journal of paleontology - west virginia...
TRANSCRIPT

Journal of Paleontologyhttp://journals.cambridge.org/JPA
Additional services for Journal of Paleontology:
Email alerts: Click hereSubscriptions: Click hereCommercial reprints: Click hereTerms of use : Click here
Generic concepts in the Actinocrinitidae Austin and Austin, 1842 (classCrinoidea) and evaluation of generic assignments of species
Elizabeth C. Rhenberg, William I. Ausich and Thomas W. Kammer
Journal of Paleontology / Volume 89 / Issue 01 / January 2015, pp 1 - 19DOI: 10.1017/jpa.2014.2, Published online: 09 March 2015
Link to this article: http://journals.cambridge.org/abstract_S002233601400002X
How to cite this article:Elizabeth C. Rhenberg, William I. Ausich and Thomas W. Kammer (2015). Generic concepts in the Actinocrinitidae Austin andAustin, 1842 (class Crinoidea) and evaluation of generic assignments of species. Journal of Paleontology, 89, pp 1-19doi:10.1017/jpa.2014.2
Request Permissions : Click here
Downloaded from http://journals.cambridge.org/JPA, IP address: 157.182.154.151 on 10 Mar 2015

Generic concepts in the Actinocrinitidae Austin and Austin, 1842(class Crinoidea) and evaluation of generic assignments of species
Elizabeth C. Rhenberg1, William I. Ausich1, and Thomas W. Kammer2
1 Geology Department, Earlham College, Richmond, VA 47374 ⟨[email protected]⟩2Department of Geology and Geography, West Virginia University, Morgantown, WV 26506-6300 ⟨[email protected]⟩; ⟨[email protected]⟩
Abstract.—The family Actinocrinitidae was a significant contributor to the global biodiversity peak of crinoids that occur-red during the Mississippian and is referred to as the “Age of Crinoids.” Although the actinocrinitids are a major componentof that high diversity, they are also a source of much taxonomic confusion. Previously, generic concepts were not appliedequally between Europe and North America creating disparity in the definition of genera. In this contribution, all genera aredefined objectively by discrete characters, and the generic assignments of all species are reevaluated. Twenty genera aredescribed. A total of 206 species were evaluated of which 56 species and one taxon in open nomenclature are reassigned todifferent genera, 21 species are designated as nomina dubia, and three species and one genus are now incertae sedis.
A phylogenetic hypothesis is presented for the relationships of the genera of Actinocrinitidae genera based on aparsimony-based analysis and plotted against stratigraphic ranges. Although groupings were revealed in this analysis,the Actinocrinitidae cannot be readily subdivided into subfamilies. Rapid diversification occurred during theTournaisian following the Hangenberg Extinction of probable fish predators. The late Devonian (Famennian) occurrenceof the highly derived genera Abactinocrinus and Physetocrinus suggests there is a more extensive, but undocumented,evolutionary history for the Actinocrinitidae during the Devonian.
Introduction
The Actinocrinitidae is an important family of Mississippian cri-noids due to their considerable genus diversity and abundance ofspecimens. Actinocrinitids encompass a large degree of morpho-logical disparity produced by the complex arrangements of theirmulti-plated calyxes. Because there are innumerable specimens inmany Mississippian faunas and because we suspect that hybridi-zation was a complicating factor as demonstrated in Eretmocrinus(Ausich and Meyer, 1994), and suggested for the Batocrinidae(Ausich and Kammer, 2010), this is a complex group. BothWachsmuth and Springer (1897) and Bowsher (1955) worked toclarify generic definitions among North American actinocrinitids,but many issues still remain. First and most significant amongthese problems is that the generic concept for Actinocrinites, astypified by Actinocrinites triacontadactylus Miller, 1821, the typespecies of the genus, has not been applied consistently to NorthAmerican faunas.
Ausich and Sevastopulo (2001) designated a neotype forActinocrinites triacontadactylus; and with this definition re-established, Ausich and Sevastopulo (2001) and Ausich andKammer (2006) made generic reassignments to all actinocrinitidsfrom Ireland, England, and Wales. Species-level systematics hasonly been completed for Ivorian faunas (Ausich and Sevastopulo,2001). A total of six actinocrinitid genera are now recognized fromIreland, England, and Wales.
The primary source of confusion regarding the systematicsof North American (and other) actinocrinitids is that genus-leveldiagnostic characters that define European actinocrinitids have
been applied as species-level diagnostic characters among NorthAmerican forms. For example, ray grouping and lobation helpsto define European genera, but this has been used to differentiateNorth American species of Actinocrinites. Unfortunately, thesolution for this is reassignment of many well-known NorthAmerican species of “Actinocrinites” to other genera. Althoughnomenclaturally messy, this situation is not surprising given thatmany nineteenth century broadly defined genera have beensubdivided into numerous new genera, such as Cyathocrinites,Phanocrinus, Platycrinites, and Poteriocrinites (see Webster,2003). Recognition of well-defined generic concepts is a pre-requisite for a modern understanding of the phylogeny, paleo-biogeography, and evolutionary paleoecology of these crinoids.
This study is a continuation of the efforts to redefine thegeneric concepts of monobathrid camerates (Ausich and Kammer,2008, 2009, 2010). Revising the camerates is an important step incompiling an accurate database to evaluate their evolutionarypaleoecology of Mississippian crinoids, including an examinationof the global, generic relationships among longevity, eurytopy, andevolutionary paleoecology (Ausich and Kammer, 2010). Thecamerates are of particular interest because they reached their peakdiversity during the Tournaisian and are one of the most diversegroups of crinoids during this time (Kammer and Ausich, 2006;Sallan et al., 2011).
In this contribution, the goals are to define all Actinocrinitidaegenera using objective characters and to assign all species to thecorrect genus. This study treated 206 species, with 57 species(and one species left in open nomenclature) reassigned to differentgenera (Table 1); 20 species were designated as nomina dubia; and
Journal of Paleontology, 89(1), 2015, p. 1–19Copyright © 2015, The Paleontological Society0022-3360/15/0088-0906doi: 10.1017/jpa.2014.2
1

Table 1. Listing of actinocrinitid species that have a revised generic assignment
Previous assignment (Webster, 2003) New assignment
?Actinocrinites polydactylus Miller, 1821 Nomen dubiumAacocrinus acylus Webster and Jell, 1999 Nomen dubiumAacocrinus algerianensis Webster et al., 2004 Thinocrinus algerianensis (Webster et al., 2004)Aacocrinus enigmaticus Webster and Lane, 1897 Thinocrinus enigmaticus (Webster and Lane, 1897)Aacocrinus triarmatus Brower, 1967 Nomen dubiumActinocrinites alatus Wright, 1955a Thinocrinus alatus (Wright, 1955a)Actinocrinites anchorensis Webster and Lane, 1987 Thinocrinus anchorensis (Webster and Lane, 1987)Actinocrinites becharensis Webster et al., 2004 Nunnacrinus becharensis (Webster et al., 2004)Actinocrinites benedicti (Miller, 1892) Thinocrinus benedicti (Miller, 1892)Actinocrinites blairi (Miller and Gurley, 1897) Thinocrinus blairi (Miller and Gurley, 1897)Actinocrinites botruosus (Miller and Gurley, 1895) Thinocrinus botruosus (Miller and Gurley, 1895)Actinocrinites brevispina (Wanner, 1924) Thinocrinus brevispina (Wanner, 1924)Actinocrinites brouweri (Wanner, 1924) Thinocrinus brouweri (Wanner, 1924)Actinocrinites c.f. brouweri (Wanner, 1924) Nomen dubiumActinocrinites carinatus (Wanner, 1937) Thinocrinus carinatus (Wanner, 1937)Actinocrinites caryocrinoides (M’Coy, 1844) Nomen dubiumActinocrinites combinatus Webster et al., 2004 Thinocrinus combinatus (Webster et al., 2004)Actinocrinites constrictus (M’Coy, 1844) Nomen dubiumActinocrinites coplowensis Wright, 1955a Sampsonocrinus coplowensis (Wright, 1955a)Actinocrinites decadactylus (Gilbertson in Portlock, 1843) Nomen dubiumActinocrinites dilatus (Wanner, 1924) Thinocrinus dilatus (Wanner, 1924)Actinocrinites erraticus (Miller and Gurley, 1893) Nunnacrinus erraticus (Miller and Gurley, 1893)Actinocrinites eximius (Kirk, 1943) Thinocrinus subpulchellus (Miller and Gurley, 1896c)Actinocrinites exornatus (Wanner, 1937) Thinocrinus exornatus (Wanner, 1937)Actinocrinites gibsoni (Miller and Gurley, 1893) Thinocrinus gibsoni (Miller and Gurley, 1893)Actinocrinites goldfussi (Schmidt, 1930) Nomen dubiumActinocrinites granulatus (Wright, 1955a) Thinocrinus granulatus (Wright, 1955a)Actinocrinites granulatus Goldfuss, 1831 Nomen dubiumActinocrinites griffithi (Wachsmuth and Springer, 1897) Thinocrinus subpulchellus (Miller and Gurley, 1896c)Actinocrinites higuchisawensis (Minato, 1951) Nomen dubiumActinocrinites intermedius Wright, 1955a Iotacrinus intermedius (Wright, 1955a)Actinocrinites laevis Miller, 1821 Nomen dubiumActinocrinites lowei (Hall, 1858) Thinocrinus lowei (Hall, 1858)Actinocrinites marcoui (Collingnon, 1924) Nomen dubiumActinocrinites multiradiatus (Shumard, 1857) Thinocrinus multiradiatus (Shumard, 1857)Actinocrinites multiramosus altidorsatus (Rowley, 1904) Thinocrinus multiramosus altidorsatus (Rowley, 1904)Actinocrinites nodosus Wright, 1955a Thinocrinus nodosus (Wright, 1955a)Actinocrinites ohmoriensis (Minato, 1951) Nomen dubiumActinocrinites permicus (Wanner, 1924) Thinocrinus permicus (Wanner, 1924)Actinocrinites pernodosus (Hall, 1858) Thinocrinus pernodosus (Hall, 1858)Actinocrinites plagosus (Miller and Gurley, 1893) Nunnacrinus plagosus (Miller and Gurley, 1893)Actinocrinites probolos Ausich and Kammer, 1991 Thinocrinus probolos (Ausich and Kammer, 1991)Actinocrinites pusillus (M’Coy, 1844) Nomen dubiumActinocrinites rotundatus Wright, 1955a Thinocrinus rotundatus (Wright, 1955a)Actinocrinites scitulus (Miller and Gurley, 1897) Thinocrinus scitulus (Miller and Gurley, 1897)Actinocrinites sedaliensis (Miller and Gurley, 1895) Sampsonocrinus sedaliensis (Miller and Gurley, 1895)Actinocrinites semimultiramosus (Whitfield, 1900) Thinocrinus semimultiramosus (Whitfield, 1900)Actinocrinites skourensis (Termier, G. and Termier, H., 1950) Nomen dubiumActinocrinites sobrinus (Miller and Gurley, 1896c) Cusacrinus sobrinus (Miller and Gurley, 1896c)Actinocrinites spectabilis (Miller and Gurley, 1896c) Cusacrinus spectabilis (Miller and Gurley, 1896c)Actinocrinites spinaetectus (Wanrer, 1937) Incertae sedisActinocrinites subpulchellus (Miller and Gurley, 1896c) Thinocrinus subpulchellus (Miller and Gurley, 1896c)Actinocrinites subscitulus (Miller and Gurley, 1896c) Cusacrinus subscitulus (Miller and Gurley, 1896c)Actinocrinites tenuis (de Koninck and Le Hon, 1854) Nomen dubiumActinocrinites tenuistriatus (Phillips, 1841) Nomen dubiumActinocrinites tricuspidatus (de Koninck and Le Hon, 1854) Dialutacrinus tricuspidatus (de Koninck and Le Hon, 1854)Actinocrinites tripus Ehlers and Kesling, 1963 Thinocrinus tripus (Ehlers and Kesling, 1963)Actinocrinites vermiculatus Wright, 1955a Thinocrinus vermiculatus (Wright, 1955a)Actinocrinites verrucosus (Hall, 1858) Thinocrinus verrucosus (Hall, 1858)Actinocrinites zhaoae Lane et al., 1997 Incertae sedisActinocrinites? batheri (Whidborne, 1896) Physetocrinus batheri (Whidborne, 1896)Blairocrinus grafensis Webster et al., 2004 Incertae sedisCactocrinus excerptus (Hall, 1861) Nomen dubiumCactocrinus fosteri (McChesney, 1861) Nomen dubiumCactocrinus reticulatus (Hall, 1861) Nunnacrinus reticulatus (Hall, 1861)Cactocrinus reticulatus ovatus (Hall, 1861a) Nunnacrinus ovatus (Hall, 1861)Cusacrinus bischoffi (Miller and Gurley, 1896c) Cactocrinus bischoffi (Miller and Gurley, 1896c)Cusacrinus gracilis (Wachsmuth and Springer, 1897) Dialutocrinus gracilis (Wachsmuth and Springer, 1897)Cusacrinus hurdianus (McChesney, 1861) Cactocrinus hurdianus (McChesney, 1861)Cusacrinus imperator (Laudon, 1933) Cactocrinus imperator Laudon, 1933Eumorphocrinus porteri (Whidborne, 1896) Incertae sedisEumorphocrinus sp. Brower, 1970 Actinocrinites sp. (Brower, 1970)Ilmocrinus Incertae sedisManillacrinus sp. Campbell and Bein, 1971 Nomen dubiumNunnacrinus armatus (de Koninck and Le Hon, 1854) Iotacrinus armatus (de Koninck and Le Hon, 1854)Nunnacrinus deornatus (de Koninck and Le Hon, 1854) Thinocrinus deornatus (de Koninck and Le Hon, 1854)Nunnacrinus sampsoni (Miller and Gurley, 1896c) Cusacrinus sampsoni (Miller and Gurley, 1896c)Nunnacrinus stellaris (de Koninck and Le Hon, 1854) Iotacrinus stellaris (de Koninck and Le Hon, 1854)Physetocrinus sampsoni Miller and Gurley, 1896c Thinocrinus? sampsoni (Miller and Gurley, 1896c)Physetocrinus smalleyi Weller, 1909 Blairocrinus smalleyi (Weller, 1909)
Journal of Paleontology 89(1):1–19 2

three species and one genus were designated incertae sedis. Thisis only a first step for a full understanding of the Actinocrinitidae,but it does allow for evaluation of regional and global patterns ofgeneric longevity, eurytopy, and evolutionary paleoecology.Significant evaluation of species-level systematics remains to becompleted but is far beyond the scope of this study.
Systematic paleontology
Where possible, all species were characterized using originaldescriptions and illustrations, subsequent descriptions andillustration of type specimens, and examination of type speci-mens. Complete synonymies are not provided as they are pub-lished in Webster (2003). Terminology follows Ubaghs (1978a)with modifications from Ausich et al. (1999) and open nomen-clature that follows Matthews (1973).
Carboniferous stratigraphy follows Heckel and Clayton(2005). Table 2 is the detailed chronostratigraphic schemeshowing the correlations used in this paper following Ausichand Kammer (2008).
Class Crinoidea Miller, 1821Subclass Camerata Wachsmuth and Springer, 1885Order Monobathrida Moore and Laudon, 1943
Suborder Compsocrinina Ubaghs, 1978bSuperfamiy Periechocrinacea Bronn, 1849
Family Actinocrinitidae Austin and Austin, 1842
Diagnosis.—The Actinocrinitidae differs from the otherthree families in the Periechocrinacea (Periechocrinidae,Amphoracrinidae, Paragaricocrinidae) in possessing a uniquecombination of characters that vary and in some cases overlapamong the four families (Ausich and Kammer, 2008, table 2). Ingeneral, actinocrinitids possess a typically conical calyx that ishigher than found in the Amphoracrinidae and Paragaricocrinidae,but somewhat lower than those in the Periechocrinidae. Thefirst primibrachial is typically hexagonal; there are few to nosecundibrachials; there are only two plates above the hexagonalprimanal; the anal tube is rarely absent; and the number of freearms ranges from 10–60, a greater range than in the other threefamilies.
Remarks.—Bowsher and Ubaghs (1978) subdivided the Acti-nocrinitidae into four subfamilies: Actinocrinitinae Austin andAustin, 1842; Eumorphocrininae Ubaghs, 1978c; CactocrininaeUbaghs, 1978d; and Physetocrininae Ubaghs, 1978e. A phylo-genetic analysis of 20 genera from the four subfamilies (andtwo outgroup genera) was performed to investigate possiblephylogenetic relationships and to test if the previously namedsubfamilies represented actual clades. Phylogenetic relationships
are inferred using parsimony-based character analysis of generausing PAUP ©4.0b10 (Swofford, 2001). The actinocrinitidgenera are defined using discrete, non-subjective characters thatdefine basic and distinctive atrributes of the calyx and tegmen.Fifteen characters with 38 character states were used to analyzethe genera (Appendices 1 and 2). These characters are unorderedand unweighted; search methods were heuristic with randomstepwise addition. The outgroup used for this analysis are twogenera from the Periechocrinidae Bronn, 1849: PeriechocrinusMorris, 1843, a Silurian and Devonian genus andMegistocrinusOwen and Shumard, 1852, a Devonian and Mississippiangenus. Characters that are used for the outgroup genera arebased on their type species, but the character states used forthe Actinocrinitidae are based on all known species for each ofthe genera. Thus, multiple character states are present for severalcharacters in most of the genera.
The subfamilies of Bowsher and Ubaghs (1978) werebased on presence or absence of arm lobes, presence or absenceof an anal tube, and how strongly the arms were groupedtogether. None of the subfamilies form a separate clade in thephylogenetic analysis (Fig. 1) completed herein. It might beargued that the genera of the subfamily Actinocrinitinae grouptogether, along with Cactocrinus, but the grouping is a subset ofthe larger clade that includes genera from the Eumorphocrininaeand Cactocrininae. As the subfamilies are scattered throughoutthe tree, the subfamily classification of the Treatise (Ubaghs,1978b) does not recognize monophyletic clades and should beabandoned. These subfamilies were each defined on four orfewer characters (Ubaghs, 1978b), in contrast to the 15 charactersused in this study (Appendix 1).
Wachsmuth and Springer (1897) subdivided the actinocri-nitids into three primary groups that included: 1) genera with ananal tube, calyx lobed, and interradials in contact with thetegmen; 2) genera with an anal tube, calyx not lobed, andinterradials not in contact with the tegmen; and 3) generawithout an anal tube, regardless of other characters. Although
Table 1. (Continued )
Previous assignment (Webster, 2003) New assignment
Sampsonocrinus cheguiaensis Webster et al., 2004 Thinocrinus cheguiaensis (Webster et al., 2004)Steganocrinus burlingtonensis Brower, 1965 Abactinocrinus burlingtonensis (Brower, 1965)Strotocrinus blairi Miller and Gurley, 1895 Teleiocrinus blairi (Miller and Gurley, 1895)Strotocrinus ornatus Miller and Gurley, 1896a Teleiocrinus ornatus (Miller and Gurley, 1896a)Strotocrinus venustus Miller and Gurley, 1893 Teleiocrinus venustus (Miller and Gurley, 1893)Teleiocrinus? sibiricus Yakovlev in Yakovlev and Ivanov, 1956 Nomen dubium
Table 2. Chronostratigraphic correlation of time units between North Americaand Europe (from Ausich and Kammer, 2008)
Time unitsNorth Americanchronostratigraphic units European chronostratigraphic units
11 upper Chesterian Serpukhovian, Arnsbergian (E2)10 middle Chesterian Serpukhovian, Pendleian (E1)9 lower Chesterian Visean, Brigantian (V3c)8 late Meramecian Visean, Asbian (V3b)7 early Meramecian Visean, Holkerian (V3a)6 late Osagean Visean, Arundian (V1b, V2a)5 late Osagean Visean, upper Chadian (V1a)4 middle Osagean Tournaisian, lower Chadian (Tn3c)3 early Osagean Tournaisian, Ivorian (Tn3a-Tn3c)2 late Kinderhookian Tournaisian, late Hastarian (Tn2)1 early Kinderhookian Tournaisian, early Hastarian (Tn1b)
3 Rhenberg et al.—Actinocrinitid concepts

these three groups are a good general way to subdivide the family,they do not reflect any phylogenetic relationship between genera.For example,Dialutocrinus lacks arm lobes but its other charactersgroup it most closely with actinocrinitids that have arm lobes, orEumorphocrinus with an anal tube relates more closely to twogenera that lack an anal tube, Strotocrinus and Physetocrinus, thanit does to the rest of its subfamily.
The generic-level diagnostic characters for genera withinActinocrinitidae are defined as follows (Table 3, Appendix 1):basal circlet height, radial circlet height, number of fixedsecundibrachials, highest brachitaxis (e.g., tertibrachials) in thevertical wall of the calyx before arm lobes, number of ranges(rows of stacked plates) in the regular interrays, number ofranges in posterior interray, interrays in contact with tegmen,plating in the proximal interrays, number of plates above theprimanal, arms grouped, arm lobes, presence or absence of fixedinterbrachials between half-rays, tegmen height compared tocalyx, number of tegmen plates, and position of anal tube.
One of the most distinctive characters that separate generawithin the actinocrinitids is the presence or absence of armlobes. Arm lobes are extensions of the theca (calyx and tegmen)away from the distal portion of the calyx, formed by fixed
brachials proximally and tegmen plates distally. All actinocri-nitids that have arm lobes have arms that are strongly groupedand interrays that are in contact with the tegmen. This was thecondition that Bowsher and Ubaghs (1978) used to distinguishthe subfamily Actinocrinitinae; the same genera are mostclosely related to each other as shown in the phylogenetictree (Fig. 1). Two other basic conditions exist among theactinocrinitids that lack arm trunks (i.e., where arms becomefree in a more-or-less circular pattern rather than being extendedinto arm lobes). Most actinocrinitids that lack arm lobes havearms that are either strongly or weakly grouped, allowing for theinterrays to be in contact with the tegmen. In contrast, a fewactinocrinitids have arm openings that are in continuous lateralcontact, not allowing for interrays to connect with the tegmen.Examples of these conditions are: Actinocrinites has arm lobes,Eumorphocrinus lacks arm lobes, but has a strong grouping ofarm openings, Cusacrinus lacks arm lobes and only has a slightgrouping of arm openings, and Cactocrinus lacks arm lobes andall the arm facets are in lateral contact with one another.
The actinocrinitids that have arm lobes are Aacocrinus,Abactinocrinus, Actinocrinites, Blairocrinus, Iotacrinus,Sampsonocrinus, Steganocrinus, and Thinocrinus. The arm
Figure 1. Strict consensus tree based on the phylogenetic analysis of the Actinocrinitidae from PAUP© 4.0b10; 22 taxa, 15 characters, 38 character states(Appendices 1 and 2), all characters were unordered and unweighted; search methods heuristic with random stepwise addition. There were seven mostparsimonious trees of length 114. Consistency index of 0.81; retention index of 0.67; rescaled consistency index of 0.54; homoplasy index of 0.75. Thesynapomorphies plotted at 17 nodes are as follows (see Appendix 1 for key): 1, C2, D2, E3, G0, H0, I0, K1, L0, M0, N1, O1; 2, A1, B1, F3, J0, I1, N0; 3, F2;4, C1, J1; 5, N2; 6, A0, J2; 7, D1, E2; 8, F1; 9, C1; 10, A0, O0; 11, F2; 12, E1; 13, K0; 14, B0; 15, O2; 16, L1; 17, O1. The polytomy is where the distributionof character states for characters E, F, and L cannot be resolved by parsimony analysis. Black boxes indicate known ranges, white indicates through ranges, andgrey indicates ranges that cannot be narrowed down to a time unit in the Serpukhovian. The asterisk (*) indicates an unknown range inside the Visean forSampsonocrinus. The open circle (○) is for the subfamily Actinocrinitinae of the Treatise, the open square (□) is for the subfamily Eumorphocrininae of theTreatise, the open diamond (◊) is for the subfamily Cactocrininae of the Treatise, and the closed circle (●) is for the subfamily Physetocrininae of the Treatise.
Journal of Paleontology 89(1):1–19 4
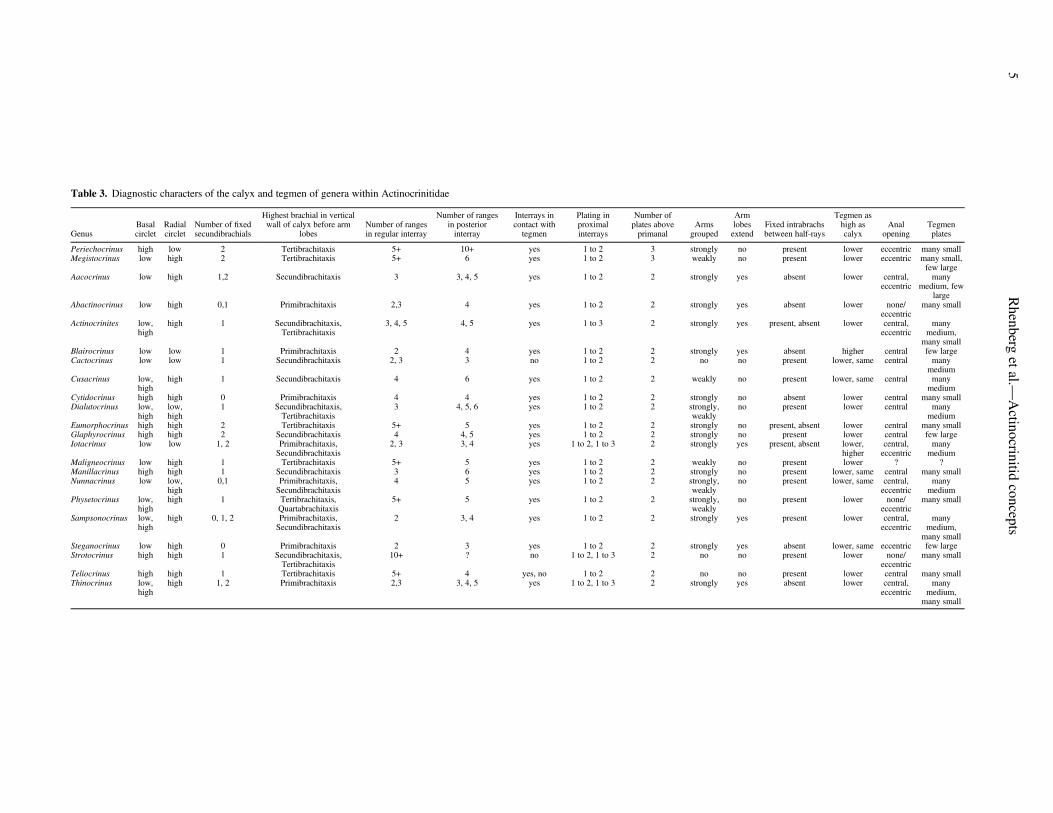
Table 3. Diagnostic characters of the calyx and tegmen of genera within Actinocrinitidae
GenusBasalcirclet
Radialcirclet
Number of fixedsecundibrachials
Highest brachial in verticalwall of calyx before arm
lobesNumber of rangesin regular interray
Number of rangesin posteriorinterray
Interrays incontact withtegmen
Plating inproximalinterrays
Number ofplates aboveprimanal
Armsgrouped
Armlobesextend
Fixed intrabrachsbetween half-rays
Tegmen ashigh ascalyx
Analopening
Tegmenplates
Periechocrinus high low 2 Tertibrachitaxis 5+ 10+ yes 1 to 2 3 strongly no present lower eccentric many smallMegistocrinus low high 2 Tertibrachitaxis 5+ 6 yes 1 to 2 3 weakly no present lower eccentric many small,
few largeAacocrinus low high 1,2 Secundibrachitaxis 3 3, 4, 5 yes 1 to 2 2 strongly yes absent lower central,
eccentricmany
medium, fewlarge
Abactinocrinus low high 0,1 Primibrachitaxis 2,3 4 yes 1 to 2 2 strongly yes absent lower none/eccentric
many small
Actinocrinites low,high
high 1 Secundibrachitaxis,Tertibrachitaxis
3, 4, 5 4, 5 yes 1 to 3 2 strongly yes present, absent lower central,eccentric
manymedium,
many smallBlairocrinus low low 1 Primibrachitaxis 2 4 yes 1 to 2 2 strongly yes absent higher central few largeCactocrinus low low 1 Secundibrachitaxis 2, 3 3 no 1 to 2 2 no no present lower, same central many
mediumCusacrinus low,
highhigh 1 Secundibrachitaxis 4 6 yes 1 to 2 2 weakly no present lower, same central many
mediumCytidocrinus high high 0 Primibrachitaxis 4 4 yes 1 to 2 2 strongly no absent lower central many smallDialutocrinus low,
highlow,high
1 Secundibrachitaxis,Tertibrachitaxis
3 4, 5, 6 yes 1 to 2 2 strongly,weakly
no present lower central manymedium
Eumorphocrinus high high 2 Tertibrachitaxis 5+ 5 yes 1 to 2 2 strongly no present, absent lower central many smallGlaphyrocrinus high high 2 Secundibrachitaxis 4 4, 5 yes 1 to 2 2 strongly no present lower central few largeIotacrinus low low 1, 2 Primibrachitaxis,
Secundibrachitaxis2, 3 3, 4 yes 1 to 2, 1 to 3 2 strongly yes present, absent lower,
highercentral,eccentric
manymedium
Maligneocrinus low high 1 Tertibrachitaxis 5+ 5 yes 1 to 2 2 weakly no present lower ? ?Manillacrinus high high 1 Secundibrachitaxis 3 6 yes 1 to 2 2 strongly no present lower, same central many smallNunnacrinus low low,
high0,1 Primibrachitaxis,
Secundibrachitaxis4 5 yes 1 to 2 2 strongly,
weaklyno present lower, same central,
eccentricmany
mediumPhysetocrinus low,
highhigh 1 Tertibrachitaxis,
Quartabrachitaxis5+ 5 yes 1 to 2 2 strongly,
weaklyno present lower none/
eccentricmany small
Sampsonocrinus low,high
high 0, 1, 2 Primibrachitaxis,Secundibrachitaxis
2 3, 4 yes 1 to 2 2 strongly yes present lower central,eccentric
manymedium,
many smallSteganocrinus low high 0 Primibrachitaxis 2 3 yes 1 to 2 2 strongly yes absent lower, same eccentric few largeStrotocrinus high high 1 Secundibrachitaxis,
Tertibrachitaxis10+ ? no 1 to 2, 1 to 3 2 no no present lower none/
eccentricmany small
Teliocrinus high high 1 Tertibrachitaxis 5+ 4 yes, no 1 to 2 2 no no present lower central many smallThinocrinus low,
highhigh 1, 2 Primibrachitaxis 2,3 3, 4, 5 yes 1 to 2, 1 to 3 2 strongly yes absent lower central,
eccentricmany
medium,many small
5Rhenberg
etal.—Actinocrinitid
concepts

lobes, along with other shared characters, keep these generaclose together in the strict consensus tree (Fig. 1, node 13).
Actinocrinites, Aacocrinus, Iotacrinus, and Thinocrinusare all very similar in appearance. To separate these genera theheight of the radial circlet, the highest brachitaxis in the verticalwall of the calyx before the arm lobes, and the size and numberof plating on the tegmen must be examined. Actinocrinites andThinocrinus both have high radial plates, but the primibrachi-taxis is the highest brachitaxis in the calyx for Thinocrinuswhereas it is either the secundibrachitaxis or tertibrachitaxis forActinocrinites. Iotacrinus has a low radial plate and Aacocrinushas a few large-sized plates on the tegmen interspersed withmany small-sized plates.
Sampsonocrinus has only two ranges in the regularinterrays but retains many medium-sized plates on the tegmenand fixed intrabrachials unlike the rest of the genera in the cladeof actinocrinitids with arm lobes. Blairocrinus has a tegmen thatis consistently higher than the calyx in all species (Iotacrinushas a single species with a higher tegmen, but this is rare in thegenus). Steganocrinus and Abactinocrinus only have ten freearms with Steganocrinus lacking fixed secundabrachials andAbactinocrinus having no anal tube.
Other genera in the same clade containing those with armlobes are Cactocrinus, Dialutocrinus, Nunnacrinus, Cusacrinus,and Manillacrinus. Cactocrinus shares several characters withIotacrinus, including having low radials, but the lack of armlobes and not having interrays connected to the tegmendifferentiates it. Cactocrinus groups within the lobate actino-crinitids despite lacking arm lobes because the ranges in theinterrays are lower than those genera without arm lobes. Inthe regular interrays Cactocrinus can have either two or threeranges, while those without arm lobes have no fewer thanthree. Similarly, in the posterior interray, Cactocrinus only hasthree ranges while the others have four or more. Dialutocrinus,which has less than four ranges in the regular interrays, haseither the secundibrachitaxis or tertibrachitaxis as the highestbrachitaxis in the vertical wall of the calyx. Nunnacrinus haseither zero or one fixed secundibrachial and the secundibrachi-taxis or lower as the highest in the vertical wall of the calyx.Nunnacrinus may have weakly grouped arms, but some specieshave strongly grouped arms whereas Cusacrinus only hasweakly grouped arms. Cusacrinus also has more ranges inthe interrays than Nunnacrinus. Manillacrinus only has threeranges in the regular interray, strongly grouped arms, and manysmall-sized plates on the tegmen.
Cytidocrinus and Glaphyrocrinus group together at thebase of the larger clade by having four ranges in all interrays; theonly actinocrinitids with this character. Both have stronglygrouped arms, butCytidocrinus has fixed intrabrachials betweenhalf rays and many small-sized tegmen plates. Glaphyrocrinuslacks intrabrachials and has few large-sized tegmen plates.
The smaller clade of actinocrinitids includesMaligneocrinus,Physetocrinus, Strotocrinus, Teleiocrinus, and Eumorphocrinus.Tegmen information forMaligneocrinus is not complete, but thecalyx shares many features with Physetocrinus. However, it isunknown whether Maligneocrinus has an anal tube or not,and the size and number of tegmen plates is not known.Physetocrinus and Strotocrinus both lack anal tubes. Strotocri-nus does not have interrays that connect to the tegmen, the arms
are not grouped at all, and there are many more ranges in theinterrays than there are in Physetocrinus. Similar to Strotocrinus,Teleiocrinus has many ranges in the interrays, arms that are notgrouped, and mostly has interrays that are not in contact with thetegmen (rarely the posterior interray will connect with the tegmen),but it differs by having a prominent anal tube. Eumorphocrinus isjudged the most plesiomorphic of the actinocrinitids, havingvarious characters seen in most of the genera, but groups withinthis clade due to the synapomorphy of tertibrachitaxis as thehighest brachitaxis in the vertical wall of the calyx, which is foundonly in this smaller clade and the outgroup.
With only seven most parsimonious trees, the phylogeny ofthe Actinocrinitidae is reasonably well resolved, although thereis a large polytomy among the genera with arm lobes. Thispolytomy indicates considerable homoplasy within the Actino-crinitidae. Indeed, if the genera with lobate arms are removed,the remaining tree exhibits less homoplasy (H.I. = 0.56 [vs.0.75]) and retains its previous topology. As shown by the nearlysimultaneous first appearances of genera in the Tournaisian(Fig. 1), there was rapid evolutionary diversification within thefamily, at the resolution available. This rapid diversification waslikely associated with predatory release following the Hangen-berg Extinction event of durophagous fishes, especially theplacoderms (Sallan et al., 2011). Evolutionary experimentationwith calyx construction may have produced similar calyxcharacter states independently. Paradoxically, the oldest occur-rences, in the Famennian, are for the highly-derived generaAbactinocrinus and Physetocrinus, rather than less-derivedgenera shown on the tree (Fig. 1). This suggests that theActinocrinitidae may have had a more extensive evolutionaryhistory in the Devonian than currently known (Lane et al., 1997,2001; Waters et al., 2003), but it only became prominent in theTournaisian when taxa became abundant after predatory release.
The two clades recognized by cladistic analysis do notjustify naming two new subfamilies. Although mapping ofsynapomorphies indicates the smaller clade is characterized byfive ranges of plates in the posterior interray (node 3), and thelarger clade by secundibrachials as the highest brachitaxis inthe vertical wall of the calyx and four ranges of plates in theregular interrays (node 7), the morphological differences aretoo subtle to recognize these clades as subfamilies within theActinocrinitidae.
The Actinocrinitidae mostly existed within the Mississip-pian, although it originated during the Famennian and ranged atleast through the Bashkirian. Geographically, the actinocrinitidsare recognized in North America, Western Europe, NorthAfrica, Russia, China, Japan, and Australia. There are alsoreports of this family (Actinocrinites and Thinocrinus) from thePermian of Indonesia and Australia, but with such a large timegap between the Bashkirian and Artinskian occurrences it isquestionable how these few managed to survive. However, asthe characters of the Permian actinocrinitids fit well within thediagnosis of the family, they are assumed to be holdovers. Morework needs to be done on the young members of the family, butthat is not within the scope of this paper.
Genus Actinocrinites Miller, 1821
Type species.—Actinocrinites triacontadactylus Miller, 1821,by subsequent designation (Wachsmuth and Springer, 1881).
Journal of Paleontology 89(1):1–19 6

Other species.—European species include: Actinocrinitesadversus (Wright, 1955a); Actinocrinites anglicus (Wright,1955a); Actinocrinites angustus (Wright, 1955a); Actinocrinitescomptus Wright, 1955a; Actinocrinites depressus Wright,1955a; Actinocrinites elongatus Wright, 1955a; Actinocriniteslaevissimus (Austin and Austin, 1843); Actinocrinites parkin-soni Wright, 1955a; Actinocrinites triplus Wright, 1955a; andActinocrinites venustus (Wright, 1955a).
North American species include Actinocrinites grandissimusVan Sant, 1965;Actinocrinites jugosus (Hall, 1859);Actinocriniteslobatus (Hall, 1859); Actinocrinites magnificus (Wachsmuth andSpringer, 1897); Actinocrinites monticuliferus (Miller and Gurley,1894);Actinocrinites spergenensis (Miller andGurley, 1896a); andActinocrinites sp. (Brower, 1969) [new combination].
Additionally, Actinocrinites timoricus (Wanner, 1924) isknown from the Permian of Timor.
Diagnosis.—Basal circlet low or high; radial circlet high; onefixed secundibrachial; secundibrachitaxis or tertibrachitaxis high-est brachitaxis in vertical wall of calyx; three, four, or five rangesin regular interray; four or five ranges in posterior interray; inter-rays in contact with tegmen; plating in proximal interrays 1 to 3;fixed intrabrachials between half-rays present or absent; armsstrongly grouped; arm lobes present and extend laterally; tegmenlower than or equal to calyx height; many medium-sized or manysmall-sized plates on tegmen; anal tube central or eccentric.
Occurrence.—Mississippian (Tournaisian) through Permian(Artinskian) of Europe, North America, and Timor. In EuropeIvorian to early Chadian (time units 3 and 4) from Ireland andUnited Kingdom; in North America late Osagean to earlyMeramecian (Visean) (time units 6 and 7) from Illinois, Indiana,Iowa, Kentucky, and Missouri; in Timor Artinskian.
Remarks.—Many species previously assigned to Actinocrinitesnow have revised generic assignments (Table 1). Brower (1969)tentatively assigned a specimen to the genus Eumorphocrinus,but the known characters of this specimen better fit the genusActinocrinites. Actinocrinites sp. has a conical calyx, the halfrays are separated by intrabrachials, and what is known of thetegmen indicates that it is a low cone. Eumorphocrinus does nothave these characters, having a medium bowl shaped calyx, nofixed intrabrachials between half-rays, and a low inverted bowlshaped tegmen.
Genus Aacocrinus Bowsher, 1955
Type species.—Aacocrinus nododorsatus Bowsher, 1955, byoriginal designation.
Other species.—North American species include Aacocrinusboonensis (Peck and Keyte, 1938); Aacocrinus chouteauensis(Miller, 1892); Aacocrinus protuberoarmatus Brower, 1967;Aacocrinus sampsoni (Miller and Gurley, 1895); Aacocrinussenectus (Miller and Gurley, 1897); Aacocrinus spinosulus (Millerand Gurley, 1893); and Aacocrinus tetradactylus Brower, 1967.
Diagnosis.—Basal circlet low; radial circlet high; one or two fixedsecundibrachials; secundibrachitaxis highest brachitaxis in verticalwall of calyx; three ranges in regular interray; three, four, or five
ranges in posterior interray; interrays in contact with tegmen; plat-ing in proximal interrays 1 to 2; fixed intrabrachials between half-rays absent; arms strongly grouped; arm lobes present and extendlaterally; tegmen lower than calyx; few large-sized and manymedium-sized plates on tegmen; anal tube central or eccentric.
Occurrence.—Mississippian (Tournaisian) of North America.Early Kinderhookian throughmiddle Osagean (time units 1 to 4) ofMissouri, Nevada, and New Mexico.
Remarks.—Aacocrinus is known only from North America.Brower (1967) thoroughly described this genus, which will notbe repeated here.
Genus Abactinocrinus Laudon and Severson, 1953
Type species.—Abactinocrinus rossei Laudon and Severson,1953, by original designation.
Other species.—Abactinocrinus devonicus Waters et al., 2003and Abactinocrinus burlingtonensis (Brower, 1965) [newcombination].
Diagnosis.—Basal circlet low; radial circlet high; zero or onefixed secundibrachials; primibrachitaxis highest brachitaxis invertical wall of calyx; two or three ranges in regular interray;four ranges in the posterior interray; interrays in contact withtegmen; plating in proximal interrays 1 to 2; fixed intrabrachialsbetween half-rays absent; arms strongly grouped; arm lobespresent and extend laterally; tegmen lower than calyx; manysmall-sized plates on tegmen; anal tube absent, opening central.
Occurrence.—Devonian (Famennian) through Mississippian(Tournaisian) of China and North America. Famennian fromChina; late Kinderhookian (time unit 2) from Montana; middleOsagean (time unit 4) from Iowa.
Remarks.—Abactinocrinus is the only actinocrinitid with armlobes that lacks an anal tube, but has previously been describedas having an anal tube. However, when available specimenswere closely examined, there was no evidence of an anal tube.Abactinocrinus rossei has a small swelling at the anal openingwhile A. devonicus and A. burlingtonensis have an anal openingflush with the tegmen.
Genus Blairocrinus Miller, 1891
Type species.—Blairocrinus trijugis Miller, 1891, by originaldesignation.
Other species.—North American species include Blairocrinusarrosus Miller, 1892; Blairocrinus macurdai Rhenberg andKammer, 2013; Blairocrinus protuberus Lee et al., 2005; andBlairocrinus smalleyi (Weller, 1909) [new combination].
Diagnosis.—Basal circlet low; radial circlet low; one fixedsecundibrachial; primibrachitaxis highest brachitaxis in verticalwall of calyx; two ranges in regular interray; four ranges inposterior interray; interrays in contact with tegmen; plating in
7 Rhenberg et al.—Actinocrinitid concepts

proximal interrays 1 to 2; fixed intrabrachials between half-raysabsent; arms strongly grouped; arm lobes present and extendlaterally; tegmen higher than calyx; few large-sized tegmenplates; anal tube central.
Occurrence.—Mississippian (Tournaisian) of North America.Early Kinderhookian throughmiddle Osagean (time units 1 to 4) ofKentucky, Montana, and New Mexico.
Remarks.—Blairocrinus is the only actinocrinitid that has ategmen that is consistently higher than the calyx, which makes iteasy to distinguish from other actinocrinitids.
Physetocrinus smalleyi is reassigned to Blairocrinus basedon the characters that link it to the latter genus. These charactersare the tegmen being higher than the calyx and the arms that arestrongly grouped and occur in short arm lobes. In Weller’s(1909) description, he stated that there was no anal tube, but hisillustrations indicate that the beginning of one located centrallyon the tegmen and broken off.
Blairocrinus arrosus is a species that is subject to muchconfusion, which arises from S. A. Miller assigning twoactinocrinitids with the species name “arrosus”: Blairocrinusarrosus Miller, 1892 and Actinocrinites arrosus Miller andGurley, 1893. Without presenting a complete history of thetaxonomic confusion, Bassler and Moodey (1943) assigned the1892 species to Actinocrinites, and they left the 1893 species inActinocrinites with a note stating that it probably belongedin Cactocrinus. Webster (2003) considered both species to be inCactocrinus (with B. arrosus also considered in the genusAacocrinus), and presumably, synonymous homonyms. How-ever, B. arrosus was correctly assigned to Blairocrinus byMiller (1892), a view endorsed by Brower (1967) and Lee et al.(2005). Actinocrinus arrosus Miller and Gurley, 1893 wascorrectly assigned to Cactocrinus.
Genus Cactocrinus Wachsmuth and Springer, 1897
Type species.—Actinocrinus proboscidialis Hall, 1858, by ori-ginal designation.
Other species.—Cactocrinus arrosus (Miller and Gurley,1893); Cactocrinus bacatusWood, 1914; Cactocrinus bischoffi(Miller and Gurley, 1896c) [new combination]; Cactocrinusclarus (Hall, 1861a); Cactocrinus extensus Wachsmuth andSpringer, 1897; Cactocrinus fossatus (Miller, 1892); Cactocrinusglans (Hall, 1859); Cactocrinus hurdianus (McChesney, 1861)[new combination]; Cactocrinus imperator (Laudon, 1933)[new combination]; Cactocrinus lucina (Hall, 1861b); Cacto-crinus magnidactylus Laudon and Severson, 1953; Cactocrinusmultibrachiatus (Hall, 1858); Cactocrinus obesus (Keyes,1894); Cactocrinus opusculus (Hall, 1859); Cactocrinus platy-brachiatus (Wood, 1914); Cactocrinus sexarmatus (Hall,1859); Cactocrinus springeri (Rowley, 1900); and Cactocrinusthalia (Hall, 1861b).
Diagnosis.—Basal circlet low; radial circlet low; one fixedsecundibrachial; secundibrachitaxis highest brachitaxis in ver-tical wall of calyx; two or three ranges in regular interray; threeranges in posterior interray; interrays not in contact with
tegmen; plating in proximal interrays 1 to 2; fixed intrabrachialsbetween half-rays present; arms not grouped; arm lobes absent;tegmen lower than or same height as calyx; many medium-sizedplates on tegmen; anal tube central.
Occurrence.—Mississippian (Tournaisian) of North America.Early Kinderhookian through middle Osagean (time units 1 to 4)from Illinois, Iowa, Missouri, New Mexico, and Utah.
Remarks.—Cusacrinus bischoffi, C. hurdianus, and C. imperatorare reassigned to Cactocrinus because the arms are not groupedand the arm facets are connected to each other, not allowing theinterrays to be in contact with the tegmen.
Genus Cusacrinus Bowsher, 1955
Type species.—Actinocrinites nodobrachiatus Wachsmuth andSpringer in Miller, 1889, by original designation.
Other species.—Cusacrinus arnoldi (Wachsmuth and Springerin Miller, 1889); Cusacrinus asperrimus (Meek and Worthen,1870); Cusacrinus chloris (Hall, 1861a); Cusacrinus coelatus(Hall, 1858); Cusacrinus daphne (Hall, 1863); Cusacrinusdenticulatus (Wachsmuth and Springer, 1897); Cusacrinusectypus (Meek and Worthen, 1870); Cusacrinus kuenzii (Laudonet al., 1952);Cusacrinus limabrachiatus (Hall, 1861a);Cusacrinuslongus (Meek and Worthen, 1870); Cusacrinus ornatissmus(Wachsmuth and Springer in Miller, 1889); Cusacrinus penicillus(Meek and Worthen, 1870); Cusacrinus sampsoni (Miller andGurley, 1896c) [new combination]; Cusacrinus sobrinus(Miller and Gurley, 1896c); Cusacrinus spectabilis (Miller andGurley, 1896c) [new combination]; Cusacrinus spinotentaculus(Hall, 1859); Cusacrinus subscitulus (Miller and Gurley, 1896c)[new combination]; Cusacrinus tenuisculptus (McChesney,1861); Cusacrinus thetis (Hall, 1861b); Cusacrinus tuberculosus(Wachsmuth and Springer, 1897); Cusacrinus viaticus (White,1874).
Diagnosis.—Basal circlet high or low; radial circlet high; onefixed secundibrachial; secundibrachitaxis highest brachitaxis invertical wall of calyx; four ranges in regular interray; six rangesin posterior interray; interrays in contact with tegmen; plating inproximal interrays 1 to 2; fixed intrabrachials between half-rayspresent; arms weakly grouped; arm lobes absent; tegmen lowerthan or same height as calyx; many medium-sized plates ontegmen; anal tube central.
Occurrence.—Mississippian (Tournaisian) of North America.Early Kinderhookian through middle Osagean (time units 1 to 4)of Iowa; Missouri; Montana; New Mexico; Nevada; Utah; andAlberta, Canada.
Remarks.—Actinocrinites spectabilis, and A. subscitulus arereassigned to Cusacrinus due to the lack of arm lobes, arms thatare weakly grouped, appropriate number of ranges in the regularinterrays, and the tegmen plate sculpture. It is possible thatthese species are variations on one species that also includesC. sobrinus. They all occur in the Burlington Limestone, and thedifferences between them appear to be minor. Miller and Gurley
Journal of Paleontology 89(1):1–19 8

(1896c) even noted that they frequently found differences in thenumber of interradial plates among specimens of the samespecies, which was one of the characters they used to separate C.spectabilis [new comb.] and C. sobrinus. The other determiningcharacter between the two species is that the regular interrays arenot in contact with the tegmen.
Cusacrinus arnoldi andC. ornatissimuswere synonymizedwith C. nodobrachiatus in Webster’s Index (2003); however,there is no explanation as to why these three species should beconsidered the same. All three species are distinguishable fromeach other and should remain separate. Cusacrinus nodobra-chiatus has steep-sided, medium-cone shaped calyx, low basals,and spinose plates on the tegmen, whereas C. arnoldi has amedium-cone shaped calyx, the sides are not so steep, the basalsare higher, and the tegmen plates are smooth. Cusacrinusornatissimus has a low-cone shaped calyx, making it differentfrom the previous two and low basals; the tegmen is unknown.
Nunnacrinus sampsoni has been placed in Cusacrinusbecause the of the high basal circlet and the highest brachitax isin the calyx is the secundibrachitaxis. The highest brachitaxis inNunnacrinus is usuallythe primibrachitaxis and only rarely thesecundibrachitaxis. Nunnacrinus has a low basal circlet.
Genus Cytidocrinus Kirk, 1944
Type species.—Actinocrinus sculptus Hall, 1858, by originaldesignation.
Diagnosis.—Basal circlet high; radial circlet high; zero fixedsecundibrachials; primibrachitaxis highest brachitaxis in ver-tical wall of calyx; four ranges in regular interray; four ranges inposterior interray; interrays in contact with tegmen; plating inproximal interrays 1 to 2; fixed intrabrachials between half-raysabsent; arms strongly grouped; arm lobes absent; tegmen lowerthan calyx; many small-sized plates on tegmen; anal tube central.
Occurrence.—Mississippian (Tournaisian) of North America.Early to middle Osagean (time units 3 to 4) of Iowa.
Remarks.—The genus Cytidocrinuswas fully described by Kirk(1943, p. 263).
Genus Dialutocrinus Wright, 1955a
Type species.—Dialutocrinus milleri Wright, 1955a.
Other species.—European species include Dialutocrinus acu-leatus (Austin and Austin, 1843); Dialutocrinus austini Ausichand Sevastopulo, 2001;Dialutocrinus cataphractus (Austin andAustin, 1843); Dialutocrinus icosidactylus (Portlock, 1843);Dialutocrinus mcoyi Ausich and Sevastopulo, 2001; Dialuto-crinus polydactylus (Miller, 1821); Dialutocrinus tessellatus(Phillips, 1836); and Dialutocrinus tricuspidatus (de Koninckand Le Hon, 1854) [new combination].
North American species include Dialutocrinus gracilis(Wachsmuth and Springer, 1897) [new combination].
Diagnosis.—Basal circlet high or low; radial circlet high or low;one fixed secundibrachial; secundibrachitaxis or tertibrachitaxishighest brachitaxis in vertical wall of calyx; three ranges in
regular interrays; four, five, or six ranges in posterior interray;interrays in contact with tegmen; plating in proximal interrays1 to 2; fixed intrabrachials between half-rays present; armsstrongly or weakly grouped; arm lobes absent; tegmen lower thancalyx; many medium-sized plates on tegmen; anal tube central.
Occurrence.—Mississippian (Tournaisian) of Europe and NorthAmerica. Ivorian and lower Chadian (time units 3–4) of Belgium,Ireland, England, and Wales; early Osagean (time unit 3) of Iowa.
Remarks.—Cusacrinus gracilis is reassigned to Dialutocrinusbecause the calyx shape is a bowl and not a cone. Though theshape of the calyx is not a diagnostic character for the whole ofthe family, there are some genera that only have a particularshape for the calyx. Cusacrinus has a conical calyx while Dia-lutocrinus only has a bowl-shaped calyx. There are also onlythree ranges of plates in the regular interray, not four as inCusacrinus. The reassignment of D. gracilis extends the rangeof Dialutocrinus to North America.
Actinocrinites tricuspidatus is also reassigned to Dialuto-crinus. As with Dialutocrinus gracilis, it is the medium bowl-shaped calyx that makes it more closely related toDialutocrinusbecause Actinocrinites is typically more conical; where it isbowl shaped, the bowl is very high.
Genus Eumorphocrinus Wright, 1955a
Type species.—Eumorphocrinus erectus Wright, 1955a.
Other species.—Eumorphocrinus elongatus Lindley, 1979;Eumorphocrinus excelsus Wright, 1955a; and Eumorphocrinushibernicus Wright, 1955a.
Diagnosis.—Basal circlet high; radial circlet high; two fixedsecundibrachials; tertibrachitaxis highest brachitaxis in verticalwall of calyx; five or more ranges in regular interrays; five rangesin posterior interray; interrays in contact with tegmen; plating inproximal interrays 1 to 2; fixed intrabrachials between half-rayspresent or absent; arms strongly grouped; arm lobes absent; teg-men lower than calyx; many small-sized plates on tegmen; analtube central.
Occurrence.—Mississippian (Tournasian to Visean) of Europeand Australia. Late Tournaisian to Visean (time units 4 to 5) ofEngland and Ireland; Late Tournaisian (time unit 4) from NewSouth Wales.
Remarks.—The genus is described by Wright (1955a, p. 232).
Genus Glaphyrocrinus Lindley, 1988
Type species.—Glaphyrocrinus expansus Lindley, 1988.
Other species.—Glaphyrocrinus minutus Lindley, 1988.
Diagnosis.—Basal circlet high; radial circlet high; two fixedsecundibrachials; secundibrachitaxis highest brachitaxis in verticalwall of calyx; four ranges in regular interray; four or five ranges inposterior interray; interrays in contact with tegmen; plating inproximal interrays 1 to 2; fixed intrabrachials between half-rays
9 Rhenberg et al.—Actinocrinitid concepts

present; arms strongly grouped; arm lobes absent; tegmen lowerthan calyx; few large-sized plates on tegmen; anal tube central.
Occurrence.—Mississippian (late Tournaisian, time unit 4) ofAustralia.
Genus Iotacrinus Ausich and Sevastopulo, 2001
Type species.—Actinocrinus dorsatus (de Koninck and Le Hon,1854), by original designation.
Other species.—European species include: Iotacrinus armatus(de Koninck and Le Hon, 1894) [new combination]; Iotacrinusintermedius (Wright, 1955a) [new combination]; Iotacrinusmoderatus (Wright, 1955a); and Iotacrinus stellaris (deKoninck and Le Hon, 1894) [new combination]. North Amer-ican species include Iotacrinus novamexicanus Rhenberg andKammer, 2013.
Diagnosis.—Basal circlet low; radial circlet low; one or twofixed secundibrachials; primabrachitaxis or secundabrachitaxishighest brachitaxis in vertical wall of calyx; two or three rangesin regular interrays; three or four ranges in posterior interray;interrays in contact with tegmen; plating in proximal interrays1 to 2 or 1 to 3; fixed intrabrachials between half-rays present orabsent; arms strongly grouped; arm lobes present and extendlaterally; tegmen lower or higher than calyx; many medium-sized sized tegmen plates; anal tube central or eccentric.
Occurrence.—Mississippian (Tournaisian) of Europe and NorthAmerica. Ivorian to early Chadian (time units 3 and 4) fromBelgium, Ireland, and England; early Osagean (time unit 3)from New Mexico.
Remarks.—Until a recent review of the Lake Valley Formationcrinoids (Rhenberg and Kammer, 2013), Iotacrinus was onlyknown in the “Mountain Limestone” of Belgium and in theHook Head Formation of Ireland (Ausich and Sevastopulo,2001). The North American species, Iotacrinus novamexicanus,is similar to the European species but has a tegmen that is muchhigher and is more distinctly lobed.
Actinocrinites rotundatus has been moved to Iotacrinusbecause it has a low radial circlet and two fixed secundibrachialsin the calyx wall. Two species from Nunnacrinus have beenmoved to this genus. These are N. armatus and N. stellaris.These species have low radials which places them in Iotacrinusand not Thinocrinus and fewer ranges in the interrays thanNunnacrinus species have.
Genus Maligneocrinus Laudon, Parks, and Spreng, 1952
Type species.—Maligneocrinus medicinensis Laudon, Parks,and Spreng, 1952.
Diagnosis.—Basal circlet low; radial circlet high; one fixedsecundibrachial; tertibrachitaxis highest brachitaxis in verticalwall of calyx; five or more ranges in regular interray; five rangesin posterior interray; interrays in contact with tegmen; plating inproximal interrays 1 to 2; fixed intrabrachials between half-ray
present; arms weakly grouped; arm lobes absent; tegmen lowerthan calyx; tegmen plate size unknown; anal tube unknown.
Occurrence.—Mississippian (Tournaisian) of North America.Early Osagean (time unit 3) of Alberta, Canada.
Remarks.—Although this genus is known only frompartial crowns,the calyx shows diagnostic characteristics of actinocrinitids, con-firming its placement in the family.Maligneocrinus does not fit intoother actinocrinitid genera due to its globe-shaped calyx.With littleknown about the tegmen, Maligneocrinus groups mostly closelywith Physetocrinus, although additional specimens for study maychange the interpretation of its relationships within the family.
Genus Manillacrinus Campbell and Bein, 1971
Type species.—Cactocrinus? brownei Dun and Benson, 1920,by original desgination.
Other species.—Manillacrinus acanthusWebster and Jell, 1999.
Diagnosis.—Basal circlet high; radial circlet high; one fixedsecundibrachial; secundibrachitaxis highest brachitaxis in ver-tical wall of calyx; three ranges in regular interray; six ranges inposterior interray; interrays in contact with tegmen; plating inproximal interrays 1 to 2; fixed intrabrachials between half-rayspresent; arms strongly grouped; arm lobes absent; tegmen lowerthan or same height as calyx; many small-sized plates on teg-men; anal tube central.
Occurrence.—Mississippian (Tournaisian) of Australia. LateTournaisian (time units 3 or 4) of New South Wales.
Remarks.—It should be noted that Bowsher (in Moore andTeichert, 1978, p. T458) erroneously referred to this genus asManillocrinus.
Genus Nunnacrinus Bowsher, 1955
Type species.—Nunnacrinus mammillatus Bowsher, 1955, byoriginal designation.
Other species.—North American species include Nunnacrinusdalyanus (Miller, 1881); Nunnacrinus erraticus (Miller andGurley, 1893) [new combination]; Nunnacrinus foveatus(Miller and Gurley, 1895); Nunnacrinus locellus (Hall, 1861b);Nunnacrinus olsoni Ausich, 2003; Nunnacrinus ovatus (Hall,1861b) [new combination]; Nunnacrinus pallubrum (Miller andGurley, 1896b); Nunnacrinus pettisensis (Miller and Gurley,1896c); Nunnacrinus plagosus (Miller and Gurley, 1893) [newcombination]; Nunnacrinus puteatus (Rowley and Hare, 1891);Nunnacrinus reticulatus (Hall, 1861a) [new combination]; andNunnacrinus rubra (Weller, 1909).
One North African species is included in Nunnacrinus,N. becharensis (Webster, et al., 2004) [new combination] fromAlgeria.
Diagnosis.—Basal circlet low; radial circlet low or high; zero orone fixed secundibrachial; primibrachitaxis or secundabrachi-taxis highest brachitaxis in vertical wall of calyx; four ranges in
Journal of Paleontology 89(1):1–19 10

regular interray; five ranges in posterior interray; interrays incontact with tegmen; plating in proximal interrays 1 to 2; fixedintrabrachials between half-rays present; arms strongly orweakly grouped; arm lobes absent; tegmen lower than or sameheight as calyx; many medium-sized plates on tegmen; anal tubecentral or eccentric.
Occurrence.—Mississippian (Tournaisian to Serpukhovian)of North America and Africa. Late Kinderhookian to earlyOsagean (time units 2 and 3) of Iowa, Missouri, and NewMexico; Serpukhovian (time unit 10) of Algeria.
Remarks.—When Bowsher (1955) named the genus Nunnacri-nus, he designatedN.mammillatus as the type species. This speciesis known only from the Nunn Member of the Lake Valley For-mation. However, it is a junior synonym of a previously namedLake Valley Formation crinoid, N. dalyanus (Miller, 1881).Bowsher (1955, p. 20) stated that the two species are distinguish-able from each other most readily by the number of plates thatseparate the rays: two in N. mammillatus and one in N. dalyanus.This distinction is not valid as there is variation in plate numberbetween rays in more than 700 Nunnacrinus specimens studiedfrom the Lake Valley, including the types for N. mammillatus (seeRhenberg and Kammer, 2013). Most of the specimens have bothsingle and double plates between the rays and not just one or theother. The other characters that Bowsher used to separate N.mammillatus and N. dalyanus also have great variation among thespecimens studied. These includeN.mammillatus being smaller insize, having a less conical calyx and tegmen, weaker plate structureon the calyx, and great irregularity of size of tegmen plates. All ofthese features have been found to have the range of variabilityjudged to be within a single species of crinoid. Because of this, N.mammillatus is a junior synonym of N. dalyanus.
Actinocrinus erraticus and A. plagosus are reassigned toNunnacrinus because the arms are weakly grouped, do not formlobes, and have the appropriate number of ranges in theinterrays. Although the tegmen is not known for N. plagosus,the calyx characters best fit Nunnacrinus.
Cactocrinus reticulatus and C. reticulatus ovatus arereassigned to Nunnacrinus because the interrays connect withthe tegmen, which does not occur in Cactocrinus. Furthermore,N. ovatus is regarded as a separate species and is not a variationof N. reticulatus, as contended by Wachsmuth and Springer(1897). Both species have a medium cone-shaped calyx, butN. ovatus has a more rounded overall shapeand a nearly bowl-shaped calyx. Nunnacrinus reticulatus has a medium cone-shaped tegmen that is about the same height as the calyx withlong spines on most of the tegmen plates, including the platesthat form the base of the anal tube. Nunnacrinus ovatus has a lowbowl-shaped tegmen with short spines on a few of the tegmenplates; the spines do not appear to continue on the anal tube.
Nunnacrinus jessieae is a problematic species of Nunna-crinus. In 1896, Miller and Gurley described several newcamerate crinoids, and two were given the trivial name“jessieae”: Actinocrinites jessieae Miller and Gurley, 1896aand Amphoracrinus jessieaeMiller and Gurley, 1896c. Webster(2003) synonymized both of these species as N. jessieae,although they are two distinct species. Bowsher (1955) correctlyassigned Actinocrinites jessieae (Miller and Gurley, 1896a) to
Nunnacrinus and Ausich and Kammer (2008) consideredAmphoracrinus jessieae (Miller and Gurley, 1896c) to be anomen dubium.
Reassignment, herein, of Actinocrinites becharensis Web-ster et al., 2004 to Nunnacrinus extends both the geographic andstratigraphic range of the latter genus. Nunnacrinus becharensisis of early Serpukovian age of western Algeria. This Algerianspecies differs from Actinocrinites s.s. based on the Europeantype species (Ausich and Sevastopulo, 2001; Rhenberg andKammer, 2013) because of the presence of a low basal circlet,two plates above the first interradial in regular interrays, and ategmen approximately as high as the calyx.
Genus Physetocrinus Meek and Worthen, 1869
Type species.—Actinocrinus ventricosusHall, 1858, by subsequentdesignation (Wachsmuth and Springer, 1881).
Other species.—North American species include Physetocrinusasper (Meek and Worthen, 1870); Physetocrinus copei (Miller,1881); Physetocrinus dilatatus (Meek and Worthen, 1870);Physetocrinus lobatus Wachsmuth and Springer, 1897; Physeto-crinus majusculus Webster and Lane, 1987; and Physetocrinusornatus (Hall 1858).
Non-North American species include Physetocrinus batheri(Whidbourne, 1896) [new combination] and Physetocrinusbrightoni Wright, 1955a.
Diagnosis.—Basal circlet low or high; radial circlet high; onefixed secundibrachial; tertibrachitaxis or quartibrachitaxishighest brachitaxis in vertical wall of calyx; five or more rangesin regular interray; five ranges in posterior interray; interrays incontact with tegmen; plating in proximal interrays 1 to 2; fixedintrabrachials between half-rays present; arms strongly to weaklygrouped; arm lobes absent; tegmen lower than calyx; many small-sized plates on tegmen; anal tube absent, opening eccentric.
Occurrence.—Devonian (Famennian) through Mississippian(Tournaisian) of North America and Europe. Famennian of Eng-land; Ivorian (time unit 3) of Ireland; early to middle Osagean (timeunits 3–4) of Arizona, Illinois, Iowa, New Mexico, and Nevada.
Remarks.—Physetocrinus is primarily known from NorthAmerica, although P. brightoni occurs in the Hook HeadFormation of Ireland and P. batheri occurs in the MarwoodBeds of England.
Physetocrinus majusculus is tentatively maintained as aseparate species from P. copei. The main difference cited byWebster and Lane (1987) between the two species is thatP. majusculus has a more highly inflated tegmen than P. copei.A study of P. copei from the Lake Valley Formation indicatesthat there is a wide variation of tegmen inflation in thatspecies (fig. 9 in Rhenberg and Kammer, 2013). Otherwise, thedifferences between the two species are few. They includeP. majusculus having a more conical calyx, a weak overhang atthe base of the arms, fewer plates separating the ambulacraltracts between rays and half-rays, and more uniform tegmenplates. However, until a more detailed study of both species isundertaken, they are retained as separate species.
11 Rhenberg et al.—Actinocrinitid concepts

With the recognition of Physetocrinus batheri (Whidborne,1896) herein, the stratigraphic range of the genus is extendedinto the Famennian. Physetocrinus batheri is a relatively poorlypreserved specimen. However, it belongs to the Actinocriniti-dae, and the morphology of the tegmen (Whidborne, 1898, Pl.32, fig. 2a; Lane et al., 2001, Pl. 3, fig. 2) aligns the species withPhysetocrinus.
Genus Sampsonocrinus Miller and Gurley, 1895
Type species.—Sampsonocrinus hemisphericus Miller andGurley, 1895
Other species.—North American species include Sampsono-crinus? globosus (Wachsmuth and Springer, 1897); and Samp-sonocrinus sedaliensis (Miller and Gurley, 1895) [newcombination].
Non-North American species include Sampsonocrinuscannindahensis Webster and Jell, 1999 and Sampsonocrinuscoplowensis (Wright, 1955a) [new combination].
Diagnosis.—Basal circlet high or low; radial circlet high; zero,one, or two fixed secundibrachials; primibrachitaxis or secun-dibrachitaxis highest brachitaxis in vertical wall of calyx; tworanges in regular interray; three or four ranges in posteriorinterray; interrays in contact with tegmen; plating in proximalinterrays 1 to 2; fixed intrabrachials between half-rays present;arms strongly grouped; arm lobes present and extend laterally;tegmen lower than calyx; many small-sized or many medium-sized plates on tegmen; anal tube central or eccentric.
Occurrence.—Mississippian (Tournaisian to Visean?) of Aus-tralia, Europe, and North America. Early Chadian (time unit 4)from United Kingdom; early Kinderhookian (time unit 1) fromMissouri; Visean? from Australia.
Remarks.—Actinocrinus sedaliensis and A. coplowensis arereassigned to the genus Sampsonocrinus because their char-acters fit the latter genus better. Actinocrinus sedaliensis has alow basal circlet, two ranges in the regular interrays, and aneccentric anal tube and A. coplowensis has two fixed secundi-brachials, two ranges of plates in the regular interrays, andproximal plating of 1 to 2.
The species Sampsonocrinus? globosus is a problematicspecies that does not conform to any described crinoid genus.Brower (1965, pg. 789) discussed this issue, suggesting itmay be a new genus of actinocrinitid. Because the species isknown from a single specimen, he hesitated to name a newgenus and concluded that it is most similar to Sampsonocrinusand questionably placed it within this genus. With no newinformation to add, we follow Brower’s suggestion and regardthis species as S.? globosus.
Genus Steganocrinus Meek and Worthen, 1866a
Type species.—Actinocrinites pentagonus Hall, 1858, by originaldesignation.
Other species.—Steganocrinus altus Brower, 1965; Steganocri-nus concinnus (Shumard, 1855); Steganocrinus elongatus Kirk,
1943; Steganocrinus longus Brower, 1965; Steganocrinusmultistriatus Brower, 1965; Steganocrinus planus Brower,1965; Steganocrinus robustus Brower, 1965; and Steganocrinusvalidus (Meek and Worthen, 1860).
Diagnosis.—Basal circlet low; radial circlet high; zero fixedsecundibrachials; primibrachitaxis highest brachitaxis in verticalwall of calyx; two ranges in regular interray; three ranges in pos-terior interray; interrays in contact with tegmen; plating in proximalinterrays 1 to 2; fixed interbrachials between half-rays absent; armsstrongly grouped; arm lobes present and extend laterally; tegmenlower than or same height as calyx; few large-sized plates ontegmen; anal tube eccentric.
Occurrence.—Mississippian (Tournaisian) of North America.Early to middle Osagean (time units 3 and 4) from Arizona,Iowa, Missouri, Nevada, and New Mexico.
Remarks.—The species Steganocrinus altus and S. longus maybe junior synonyms of S. pentagonus. The descriptions byBrower (1965) indicated that the tegmen features of all threespecies are the same, and that it is only minor differences inthe calyx that separates them. The differences between S. altusand S. pentagonus are that the former has a more conicaldorsal cup; however, when comparing S. pentagonus (plate 92,figure 29) and S. altus (plate 93, figure 7), it is difficult to tellthe two apart. Steganocrinus longus differs from S. pentagonusby having low basals, highly tumid calyx plates, and veryhigh ambulacral tracts. Geographically, S. altus and S. longusoccur only in the Nunn Member of the Lake Valley Formation,is consistent with them being morphological variants ofS. pentagonus. Because the review of the individual species isbeyond the scope of this paper, they are retained as separatespecies.
Steganocrinus planus Brower, 1965 is least like the otherspecies in the genus in that all other species of Steganocrinushave few, large plates on the tegmen, but S. planus is coveredwith numerous smaller plates. Brower (1965) noted that thetegmen is very similar to Abactinocrinus, but all features of thecalyx compare more closely with Steganocrinus. Therefore, wefollow Brower and retain S. planus within this genus.
Genus Strotocrinus Meek and Worthen, 1866b
Type species.—Actinocrinites perumbrosus Hall, 1860, bysubsequent designation (Meek and Worthen, 1866a).
Diagnosis.—Basal circlet high; radial circlet high; one fixedsecundibrachial; secundibrachitaxis or tertibrachitaxis highestbrachitaxis in vertical wall of calyx; 10 or more ranges in regularinterrays; number of ranges in posterior interray unknown;interrays not in contact with tegmen; plating in proximal inter-rays 1 to 2 or 1 to 3; fixed intrabrachials between half-rayspresent; arms not grouped; arm lobes absent; tegmen lower thancalyx; many small-sized plates on tegmen; anal tube absent,opening eccentric.
Occurrence.—Mississippian (Tournaisian) of North America.Middle Osagean (time unit 4) from Iowa and Missouri.
Journal of Paleontology 89(1):1–19 12

Discussion.—Strotocrinus glyptus (Hall, 1860), is the onlyrecognized species in this genus. Macurda (1974) synonymizedall reported members of the genus into S. glyptus, includingS. perumbrosus; stating that S. glyptus is the valid name due topage priority. Ausich and Kammer (1991b) also discussednomenclatural history of S. glyptus; and therefore, it will not berepeated here.
Genus Teleiocrinus Wachsmuth and Springer, 1881
Type species.—Actinocrinites umbrosus Hall, 1858, by originaldesignation.
Other species.—Teleiocrinus adolescensWachsmuth and Springer,1897; Teleiocrinus blairi (Miller and Gurley, 1895) [new combina-tion]; Teleiocrinus liratus (Hall, 1859); Teleiocrinus ornatus(Miller and Gurley, 1896a) [new combination]; and Teleiocrinusvenustus (Miller and Gurley, 1893) [new combination].
Diagnosis.—Basal circlet high; radial circlet high; one fixedsecundibrachial; tertibrachitaxis highest brachitaxis in verticalwall of calyx; five or more ranges in regular interray; four rangesin posterior interray; interrays may or may not be in contact withtegmen; plating in proximal interrays 1 to 2; fixed intrabrachialsbetween half-rays present; arms not grouped; arm lobes absent;tegmen lower than calyx; many small-sized plates on tegmen;anal tube central.
Occurrence.—Mississippian (Tournaisian) of North America.Early to middle Osagean (time units 3 and 4) of Iowa, Missouri,Illinois, and New Mexico.
Remarks.—Strotocrinus blairi and S. ornatus have been reas-signed to Teleiocrinus because they have an anal tube, whichis lacking in Strotocrinus. Strotocrinus venustus isreassigned to Teleiocrinus based on characters of the calyx (thetegmen is unknown; and therefore, it is unknown whether ananal tube exists). The calyx of T. venustus lacks distal flaringand has fewer ranges in the interray than Strotocrinus. Macurda(1974) placed this species with Cusacrinus ectypus (Meek andWorthen, 1870), but the original description (Miller and Gurley,1893, p. 27) and illustration (pl. 6, fig. 1) indicate that thistaxon has at least 60 arm facets that separate the calyx fromthe tegmen, not allowing for the interrays to be in contactwith the tegmen. This feature distinguishes this species fromCusacrinus, which has fewer arms and interrays that connectwith the tegmen.
Genus Thinocrinus Ausich and Sevastopulo, 2001
Type species.—Sampsonocrinus westheadi (Wright, 1947), byoriginal designation.
Other species.—European species include Thinocrinus alatus(Wright, 1955a) [new combination]; Thinocrinus deornatus (deKoninck and Le Hon, 1854) [new combination]; Thinocrinusgranulatus (Wright, 1955b) [new combination]; Thinocrinusloricatus (Schlotheim, 1820); Thinocrinus nodosus (Wright,1955a) [new combination]; Thinocrinus rotundus (Wright,
1955a) [new combination]; Thinocrinus smythi Ausich andSevastopulo, 2001; Thinocrinus vermiculatus (Wright, 1955a)[new combination]; and Thinocrinus wexfordensis Ausich andSevastopulo.
North American species include Thinocrinus anchorensis(Webster and Lane, 1987) [new combination]; Thinocrinusbenedicti (Miller, 1892) [new combination]; Thinocrinus blairi(Miller and Gurley, 1897) [new combination]; Thinocrinusbotruosus (Miller and Gurley, 1895) [new combination];Thinocrinus enigmatus (Webster and Lane, 1987) [newcombination]; Thinocrinus gibsoni (Miller and Gurley, 1893)[new combination]; Thinocrinus lowei (Hall, 1858) [newcombination]; Thinocrinus multiradiatus (Shumard, 1857)[new combination]; Thinocrinus multiramosus altidorsatus (Row-ley, 1904) [new combination]; Thinocrinus pernodosus (Hall, 1858)[new combination]; Thinocrinus probolos (Ausich and Kammer,1991a) [new combination]; Thinocrinus? sampsoni (Miller andGurley, 1896c) [new combination]; Thinocrinus scitulus (Miller andGurley, 1897) [new combination]; Thinocrinus semimultiramosus(Whitfield, 1900) [new combination]; Thinocrinus subpulchellus(Miller and Gurley, 1896c) [new combination]; Thinocrinus tripus(Ehlers and Kesling, 1963) [new combination]; and Thinocrinusverrucosus (Hall, 1858) [new combination].
Additionally, Thinocrinus algerianensis (Webster et al., 2004)[new combination]; Thinocrinus cheguiaensis (Webster et al.,2004) [new combination]; Thinocrinus combinatus (Webster, et al.,2004) [new combination] fromAlgeria; andThinocrinus brevispina(Wanner, 1924) [new combination];Thinocrinus brouweri (Wanner,1924) [new combination]; Thinocrinus carinatus (Wanner, 1937)[new combination]; Thinocrinus dilatus (Wanner, 1924) [newcombination]; Thinocrinus exornatus (Wanner, 1937) [newcombination]; and Thinocrinus permicus (Wanner, 1924)[new combination] from Timor.
Diagnosis.—Basal circlet high or low; radial circlet high; one ortwo fixed secundibrachials; primibrachitaxis highest brachitaxisin vertical wall of calyx; two or three ranges in regular interray;three, four, or five ranges in posterior interray; interrays incontact with tegmen; plating in proximal interrays 1 to 2 or 1 to3; fixed intrabrachials between half-rays present or absent; armsstrongly grouped; arm lobes present and extend laterally;tegmen lower than or equal to calyx; many small-sized or manymedium-sized on tegmen; anal tube central.
Occurrence.—Mississippian (Tournaisian) through Permian(Artinskian) of Africa, Australia, Europe, and North America.Ivorian and early Chadian (time units 3 and 4) of Belgium,England, and Ireland; early Osagean to early Meramecian (timeunit 3 to 7) of Illinois, Indiana, Iowa, Kentucky, Missouri, NewMexico, and Nevada; Serpukhovian to middle Bashkirian ofAlgeria; and Artinskian of Timor.
Remarks.—The separation of actinocrinitid genera on the basisof the strict application of diagnostic characters has resulted inThinocrinus being one of the most widespread actinocrinitidsrecognized.
Actinocrinites combinatus Webster et al., 2004 is reas-signed to Thinocrinus, making this the first late Mississippian(Serpukhovian) species recognized in Thinocrinus. Thinocrinus
13 Rhenberg et al.—Actinocrinitid concepts

combinatus has only primibrachials as part of the vertical wallof the calyx, which distinguishes it from Actinocrinites thathas either secundibrachials or higher brachitaxes fixed into thecalyx wall. The same criteria result in the PennsylvanianT. algerianensis and the six Actinocrinites species fromWanner(1924, 1937) being reassigned to Thinocrinus, i.e. T. brevispina(Wanner, 1924), T. broweri (Wanner, 1924), T. carinatus(Wanner, 1937), T. dilatus (Wanner, 1924), T. exornatus(Wanner, 1937), and T. permicus (Wanner, 1924).
The holotype of Thinocrinus lowei has three plates abovethe primanal, which is not a characteristic of the actinocrinitids.However, Ausich and Kammer (1991a) state that all otherspecimens have two plates above the primanal. Because only theholotype has the aberrant character, T. lowei is retained inthe family.
The calyx characters of Thinocrinus? sampsoni [newcomb.] are three ranges in the regular interray, fixedintrabrachials between the half-rays absent, and arm lobespresent. Physetocrinus has more ranges in the regularinterrays, fixed intrabrachials between half-rays, and the armsdo not form lobes. The feature that Thinocrinus? sampsonishares with to Physetocrinus is the lack of an anal tube onthe tegmen. Only three genera of actinocrinitids are known tolack an anal tube (Abactinocrinus, Physetocrinus, andStrotocrinus), but the other features of the species do not fitthose genera. Thinocrinus? sampsoni is also similar toAbactinocrinus burlingtonensis in that both have arm lobesand lack an anal tube. However, they lack other similarities.The calyx shape, basal height, and interray ranges are allvery different and it would not make sense to assignT.? sampsoni to the genus Abactinocrinus. Until moreinformation can be found on this species, Thinocrinus?sampsoni will be retained with this genus.
There is some confusion with the species Actinocrinitesscitulus. Kirk (1943) stated that there is some question as towhether or not Actinocrinites sharonensis (Miller and Gurley,1897) is a valid species but listed it as being synonymouswith A. scitulus. Brower (1965) stated that A. sharonensisshould be a separate species but listed A. scitulus in thesynonymy list. On the same page, Brower listed A. scitulus as aseparate species. Webster (2003) followed Brower’s firstsynonymy list, naming A. sharonensis as the valid name in theindex and A. scitulus as a junior synonym. Herein, both speciesare considered variations of one species. Because A. scitulus haspriority, the valid name for this species is Thinocrinus scitulus[new comb.].
Webster (2003) maintained Steganocrinus griffithi Millerand Gurley, 1897 (non Wachsmuth and Springer, 1897) as aseparate species, even though Kirk, (1943) and Brower (1965)synonymized it with Actinocrinites scitulus. Herein, wefollow Webster (2003) in considering it as a separate speciesfrom T. scitulus; however, it is with a junior synonym ofT. subpulchellus.
Thinocrinus subpulchellus (Miller and Gurley, 1896c)
1896c Actinocrinus subpulchellus Miller and Gurley, p. 13.1897 Actinocrinus subpulchellus; Miller, p. 732.1898 Actinocrinus subpulchellus; Weller, p. 63.
1943 Actinocrinites subpulchellus; Bassler and Moodey,p. 275.
1897 Actinocrinus griffithi Wachsmuth and Springer,p. 568.
1897 Actinocrinus griffithi; Miller, p. 731.1898 Actinocrinus griffithi; Weller, p. 59.1943 Actinocrinites griffithi; Bassler and Moodey, p. 270.1973 Actinocrinites griffithi; Laudon, p. 29.1977 Actinocrinites griffithi; Webster, p. 28.1943 Actinocrinus eximius Kirk, p. 264.1973 Actinocrinites eximius; Webster, p. 35.1897 Steganocrinus griffithi Miller and Gurley, p. 34.1897 Steganocrinus griffithi; Miller, p. 753.1898 Steganocrinus griffithi; Weller, p. 600.1943 Steganocrinus griffithi; Bassler and Moodey, p. 683.
Remarks.—Kirk (1943) resassigned Actinocrinites griffithi(Wachsmuth and Springer, 1897) to A. eximius due to namepriority. However, this was not a necessary action as Kirk hadsynonymized Steganocrinus griffithi with A. scitulus. In Kirk’ssynonymy list for A. scitulus, he redesignated S. griffithi asA. griffithi, which was an unnecessary step. Due to this, A. eximiusis considered a junior synonym to A. griffithi (Wachsmuth andSpringer, 1897, non Miller and Gurley, 1897).
Despite the confusion created by Kirk, both speciesActinocrinites griffithi and Steganocrinus griffithi are the samespecies which are synonymized with A. subpulchellus, now-Thinocrinus subpulchellus. All three nominal species occur inthe same time unit (time unit 3) in the Burlington Limestone andthe slight differences in size and ornamentation are not enoughto justify three separate species.
Nomina dubia.—Twenty-one species of actinocrinitids areconsidered to be nomina dubia because of poor illustrations orlack of information on the species. Most of these species arefrom the genus Actinocrinites.
Actinocrinites tenuistriatus (Phillips, 1841) is known onlyfrom the stem and, therefore, cannot be placed into a genus.Actinocrinites granulatus Goldfuss, 1831 is known fromonly non-descript calyx plates and at least two types ofcolumnals. Because there is such little information on thespecimens assigned to the name, we consider it to be nomendubia. As noted above, Actinocrinites granulatus Wright,1955b was transferred to Thinocrinus, keeping the name valid.Actinocrinites cf. A. brouweri (Wanner, 1924), A. laevisMiller, 1821, and Teleiocrinus? sibircus (Yakovlev in Yakovlevand Ivanov, 1956) are known only by a few plates, whichdo not give enough information to assign them confidently toa genus.
Several species of actinocrinitids have more than just a fewplates preserved, but lack enough information to assign themcorrectly anywhere, mostly due to the lack of any tegminalinformation. Actinocrinites pusillus (M’Coy, 1844) has only apartial calyx that has been illustrated and described. The tegmenis missing and only one ray with partial interrays on either sideare preserved.? Actinocrinites polydactylus Miller, 1821 andActinocrinites constrictus (M’Coy, 1844) are nothing butinternal molds of the calyx. Although individual plates can beseen in the molds, there is no information on whether or
Journal of Paleontology 89(1):1–19 14

not there are arm lobes or any information on the tegmens;therefore, there is not enough information to assign them to a genus.Both Japanese species of Actinocrinitidae, A. higuchisawensisMinato, 1951 and A. ohmoriensis Minato, 1951, consist only of acalyx, which are generic enough that both could be placed into anynumber of actinocrinitid genera. Manillacrinus sp. Campbell andBein, 1971, though in open nomenclature, only has a partial thecapreserved that could be part of several different genera ofactinocrinitids that lack arm lobes.
Actinocrinites caryocrinoides M’Coy, 1844 is based on aspecimen with only four basals, making it abnormal for anactinocrinitid. The characters that have been illustrated do notagree with a single genus of actinocrinitid. The arms are brokenoff above the second primibrachial (M’Coy, 1844, pl. 26, fig. 5);therefore, it is unknown if arm lobes exist or how manysecundibrachials may be fixed into the lobes and therefore thecalyx. Unlike other actinocrinitids, there appears to be only oneplate in the interrays. The type is missing, and there is noinformation on where the specimen was collected. Therefore,further study is unlikely to occur and this species should beconsidered nomen dubium.
The specimen associated with Actinocrinites goldfussi(Schmidt, 1930) is a circular partial theca showing the CDinterray and part of the tegmen. There is no information on therays, the arms or most of the tegmen, which is why this speciesis placed into nomen dubium.
Actinocrinites marcoui Collignon, 1924 is considered anomen dubium because the only illustration of it is a platediagram. Although the plate diagram may be accurate, it doesnot illustrate specimens of the species; and therefore, it cannotbe compared to other species within the genus. A. decadactylus(Gilbertson, 1843) is not figured, thus becoming nomendubium.
Actinocrinites skourensis (Termier, G. and Termier, H.,1950) is illustrated only as a line drawing of a partial theca withvery little information on the crinoid. Unless more informationcan be discovered, this species will remain a nomen dubium.
Actinocrinites tenuis (de Koninck and Le Hon, 1854) isillustrated only as a part of the calyx. Based on the illustration,A. tenuis appears to belong in the genus Aryballocrinus due tothe wide interrays and appearance of thin plates. However,without knowing what the posterior interray looks like orknowing that the plates are thin, this species is regarded asnomen dubium.
Aacocrinus triarmatus Brower, 1967 was named for twospecimens that are poorly preserved. One specimen is very small(only 4 mm), a possible juvenile, and lacks a tegmen, and theother is a set of isolated arms. The preservation of these twospecimens is too poor to have been used as the basis of a newspecies and therefore A. triarmatus is designated as nomendubium.
Aacocrinus acylus Webster and Jell, 1999 is considerednomen dubium until the specimens can be examined. Theillustrations do not show distinct plate outlines on the calyx. Theillustration of the holotype (Webster and Jell, 1999, fig. 4c)shows a crushed calyx with fractures that appear to follow theoutline of a large calyx plate. If the facture indeed follows aplate, the holotype is more likely to belong to the familyPlatycrinitidae.
Cactocrinus excerptus (Hall, 1861a) has neither beenillustrated nor have any type specimens been located for study.Until type specimens are found, C. excerptus is regarded anomen dubium. Cactocrinus fosteri (McChesney, 1861) is alsoconsidered a nomen dubium. This species name was assigned toa specimen that did not have a tegmen and the illustration is allthat remains of the original type; the types were lost in theChicago fire of 1871 (Wachsmuth and Springer, 1897).
Incertae sedis.—Actinocrinites spinaetectus (Wanner, 1937)from the Permian of Timor appears to be a beautiful specimenbased on the illustration (Wanner, 1937, pl. 1, fig. 3–4) butassigning it to a genus is difficult. The calyx of A. spinaetectusfits the characters of Steganocrinus (e.g. the primibrachitaxisis highest in the vertical wall and there are no fixed secundi-brachials) but the lack of an anal tube makes it morelike Abactinocrinus. As neither Steganocrinus nor Abactino-crinus are known past the Tournaisian, it was decided to notforce this species into a genus until the actual specimen can beexamined.
Eumorphocrinus porteri (Whidborne, 1896) is consideredincertae sedis due to poor preservation. The illustrations ofWhidborne (1898, pls. 30, fig. 8; 31, figs. 1,2, and 5) and Laneet al. (2001, pls. 1, fig. 6; 2, figs. 4 and 6; 4, fig. 1) are not clearenough to determine where this species fits within theActinocrinitidae. Lane et al. (2001) diagnosed this species ashaving two secundibrachials in each half-ray, which is acharacter of the genus, but this does not match the illustrations.Where the plates can be distinguished on the calyx, there is onlya single secundibrachial and not two. Therefore, more completespecimens are needed before this species can be assigned toa genus.
One species and one genus previously assigned toActinocrinitidae have characters that do not occur in the family.Actinocrinites zhaoae (Lane et al., 1997) was removed fromUperocrinus byWaters et al. (2003) and placed into Actinocrinitesbased upon the hexagonal first primibrachials. However, they didnot take into account that there are three plates following theprimanal. The Actinocrinitidae only have two plates above theprimanal. There is only one specimen of this species, thus it cannotbe known if, like Thinocrinus lowei, three plates above theprimanal is an aberrant feature or the norm. Until morespecimens can be found, this species should be excluded fromthe Actinocrinitidae.
The genus Ilmocrinus has also been determined to be incertaesedis. The known specimens of I. dissymmetricus Solov’yeva,1984 (1985) are poorly preserved, but the shape of the calyx platesis preserved. Actinocrinitids have a hexagonal first primibrachialwhereas Ilmocrinus has a quadrangular one. This feature removesIlmocrinus from the Actinocrinitidae. Similarly, Blairocrinusgrafensis Webster et al., 2004, from the Bashkirian of Algeria,has a quadrangular first primibrachial.
Acknowledgments
For making specimens available for study, we thank K. Hollis,Smithsonian Institution, and D.B. Blake, University ofIllinois. G.D. Sevastopulo and D.L. Meyer made important
15 Rhenberg et al.—Actinocrinitid concepts

improvements to the final manuscript. This research was sup-ported by the National Science Foundation (EAR-02059068 [toW.I.A.] and EAR-0206307 [to T.W.K.]).
References
Ausich, W.I., 2003, Lower Mississippian crinoid (Echinodermata) fauna fromUtah County, Utah: Journal of Paleontology, v. 77, p. 139–145.
Ausich, W.I., and Kammer, T.W., 1991a, Late Osagean and MeramecianActinocrinites (Echinodermata, Crinoidea) from the Mississippianstratotype region: Journal of Paleontology, v. 65, p. 485–499.
Ausich, W.I., and Kammer, T.W., 1991b, Systematic revision of Aorocrinus,Dorycrinus, Macrocrinus, Paradichocrinus, Strotocrinus, and Uperocrinus:Mississippian camerate crinoids (Echinodermata) from the stratotype region:Journal of Paleontology, v. 65, p. 936–944.
Ausich, W.I., and Kammer, T.W., 2006, Stratigraphical and geographical distributionof Mississippian (Lower Carboniferous) Crinoidea from England and Wales:Proceedings of the Yorkshire Geological Society, v. 56, no. 1, p. 91–109.
Ausich, W.I., and Kammer, T.W., 2008, Generic concepts in the Amphor-acrinidae Bather, 1899 (Class Crinoidea) and evaluation of genericassignments of North American species: Journal of Paleontology, v. 82,p. 1139–1149.
Ausich, W.I., and Kammer, T.W., 2009, Generic concepts in the PlatycrinitidaeAustin and Austin, 1842 (Class Crinoidea): Journal of Paleontology, v. 83,p. 694–717.
Ausich, W.I., and Kammer, T.W., 2010, Generic concepts in the BatocrinidaeWachsmuth and Springer, 1881 (Class Crinoidea): Journal of Paleontology,v. 84, p. 32–50.
Ausich, W.I., and Meyer, D.L., 1994, Hybrid crinoids in the fossil record (EarlyMississippian, Phylum Echinodermata): Paleobiology, v. 20, p. 362–367.
Ausich, W.I., and Sevastopulo, G.D., 2001. The Lower Carboniferous (Tour-naisian) crinoids fromHook Head: CountyWexford, Ireland,Monograph ofthe Palaeontological Society. v. 617, 136. p.
Ausich, W.I., Brett, C.E., Hess, H., and Simms, M.J., 1999, Crinoid form andfunction, in Fossil Crinoids: Cambridge, Cambridge University Press, p. 3–30.
Austin, T., Sr., and Austin, T., Jr., 1842, XVIII. - Proposed arrangement of theEchinodermata, particularly as regards the Crinoidea, and a subdivision ofthe Class Adelostella (Echinidae): Annals and Magazine of Natural History,series 1, v. 10, p. 106–113.
Austin, T., Sr., and Austin, T., Jr., 1843–1847, A monograph on recent and fossilCrinoidea, with figures and descriptions of some recent and fossil alliedgenera. v. 1(2), p. 1–32 (1843); v. 1(3), p. 33–48 (1844); v. 1(4), p. 49–64(1845); v. 1(5), p. 65–80 (1846); v. 1(6–8), p. 81–128 (1847). London andBristol.
Bassler, R.S., and Moodey, M.W., 1943, Bibliographic and faunal index ofPaleozoic pelmatozoan echinoderms: Geological Society of America Spe-cial Paper, 45, 734 p.
Bowsher, A.L., 1955, New genera of Mississippian camerate crinoids, Echino-dermata Article 1: University of Kansas Paleontological Contributions,23 p.
Bowsher, A.L., and Ubaghs, G., 1978, Family Actinocrinitidae, T452–T462, inMoore, R.C. and Teichert, K. eds., Treatise on Invertebrate Paleontology,Echinodermata, Pt. T(2) Boulder, Colorado, and Lawrence, Kansas,Geological Society of America and University of Kansas Press.
Bronn, H.G., 1849, Index palaeontologicus, unter Mitwirking der Herren Prof.H. R. Göppert und H. vonMeyer, Handbuch einer Geschichte der Nature, 5,Abt. 1, A, Nomenclator Palaeontologicus, 1381 p.
Brower, J.C., 1965, The genus, Steganocrinus: Journal of Paleontology, v. 39, p.773–793.
Brower, J.C., 1967, The actinocrinitid genera Abactinocrinus, Aacocrinus andBlairocrinus: Journal of Paleontology, v. 41, p. 675–705.
Brower, J.C., 1969, Chapter 12, Crinoids, in McKee, E.D. and Gutschick, R.C.eds., History of the Redwall Limestone of northern Arizona. GeologicalSociety of America, Memoir, 114.
Campbell, K.S.W., and Bein, J., 1971, Some lower Carboniferous crinoids fromNew South Wales: Journal of Paleontology, v. 45, p. 419–436.
Collignon, M.M., 1924, Sur un Actinocrinide de la collection Marcou auMuseum National d 'Histoire Naturelle: Bulletin de la Société Géologiquede France, v. 24, p. 13–15.
Dun, W.S., and Benson, W.N., 1920, The geology and petrology of the GreatSerpentine Belt of New South Wales. Part IX–The geology, palaeontologyand petrography of the Currabubula District, wtih notes on adjacent regions:Section B—Paleontology: Proceedings of the Linnean Society of NewSouth Wales, v. 45, p. 337–374.
Ehlers, G.M., and Kesling, R.V., 1963, Two new crinoids from Lower Mis-sissippian rocks in southeastern Kentucky: Journal of Paleontology, v. 37,p. 1028–1041.
Gilbertson, W., 1843, In Portlock, J.E. ed., Report on the Geology of the Countyof Londonderry and of Parts of Tyrone and Fermanagh: Dublin, A. Miliken,784 p.
Goldfuss, G.A., 1826-44, Petrefacta Germaniae, tam ea, Quae in Museo Uni-versitatis Regiae Borussicae Fridericiae Wilhelmiae Rhenanea, serventur,quam alia quaecunque in Museis Hoeninghusiano Muensteriano aliisque,extant, iconibus et descriiptionns illustrata. – Abbildungen und Beschrei-bungen der Petrefacten Deutschlands und der Angränzende Länder,unter Mitwirkung des Hern Grafen Georg zu Münster, herausgegeben vonAugust Goldfuss. v. 1 (1826–1833), Divisio prima. Zoophytorumreliquiae, p. 1-114; Divisio secunda. Radiariorum reliquiae, p. 115-221[Echinodermata]; Divisio tertia. Annulatorium reliquiae, p. 222–242; v. 2(1834–40), Divisio quarta. Molluscorum acephalicorum reliquiae. I. Bivalvia,p. 65–286; II. Brachiopoda, p. 287–303; III. (1841–44), Divisio quinta. Mol-luscorum gasteropodum reliquiae, p. 1–121; atlas of plates, 1–199, Düsseldorf,Arnz & Co. v. 1, p. 1–76 (1826); p. 77–164 (1829); p. 165–240 (1831);p. 241–252 (1833); v. 2, p. 1–68 (1833); p. 69–140 (1836); p. 141–224 (1837);p. 225–312 (1840); v. 3, p. 1–128 (1844).
Hall, J., 1858, Chapter 8. Palaeontology of Iowa, in Hall, J. and Whitney, J.D.eds., Report of the Geological Survey of the state of Iowa: Embracing theresults of investigations made during portions of the years 1855, 56 & 57:v. 1, part II; Palaeontology, p. 473–724, 29 pl., index to Part II separatelypaginated, 3 p.
Hall, J., 1859, Contributions to the palaeontology of Iowa, being descriptions ofnew species of Crinoidea and other fossils: Supplement to vol. I, part II, ofthe Geological Report of Iowa, 92 p.
Hall, J., 1860, Contributions to the palaeontology of Iowa: being descriptions ofnew species of Crinoidea and other fossils: Iowa Geological Survey, v. 1,no. 2, supplement, 94, p.
Hall, J., 1861a, Descriptions of new species of Crinoidea from the Carboniferousrocks of the Mississippi Valley: Journal of the Boston Society of NaturalHistory, v. 3, p. 261–328.
Hall, J., 1861b, Descriptions of new species of Crinoidea; from investigations ofthe Iowa Geological Survey, preliminary notice: Albany, New York, C. vanBenthuysen, 18 p.
Hall, J., 1863. Preliminary notice, of some species of Crinoidea from theWaverly Sandstone series of Summit Co., Ohio, supposed to be of the age ofthe Chemung Group of New York: Preprint of Seventeenth Annual Reportof the Regents of the University of the state of New-York, on the Conditionof the State Cabinet of Natural History, and the Historical and AntiquarianCollection annexed thereto, State of New York in Senate Document 189:Albany, New York, Comstock and Cassiday Printers, p. 50–60.
Heckel, P., and Clayton, G., 2005, Official names of the Carboniferous System:Geology Today, v. 21, no. 6, p. 213–214.
Kammer, T.W., and Ausich, W.I., 2006, The “Age of Crinoids” a Mississippianbiodiversity spike coincident with widespread carbonate ramps: PALAIOS,v. 21, p. 238–248.
Keyes, C.R., 1894, Paleontology of Missouri, Pt. I: Missouri Geological Survey,v. 4, p. 143–225.
Kirk, E., 1943, A revision of the genus Steganocrinus: Journal of theWashington Academy of Science, v. 33, p. 259–265.
Kirk, E., 1944, Cytidocrinus, a new name for Cyrtocrinus Kirk: Journal of theWashington Academy of Science, v. 34, p. 85.
Koninck, L.G. de, and Le Hon, H., 1854, Recherches sur les crinoides du terrainCarbonifere de la Belgique: Academie Royal de Belgique, Memoir, v. 28,no. 3, 215 p.
Lane, N.G., Waters, J.A., and Maples, C.G., 1997, Echinoderm faunas of theHongguleleng Formation, Late Devonian (Famennian), Xinjiang-UygurAutonomous Region, People’s Republic of China: Journal of Paleontology,v. 71, Memoir (supplement to no. 2), p. 1–43.
Lane, N.G., Maples, C.G., and Waters, J.A., 2001, Revision of Late Devonian(Famennian) and some Early Carboniferous (Tournaisian) crinoids andblastoids from the type Devonian area of North Devon: Palaeontology,v. 44, p. 1043–1080.
Laudon, L.R., 1933, The stratigraphy and paleontology of the Gilmore CityFormation of Iowa: University of Iowa Studies, v. 15, no. 2, p. 1–74.
Laudon, L.R., 1973, Stratigraphic crinoid zonation in Iowa Mississippian rocks:Proceedings of the Iowa Academy of Science, v. 80, p. 25–33.
Laudon, L.R., Parks, J.M., and Spreng, A.C., 1952, Mississippian crinoid faunafrom the Banff Formation, Sunwapta Pass, Alberta: Journal of Paleontol-ogy, v. 26, p. 544–575.
Laudon, L.R., and Severson, J.L., 1953, New crinoid fauna, Mississippian,Lodgepole Formation, Montana: Journal of Paleontology, v. 27, p. 505–536.
Lee, K., Ausich, W.I., and Kammer, T.W., 2005, Crinoids from the NadaMember of the Borden Formation (Lower Mississippian) in easternKentucky: Journal of Paleontology, v. 79, p. 337–355.
Lindley, I.D., 1979, An occurrence of the camerate crinoid genus Eumorphocrinusin the Early Carboniferous of New South Wales: Journal and Proceedings ofthe Royal Society of New South Wales, v. 112, no. 3–4, p. 121–124.
Journal of Paleontology 89(1):1–19 16

Lindley, I.D., 1988, Glaphyrocrinus, a new camerate crinoid genus from theLower Carboniferous of New South Wales: Alcheringa, v. 12, p. 129–136.
Macurda, D.B., Jr., 1974, A quantitative phyletic study of the cameratecrinoid Families Actinocrinitidae and Periechocrinitidae and its taxonomicimplications: Journal of Paleontology, v. 48, p. 820–832.
Matthews, S.C., 1973, Notes on open nomenclature and on synonymy lists:Palaeontology, v. 16, no. 4, p. 713–719.
McChesney, J.H., 1861, Descriptions of new fossils from the Paleozoic rocks ofthe western states: Transactions of the Chicago Academy of Science, v. 2,p. 77–95.
M’Coy, F., 1844, Echinodermata. InGriffith, R., A Synopsis of the Characters of theCarboniferous Limestone Fossils of Ireland: Dublin, University Press, 274 p.
Meek, F.B., and Worthen, A.H., 1860, Descriptions of new species of Crinoideaand echinoidea from the Carboniferous rocks of Illinois, and otherwestern states: Proceedings of the Academy of Natural Sciences ofPhiladelphia, v. 12, p. 379–397.
Meek, F.B., and Worthen, A.H., 1866a, Descriptions of invertebrates from theCarboniferous system: Illinois Geological Survey, v. 2, no. 2, p. 143–411.
Meek, F.B., and Worthen, A.H., 1866b, Contributions to the palaeontology ofIllinois and other western states: Proceedings of the Academy of NaturalSciences of Philadelphia (1865), v. 17, p. 251–274.
Meek, F.B., and Worthen, A.H., 1869, Descriptions of new Carboniferousfossils from the western states: Proceedings of the Academy of NaturalSciences of Philadelphia, v. 22, p. 137–172.
Meek, F.B., and Worthen, A.H., 1870, Descriptions of new Carboniferousfossils from the western states: Proceedings of the Academy of NaturalSciences of Philadelphia, v. 21, p. 137–172.
Miller, J.S., 1821, A Natural History of the Crinoidea, or Lily-ShapedAnimals; with Observations on the Genera, Asteria, Euryale, Comatulaand Marsupites: Bristol, England, Bryan & Co., 150 p.
Miller, S.A., 1881, Subcarboniferous fossils from the Lake Valley MiningDistrict of New Mexico, with descriptions of new species: Journal of theCincinnati Society of Natural History, v. 4, no. 4, p. 306–315.
Miller, S.A., 1889, North American Geology and Paleontology: Cincinnati,Ohio, Western Methodist Book Concern, 664 p.
Miller, S.A., 1891, Palaeontology. Advance sheets, Indiana Department ofGeology and Natural Resources, 17th Annual Report, 103 p.
Miller, S.A., 1892. North American Geology and Paleontology, first appendix:Cincinnati, Ohio, Western Methodist Book Concern, p. 665–718.
Miller, S.A., and Gurley, W.F.E., 1893, Description of some new species ofinvertebrates from the Palaeozoic rocks of Illinois and adjacent states:Illinois State Museum Bulletin, v. 3, 81 p.
Miller, S.A., and Gurley, W.F.E., 1894, New genera and species of Echino-dermata: Illinois State Museum Bulletin, v. 5, 53. p.
Miller, S.A., and Gurley, W.F.E., 1895, New and interesting species ofPalaeozoic fossils: Illinois State Museum Bulletin, v. 7, 89 p.
Miller, S.A., and Gurley, W.F.E., 1896a, Description of new and remarkablefossils from the Palaeozoic rocks of the Mississippi Valley: Illinois StateMuseum Bulletin, v. 8, 65 p.
Miller, S.A., and Gurley, W.F.E., 1896b, New species of crinoids from Illinoisand other states: Illinois State Museum Bulletin, v. 9, 66 p.
Miller, S.A., and Gurley, W.F.E., 1896c, New species of Echinodermata and anew crustacean from the Palaeozoic rocks: Illinois State Museum Bulletin,v. 10, 91 p.
Miller, S.A., and Gurley, W.F.E., 1897, New species of crinoids, cephalopods,and other Palaeozoic fossils: Illinois State Museum Bulletin, v. 12, 69 p.
Minato, M., 1951, On the Lower Carboniferous fossils of the Kitakami Massif,NE Honsyu, Japan: Journal of the Faculty of Science, Hokkaido University,v. 7, p. 355–382.
Moore, R.C., and Laudon, L.R., 1943, Evolution and classification ofPaleozoic crinoids: Geological Society of America Special Paper, v. 46,151 p.
Moore, R.C., and Teichert, C., eds., 1978, Treatise on Invertebrate Paleontology,Part T, Echinodermata 2, Crinoidea. 3 vols. Geological Society of America andUniversity of Kansas, 1026 p.
Morris, J., 1843, A Catalogue of British Fossils. Comprising all the Genera andSpecies Hitherto Described; with Reference to their Geological Distributionand to the Localities in which They Have Been Found, 1st ed.: London,John Van Voorst, 222 p.
Owen, D.D., and Shumard, B.F., 1852, Descriptions of seven new species ofcrinoidea from the subcarboniferous of Iowa and Illinois: Journal of theAcademy of Natural Sciences of Philadelphia, series 2, v. 2, p. 89–94.
Peck, R.E., and Keyte, I.A., 1938, The Crinoidea of the Chouteau Limestone,in Branson, E.B. ed., Stratigraphy and Paleontology of the LowerMississippian of Missouri, Pt. 2. Missouri University Studies, v. 13,no. 4 p. 70–108.
Phillips, J., 1836, Illustrations of the Geology of Yorkshire, or a Description ofthe Strata and Organic Remains. Pt. 2 The Mountain Limestone districts,2nd ed: London, John Murray, p. 203–208.
Phillips, J., 1841, Figures and Descriptions of the Palaeozoic Fossils of Corn-wall, Devon, and West Somerset; Observed in the Course of the OrdinanceGeological Survey of that District: London, Longmans, Brown, Green, andLongmans, 232 p. 60 pl. (Crinoidea, p. 28–32, pl. 15–16).
Portlock, J.E., 1843, Report on the Geology of the County of Londonderry andof Parts of Tyrone and Fermanagh: Dublin, A. Milliken, 784 p.
Rhenberg, E.C., and Kammer, T.W., 2013, Camerate crinoids from the NunnMember (Tournaisian, Osagean) of the lower Mississippian Lake ValleyFormation, New Mexico: Journal of Paleontology, v. 87, p. 312–340.
Rowley, R.R., 1900, New species of crinoids, blastoids and cystoids fromMissouri: American Geologist, v. 25, p. 65–75.
Rowley, R. R., 1901–04, Description of fossils, in Green, G.K. ed., Contributionto Indiana Palaeontology: New Albany, Indiana, v. 1, no. 7, p. 50–60,pl. 19–21 (1901a); no. 8, p. 62–74, pl. 22–24 (1901b); no. 10, p. 85–97,pl. 28–30 (1902a); no. 11, p. 98–109, pl. 31–33 (1903a); no. 12, p. 110–129,pl. 34–36 (1903b); no. 13, p. 130–137, pl. 37–39 (1903c); no. 14, p. 138–145, pl. 40–42 (1903d); no. 15, p. 146–155, pl. 43–45 (1903e); no. 16,p. 156–167, pl. 46–48 (1903f); no. 17, p. 168–175, pl. 49–51 (1904a);no. 18, p. 176–184, pl. 52–54 (1904b); no. 19, p. 185–197, pl. 55–57 (1904c).
Rowley, R.R., and Hare, S.J., 1891, Description of some new species ofEchinodermata from the sub-Carboniferous rocks of Pike County, Mo:Kansas City Scientist, v. 5, no. 7, p. 97–103.
Sallan, L.C., Kammer, T.W., Ausich, W.I., and Cook, L.A., 2011, Persistentpredator-prey dynamics revealed by mass extinction: Proceedings of theNational Academy of Science, v. 108, p. 8335–8338.
Schlotheim, E.F., von, 1820, Die Petrefactenkunde auf ihrem jetzigen Stand-punkte durch die Beschreibung seiner Sammlung versteinerter und fossilerÜberreste des Thier-und Pflanzenreichs der Vorwelt erläutert: Gotha,Beckersche Buchhandlung, 437 p.
Schmidt, W.E., 1930, Die Echinodermen des deutschen Unterkarbons: Abhandlun-gen der Preussichen Geologischen Landesanstalt, n. s., v. 122, no. 1, p. 1–92.
Shumard, B.F., 1855, Description of new species of organic remains: MissouriGeological Survey, v. 2, p. 185–208.
Shumard, B.F., 1857, Description of new fossil Crinoidea from the Palaeozoicrocks of the western and southern portions of the United States:Transactions of the St. Louis Academy of Science, v. 1, p. 71–80.
Solov'Yeva, M.V., 1984, Novyi rod krinoidei kamerat iz Karbona Urala[A new genus of camerate crinoid from the Carboniferous of the Urals]:Paleontologicheskii Zhurnal, v. 4, p. 114–118.
Solov'Yeva, M.V., 1985, A new Carboniferous genus of camerate crinoid fromthe Urals: Paleontological Journal, v. 18, p. 114–118.
Swofford, D.L., 2001, PAUP*: Phylogenetic Analysis Using Parsimony, version4.0b10. Computer program distributed by Sinauer Associates, Inc.,Sunderland, MA (http://www.paup.csit.fsu.edu).
Termier, G., and Termier, H., 1950, Paléontologie Marocaine II. Invertébres del'Ere Primaire. 4. Annélides, Arthropodes, Echinodermes, Conularides etGraptolithes: Service Carte GéologiqueMorocco, Notes et Mémoires, v. 79,no. 4, p. 1–279.
Ubaghs, G., 1978a, Skeletal morphology of fossil crinoids, inMoore, R.C., andTeichert, K. eds., Treatise on Invertebrate Paleontology, Echinodermata,Pt. T(1) Boulder, Colorado, and Lawrence, Kansas, Geological Society ofAmerica and University of Kansas Press, p. T59–T216.
Ubaghs, G., 1978b, Camerata, inMoore, R.C. and Teichert, K. eds., Treatise onInvertebrate Paleontology, Echinodermata, Pt. T(2) Boulder, Colorado,and Lawrence, Kansas, Geological Society of America and University ofKansas Press, p. T408–T519.
Ubaghs, G., 1978c, Subfamily Eumorphocrininae, in Moore, R.C. andTeichert, K. eds., Treatise on Invertebrate Paleontology, Echinodermata, Pt.T(2): Boulder, Colorado, and Lawrence, Kansas, Geological Society ofAmerica and University of Kansas Press, p. T456–T458.
Ubaghs, G., 1978d, Subfamily Cactocrininae, in Moore, R.C. and Teichert, K.,eds., Treatise on Invertebrate Paleontology, Echinodermata, Pt. T(2):Boulder, Colorado, and Lawrence, Kansas, Geological Society of Americaand University of Kansas Press, p. T458–T460.
Ubaghs, G., 1978e, Subfamily Physetocrininae, p. T460–T462. InMoore, R. C.and Teichert K. eds., Treatise on Invertebrate Paleontology, Echinodermata,Pt. T(2) Geological Society of America and University of Kansas Press:Boulder and Lawerence.
Van Sant, J.F., 1965, Actinocrinites grandissimus, a new name for a cameratecrinoid from Borden (lower-Middle Mississippian) rocks of Indiana:Journal of Paleontology, v. 39, p. 290–292.
Wachsmuth, C., and Springer, F., 1880–1886, Revision of the Palaeocrinoidea.Proceedings of the Academy of Natural Sciences of Philadelphia Pt. I. Thefamilies Ichthyocrinidae and Cyathocrinidae (1880):226–378, (separaterepaged p. 1–153); Pt. II. Family Sphaeroidocrinidae, with the sub-familiesPlatycrinidae, Rhodocrinidae, and Actinocrinidae (1881):177–411 (sepa-rate repaged, p. 1–237); Pt. III, Sec. 1. Discussion of the classification andrelations of the brachiate crinoids, and conclusion of the generic descrip-tions (1885):225–364 (separate repaged, p. 1–138); Pt. III, Sec. 2.
17 Rhenberg et al.—Actinocrinitid concepts

Discussion of the classification and relations of the brachiate crinoids, andconclusion of the generic descriptions (1886):64–226 (separate repaged tocontinue with section 1, 139–302).
Wachsmuth, C., and Springer, F., 1897, The North American CrinoideaCamerata: Harvard College Museum of Comparative Zoology, Memoirs20 and 21, 897 p.
Wanner, J., 1924, Die Permischen Krinoiden von Timor. Jaarbook van netMijnwezen Nederlandes Oost-Indie, Verhandlungen (1921): Gedeelte, v. 3,p. 1–348.
Wanner, J., 1937, Neue beiträge zur kenntnis der Permischen echinodermen vonTimor, VIII-XIII: Palaeontographica, Supplement 4, IV Abteilungen,Abschnitt v. 1, p. 1–212.
Waters, J.A., Maples, C.G., Lane, N.G., Marcus, S., Liao, Z.T., Liu, L., Hou, H.-F., and Wang, J.-X., 2003, A quadrupling of Famennian pelmatozoandiversity: new Late Devonian blastoids and crinoids from northwest China:Journal of Paleontology, v. 77, no. 5, p. 922–948.
Webster, G.D., 1973, Bibliography and index of Paleozoic crinoids, 1942–1968:Geological Society of America, Memoir 137, 341 p.
Webster, G.D., 1977, Bibliography and index of Paleozoic crinoids, 1969–1973:Geological Society of America, Microform Publication 8, 235 p.
Webster, G.D., 2003, Bibliography and index of Paleozoic crinoids, coronates,and hemistreptocrinoids 1758–1999: Geological Society of America Spe-cial Paper 363.
Webster, G.D., and Jell, P.A., 1999, New Carboniferous crinoids from easternAustralia: Memoirs of the Queensland Museum, v. 43, no. 1, p. 237–278.
Webster, G.D., and Lane, N.G., 1987, Crinoids from the Anchor Limestone(Lower Mississippian) of the Monte Cristo Group, Southern Nevada: Uni-versity of Kansas Paleontological Contributions Paper, v. 119, 55 p.
Webster, G.D., Maples, C.G., Sevastopulo, G.D., Frest, T.J., and Waters, J.A.,2004, Carboniferous (Visean–Moscovian) echinoderm faunas from theBechar Basin area of western Algeria: Bulletin of American Paleontology,v. 368, 98 p.
Weller, S., 1898, A bibliographic index of Carboniferous invertebrates: U. S.Geological Survey, Bulletin 153, 653 p.
Weller, S., 1909, Kinderhook faunal studies - V, the fauna of the Fern GlenFormation: Geological Society of America, Bulletin 20, p. 265–332.
Whidborne, G.F., 1896, A preliminary synopsis of the faunas of the PickwellDown, Baggy, and Pilton Beds: Proceedings of the Geological Association,v. 14, p. 371–377.
Whidborne, G.F., 1898, A monograph of the Devonian fauna of the south ofEngland. The fauna of the Marwood and Pilton Beds. PalaeontographicalSociety, Monograph 52, v. 3, p. 214–236.
White, C.A., 1874, Preliminary Report upon Invertebrate Fossils Collected bythe Expeditions of 1871, 1872, and 1873. Geographical and GeologicalExploration and Surveys West of the 100th Meridian: Washington, D.C.,Government Printing Office, p. 1–27.
Whitfield, R.P., 1900, Description of a new crinoid from Indiana: AmericanMuseum of Natural History, Bulletin 13, p. 23–24.
Wood, E., 1914, The use of crinoid arms in studies of phylogeny: Annals of theNew York. Academy of Science, v. 24, p. 1–17.
Wright, J., 1947, Steganocrinus westheadi n. sp. and note on a rare crinoid and ablastoid from the Carboniferous Limestone of Coplow Knoll, Clitheroe:Geological Magazine, v. 84, p. 101–105.
Wright, J., 1950–1960, The British Carboniferous Crinoidea. PalaeontographicalSociety, Monograph, v. 1(1), p. 1–24, 1950; v. 1, no. 2, p. 25–46, 1951a; v. 1,no. 3, p. 47–102, 1951b; v. 1, no. 4, p. 103–148, 1952a; v. 1, no. 5, p. 149–190,1954a; v. 2, no. 1, p. 191–254, 1955a; v. 2, no. 2, p. 255–272, 1955b; v. 2, no. 3,p. 273–306, 1956b; v. 2, no. 4, p. 307–328,1958; v. 2, no. 5, p. 329–347 1960.
Yakovlev, N.N., and Ivanov, A.P., 1956, Morskie lilii i blastoidei Kamennou-golnykh i Permskikh otlozhenii SSSR [Marine crinoids and blastoids of theCarboniferous and Permian deposits of Russia]: Vsesoyuznogo Nauchno-Issledovatelskii Geologicheskogo Institut Trudy, v. 11, p. 1–142.
Accepted 21 January 2014
Appendix 1. Diagnostic characters for genera of theActinocrinitidae (see also Table 3)
A. Basal Circlet: 0, low; 1, high.B. Radial Circlet: 0, low; 1, high.C. Number of fixed secundibrachials: 0, 0; 1, 1; 2, 2; 3, 3.D. Highest brachial in the vertical wall of the calyx before arm
lobes: 0, Primibrachials; 1, Secundibrachials; 2, Tertibrachials; 3,Quartibrachials.
E. Number of ranges in regular interrays: 0, 2; 1, 3; 2, 4; 3, 5+;4, 10+.
F. Number of ranges in posterior interray: 0, 3; 1, 4; 2, 5; 3,6; 4, 10.
G. Interrays in contact with tegmen: 0, yes; 1, no.H. Plating in proximal interrays: 0, 1 to 2; 1 to 3.I. Number of plates above primanal: 0, 3; 1, 2.J. Arms grouped: 0, strongly; 1, no; 2 weakly.K. Arm lobes present: 0, yes; 1, no.L. Fixed intrabrachials between half-rays: 0, present; 1, absent.M. Tegmen as high as calyx: 0, lower; 1, higher; 2, same.N. Anal tube: 0, central; 1, eccentric; 2, none.O. Tegmen plates: 0, many medium; 1, many small; 2, few large.
Journal of Paleontology 89(1):1–19 18

Appendix 2. Character states for phylogenetic analysis of the Actinocrinitidae
Characters
Genera A B C D E F G H I J K L M N O
Periechocrinus 1 0 2 2 3 4 0 0 0 0 1 0 0 1 1Megistocrinus 0 1 2 2 3 3 0 0 0 2 1 0 0 1 (1,2)Aacocrinus 0 1 (1,2) 1 1 (0,1,2) 0 0 1 0 0 1 0 (0,1) (0,2)Abactinocrinus 0 1 (0,1) 0 (0,1) 1 0 (0,1) 1 0 0 1 0 2 1Actinocrinites (0,1) 1 1 (1,2) (1,2,3) (1,2) 0 1 1 0 0 (0,1) 0 (0,1) (0,1)Blairocrinus 0 0 1 0 0 1 0 0 1 0 0 1 1 0 2Cactocrinus 0 0 1 1 (0,1) 0 1 0 1 1 1 0 (0,2) 0 0Cusacrinus (0,1) 1 1 1 2 3 0 0 1 2 1 0 (0,2) 0 0Cytidocrinus 1 1 0 0 2 1 0 0 1 0 1 1 0 0 1Dialutocrinus (0,1) (0,1) 1 (1,2) 1 (1,2,3) 0 0 1 (0,2) 1 0 0 0 0Eumorphocrinus 1 1 2 2 3 2 0 0 1 0 1 (0,1) 0 0 1Glaphyrocrinus 1 1 2 1 2 (1,2) 0 0 1 0 1 0 0 0 2Iotacrinus 0 0 (1,2) (0,1) (0,1) (0,1) 0 (0,1) 1 0 0 (0,1) (0,1) (0,1) 0Maligneocrinus 0 1 1 2 3 2 0 0 1 2 1 0 0 ? ?Manillacrinus 1 1 1 1 1 3 0 0 1 0 1 0 (0,2) 0 1Nunnacrinus 0 (0,1) (0,1) (0,1) 2 2 0 0 1 (0,2) 1 0 (0,2) (0,1) 0Physetocrinus (0,1) 1 1 (2,3) 3 2 0 0 1 (0,2) 1 0 0 2 1Sampsonocrinus (0,1) 1 (0,1,2) (0,1) 0 (0,1) 0 0 1 0 0 0 0 (0,1) (0,1)Steganocrinus 0 1 0 0 0 0 0 0 1 0 0 0 (0,2) 1 2Strotocrinus 1 1 1 (1,2) 4 ? 1 (0,1) 1 1 1 0 0 2 1Teleiocrinus 1 1 1 2 3 1 (0,1) 0 1 1 1 0 0 0 1Thinocrinus (0,1) 1 (1,2) 0 (0,1) (0,1,2) 0 (0,1) 1 0 0 1 0 (0,1) (0,1)
19 Rhenberg et al.—Actinocrinitid concepts