midkine lacking its last 40 amino acids acts on...
TRANSCRIPT

Cancer Biology and Signal Transduction
Midkine Lacking Its Last 40 Amino Acids Acts onEndothelial and Neuroblastoma Tumor Cells andInhibits Tumor DevelopmentNoushin Dianat1, Barbara Le Viet1, Emilie Gobbo2, Nathalie Auger3, Ivan Bi�eche4,5,Annelise Bennaceur-Griscelli2,6, and Frank Griscelli2,3,5
Abstract
Midkine (MDK) is a member of a new family of neurotrophicfactors considered as rate-limiting growth and angiogenic fac-tors implicated in the onset, invasion, and metastatic process ofneuronal tumors, including neuroblastoma. We showed that allneuroblastoma cell lines highly expressed MDK, indicating thatit is a critical player in tumor development, which may hence-forth represent an attractive therapeutic target. We showed thatthe knockdown of MDK expression by siRNA led to a markedand significant decrease in neuroblastoma (IGR-N91 and SH-SY5Y) cell proliferation in vitro. Using a new strategy, we thenevaluated the antitumor effect of a truncated MDK protein,lacking the C-terminal 81–121 portion of the molecule(MDKD81-121), which may act as a dominant-negative effectorfor its mitogenic, angiogenic, and tumorigenic activities by
heterodimerizing with the wild-type protein. In vitro studiesshowed that MDKD81-121 selectively inhibited MDK-depen-dent tumor cells and was able to strongly reduce endothelialcell proliferation and migration and to induce ER stress–medi-ated apoptosis. We then investigated the effects of MDKD81-121 in vivo using electrotransfer of a plasmid encoding asecretable form of MDKD81-121 into tibialis cranialis musclesof nude mice. We showed that MDKD81-121 dramaticallyinhibited (up to 91%) tumor development and growth. Thisinhibition was correlated with the detection of the MDKD81-121 molecule in plasma and the suppression of intratumorneovascularization. Our findings demonstrate that MDK inhi-bition is a tractable therapeutic target for this lethal pediatricmalignancy. Mol Cancer Ther; 14(1); 213–24. �2014 AACR.
IntroductionOver the last decade, a number of new antiangiogenic agents
have demonstrated their ability to retard tumor growth, and someof them are physiologic angiostatic factors that are derived uponproteolytic cleavage of native proteins. In this field, we previouslyevaluated antiangiogenesis and antitumor activities of a 38-kDainternal fragment of plasminogen, a 24-kDa noncollagenousdomain of collagen type IV a-chains, and a 20-kDa fragment ofcollagen XVIII, called, respectively, angiostatin, canstatin, andendostatin (1–3). All these fragments are known to silence cyto-kine-induced angiogenesis pathways by binding to integrin recep-tors and triggering mitochondrial apoptosis (4–6). They specif-ically target endothelial cells, do not possess antineoplastic activ-ities and should therefore be administered in combination with
chemotherapy or radiotherapy to achieve a major clinicalresponse (4, 7, 8). Thus, the development of new anticancerstrategies targeting both endothelial and neoplastic compart-ments should be advantageous. In this field, we already designedtruncated proteins that are dominant-negative effectors of nativegrowth factors specific to endothelial and tumor cell proliferation.They are capable of silencing the native protein by forminginactive heterodimeric molecules (9). In particular, we demon-strated that a truncated Pleiotrophin (PTN), lacking the C-termi-nal 111–136 portion of the molecule (PTND111-136), acted as adominant-negative mutant that selectively inhibited in vitro theproliferationof a PTN-dependentMDA-MB-231breast tumor andendothelial cells by silencing the VEGF-A and HIF1a pathwaysand breast tumor growth in vivo by 81% (10).
In the current study, we assessed the antiangiogenesis andantitumor activities of a new dominant-negative mutant derivedfrom theMidkine (MDK) growth factor. MDK is a 121 amino acidprotein that like PTNbelongs to the heparin-binding growth factor(HBGF) family and is constitutively active when it forms homo-dimers that are stabilized by heparin (11). Although MDKis composed of two major N- and C-terminal domains heldtogether by disulfide linkages, most protein activities are per-formed by C-terminal domain. That is the case of the lysine-richdomain of PTN111-136C-terminal residues, which is required forits mitogenic and tumorigenic activities (12). MDK was identifiedas the ligand of different tyrosine kinase receptors such as ana-plastic lymphomakinase (ALK), aswell as the receptor-like proteintyrosine phosphatase (RPTP) and the LDL-receptor–related pro-tein families (LRP; refs. 13–15). Through these interactions, MDK
1INSERM U745, Laboratoire de G�en�etique Mol�eculaire, Facult�e desSciences Pharmaceutiques et Biologiques, Universit�e Paris Descartes,Paris, France. 2INSERM U935, Human Embryonic Stem Cell CoreFacility, Villejuif, France. 3D�epartements de Biologie et PathologieM�edicale, Institute Gustave-Roussy, Villejuif, France. 4Laboratoired'Oncog�en�etique, Institut Curie, Hopital Ren�e Huguenin, St-Cloud,France. 5Universit�e Paris Descartes, Sorbonne Paris Cit�e, Facult�e deSciences Pharmaceutiques et Biologiques, Paris, France. 6Universit�eParis-Sud 11, Campus Orsay, Orsay, France.
Corresponding Author: Frank Griscelli, INSERM 935, Institut Andr�e Lwoff,Batiment A CNRS, 7 Rue Guy Moquet, Villejuif 94802, France. Phone: 3301-4211-5193; Fax: 3301-4211-5313; E-mail: [email protected]
doi: 10.1158/1535-7163.MCT-14-0226
�2014 American Association for Cancer Research.
MolecularCancerTherapeutics
www.aacrjournals.org 213
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226

is involved in important cellular functions, promoting the growthof tumor cell lines (16) and cell survival by inhibiting apoptosis(17) and was shown to promote tumor angiogenesis (18). Inparticular, MDK was shown to correlate with tumor progressionand invasiveness of neuroblastoma. Indeed, plasma MDK levelswere found to be significantly correlated with the status of N-mycamplification, TRKA expression, and stage, which are knownprognostic factors in neuroblastoma (19). In this study, weassessed the effects of the 81-121 deleted MDK (MDKD81-121)lacking its last 40 amino acids, and for the first time, demonstratethat this truncated protein could act as a potent inhibitor of tumorgrowth and angiogenesis in a neuroblastoma tumor model.
Materials and MethodsPlasmid DNA
The pcDNA3-LacZ control plasmid encoding b-galactosidasewas constructed as previously described (10). The pcDNA3-MDKD81-121-MycHis plasmid was obtained by cloning theMDKD81-121 fragment between the BamHI and the EcoRI sitesof the pcDNA3.1(þ)/myc-HisA plasmid (Invitrogen). All plas-mids were purified using endofree Plasmid Maxi Kit (Qiagen),and solubilized in endotoxin-free 0.9% NaCl.
Cell culture and conditioned mediumHuman neuroblastoma cell lines KCNR, CLB-PE, LAN-1, and
GIMen were a kind gift of Dr. Isabelle Janoueix-Lerosey (InsermU830, Institut Curie, Paris, France; ref. 20). The human IGR-N91cell line was a kind gift of Dr. B�enard (CNRS UMR 8126, InstitutGustave Roussy, Villejuif, France) and was established from aninfiltrated bone marrow collected from a stage IV neuroblastoma(21). These neuroblastoma cell lines as well as SK-N-AS (ATCCCRL2137), SH-SY5Y (ATCC CRL2266), and BE(2)-M17 (ATCCCRL2267), MRC-5 cell line (ATCC CCL-171) and monkey Cos7cells (ATCC CRL1651) were grown in DMEM supplemented with10% FBS. Human umbilical smooth muscle cells (SMC) were akind gift of Dr. Georges Uzan (INSERM U972, Hopital Paul-Brousse, Villejuif, France) and were isolated and characterized asdescribed previously (22). Human umbilical vein endothelialcells (HUVEC, Lonza CC-2517) and SMCs were cultured in theiradequate medium from Lonza and promocell, respectively (10).Experiments involving ATCC or Lonza cell lines were mostlyperformed in the same year they were purchased. Authors per-formed no authentification on the cell lines used in this article. Toobtain Cos7-MDKD81-121 and Cos7-LacZ cells, Cos7 cells weretransfected with 8 mg of either pcDNA3-MDKD81-121 orpcDNA3-LacZ plasmids, using jetPEI reagent (Polyplus Transfec-tion). The medium was changed 4 hours after transfection andstably transfected cells were selected with Geneticin/G-418 (700mg/mL; Invitrogen). After 2 weeks of culture, single cells wereadded to a 96-well cell culture plate for subcloning and expandedin presence of G-418. Conditioned medium was harvested, ali-quoted, and stored at �20�C until use.
Small interfering RNAIGR-N91 and SH-SY5Y cells were seeded in 24-well tissue
culture dishes, at a density of 5 � 104 cells per well in theappropriate medium. After 24 hours, MDK siRNA (SI02663059;Qiagen) alone or in combination with PTN siRNA (SI00043764;Qiagen) and scrambled siRNA (control) were added at a finalconcentration of 10 nmol/L. Three days after transfection, cellswere collected for real-time RT-PCR, or were used to perform
proliferation assay. Cell supernatantswere harvested and stored at�20�C for an ELISA assay.
Western blot analysis, ELISA, and FISHSamples protein concentrationwas quantified usingDc Protein
Assay (Bio-Rad). PTN expression in neuroblastoma cell pelletswas assessed as previously described (10). For MDK detectionin cell pellets by Western blot analysis, proteins were transferredto a nitrocellulose membrane (Pall) that was blocked in PBS0.2% Tween and 5% powdered milk. Membranes were thenprobed with rabbit polyclonal anti-MDK antibody (1:5,000;American Antigenix). Swain anti-rabbit peroxidase-linked anti-body (1:1,000; Dako) was used as secondary antibody. ALKWestern blot was performed as previously described (10). Forthe RPTPb/z Western blot analysis, the protein was detectedusing a primary rat anti-RPTPb/z antibody (1:500, R&D Sys-tems), and a secondary anti-rat antibody. Actin was detectedwith monoclonal anti–a-actin peroxidase conjugate antibody(1:50,000). MDK expression in cell supernatants was quantifiedby an ELISA assay using a Human Midkine Construction Kit(America antigenix), according to manufacturer's instructions.FISH was performed using the Vysis LSI ALK (2q23) dual color,break-apart rearrangement probe (Abbott Molecular).
Real-time RT-PCRThe theoretical and practical aspects of real-time quantitative
RT-PCR using the ABI Prism 7900 Sequence Detection System(Perkin-Elmer Applied Biosystems) as well as the RNA extractionmethod, cDNA synthesis, and PCR conditions, were described indetail elsewhere (10). Target gene expression was normalizedrelative to an endogenous RNA control (TBP gene, which encodesthe TATA box-binding protein), as previously described (10).
Virus construction and infectionAdMDKD81-121 is a DE1–DE3 recombinant adenovirus that
expresses the truncated protein MDKD81-121 lacking the last 40amino acids. The MDKD81-121 fragment was retrieved frompcDNA3-MDKD81-121 by digestion with AflII and AvrII andinserted into the pCA350 shuttle plasmid. The recombinantadenoviral genome (Ad5DE1E3) encoding the truncated proteinwas generated by homologous recombination between plasmidspCA350 andpOSE1700 in competent E. coli JM83 (ATCC35607).After recombination, the adenoviral genome was excised by PacIand the AdMDKD81-121 virus was obtained by transfecting 293cells (CRL-1573; ATCC) using Lipofectamine (Invitrogen).Recombinant adenovirus expressing no transgene (AdCO1) wasused as negative control. Recombinant viruses were expandedin 293 cells and purified by two-step CsCl gradient ultracentri-fugation. Viral stocks were desalted using G50 columns (Phar-macia), frozen, and preserved at�80�C in PBS 10% glycerol. Viraltiter was quantified as plaque-forming units (PFU)/mL followinginfection of 293 cells and infection was carried out with amultiplicity of infection (MOI) of 100 PFU per cell.
Proliferation assayHuman neuroblastoma cell lines IGR-N91, SH-SY5Y, and
HUVEC were seeded in 24-well tissue culture dishes at a densityof 5 � 104 cells per well in complete media. After 24 hours, themedium was replaced by 420 mL of conditioned medium Cos7containing MDKD81-121, and 420mL of medium containing 1%or 5%of FBS, respectively, for neuroblastoma and endothelial cell
Dianat et al.
Mol Cancer Ther; 14(1) January 2015 Molecular Cancer Therapeutics214
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226

lines. The culture medium was refreshed every 24 hours over 4days, and cell survival was then quantified by a cell proliferationassay using MTT (0.5 mg/mL), as previously described (10). IGR-N91, SH-SY5Y, MRC-5 cell lines, HUVECs, and SMCs were trans-ducedwith AdMDKD81-121 or AdCO1 used as control and after 4days cell proliferationwas performed as previously described (10).
Coculture of SMCs and HUVECsA endothelial SMC coculture experiment was performed as
previously described, using the 24-well Transwell culture plates(0.4 mm pore size, Corning; ref. 23). Briefly, SMCs were seeded in24-well culture plates at 5� 104 cell density perwell, infectedwith200 MOI of AdMDKD81-121 and cultured during 24 hours withDMEM supplemented with 5% FBS. Then, HUVECs were seededonto the culture inserts at a density of 2.0� 105 cells per well andcultured with DMEM supplemented with 5% of FBS. A total of100 ng/mL of MDK was added or not to the medium. After 48hours, aMTT assay was performed to quantify the endothelial cellproliferation.
Endothelial tube formation assayTwo-hundred microliters of growth factor–reduced Matrigel (8
mg/mL; BDBiosciences)was added to a 48-well tissue culture dishand was polymerized during 1 hour at 37�C. HUVEC cells werethen seeded at a density of 6� 104 cells per well in 250 mL of EBM-2 1% FBS and 100 or 250 mL of conditioned medium (control orMDKD81-121 medium). After 18 hours of incubation undernormal culture conditions, the capillary-like structures werestained with Giemsa and the length of networks was quantifiedunder phase-contrast microscopy, using Archimed and Videomatsoftwares (GT Vision).
Plasmid electrotransferBefore the electrotransfer procedure, 6-week-old female athy-
mic mice were anesthetized with an intraperitoneal injection of100 mg/kg ketamine (Ketalar, Panpharma), and 40 mg/kg xyla-zine (Rompun, Bayer). Forty micrograms of pcDNA3-MDKD81-121 or pcDNA3-LacZ plasmids was resuspended in 30 mL sterile0.9%normal saline and injected inboth tibialis cranialismuscles ofmice using a Hamilton syringe. The electrotransfer was performedwith a Cliniporator system (IGEA), as previously described (10).
Tumor cell inoculationIGR-N91 cells were harvested, washed, and resuspended in
sterile 0.9% normal saline at a concentration of 10 � 106 cells in100 mL andwere injected subcutaneously into the athymicmice, 7days after plasmid electrotransfer. The tumor size was monitoredafter 41 days by measuring the largest (a) and the longest (b)tumor diameters and the tumor volume was calculated using thefollowing formula: [(aþb)/2]3� p/6. Mice were sacrificed using alethal dose of isofluorane: for each mouse, the electrotransferredmuscles and the tumor were collected and stored under appro-priate conditions for a further immunohistochemical analysis.Serum was collected at day 41 for the quantification of MDKD81-121 by a specific ELISA.
Immunohistochemistry and image analysisAfter mice were sacrificed, muscles and tumors were collected
and fixed in FineFix (Milestone). Paraffin sections of IGR-N91tumor cells were prepared to quantify the number of intratumoralvessels by CD31 immunostaining, as well as Ki67, caspase-3, and
SMA immunostaining, as previously described (4, 10). Images ofhistologic slides were recordedwith aNikon SuperCoolscan 8000EDscanner, and analyzedwith thePixCyt image analysis software.Paraffin sections of electroporated muscle were stained for thedetection of b-galactosidase and MDKD81-121, as previouslydescribed (10).
Statistical analysisEach experiment was done at least twice. Statistical signifi-
cance was evaluated using the Student t test (unilateral andunpaired).
ResultsExpression of MDK, PTN, and their receptors
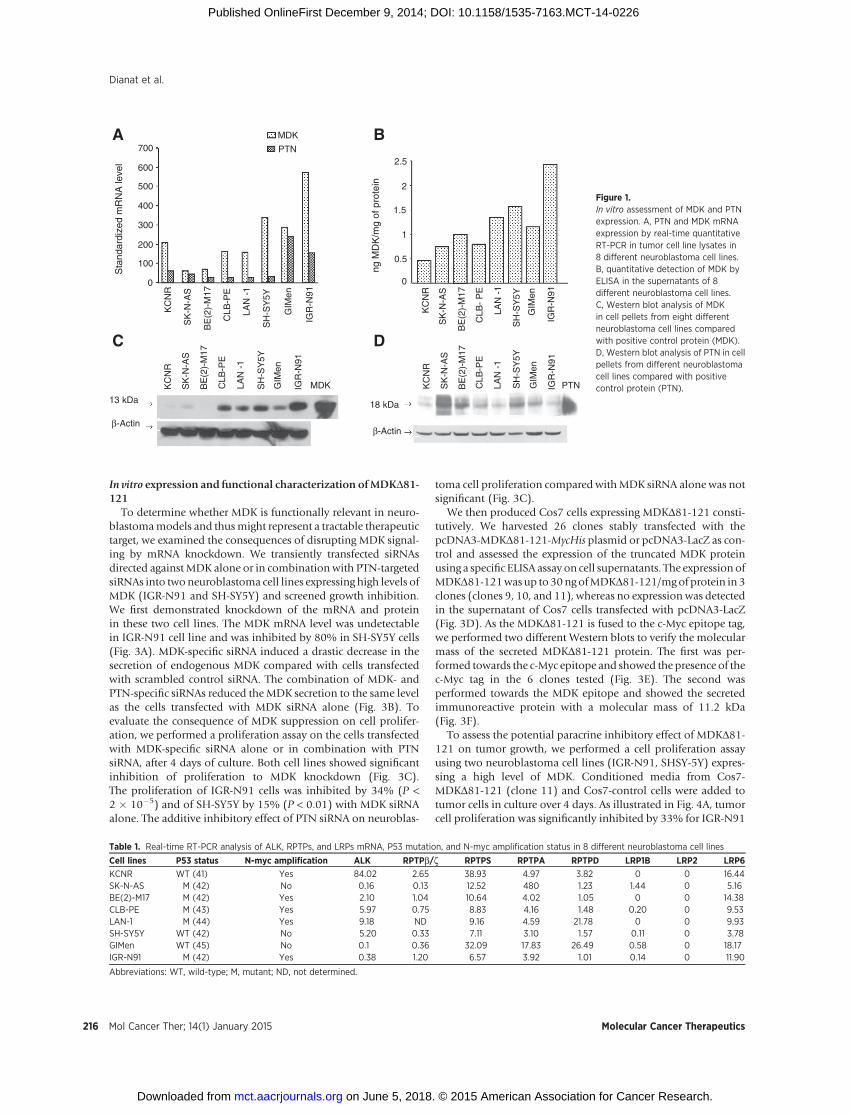
To analyze the expression of MDK and PTN in neuroblastomacell lines (KCNR, SK-N-AS, BE(2)-M17, CLB-PE, LAN-1, SH-SY5Y,GIMen, and IGR-N91), we first quantified endogenous MDK andPTN mRNA expression by real-time RT-PCR. We showed thatMDK and PTN mRNAs were detected in all of the 8 cell lines andthat MDK mRNA level was significantly higher (P ¼ 0.039) thanPTN mRNA (Fig. 1A). Indeed, in five cell lines (KCNR, CLB-PE,LAN-1, SH-SY5Y, and IGR-N91) MDKmRNA was 3.5- to 11-foldhigher than PTN mRNA. To study MDK protein expression after48 hours of culture, cell supernatants and cell extracts wereanalyzed using ELISA and Western blot, respectively. SecretionofMDK in the culturemediawasdemonstrated for all the cell lines(Fig. 1B). The IGR-N91 cell line was found to produce and secretethe highest level of MDK and was also found to produce a highlevel of MDK mRNA (Fig. 1A and B). Furthermore, immunore-active MDK protein was also detected at a high level in IGR-N91cell extracts that also expressed a low level of PTN compared withother cell lines (Fig. 1C and D). All these results clearly showedheterogeneous expression of MDK and PTN in all the cell linestested and that IGR-N91 and SH-SY5Y cell lines appeared toproduce and secrete high levels of MDK certainly contributingto their proliferation.
To evaluate the potential contribution of HBGH signaling, wethen studied the expression of its receptors (ALK, RPTPs, andLRPs) using quantitative RT-PCR. Table 1 shows that ALK mRNAwas present in all neuroblastoma cell lines tested with clearheterogeneity of expression as 5 of 8 cell lines (KCNR, BE(2)-M17, CLB-PE, LAN-1, SH-SY5Y) expressed higher levels of ALKmRNA. Our results were confirmed by Western blot analysisshowing the presence of two ALK isoforms, a 220-kDa nativetransmembrane protein and a 140-kDa cleaved soluble ALK (Fig.2A). We then focused on the ALK gene to verify whether there wasa correlation betweenALKprotein expression andpossible somat-ic amplification of the ALK genomic locus. No gene amplification(<5 copies) was found in IGR-N91 (Fig. 2C) and SH-SY5Y (Fig.2D). Moreover, the level of ALK expression was not stronglycorrelated with the N-myc status as SH-SY5Y (not amplified forN-myc) and IGR-N91 (amplified for N-myc), respectively,expressed high and low ALK levels. The second well-knownHBGH receptor, RPTPb/z, was also detected by RT-PCR (Table1) and Western blot analysis (Fig. 2B) in all cell lines at the samelevels and thus seemed to be highly ubiquitous in all cell linestested. Concerning other RPTP isoforms and LRP receptors, all theneuroblastoma cell lines exhibited RPTPS, RPTPA, RPTPD, andLRP6 mRNA. Some cell lines expressed LRP1B but none of thecells expressed LRP2 (Table 1).
Inhibition of Neuroblastoma Development by Targeting Midkine
www.aacrjournals.org Mol Cancer Ther; 14(1) January 2015 215
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226
In vitro expression and functional characterization ofMDKD81-121
To determine whether MDK is functionally relevant in neuro-blastomamodels and thusmight represent a tractable therapeutictarget, we examined the consequences of disrupting MDK signal-ing by mRNA knockdown. We transiently transfected siRNAsdirected against MDK alone or in combination with PTN-targetedsiRNAs into twoneuroblastoma cell lines expressing high levels ofMDK (IGR-N91 and SH-SY5Y) and screened growth inhibition.We first demonstrated knockdown of the mRNA and proteinin these two cell lines. The MDK mRNA level was undetectablein IGR-N91 cell line and was inhibited by 80% in SH-SY5Y cells(Fig. 3A). MDK-specific siRNA induced a drastic decrease in thesecretion of endogenous MDK compared with cells transfectedwith scrambled control siRNA. The combination of MDK- andPTN-specific siRNAs reduced the MDK secretion to the same levelas the cells transfected with MDK siRNA alone (Fig. 3B). Toevaluate the consequence of MDK suppression on cell prolifer-ation, we performed a proliferation assay on the cells transfectedwith MDK-specific siRNA alone or in combination with PTNsiRNA, after 4 days of culture. Both cell lines showed significantinhibition of proliferation to MDK knockdown (Fig. 3C).The proliferation of IGR-N91 cells was inhibited by 34% (P <2 � 10�5) and of SH-SY5Y by 15% (P < 0.01) with MDK siRNAalone. The additive inhibitory effect of PTN siRNA on neuroblas-
toma cell proliferation compared withMDK siRNA alone was notsignificant (Fig. 3C).
We then produced Cos7 cells expressing MDKD81-121 consti-tutively. We harvested 26 clones stably transfected with thepcDNA3-MDKD81-121-MycHis plasmid or pcDNA3-LacZ as con-trol and assessed the expression of the truncated MDK proteinusing a specific ELISA assay on cell supernatants. The expression ofMDKD81-121was up to30ngofMDKD81-121/mgof protein in 3clones (clones 9, 10, and 11), whereas no expression was detectedin the supernatant of Cos7 cells transfected with pcDNA3-LacZ(Fig. 3D). As the MDKD81-121 is fused to the c-Myc epitope tag,we performed two different Western blots to verify the molecularmass of the secreted MDKD81-121 protein. The first was per-formed towards the c-Myc epitope and showed the presence of thec-Myc tag in the 6 clones tested (Fig. 3E). The second wasperformed towards the MDK epitope and showed the secretedimmunoreactive protein with a molecular mass of 11.2 kDa(Fig. 3F).
To assess the potential paracrine inhibitory effect of MDKD81-121 on tumor growth, we performed a cell proliferation assayusing two neuroblastoma cell lines (IGR-N91, SHSY-5Y) expres-sing a high level of MDK. Conditioned media from Cos7-MDKD81-121 (clone 11) and Cos7-control cells were added totumor cells in culture over 4 days. As illustrated in Fig. 4A, tumorcell proliferation was significantly inhibited by 33% for IGR-N91
Table 1. Real-time RT-PCR analysis of ALK, RPTPs, and LRPs mRNA, P53 mutation, and N-myc amplification status in 8 different neuroblastoma cell lines
Cell lines P53 status N-myc amplification ALK RPTPb/z RPTPS RPTPA RPTPD LRP1B LRP2 LRP6
KCNR WT (41) Yes 84.02 2.65 38.93 4.97 3.82 0 0 16.44SK-N-AS M (42) No 0.16 0.13 12.52 480 1.23 1.44 0 5.16BE(2)-M17 M (42) Yes 2.10 1.04 10.64 4.02 1.05 0 0 14.38CLB-PE M (43) Yes 5.97 0.75 8.83 4.16 1.48 0.20 0 9.53LAN-1 M (44) Yes 9.18 ND 9.16 4.59 21.78 0 0 9.93SH-SY5Y WT (42) No 5.20 0.33 7.11 3.10 1.57 0.11 0 3.78GIMen WT (45) No 0.1 0.36 32.09 17.83 26.49 0.58 0 18.17IGR-N91 M (42) Yes 0.38 1.20 6.57 3.92 1.01 0.14 0 11.90
Abbreviations: WT, wild-type; M, mutant; ND, not determined.
BA
C
BE
(2)-
M17
CLB
-PE
LAN
-1
IGR
-N91
KC
NR
GIM
en
SH
-SY
5Y
SK
-N-A
S
Sta
ndar
dize
d m
RN
A le
vel
PTN
MDK
ng M
DK
/mg
of p
rote
in
0
0.5
1
1.5
2
2.5
MDK
13 kDa
β-Actin
D
PTN
β-Actin
18 kDa
0
100
200
300
400
500
600
700
BE
(2)-
M17
CLB
-P
E
LAN
-1
IGR
-N91
KC
NR
GIM
en
SH
-SY
5Y
SK
-N-A
S
BE
(2)-
M17
CLB
-PE
LAN
-1
IGR
-N91
KC
NR
GIM
en
SH
-SY
5Y
SK
-N-A
S
BE
(2)-
M17
CLB
-PE
LAN
-1
IGR
-N91
KC
NR
GIM
en
SH
-SY
5Y
SK
-N-A
S
Figure 1.In vitro assessment of MDK and PTNexpression. A, PTN and MDK mRNAexpression by real-time quantitativeRT-PCR in tumor cell line lysates in8 different neuroblastoma cell lines.B, quantitative detection of MDK byELISA in the supernatants of 8different neuroblastoma cell lines.C, Western blot analysis of MDKin cell pellets from eight differentneuroblastoma cell lines comparedwith positive control protein (MDK).D, Western blot analysis of PTN in cellpellets from different neuroblastomacell lines compared with positivecontrol protein (PTN).
Dianat et al.
Mol Cancer Ther; 14(1) January 2015 Molecular Cancer Therapeutics216
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226

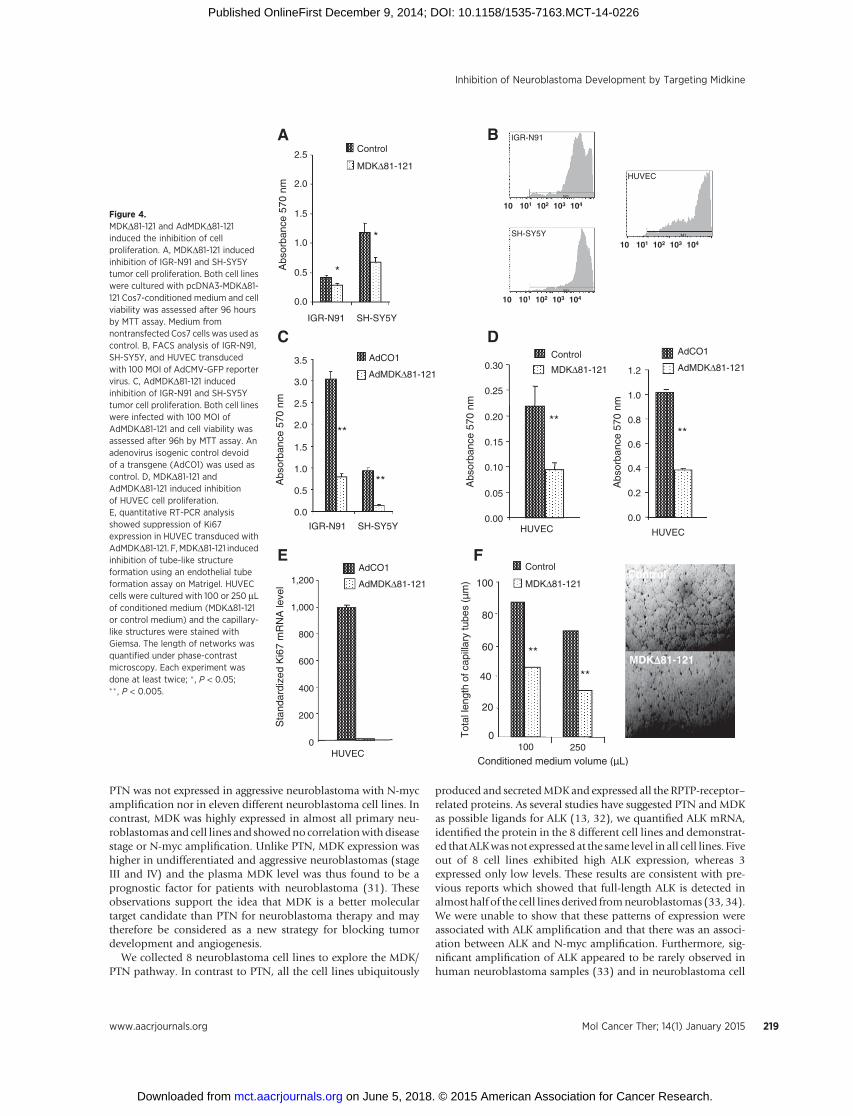
cells (P < 3� 10�7), and by 43% for SH-SY5Y cells (P < 1� 10�6).To confirm these results, we used a recombinant adenoviruscarrying a CMV-driven MDKD81-121 peptide (AdMDKD81-121)and an isogenic adenovirus (AdCO1) as control. The efficacy ofadenovirus-mediated gene delivery was first evaluated by quanti-fying the proportion of cells infectedwith the AdCMV-GFP reportervirus after 48 hours by FACS analysis (Fig. 4B). The dose of 100multiplicity of infection (MOI) enabled us to efficiently transduceall IGR-N91 and SH-SY5Y cell lines as well as human primaryendothelial cells (HUVECs). Under these conditions, transductionof IGR-N91 and SH-SY5Y cell lines by AdMDKD81-121 resultedin stronger significant inhibition of cell proliferation comparedwith the use of the Cos7-conditioned medium (Fig. 4C).
Effect of MDKD81-121 on angiogenesisTo evaluate a possible antiangiogenic role of MDKD81-121, we
first performed a proliferation assay on HUVEC, under the sameconditions as for the tumor cells. We observed 60% inhibition ofHUVEC proliferation (P < 5 � 10�7) with the supernatant con-taining MDKD81-121, compared with the control or after theirinfection with 100 MOI of AdMDKD81-121 (Fig. 4D). Theseresults were confirmed by quantitative RT-PCR with completerepression of Ki67 expression in AdMDKD81-121 transducedHUVECs compared with control (AdCO1-transduced) cells(Fig. 4E).
As endothelial cell migration is also an important processin angiogenesis, we then used an in vitro Matrigel angiogenesisassay based on the differentiation of endothelial cells and theformation of tube-like structures on an extracellular matrix.HUVEC cells were seeded on Matrigel in the presence of 100 or250 mL of conditioned medium (MDKD81-121 or control medi-um). As shown in Fig. 4F, we observed dose-dependent inhibitionof tube-like structure formation with a 59%mean reduction afteradding 250 mL of MDKD81-121 conditioned medium.
To obtain further evidence for antiangiogenic activity ofMDKD81-121, we studied the endoplasmic reticulum (ER)stress-mediated apoptosis in endothelial cells transduced withAdMDKD81-121. C/EBP homologous protein (CHOP), alsoknown as DNA damage-inducible transcript 3 (DDIT3) is thekey proapoptotic effector of ER stress–mediated apoptosis. CHOPexpression was significantly upregulated in HUVECs transduced
with AdMDKD81-121 (70-fold increase, P < 0.01) as assessed byquantitative RT-PCR (Table 2).We observed significant activationof a set of genes implicated in anunfoldedprotein response (UPR)pathway, which triggers apoptosis in the case of intensive ER-stress, such as heat shock 70-kDa Protein 5 (HSPA5), growtharrest- andDNAdamage-inducible protein (GADD34), activatingtranscription factor 2 and 3 (ATF2 and ATF3), and the splicedvariant of X-box binding protein 1 (XBP1S) in endothelial cellsexpressing truncated MDKD81-121 protein (Table 2).
As endothelial cells boarding blood vessels are surrounded bySMCs and fibroblasts, we therefore investigated the inhibitoryeffect of MDKD81-121 on SMC (Fig. 5B) and MRC-5 fibroblast(Fig. 5A) using theMTT proliferation assay. Cell proliferation wassignificantly inhibited when MRC5 (P < 0.0001) and SMC (P ¼0.005) were infected with AdMDKD81-121 with 57% and 38%ofinhibition, respectively. In contrast, when SMC were treated withMDK (100 ng/mL), no increase in cell proliferation was observed(Fig. 5C).
We further studied the effect of AdMDKD81-121on endothelialcells proliferation when cocultured with SMC during 48 hours inthe presence or not of 100 ng/mL of MDK (Fig. 5D). In theseexperiments, when HUVEC were treated only with MDK noincrease in cell proliferation was observed (P ¼ 0.65). However,when HUVEC were cocultured with SMC, MDK significantlyincreased the cell proliferation (P ¼ 0.0002). Furthermore cocul-ture of HUVEC with AdMDKD81-121–transduced SMC withor without MDK repressed significantly HUVEC proliferation(Fig. 5D).
Inhibition of tumor growth by MDKD81-121 in vivoTo confirm the antitumor potency of MDKD81-121 in vivo, we
used a prophylactic protocol after muscle electrotransfer of thepcDNA3-MDKD81-121-MycHis plasmid. We first electrotrans-ferred 30 mg pcDNA3-MDKD81-121-MycHis or pcDNA3-LacZinto the tibialis cranialismuscles of nude mice (n¼ 10 per group).Seven days later, 10 � 106 IGR-N91 cells were inoculated sub-cutaneously. Serum was collected from mice 41 days after theelectrotransfer to quantify the systemic amount of MDKD81-121secreted by the electrotransferred muscle cells. As shownin Fig. 6A, IGR-N91 tumors from theMDKD81-121–treated groupdid not grow and after 2 weeks the tumor size reduced drastically,
250 kDa 140 kDa
β-Actin β-Actin
220 kDa
A B
C
BE
(2)-
M17
CLB
-PE
LAN
-1
IGR
-N91
KC
NR
GIM
en
SH
-SY
5Y
SK
-N-A
S
BE
(2)-
M17
CLB
-PE
LAN
-1
IGR
-N91
KC
NR
GIM
en
SH
-SY
5Y
SK
-N-A
S
DIGR-N91 SH-SY5Y
Figure 2.In vitro assessment of MDK receptorsexpression. A, immunoblots of ALK invarious neuroblastoma cell lines. B,immunoblots of RPTPb/z in variousneuroblastoma cell lines. C andD, FISHwas performed on interphase nucleifrom IGR-N91 and SH-SY5Y culturedlines.We used 2 probes (Vysis LSI ALKDual Color, Abbott Molecular) specificfor the 2p23 chromosomal region(SHGC-56576 spectrum orange,SHGC-104192 spectrum green),showing two signals for each probe inall the cells analyzed corresponding tothe presence of the ALK chromosomeregion.
Inhibition of Neuroblastoma Development by Targeting Midkine
www.aacrjournals.org Mol Cancer Ther; 14(1) January 2015 217
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226

whereas the tumors from the control group treated with pcDNA3-LacZ increased in size. This inhibition correlated with the detec-tion of MDKD81-121 immunoreactive material in the serum(Fig. 6B) and in the muscles (Fig. 6C). IGR-N91 tumors treatedwith MDKD81-121 appeared much less vascularized than theb-galactosidase–treated control group (Fig. 6A). Intratumoralangiogenesis was thus assessed by CD31 and a-SMA immu-nostaining of tumor sections (Fig. 6F). The results indicated amarked reduction (37%) of intratumoral vascularization with-in MDKD81-121–treated tumors (score of 3.15 � 0.15) ascompared with the b-galactosidase–treated group (score of2.14 � 0.05; Fig. 6D and E). These results correlated with SMAimmunostaining showing again the presence of large vesselsonly in tumors treated with MDKD81-121 (Fig. 6F). Tumorproliferation activity was assessed by Ki67 immunostaining,which is widely used in clinical pathology diagnostics. Asshown in Fig. 6F, the proliferation appeared to be drasticallydecreased within MDKD81-121–treated tumors (50% vs. 90%of positive tumor cells) with a higher proportion of apoptotic
cells through activation of caspase-3–mediated apoptosis path-way (35 � 8 vs. 3 � 2 positive nuclei per field; Fig. 6F).
DiscussionPTN and MDK are members of a new family of neurotrophic
factors known to be expressed in many human tumors (24–27).They participate in tumorigenesis through their action on angio-genesis and cell proliferation (18, 28), and their expression ishighly correlated with metastasis development (29).
MDK and PTN were found to play an important role inregulating growth and differentiation in neuroblastoma, the thirdmost common childhood solid tumor, which originates from thesympathoadrenal lineage of the neural crest. Both factors werefound to be expressed inmany primary neuroblastomas, but theirpatterns of expression were found to be quite different. PTN wasfound to be highly expressed in favorable neuroblastomas (stagesI, II, and IV-S), whereas it was expressed at a significantly lowerlevel in advanced tumors (stages III and IV; ref. 30). Furthermore,
11.2 kDa
10 Con
trol
20 kDa
11.2 kDa
E
Cos7-MDKΔ81-121 clones
Qua
ntity
of M
DK
Δ81-
121
(ng/
mL)
D
Control
30
15
01 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26
F
0
0.5
1
1.5
2
2.5
IGR-N91 SH-SY5YA
bsor
banc
e 57
0 nm
**
Qua
ntity
of M
DK
(ng
/mL)
0
1
2
3
IGR-N91 SH-SY5Y
B
C
*
0
0.2
0.4
0.6
0.8
1
1.2S
tand
ardi
zed
mR
NA
leve
l
IGR-N91 SH-SY5Y
Control siRNA
MDK siRNA
A
2 7 9 10 11 13 Con
trol
M
MDKΔ81-121
**MDK/PTN siRNA
Control siRNA
MDK siRNA
MDK/PTN siRNA
Control siRNA
MDK siRNA
MDK/PTN siRNA
NS
*
NS
Figure 3.Transfection of IGR-N91 and SH-SY5Ycell lines by siRNAs directed againstMDK, PTN, or nonsilencing sequencesiRNAs (control) and Cos7 cells bypcDNA3-MDKD81-121-MycHis orpcDNA3-LacZ plasmids. A, real-timeRT-PCR expression of MDK mRNA, 4days after the transfection of IGR-N91and SH-SY5Y cell lines with 10 nmol/LsiRNAs targeting MDK alone or incombination with PTN-targeted siRNAsand with nonsilencing sequence siRNAs.B, quantitative detection of MDK byELISA in the supernatant of IGR-N91 andSH-SY5Y cell lines transfected withsiRNAs directed against MDK alone or incombination with PTN siRNAs ornonsilencing sequence siRNAs after4 days of culture. C, siRNAs directedagainst MDK induced inhibition ofIGR-N91 and SH-SY5Y cell linesproliferation compared with thetransfection of nonsilencing siRNAs.PTN-targeted siRNA did not induced anysignificant additive effect onproliferationof neuroblastoma cell lines whentransfected in combination with MDK-targeted siRNAs; NS, not significant;� ,P <0.05; �� ,P <0.005. D, quantificationof MDKD81-121 in the supernatant ofCos7-transfected cells by ELISA. Cos7cells were transfected with 8 mg of eitherpcDNA3-MDKD81-121 or pcDNA3-LacZplasmids and subclones were expanded.Supernatants of 26 different Cos7-MDKD81-121 subclones were tested byELISA compared with Cos7-LacZ controlcells. E, Western blot analysis of thec-Myc epitope in 6 different Cos7-MDKD81-121 subclones. F, MDKD81-121immunoblotting of the culturesupernatant of Cos7-transfected cells(subclones 10).
Dianat et al.
Mol Cancer Ther; 14(1) January 2015 Molecular Cancer Therapeutics218
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226
PTN was not expressed in aggressive neuroblastoma with N-mycamplification nor in eleven different neuroblastoma cell lines. Incontrast, MDK was highly expressed in almost all primary neu-roblastomas and cell lines and showedno correlationwithdiseasestage or N-myc amplification. Unlike PTN, MDK expression washigher in undifferentiated and aggressive neuroblastomas (stageIII and IV) and the plasma MDK level was thus found to be aprognostic factor for patients with neuroblastoma (31). Theseobservations support the idea that MDK is a better moleculartarget candidate than PTN for neuroblastoma therapy and maytherefore be considered as a new strategy for blocking tumordevelopment and angiogenesis.
We collected 8 neuroblastoma cell lines to explore the MDK/PTN pathway. In contrast to PTN, all the cell lines ubiquitously
produced and secretedMDKand expressed all the RPTP-receptor–related proteins. As several studies have suggested PTN and MDKas possible ligands for ALK (13, 32), we quantified ALK mRNA,identified the protein in the 8 different cell lines and demonstrat-ed that ALKwasnot expressed at the same level in all cell lines. Fiveout of 8 cell lines exhibited high ALK expression, whereas 3expressed only low levels. These results are consistent with pre-vious reports which showed that full-length ALK is detected inalmost half of the cell lines derived fromneuroblastomas (33, 34).We were unable to show that these patterns of expression wereassociated with ALK amplification and that there was an associ-ation between ALK and N-myc amplification. Furthermore, sig-nificant amplification of ALK appeared to be rarely observed inhuman neuroblastoma samples (33) and in neuroblastoma cell
M1
M1
M1
10 101 102 103 104
10 101 102 103 104
10 101 102 103 104
IGR-N91
SH-SY5Y
HUVEC
A
*
*
0.0
0.5
1.0
1.5
2.0
2.5
SH-SY5YIGR-N91
Abs
orba
nce
570
nm
Control
MDKΔ81-121
C
B
0.00
0.05
0.10
0.15
0.20
0.25
0.30
HUVEC
Abs
orba
nce
570
nm
Control
MDKΔ81-121
**
0.0
0.2
0.4
0.6
0.8
1.0
1.2
HUVEC
Abs
orba
nce
570
nm
**
AdCO1
AdMDKΔ81-121
0.0
0.5
1.0
1.5
2.0
2.5
3.0
SH-SY5YIGR-N91
Abs
orba
nce
570
nm
AdCO1
AdMDKΔ81-121
**
**
3.5
D
0
20
40
60
80
100
100 250
Conditioned medium volume (µL)
Tot
al le
ngth
of c
apill
ary
tube
s (µ
m)
Control
MDKΔ81-121
**
**
F
MDKD81-121
Control
E
0
200
400
600
800
1,000
1,200
Sta
ndar
dize
d K
i67
mR
NA
leve
l
HUVEC
AdCO1
AdMDKΔ81-121
Figure 4.MDKD81-121 and AdMDKD81-121induced the inhibition of cellproliferation. A, MDKD81-121 inducedinhibition of IGR-N91 and SH-SY5Ytumor cell proliferation. Both cell lineswere cultured with pcDNA3-MDKD81-121 Cos7-conditioned medium and cellviability was assessed after 96 hoursby MTT assay. Medium fromnontransfected Cos7 cells was used ascontrol. B, FACS analysis of IGR-N91,SH-SY5Y, and HUVEC transducedwith 100 MOI of AdCMV-GFP reportervirus. C, AdMDKD81-121 inducedinhibition of IGR-N91 and SH-SY5Ytumor cell proliferation. Both cell lineswere infected with 100 MOI ofAdMDKD81-121 and cell viability wasassessed after 96h by MTT assay. Anadenovirus isogenic control devoidof a transgene (AdCO1) was used ascontrol. D, MDKD81-121 andAdMDKD81-121 induced inhibitionof HUVEC cell proliferation.E, quantitative RT-PCR analysisshowed suppression of Ki67expression in HUVEC transduced withAdMDKD81-121. F,MDKD81-121 inducedinhibition of tube-like structureformation using an endothelial tubeformation assay on Matrigel. HUVECcells were cultured with 100 or 250 mLof conditioned medium (MDKD81-121or control medium) and the capillary-like structures were stained withGiemsa. The length of networks wasquantified under phase-contrastmicroscopy. Each experiment wasdone at least twice; � , P < 0.05;�� , P < 0.005.
Inhibition of Neuroblastoma Development by Targeting Midkine
www.aacrjournals.org Mol Cancer Ther; 14(1) January 2015 219
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226

lines (20). Recently, somatic and germline activatingmutations ofthe ALK kinase domain of the receptor were documented inhuman neuroblastoma samples and in neuroblastoma cell lines.Thesemutations weremainly clustered in two hotspots located inexon 23 (F1174V) or exon 25 (R1275Q). The F1174V mutationwas observedmore frequently in cell lines than inprimary tumors,suggesting that the former may provide a selective in vitro growth
advantage (20). Interestingly, the knockdown of ALK mRNA bysiRNA resulted in profound inhibition of cell proliferation in allcell lines harboring mutant or wild-type ALK (20).
In the current study, we investigated a new strategy evaluatingthe antitumor and antiangiogenesis effects in a neuroblastomamodel of a truncatedMDK lacking the COOH-terminal 81 to 121portion of the molecule (MDKD81-121), that might act as adominant-negative effector for its mitogenic, angiogenic, andtumorigenic activities by interfering with the wild-type nativeprotein.
We decided to use two cell lines (SH-SY5Y and IGR-N91) toperform a further analysis of the effect ofMDKD81-121 for severalreasons. Both cell lines expressed a high level of MDK and a lowlevel of PTN compared with other cell lines, suggesting theexistence of a dominant autocrine MDK signaling loop. In addi-tion, ALK mRNA was highly expressed in SH-SY5Y cells and was52-fold higher than themRNA level detected in IGR-N91 cells andboth cell lines identically expressed the other tyrosine kinasereceptors (RPTPs and LRPs).
Table 2. Real-time RT-PCR analysis of CHOP pathway–mediated apoptosis inHUVEC cells transduced with AdMDKD81-121 or AdCO1.
AdCO1 AdMDKD81-121 P value
MDK 8,862 � 941 65,387 � 6,013 0.009HSPA5 33,195 � 2854 237,304 � 24,156 0.015DOIT3 (CHOP) 5 � 0.28 353 � 46 0.023PPP1R15A(GADD34) 307 � 21 2,326 � 206 0.008XEP1L 2,454 � 133 1,102 � 257 0.11XBP1S 351 � 27 2,815 � 247 0.0075ATF2 482 � 42 771 � 40 0.02ATF3 74 � 4.5 4,789 � 567 0.015
AdMDKΔ81-121
A
0
0.5
1
1.5
2
2.5
3
**
50 MOI
AdCO1
Abs
orba
nce
570
nm
**
100 MOI
D
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
HUVECSMC
MDK 100 ng/mLAdMDKΔ81-121–transduced SMC
+ + + + + ++ + + +−−
++ −+ −−+ + −−−−
NS P = 0.0002
P = 0.0008
P = 0.01
0.40
0.45
Abs
orba
nce
570
nm
AdMDKΔ81-121
0
0.2
0.4
0.6
0.8
1
Control MDK100 ng/mL
NS
C
0
0.2
0.4
0.6
0.8
1
AdCO1B
200 MOI
**
NS
SMCMRC-5
Figure 5.In vitro assessment of AdMDKD81-121effect on HUVEC and SMC apoptosisand proliferation. A, proliferation ofMRC-5 cells transduced with 50 or 100MOI AdMDKD81-121 was assessed withMTT assay after four days. B, cellproliferation of SMCs transduced with200 MOI AdMDKD81-121 or AdCO1 ascontrol. C, proliferation of SMC treatedwith MDK (100 ng/mL) was assessedafter 96 hours by MTT assay. D,endothelial cell proliferation assayafter coculture with SMC orAdMDKD81-121-transduced SMC withor without MDK treatment. Eachexperiment was done at least twice.NS, not significant; � , P < 0.05;��, P < 0.005.
Dianat et al.
Mol Cancer Ther; 14(1) January 2015 Molecular Cancer Therapeutics220
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226

Our study was completed with the use of specific MDK siRNA,to downregulate the level of MDK mRNA in both cell lines. AsMDK and PTN share same receptors and have 45% structuralhomology, we investigated whether knockdown of PTN expres-sion together with MDK could have a synergic effect on cellproliferation. Our results showed that MDK siRNA alone ortogether with PTN siRNA induced significant specific inhibitionof proliferation of both cell lines. That was more pronounced forIGR-N91 cell lines which expressed the highest level of MDK. Wethus believe in the existence of a strong autocrine MDK signalingloop in both cell lines. However, extinction of PTN expressiontogether with MDK did not have any additive or synergic inhib-itory effect on IGR-N91 and SH-SY5Y proliferation. This obser-vation is consistent with the data regarding the low PTN expres-
sion level in these two cell lines (Fig. 1A) and our observation thatPTN protein was not detectable in these two cell lines supernatantonce assessed by ELISA (data not shown).
We demonstrated that MDKD81-121 is able to significantlyinhibit the proliferation of both IGR-N91 and SH-SY5Y cell linesin vitro. In this case, it might be probably due to the heterodimer-ization of MDKD81-121 with endogenous MDK which mainlyexplains its dominant-negative effect and that MDKD81-121might be able to dissociate MDK homodimers, thereby mightinduce the formation of nonfunctional heterodimers. Thus, theinhibition of MDK in both cell lines exerts several effects ondifferent important cellular systems. In this context, we believethat the inhibition of MDK byMDKD81-121 abolishes the MDK/ALK and/or the MDK/RPTPb/z signaling pathways, as previously
MDKΔ81-121
0
0.5
1
1.5
2
2.5
3
3.5
4
Sco
re
MDKΔ81-121
β-Galactosidase
**
β-Galactosidase β-Galactosidase
MDKΔ81-121
β-Galactosidase
MDKΔ81-121
A
CB
F
0
2
4
6 MDKΔ81-121
β-Galactosidase
ng M
DK
Δ81-
121/
mg
of p
rote
in **
E
0
10
20
30
40
50
60
70
2514107day
β-GalactosidaseMDKΔ81-121
Tum
or v
olum
e (m
m3 )
Ki67 SMACaspase-3
β-Gal
MDKΔ81-121
D
Figure 6.Tumor growth and inhibition ofintratumoral vascularization. A, effectof MDKD81-121 transgene expressionafter in vivo muscle pDNAelectrotransfer on IGR-N91 tumorgrowth in nudemice. Animals receiveda subcutaneous injection of 10 � 106
IGR-N91 cells 7 days afterelectrotransfer. The mean tumorvolume evolution is shown until day25 in MDKD81-121–treated versusb-galactosidase–treated mice(P < 0.05). Tumors from MDKD81-121–treated and b-galactosidase–treatedgroups are shown at day 41 after cellinoculation. B, quantification ofMDKD81-121 in serum by ELISA.C, detection of b-galactosidase andMDKD81-121 in electroporated muscleby immunohistochemistry.D, the extent of intratumoralvascularization was assessed by CD31immunostaining and quantified byassessing the mean immunoreactivescore within the whole tumor area atday 41. E, the extent of vascularizationof a representative tumor treated withMDKD81-121 and the control(b-galactosidase) is shown 41 daysafter the subcutaneous injection ofcells. F, antiproliferative,proapoptotic, and antiangiogeniceffect of MDKD81-121 is demonstratedby immunohistochemitry on tumorslices for Ki67, caspase-3 and SMA,respectively.
Inhibition of Neuroblastoma Development by Targeting Midkine
www.aacrjournals.org Mol Cancer Ther; 14(1) January 2015 221
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226

suggested with a PTN-derived peptide lacking the C-terminalresidues (PTND111-136). PTND111-136 was shown to act as apotent inhibitor of PTN biologic activities by the formation ofinactive heretodimers (PTN/PTND111-136; ref. 9). In particular,binding studies using a biotinylated peptide showed thatPTND111-136 could interfere with PTN binding to DU145 pros-tatic tumor cells, in an RPTPb/z-dependent fashion (35). Further-more, PTND111-136 was shown to bind to the extracellulardomain of ALK and to inhibit PTN-induced mitogenesis(9, 12) and thus is also able to interfere with the PTN/ALKpathway. MDK and PTN are members of the same neurotrophicfamily factors and are both basic, cysteine-rich polypeptidescontaining identical heparin-binding motifs, which are essentialfor the dimerization of PTN andMDK. They exhibit about 50% ofhomology and no sequence homology with other heparin-bind-ing proteins, such as fibroblast growth factors. In this context, theformation of MDKD81-121/PTN heterodimers is not excludedand further experiments are warranted to explore this hypothesis.
As angiogenesis is also known to be a keymechanism viawhichMDK promotes tumorigenesis, we then investigated the effects ofMDKD81-121on endothelial cell proliferation andmigration.Wewere able to show that the inhibition of MDK by MDKD81-121has major consequences on different stages of angiogenesis,because MDKD81-121 strongly inhibited the proliferation andthe migration of endothelial cells in vitro. We also showed thatMDKD81-121 could have major effects on their structural orga-nization in a three-dimensional model. MDKD81-121 is able toreduce the formation of capillary-like structures, a crucial step intumor neovascularization. The effects of MDKD81-121 onHUVEC could be mediated by the inhibition of ALK and/orRPTPb/z tyrosine kinase receptors as HUVECs have been shownto express MDK and both receptors (36, 37) enabling the stim-ulation of the phosphorylation of ALK (36) and the activation ofthe Akt and ERK pathways (38). We also showed that MDKD81-121 inhibits endothelial cell cycling with a complete extinction ofKi67 expression, while promotes cell apoptosis via activation of aset of genes, including CHOP, implicated in ER stress–mediatedapoptosis pathway. These data are consistent with the observa-tions from U87MG glioblastoma cell line and the truncatedprotein PTND111-136 which enhances apoptosis via CHOP pro-tein (39).
Angiogenesis process involves interactive dialog between endo-thelial and SMCs. These interactions were previously studied bySumi and colleagues using an artificial blood vessel model (23).They documented that MDK had no effect on the human aorticSMC or endothelial cells when cultured separately and theseobservations are in concordance with our results showing noeffect of MDK on SMCs and HUVECs. They also clearly showedthat MDK promotes growth and glycosaminoglycan synthesis ofHUVEC through its action on smoothmuscle cells that are able toproduce IL8. To evaluate whether MDKD81-121 is able to disruptthis interaction, we have cocultured HUVECs and SMCs that weretransduced or not with an adenovirus expressing MDKD81-121.Wewere able to show that only the coculture of HUVEC and SMCtreated withMDK could trigger the proliferation of HUVECs. Thiseffect was abolished by MDKD81-121.
To complete and confirm our in vitro experiments, we investi-gated the antitumoral efficacy and antiangiogenic properties ofMDKD81-121 using an in vivo therapeutic approach, namely theintramuscular electrotransfer of a plasmid delivering a secretableform of MDKD81-121. A single intramuscular injection of
pcDNA3-MDKD81-121 but not the control plasmid was shownto inhibit primary tumor growth without apparent toxicity. Thisantitumor inhibitory effect was tightly correlated with markedlydecreased intratumoral vascularization and with the detection ofMDKD81-121 immunoreactive material in muscle sections andblood. As MDK is known to both promote proliferation andinhibit caspase-dependent apoptosis (40), MDKD81-121 can beconsidered to induce these two mechanisms, and thereby, toinhibit the proliferation of tumor cells. TheMDKD81-121–treatedtumors apparently failed to establish a vascular network withinthe tumor mass to support rapid and extensive growth, and thisfailure translated into >80% inhibition of tumor growth. Thetreated tumors were shown to be less aggressive and less vascu-larized and to have a higher apoptotic index. This effect wasattributed to the efficient action of MDKD81-121 on endothelialcell growth and migration and on SMCs which participate in theformation to the vascular network.
In conclusion, we showed that specific targeting of MDK byMDKD81-121 results in strong inhibition of a neuroblastomamodel and inhibition of the angiogenesis process. Nevertheless,the fact that cDNA muscle electrotransfer failed to completelyabolish tumorigenesis emphasizes the importance of improvingthe gene transfer method, so that sustained expression canbe achieved for maximum clinical benefits. We also intend tocombine PTN inhibitors or tyrosine kinase inhibitors targetingALK and/or RPTPb/z receptors to enhanceMDK inhibitor potencyand thus improve the clinical outcome of children withneuroblastoma.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: N. Dianat, B. Le Viet, F. GriscelliDevelopment of methodology: B. Le Viet, E. Gobbo, F. GriscelliAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): N. Dianat, B. Le Viet, N. Auger, I. Bieche, F. GriscelliAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): N. Dianat, B. Le Viet, F. GriscelliWriting, review, and/or revision of the manuscript: N. Dianat, B. Le Viet,I. Bieche, A. Bennaceur-Griscelli, F. GriscelliAdministrative, technical, or material support (i.e., reporting or organizingdata, constructing databases): B. Le Viet, E. Gobbo, F. GriscelliStudy supervision: B. Le Viet, I. Bieche, F. Griscelli
AcknowledgmentsThe authors thank Dr. Isabelle Janoueix-Lerosey for providing the neuro-
blastoma cell lines, Dr. Jean B�enard for IGR-N91 cell line, Dr. Georges Uzan forSMCs, and Lorna Saint Ange for editing. The authors also thank Aur�elie Duc�esfor providing pcDNA3-MDKD81-121-MycHis plasmid construct and DelphineDargere for technical assistance with siRNA experiments. L'Institut National dela Sant�e et de la Recherche M�edicale (INSERM), l'Association pour la Recherchesur le Cancer (ARC), and le Minist�ere de l'Enseignement Sup�erieur et de laRecherche are acknowledged for financial support.
Grant SupportThis work was supported by grant from "Fondation ARC pour la recherche
sur le cancer" (grant no. 4882). All the authors were financed by the same grant.The costs of publication of this articlewere defrayed inpart by the payment of
page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received April 28, 2014; revised October 20, 2014; accepted November 5,2014; published OnlineFirst December 9, 2014.
Dianat et al.
Mol Cancer Ther; 14(1) January 2015 Molecular Cancer Therapeutics222
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226

References1. Griscelli F, Li H, Bennaceur-Griscelli A, Soria J, Opolon P, Soria C, et al.
Angiostatin gene transfer: inhibition of tumor growth in vivo by blockage ofendothelial cell proliferation associated with a mitosis arrest. Proc NatlAcad Sci U S A 1998;95:6367–72.
2. Magnon C, Galaup A, Mullan B, Rouffiac V, Bouquet C, Bidart J-M, et al.Canstatin acts on endothelial and tumor cells via mitochondrial damageinitiated through interaction with alphavbeta3 and alphavbeta5 integrins.Cancer Res 2005;65:4353–61.
3. Dkhissi F, Lu H, Soria C, Opolon P, Griscelli F, Liu H, et al.Endostatin exhibits a direct antitumor effect in addition to its anti-angiogenic activity in colon cancer cells. Hum Gene Ther 2003;14:997–1008.
4. Magnon C, Opolon P, Ricard M, Connault E, Ardouin P, Galaup A, et al.Radiation and inhibition of angiogenesis by canstatin synergize to induceHIF-1alpha-mediated tumor apoptotic switch. J Clin Invest 2007;117:1844–55.
5. Tarui T, Miles LA, Takada Y. Specific interaction of angiostatin withintegrin alpha(v)beta(3) in endothelial cells. J Biol Chem 2001;276:39562–8.
6. Sudhakar A, Sugimoto H, Yang C, Lively J, Zeisberg M, Kalluri R. Humantumstatin and human endostatin exhibit distinct antiangiogenic activitiesmediated by alpha v beta 3 and alpha 5 beta 1 integrins. Proc Natl Acad SciU S A 2003;100:4766–71.
7. Galaup A, Opolon P, Bouquet C, Li H, Opolon D, Bissery M-C,et al. Combined effects of docetaxel and angiostatin gene therapyin prostate tumor model. Mol Ther J Am Soc Gene Ther 2003;7:731–40.
8. Griscelli F, Li H, Cheong C, Opolon P, Bennaceur-Griscelli A, VassalG, et al. Combined effects of radiotherapy and angiostatin genetherapy in glioma tumor model. Proc Natl Acad Sci U S A 2000;97:6698–703.
9. Bernard-Pierrot I, Delb�e J, Rouet V, Vigny M, Kerros M-E, Caruelle D,et al. Dominant negative effectors of heparin affin regulatory peptide(HARP) angiogenic and transforming activities. J Biol Chem 2002;277:32071–7.
10. Duc�es A, Karaky R, Martel-Renoir D, Mir L, Hamma-Kourbali Y, Bi�eche I,et al. 16-kDa fragment of pleiotrophin acts on endothelial and breasttumor cells and inhibits tumor development. Mol Cancer Ther 2008;7:2817–27.
11. Iwasaki W, Nagata K, Hatanaka H, Inui T, Kimura T, Muramatsu T, et al.Solution structure of midkine, a new heparin-binding growth factor.EMBO J 1997;16:6936–46.
12. Bernard-Pierrot I,Delbe J, CaruelleD, BarritaultD,Courty J,Milhiet PE. Thelysine-richC-terminal tail of heparin affin regulatory peptide is required formitogenic and tumor formation activities. J Biol Chem 2001;276:12228–34.
13. Stoica GE, Kuo A, Powers C, Bowden ET, Sale EB, Riegel AT, et al. Midkinebinds to anaplastic lymphoma kinase (ALK) and acts as a growth factor fordifferent cell types. J Biol Chem 2002;277:35990–8.
14. M€uller S, Kunkel P, Lamszus K, Ulbricht U, Lorente GA, Nelson AM, et al. Arole for receptor tyrosine phosphatase zeta in glioma cell migration.Oncogene 2003;22:6661–8.
15. Chen S, Bu G, Takei Y, Sakamoto K, Ikematsu S, Muramatsu T, et al.Midkine and LDL-receptor-related protein 1 contribute to the anchor-age-independent cell growth of cancer cells. J Cell Sci 2007;120:4009–15.
16. Kurtz A, Schulte AM, Wellstein A. Pleiotrophin and midkine innormal development and tumor biology. Crit Rev Oncog 1995;6:151–77.
17. Friedrich C, Holtkamp N, Cinatl JJr, Sakuma S, Mautner VF, Wellman S,et al. Overexpression of Midkine in malignant peripheral nerve sheathtumor cells inhibits apoptosis and increases angiogenic potency. Int JOncol 2005;27:1433–40.
18. Choudhuri R, Zhang HT, Donnini S, Ziche M, Bicknell R. An angiogenicrole for the neurokinesmidkine and pleiotrophin in tumorigenesis. CancerRes 1997;57:1814–9.
19. Ikematsu S, Nakagawara A, Nakamura Y, Sakuma S, Wakai K, Mura-matsu T, et al. Correlation of elevated level of blood midkine withpoor prognostic factors of human neuroblastomas. Br J Cancer 2003;88:1522–6.
20. Janoueix-Lerosey I, Lequin D, Brugi�eres L, Ribeiro A, de Pontual L, Com-baret V, et al. Somatic and germline activatingmutations of the ALK kinasereceptor in neuroblastoma. Nature 2008;455:967–70.
21. Ferrandis E, Da Silva J, Riou G, B�enard I. Coactivation of the MDR1 andMYCN genes in human neuroblastoma cells during the metastatic processin the nude mouse. Cancer Res 1994;54:2256–61.
22. Martín de Llano JJ, Fuertes G, García-Vicent C, Torr�o I, Fayos JL, Lurbe E.Procedure to consistently obtain endothelial and smooth muscle cellcultures from umbilical cord vessels. Transl Res J Lab Clin Med2007;149:1–9.
23. Sumi Y, Muramatsu H, Takei Y, Hata K-I, Ueda M, Muramatsu T.Midkine, a heparin-binding growth factor, promotes growth and gly-cosaminoglycan synthesis of endothelial cells through its action onsmooth muscle cells in an artificial blood vessel model. J Cell Sci2002;115:2659–67.
24. SouttouB, JuhlH,Hackenbruck J, R€ockseisenM,KlompHJ, RaulaisD, et al.Relationship between serum concentrations of the growth factor pleio-trophin and pleiotrophin-positive tumors. J Natl Cancer Inst 1998;90:1468–73.
25. Souli�e P, H�eroult M, Bernard-Pierrot I, Caruelle D, Oglobine J,Barritault D, et al. Correlation of elevated plasma levels of twostructurally related growth factors, heparin affin regulatory peptideand midkine, in advanced solid tumor patients. Cancer Detect Prev2004;28:319–24.
26. RuanM, Ji T,WuZ, Zhou J, ZhangC. Evaluationof expression ofmidkine inoral squamous cell carcinoma and its correlation with tumour angiogen-esis. Int J Oral Maxillofac Surg 2007;36:159–64.
27. Kaifi JT, Fiegel HC, Rafnsdottir SL, Aridome K, Schurr PG, Reichelt U, et al.Midkine as a prognostic marker for gastrointestinal stromal tumors.J Cancer Res Clin Oncol 2007;133:431–5.
28. Jin Z, Lahat G, Korchin B, Nguyen T, Zhu Q-S, Wang X, et al. Midkineenhances soft-tissue sarcoma growth: a possible novel therapeutic target.Clin Cancer Res 2008;14:5033–42.
29. Czubayko F, Schulte AM, Berchem GJ, Wellstein A. Melanoma angio-genesis and metastasis modulated by ribozyme targeting of the secret-ed growth factor pleiotrophin. Proc Natl Acad Sci U S A 1996;93:14753–8.
30. NakagawaraA,Milbrandt J,Muramatsu T,Deuel TF, ZhaoH,CnaanA, et al.Differential expression of pleiotrophin and midkine in advanced neuro-blastomas. Cancer Res 1995;55:1792–7.
31. Ikematsu S, Nakagawara A, Nakamura Y, Ohira M, Shinjo M, Kishida S,et al. Plasma midkine level is a prognostic factor for human neuroblasto-ma. Cancer Sci 2008;99:2070–4.
32. Stoica GE, Kuo A, Aigner A, Sunitha I, Souttou B, Malerczyk C, et al.Identification of anaplastic lymphoma kinase as a receptor for the growthfactor pleiotrophin. J Biol Chem 2001;276:16772–9.
33. Osajima-Hakomori Y, Miyake I, Ohira M, Nakagawara A, Nakagawa A,Sakai R. Biological role of anaplastic lymphoma kinase in neuroblastoma.Am J Pathol 2005;167:213–22.
34. Lamant L, Pulford K, Bischof D, Morris SW, Mason DY, Delsol G, et al.Expression of the ALK tyrosine kinase gene in neuroblastoma. Am J Pathol2000;156:1711–21.
35. Bermek O, Diamantopoulou Z, Polykratis A, Dos Santos C, Hamma-Kourbali Y, Burlina F, et al. A basic peptide derived from the HARP C-terminus inhibits anchorage-independent growth of DU145 prostate can-cer cells. Exp Cell Res 2007;313:4041–50.
36. Zhang N, Deuel TF. Pleiotrophin and midkine, a family of mitogenic andangiogenic heparin-binding growth and differentiation factors. Curr OpinHematol 1999;6:44–50.
37. Polykratis A, Katsoris P, Courty J, Papadimitriou E. Characterization ofheparin affin regulatory peptide signaling inhuman endothelial cells. J BiolChem 2005;280:22454–61.
38. Takenaka H, Horiba M, Ishiguro H, Sumida A, Hojo M, Usui A, et al.Midkine prevents ventricular remodeling and improves long-term survivalafter myocardial infarction. Am J Physiol Heart Circ Physiol 2009;296:H462–9.
39. Karaky R, Gobbo E, Opolon P, Delb�e J, Courty J, Griscelli F, et al.HARPD111-136 enhances radiation-induced apoptosis of U87MG glio-blastoma by induction of the proapoptotic protein CHOP. Int J Oncol2011;38:179–88.
Inhibition of Neuroblastoma Development by Targeting Midkine
www.aacrjournals.org Mol Cancer Ther; 14(1) January 2015 223
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226

40. OwadaK, SanjoN,Kobayashi T,MizusawaH,MuramatsuH,MuramatsuT,et al. Midkine inhibits caspase-dependent apoptosis via the activation ofmitogen-activated protein kinase and phosphatidylinositol 3-kinase incultured neurons. J Neurochem 1999;73:2084–92.
41. Li Z, Tan F, Liewehr DJ, Steinberg SM, Thiele CJ. In vitro and in vivoinhibition of neuroblastoma tumor cell growth by AKT inhibitor perifo-sine. J Natl Cancer Inst 2010;102:758–70.
42. Goldschneider D, Horvilleur E, Plassa L-F, Guillaud-Bataille M, Million K,Wittmer-Dupret E, et al. Expression of C-terminal deleted p53 isoforms inneuroblastoma. Nucleic Acids Res 2006;34:5603–12.
43. Mergui X, Leteurtre F, Lipinski M, B�enard J, Amor-Gu�eret M. Two distinctlyaltered cellular responses to DNA double-strand breaks in human neuro-blastoma. Biochimie 2008;90:1656–66.
44. Davidoff AM, Pence JC, Shorter NA, Iglehart JD, Marks JR. Expression ofp53 in human neuroblastoma- and neuroepithelioma-derived cell lines.Oncogene 1992;7:127–33.
45. Carr J, Bell E, Pearson ADJ, Kees UR, Beris H, Lunec J, et al. Increasedfrequency of aberrations in the p53/MDM2/p14(ARF) pathway inneuroblastoma cell lines established at relapse. Cancer Res 2006;66:2138–45.
Mol Cancer Ther; 14(1) January 2015 Molecular Cancer Therapeutics224
Dianat et al.
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226

2015;14:213-224. Published OnlineFirst December 9, 2014.Mol Cancer Ther Noushin Dianat, Barbara Le Viet, Emilie Gobbo, et al. Neuroblastoma Tumor Cells and Inhibits Tumor DevelopmentMidkine Lacking Its Last 40 Amino Acids Acts on Endothelial and
Updated version
10.1158/1535-7163.MCT-14-0226doi:
Access the most recent version of this article at:
Cited articles
http://mct.aacrjournals.org/content/14/1/213.full#ref-list-1
This article cites 45 articles, 20 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mct.aacrjournals.org/content/14/1/213To request permission to re-use all or part of this article, use this link
on June 5, 2018. © 2015 American Association for Cancer Research. mct.aacrjournals.org Downloaded from
Published OnlineFirst December 9, 2014; DOI: 10.1158/1535-7163.MCT-14-0226