light and electron microscopic observations on the pigmented layers of the developing human eye
TRANSCRIPT

LIGHT AND ELECTRON MICROSCOPIC OBSERVATIONS ON T H E PIGMENTED LAYERS O F T H E DEVELOPING HUMAN EYE
MICHAEL L. MUND, M.D., MERLYN M. RODRIGUES, M.D., AND BEN S. F INE, M.D Washington, D.C.
There is considerable interest in the retinal pigment epithelium, yet relatively few studies have been carried out on the embryonic development of this tissue in the human eye.1
While observations by light microscopy have been well documented previously,2·3
similar observations by electron microscopy have been limited to a few selected periods of gestation.4"7
The purpose of this paper is to describe the developing pigment epithelium and cho-roid in a series of human embryos and fetuses. Melanogenesis in the pigment epithelium is compared to that in the choroid. The developing pigment epithelium of the retina is also compared with that of the iris. To the best of our knowledge, a study of this range of development by light and electron microscopy has not been previously reported.
MATERIALS AND METHODS
Sixty-two eyes ranging in age from six to 40 weeks' gestation were obtained from surgical specimens, stillbirths, or neonatal deaths. Spontaneous abortions were excluded, and all fetuses appeared normal.
From the Ophthalmic Pathology Branch, Armed Forces Institute of Pathology, and The George Washington University Medical Center, Washington, D.C. This study was supported in part by Public Health Service Special Fellowship Award EY-1-01; Research Grants EY-00397, EY-00133, and Training Grant EY-00032, from the National Eye Institute, National Institutes of Health, Bethesda, Maryland. Dr. Mund's present address is Beth Israel Medical Center, New York, New York. Dr. Rodrigues' present address is Wills Eye Hospital, Philadelphia, Pennsylvania.
The opinions and assertions contained herein are the private views of the authors and are not to be construed as official or as reflecting the views of the Department of the Army or the Department of Defense.
Reprint requests to Ben S. Fine, M.D., Ophthalmic Pathology Branch, Armed Forces Institute of Pathology, Washington, D.C. 20305.
Ages of the specimens were obtained from measurements and/or the mother's menstrual history.
For the purpose of comparison, postnatal irides were obtained from eyes removed for retinoblastoma at five months and at 10 months of age.
Of the 64 eyes, 36 were studied by both light microscopy and electron microscopy, and 28 eyes were studied by light microscopy only. Specimens obtained between the 27th and 40th weeks of gestation were studied only by light microscopy, since adequate material for electron microscopy was either unavailable or unsuitable.
Most of the specimens were placed in fixative within 15 to 50 minutes after the fetus or embryo was delivered from the mother. Of these specimens, most were immediately opened in a frontal plane for about 120 degrees between the equator and the limbus, and the lens and vitreous were removed to facilitate fixation, which was accomplished by immersion. Autopsy material was used up to 41 hours after death and correlated for usefulness with the material prepared after shorter delay. Various solutions were used for fixation for light microscopy: 2 or 4% glutaraldehyde for one hour ; or Zenker's solution, Helly's solution, Bouin's solution, or 10% formalin for at least 48 hours each. Helly's solution produced the best cytologie detail. Sections of the whole eye for light microscopy were cut 8 μ thick, and samplings were taken at several levels.
In general, tissues from the posterior pole, periphery, equator (size permitting), and sector portions of the anterior segment were selected for study and fixed for electron microscopy in 2 or 4% glutaraldehyde for 20 minutes to five days and postfixed in Dalton's solution for 30 to 45 minutes, then dehydrated in ascending concentrations of
167
AMERICAN JOURNAL OF OPHTHALMOLOGY
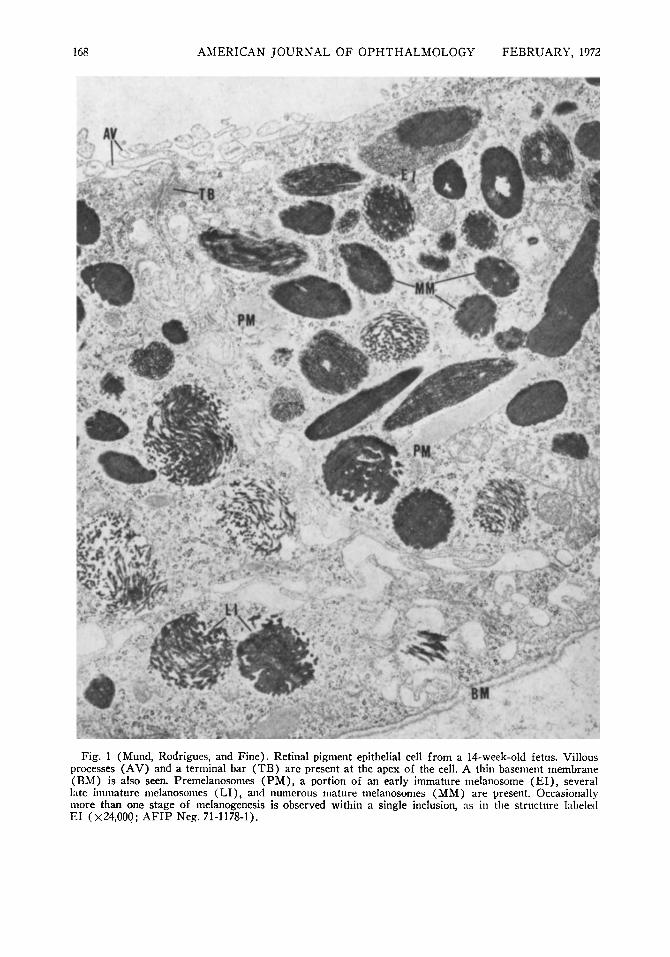
Fig. 1 (Mund, Rodrigues, and Fine). Retinal pigment epithelial cell from a 14-week-old fetus. Villous processes (AV) and a terminal bar (TB) are present at the apex of the cell. A thin basement membrane (BM) is also seen. Premelanosomes (PM), a portion of an early immature melanosome (E l ) , several late immature melanosomes (LI), and numerous mature melanosomes (MM) are present. Occasionally more than one stage of melanogenesis is observed within a single inclusion, as in the structure labeled EI (X24.000; AFIP Neg. 71-1178-1).
166 FEBRUARY, 1972

VOL. 73, NO. 2 PIGMENTATION IN THE DEVELOPING EYE 169
ethyl alcohol. The tissues were embedded in Epon and sectioned on a Reichert microtome. A few eyes that previously had been partially fixed in formalin were washed in Sorenson's buffer before starting the above procedure. Sections for electron microscopy were stained with uranyl acetate and lead citrate and examined with a Siemens I Elmis-kop. Thin sections (about 1.5 μ) for light microscopy were cut from the Epon block and stained with either toluidine blue or paraphe-nylenediamine.
We initially employed various combinations of glutaraldehyde (2 or 4%) and Dal-ton's solution. Initial fixation for 20 minutes in 2% glutaraldehyde followed by postfixation in Dalton's solution for 30 minutes was found to be most satisfactory, producing the least amount of artifacts and tissue destruction. Fetal tissue is more susceptible to distortion by fixatives than mature tissue. All electron micrographs in this paper except Figure 9 are from specimens fixed in both glutaraldehyde and Dalton's solution.
RESULTS
RETINAL PIGMENT EPITHELIUM
Cell morphology·—From six to 40 weeks' gestation, the retinal pigment epithelium was always a one-cell-thick layer of cuboidal to columnar cells with definite orientation of the base externally and the apex internally (Figs. 1 and 2) . From seven to at least 27 weeks' gestation, retinal pigment epithelial cells had basal infoldings, apical processes, and terminal bars. Lateral infoldings were found also from seven to 22 weeks' gestation, but were less prominent later. The nucleus was relatively large. Since fixation of the single specimen of six weeks' gestation was not satisfactory, a gestational age of seven weeks is considered our youngest specimen.
Cell inclusions and/or organelles— Mitochondria, prominent rough endoplasmic reticulum, smooth endoplasmic reticulum, many ribosomes and polysomes, premelano-somes, and melanosomes were found in all
cells of the retinal pigment epithelium at seven weeks' gestation, but at no stage was glycogen seen.
From the spectrum of cytoplasmic inclusions observed, a maturation sequence of the melanin granule (Figs. 3-5) could be reconstructed that began with a membrane-bounded collection of highly oriented, generally parallel, usually weakly electron-dense filaments, apparently free of melanin (pre-melanosome), and progressed to stages characterized by the appearance of melanin (mel-anosome). The immature melanosome initially (early, or filament, stage) showed sporadic development of focal densities (assumed to be melanin) on the filaments, and it progressed to completion of densification of the filaments. Then the filaments thickened to form dense rodlets (late immature, or rodlet, stage), thus retaining an internal structure. The final stage was formed by compaction of the rodlets to form an almost homogeneously dense inclusion (mature melanosome).
Particularly at the early gestational ages examined, the premelanosomes and melanosomes appeared evenly distributed throughout the cell with no preference for the apical region. Pigment granules were seen at seven weeks of development, and by light microscopy the cells appeared fully pigmented ( Fig. 6, inset).
The full range in melanosome development could be found from seven to 27 weeks of gestation. At term, only late immature and mature melanosomes were seen. No essential difference in the sequence of melano-genesis was noted between peripheral, equatorial, or posterior retinal pigment epithelium.
IRIS EPITHELIUM
Cellular configurations, inclusions, and/or organelles—Before 10 weeks of development, the inner layer of the anterior portion of the optic cup, representing the anläge of the posterior epithelial layer of the iris, was nonpigmented, after which time premelano-

170 AMERICAN JOURNAL OF OPHTHALMOLOGY FEBRUARY, 1972
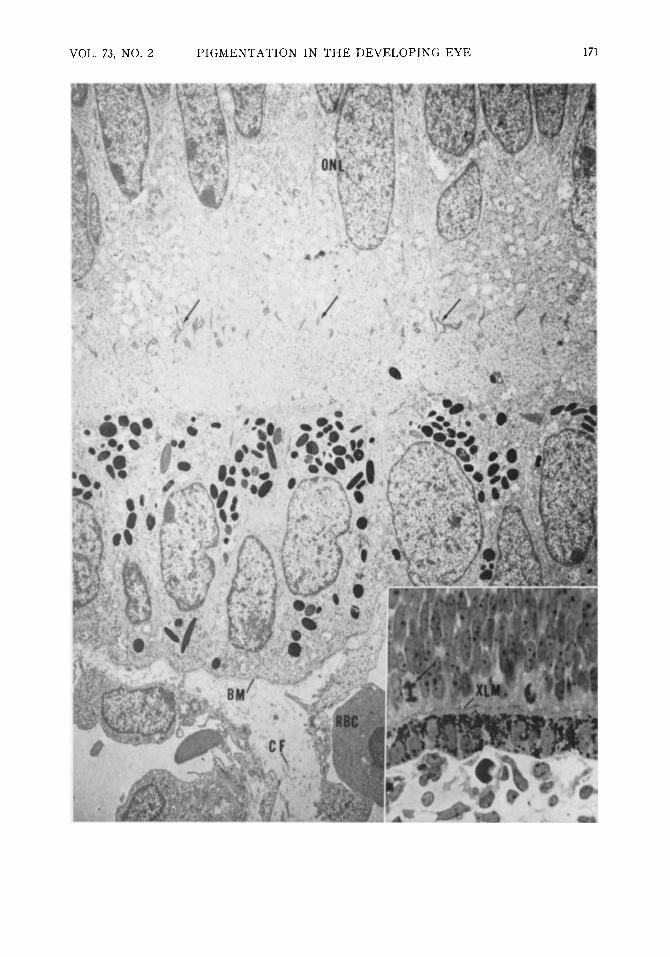
Fig. 2 (Mund, Rodrigues, and Fine). Both layers of the optic cup can be seen in apposition in this nine-week-old embryo. The outer neuroblastic layers (ONL) and external limiting membrane (arrows) are in the neural retina. Premelanosomes and melanosomes are distributed throughout the pigment epithelial cells, which appear columnar and contain relatively large nuclei. At this early stage Bruch's membrane is composed only of the basement membrane (BM) of the pigment epithelial cells and a space of variable width containing collagen fibrils (CF) and amorphous material. An immature erythrocyte (RBC) is present in the lumen of the choriocapillaris (X6000; AFIP Neg. 71-1178-2). Inset: Light microscopic picture to show apposition of the two neuroepithelial layers and the adjacent developing choriocapillaris in a nine-week-old embryo. Note the mitotic figure (arrow) in the developing outer neural retina; external limiting membrane is labeled XLM (toluidine blue X600; AFIP Neg. 70-10996).
77777r
somes and all stages of melanosomes appeared in the most marginal cells of the cup. At 10 weeks' gestation, pigment was visible by light microscopy. Pigmentation progressed by sequential melanization of cells, starting at the margin of the cup and proceeding toward the ciliary region.
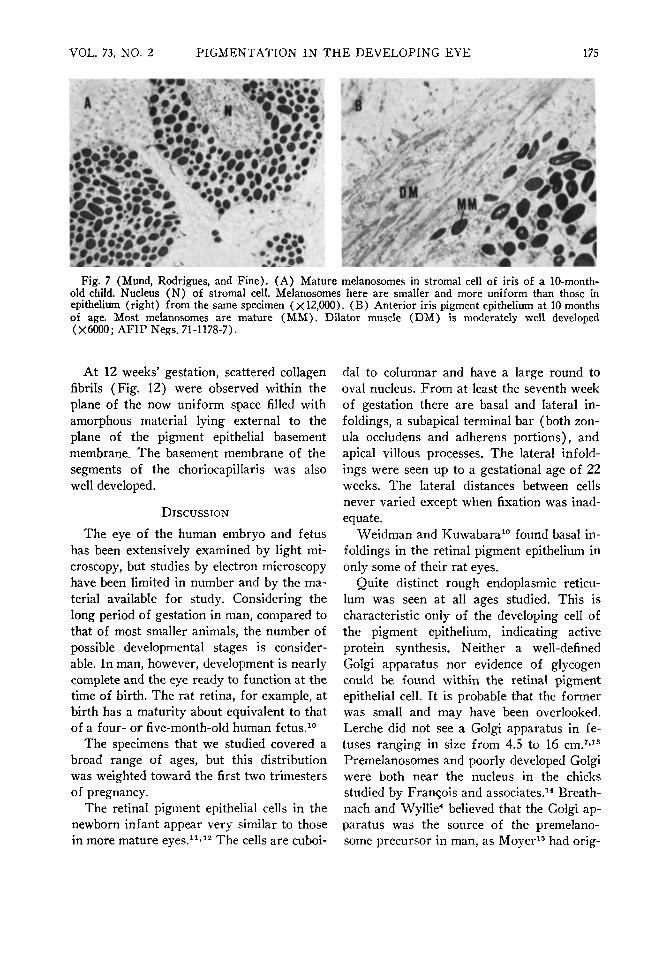
The anterior layer and most of the posterior layer of iris pigment epithelium showed similar stages of melanogenesis. Premelanosomes were found in these layers until 27 weeks of gestation. Similar stages of melanogenesis were present in both the iris epithelium and the retinal pigment epithelium. By term, only late immature and mature melanosomes were found in any portion of the epithelium of the iris. In the infants' eyes, at five months of age a greater portion of melanosomes were mature, and at 10 months of age most were mature (Fig. 7-B).
Other cytologie details in both layers of the iris epithelium (Fig. 6) could not be as well characterized as in the retinal pigment epithelium, but the cells tended to be more columnar and palisaded with similar inclusions and/or organelles at 10, 16, and 19 weeks of gestational age.
Epithelial derivatives—By light microscopy at 11 weeks' gestation, sphincter muscle appeared to arise initially from the cells of the anterior layer of neuroepithelium adjacent to the periphery of the marginal sinus. As differentiation progressed, the muscle cells appeared to develop forward from the anterior epithelial border of the marginal sinus and became more sharply demarcated from the underlying pigment cell layer from
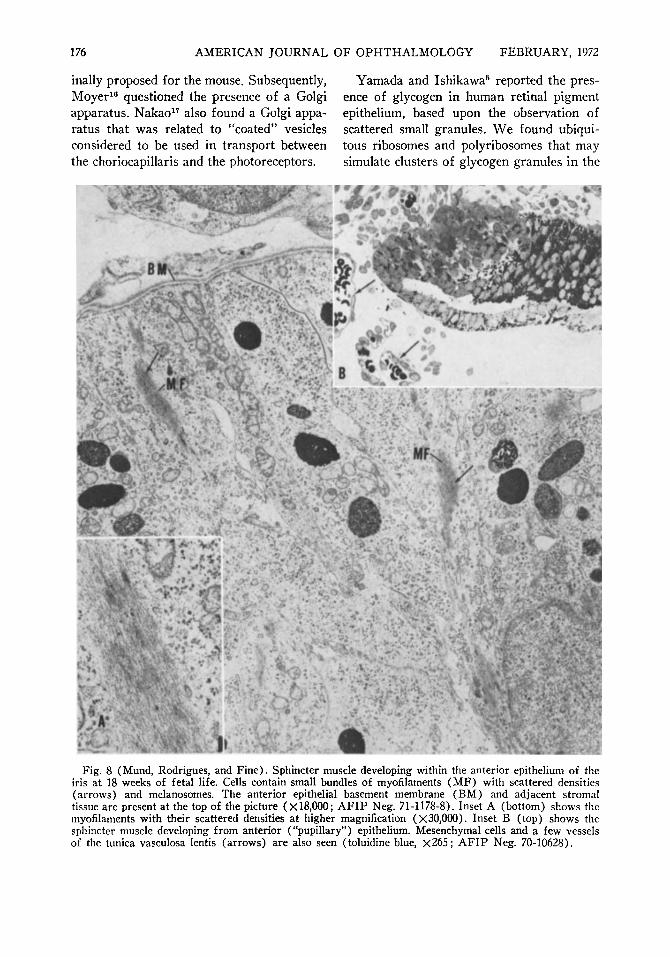
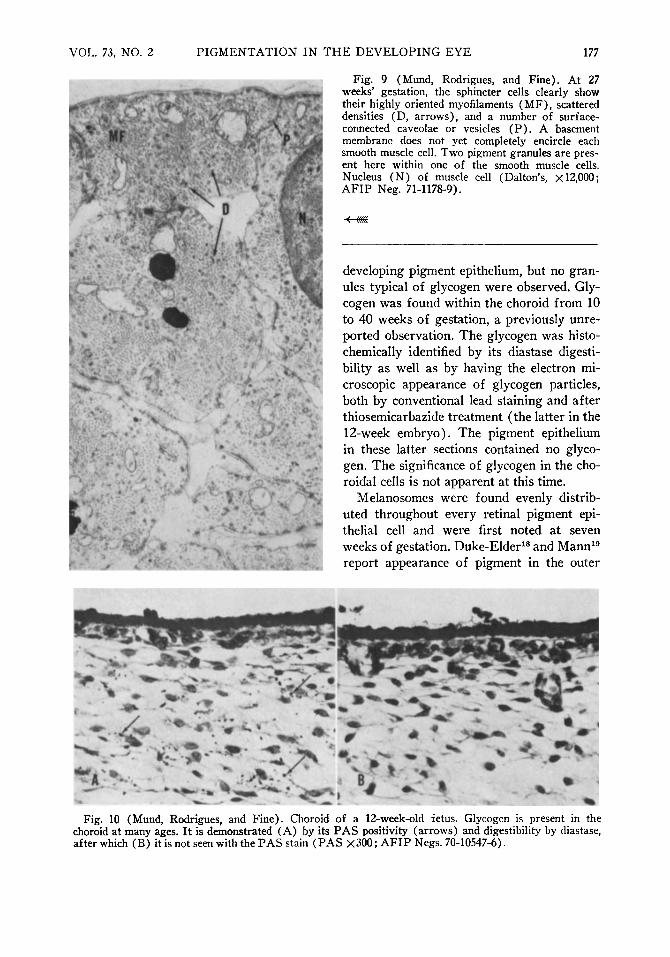
which they were derived. Though the sphincter muscle displayed very sparse deposits of pigment and later appeared relatively non-pigmented by light microscopy, immature and mature melanosomes could be identified in the earliest stage of developing muscle cells by electron microscopy (Fig. 8) . The earliest evidence of developing sphincter muscle was the presence of bundles of filaments with scattered densities (Fig. 8) lying in the cytoplasm of the cell. At a later stage, the cells displayed numbers of highly oriented filaments, scattered densities, and surface-connected caveolae or vesicles (Fig. 9) and densities. The basement membrane around each cell was frequently poorly developed or even partially lacking. At 27 weeks' gestation, mature melanosomes were still present in small numbers within the muscle cells.
PREMELANOSOME
' MELANOSOME
EARLY LATE
« v - MA TÜRE IMMATURE
Fig. 3 (Mund, Rodrigues, and Fine). Drawing of morphologic stages in the sequence of prenatal melanogenesis (AFIP Neg. 71-1178-3).
VOL. 73, NO. 2 PIGMENTATION IN THE DEVELOPING EYE 171
172 AMERICAN JOURNAL OF OPHTHALMOLOGY FEBRUARY, 1972
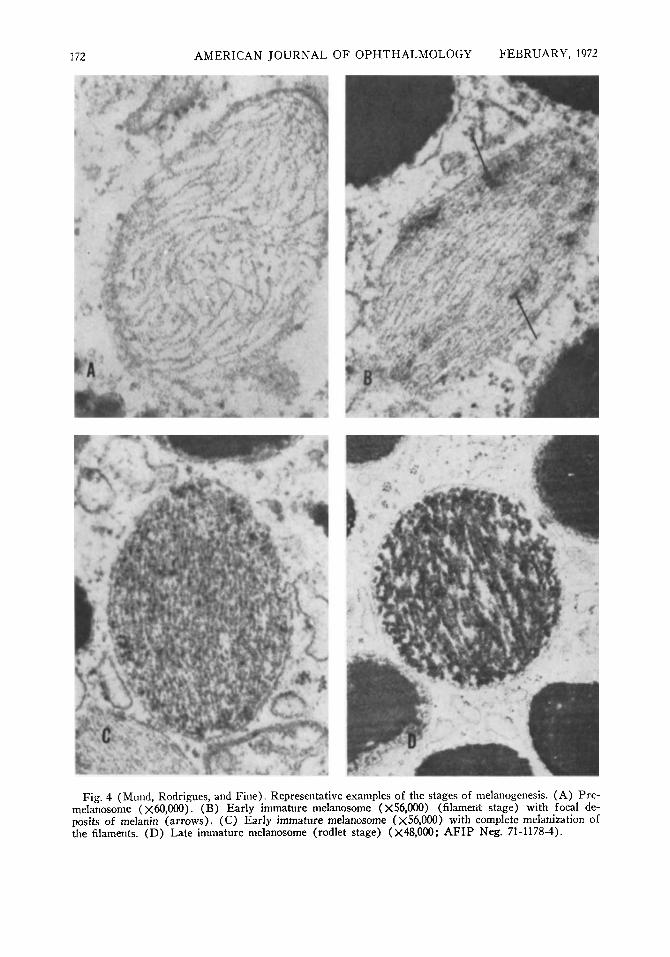
Fig. 4 (Mund, Rodrigues, and Fine). Representative examples of the stages of melanogenesis. (A) Pre-melanosome (χόΟ,ΟΟΟ). (B) Early immature melanosome (x56,000) (filament stage) with focal deposits of melanin (arrows). (C) Early immature melanosome (χ56,000) with complete melanization of the filaments. (D) Late immature melanosome (rodlet stage) (X48,000; AFIP Neg. 71-1178-4).
VOL. 73, NO. 2 PIGMENTATION IN THE DEVELOPING EYE 173
.4 :··.>*" W L vat» * f
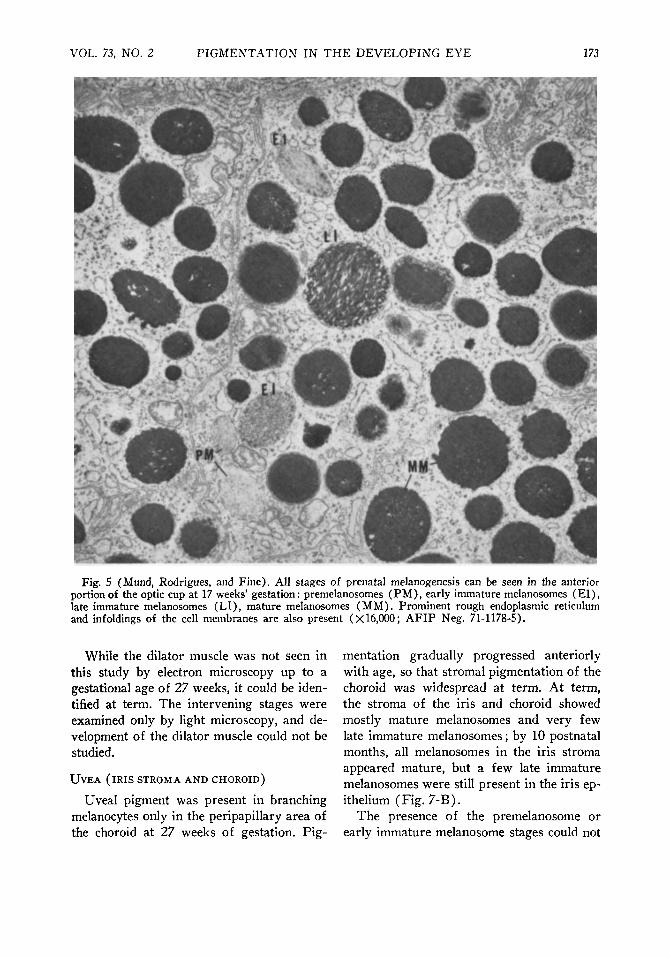
Fig. S (Mund, Rodrigues, and Fine). All stages of prenatal melanogenesis can be seen in the anterior portion of the optic cup at 17 weeks' gestation : premelanosomes (PM), early immature melanosomes (El) , late immature melanosomes (LI), mature melanosomes (MM). Prominent rough endoplasmic reticulum and infoldings of the cell membranes are also present (X 16,000; AFIP Neg. 71-1178-5).
While the dilator muscle was not seen in this study by electron microscopy up to a gestational age of 27 weeks, it could be identified at term. The intervening stages were examined only by light microscopy, and development of the dilator muscle could not be studied.
UVEA (IRIS STROMA AND CHOROID)
Uveal pigment was present in branching melanocytes only in the peripapillary area of the choroid at 27 weeks of gestation. Pig
mentation gradually progressed anteriorly with age, so that stromal pigmentation of the choroid was widespread at term. At term, the stroma of the iris and choroid showed mostly mature melanosomes and very few late immature melanosomes ; by 10 postnatal months, all melanosomes in the iris stroma appeared mature, but a few late immature melanosomes were still present in the iris epithelium (Fig. 7-B).
The presence of the premelanosome or early immature melanosome stages could not
174 AMERICAN JOURNAL
be evaluated in the uvea because of the hiatus in the specimens available for study by electron microscopy.
Lipofuscin was first seen in the iris stroma and choroid at term by electron microscopy, and such granules were not seen in the iris pigment epithelium.
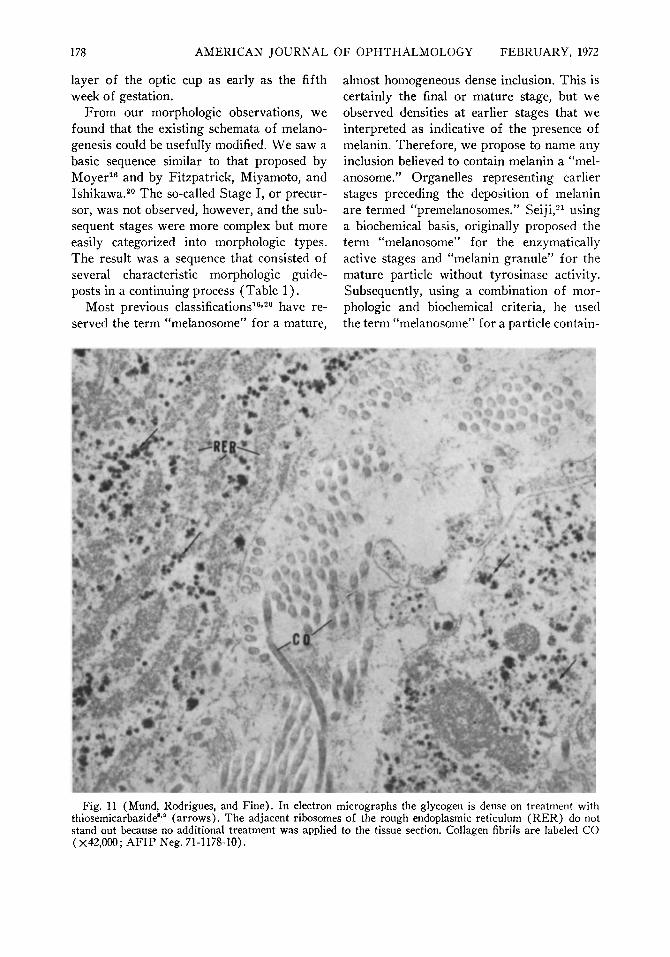
Glycogen was prominent in the cells of the choroid, especially posteriorly, at 10, 12, 19, 24, 27, 28, and 40 weeks of gestation, but was not found in all specimens in this range. The glycogen was identified by its PAS pos-itivity and digestibility with diastase by light microscopy (Fig. 10). By electron micros-
OF OPHTHALMOLOGY FEBRUARY, 1972 copy this material also stained densely with thiosemicarbazide (Fig. 11).
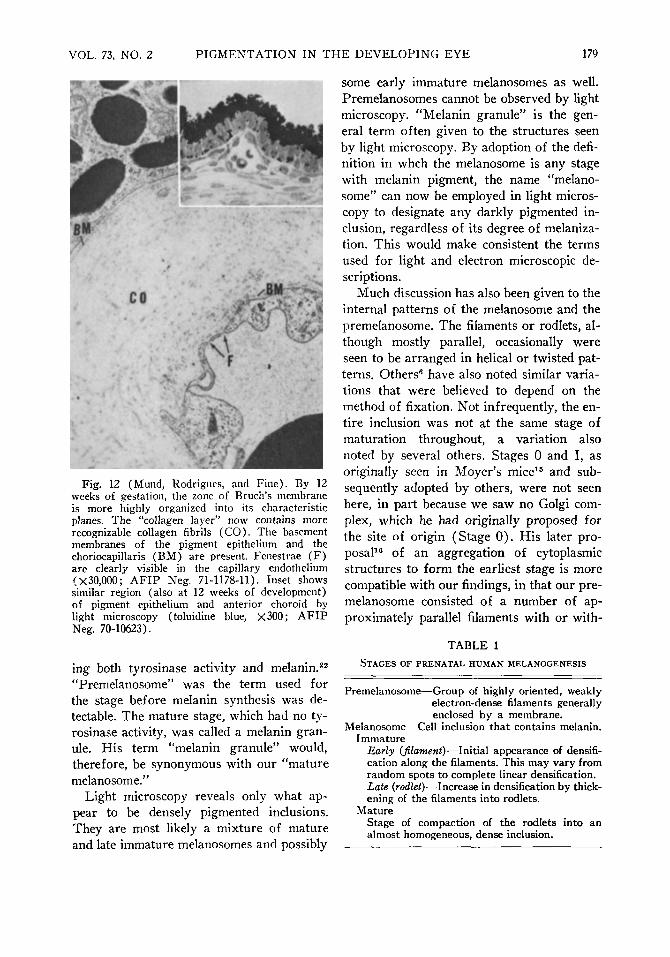
The choriocapillaris was first seen at seven weeks as an incomplete chain of capillaries outside the retinal pigment epithelium. Fenestrae were not consistently demonstrated, for reasons of technical sampling at least partly, but were clearly seen as early as 12 weeks of gestation (Fig. 12). As the fetus matured, the capillaries appeared closer together, particularly posteriorly, and a basement membrane around each one became more distinct.
Bruch's membrane developed by stages: At seven weeks of gestation, a basement membrane of the retinal pigment epithelium could be identified, and a variable space between it and the choriocapillaris was seen by electron microscopy.
At nine weeks of gestation, the basement membrane of the pigment epithelium was more dense, the space between it and the choriocapillaris was more uniform, and a delicate basement membrane was appearing around the choriocapillaris. Amorphous granular material occupied the space between the planes of the two basement membranes (Fig. 2).
At 11 weeks of gestation, Bruch's membrane first became recognizable by light microscopy, most readily seen with the aid of the Masson trichrome stain.
~4 w i
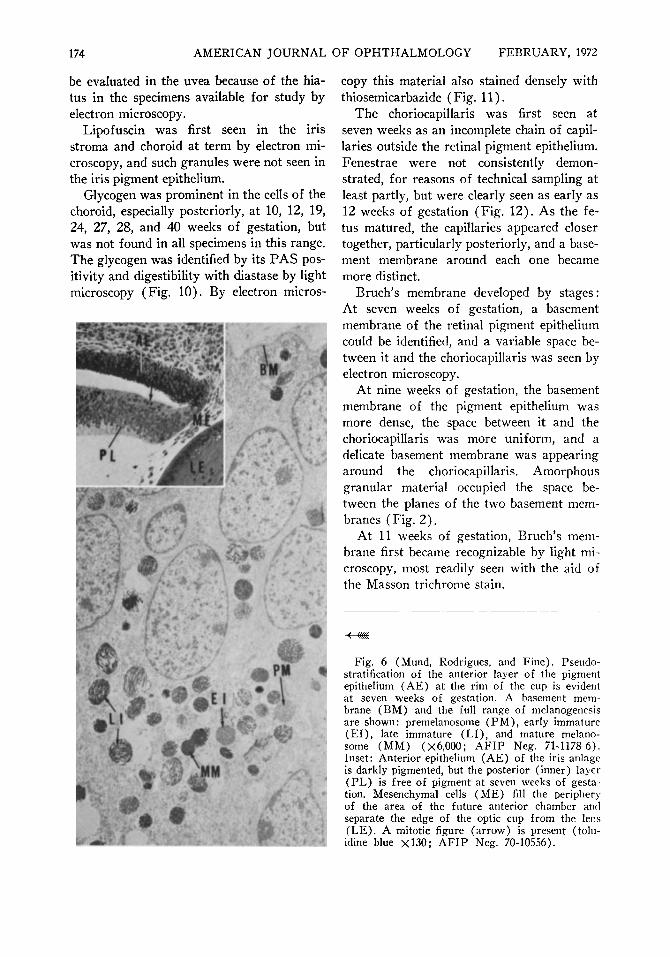
Fig. 6 (Mund, Rodrigues, and Fine). Pseudo-stratification of the anterior layer of the pigment epithelium (AE) at the rim of the cup is evident at seven weeks of gestation. A basement membrane (BM) and the full range of melanogenesis are shown: premelanosome (PM), early immature (El ) , late immature (LI), and mature melano-some (MM) (Χό,ΟΟΟ; AFIP Neg. 71-1178-6). Inset: Anterior epithelium (AE) of the iris anläge is darkly pigmented, but the posterior (inner) layer (PL) is free of pigment at seven weeks of gestation. Mesenchymal cells (ME) fill the periphery of the area of the future anterior chamber and separate the edge of the optic cup from the lens (LE). A mitotic figure (arrow) is present (tolu-idine blue χ130; AFIP Neg. 70-10556).
VOL. 73, NO. 2 PIGMENTATION IN THE DEVELOPING EYE 175
Fig. 7 (Mund, Rodrigues, and Fine). (A) Mature melanosomes in stromal cell of iris of a 10-month-old child. Nucleus (N) of stromal cell. Melanosomes here are smaller and more uniform than those in epithelium (right) from the same specimen (X 12,000). (B) Anterior iris pigment epithelium at 10 months of age. Most melanosomes are mature (MM). Dilator muscle (DM) is moderately well developed (X6000; AFIP Negs. 71-1178-7).
At 12 weeks' gestation, scattered collagen fibrils (Fig. 12) were observed within the plane of the now uniform space filled with amorphous material lying external to the plane of the pigment epithelial basement membrane. The basement membrane of the segments of the choriocapillaris was also well developed.
DISCUSSION
The eye of the human embryo and fetus has been extensively examined by light microscopy, but studies by electron microscopy have been limited in number and by the material available for study. Considering the long period of gestation in man, compared to that of most smaller animals, the number of possible developmental stages is considerable. In man, however, development is nearly complete and the eye ready to function at the time of birth. The rat retina, for example, at birth has a maturity about equivalent to that of a four- or five-month-old human fetus.10
The specimens that we studied covered a broad range of ages, but this distribution was weighted toward the first two trimesters of pregnancy.
The retinal pigment epithelial cells in the newborn infant appear very similar to those in more mature eyes.11'12 The cells are cuboi-
dal to columnar and have a large round to oval nucleus. From at least the seventh week of gestation there are basal and lateral in-foldings, a subapical terminal bar (both zon-ula occludens and adherens portions), and apical villous processes. The lateral infold-ings were seen up to a gestational age of 22 weeks. The lateral distances between cells never varied except when fixation was inadequate.
Weidman and Kuwabara10 found basal in-foldings in the retinal pigment epithelium in only some of their rat eyes.
Quite distinct rough endoplasmic reticu-lum was seen at all ages studied. This is characteristic only of the developing cell of the pigment epithelium, indicating active protein synthesis. Neither a well-defined Golgi apparatus nor evidence of glycogen could be found within the retinal pigment epithelial cell. It is probable that the former was small and may have been overlooked. Lerche did not see a Golgi apparatus in fetuses ranging in size from 4.5 to 16 cm.7'13
Premelanosomes and poorly developed Golgi were both near the nucleus in the chicks studied by François and associates.14 Breath-nach and Wyllie4 believed that the Golgi apparatus was the source of the premelano-some precursor in man, as Moyer15 had orig-
176 AMERICAN JOURNAL OF OPHTHALMOLOGY FEBRUARY, 1972
inally proposed for the mouse. Subsequently, Moyer18 questioned the presence of a Golgi apparatus. Nakao17 also found a Golgi apparatus that was related to "coated" vesicles considered to be used in transport between the choriocapillaris and the photoreceptors.
Yamada and Ishikawa5 reported the presence of glycogen in human retinal pigment epithelium, based upon the observation of scattered small granules. We found ubiquitous ribosomes and polyribosomes that may simulate clusters of glycogen granules in the
%fj* ,.; ism
Fig. 8 (Mund, Rodrigues, and Fine). Sphincter muscle developing within the anterior epithelium of the iris at 18 weeks of fetal life. Cells contain small bundles of myofilaments (MF) with scattered densities (arrows) and melanosomes. The anterior epithelial basement membrane (BM) and adjacent stromal tissue are present at the top of the picture (X18.000; AFIP Neg. 71-1178-8). Inset A (bottom) shows the myofilaments with their scattered densities at higher magnification (X30,000). Inset B (top) shows the sphincter muscle developing from anterior ("pupillary") epithelium. Mesenchymal cells and a few vessels of the tunica vasculosa lentis (arrows) are also seen (toluidine blue, χ265 ; AFIP Neg. 70-10628).
VOL. 73, NO. 2 PIGMENTATION IN THE DEVELOPING EYE 177
Fig. 9 (Mund, Rodrigues, and Fine). At 27 weeks' gestation, the sphincter cells clearly show their highly oriented myofilaments (MF), scattered densities (D, arrows), and a number of surface-connected caveolae or vesicles (P) . A basement membrane does not yet completely encircle each smooth muscle cell. Two pigment granules are present here within one of the smooth muscle cells. Nucleus (N) of muscle cell (Dalton's, X 12,000; AFIP Neg. 71-1178-9).
^ WS
developing pigment epithelium, but no granules typical of glycogen were observed. Glycogen was found within the choroid from 10 to 40 weeks of gestation, a previously unre-ported observation. The glycogen was histo-chemically identified by its diastase digestibility as well as by having the electron microscopic appearance of glycogen particles, both by conventional lead staining and after thiosemicarbazide treatment (the latter in the 12-week embryo). The pigment epithelium in these latter sections contained no glycogen. The significance of glycogen in the cho-roidal cells is not apparent at this time.
Melanosomes were found evenly distributed throughout every retinal pigment epithelial cell and were first noted at seven weeks of gestation. Duke-Elder18 and Mann19
report appearance of pigment in the outer
Fig. 10 (Mund, Rodrigues, and Fine). Choroid of a 12-week-old fetus. Glycogen is present in the choroid at many ages. It is demonstrated (A) by its PAS positivity (arrows) and digestibility by diastase, after which (B) it is not seen with the PAS stain (PAS X300; AFIP Negs. 70-10547-6).
178 AMERICAN JOURNAL OF OPHTHALMOLOGY FEBRUARY, 1972
layer of the optic cup as early as the fifth week of gestation.
From our morphologic observations, we found that the existing schemata of melano-genesis could be usefully modified. We saw a basic sequence similar to that proposed by Moyer16 and by Fitzpatrick, Miyamoto, and Ishikawa.20 The so-called Stage I, or precursor, was not observed, however, and the subsequent stages were more complex but more easily categorized into morphologic types. The result was a sequence that consisted of several characteristic morphologic guide-posts in a continuing process (Table 1).
Most previous classifications16,20 have reserved the term "melanosome" for a mature,
almost homogeneous dense inclusion. This is certainly the final or mature stage, but we observed densities at earlier stages that we interpreted as indicative of the presence of melanin. Therefore, we propose to name any inclusion believed to contain melanin a "melanosome." Organelles representing earlier stages preceding the deposition of melanin are termed "premelanosomes." Seiji,21 using a biochemical basis, originally proposed the term "melanosome" for the enzymatically active stages and "melanin granule" for the mature particle without tyrosinase activity. Subsequently, using a combination of morphologic and biochemical criteria, he used the term "melanosome" for a particle contain-
afi». * «t» " -Mr»-wr -%*x.· ■**
Fig. 11 (Mund, Rodrigues, and Fine). In electron micrographs the glycogen is dense on treatment with thiosemicarbazide8,0 (arrows). The adjacent ribosomes of the rough endoplasmic reticulum (RER) do not stand out because no additional treatment was applied to the tissue section. Collagen fibrils are labeled CO (X42,000; AFIP Neg. 71-1178-10).
VOL. 73, NO. 2 P I G M E N T A T I O N IN T H E D E V E L O P I N G E Y E 179
Fig. 12 (Mund, Rodrigues, and Fine) . By 12 weeks of gestation, the zone of Bruch's membrane is more highly organized into its characteristic planes. The "collagen layer" now contains more recognizable collagen fibrils ( C O ) . The basement membranes of the pigment epithelium and the choriocapillaris (BM) are present. Fenestrae ( F ) are clearly visible in the capillary endothelium (X30,000; A F I P Neg. 71-1178-11). Inset shows similar region (also at 12 weeks of development) of pigment epithelium and anterior choroid by light microscopy (toluidine blue, X300; A F I P Neg. 70-10623).
ing both tyrosinase activity and melanin.22
"Premelanosome" was the term used for the stage before melanin synthesis was detectable. The mature stage, which had no tyrosinase activity, was called a melanin granule. His term "melanin granule" would, therefore, be synonymous with our "mature melanosome."
Light microscopy reveals only what appear to be densely pigmented inclusions. They are most likely a mixture of mature and late immature melanosomes and possibly
some early immature melanosomes as well. Premelanosomes cannot be observed by light microscopy. "Melanin granule" is the general term often given to the structures seen by light microscopy. By adoption of the definition in whch the melanosome is any stage with melanin pigment, the name "melanosome" can now be employed in light microscopy to designate any darkly pigmented inclusion, regardless of its degree of melaniza-tion. This would make consistent the terms used for light and electron microscopic descriptions.
Much discussion has also been given to the internal patterns of the melanosome and the premelanosome. The filaments or rodlets, although mostly parallel, occasionally were seen to be arranged in helical or twisted patterns. Others6 have also noted similar variations that were believed to depend on the method of fixation. Not infrequently, the entire inclusion was not at the same stage of maturation throughout, a variation also noted by several others. Stages 0 and I, as originally seen in Moyer's mice15 and subsequently adopted by others, were not seen here, in part because we saw no Golgi complex, which he had originally proposed for the site of origin (Stage 0) . His later proposal16 of an aggregation of cytoplasmic structures to form the earliest stage is more compatible with our findings, in that our premelanosome consisted of a number of approximately parallel filaments with or with-
TABLE 1 STAGES OF PRENATAL HUMAN MELANOGENESIS
Premelanosome—Group of highly oriented, weakly electron-dense filaments generally enclosed by a membrane.
Melanosome—Cell inclusion that contains melanin. Immature
Early (filament)—Initial appearance of densifi-cation along the filaments. This may vary from random spots to complete linear densification. Late (rodlet)—Increase in densification by thickening of the filaments into rodlets.
Mature Stage of compaction of the rodlets into an almost homogeneous, dense inclusion.

180 AMERICAN JOURNAL OF OPHTHALMOLOGY FEBRUARY, 1972
out an encircling membrane. There is no direct evidence for this origin, however. Fee-ney and associates6 were unable to find a Stage I by Moyer's classification. Lerche7
saw in one case an outpouching from the cell nucleus that he thought was a precursor to the premelanosome, but we could not make a similar correlation. Outpouching of only one nuclear membrane was seen in all of our specimens, and this was not associated with pigment granules. We do not believe that the absence of a vesicle precursor or the presence of premelanosomes and melanosomes without definite limiting membranes is an artifact of processing or fixation, as discussed by Breathnach and Wyllie.4 Membranes were seen in both well-fixed and poorly fixed specimens and in cells with intact adjacent structures.
The concept of a possible mitochondrial origin for the premelanosome has been generally discarded, based upon a number of studies involving experimental animals. Seiji and associates21 separated the mitochondrial and melanosomal fractions by ultracentrifu-gation over a specific gravity gradient and verified this separation by electron microscopy. Tyrosinase activity was found mainly in the melanosomal fraction, and succinoxi-dase mainly in the mitochondrial fraction.
Maul23 believed that formation of the premelanosome is not necessarily dependent on the Golgi apparatus. He demonstrated a direct membranous continuity between premelanosomes and tubular smooth endoplasmic reticulum in vitro. Subsequently, Maul and Brumbaugh24 assessed the role of coated vesicles in the transport of tyrosinase in fowls. They were unable to demonstrate a reaction product of DOPA in early premelanosomes, but this reaction was present in the anastomosing membrane tubules and coated vesicles, most of which were connected with the Golgi apparatus.
The anterior continuation of the outer layer of the optic cup develops into the pig-mented ciliary epithelium and anterior epi
thelium of the iris. This turns around at the edge of the cup to form the posterior epithelium of the iris, the only pigmented portion of the inner layer of the cup. The structural characteristics of these cells are similar to those of the retinal pigment epithelium.
Up to 10 weeks of gestation, this posterior layer is nonpigmented and the anterior layer fully pigmented, as seen by light microscopy. Pigment can then be seen posteriorly by both light and electron microscopy. A full range of melanogenesis is present near the margin of the cup. The pattern of pigmentation of the remainder of this layer is variable, in some fetuses being complete by 22 weeks but in most of them not until 32 weeks.
According to Duke-Elder,18 pigmentation starts at the 20th week of fetal life and reaches the ciliary body by the 28th week.
The development of the melanosome in the epithelia of the iris was identical to that in the retinal pigment epithelium. Premelanosomes were present in this study until 27 weeks of gestation in both areas. Because of the incompleteness of our series, we cannot determine more precisely the latest appearance of premelanosomes and the earliest appearance of immature melanosomes. In our observations the maturity of the pigment inclusions in the anterior segment was never more advanced than in the posterior segment, as reported by Feeney and associates.6
Duke-Elder18 also records that the first sign of iris sphincter occurs at the 12th week of gestation and of the dilator muscle at the 24th week. By light microscopy, sphincter muscle arising from the anterior surface of the most medial cells of the anterior epithelium is seen at 11 weeks' gestation, but consistently seen only from the 15th week, correlating well with classic concepts. Interestingly, striated muscle has been found arising from those same regions in the second week in the chick embryo.25 By electron microscopy, structures typical of smooth muscle appear to carry some pigment with them as they grow into the loose mesenchyme found

VOL. 73, NO. 2 PIGMENTATION IN THE DEVELOPING EYE 181
in the anterior chamber, from which the iris stroma will be derived. The initial finding of structures typical of smooth muscle within the epithelial cell and the continued presence of pigment in the cell confirms the origin of the muscle from the pigment epithelial cells.
Dilator muscle was seen only at term by light and electron microscopy in this study, but it is reported to appear at six months of gestation.18
Uveal pigmentation is more erratic. By light microscopy choroidal melanocytes contained pigment from 27 weeks of fetal life onward, regardless of race. Electron microscopy of the posterior choroid at 24 weeks' gestation failed to reveal the presence of melanin, suggesting that choroidal stromal mel-anization probably occurs between these periods with fairly rapid progression, so that the early stages are not identified. Electron microscopy of posterior choroid taken from a formalin-fixed paraffin block at 32 weeks' gestation showed late immature (rodlet) and mature melanosomes in the stroma, but tissue taken at term revealed mainly mature melanosomes, with very few rodlet forms.
Lipofuscin, the "wear-and-tear" pigment, was initially noted in the retinal pigment epithelium in specimens two years of age by Feeney, Grieshaber, and Hogan" and in specimens aged 16 months by Streeten.26 This was consistent with our findings because we noted definite lipofuscin in the choroid and iris stroma at 40 weeks' gestation. In our cases no lipofuscin was seen in any of the epithelial layers or up to 10 months of age in the iris epithelium. It is worthwhile to emphasize their presence at birth if they truly are related to aging.
SUMMARY
The development of the pigment epithelium of the retina and iris was studied in 64 human eyes. Specimens ranged from a seven-week-old embryo to the iris of a 10-month-old child. A sequence of melanogen-esis is proposed, based on distinct morpho
logic criteria. Glycogen was not detected in the retinal pigment epithelium but was found for the first time in the choroid of the embryo and the fetus. Development of the sphincter smooth muscle from neuroepithe-lial cells was confirmed.
REFERENCES
1. L'Espérance, F. A., Jr. : Annual review. The retina and optic nerve. Arch. Ophth. 83:771, 1970.
2. Duke-Elder, S. (ed.) : System of Ophthalmology, vol. I l l , part 1. London, Kimpton, 1963.
3. Mann, I. : The Development of the Human Eye, 3rd ed. New York, Grune and Stratton, 1969.
4. Breathnach, A. S., and Wyllie, L. M. : Ultra-structure of retinal pigment epithelium of the human fetus. J. Ultrastruct. Res. 16:584, 1966.
5. Yamada, E., and Ishikawa, T. : Some observations on the submicroscopic morphogenesis of the human retina. In Rohen, J. W. (ed.) : Eye Structure, II Symposium. Stuttgart, Schattauer-Verlag, 1965, p. 5.
6. Feeney, L., Grieshaber, J. A., and Hogan, M. J. : Studies on human ocular pigment. In Rohen, J. W. (ed.): Eye Structure, II Symposium. Stuttgart, Schattauer-Verlag, 1965, p. 535.
7. Lerche, W., and Wulle, K. G. : Über die genese der melaningranula in der embryonalen menschlichen retina. Z. Zellforsch. 76:452, 1967.
8. Seligman, A. M., Hanker, J. S., Wasserkrug, H., Dmochowski, H., and Katzoff, L. : Histochemi-cal demonstration of some oxidized macromolecules with thiocarbohydrazide (TCH) or thiosemicarba-zide (TSC) and osmium tetroxide. J. Histochem. Cytochem. 13:629, 1965.
9. Berkow, J. W., and Fine, B. S. : Glycogen in normal human iris pigment epithelium. Am. J. Ophth. 69 :994, 1970.
10. Weidman, T. A., and Kuwabara, T. : Postnatal development of the rat retina. Arch. Ophth. 79 : 470, 1968.
11. Fine, B. S. : Limiting membranes of sensory retina and pigment epithelium. Arch. Ophth. 66:847, 1961.
12. Garron, L. K. : The ultrastructure of the retinal pigment epithelium with observations on the choriocapillaris and Bruch's membrane. Tr. Am. Ophth. Soc. 61:545, 1963.
13. Lerche, W. : Electronenmikroskopische Untersuchungen zur differenzierung des pigmentepi-thels und der ausseren kornerzellen (sinneszellen) in menschlichen auge. Z. Zellforsch. 58:953, 1963.
14. François, J., Rabaey, M., and Lagassee, A. : Electron microscopic observations on choroid, pigment epithelium and pecten of the developing chick in relation to melanin svnthesis. Ophthalmologica 146:415,1963.
15. Moyer, F. : Electron microscopic observations on the origin, development and genetic control of

182 AMERICAN JOURNAL OF OPHTHALMOLOGY FEBRUARY, 1972
melanin granules in the mouse eye. In Smelser, G. K. (ed.) : Structure of the Eye. New York, Academic Press, 1961, p. 469.
16. Moyer, F. : Genetic effects of melanosome fine structure and ontogeny in normal and malignant cells. Ann. New York Acad. Sei. 100:584, 1963.
17. Nakao, F. : Electron microscopic studies of the retinal pigment epithelium. Acta Soc. Ophtli. Jap. 72:1049, 1968.
18. Duke-Elder, S. (ed.) : System of Ophthalmology, vol. I l l , part 1. London, Kimpton, 1963, p. 101.
19. Mann, I. : The Development of the Human Eye, 3rd ed. New York, Grune and Stratton, 1969, p. 79.
20. Fitzpatrick, T. B., Miyamoto, M., and Ishi-kawa, K. : The evolution of concepts of melanin biology. Arch. Derm. 96:305, 1967.
21. Seiji, M., Fitzpatrick, T. B., and Birbeck, M. S. C. : The melanosome : A distinctive subcellular particle of mammalian melanocytes and the site of melanogenesis. J. Invest. Derm. 36:243, 1961.
22. Seiji, M. : Formation of mammalian melanin. Jap. J. Derm. (Ser. B) 73:4, 1963.
23. Maul, G. G. : Golgi-melanosome relationships in human melanoma in vitro. J. Ultrastruct. Res. 26:163, 1969.
24. Maul, G. G., and Brumbaugh, J. A. : On the possible function of coated vesicles in melanogenesis of the regenerating fowl feathers. J. Cell Biol. 48:41, 1971.
25. Jensen, G. E., Zimmerman, B. L., and Ts'o, M. O. M. : Embryogenesis of striated muscle in the chick iris. In preparation.
26. Streeten, B. W. : The sudanophilic granules of the human retinal pigment epithelium. Arch. Ophth. 66:391, 1961.
OPHTHALMIC MINIATURE
The theory of colours, with all these marvellous and complicated relations, was a riddle which Goethe in vain attempted to solve ; nor were we physicists and physiologists more successful. I include myself in that number ; for I long toiled at the task, without getting any nearer my object, until I at last discovered that a wonderfully simple solution had been discovered at the beginning of this century, and had been in print ever since for anyone to read who chose. This solution was found and published by the same Thomas Young who first showed the right method or arriving at the interpretation of Egyptian hieroglyphics. He was one of the most acute men who ever lived, but had the misfortune to be too far in advance of his contemporaries. They looked on him with astonishment, but could not follow his bold speculations, and thus a mass of his most important thoughts remained buried and forgotten in the Transactions of the Royal Society until a further generation by slow degrees arrived at the rediscovery of his discoveries, and came to appreciate the force of his arguments and the accuracy of his conclusions.
Hermann von Helmholtz Popular Lectures on Scientific Subjects
Longmans, Green, 1893