known and putative function

Lamination
reelin LCH reeler Glycoprotein
dab1 - scambler Adaptor protein
Apoer2 - Inverted cortex Reelin receptor
Vldlr - Inverted cortex Reelin receptor
p35 - Inverted cortex Activator of cdk5
cdk5 - Inverted cortex Serine-threonine
kinase
Brn1/Brn2 - Inverted cortex Transcriptional
activation of cdk5
and dab1

Stop signal
Fak -- Disrupted migration Focal-adhesion
kinase
POMT1 Walker-Warburg
syndrome
-- -dystroglycan O-
linked glycosylation
POMGnT1 Muscle-eye-brain
disease
-- -dystroglucan O-
linked glycosylation
fukutin Fukuyamu MD Disrupted migration Phospholigand
transferase
Abbreviations: PVNH, periventricular nodular heterotopia; PVH, periventricular heterotopia;
DC/XLIS, double cortex/X-linked lissencephaly; LIS, lissencephaly; LCH, lissencephaly
cerebellar hypoplasia; MD, muscular dystrophy; , is not described. Other abbreviations are
indicated in the text.
Oligophrénine= GAP (G Activating Protein) de la famille Rho.
Désordres neurologiques et retard mental