major microbiology department of biology, ipb...fisiologi molekuler (nisa rm) 4 « all enzymes are...
TRANSCRIPT

Fisiologi Molekuler (Nisa RM) 1
Nisa Rachmania MubarikMajor Microbiology Department of Biology, IPB

Fisiologi Molekuler (Nisa RM) 2
ENZYMES ARE: Proteins (note that recent developments indicate that both RNA and antibodies may have catalytic activity, these are called ribozymes, and catalytic antibodies or abzymes, respectively)
Biological catalysts, critical components of cell metabolism & biological processes
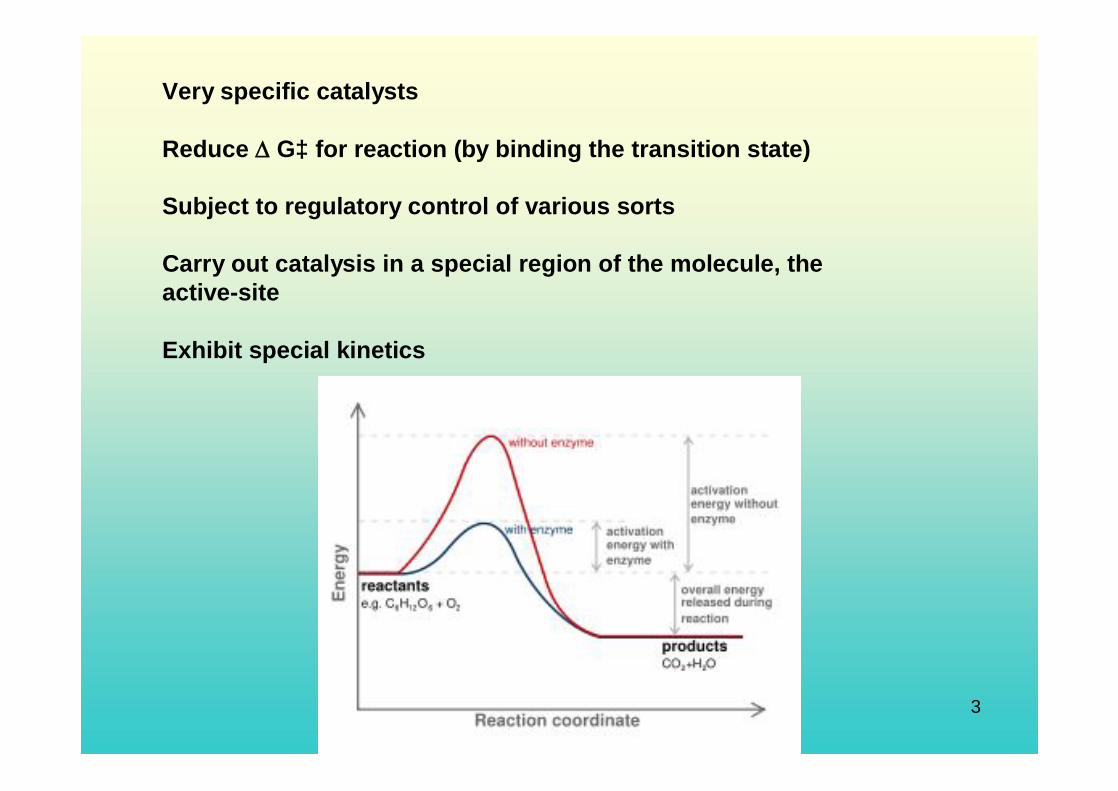
Very efficient catalysts Like other catalysts, enzymes do not alter the position of equilibrium between substrates and products. However, unlike normal chemical reactions, enzymes are saturable. This means as more substrate is added, the reaction rate will increase, because more active sites become occupied.
Fisiologi Molekuler (Nisa RM) 3
Very specific catalysts
Reduce G‡ for reaction (by binding the transition state)
Subject to regulatory control of various sorts
Carry out catalysis in a special region of the molecule, the active-site
Exhibit special kinetics

Fisiologi Molekuler (Nisa RM) 4
« All enzymes are proteins, with the exception of some small catalytic RNAs and RNA/protein complexes.
« MW's range from 104 to 106 daltons.
« May be single polypeptide chains, or oligomers of several subunits (most commonly oligomers are dimers, or tetramers, some multienzyme complexes as many as 48 protomers).
« May have more than one activity associated with the same protein (i. e. there are some large enzymes which catalyze more than onereaction (frequently successive steps in a metabolic pathway).
« Often contain a prosthetic group (or cofactor): Typical examplesare: metal ions, heme, Fe-S clusters, coenzymes (e.g. NADH, FAD, FMN, PLP) .
« Coenzymes usually are vitamins, or derived from vitamins, and act as carriers (e.g. of H, e-, CO2).
« Enzymes are usually named after their substrate by adding ase, e. g. protease (proteinase), esterase, -glucosidase, alcohol dehydrogenase, -lactamase.

Fisiologi Molekuler (Nisa RM) 5
The four levels of protein structure are illustrated in this diagram. http://www.genome.gov/page.cfm?pageID=10000552
For example, the enzyme ß-galactosidase is a tetramer, meaning that it is composed of four subunits, and, in this case, the subunits are identical - each polypeptide chain has the same sequence of amino acids. For some proteins, quaternary structure is required for full activity (function) of the protein.

Fisiologi Molekuler (Nisa RM) 6
Tertiary structure refers to the three dimensional globular structure formed by bending and twisting of the polypeptide chain. This process often means that the linear sequence of amino acids is folded into a compact globular structure. The folding of the polypeptide chain is stabilized by multiple weak, noncovalentinteractions. These interactions include:
Hydrogen bonds that form when a Hydrogen atom is shared by two other atoms.
Electrostatic interactions that occur between charged amino acid side chains. Electrostatic interactions are attractions between positive and negative sites on macromolecules.
Hydrophobic interactions: During folding of the polypeptide chain, amino acids with a polar (water soluble) side chain are often found on the surface of the molecule while amino acids with non polar (water insoluble) side chain are buried in the interior. This means that the folded protein is soluble in water or aqueous solutions.
Covalent bonds may also contribute to tertiary structure. The amino acid, cysteine, has an SH group as part of its R group and therefore, the disulfide bond (S-S ) can form with an adjacent cysteine.

Fisiologi Molekuler (Nisa RM) 7
Enzyme Assays
All enzyme assays measure either the consumption of substrate orproduction of product over time.
Methode:Spectrophotometric assays (colorimetric assays): the course of the reaction by measuring a change in how much light the assay solution absorbs. If this light is in the visible region you can actually see a change in the color of the assay, these are called
Diagram of a single-beam UV/vis spectrophotometer

Fisiologi Molekuler (Nisa RM) 8
Fluorimetric assays
Fluorescence is when a molecule emits light of one wavelength after absorbing light of a different wavelength.
Fluorometric assays use a difference in the fluorescence of substrate from product to measure the enzyme reaction. These assays are ingeneral much more sensitive than spectrophotometric assays, but can suffer from interference caused by impurities and the instability of many fluorescent compounds when exposed to light.
An example of these assays is again the use of the nucleotide coenzymes NADH and NADPH. The reduced forms are fluorescent and the oxidised forms non-fluorescent. Oxidation reactions can therefore be followed by a decrease in fluorescence and reduction reactions by an increase.
Synthetic substrates that release a fluorescent dye in an enzyme-catalyzed reaction are also available, such as 4-methylumbelliferyl-β-D-glucuronide for assaying β-galactosidase.

Fisiologi Molekuler (Nisa RM) 9
Factors to control in assays
Salt Concentration: Most enzymes can not tolerate extremely high salt concentrations. The ions interfere with the weak ionic bonds of proteins. Typical enzymes are active in salt concentrations of 1-500 mM.
Effects of Temperature: All enzymes work within a range of temperature specific to the organism. Increases in temperature generally lead to increases in reaction rates. There is a limit to the increase because higher temperatures lead to a sharp decrease in reaction rates. This is due to the denaturating (alteration) of protein structure resulting from the breakdown of the weak ionic and hydrogen bonding that stabilize the three dimensional structure of the enzyme. However, the idea of an "optimum" rate of an enzyme reaction is misleading, as the rate observed at any temperature is the product of two rates, the reaction rate and the denaturation rate.

Fisiologi Molekuler (Nisa RM) 10
Effects of pH: Most enzymes are sensitive to pH and have specific ranges of activity. All have an optimum pH. The pH can stop enzyme activity by denaturating (altering) the three dimensional shape of the enzyme by breaking ionic, and hydrogen bonds.
Substrate Saturation: Increasing the substrate concentration increases the rate of reaction (enzyme activity). However, enzyme saturation limits reaction rates. An enzyme is saturated when the active sites of all the molecules are occupied most of the time. At the saturation point, the reaction will not speed up, no matter how much additional substrate is added. The graph of the reaction rate will plateau.

Fisiologi Molekuler (Nisa RM) 11
Protein Properties and SeparationProteins are typically characterized by their size (molecular weight) and shape, amino acid composition and sequence, isolelectric point (pI), hydrophobicity, and biological affinity. Differences in these properties can be used as the basis for separation methods in a purification strategy.
The chemical composition of the unique R groups is responsible for the important characteristics of amino acids, chemical reactivity, ionic charge and relative hydrophobicity. Therefore protein properties relate back to number and type ofamino acids that make up the protein. Size: Size of proteins is usually measured in molecular weight (mass) although occasionally the length or diameter of a protein is given in Angstroms. The molecular weight of a protein is the mass of one mole of protein, usually measured in units called daltons. One dalton is the atomic mass of one proton or neutron. The molecular weight can be estimated by a number of different methods including electrophoresis, gel filtration, and more recently by mass spectrometry. The molecular weight of proteins varies over a wide range. The average molecular weight of a protein is between 40,000 to 50,000 daltons. Separation methods that are based on size and shape include gel filtration chromatography (size exclusion chromatography) and polyacrylamide gel electrophoresis.

Fisiologi Molekuler (Nisa RM) 12
Charge: Each protein has an amino group at one end and a carboxyl group at the other end as well as numerous amino acid side chains, some of which are charged. Therefore each protein carries a net charge. The net protein charge is strongly influenced by the pH of the solution.
Different proteins have different numbers of each of the amino acid side chains and therefore have different isoelectric points. So, in a buffer solution at a particular pH, some proteins will be positively charged, some proteins will be negatively charged and some will have no charge.
Separation techniques that are based on charge include ion exchange chromatography, isoelectric focusing and chromatofocusing.

Fisiologi Molekuler (Nisa RM) 13
The pI is the pH at which there is no net charge on the protein. At lower pH readings, there are more positive charges in the environment and therefore, the protein has an increased cationic character. The reverse is true at pH readings above the pI.
At pH 6.8, this protein has an equal number of positive and negative charges and so there is no net charge on the protein. As the pH drops, more H+ ions are available in the solution. These hydrogen ions bind to negative sites on the amino acids. Therefore, as the pH drops, the protein as a whole becomes positively charged. Conversely, at a basic pH, the protein becomes negatively charged. pH 6.8 is called the pI, or isoelectric point, for this protein; that is, the pH at which there are an equal number of positive and negative charges.
http://matcmadison.edu/biotech/resources/proteins/labManual/chapter_2.htm

Fisiologi Molekuler (Nisa RM) 14
Hydrophobicity: Literally, hydrophobic means fear of water. In aqueous solutions, proteins tend to fold so that areas of the protein with hydrophobic regions are located in internal surfaces next to each other and away from the polar water molecules of the solution. Polar groups on the amino acid are called hydrophilic (water loving) because they will form hydrogen bonds with water molecules. The number, type and distribution of nonpolar amino acid residues within the protein determines its hydrophobic character.
A separation method that is based on the hydrophobic character of proteins is hydrophobic interaction chromatography.

Fisiologi Molekuler (Nisa RM) 15
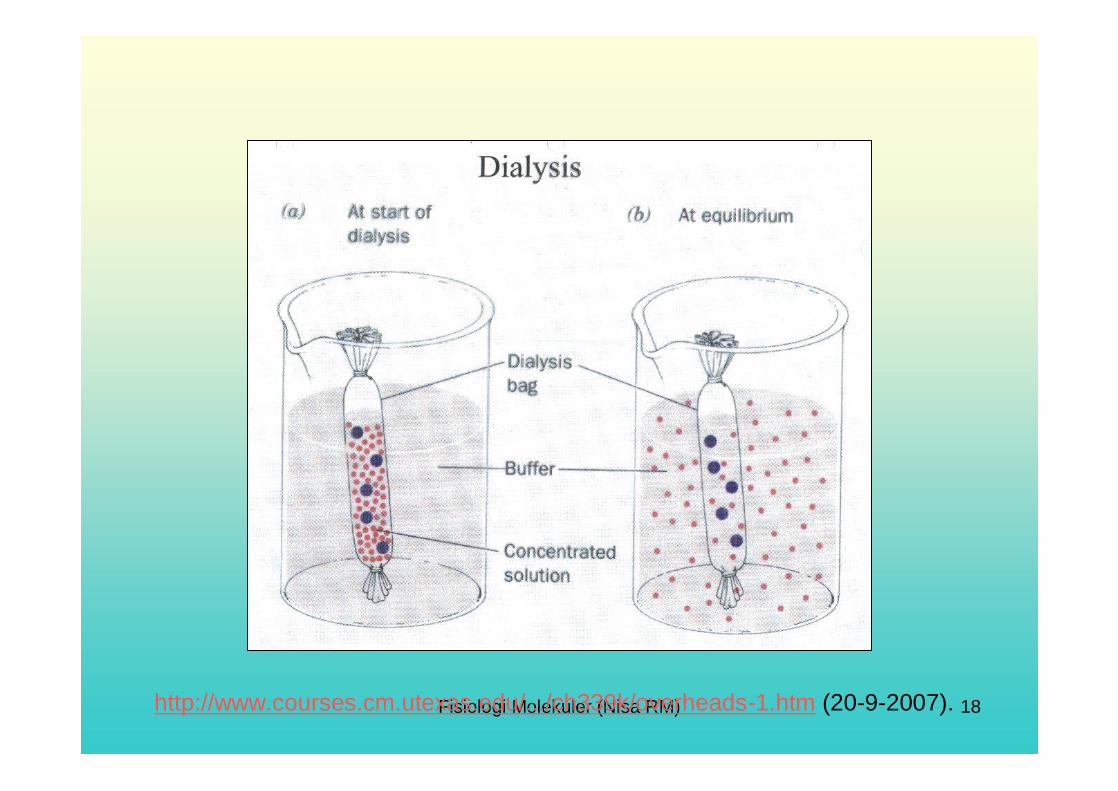
Solubility: As the name implies, solubility is the amount of a solute that can be dissolved in a solvent. The 3-D structure of a protein affects its solubility properties. Cytoplasmicproteins have mostly hydrophilic (polar) amino acids on their surface and are therefore water soluble, with more hydrophobic groups located on the interior of the protein, sheltered from the aqueous environment. Each protein has a distinct and characteristic solubility in a defined environment and any changes to those conditions (buffer or solvent type, pH, ionic strength, temperature, etc.) can cause proteins to lose the property of solubility and precipitate out of solution. The environment can be manipulated to bring about a separation of proteins- for example, the ionic strength of the solution can be increased or decreased, which will change the solubility of some proteins.
Ionic Strength and Protein Folding. This figure shows the effect of ion concentration on protein folding.

Fisiologi Molekuler (Nisa RM) 16
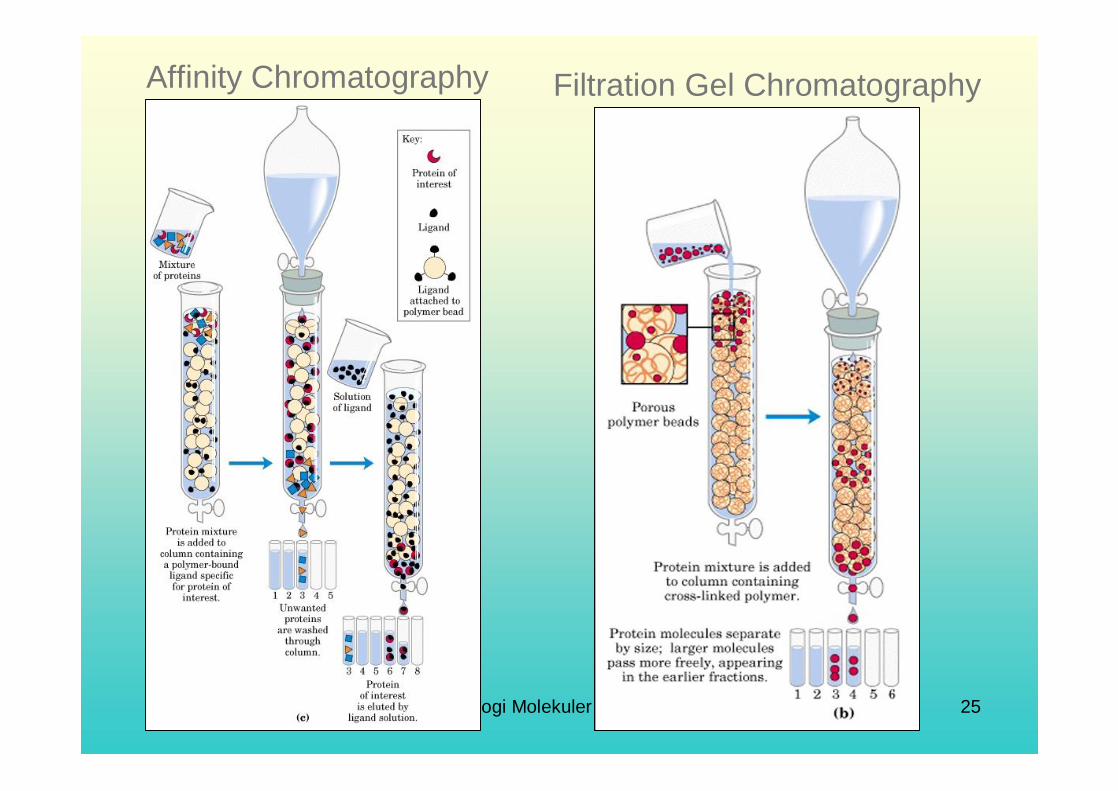
Biological Affinity (Function): Proteins often interact with other molecules in vivo in a specific way- in other words, they have a biological affinity for that molecule. These molecular counterparts, termed ligands, can be used as “bait” to “fish” out the target protein.Affinity chromatography is a very common method for purifying recombinant proteins (proteins produced by genetic engineering). Several histidine residues can be engineered at the end of a polypeptide chain. Since repeated histidines have an affinity for metals, a column of the metal can be used as bait to “catch” the recombinant protein.
Fisiologi Molekuler (Nisa RM) 17
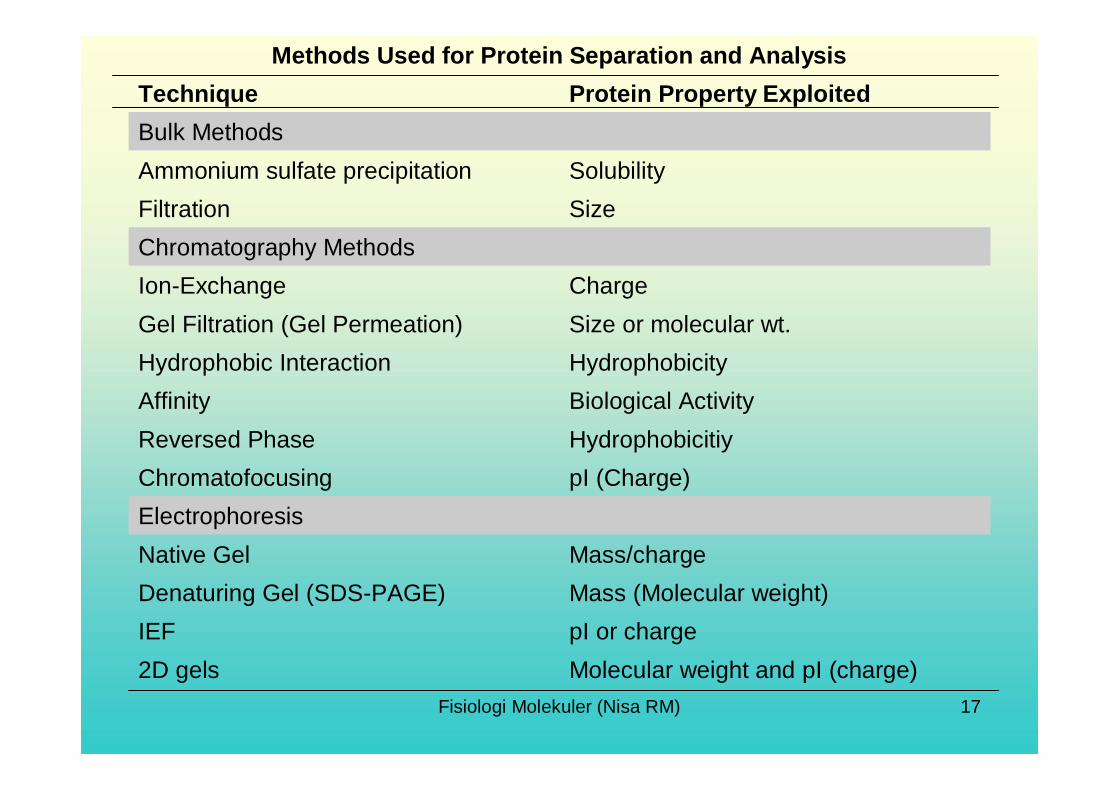
Molecular weight and pI (charge)2D gelspI or chargeIEFMass (Molecular weight)Denaturing Gel (SDS-PAGE)Mass/chargeNative Gel
ElectrophoresispI (Charge)ChromatofocusingHydrophobicitiyReversed PhaseBiological ActivityAffinityHydrophobicityHydrophobic InteractionSize or molecular wt.Gel Filtration (Gel Permeation)ChargeIon-Exchange
Chromatography Methods SizeFiltrationSolubilityAmmonium sulfate precipitation
Bulk MethodsProtein Property ExploitedTechnique
Methods Used for Protein Separation and Analysis
Fisiologi Molekuler (Nisa RM) 18http://www.courses.cm.utexas.edu/.../ch339k/overheads-1.htm (20-9-2007).

Fisiologi Molekuler (Nisa RM) 19
http://www.ucl.ac.uk/~ucbcdab/enzpur/amso4.htm

Fisiologi Molekuler (Nisa RM) 20
A number of different methods can be used to reduce hydrophilic interactions and precipitate out proteins reasonably selectively, including:Salting out with ammonium sulphate
Selective precipitation with an organic solvent that both reduces available water and also decreases the dielectric constant of the solution. Ethanol fractionation is commonly used to separate protein fractions from blood plasma. Historically, acetone precipitation was an important way of obtaining a dry powder that could be stored for some time without losing activity;
Selective precipitation using non-ionic polymers such as polyethylene glycol, which both reduce the available water and may also interact directly with some of the proteins in the mixture, in much the same way as do the polymers used in gel filtration. Polyethylene glycol (PEG) is a polymer with the structure (–CH2CH2O–)nthat is synthesized normally by ring opening polymerization of ethylene oxide. The polymer is usually linear at molecular weights (MWs) 10 kD.

Fisiologi Molekuler (Nisa RM) 21
Ammonium sulphate is highly hydrated, and a concentrated ammonium sulphate solution reduces the available water very considerably.The diagram on the right shows two proteins, with their hydrophilic regions coloured blue.The protein on the left has relatively few hydrophilic regions, andhence will aggregate and precipitate at a relatively low concentration of ammonium sulphate - perhaps around 20 - 30% saturation. By contrast, the protein on the right has considerably more hydrophilic regions, and hence will remain in solution until the concentration of ammonium sulphate is considerably higher - perhaps around 50 - 60% saturation. This means that it is possible to separate proteins from a mixture on the basis of their relative hydrophilicity by gradually increasing the concentration of ammonium sulphate.

Fisiologi Molekuler (Nisa RM) 22
Ion Exchange Chromatography

Fisiologi Molekuler (Nisa RM) 23
Ion Exchange Chromatography

Fisiologi Molekuler (Nisa RM) 24
Ion chromatography is used for the separation and analysis of ions (both anions and cations).
The mode of separation is called ion exchange and it is based on the premise that different sample ions migrate through the separator column at different rates, depending upon their interactions with the ion exchange sites of the packing material (ion-exchange resins bonded to inert polymeric particles).
The ion-resin interaction is unique and characteristic for each ion for a given resin. By comparing the data obtained from a sample to those obtained using known standard solutions, sample ions can be identified and quantitated.
The common detector is a conductivity cell, which measures the electrical conductance of the sample ions. Amperometry, UV-VIS absorbance, photodiode array, and mass spectrometry detectors can also be used.
Fisiologi Molekuler (Nisa RM) 25
Affinity Chromatography Filtration Gel Chromatography

Fisiologi Molekuler (Nisa RM) 26

Fisiologi Molekuler (Nisa RM) 27
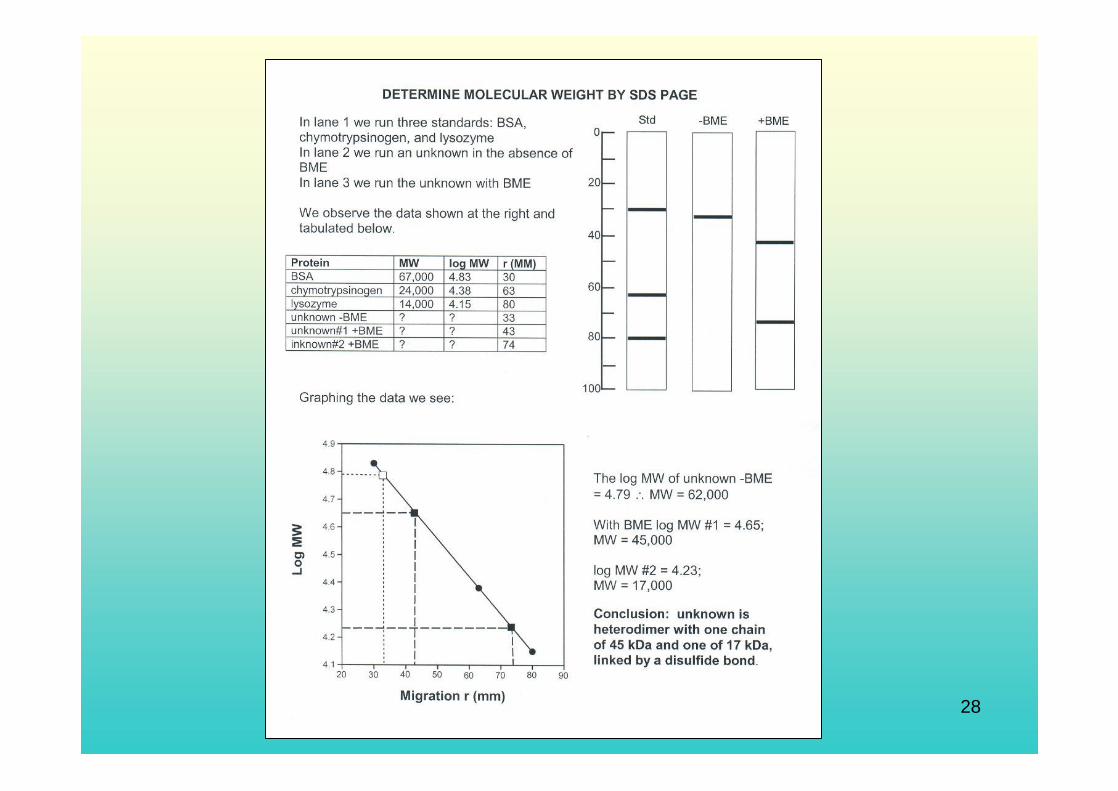
Fisiologi Molekuler (Nisa RM) 28

Fisiologi Molekuler (Nisa RM) 29

Fisiologi Molekuler (Nisa RM) 30

Fisiologi Molekuler (Nisa RM) 31

Fisiologi Molekuler (Nisa RM) 32

Fisiologi Molekuler (Nisa RM) 33

Fisiologi Molekuler (Nisa RM) 34
Purification• Make your protein as pure as possible• One single band on an overloaded gel• Make sure that no aggregates are present, or
mixtures of e.g. monomers and dimers (check with e.g. dynamic light scattering)
• Concentrate your protein to about 10 mg/ml• Do not use high buffer and salt concentrations in
your final prep for crystallization• Researchers have been successful with
crystallizing His-tagged proteins
(Dijkstra 2006)
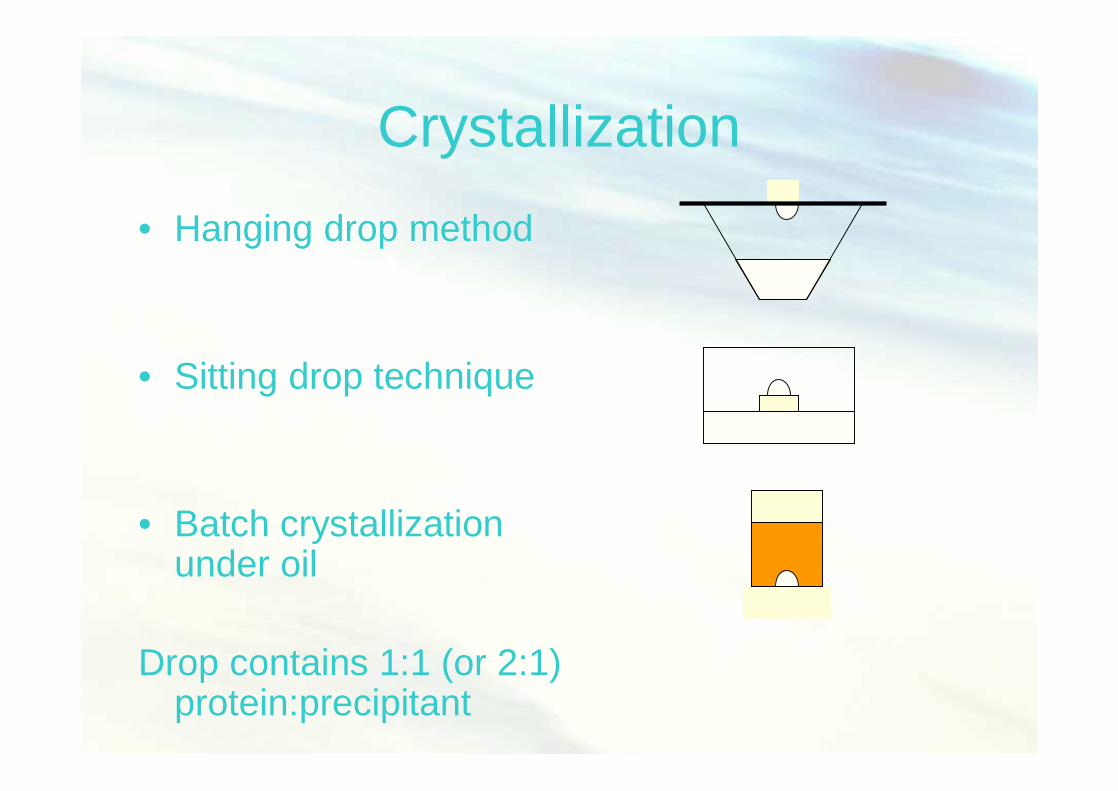
Fisiologi Molekuler (Nisa RM) 35
Crystallization• Hanging drop method
• Sitting drop technique
• Batch crystallizationunder oil
Drop contains 1:1 (or 2:1)protein:precipitant

Fisiologi Molekuler (Nisa RM) 36
Crystallization screens
• Crystal Screens 1 and 2• Wizard screens 1 and 2• PEG screens• Additive screen
• Beware: calcium + phosphate easily gives calcium-phosphate crystals!

Fisiologi Molekuler (Nisa RM) 37
What is a good crystal?
• Dimension 0.2 x 0.2 x 0.2 mm• No scratches or weird outgrows• Sharp edges
• It should diffract to high resolution• It should give a “clean” diffraction pattern• It should have a low “mosaicity”

Fisiologi Molekuler (Nisa RM) 38
Aquifex aeolicus amylomaltase
• Best condition: 100 mM Tris7.5, 200 mM A.S., 35% MPD (condition 11)

Fisiologi Molekuler (Nisa RM) 39
Crystals
• Are built up in 3 dimensions of repeating units (unit cell)
• in each unit cell exactly the same atoms are present at exactly the same positions

Fisiologi Molekuler (Nisa RM) 40
Crystals behave like a grating
• the atoms in a crystal can be considered to be the grooves in a three-dimensional grating
• The distances between the atoms is about 0.15 nm (0.15 x 10-9 m)
• With normal visible light ( = 400 – 650 nm) such details are not visible, but with X-rays ( = 0.1 nm) they are!

Fisiologi Molekuler (Nisa RM) 41

Fisiologi Molekuler (Nisa RM) 42
How about bio-macromolecules?
• They can also be crystallized
• They have a size of only 0.1-0.3 mm
• They contain about 50% water
• They diffract weakly -> strong X-ray sources needed!

Fisiologi Molekuler (Nisa RM) 43
X-ray diffraction of protein crystals

Fisiologi Molekuler (Nisa RM) 44
Xray diffraction patterns have symmetry
• This symmetry is caused by the symmetry in the crystal
• There are exactly 230 ways how to combine symmetry operations (2,3,4,6-fold rotations, translations, inversions, mirrors) (space groups)
• For proteins only 75 possibilities are allowed (No mirrors or inversions: L-amino acids -> D-amino not allowed
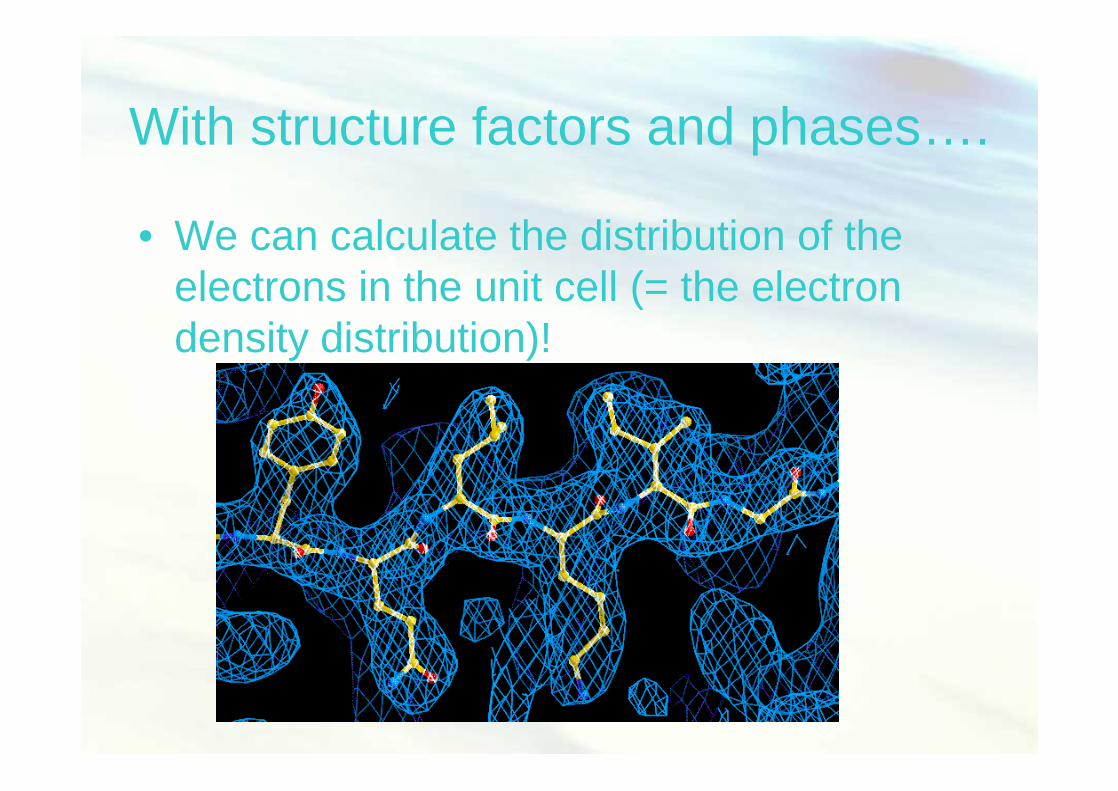
Fisiologi Molekuler (Nisa RM) 45
With structure factors and phases….
• We can calculate the distribution of the electrons in the unit cell (= the electron density distribution)!

Fisiologi Molekuler (Nisa RM) 46
Model building

Fisiologi Molekuler (Nisa RM) 47
Result of X-ray diffraction

Fisiologi Molekuler (Nisa RM) 48
Why is protein crystallography useful?
• X-ray diffraction experiments yield –a better understanding of the
biochemistry of processes in living organisms
–a better understanding of the molecular basis of some diseases
–new medicines for some diseases–new enzymes for industrial applications– ...