maternal deprivation effects on brain plasticity and recognition memory in adolescent male and...
TRANSCRIPT

at SciVerse ScienceDirect
Neuropharmacology 68 (2013) 223e231
Contents lists available
Neuropharmacology
journal homepage: www.elsevier .com/locate/neuropharm
Maternal deprivation effects on brain plasticity and recognition memory inadolescent male and female rats
Eva M. Marco a, Manuel Valero a, Oscar de la Serna b, Barbara Aisa b, Erika Borcel a,Maria Javier Ramirez b,*, María-Paz Viveros a,**
aDepartamento de Fisiología (Fisiología Animal II), Facultad de Ciencias Biológicas, Universidad Complutense de Madrid, Instituto de Investigación Sanitaria del Hospital Clínico SanCarlos (IdISSC), Madrid, SpainbDepartamento de Farmacología, Centro de Investigación Médica Aplicada (CIMA), Universidad de Navarra, Pamplona, Spain
a r t i c l e i n f o
Article history:Received 12 March 2012Received in revised form1 August 2012Accepted 16 August 2012
Keywords:Animal modelMaternal deprivationPsychopathologyCognitionSynaptic plasticityBehaviorSexual dimorphisms
* Corresponding author. Departamento de FarmacoMédica Aplicada (CIMA), Universidad de Navarra, C/IrSpain. Tel.: þ34 948425600; fax: þ34 948425740.** Corresponding author. Departamento de Fisiologultad de CC. Biológicas, Universidad Complutense de MNovais, n� 2, 28040 Madrid, Spain. Tel.: þ34 91 394 4
E-mail addresses: [email protected] (M.J. Ram(M.-P. Viveros).
0028-3908/$ e see front matter � 2012 Elsevier Ltd.http://dx.doi.org/10.1016/j.neuropharm.2012.08.014
a b s t r a c t
Data from both human and animal studies suggest that exposure to stressful life events at neonatal stagesmay increase the risk of psychopathology at adulthood. In particular, early maternal deprivation, 24 h atpostnatal day (pnd) 9, has been associated with persistent neurobehavioural changes similar to thosepresent in developmental psychopathologies such as depression and schizophrenic-related disorders. Mostneuropsychiatric disorders first appear during adolescence, however, the effects of MD on adolescentanimals’ brain and behaviour have been scarcely explored. In the present study, we aimed to investigate theemotional and cognitive consequences of MD in adolescent male and female rats, as well as possibleunderlying neurobiological mechanisms within frontal cortex and hippocampus. Animals were exposed toa battery of behavioural tasks, from pnd 35 to 42, to evaluate cognitive [spontaneous alternation task (SAT)and novel object test (NOT)] and anxiety-related responses [elevated plus maze (EPM)] during adolescence.Changes in neuronal and glial cells, alterations in synaptic plasticity as well as modifications in cannabinoidreceptor expression were investigated in a parallel group of control and adolescent (pnd 40) male andfemale animals. Notably, MD induced a significant impairment in recognition memory exclusively amongfemales. A generalized decrease in NeuN expressionwas found in MD animals, together with an increase inhippocampal glial fibrillar acidic protein (GFAP) expression exclusively among MD adolescent males. Inaddition, MD induced in the frontal cortex and hippocampus of male and female adolescent rats a signif-icant reduction in brain derived neurotrophic factor (BDNF) and postsynaptic density (PSD95) levels,together with a decrease in synaptophysin in frontal cortex and neural cell adhesion molecule (NCAM) inhippocampus. MD induced, in animals of both sexes, a significant reduction in CB1R expression, but anincrease in CB2R that was statistically significant only for the frontal cortex. Taken together, these resultsindicate that adolescent females are more vulnerable than males to the cognitive deficits derived fromMDdespite the changes in neural cells, cannabinoid receptors, as well as the reduction in neural plasticity seemto be similar in both sexes. Further investigation is needed to understand the neurobiological mechanismsunderlying the sexual dimorphisms associated to theMD effects, and thus, for a better understanding of thespecific sex-dependent vulnerabilities to early life stress.
This article is part of the Special Issue entitled ‘Neurodevelopmental Disorders’.� 2012 Elsevier Ltd. All rights reserved.
logía, Centro de Investigaciónunlarrea 1, 31008 Pamplona,
ía (Fisiología Animal II), Fac-adrid, U.C.M., C/Jose Antonio993; fax: þ34 91 394 4935.irez), [email protected]
All rights reserved.
1. Introduction
Increasing evidence gives support to the fact that early-lifestress induces permanent alterations in neurodevelopment(Plotsky and Meaney, 1993) that may increase the risk of psycho-pathology at adulthood (Heim and Nemeroff, 2001; Morgan andFisher, 2007; Talge et al., 2007). Actually, data from both humanand animal studies suggest a solid association between early-lifetrauma and psychopathology (Bremne and Vermetten, 2001;Fatemi and Folsom, 2009; Fumagalli et al., 2007; Teicher et al.,

E.M. Marco et al. / Neuropharmacology 68 (2013) 223e231224
2003). Manipulation of mothereinfant interactions has beenextensively used as animal models of early-life stress (Cirulli et al.,2009). In the rat, separations of pups from their dam are typicallycarried out during the stress-hypo-responsive-period (SHRP),between postnatal days (pnd) 2 and 14. Notably, among thesemodels, maternal deprivation (MD), 24 h at pnd 9, has been used asan animal model of early-life stress highly suitable for the inves-tigation of the developmental origin of certain psychiatric disorderssuch as schizophrenia and depression (Ellenbroek and Riva, 2003;Marco et al., 2009).
MD has been reported to provoke behavioural abnormalitiesthat resemble psychotic-like symptoms such a disruption in thepre-pulse inhibition (PPI) response (Ellenbroek et al., 2005;Ellenbroek and Riva, 2003), neuroendocrine alterations related tostress reactivity (Lehmann et al., 2002; Levine et al., 1991; Rentesiet al., 2010), as well as cognitive impairments in adult animals(Llorente et al., 2011). Despite most neuropsychiatric disorderspresent early developmental onsets (i.e. schizophrenia in youngadulthood) (Paus et al., 2008; Steinberg, 2005) the investigation ofMD effects in adolescent animals has been scarcely explored. Onlyrecent studies have described deviant behaviours in adolescentanimals following neonatal exposure to MD, i.e. depressive-likeresponses (Llorente et al., 2007) and increased impulsivity (Marcoet al., 2007). Nonetheless, the emotional and cognitive effects ofMD in adolescent animals remain unexplored.
Previous studies have shown alterations in several neurotrans-mitter systems, including glutamatergic, dopaminergic and sero-tonergic activity, following MD (Llorente et al., 2010, 2012; Rentesiet al., 2010; Roceri et al., 2002). Neurodegenerative changestogether with alterations in glial cells have been reported shortafter MD in the hippocampus of neonate rat pups (Llorente et al.,2009, 2008). Changes in the levels of brain derived neurotrophicfactor (BDNF), as well as in other markers of synaptic plasticity,have also been observed in the brain of adult MD animals (Llorenteet al., 2011; Roceri et al., 2002). More recently, changes in theendocannabinoid system have been described following early MD(Llorente et al., 2008; Suarez et al., 2009, 2010). However, neitherneuronal, glial cells, markers of synaptic plasticity nor cannabinoidreceptors have been investigated in adolescent animals neonatallyexposed to 24 h of MD.
Notably, important sex differences have been consistentlydescribed in relation to both behavioural and neurobiologicalconsequences of MD. Actually, sex differences have been reportedfrom early neonatal stages (pnd 13) to adulthood (Viveros et al.,2010). Moreover, our present investigation focused on frontalcortex and hippocampus given the central role of these two brainregions in emotional and memory processes (Buffalo et al., 2006;Squire et al., 2007). Consequently, in the present study we inves-tigated the emotional and cognitive consequences of early MD inadolescentmale and female rats, as well as possible neurobiologicalunderling mechanisms within frontal cortex and hippocampus.
2. Material & methods
Experiments are in compliance with the Spanish Royal Decree 1201/2005,October 21, 2005 (BOE n� 252) about protection of experimental animals, in closeagreement with the European Communities Council Directive of 24 November 1986(86/609/EEC). In addition, the local Animal Ethics Committee approved the presentexperimental protocol. All efforts were made to minimise animal suffering anddistress.
2.1. Animals
Experimental subjects were the offspring of Albino Wistar rats purchased fromHarlan Laboratories Inc. Animals were housed in plastic Macrolon� III cages. Theparental generation was mated (one male � two females) in our animal facilitiesapproximately 2 weeks after their arrival. After 10 days, females were isolated andthe day of delivery strictly controlled. On the day of birth ( pnd 0), litters were sex-
balanced and culled to 8 pups per dam (4males and 4 females). Thereafter, dams andlitters were left undisturbed until weaning (pnd 22), when animals were separatedby sex and housed in groups of four siblings per cage. All animals weremaintained ata constant temperature (22 � 1 �C) and humidity (50 � 1%) in a reverse 12-h darkelight cycle (lights on at 20.00 h), with free access to food (commercial diet forrodents A04/A03; SAFE, Augy, France) and water.
2.2. Early maternal deprivation
Early maternal deprivation (MD) was performed as previously described(Llorente et al., 2007). In brief, on pnd 9, litters were submitted to 24 h of maternaldeprivation, i.e. dams were removed from their home-cages at 09.00 h and pupswere left undisturbed (in the same room) until the next day (pnd 10, 09.00 h) whendams were placed back in their corresponding home-cages. Animals from thecontrol group (Co) were submitted to the same manipulation except for thematernal deprivation episode.
2.3. Behavioural testing
2.3.1. Spontaneous alternation test (SAT)Continuous spontaneous alternation was assessed in a symmetrical Y-maze
with three equal arms (40 cm long, 15 cm wide and 30 cm high) made of opaqueblack polyvinyl chloride (PVC) (Hughes, 2004). Spontaneous alternation in the Y-maze is regarded as an index of spatial working memory based on a naturaltendency of rats to explore a less recently visited arm (Dember and Fowler, 1958).Each rat was allowed to freely explore the three arms of the maze for 8 min underdim light conditions. Animals’ behaviour was video recorded (Sony DCR-DVD310E), and later analysed by an experienced observer. The total number ofarm entries was registered, considering an arm entry whenever a rat entered thearm with the four paws. Spontaneous alternation behaviour was defined assuccessive entries into the three arms in overlapping triplet sets, and the alter-nation index calculated as the ratio of actual to possible alternations [(number ofactual alternations/(total number of arm entries � 2)) � 100]. An alternation indexof 22% indicates chance performance. Animals that failed to perform any armalternation, and those entering less than 5 times to the arms were not included inthe statistical analysis.
2.3.2. Novel object test (NOT)The novel object test (NOT) was performed in a squared arena
(60 cm � 60 cm � 45 cm) with matte-painted metallic walls and a plastic-coveredwooden floor divided into 36 squares (10 cm � 10 cm) by white painted lines. Thetest was performed as previously described by Ennaceur and Delacour (1988) withsome minor modifications (Mateos et al., 2011). Animals were allowed to freelyexplore the arena, under dim light conditions, for 5 min during 3 consecutive days(habituation period). On the first day of habituation, animals’ general activity wasevaluated by direct observation. Total ambulation (horizontal motor activity), wascalculated as peripheral (by the walls) and internal (by the squares not adjacent tothe walls) line crossings. Rearing frequency (vertical motor activity) and percentageof internal ambulation, considered as an index of emotionality (Archer, 1973) andcalculated as the percentage of internal ambulation refereed to total ambulation[(internal line crossings/total line crossings) � 100], were also analysed. On the testday, rats were firstly exposed to two identical objects (two plastic boxes) during3 min, training session. Rats were thereafter exposed, following a 1 h inter-trialinterval, to one of the previously encountered objects (familiar object, F1 or F2)and to a novel, non-familiar object (metallic coloured box, N), test session. Objectswere not bigger than twice the size of a rat, and were located in contiguouscorners, at a distance of 10 cm from the walls. For each animal, the position of theobjects was not changed between the training and the test session. However,objects’ position was changed between animals in order to avoid spatial preference.Both training and test sessions were video recorded (Sony DCR-DVD310E), andanimals’ behaviour was later evaluated by an experienced observer by means of anevent-recorder software (RCV, Cibertec, Madrid, Spain). Time spent exploring theobjects during the two sessions was registered. Exploration of an object wasconsidered whenever animals pointed their nose toward an object at a distance�1 cm, whereas turning around, climbing and/or biting the objects were notconsiderer as exploration (Barker et al., 2007; Dix and Aggleton, 1999; Mitchell andLaiacona, 1998; Rossato et al., 2007). In the test session, the discrimination index(DI) was calculated as the difference between the time spent exploring the novelobject (N) and the familiar one (F1 or F2) in relation to the total time spentexploring the objects [(N � F)/(N þ F)]. Animals that during the training and/or thetest session explored the two objects less than 10 s were excluded from thestatistical analysis, as did animals that explored less than 1 s one of the objectsduring the test session. Since animals’ capacity to discriminate between the noveland the familiar objects diminishes with time (Dix and Aggleton, 1999; Mumbyet al., 2002), and in the present conditions animals explored the objects duringthe first minute more that half the time spent in exploration during the wholetesting session, data from the first minute of the test session were employed for thestatistical analyses.

E.M. Marco et al. / Neuropharmacology 68 (2013) 223e231 225
2.3.3. Elevated plus-maze (EPM)The elevated plus-maze (EPM) was formed by two open arms (50 cm � 10 cm)
and two equally sized enclosed arms with 40 cm high walls, arranged so that thearms of the same type are opposite to each other. The junction of the four armsformed a central square area (10 cm� 10 cm). The apparatus was made of black hardplastic material and elevated to a height of 62 cm. Animals were allowed to freelyexplore the maze for 5 min under dim red light conditions. Frequency and durationof open and closed arm visits were separately recorded, considering an arm visitwhenever an animal entered it with the four limbs. Percentages of open arm entriesand time spent in the open arms were calculated as referred to total arm entries andtotal time in arms, respectively, and were considered as the most relevant param-eters related to anxiety. In contrast, total arms entries were considered as an index ofgeneral motor activity (Hogg, 1996; Pellow et al., 1985). Animals that fell off the EPMwere not included in the statistical analysis.
In the different behavioural tests, animals were placed in a neutral part of thearena (e.g. facing an enclosed arm or opposite to the objects) so that animals werenot artificially induced to perform a significant pattern. After each trial, animalswere placed back into their home cageswith littermates and both arenas and objectswere thoroughly cleaned with ethanol (20%) after each session in order to removeany odour contamination.
2.4. Brain biochemical analysis
2.4.1. Western blottingBrain samples were homogenized in a 50 mM Tris buffer (pH 7.2, 4 �C), and each
sample adjusted to a final protein concentration of 4 mg/ml (DC protein assay; Bio-Rad, Hercules, CA). Extracts were mixed with Laemmeli’s sample buffer boiled for5 min. Samples (30 mg) were loaded into 7.5% bisacrylamide gels and separated bySDS-PAGE. Separated proteins were electrophoretically transferred from gels tonitrocellulose membranes. Proteins of interest were detected by specific antibodies(see Table 1 for details). b-actin was used as internal control. Immunopositive bandswere visualized by a chemiluminescent method (ECL; Amersham, Arlington Heights,IL). The optical density (O.D.) of reactive bands visible on x-ray filmwas determineddensitometrically. In each region, results were expressed as percentage of O.D.values from control male rats.
2.4.2. ELISACrude synaptosomal pellets were obtained (Sandi et al., 2005), and levels of
neural cell adhesion molecule (NCAM) measured by ELISA (Venero et al., 2006). Inbrief, polystyrene flat bottom 96-well microplates were allowed to adsorb a coatingsolution (Na2CO3 0.1 M/NaHCO3, 0.1 M) for 2 h at room temperature. The solutionwas removed and 50 mL of pellet samples added to each well at a concentration of10 mg/mL. Plates were incubated overnight at 4 �C and thenwashed three times with1 M PBS containing 0.05% Tween 20, pH 7.4. Additional binding sites were blockedwith BSA (3%) for 2 h at room temperature.Wells were incubatedwith 50 mL aliquotsof primary antibody Ab5032 (1:15000 TBST; Chemicon, USA) for 20e24 h at 4 �C andsubsequently, 50 mL aliquots of anti-rabbit IgG peroxidase conjugate antibody(1:500; SigmaeAldrich, St. Louis, MO, USA) were added for a 2 h incubation period.TMB (Promega, Madison, WI, USA) was used as a chromogenic substrate. Thereactionwas terminated by the addition of 1 N hydrochloric acid. Absorbancies weremeasured at 450 nm using an automatic ELISA microplate reader. Results wereexpressed as a percentage of change from control males.
Table 1Specific antibodies used in Western blotting to detect proteins of interest.
Protein Primary antibody Secondaryantibody
BDNF, marker of synapticplasticity
Anti-BDNF (1:1000)a Rabbit (1:10000)
Synaptophysin, marker of pre-synaptic density
Anti-Sinaptophysin(1:1000)a
Rabbit (1:10000)
PSD95, marker of post-synapticdensity
Anti-PSD95 (1:1000)a Mouse (1:10000)
GFAP, glial marker Anti-GFAP (1:1000)a Mouse (1:10000)NeuN, neuronal marker Anti-NeuN (1:1000)a Mouse (1:10000)APC, marker of mature
oligodendrocytesAnti-APC (1:1000)b Rabbit (1:10000)
NG2, marker of oligodendrocyteprecursor cells
Anti-NG2 (1:1000)b Mouse (1:10000)
CB1, Cannabinoid receptortype-1
Anti-CB1 (1:500)c Rabbit (1:5000)
CB2, Cannabinoid receptortype-2
Anti-CB2 (1:500)c Rabbit (1:5000)
Actin Anti-Actin (1:10000)d Mouse (1:10000)
Source of antibodies: aAbcam Inc., Cambridge, MA, USA; bCell Signaling Technology,Beverly, MA, USA; cPierce, Rockford, LI, USA; dSigmaeAldrich, St. Louis, MO, USA.Secondary antibody came from DakoCytomation, Glostrup, Denmark.
2.5. Experimental design
Separate batches of animals were used for the behavioural testing andbiochemical analysis. 1) In a first batch, a total of 48 rats coming from 6 litters wereused. Half of the litters were exposed to the MD protocol, while the other half wasdevoted to the control (Co) group. Male and female rats were exposed to the batteryof behavioural testing throughout adolescence: SAT at pnd 35, NOT at pnd 40 andEPM at pnd 42. Body weight was registered from both Co and MD rat pups at pnd 9,10 and every 6 days until pnd 34 at 09.00 h, approximately. 2) In the second batch,additional 48 rats coming from 6 litters were used. Half of the litterswere exposed tothe MD protocol, while the other half was devoted to the control (Co) group. Nobehavioural analyses were performed, and animals were sacrificed by decapitationat pnd 40. Brains were rapidly extracted; frontal cortex and hippocampi bilaterallydissected on ice, and samples were then frozen in dry ice and stored at �80 �C untilthe biochemical analysis.
2.6. Statistical analyses
Behavioural and biochemical data were analysed using a two-way analysis ofvariance (ANOVA), considering sex (male or female) and neonatal condition (Co orMD) as independent factors. ANOVA for repeated measures were used for theanalysis of age-dependent body weight changes. Saphiro-Wilk and Levene testswere used to confirm normality and homocedasticity of the data. Tukey post hoccomparisons were performed only for significant interactions, as well as additionalone-way ANOVAs. Significance level was set at p < 0.05 for all comparisons.Statistical analyses were performed by the SPSS 19.0 software package (SPSS Inc.,Chicago, IL, USA).
3. Results
3.1. Body weight
No differences in bodyweight were observed at the beginning ofthe experiments (pnd 9) [Co-males, 20.60 � 0.67 g, MD-males,19.97 � 0.61 g and Co-females, 19.97 � 0.73 g, MD-females,19.60 � 0.61 g]. When analysing pre-weaning body weight data,Mauchly’s test indicated that the assumption of sphericity had beenviolated [two-way repeated measures ANOVA, c2 (2) ¼ 14.34,p < 0.05]; therefore, degrees of freedom were corrected usingGreenhouse-Geisser estimates of sphericity (ε ¼ 0.53). The two-way repeated measures ANOVA showed a significant overalleffect of age [F(1.07, 8.55) ¼ 597.29, p < 0.001] as well as a signifi-cant age by neonatal condition interaction [F(1.07, 8.55) ¼ 20.83,p ¼ 0.001]. Tukey post-hoc comparisons within each day of agerevealed significant differences between Co and MD animalsamong each sex condition at pnd 16 (see Fig. 1).
Regarding the analysis of post-weaning body weight data, sphe-ricity could be neither assumed [two-way repeated measuresANOVA, c2 (2) ¼ 32.20, p < 0.01] and degrees of freedomwere cor-rected using Greenhouse-Geisser estimates of sphericity (ε ¼ 0.66).In this occasion, the two-way repeated measures ANOVA showeda significant overall effect of age F(1.32, 57.9)¼ 6090.17, p< 0.001] aswell as significant age per sex [F(1.32, 57.9) ¼ 31.95, p < 0.001] andage per neonatal condition [F(1.32, 57.9) ¼ 8.85, p ¼ 0.002] interac-tions. Tukey post-hoc comparisons within each day of age revealedsignificant differences between Co and MD animals among each sexcondition at every day age analysed, and a significant sex effectsappeared on the last day of measures, pnd 34 (see Fig. 1).
Taken together, the neonatal stressful condition of MD induceda reduction in body weight that achieved statistical significancefrom pnd 16 and lasted until the end of measurements, pnd 34, atearly adolescence. In contrast, sex differences only became evidentat adolescence, pnd 34, with females weighting significantly lessthat their sibling males.
3.2. Spontaneous alternation task (SAT)
No differences in spontaneous alternationwere observed due tosex or as a consequence of the neonatal manipulation at this age

Fig. 1. e Body weight gain (g.) of control (Co) and maternally deprived (MD, 24 h onpnd 9) animals from neonatal (pnd 9) to adolescence (pnd 34). Mean � SEM of male(upper panel) and female (lower panel) animals; pnd, postnatal day. Two-wayrepeated measures ANOVA, *p < 0.05, **p < 0.01, ***p < 0.001 denotes a significantdifference between MD vs. Co rats at each age; {p < 0.01 denotes a significantdifference between male and female rats.
E.M. Marco et al. / Neuropharmacology 68 (2013) 223e231226
(pnd 35), nor a significant interaction between these factors.Similarly, the two-way ANOVA from the total arm entries analysisrendered no significant differences, but a trend for an effect of theneonatal condition [two-way ANOVA, F(1,39)¼ 4.09, p¼ 0.05]; thatappeared to be stronger among female animals (Fig. 2). In resume,neither sex nor neonatal condition seemed to have affectedanimals’ SAT performance; no differences inworkingmemorywere
Fig. 2. e Spontaneous alternation test (SAT). Spontaneous arm alternation (upperpanel) and total arm entries (bottom panel) were registered in control (Co) and MDanimals at postnatal day 35. Mean � SEM (n ¼ 9e12 animals per experimental group).
observed in this study, and only a subtle although not significanttendency for increased locomotor activity among maternallydeprived animals could be reported.
3.3. General activity
General activity parameters were evaluated during the first dayof habituation to the NOT (pnd 37), when the arena was used as anopen-field. No significant effects of the neonatal condition werefound in any of the parameters analysed, neither significant inter-actions between factors. In contrast, a significant overall effect of sexwas found for total ambulation [two-wayANOVA,main effect of sex,[F(1,44) ¼ 4.09, p < 0.05], as well as similar trends for rearingfrequency [two-way ANOVA, F(1,44) ¼ 4.07, p ¼ 0.05] andpercentage of internal ambulation [two-wayANOVA, F(1,44)¼ 3.62,p¼ 0.06] (Fig. 3). Adolescent female rats seemed to displayed higherlevels of motor activity, and showed a trend to prefer the internalarea of the arena (internal ambulation), thus reflecting decreasedemotionality levels compared to their male counterparts.
3.4. Novel object test (NOT)
No differences were found for the total time animals spentexploring the objects during the training session, so that neithersex nor the neonatal manipulation significantly affected generalexploratory behaviour (Fig. 4, upper panel). The two-way ANOVA ofthe discrimination index data rendered a significant effect of theneonatal condition [two-way ANOVA, main effect of neonatalcondition, F(1,30) ¼ 7.84, p < 0.01)] that was accompanied bya significant interaction between sex and neonatal condition [two-way ANOVA, sex by neonatal condition interaction, F(1,30) ¼ 8.78,p < 0.01)]. Tukey post-hoc comparisons revealed a significantdifference between Co and MD females (p ¼ 0.002) in the absenceof changes amongmale animals (Fig. 4, lower panel). Therefore, MDseemed to have caused a sex-specific cognitive impairment, onlyevident among female adolescent rats.
3.5. Elevated plus-maze (EPM)
No differences were observed in the EPM in any of the param-eters analysed: percentages of open arm entries and time spent inthe open arms as well as frequency of total arms entries. However,it might be worth mentioning a trend for increased open armsexploration by MD-females, as revealed by additional statisticalanalysis (one-way ANOVA split by sex, p ¼ 0.057 and p ¼ 0.064, forpercentage of time and entries in open arms, respectively withinfemales) (Fig. 5). Therefore, neither sex nor the neonatal conditionaffected animals’ EPM performance at this age (pnd 42), althoughMD among females seemed to have slightly increased open armexploration.
3.6. Markers of synaptic plasticity
A significant effect of the neonatal manipulation was observedfor BDNF levels both in the frontal cortex [two-way ANOVA, maineffect of neonatal condition, F(1,22) ¼ 7.91, p < 0.05] and thehippocampus [two-way ANOVA, main effect of neonatal condition,F(1,23) ¼ 8.51, p < 0.005], in the absence of differences due to sex[two-way ANOVA, main effect of sex, F(1,22) ¼ 0.92, p ¼ 0.35 andF(1,23) ¼ 0.05, p ¼ 0.82, for frontal cortex and hippocampus,respectively]. The MD episode induced a significant decrease inBDNF levels in both males and females in the two brain regionsanalysed (Fig. 6, left-upper panel). Similarly, the neonatal manip-ulation modified PSD-95 levels, a marker of post-synaptic density,in the two brain regions analysed [two-way ANOVA, main effect of
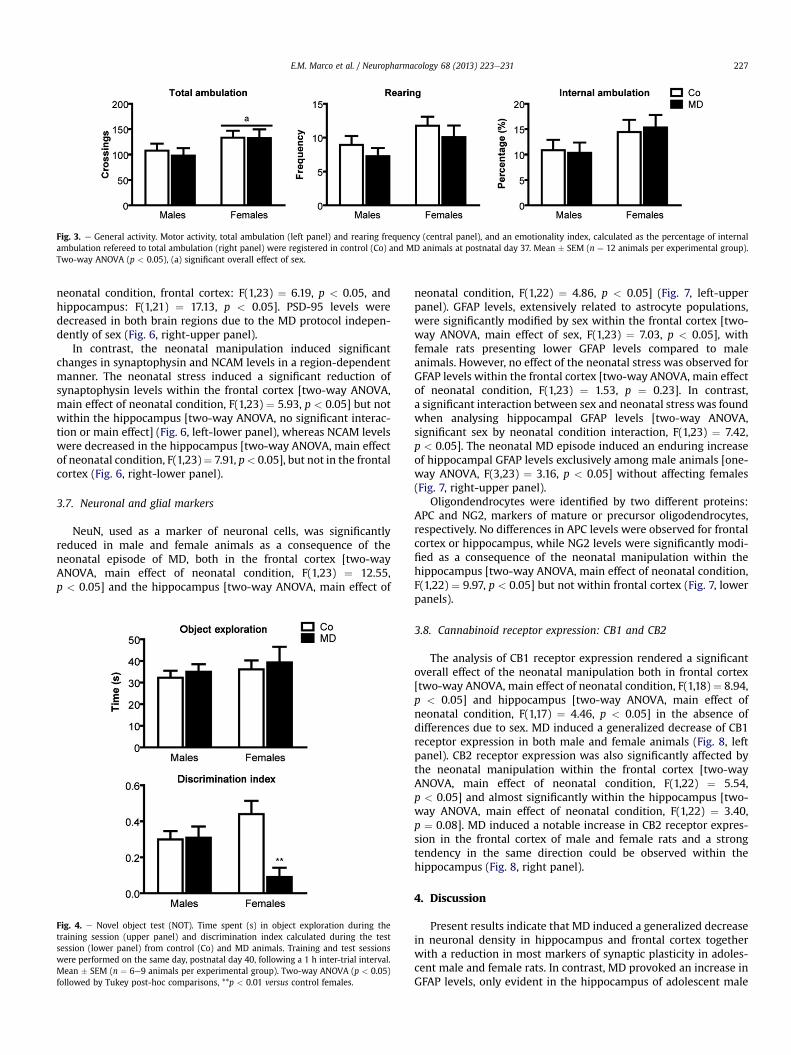
Fig. 3. e General activity. Motor activity, total ambulation (left panel) and rearing frequency (central panel), and an emotionality index, calculated as the percentage of internalambulation refereed to total ambulation (right panel) were registered in control (Co) and MD animals at postnatal day 37. Mean � SEM (n ¼ 12 animals per experimental group).Two-way ANOVA (p < 0.05), (a) significant overall effect of sex.
E.M. Marco et al. / Neuropharmacology 68 (2013) 223e231 227
neonatal condition, frontal cortex: F(1,23) ¼ 6.19, p < 0.05, andhippocampus: F(1,21) ¼ 17.13, p < 0.05]. PSD-95 levels weredecreased in both brain regions due to the MD protocol indepen-dently of sex (Fig. 6, right-upper panel).
In contrast, the neonatal manipulation induced significantchanges in synaptophysin and NCAM levels in a region-dependentmanner. The neonatal stress induced a significant reduction ofsynaptophysin levels within the frontal cortex [two-way ANOVA,main effect of neonatal condition, F(1,23) ¼ 5.93, p < 0.05] but notwithin the hippocampus [two-way ANOVA, no significant interac-tion or main effect] (Fig. 6, left-lower panel), whereas NCAM levelswere decreased in the hippocampus [two-way ANOVA, main effectof neonatal condition, F(1,23)¼ 7.91, p< 0.05], but not in the frontalcortex (Fig. 6, right-lower panel).
3.7. Neuronal and glial markers
NeuN, used as a marker of neuronal cells, was significantlyreduced in male and female animals as a consequence of theneonatal episode of MD, both in the frontal cortex [two-wayANOVA, main effect of neonatal condition, F(1,23) ¼ 12.55,p < 0.05] and the hippocampus [two-way ANOVA, main effect of
Fig. 4. e Novel object test (NOT). Time spent (s) in object exploration during thetraining session (upper panel) and discrimination index calculated during the testsession (lower panel) from control (Co) and MD animals. Training and test sessionswere performed on the same day, postnatal day 40, following a 1 h inter-trial interval.Mean � SEM (n ¼ 6e9 animals per experimental group). Two-way ANOVA (p < 0.05)followed by Tukey post-hoc comparisons, **p < 0.01 versus control females.
neonatal condition, F(1,22) ¼ 4.86, p < 0.05] (Fig. 7, left-upperpanel). GFAP levels, extensively related to astrocyte populations,were significantly modified by sex within the frontal cortex [two-way ANOVA, main effect of sex, F(1,23) ¼ 7.03, p < 0.05], withfemale rats presenting lower GFAP levels compared to maleanimals. However, no effect of the neonatal stress was observed forGFAP levels within the frontal cortex [two-way ANOVA, main effectof neonatal condition, F(1,23) ¼ 1.53, p ¼ 0.23]. In contrast,a significant interaction between sex and neonatal stress was foundwhen analysing hippocampal GFAP levels [two-way ANOVA,significant sex by neonatal condition interaction, F(1,23) ¼ 7.42,p < 0.05]. The neonatal MD episode induced an enduring increaseof hippocampal GFAP levels exclusively among male animals [one-way ANOVA, F(3,23) ¼ 3.16, p < 0.05] without affecting females(Fig. 7, right-upper panel).
Oligondendrocytes were identified by two different proteins:APC and NG2, markers of mature or precursor oligodendrocytes,respectively. No differences in APC levels were observed for frontalcortex or hippocampus, while NG2 levels were significantly modi-fied as a consequence of the neonatal manipulation within thehippocampus [two-way ANOVA, main effect of neonatal condition,F(1,22) ¼ 9.97, p < 0.05] but not within frontal cortex (Fig. 7, lowerpanels).
3.8. Cannabinoid receptor expression: CB1 and CB2
The analysis of CB1 receptor expression rendered a significantoverall effect of the neonatal manipulation both in frontal cortex[two-way ANOVA, main effect of neonatal condition, F(1,18) ¼ 8.94,p < 0.05] and hippocampus [two-way ANOVA, main effect ofneonatal condition, F(1,17) ¼ 4.46, p < 0.05] in the absence ofdifferences due to sex. MD induced a generalized decrease of CB1receptor expression in both male and female animals (Fig. 8, leftpanel). CB2 receptor expression was also significantly affected bythe neonatal manipulation within the frontal cortex [two-wayANOVA, main effect of neonatal condition, F(1,22) ¼ 5.54,p < 0.05] and almost significantly within the hippocampus [two-way ANOVA, main effect of neonatal condition, F(1,22) ¼ 3.40,p ¼ 0.08]. MD induced a notable increase in CB2 receptor expres-sion in the frontal cortex of male and female rats and a strongtendency in the same direction could be observed within thehippocampus (Fig. 8, right panel).
4. Discussion
Present results indicate that MD induced a generalized decreasein neuronal density in hippocampus and frontal cortex togetherwith a reduction in most markers of synaptic plasticity in adoles-cent male and female rats. In contrast, MD provoked an increase inGFAP levels, only evident in the hippocampus of adolescent male

Fig. 5. e Elevated plus maze (EPM). Time spent in open arms (left panel), open arm entries (central panel) and total entries (right panel) of control (Co) and MD animals exposed tothe EPM at postnatal day 43. Mean � SEM (n ¼ 7e11 animals per experimental group).
E.M. Marco et al. / Neuropharmacology 68 (2013) 223e231228
rats as well as important and opposite changes in the expression ofcannabinoid receptors in the two brain regions analysed, i.e. CB1receptors were decreased whereas the contrary was observed forCB2 receptors, an up-regulation. Surprisingly, at the behaviourallevel, MD consequences were exclusively observed among adoles-cent females, that exhibited a marked deficit in recognitionmemory.
MD induced a remarkable deficit in recognition memory (NOT)among females (not in males), in the absence of changes inworkingmemory (SAT). Previously, cognitive impairments in recognitionmemory have been described among male and female adultanimals due to MD (Llorente-Berzal et al., 2012; Llorente et al.,2011). Therefore, the detrimental consequences of MD in recogni-tion memory might be sex-specific, at least during the adolescenceperiod, since only adolescent females appeared to be affected in thepresent study. Most sex differences in learning processes aredependent on organizational effects of gonadal hormones duringearly development of the brain, but also due to its modulation byactivational factors during puberty and/or adolescence (Dalla andShors, 2009). Actually, detection of novelty, essential for recogni-tion memory, seems to develop throughout brain maturation(Reger et al., 2009). Consequently, the increased vulnerability ofadolescent females to the effects of MD on recognition memorymay rely on the sexually different brain maturation pathways.However, further research is needed to better understand theconsequences of the developmental disruption - by early life stress,i.e. MD - of these sex-dependent maturational processes.
Fig. 6. e Markers of synaptic plasticity in the frontal cortex and hippocampus of control (Coa marker of post-synaptic density, synaptophysin and neural cell adhesion molecule (NCAMANOVA (p < 0.05), (b) significant overall effect of neonatal condition.
Amodest although not significant trend for increased locomotoractivity was observed among MD animals, particularly females (inthe SAT), which also exhibited a tendency for increased open armsexploration (in the EPM). Classically, increased open-arm explora-tion in the EPM has been related to anxiolytic-like responses,despite alternative interpretations such as increased risk-takingbehaviour and/or impulsivity have been proposed (Davis et al.,2009; Llorente et al., 2011). Additional experiments would beneeded to appropriately address this question that is furtherobscured by the ontogenetic sex-differences previously describedfor rats’ performance in the EPM (Imhof et al., 1993). As in previousstudies (Ellenbroek et al., 2005; Gruss et al., 2008; Llorente et al.,2007; Rentesi et al., 2010; Viveros et al., 2009), a long-lastingreduction in body weight due to MD was observed in both maleand female animals, with sex-differences becoming significantfrom mid adolescence. Nevertheless, from the behavioural point ofview, adolescent female rats appeared to be more vulnerable to thedetrimental consequences of MD, particularly as regards cognitiveimpairments.
Hippocampus and frontal cortex were investigated as potentialneurobiological substrates of the reported sex-dependent deficitsin cognitive function. Previous studies had already showed a sex-dependent effect of MD on hippocampal GFAP levels; MD exclu-sively augmented GFAP levels within the hippocampus of maleanimals (Lopez-Gallardo et al., 2012; Llorente et al., 2008). In thepresent study, we further confirmed this sex and region depen-dent effect among adolescent animals, despite a similar although
) and MD animals, postnatal day 40. Brain derived neurotrophic factor (BDNF), PSD95,) were analysed. Mean � SEM (n ¼ 4e6 animals per experimental group). Two-way

Fig. 7. e Neuronal and glial markers in the frontal cortex and hippocampus of control (Co) and MD animals, postnatal day 40. NeuN and GFAP were employed as neuronal and glialmarkers, respectively (upper panel), while APC and NG2 were used as markers of mature oligodendrocytes and oligodendrocyte precursor cells, respectively (bottom panel).Mean � SEM (n ¼ 4e6 animals per experimental group). Two-way ANOVA (p < 0.05), (a) significant overall effect of sex, and (b) significant overall effect of neonatal condition; one-way ANOVA, &&p < 0.01 vs. control males.
E.M. Marco et al. / Neuropharmacology 68 (2013) 223e231 229
non-statistically significant trend was observed for the frontalcortex. In this regard, males’ astrocytes seem to be more vulner-able to the effects of MD. Actually, female cortical astrocytes seemto be more resistant to in vitro oxidative insults than their malecounterparts, probably due to the enhanced aromatization andestradiol formation provided by P450 aromatase in female cells(Liu et al., 2007). Accordingly, a higher aromatization capacity infemales’ astrocytes may represent one of the mechanisms under-lying the sex-dependent differences observed in vivo for theeffects of early life stress, i.e. MD, on the astrocyte population.Differences in oligodendrocytes precursors, indicated by NG2levels, were also found in the hippocampus of adolescent maleand female MD animals, without changes in frontal cortex or inthe levels of mature oligodendrocytes. In line with this finding, anincrease in the rate of cell death, mostly of oligodendrocytes inwhite matter tracts, was observed following a similar protocol ofearly life stress, MD at pnd 11 (Zhang et al., 2002). Thus, oligo-dendrocytes seem to be a sensitive target to the MD effects,although additional research is still needed in order to investigatethe consequences of early life stress in the temporal and regionalmaturation of oligodendrocytes as well as in mielinizationprocesses.
MD also induced a generalized decrease in the number of NeuN-positive cells in both brain regions. NeuN immunoreactivity isbroadly used as a specific marker for neurons (Mullen et al., 1992),
Fig. 8. e Expression of cannabinoid receptors, CB1 and CB2 receptors, in the frontal cortex an6 animals per experimental group). Two-way ANOVA (p < 0.05): (b) significant overall effe
and thus, such a decrease might be considered as an evidence ofneuronal loss. Additional experiments are guaranteed to determinewhether this neuronal loss results from increased apoptosis and/orfrom a decrease in neurogenesis during neurodevelopment. Actu-ally, an increase in neuronal degeneration in the hippocampus andcerebellum of neonate rat pups has been described (at pnd 13)following MD, indicated by an increase in the number of Fluoro-Jade C positive cells (Llorente et al., 2009). In contrast, a decreasein hippocampal neurogenesis has also been reported followingseveral protocols early life stress (Lajud et al., 2012; Mirescu et al.,2004; Oreland et al., 2010).
Present data provide evidence for alterations in synaptic plas-ticity as a consequence of early life stress at adolescence (pnd 40),as previously described in adult animals (Llorente et al., 2011;Roceri et al., 2002). A decrease in BDNF and PSD95 expression, aswell as region-specific decrements in synaptophysin and NCAMlevels, was observed in male and female adolescent animalsexposed to MD. These changes in neurotrophin factors and insynaptic density, indicated by PSD-95 expression, may result fromthe disruption of developing neurotransmitters circuitries andhormonal systems involved in the control of neurotrophinproduction as previously suggested by Roceri et al. (2002).
The impact of early life stress on the endocannabinoid systemextends previous existing literature (Llorente et al., 2008; Suarezet al., 2009, 2010; Llorente-Berzal et al., 2012). MD diminished
d hippocampus of control (Co) and MD animals, postnatal day 40. Mean � SEM (n ¼ 4ects of neonatal condition.

E.M. Marco et al. / Neuropharmacology 68 (2013) 223e231230
hippocampal and cortical CB1 receptor expression, whereas theopposite was observed for CB2 receptor expression - an increase -in the frontal cortex of adolescent animals. This profile was alreadyobserved short after the MD protocol (at pnd 13) in the hippo-campus of male rat pups, although no changes in CB1 receptorexpression were observed in females’ hippocampus (Suarez et al.,2009). Similarly, MD induced a long-lasting reduction in CB1receptor expression in the hippocampus of adult male and femaleanimals (Llorente-Berzal et al., 2012). Recently, 2-arachidonylglycerol (2-AG) has been proposed to play a critical role in thewiring of emergent neuronal networks in the foetal brain anddevelopmental 2-AG concentrations might substantially influenceCB1 receptor expression as these receptors are being trafficked inaxons (Keimpema et al., 2011). Given the fact that MD has beenreported to increase 2-AG content and metabolism in the hippo-campus of neonatal rat pups (Llorente et al., 2008; Suarez et al.,2010), the developmental fluctuations of 2-AG levels during earlyneonatal stages may have determined the long-lasting changesobserved in CB1 receptor expression. In contrast, the strikingopposite pattern observed for CB2 receptor expression may indi-cate, as previously suggested (Suarez et al., 2009), some kind offunctional compensation and/or interaction between these twocannabinoid receptors.
Sexual dimorphisms are present in response to early lifestress. In our hands, sex differences were observed in thebehavioural studies, i.e. cognitive deficit, but not in the neuro-biological parameters analysed. Changes in markers of synapticplasticity, neuronal and glial populations and cannabinoidreceptor expression within hippocampus and frontal cortex werecomparable between sexes, but the impairment in recognitionmemory was only observed among adolescent female rats. Itmight be worth mentioning that, although not significant, thedecrease in NeuN expression seemed to be more drastic inadolescent female animals, and such a sex difference may influ-ence, at least in part, animals’ cognitive function. More recently,sex differences in the effects of MD in the hippocampal expres-sion of calcium-binding proteins (calretinin and calbindin) havebeen reported. MD provoked an increase in both calretinin andcalbindin proteins in the hippocampus of males, while MDfemales experienced an increase in calretinin levels but a notabledecrease in calbindin expression (Xu et al., 2011). Since a declinein calbindin levels may represent alterations in the neuro-protective properties against apoptosis (Nowak et al., 2010), thissexually dimorphic effect may account for the presently observedsex differences in NeuN expression, and consequently in thedifferential effects of MD on males and females’ cognitive func-tion. However, further research is needed to understand theneurobiological mechanisms underlying the sexual dimorphismsassociated to the MD effects.
5. Concluding remarks
Early life stress interferes with brain developmental trajecto-ries modifying both behavioural and neurobiological parameters.However, future research on the sex-dependent ontogeneticprofile of early-life stress consequences is urgently needed.Further investigation of the specific sex-dependent vulnerabilitiesto early life stress is critical for the understanding the aetiologyof psychopathological conditions with a neurodevelopmentalorigin.
Acknowledgements
Instituto de Salud Carlos III, Redes temáticas de InvestigaciónCooperativa en salud RD06/0001/1013; GRUPOS UCM-BSCH
(GRUPO UCM 951579); Ministerio de Ciencia e Innovación:BFU2009-10109; FIS (PI10/01748).
References
Archer, J., 1973. Tests for emotionality in rats and mice: a review. Anim. Behav. 21,205e235.
Barker, G.R., Bird, F., Alexander, V., Warburton, E.C., 2007. Recognition memory forobjects, place, and temporal order: a disconnection analysis of the role of themedial prefrontal cortex and perirhinal cortex. J. Neurosci. 27, 2948e2957.
Bremne, J.D., Vermetten, E., 2001. Stress and development: behavioral and biolog-ical consequences. Dev. Psychopathol. 13, 473e489.
Buffalo, E.A., Bellgowan, P.S., Martin, A., 2006. Distinct roles for medial temporal lobestructures in memory for objects and their locations. Learn. Mem. 13, 638e643.
Cirulli, F., Francia, N., Berry, A., Aloe, L., Alleva, E., Suomi, S.J., 2009. Early life stress asa risk factor for mental health: role of neurotrophins from rodents to non-human primates. Neurosci. Biobehav Rev. 33, 573e585.
Dalla, C., Shors, T.J., 2009. Sex differences in learning processes of classical andoperant conditioning. Physiol. Behav. 97, 229e238.
Davis, J.F., Krause, E.G., Melhorn, S.J., Sakai, R.R., Benoit, S.C., 2009. Dominant rats arenatural risk takers and display increased motivation for food reward. Neuro-science 162, 23e30.
Dember, W.N., Fowler, H., 1958. Spontaneous alternation behavior. Psychol. Bull. 55,412e428.
Dix, S.L., Aggleton, J.P., 1999. Extending the spontaneous preference test of recog-nition: evidence of object-location and object-context recognition. Behav. BrainRes. 99, 191e200.
Ellenbroek, B.A., Derks, N., Park, H.J., 2005. Early maternal deprivation retardsneurodevelopment in Wistar rats. Stress 8, 247e257.
Ellenbroek, B.A., Riva, M.A., 2003. Early maternal deprivation as an animal model forschizophrenia. Clin. Neurosci. Res. 3, 297e302.
Ennaceur, A., Delacour, J., 1988. A new one-trial test for neurobiological studies ofmemory in rats. 1: Behavioral data. Behav. Brain Res. 31, 47e59.
Fatemi, S.H., Folsom, T.D., 2009. The neurodevelopmental hypothesis of schizo-phrenia, revisited. Schizophr Bull. 35, 528e548.
Fumagalli, F., Molteni, R., Racagni, G., Riva, M.A., 2007. Stress during development:impact on neuroplasticity and relevance to psychopathology. Prog. Neurobiol.81, 197e217.
Gruss, M., Braun, K., Frey, J.U., Korz, V., 2008. Maternal separation during a specificpostnatal time window prevents reinforcement of hippocampal long-termpotentiation in adolescent rats. Neuroscience 152, 1e7.
Heim, C., Nemeroff, C.B., 2001. The role of childhood trauma in the neurobiology ofmood and anxiety disorders: preclinical and clinical studies. Biol. Psychiatry 49,1023e1039.
Hogg, S., 1996. A review of the validity and variability of the elevated plus-maze asan animal model of anxiety. Pharmacol. Biochem. Behav. 54, 21e30.
Hughes, R.N., 2004. The value of spontaneous alternation behavior (SAB) as a test ofretention in pharmacological investigations of memory. Neurosci. Biobehav Rev.28, 497e505.
Imhof, J.T., Coelho, Z.M., Schmitt, M.L., Morato, G.S., Carobrez, A.P., 1993. Influence ofgender and age on performance of rats in the elevated plus maze apparatus.Behav. Brain Res. 56, 177e180.
Keimpema, E., Mackie, K., Harkany, T., 2011. Molecular model of cannabis sensitivityin developing neuronal circuits. Trends Pharmacol. Sci. 32, 551e561.
Lajud, N., Roque, A., Cajero, M., Gutierrez-Ospina, G., Torner, L., 2012. Periodicmaternal separation decreases hippocampal neurogenesis without affectingbasal corticosterone during the stress hyporesponsive period, but alters HPA axisand coping behavior in adulthood. Psychoneuroendocrinology 37, 410e420.
Lehmann, J., Russig, H., Feldon, J., Pryce, C.R., 2002. Effect of a single maternalseparation at different pup ages on the corticosterone stress response in adultand aged rats. Pharmacol. Biochem. Behav. 73, 141e145.
Levine, S., Huchton, D.M., Wiener, S.G., Rosenfeld, P., 1991. Time course of the effectof maternal deprivation on the hypothalamic-pituitary-adrenal axis in theinfant rat. Dev. Psychobiol. 24, 547e558.
Liu, M., Hurn, P.D., Roselli, C.E., Alkayed, N.J., 2007. Role of P450 aromatase in sex-specific astrocytic cell death. J. Cereb. Blood Flow Metab. 27, 135e141.
Lopez-Gallardo, M., Lopez-Rodriguez, A.B., Llorente-Berzal, A., Rotllant, D.,Mackie, K., Armario, A., Nadal, R., Viveros, M.P., 2012. Maternal deprivation andadolescent cannabinoid exposure impact hippocampal astrocytes, CB1 recep-tors and brain-derived neurotrophic factor in a sexually dimorphic fashion.Neuroscience 204, 90e103.
Llorente-Berzal, A., Mela, V., Borcel, E., Valero, M., Lopez-Gallardo, M., Viveros, M.P.,Marco, E.M., 2012. Neurobehavioral and metabolic long-term consequences ofneonatal maternal deprivation stress and adolescent olanzapine treatment inmale and female rats. Neuropharmacology 62, 1332e1341.
Llorente, R., Arranz, L., Marco, E.M., Moreno, E., Puerto, M., Guaza, C., De laFuente, M., Viveros, M.P., 2007. Early maternal deprivation and neonatal singleadministration with a cannabinoid agonist induce long-term sex-dependentpsychoimmunoendocrine effects in adolescent rats. Psychoneuroendocrinology32, 636e650.
Llorente, R., Gallardo, M.L., Berzal, A.L., Prada, C., Garcia-Segura, L.M., Viveros, M.P.,2009. Early maternal deprivation in rats induces gender-dependent effects ondeveloping hippocampal and cerebellar cells. Int. J. Dev. Neurosci. 27, 233e241.

E.M. Marco et al. / Neuropharmacology 68 (2013) 223e231 231
Llorente, R., Llorente-Berzal, A., Petrosino, S., Marco, E.M., Guaza, C., Prada, C.,Lopez-Gallardo, M., Di Marzo, V., Viveros, M.P., 2008. Gender-dependentcellular and biochemical effects of maternal deprivation on the hippocampus ofneonatal rats: a possible role for the endocannabinoid system. Dev. Neurobiol.68, 1334e1347.
Llorente, R., Miguel-Blanco, C., Aisa, B., Lachize, S., Borcel, E., Meijer, O.C.,Ramirez, M.J., De Kloet, E.R., Viveros, M.P., 2011. Long term sex-dependentpsychoneuroendocrine effects of maternal deprivation and juvenile unpre-dictable stress in rats. J. Neuroendocrinol 23, 329e344.
Llorente, R., O’Shea, E., Gutierrez-Lopez, M.D., Llorente-Berzal, A., Colado, M.I.,Viveros, M.P., 2010. Sex-dependent maternal deprivation effects on brainmonoamine content in adolescent rats. Neurosci. Lett. 479, 112e117.
Llorente, R., Villa, P., Marco, E.M., Viveros, M.P., 2012. Analyzing the effects ofa single episode of neonatal maternal deprivation on metabolite profiles in ratbrain: a proton nuclear magnetic resonance spectroscopy study. Neuroscience201, 12e19.
Marco, E.M., Adriani, W., Canese, R., Podo, F., Viveros, M.P., Laviola, G., 2007.Enhancement of endocannabinoid signalling during adolescence: modulationof impulsivity and long-term consequences on metabolic brain parameters inearly maternally deprived rats. Pharmacol. Biochem. Behav. 86, 334e345.
Marco, E.M., Adriani, W., Llorente, R., Laviola, G., Viveros, M.P., 2009. Detrimentalpsychophysiological effects of early maternal deprivation in adolescent andadult rodents: altered responses to cannabinoid exposure. Neurosci. Biobehav.Rev. 33, 498e507.
Mateos, B., Borcel, E., Loriga, R., Luesu, W., Bini, V., Llorente, R., Castelli, M.,Viveros, M.P., 2011. Adolescent exposure to nicotine and/or the cannabinoidagonist CP 55,940 induces gender-dependent long-lasting memory impair-ments and changes in brain nicotinic and CB1 cannabinoid receptors.J. Psychopharmacol. 25, 1676e1690.
Mirescu, C., Peters, J.D., Gould, E., 2004. Early life experience alters response of adultneurogenesis to stress. Nat. Neurosci. 7, 841e846.
Mitchell, J.B., Laiacona, J., 1998. The medial frontal cortex and temporal memory: testsusing spontaneous exploratory behaviour in the rat. Behav. Brain Res. 97,107e113.
Morgan, C., Fisher, H., 2007. Environment and schizophrenia: environmentalfactors in schizophrenia: childhood traumaea critical review. Schizophr. Bull.33, 3e10.
Mullen, R.J., Buck, C.R., Smith, A.M., 1992. NeuN, a neuronal specific nuclear proteinin vertebrates. Development 116, 201e211.
Mumby, D.G., Gaskin, S., Glenn, M.J., Schramek, T.E., Lehmann, H., 2002. Hippo-campal damage and exploratory preferences in rats: memory for objects, pla-ces, and contexts. Learn. Mem. 9, 49e57.
Nowak, B., Zadrozna, M., Ossowska, G., Sowa-Kucma, M., Gruca, P., Papp, M.,Dybala, M., Pilc, A., Nowak, G., 2010. Alterations in hippocampal calcium-binding neurons induced by stress models of depression: a preliminaryassessment. Pharmacol. Rep. 62, 1204e1210.
Oreland, S., Nylander, I., Pickering, C., 2010. Prolonged maternal separationdecreases granule cell number in the dentate gyrus of 3-week-old male rats. Int.J. Dev. Neurosci. 28, 139e144.
Paus, T., Keshavan, M., Giedd, J.N., 2008. Why do many psychiatric disorders emergeduring adolescence? Nat. Rev. Neurosci. 9, 947e957.
Pellow, S., Chopin, P., File, S.E., Briley, M., 1985. Validation of open:closed armentries in an elevated plus-maze as a measure of anxiety in the rat. J. Neurosci.Methods 14, 149e167.
Plotsky, P.M., Meaney, M.J., 1993. Early, postnatal experience alters hypotha-lamic corticotropin-releasing factor (CRF) mRNA, median eminence CRF
content and stress-induced release in adult rats. Brain Res. Mol. Brain Res.18, 195e200.
Reger, M.L., Hovda, D.A., Giza, C.C., 2009. Ontogeny of Rat Recognition Memorymeasured by the novel object recognition task. Dev. Psychobiol. 51, 672e678.
Rentesi, G., Antoniou, K., Marselos, M., Fotopoulos, A., Alboycharali, J., Konstandi, M.,2010. Long-term consequences of early maternal deprivation in serotonergicactivity and HPA function in adult rat. Neurosci. Lett. 480, 7e11.
Roceri, M., Hendriks, W., Racagni, G., Ellenbroek, B.A., Riva, M.A., 2002. Earlymaternal deprivation reduces the expression of BDNF and NMDA receptorsubunits in rat hippocampus. Mol. Psychiatry 7, 609e616.
Rossato, J.I., Bevilaqua, L.R., Myskiw, J.C., Medina, J.H., Izquierdo, I., Cammarota, M.,2007. On the role of hippocampal protein synthesis in the consolidation andreconsolidation of object recognition memory. Learn. Mem. 14, 36e46.
Sandi, C., Woodson, J.C., Haynes, V.F., Park, C.R., Touyarot, K., Lopez-Fernandez, M.A.,Venero, C., Diamond, D.M., 2005. Acute stress-induced impairment ofspatial memory is associated with decreased expression of neural celladhesion molecule in the hippocampus and prefrontal cortex. Biol. Psychiatry57, 856e864.
Squire, L.R., Wixted, J.T., Clark, R.E., 2007. Recognition memory and the medialtemporal lobe: a new perspective. Nat. Rev. Neurosci. 8, 872e883.
Steinberg, L., 2005. Cognitive and affective development in adolescence. TrendsCogn. Sci. 9, 69e74.
Suarez, J., Llorente, R., Romero-Zerbo, S.Y., Mateos, B., Bermudez-Silva, F.J., deFonseca, F.R., Viveros, M.P., 2009. Early maternal deprivation induces gender-dependent changes on the expression of hippocampal CB(1) and CB(2)cannabinoid receptors of neonatal rats. Hippocampus 19, 623e632.
Suarez, J., Rivera, P., Llorente, R., Romero-Zerbo, S.Y., Bermudez-Silva, F.J., deFonseca, F.R., Viveros, M.P., 2010. Early maternal deprivation induces changes onthe expression of 2-AG biosynthesis and degradation enzymes in neonatal rathippocampus. Brain Res. 1349, 162e173.
Talge, N.M., Neal, C., Glover, V., 2007. Antenatal maternal stress and long-termeffects on child neurodevelopment: how and why? J. Child. Psychol. Psychi-atry 48, 245e261.
Teicher, M.H., Andersen, S.L., Polcari, A., Anderson, C.M., Navalta, C.P., Kim, D.M.,2003. The neurobiological consequences of early stress and childhoodmaltreatment. Neurosci. Biobehav Rev. 27, 33e44.
Venero, C., Herrero, A.I., Touyarot, K., Cambon, K., Lopez-Fernandez, M.A.,Berezin, V., Bock, E., Sandi, C., 2006. Hippocampal up-regulation of NCAMexpression and polysialylation plays a key role on spatial memory. Eur. J.Neurosci. 23, 1585e1595.
Viveros, M.P., Llorente, R., Diaz, F., Romero-Zerbo, S.Y., Bermudez-Silva, F.J., Rodri-guez de Fonseca, F., Argente, J., Chowen, J.A., 2010. Maternal deprivation hassexually dimorphic long-term effects on hypothalamic cell-turnover, bodyweight and circulating hormone levels. Horm. Behav. 58, 808e819.
Viveros, M.P., Llorente, R., Lopez-Gallardo, M., Suarez, J., Bermudez-Silva, F., De laFuente, M., Rodriguez de Fonseca, F., Garcia-Segura, L.M., 2009. Sex-dependentalterations in response to maternal deprivation in rats. Psychoneur-oendocrinology 34 (1), S217eS226.
Xu, H., Hu, W., Zhang, X., Gao, W., Liang, M., Chen, T., He, X., Hu, Z., 2011. The effect ofdifferent maternal deprivation paradigms on the expression of hippocampalglucocorticoid receptors, calretinin and calbindin-D28k in male and femaleadolescent rats. Neurochem. Int. 59, 847e852.
Zhang, L.X., Levine, S., Dent, G., Zhan, Y., Xing, G., Okimoto, D., Kathleen Gordon, M.,Post, R.M., Smith, M.A., 2002. Maternal deprivation increases cell death in theinfant rat brain. Brain Res. Dev. Brain Res. 133, 1e11.