measurements and models of primary productivity
TRANSCRIPT

Cullen C-MORE 2014
1Provided by the SeaWiFS Project, NASA/Goddard Space Flight Center and ORBIMAGE
Measurements and Models of Primary Productivity
Supported by NSERC including OTN
John J. Cullen !
Department of Oceanography, Dalhousie University Halifax, Nova Scotia, Canada B3H 4R2
!2014 C-MORE Summer Training Course
“Microbial Oceanography: Genomes to Biomes”

John Cullen: C-MORE 2013
Source of some reading
2
http://www.ceotr.ca/publications/
for reading lists, [email protected]
MacIntyre and Cullen 2005
Parkhill et al. 2001

Cullen C-MORE 2014
3
What is marine primary productivity?
Net Primary Productivity (Production)!!
Net rate of synthesis of organic material from inorganic compounds such as CO2 and water!
!Chemosynthesis: chemical reducing power comes from reduced inorganic compounds such as H2S and NH3!!Photosynthesis: reducing power comes from light energy!!Photosynthetic primary production is usually measured and considered to dominate. !!
g C m-3 h-1 !! !g C m -2 d-1!

Cullen C-MORE 2014
4
More to it than that, of course
Doney, S. C.,et al. (2004), Frontiers in Ecology and the Environment, 2(9), 457-466.

John Cullen: C-MORE 2013
Biological oceanography and phytoplankton ecologyDescribe the causes and consequences of variations in
primary productivity (and food web structure)

Cullen C-MORE 2014
6
Ecological prediction!
Biogeochemical models!
Climate change scenarios
Why measure and model primary productivity?
Behrenfeld et al. Nature 2006

Cullen C-MORE 2014
7
Oxygenic Photosynthesis
This process can be quantified directly by measuring the increase of oxygen, the decrease of CO2, or the increase of organic carbon. Incubations can be conducted with 14C, 13C or H218O added as tracers.!!The process has many chemical consequences that can be traced with measurements of the concentrations of oxygen and carbon, and their isotopic signatures !!

Cullen C-MORE 2014Annual Review of Marine Science 2013
There are measurements and there are measurements…

Cullen C-MORE 2014
in vitro vs in situ

Cullen C-MORE 2014
In situ estimates need calibration

Cullen C-MORE 2014
Cullen, J.J., 2001. Plankton: Primary production methods. In J. Steele, S. Thorpe, K. Turekian (Eds.), Encyclopedia of Ocean Sciences (pp. 2277-2284): Academic Press.

Biological Oceanography (4): John Cullen
Effects of Light on PhotosynthesisNet Photosynthesis = Gross Photosynthesis - Respiration
Respiration is measured directly only in the dark. The light-dependent component is much harder to measure (18O-tracer can be used for that). 12

Biological Oceanography (4): John Cullen
Effects of Light on PhotosynthesisNet Photosynthesis = Gross Photosynthesis - Respiration
Net photosynthesis is negative when irradiance is below the compensation irradiance.13

Biological Oceanography (4): John Cullen
Relating oxygen evolution to carbon assimilation with the photosynthetic quotient
Note that more photosynthesis is required for growth on nitrate because the nitrate must be reduced.
1.0 NO3- + 5.7CO2 + 5.4H 2O! → ! (C5.7H9.8O2.3N) + 8.5 O2 +1.0 OH-
P.Q. = 1.49 (O2 evolved /CO2 consumed)
1.0 NH4+ + 5.7CO2 + 3.4H2O ! → ! (C5.7H9.8O2.3N) + 6.25 O2 +1.0 H+
P.Q. = 1.10
Generalized reactions for growth on nitrate and ammonium
14

Cullen C-MORE 2014
15
Inevitable comparisons with satellite-based estimates (models)

Cullen C-MORE 2014
16
What is behind this?

Cullen C-MORE 2014
17
Still a benchmark: The 14C method for measuring primary productivity
HOT website

Cullen C-MORE 2014
18
0
10
20
30
40
50
0 1 2 3 4 5D
epth
(m)
Productivity (mgC m-3 d-1)
± s.e.
The 14C method for measuring primary productivity

Cullen C-MORE 2014
19
An incredibly useful tool for time series and process studies
HOT website

Cullen C-MORE 2014
20
Ideally, the 14C method measures net primary productivity
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)
Productivity (mgC m-3 d-1)
± s.e.
12 - 24 h incubations
Sometimes, it does

Cullen C-MORE 2014
21
The measurement is subject toartifacts and biases
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)
Productivity (mgC m-3 d-1)
± s.e.
12 - 24 h incubations
Hypothesis: uncertainties inherent in the 14C method are being forgotten with important consequences for the estimation of primary productivity using other methods.
Cullen, J.J., 2001. Plankton: Primary production methods. In J. Steele, S. Thorpe, K. Turekian (Eds.), Encyclopedia of Ocean Sciences (pp. 2277-2284): Academic Press.

Cullen C-MORE 2014
22
The measurement is subject toartifacts and biases
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)
Productivity (mgC m-3 d-1)
± s.e.
12 - 24 h incubations
Overestimation due to exclusion of UV-B

Cullen C-MORE 2014
23
The measurement is subject toartifacts and biases
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)
Productivity (mgC m-3 d-1)
± s.e.
12 - 24 h incubations
Underestimation due to static incubation at excessive irradiance

Cullen C-MORE 2014
24
The measurement is subject toartifacts and biases
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)
Productivity (mgC m-3 d-1)
± s.e.
12 - 24 h incubations
Underestimation due to dilution of intracellular DIC with respired cellular C

Cullen C-MORE 2014
25
The measurement is subject toartifacts and biases
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)
Productivity (mgC m-3 d-1)
± s.e.
12 - 24 h incubations
Overestimation due to unnatural accumulation of biomass (disruption or exclusion of grazers)

Cullen C-MORE 2014
26
The measurement is subject toartifacts and biases
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)
Productivity (mgC m-3 d-1)
± s.e.
12 - 24 h incubations
Underestimation due to food-web cycling!(microbial respiration and excretion)

Cullen C-MORE 2014
27
The measurement is subject toartifacts and biases
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)
Productivity (mgC m-3 d-1)
12 - 24 h incubations
Overestimation due to inadequate time for respiration to be measured

Cullen C-MORE 2014
28
…and that’s not all:
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)
Productivity (mgC m-3 d-1)
12 - 24 h incubations
Toxicity!
Relief of iron limitation!
Exposure to bright light!
Disruption of fragile cells!for Simulated in situ:!
Poor match of irradiance!
Inappropriate temperature!
Possible diel bias

Cullen C-MORE 2014
29
Productivity normalized to Chl is biased by:
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)
Productivity (mgC m-3 d-1)
12 - 24 h incubations
Changes in Chl during incubations!
Inadequate extraction of Chl by 90% acetone!
Interference from Chl b(fluorometric acid-ratio method)

Cullen C-MORE 2014
30
and in general…
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)
Productivity (mgC m-3 d-1)
12 - 24 h incubations
Irradiance is not controlled!
Respiration is not accurately measured

Cullen C-MORE 2014
31
The measurement has its limitations
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)Productivity (mgC m-3 d-1)
± s.e.
Net production?
!Gross production?
!Overestimate?
!Underestimate?

Cullen C-MORE 2014
A less skeptical view

Cullen C-MORE 2014
33
But we use it routinely
0
10
20
30
40
50
0 1 2 3 4 5
Dep
th (m
)
Productivity (mgC m-3 d-1)
± s.e.
Water column integral is something like !net primary production!(photosynthesis less losses to respiration)
(extent depends on temperature, light and maybe nutrients)

Cullen C-MORE 2014
34
Regardless, it has served us very well
Time Series
Karl et al. in Williams et al. 2002

Cullen C-MORE 2014
35
Process studies

Cullen C-MORE 2014
36
Solving mysteries
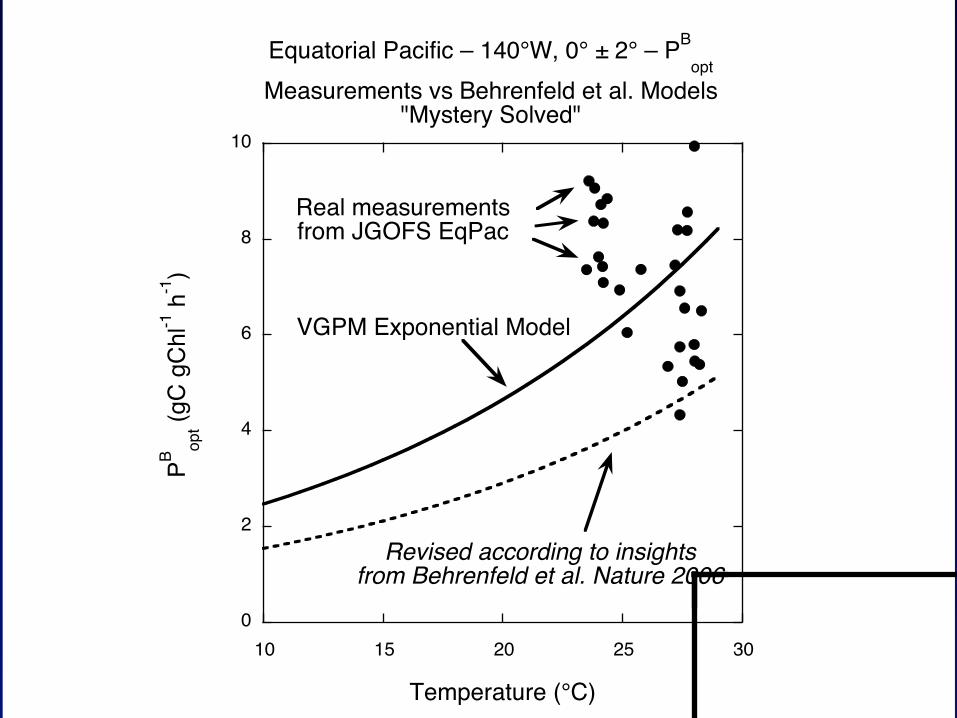
Cullen C-MORE 2014
0
2
4
6
8
10
10 15 20 25 30
Equatorial Pacific – 140°W, 0° ± 2° – PBopt
Measurements vs Behrenfeld et al. Models"Mystery Solved"
PB opt (g
C gC
hl-1
h-1
)
Temperature (°C)
VGPM Exponential Model
Revised according to insightsfrom Behrenfeld et al. Nature 2006
Real measurementsfrom JGOFS EqPac

Cullen C-MORE 2014
38
Direct Measurements will Never Provide Synoptic Estimates of
Productivity
marine.rutgers.edu/opp/

Cullen C-MORE 2014
39
Continuous measures of optics reveal rates
see

Cullen C-MORE 2014
40
Ocean chemistry tells the tale — even in the most oligotrophic waters
3 years of O2 at POT: Ken Johnson

Cullen C-MORE 2014
41
Models are required for many applications
Productivity from ocean color!
Ecological prediction!
Biogeochemical models!
Climate change scenarios

Cullen C-MORE 2014
42
Photosynthesis normalized to chlorophyll is the foundation of productivity modeling
Pmaxchl ⋅ (1− exp −(α chl ⋅E)
Pmaxchl
$%&
'())
Pchl vs. E relationship! "##### $#####
Behrenfeld and Falkowski L&O 1997

Cullen C-MORE 2014
43
One approach: model the measurements
Behrenfeld and Falkowski 1997a L&O
! Opt
ical
Dep
th

Cullen C-MORE 2014
44
Simplified functions describe major patterns
Behrenfeld and Falkowski 1997a L&O

Cullen C-MORE 2014
45
Is the measured/modeled pattern real?
0
20
40
60
80
100
0 10 20 30 40
Dep
th (m
)
PB (mg C mg Chl-1 d-1)
Measured near-surface inhibition

Cullen C-MORE 2014
46
Is the measured/modeled pattern real?
0
20
40
60
80
100
0 10 20 30 40
Dep
th (m
)
PB (mg C mg Chl-1 d-1)
or an artifact of fixed-depth incubation?

Cullen C-MORE 2014
47
Measurements must be made on a shorter time scale
Photosynthetron: Controlled laboratory incubation (Lewis and Smith 1983)

Cullen C-MORE 2014
48
A comprehensive approach
Approach introduced by Jitts, H. R., A. Morel, and Y. Saijo. 1976. The relation of oceanic primary production to available photosynthetic irradiance. Aust. J. Mar. Freshwater Res. 27: 441-454.
Cullen, J. J., M. R. Lewis, C. O. Davis, and R. T. Barber. 1992. Photosynthetic characteristics and estimated growth rates indicate grazing is the proximate control of primary production in the equatorial Pacific. Journal of Geophysical Research 97: 639-654.

Cullen C-MORE 2014
49
0
10
20
30
40
50
0 0.08 0.2
Dep
th (m
)
Chlorophyll a (mg m-3)
0
10
20
30
40
50
0 0.4 0.8
Dep
th (m
)
Short-term measurements can detect near surface inhibition
0630 h: Chlorophyll and Pmax are nearly uniform in the 50-m mixed layer
MORNING
PBm . Chl (mg C m-3 h-1)
Principles described by Marra, Harris, Yentsch -- all prior to 1980

Cullen C-MORE 2014
50
0
10
20
30
40
50
0 0.08 0.2
Dep
th (m
)
Chlorophyll a (mg m-3)
0
10
20
30
40
50
0 0.4 0.8
Dep
th (m
)
PBm . Chl (mg C m-3 h-1)
1215 h: Chlorophyll and Potential Productivity are still nearly uniform in the 50-m mixed layer
Is measured near-surface inhibition real?
Assessment with short-term P vs E
MIDDAY

Cullen C-MORE 2014
51
0
10
20
30
40
50
0 0.08 0.2
Dep
th (m
)
Chlorophyll a (mg m-3)
0
10
20
30
40
50
0 0.4 0.8
Dep
th (m
)
Surface incubation led to artifact
1530 h: Fixed-depth incubations at the surface are “fried” — an artifact of sustained exposure 35% underestimation of ML productivity
INCUBATED
PBm . Chl (mg C m-3 h-1)

Cullen C-MORE 2014
52
Conventional 14C: Near-surface inhibition is overestimated
Error at the surface: big
Error for integral: 8%
0
20
40
60
80
100
0 10 20 30 40
Dep
th (m
)
PB (mg C mg Chl-1 d-1)
Artifact

Cullen C-MORE 2014
53
Parameterization of an Artifact?
Consequences for a model: minor except for irradiance dependence of ΣP at higher daily irradiance
Behrenfeld and Falkowski Consumer’s Guide: L&O 1997

Cullen C-MORE 2014
54
Many models calculate P from measured relationships
From! ! PB vs E!
and ! E vs depth!
and ! ! Chl!
to! daily water column net primary productivity
0
20
40
60
80
100
0 0.2 0.4 0.6 0.8 1
P (mg C m-3 d-1)
Dept
h (m
)

Cullen C-MORE 2014
55
Results can be generalized
0
0.2
0.4
0.6
0.8
1
0 0.2 0.4 0.6 0.8 1
PB / PBopt
Z /
Z 1%(P
AR)
€
PZ =PoptB ⋅BKPAR
⋅ f (EPAR (0)Ek)

Cullen C-MORE 2014
56
Maximum PB is still a key parameter for most models
0
20
40
60
80
100
0 2 4 6 8 10PB (g C g Chl-1 h-1)
Z (m
)
Even the spectral ones — even the quantum-yield models

Cullen C-MORE 2014
57
Compared to other measures, PBopt is
relatively insensitive to artifact
0
20
40
60
80
100
0 10 20 30 40D
epth
(m)
PB (mg C mg Chl-1 d-1)

Cullen C-MORE 2014
58
Global assessment of primary productivity requires global assessment of maximum PB
Behrenfeld and Falkowski 1997b L&O

Cullen C-MORE 2014
59
PBopt modelled as a function of
temperature
Behrenfeld and Falkowski 1997b L&O

Cullen C-MORE 2014
60
Two commonly used functions
Behrenfeld and Falkowski 1997b L&O
0
2
4
6
8
10
0 5 10 15 20 25 30
Max
imum
PB (g
C g
Chl-1
h-1
)
Temperature (°C)
B & F VGPM
Eppley - type(e.g., Antoine et al 1996)

Cullen C-MORE 2014
61
You may have seen the figure
Behrenfeld and Falkowski 1997b L&O

Cullen C-MORE 2014
62
Here are the measurements
Rutgers OPP Study Data Base
0
5
10
15
20
25
30
35
40
0 5 10 15 20 25 30
Max
imum
PB (g
C g
Chl-1
h-1
)
Temperature (°C)

Cullen C-MORE 2014
63
Compared with more measurements
Texas Shelf
0
5
10
15
20
25
30
35
40
0 5 10 15 20 25 30
Max
imum
PB (g
C g
Chl-1
h-1
)
Temperature (•C)

Cullen C-MORE 2014
64
and more measurements!
0
5
10
15
20
25
30
35
40
0 5 10 15 20 25 30
Eppley Type ModelVGPMRutgers OPP DataTexas Shelf & SlopeHOTBATSEqPacArabian Sea JGOFSRoss Sea JGOFSAntarctic PFZ JGOFSPBmax Texas (SL)
Max
imum
PB (g
C g
Chl-1
h-1
)
Temperature (•C)

The foundations of VGPM
Behrenfeld and Falkowski 1997 L&O Underlying data + a few more data sets
Published Statistical Fit Measurements (note scale)

Cullen C-MORE 2014
66
General functions of T cannot capture major causes of variability in water column productivity
± 2x error
-3
-2
-1
0
1
2
3
-5 0 5 10 15 20 25 30 35
ln(m
easu
red/
mod
eled
)
Temperature (°C)

Cullen C-MORE 2014
67
Conclusion: General functions of T cannot capture major causes of variability in water column productivity
Relative Error

Cullen C-MORE 2014
68
We must appreciate the implicationsof this unexplained variability

Carbon cancels out!
…and implications of a carbon-based model that isn’t
!Times Cited: 353 (from Google Scholar, May 2014)

Cullen C-MORE 2014
How much confidence should we have?
Juranek and Quay 2013 Ann Rev Mar Sci

Cullen C-MORE 2014
71
Estimates like these (NPP) do not yet account for variable physiology beyond central tendencies
Behrenfeld et al. Nature 2006

Cullen C-MORE 2014
72
The need to compare models with measurements

Cullen C-MORE 2014
73
Validation is important

Cullen C-MORE 2014
74
Model

Cullen C-MORE 2014
75
Headline

Cullen C-MORE 2014
76
Published model result
Measurements!show the !
opposite trend

Cullen C-MORE 2014
77
Time Frame
& Statistics
Matter
Measurements

Cullen C-MORE 2014
78
Measurements !
(no headlines)

Cullen C-MORE 2014
79
It is also useful to remember the distinction between models and observations
Follows et al. ModelVGPM model

Cullen C-MORE 2014
80
Many models cannot be directly verified
0
20
40
60
80
100
1200.4 0.5 0.6 0.7 0.8 0.9 1.0
Mix
ing
Dep
th (m
)
PT / PTpot
20, 40, 60min
Nomixing
8 h4 h
2 h
b
No one is immune!

Cullen C-MORE 2014
81
Conclusions• Models of primary productivity are a fundamental
requirement for describing and explaining ecosystem dynamics and biogeochemical cycling in the sea
• The models are based on measurements: – The measurements are not perfect – The models are not perfect
• Capabilities and limitations of models should always be considered when they are applied
• Effects of underlying assumptions • Comparison with real measurements when available
• Know your model!