isbn978-951-27-1183-3
TRANSCRIPT

ELENA AUTIO
Loose Housing of Horses in a Cold Climate
Effects on Behaviour, Nutrition, Growthand Cold Resistance
JOKAKUOPIO 2008
KUOPION YLIOPISTON JULKAISUJA C. LUONNONTIETEET JA YMPÄRISTÖTIETEET 245KUOPIO UNIVERSITY PUBLICATIONS C. NATURAL AND ENVIRONMENTAL SCIENCES 245
Doctoral dissertation
To be presented by permission of the Faculty of Natural and Environmental Sciences
of the University of Kuopio for public examination in Auditorium,
Mediteknia building, University of Kuopio
on Friday 5th December 2008, at 12 noon
Department of BiosciencesUniversity of Kuopio

Distributor : Kuopio University Library P.O. Box 1627 FI-70211 KUOPIO FINLAND Tel. +358 40 355 3430 Fax +358 17 163 410 http://www.uku.fi/kirjasto/julkaisutoiminta/julkmyyn.html
Series Editors: Professor Pertti Pasanen, Ph.D. Department of Environmental Science
Professor Jari Kaipio, Ph.D. Department of Physics
Author’s address: Equine Information Centre P.O. Box 1627 FI-70211 KUOPIO FINLAND Tel. +358 440 986 622 E-mail : [email protected] i
Supervisors: Minna-Liisa Heiskanen, Ph.D. Equine Information Centre Kuopio
Professor Jaakko Mononen, Ph.D. Department of Biosciences University of Kuopio
Reviewers: Professor Esa Hohtola, Ph.D. Department of Biology University of Oulu
Karin Morgan, Ph.D. Swedish Equestrian Educational Centre Strömsholm, Sweden
Opponent: Professor Hannu Saloniemi, Ph.D. Department of Production Animal Medicine University of Helsinki
ISBN 978-951-27-1183-3ISBN 978-951-27-1098-0 (PDF)ISSN 1235-0486
KopijyväKuopio 2008Finland

Autio, Elena. Loose Housing of Horses in a Cold Climate. Effects on Behaviour, Nutrition, Growth and Cold Resistance. Kuopio University Publications C. Natural and Environmental Sciences 245. 2008. 76 p. ISBN 978-951-27-1183-3 ISBN 978-951-27-1098-0 (PDF) ISSN 1235-0486
ABSTRACT Housing of weanling horses in cold loose housing systems is common in the Nordic countries, although its suitability as a winter environment has not been studied. This thesis aimed to study the effects of a cold loose housing environment on weanling horses. The loose housing system consisted of insulated halls with a deep-litter bed, entrance shelters and paddocks to which the group-housed horses had free access. The effects on the behaviour, nutrition, growth and cold resistance of the horses were studied.
The time budget of the horses consisted mainly of eating, resting and standing (37, 32, and 26 % of daily time, respectively); only one hour (5 %) was spent on locomotor activity. Their circadian rhythm largely resembled that of feral horses. The horses acclimatized to the cold by behavioural, metabolic and insulation adjustments. Their activity was low and huddling ex-isted commonly. Energy intake increased by 1.8, 0.5 and 0.2 % in November, December and January, respectively, per 1 °C decrease in ambient temperature below the lower critical tem-perature of –11 ºC. When this demand was taken into account in the feeding, the horses gained weight at expected rates. The results for radiative heat loss and winter coat develop-ment indicated that differences existed in the cold resistance of the horses, coldblood and pony types being more resistant than light and warmblood types.
A loose housing system contributes to horse welfare by allowing horses to follow the natural circadian rhythm of behavioural patterns, but does not necessarily encourage activity. The opportunity to behave unrestrictedly rather than the opportunity for high locomotor ac-tivity seems to be more important for horses. Weanling horses cope in the cold and thus can be reared in loose housing systems provided their level of acclimatization is taken into ac-count in the feeding and that the loose housing facilities are adequate. However, due to chan-ging animal- and environment-related factors during the winter and ad libitum forage feeding, the regulation of nutrient intake is complicated and the risk of overfeeding and imbalanced nutrition is high in late winter.
With regard to horse welfare, this thesis indicates that loose housing may involve both ad-vantages and disadvantages. The suitability of loose housing for weanling horses depends on which welfare-related features are emphasized. Universal Decimal Classification: 591.615, 599.723.2, 636.083.312, 636.1 CAB Thesaurus: animal housing; loose housing; cold zones; horses; animal welfare; ther-moregulation; acclimatization; animal behaviour; animal nutrition; circadian rhythm; energy intake; growth; cold resistance; heat loss; thermography


ACKNOWLEDGEMENTS The studies in this thesis were performed at the Equine Information Centre in Kuopio and at the school farm of the Ylä-Savo Vocational Institute in Kiuruvesi. I would like to thank the Employment and Economic Development Centre of Northern Savo (The European Agricultu-ral Guidance and Guarantee Fund) and the State Provincial Office of Eastern Finland (The European Social Fund) for the opportunity to carry out these studies in connection with de-velopment projects in the horse industry. The Finnish Cultural Foundation, the University of Kuopio and the Erkki Rajakoski Foundation also provided financial support for this thesis, for which I am grateful.
My deepest gratitude goes for my supervisor, Minna-Liisa Heiskanen, PhD, (Equine In-formation Centre), whose dedication to the horse welfare generated the idea for this thesis, and whose inspiring and determined guidance made this thesis possible. I am also grateful to my supervisor, Professor Jaakko Mononen, PhD, (Department of Biosciences, University of Kuopio) for his expertise and enthusiastic support on this thesis.
I sincerely thank the reviewers, Professor Esa Hohtola, PhD, (University of Oulu) and Karin Morgan, PhD, (Swedish Equestrian Educational Centre) for their expert review and valuable comments of this thesis. I also thank William Ansell, BA, for revising the language of this thesis, and Martti Niskanen and Ari Ekholm for their technical support.
I wish to express my gratitude to the Counsellor of Vocational Education Matti Notko (Di-rector of the Ylä-Savo Federation of Municipalities for Education) for the opportunity to per-form the studies using the facilities and horses of the Ylä-Savo Vocational Institute. I also thank the personnel of the Ylä-Savo Vocational Institute, especially Arja Aalto, Timo Vääränen, Kirsi Kettunen and Terho Partanen for their assistance with the experiments. I am also grateful to my co-authors Ulla Sihto and Riitta Neste for the great effort they put into the experiments.
My special thanks go to the staff of the Equine Information Centre, to all the people who have helped me during these years. In particular, I am deeply grateful to Sanna Airaksinen and Arja Lehmuskero for their valuable advice and encouragement. I also want to express my gratitude to the Research School for Animal Welfare for the inspiring atmosphere and profit-able discussions during the seminars and courses, which have greatly contributed to my work.
Finally, I owe my warmest thanks to my dear parents and sisters for their support and un-derstanding. I am especially grateful to Sanna for the company and help during the long re-search periods in Kiuruvesi. And Venla, Marja and Lassi, I cannot even describe my gratitude to you for being there for me during these years. Kuopio, 2008 Elena Autio


ABBREVIATIONS
ADG Average daily gain BAT Brown adipose tissue BCS Body condition score BMR Basal metabolic rate BW Body weight C Coldblood horse type CED Climatic energy demand DM Dry matter FC Finnish coldblood horse, i.e. Finnhorse GPS Global Positioning System HI Heat increment HP Heat production HR Heart rate L Light horse type LCT Lower critical temperature LMM Linear Mixed Model ME Metabolizable energy MR Metabolic rate NST Non-shivering thermogenesis P Pony type QH Quarter Horse RH Relative humidity RQ Respiratory quotient SB Standardbred horse Ta Ambient temperature Tb Body temperature TB Thoroughbred horse TNZ Thermoneutral zone W Warmblood horse type
DEFINITIONS
Foal A young horse from the time of birth until weaning (Belknap, 1997) Weanling A foal separated from its dam until the first birthday (Belknap, 1997) Yearling A foal between one and two years of age (Belknap, 1997)


LIST OF ORIGINAL PUBLICATIONS
This dissertation is based on the following original publications, which are referred in the text by their Roman numerals I–IV:
I Autio, E., & Heiskanen, M-L. (2005). Foal behaviour in a loose housing/paddock envi-ronment during winter. Applied Animal Behaviour Science, 91, 277–288.
II Autio, E., Sihto, U., Mononen, J., & Heiskanen, M-L. (2008). Energy intake and growth of weanling horses in a cold loose housing system. Agricultural and Food Science. In press.
III Autio, E., Neste, R., Airaksinen, S., & Heiskanen, M-L. (2006). Measuring the heat loss in horses in different seasons by infrared thermography. Journal of Applied Animal Wel-fare Science, 9 (3), 211–221.
IV Autio, E., Mononen, J., & Heiskanen, M-L. (2007). Thermographic evaluation of the lower critical temperature in weanling horses. Journal of Applied Animal Welfare Sci-ence, 10 (3), 207–216.
This thesis also contains previously unpublished data referred to in the text as U1 and U2.


CONTENTS
1. INTRODUCTION ............................................................................................................... 13
2. REVIEW OF THE LITERATURE ..................................................................................... 152.1 Horse housing alternatives in relation to health and welfare ......................................... 15
2.1.1 Stabling in loose boxes ........................................................................................... 152.1.2 Loose housing ......................................................................................................... 16
2.2 General aspects of thermoregulation in the cold............................................................ 182.2.1 Types of thermal responses..................................................................................... 182.2.2 Body temperature.................................................................................................... 182.2.3 Heat balance............................................................................................................ 192.2.4 Heat production....................................................................................................... 202.2.5 Heat loss.................................................................................................................. 212.2.6 Thermoneutrality and lower critical temperature.................................................... 23
2.3 Thermoregulation of horses in the cold ......................................................................... 252.3.1 Lower critical temperatures for horses ................................................................... 252.3.2 Physiological responses .......................................................................................... 272.3.3 Behavioural responses ............................................................................................ 31
3. OBJECTIVES ...................................................................................................................... 33
4. MATERIALS AND METHODS......................................................................................... 354.1 Animals and housing...................................................................................................... 354.2 Animal care .................................................................................................................... 364.3 Behavioural measurements ............................................................................................ 364.4 Energy intake and growth .............................................................................................. 374.5 Thermographic examinations......................................................................................... 384.6 Statistics and the presentation of results ........................................................................ 40
5. RESULTS ............................................................................................................................ 435.1 Behavioural patterns of horses....................................................................................... 43
5.1.1 Time budget and circadian rhythm ......................................................................... 435.1.2 Distances travelled under different management conditions .................................. 44
5.2 Effects of a cold housing environment on horses .......................................................... 455.2.1 Behaviour ................................................................................................................ 455.2.2 Energy intake and growth ....................................................................................... 46
5.3 Cold resistance ............................................................................................................... 495.3.1 Radiative heat loss .................................................................................................. 495.3.2 Thermal insulation .................................................................................................. 50
6. DISCUSSION ...................................................................................................................... 536.1 Loose housing and behavioural needs ........................................................................... 546.2 Effects of a cold housing environment on horses .......................................................... 586.3 Cold resistance of different horse types......................................................................... 63
7. SUMMARY AND CONCLUSIONS .................................................................................. 67
8. PRACTICAL RECOMMENDATIONS.............................................................................. 69
9. REFERENCES .................................................................................................................... 71
APPENDIX: ORIGINAL PUBLICATIONS


Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 13
1. INTRODUCTION
A great proportion of the world’s horse population is located in cold climate count-ries or at least in countries with a cold win-ter (Langlois, 1994). The most common way of housing horses in these regions dur-ing the long indoor feeding period is con-finement in stables in loose boxes (Mills & Clarke, 2002). However, stabling limits the amount of free exercise horses can take and their social contacts, and reduces environ-mental stimulation (Fader & Sambraus, 2004). Therefore, an alternative housing system, loose housing, has been established to solve the problems caused by stabling. A loose house is a group housing system where horses have free access to an un-heated hall with bedding and a paddock (Ventorp, 1994). Free outdoor access is expected to encourage voluntary daily exer-cise of horses, which is limited in box sta-bling (Zeeb & Schnitzer, 1997).
In Finland, the number of registered horses is about 68,000 (Suomen Hippos, 2008). This number, and especially the pro-portion of horses raised for hobby and lei-sure purposes, is continuously increasing. Horse breeding is mainly carried on by small breeders, since over 80 % of Finnish breeders have only one or two broodmares (Heiskanen et al., 2002). Nowadays, it is common for small breeders to rear weaned foals during their first winter in a commer-cial group housing system, i.e. a loose housing system, in which foals are collected from several different stables in autumn. In Finland, the number of loose housing sys-tems with five to ten horses is estimated to be about 200 (executive directors of the regional horse breeding associations, per-sonal communication, October 2, 2007). In addition, there is a large, undefined number of loose houses with one to four horses.
Loose housing of horses in regions with a long and cold winter, such as Finland, exposes the horses to considerable weather fluctuations and low ambient temperatures. Scientific knowledge on the effects of the cold, especially on growing horses during prolonged exposure to cold, is limited. Re-cently, feedback from Finnish veterinarians and the executive directors of the regional horse breeding associations revealed that a lack of knowledge and skills in taking care of horses in a cold environment has caused health and welfare problems, especially in young, loose-housed horses. The concern about the impacts of winter conditions on horse welfare has initiated discussion about the suitability and acceptability of loose housing of horses in a cold, northern cli-mate. Since loose housing provides a useful alternative to stabling with several benefits, e.g. the opportunity for free exercise and social contacts, this housing method should be studied and developed further to enhance the health and welfare of loose-housed horses.
In the late 1990s, the Ylä-Savo Feder-ation of Municipalities for Education founded a development project “Equine Information Centre” in Eastern Finland to advance the Finnish horse industry and cur-rent horse management practices. The Eq-uine Information Centre designed a re-search loose housing system on the school farm of the Ylä-Savo Vocational Institute in Kiuruvesi, where an extensive research pro-ject was conducted in 2002–2006 into the effects of a cold housing environment on horses. The present study, which is based on this research project, aimed to determine the suitability of a loose housing system as a winter environment for weanling horses.


Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 15
2. REVIEW OF THE LITERATURE
2.1 Horse housing alternatives in relation to health and welfare The horse is a herding, free-ranging herbi-vore, which spends most of the day forag-ing (McGreevy, 2004). The herd, which is characterized by a strong social hierarchy, provides safety and social comfort for the horse. As a prey animal, the self-preservation of the horse relies mainly on an open environment that enables early de-tection and fast escape from predators. The possibility to seek shelter and relief from adverse weather conditions, such as storms, wind, cold, heat and sun, is important for the horse (McDonnell, 2003). The herd also contributes to the heat balance of the horse by providing shelter from the adverse ef-fects of weather (Morgan, 1996; Ingólfsdót-tir & Sigurjónsdóttir, 2008).
Current horse housing environments dif-fer to a great extent from the horse’s natural habitat and social environment. In order to ensure the health and welfare of domestic horses, the horse’s natural characteristics should be taken into account in horse man-agement. In addition to behavioural charac-teristics, also nutritional needs, cold resis-tance, type of use, individual features (e.g. health and temperament) as well as general and regional climatic conditions should be considered when selecting a winter housing system for the horse. Since the selection of a housing system, which contributes to horse welfare, is a multidimensional ques-tion, alternatives should be considered from different standpoints.
Horses can be generally classified into three groups according to the type of use: growing horses, mature horses used for various purposes, e.g. recreation, sport or competition, and breeding horses. These groups have unique demands in terms of housing and care. In growing horses, one of
the most important things is to ensure the normal growth and development of the musculoskeletal system. In particular, ad-equate exercise (Raub et al., 1989; Bell et al., 2001; van de Lest et al., 2002; Rogers et al., 2008; van Weeren et al., 2008) and bal-anced nutrition (Glade & Belling, 1984; Glade et al., 1984; Cymbaluk et al., 1989a, 1989b, 1990) are essential. The company of age-mates is important since it may im-prove the sociability (Christensen et al., 2002) and manageability (Søndergaard & Ladewig, 2004) of growing horses.
The most common housing alternatives for the indoor feeding period are stabling in single, loose boxes and loose housing in groups (Zeeb & Schnitzer, 1997). In addi-tion, tethering in stalls is used to some ex-tent in Europe, but, for example, according to the Finnish Ministry of Agriculture and Forestry decree on animal welfare require-ments for horses (14/EEO/1998), tethering should nowadays be avoided. None of these housing options is ideal, with each system having advantages and disadvantages and thus posing different risks to horse welfare (Clarke, 1994; Mills & Clarke, 2002). In the following, the features of the most common housing systems in relation to horse health and welfare are discussed.
2.1.1 Stabling in loose boxes Stabling in loose boxes with restricted turn-out in paddocks has several advantages (Mills & Clarke, 2002). Stabling protects the horse from adverse weather conditions (McGreevy, 2004), and allows each horse to have personal space and a management routine (Mills & Clarke, 2002). The horse’s feed quality and intake, and the amount of exercise can be controlled, and the risk of injuries is low. The manageability of sta-bled young horses has also been reported to

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 16
be better than that of horses in more natural environments due to daily handling (Jezier-ski et al., 1999).
Stabling has several disadvantages re-lated especially to horses’ behavioural needs (Mills & Clarke, 2002). Stabled horses may have rather different time budg-ets than horses in more natural envi-ronments depending on feeding practices (Kiley-Worthington, 1987), and also differ-ent circadian rhythms of behaviour, e.g. sleep patterns (Dallaire, 1986). The most remarkable differences in time budgets are in the time spent eating and standing (Table 1). Stabled horses spend much time stand-ing and less time eating than free-ranging horses due to the limited opportunity for exercise and foraging, which may cause frustration and affect the gastrointestinal function (McGreevy, 2004). Stabled horses also lack an opportunity to interact with other horses (Rivera et al., 2002), which may, for example, increase aggression to-wards other horses (McGreevy, 2004). Chronic frustration and distress, and man-agement and feeding practices associated with stabling may further lead to psy-chological reactions (Mills & Clarke, 2002) in the form of aberrant or unwanted (Hele-ski et al., 2002; Rivera et al., 2002), aggres-sive (Christensen et al., 2002; Søndergaard & Ladewig, 2004) and stereotypic behav-iour, such as weaving, crib-biting, wind-sucking and box-walking (McGreevy et al., 1995; Normando et al., 2002; Waters et al., 2002; Ninomiya et al., 2007; Parker et al.,
2008). The major physical health concern is
respiratory health (Mills & Clarke, 2002). Stabling may predispose the horse to high levels of ammonia, organic dusts and mould spores originating from bedding materials and forages, especially when the ventilation in the stable is poor (Tuomivaara, 1992; Clarke, 1994; Airaksinen, 2006; Hotchkiss et al., 2007). Therefore, stabling has been associated with airway inflammations (Tremblay et al., 1993; Holcombe et al., 2001). In addition, stabling without the op-portunity for exercise may retard the mus-culoskeletal development of growing horses (Raub et al., 1989; Bell et al., 2001; van de Lest et al., 2002; Hiney et al., 2004).
These facts suggest that stabling may cause horses both psychological and phys-ical stress (Mills & Clarke, 2002). How-ever, the negative impacts of stabling could be greatly minimized by management changes, e.g. by avoiding dusty bedding materials and poor-quality forages, and by adequate ventilation (Mills & Clarke, 2002; Hotchkiss et al., 2007).
2.1.2 Loose housing A loose house is typically a group housing system where horses have free access to an unheated sleeping hall with bedding and an outdoor paddock (Ventorp, 1994). Loose housing became common in many parts of Europe, e.g. in Germany (Fader & Sam-braus, 2004) and Sweden (Ventorp, 1994) in the early 1990s, and in Finland in the late
Table 1. Examples of the time budgets of free-ranging and stabled horses (Kiley-Worthington, 1987). Behaviour (%) Free-ranging horses Stabled horses, fed
ad libitum Stabled horses, controlled feeding
Eat 60 47 15 Stand 20 40 65 Lie 10 10 15 Other 10 3 5

Review of the Literature
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 17
1990s. A loose house is considered a more appropriate housing system for horses than a box stable because it is expected to pro-vide a better opportunity to satisfy behav-ioural needs (Bender, 1992; Zeeb & Schnit-zer, 1997). The time budgets of outdoor-housed horses may thus resemble time budgets of feral horses (Heleski et al., 2002). Furthermore, horses have the oppor-tunity to exercise freely, which should en-courage daily locomotor activity (Zeeb & Schnitzer, 1997). Since adequate, regular exercise enhances the development of mus-culoskeletal system (Raub et al., 1989; Bell et al., 2001; van de Lest et al., 2002, 2003; Hiney et al., 2004), loose housing is ex-pected to yield more healthy, athletic horses that are not as prone to strain injuries as stabled horses (Bender, 1992). The lower investment costs as well as the relatively low maintenance costs are also generally considered to be an advantage (Ventorp, 1994) since in Finland, for example, the space requirement per horse is smaller in a loose housing system than in a box stable (Ministry of Agriculture and Forestry, 1998). Space requirement in a loose hous-ing system is 80 % of the area for a loose box for mature horses, 60 % for yearling horses and 40 % for weanling horses.
However, loose housing may also have several shortcomings. For example, a high rate of voluntary exercise in the loose hous-ing system cannot be regarded as self-evident. When paddocks are too small, horses do not exercise enough in order to ensure normal musculoskeletal develop-ment (Hiney et al., 2004) and aggressive behaviour towards other horses may in-crease (Hogan et al., 1988). In addition, the consequences of group housing may be different for horses located differently in the social ranking order, because low-ranking horses may not, for example, be able to rest adequately or at all in a lying position (Fader & Sambraus, 2004) and
may have limited access to food and shelter (Ingólfsdóttir & Sigurjónsdóttir, 2008) due to interference by higher ranking horses. Moreover, the risk of injuries caused by kicking and biting may be substantial (Fürst et al., 2006). Situations such as establish-ment of the ranking order within the group hierarchy, agitation among the horses, and introduction of a new horse into an estab-lished group may provoke kicking and bit-ing. Since horses have a strong social hier-archy (McGreevy, 2004), the smaller space requirements in a loose housing system compared to a box stable may not therefore be adequate and more space per horse may be required, thus diminishing the advantage in the investment costs.
Group housing may also affect horses’ reactions towards humans, since group-housed horses have been observed to seek contact with humans later and to be more difficult to approach than singly-housed horses (Søndergaard & Halekoh, 2003). Therefore, group housing may impair the human-animal relationship if the horses are not adequately handled (Søndergaard & Halekoh, 2003), although it should be noted that contradictory results also exist (Chris-tensen et al., 2002; Søndergaard & Ladewig, 2004).
Outdoor-housed horses are also exposed to fluctuating weather conditions, such as variations in ambient temperature (Ta), pre-cipitation, relative humidity (RH) and wind speed. The cold housing environment in-creases energy requirements of horses (Cymbaluk & Christison, 1989a; Cymba-luk, 1990), and hence also feed intake and feeding costs, but may also indirectly affect the health of the animals (Clark & McAr-thur, 1994). In a recent study, being outside in cold winter weather increased the risk of airway inflammations in horses (Robinson et al., 2006). The longer the horses were kept outdoors in a day, the greater was the risk of airway inflammation. It was specu-

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 18
lated that cooling and drying of the airway mucosa in the cold may be responsible for inflammations. Cold-induced deaths have been reported in only a few cases. Deaths have occurred in horses living under close-to-natural conditions, and have been caused by severe nutritional and environmental stress (Dieterich & Holleman, 1973; van Dierendonck et al., 1996). Generally, the magnitude of the thermal challenge of the environment depends on how the horse can maintain its thermal balance with the aid of its thermal properties and its physiological regulatory mechanism in relation to the physical environment (Morgan et al., 1997).
2.2 General aspects of thermoregula-tion in the cold 2.2.1 Types of thermal responses Thermal responses comprise all those ad-justments of an organism’s anatomy, physi-ology and behaviour that reduce the physio-logical strain caused by the stressful com-ponents of the total environment (Clarke, 1991; IUPS Thermal Commission, 2003). The animal reacts to an immediate change in the environment by acute, instant re-sponses (Young, 1975; Clarke, 1991 in Clarke, 1991), e.g. by activating mecha-nisms to regulate heat loss by vasoconstric-tion, piloerection and behaviour, and heat production (HP) by feed intake and shiver-ing (Young, 1975). The acute responses depend on the animal’s acclimatization (i.e. phenotypic adaptation), which comprises the adjustments at the individual level within the lifetime of the animal in the natural climate (IUPS Thermal Commis-sion, 2003), e.g. hormonal, metabolic and morphological adjustments (Young, 1975). Ultimately, genotypic adaptation, which encompasses the adjustments at the species level and is a result of natural selection, determines the type and magnitude of ther-mal responses by which an individual of a
certain species is able to react (Davenport, 1992). Adaptive changes may appear in behaviour, insulation, metabolism and mor-phology (Young, 1975). Cold adaptation is hence a deep-seated adaptation in the spe-cies (Langlois, 1994).
2.2.2 Body temperature Homeothermic endotherms, such as the horse, maintain body temperature (Tb) within a narrow range despite fluctuations in Ta (Young & Coote, 1973; Cross et al., 1991; Mogg & Pollitt, 1992; Ousey et al., 1992). Endotherms have significant internal sources of heat from metabolic processes, and have anatomical, physiological and behavioural mechanisms to control the heat loss so that a relatively constant Tb is main-tained (Bicego et al., 2007). In mature horses, the normal core Tb is about 38.0 °C (Hines, 2004). Neonatal, rapidly growing, pregnant and lactating animals have a higher Tb (Sjaastad et al., 2003; Hines, 2004). It is considered that a deviation from normothermia of more than 1 °C leads to discomfort, and that a decrease of more than 10 °C or an increase by more than 5 °C is fatal (Langlois, 1994).
Tb is not constant throughout the body (Bligh, 1998). The body core has nearly constant temperature, but the temperature of the outer shell (i.e. skin and subcutaneous adipose tissue) varies considerably depend-ing on the body site and Ta. Generally, sur-face temperatures are lower on body areas with poor vascular supply, with bone un-derneath the skin and at the limbs (Young & Coote, 1973; Palmer, 1983; Mogg & Pollitt, 1992; Verschooten et al., 1997). The fluctuation in surface temperature in direct proportion to Ta is a thermoregulatory mechanism regulated by vasoconstriction or vasodilatation of cutaneous blood vessels (Palmer, 1983; Mogg & Pollitt, 1992).
Tb is regulated by nervous feedback mechanisms, which operate mostly through

Review of the Literature
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 19
temperature-regulating centres located in the hypothalamus (Bicego et al., 2007). The preoptic-anterior hypothalamus, skin and deep tissues, e.g. spinal chord, brain stem and the great veins, contain large numbers of temperature-sensitive neurons that moni-tor Tb (Boulant, 2000). The anterior hypo-thalamus contains large numbers of heat-sensitive neurons (Davenport, 1992; Hines, 2004), whereas peripheral receptors in the skin are most sensitive to cool and cold temperatures (Bicego et al., 2007). The pre-optic region in the hypothalamus acts as coordinating centre that compares and inte-grates information from the central and pe-ripheral neurons (Boulant, 2000).
A defended body temperature is the Tb that the body attempts to maintain (Bligh, 2006). It is thought to arise from the recip-rocal crossing inhibition between the cold-sensitive and warm-sensitive neurons in the central nervous system. Therefore, a bal-ance between the signals from cold-sensitive and warm-sensitive neurons de-termines the thermoregulatory set-level at which Tb is regulated by acute physiological responses, i.e. through heat-production or heat-loss effector pathways. Hence, the temperature difference between the body core and the skin is important in Tb regula-tion (Morgan, 1996).
2.2.3 Heat balance The total heat content of the body deter-mines Tb (Clark & McArthur, 1994). Heat is continuously generated as a by-product of metabolism, and thus Tb depends on the metabolic rate (MR) (Bicego et al., 2007). This metabolic heat is transferred from the deeper tissues to the skin, where it is lost to the air and other surroundings in a con-trolled manner (Morgan, 1997b). Tb re-mains constant when total heat loss, com-prising non-evaporative (radiation, convec-tion and conduction) and evaporative heat loss, equals heat gain, and changes in Tb are
caused by alterations in this ratio (Clark & McArthur, 1994). The following equation (IUPS Thermal Commission, 2003) de-scribes the body heat balance for a standing animal:
Storage of body heat = MR ± evaporation ± convection ± conduction ± radiation
When Tb decreases, the hypothalamus is activated and induces mechanisms which reduce heat loss and increase HP (Boulant, 2000). Heat loss-reducing mechanisms strive to increase body insulation and re-duce the exposed surface area, while heat-producing mechanisms aim at increasing the metabolic HP. When the mechanisms are not sufficient to maintain heat balance, core Tb will fall below the normal level (hypothermia) (IUPS Thermal Commission, 2003).
Allometry, the systematic change in body proportions with increasing body size, has a great effect on heat balance both be-tween and within animal species (Reiss, 1991). Changes within species occur as animals grow and develop but exist also between breeds of species (Reiss, 1991), e.g. in horses (Langlois, 1994). Generally, large body size is an advantage with respect to thermoregulation in the cold, since the ratio of heat-dissipating surface area to heat-producing/retaining body mass de-creases with increasing body size (Phillips & Heath, 1995; Bligh, 1998). Therefore, large animals have less relative surface area available for heat exchange and experience proportionately less heat loss in the cold than small animals. Body size also affects the animal’s MR, which is approximately proportional to body mass by the exponent 0.75 (Bligh, 1998; Cannon & Nedergaard, 2004). Therefore, the mass-specific MR declines as body mass increases. Small animals usually depend on an ability to change the MR to regulate Tb, whereas

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 20
large animals rely on adjustment of surface temperature to regulate heat exchange (Phillips & Heath, 1995; Lovegrove, 2005). In addition to large body size, a spherical body shape reduces the surface area to body mass ratio (Langlois, 1994). According to Allen’s rule, the relative size of exposed portions of the body decreases as Ta de-creases (Davenport, 1992). Accordingly, it is considered that northern types of a given mammalian species have smaller/shorter limbs, tails, ears and/or wings than southern types (Langlois, 1994).
2.2.4 Heat production Heat is mainly produced as a by-product of catabolism – the breakdown of biochemical substrates (carbohydrates, fatty acids and proteins), but there is an inevitable produc-tion of heat from all metabolic processes, i.e. also from anabolic processes (Daven-port, 1992). MR describes the total conver-sion of chemical energy into mechanical work and heat per unit time (IUPS Thermal Commission, 2003). The main ways of generating heat are the basal metabolic rate (BMR), shivering thermogenesis and cold-exposure induced non-shivering ther-mogenesis (NST) (Cannon & Nedergaard, 2004) and physical work, i.e. heat produced in the muscles during exercise (Hodgson et al., 1994). The BMR is the stable rate of energy metabolism measured under condi-tions of minimum environmental and physiological stress, i.e. in a rested, awake, fasting animal in a thermoneutral zone (TNZ) (IUPS Thermal Commission, 2003). The BMR represents the energy expendi-ture needed to maintain cell and organ func-tions (Bligh, 1998). At rest, about 60 % of the body’s HP occurs in the heart, liver, kidneys and brain (Sjaastad et al., 2003). The remaining 40 % is formed in the mus-cles, skin and skeleton.
The hypothalamus regulates the secre-tion of thyroid-stimulating hormone (TSH)
from the pituitary gland, which further regulates the secretion of thyroid hormones, thyroxine and triiodothyronine, from the thyroid gland (Storer et al., 1979). Thyroid hormones raise the cellular metabolism in most body tissues, and thus the BMR is related to thyroid activity (Irvine, 1967). However, an increase in the BMR does not occur instantly after, say, acute cold expo-sure since the thyroid gland must increase in size before reaching a higher level of thyroid hormone secretion. Therefore, in-creased HP by means of thyroid hormones requires prolonged cold exposure (Sjaastad et al., 2003).
During moderate and maximal activity, HP increases greatly (Hodgson et al., 1994). Most of the body’s HP occurs in the mus-cles, since 80 % of the energy used during exercise is given off as heat. For example, in exercising horses, the heat generated was about 2,300 kcal at an exercise intensity of 40 % (38 min) of maximum O2 uptake, and about 4,200 kcal at an exercise intensity of 90 % (9 min) (Hodgson et al., 1993), the power in the latter case being about eight times that in the former. About 7 to 20 % of the heat generated remained as stored heat post-exercise, which consequently in-creased the Tb of the horses. But even minimal muscular activity may increase HP. For example, slight body movements increased the MR of a sleeping foal by about 18 % compared to inactive sleeping (Ousey et al., 1997).
In shivering thermogenesis, heat is rap-idly produced by breaking down ATP in the muscles (Langlois, 1994). Shivering con-sists mainly of involuntary, contractile ac-tivity of skeletal muscles (IUPS Thermal Commission, 2003). As shivering pro-gresses, its intensity may increase from muscle tone to microvibrations and even to visible contractions of both flexor and ex-tensor muscles. Shivering is facilitated by the posterior area of hypothalamus and

Review of the Literature
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 21
suppressed by the preoptic area (Tanaka et al., 2001). Thus, the appearance of shiver-ing depends on the balance between the signals from these two hypothalamic re-gions. Shivering is usually an acute re-sponse to a sudden cold exposure, but may be maintained even several months pro-vided that the animal is able to develop the physical capacity (lung, heart and muscle capacity) necessary for a sustained shiver-ing thermogenesis (Cannon & Nedergaard, 2004). Shivering may increase HP about fourfold as compared to the BMR. On the other hand, body movements associated with shivering may lower the thermal re-sistance of the body and increase heat loss (McArthur, 1991).
In NST, the MR increases during cold exposure without increased muscle activity (IUPS Thermal Commission, 2003). NST is under the control of norepinephrine re-leased from sympathetic nerves (Cannon & Nedergaard, 2004). The principle effector organ is brown adipose tissue (BAT), where most of the energy released as a result of oxidation of glucose and lipids is not util-ized to synthesize ATP as in most body tissues, but is instead dissipated entirely as heat. The capacity for HP by NST increases adaptively in the course of acclimatization to cold (IUPS Thermal Commission, 2003), and replaces shivering in the long term (Cannon & Nedergaard, 2004). BAT exists in mammalian species of smaller body size, e.g. rodents (Klaus et al., 1988; Trayhurn & Jennings, 1988; Haim et al., 1993; Saarela & Hissa, 1993), but is also present in new-borns of larger species, e.g. calves (Alexan-der et al., 1975; Vermorel et al., 1983) and reindeers (Markussen et al., 1985; Soppela et al., 1991, 1992).
2.2.5 Heat loss Heat transfer between an animal and its environment depends on the temperature gradient between the body core and the
surroundings, and the surface area through which transfer takes place (Duncan, 1990). Heat transfer occurs by two main routes: non-evaporative heat loss (dry heat loss) by convection, conduction and thermal radia-tion, and evaporative heat loss (Clark & McArthur, 1994). Heat exchange can be regulated by several mechanisms: changes in circulation, hair coat, exposed surface area, respiration rate and sweating rate (McArthur, 1991; Morgan et al., 1997). In particular, body insulation, which causes the surface temperature to fall near to the level of Ta, efficiently reduces heat loss (Langlois, 1994).
The total body insulation consists of the body tissue, hair coat and air acting in se-ries (McArthur, 1991). Thermal insulation of tissue acts between the body core and skin surface and depends on the thickness of the skin, on the amount of subcutaneous fat and on peripheral blood flow (Ousey et al., 1992). Tissue thermal insulation is regu-lated by vasomotor control of the blood flow to the periphery (Morgan, 1997a). The amount of fat is also important, since fat is three times more insulating than other tis-sues (Guyton, 1991). Coat insulation de-pends on the depth and thickness of the hair layer, the wind speed and the temperature and humidity gradients within the coat (Ousey et al., 1992). The coat insulation is controlled by bristling of the hair with the aid of hair erector muscles (Kainer et al., 1994; Morgan, 1997a). This piloerection, which is an autonomic response to cold, increases the thickness of the insulating, stationary air layer between hairs (Bligh, 1998).
Heat loss by convection occurs between an organism and its external environment in a moving gas or fluid, and depends on the temperature gradient (IUPS Thermal Com-mission, 2003). When the body surface temperature is higher than Ta, air in contact with the skin will be heated by conduction

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 22
and forms a stationary layer a few milli-metres thick (Duncan, 1990). Outside this layer, warm air is moved away from the body by convective currents and replaced by cooler air. This natural convection ac-celerates as the Ta decreases (McArthur, 1991). However, in animals with a hair coat, natural convection is of limited sig-nificance for heat loss, whereas convection due to wind or body movements, i.e. forced convection, is of greater importance (Dav-enport, 1992). Convection also occurs to some extent from the respiratory tract be-cause the exhaled air is usually warmer than the inhaled air (Clark & McArthur, 1994; Morgan, 1997b). It has been estimated that convection by this route is about 3 % of the total heat loss in housed stock (Clark & McArthur, 1994).
Conduction encompasses heat transfer between objects in contact with each others and may occur to solid material, gas or fluid between an organism and its external envi-ronment (IUPS Thermal Commission, 2003). In a cold environment, heat conduc-tion to the air can be considerable since conduction follows the temperature gradi-ent. However, when the temperature of the air layer immediately adjacent to the skin equals the temperature of the skin, no fur-ther heat conduction occurs (Guyton, 1991). In addition to the temperature gradient, heat conduction depends on the surface area of contact and the thermal properties of the object (Langlois, 1994). In a standing ani-mal, only small quantities of heat are lost by conduction to objects, but when the animal is lying, heat conduction can be re-markable (Clark & McArthur, 1994). A wet body surface or lying surface increases conduction since the thermal conductivity of water is high.
In thermal radiation, heat transfer occurs in the form of electromagnetic waves in the infrared part of the spectrum (Danielsson et al., 1998). Also heat loss by radiation fol-
lows the heat gradient, i.e. when the ani-mal’s body is warmer than the environment it loses heat by radiation (Langlois, 1994). It has been estimated that in large animals exposed to slowly moving air, about 50 % of the non-evaporative heat loss will occur by radiation (Clark & McArthur, 1994). Radiative cooling may be high especially at night during the cold winter months (Cym-baluk & Christison, 1989a; Clark & McAr-thur, 1994). When Ta is higher than the skin, the net transfer of heat by radiation is towards the animal, which may increase Tb
(Clark & McArthur, 1994). Animals can reduce heat loss by radiation by standing or lying close to each other so as to reduce the surface area exposed to the external envi-ronment (Bligh, 1998), and by seeking shel-ter (Mejdell & Boe, 2005), and increase absorbed radiation by changing body pos-ture and orientation (Keren & Olson, 2007). For example, solar radiation may effec-tively diminish the cooling effect of low Ta and high wind velocities.
Evaporative heat transfer is associated with the loss of water vapour from the body surface and respiratory system (Clark & McArthur, 1994). Evaporation and its cool-ing effect depend on the difference in water vapour pressure between the skin and Ta, and the exhaled and inhaled air, and on the respiration rate (Morgan, 1997b; Morgan et al., 1997). Evaporation of sweat is a con-trolled and effective mechanism of heat loss when Ta is higher than Tb (Clark & McAr-thur, 1994) and during exercise (Hodgson et al., 1994; Morgan et al., 2002). Evaporation also occurs insensibly (insensible water loss), causing continuous heat loss by evaporation, in the cold too (McArthur, 1991). Insensible water loss cannot be con-trolled for the purposes of thermoregula-tion, since it results from continuous diffu-sion of water molecules through the skin and respiratory surfaces regardless of Tb. At low Ta’s, insensible evaporation is almost

Review of the Literature
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 23
constant and has been observed to encom-pass about 20 % of total heat loss (Morgan, 1996). The proportion of respiratory water loss may be relatively high since inhaled cold air contains little water vapour (Robin-son et al., 2006). Wetting of the hair coat (Ousey et al., 1991; Kohn et al., 1999) and wind (Davenport, 1992) will further in-crease the cooling effect of evaporation. For example, Ousey et al. (1991) calculated that the warm wet surface of a horse at 25 °C loses heat at a rate of about 200 W/m2 by evaporation.
2.2.6 Thermoneutrality and lower critical temperature A general model of the relationship be-tween Ta, Tb, HP and heat loss is presented in a thermoneutral diagram (Fig. 1). How-ever, the true pattern of heat balance is more complex and dynamic because the
actual values and relative positions of the curves depend on the animal’s species, age, plane of nutrition, acclimatization and envi-ronmental factors (Mount, 1973). There-fore, the model presents only general rela-tionships between different quantities and zones. In addition, the zones of the model can be interpreted in a number of ways since they may be neutral in different re-spects. For example, the zone of least ther-moregulatory effort is bounded at the colder limit by rising MR and at the warmer limit by increasing evaporative heat loss, and the zone of minimal metabolism is bounded on each side by rising MR. The zones may also be defined for particular purposes e.g. pre-ferred thermal environment, animal produc-tivity or optimal growth rate.
The TNZ is usually defined as the range of Ta at which a homeothermic endotherm does not have to expend more energy than
Ambient temperature
B
Body temperature
Rat
e of
hea
t pro
duct
ion
or lo
ss
Non-evaporative heat loss
Metabolic heat production
Evaporative heat loss
A F D E C
Figure 1. Diagrammatic representation of the relationship between ambient temperature, heat pro-duction, evaporative and non-evaporative heat loss and body temperature. A = zone of hypother-mia, B = temperature of maximal metabolism, C = lower critical temperature, D = temperature of marked increase in evaporative heat loss, E = temperature of increase in metabolic rate, i.e. upper critical temperature, F = zone of hyperthermia, CD = zone of least thermoregulatory effort, CE = zone of minimal metabolism, i.e. thermoneutral zone (Mount, 1973).

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 24
that required for maintenance metabolism in order to compensate heat loss and to maintain constant Tb (zone of minimal me-tabolism, C–E in Fig. 1) (Mount, 1973; Clark & McArthur, 1994). In this case, thermal balance is achieved by adjusting the rate of non-evaporative and evaporative heat loss through metabolically inexpensive adjustments in the thermal conductance of the body surfaces, including vasomotor responses, i.e. blood flow to the skin, pos-tural changes and insulation adjustments, i.e. the thickness of the insulating air layer within the hair coat. However, the upper limit of the TNZ, i.e. the upper critical tem-perature, has also been defined on the basis of increasing evaporative heat loss (D in Fig. 1) (e.g. IUPS Thermal Commission, 2003). Therefore, the zone C–D is called the zone of least thermoregulatory effort (Mount, 1973). Between C and D, the line of non-evaporative heat loss is undefined since the rate changes as a result of periph-eral vasomotor control to regulate the sur-face temperature. Above zone C–D, the rate of non-evaporative heat loss decreases be-cause it is limited by the small temperature difference between the surface temperature of the horse and Ta (Clark & McArthur, 1994; Morgan 1997b).
At and below the lower limit of the TNZ (C in Fig, 1), the body insulation is maxi-mal (Morgan, 1995; Morgan et al., 1997). The non-evaporative heat loss, which oc-curs through convection, conduction and radiation, increases linearly with decreasing Ta (towards B in Fig. 1) and the BMR is insufficient to balance increasing non-evaporative heat loss (Mount, 1973). There-fore, metabolic HP also has to increase in order to maintain a constant Tb (zone B–C in Fig. 1). Below the lower critical tempera-ture (LCT, point C in Figure 1), heat loss from the body surface in a standing animal occurs mainly by convection and radiation, because conduction is assumed to be negli-
gible (Clark & McArthur, 1994) and evapo-rative heat loss is minimal and constant (Davenport, 1992; Morgan, 1995; Morgan et al., 1997). Both convection and radiation follow the temperature gradient (Langlois, 1994; Randall et al., 2001). However, it has been proposed that in still air conditions and especially in animals with an intact hair coat, convection may be of limited signifi-cance in the cold (Davenport, 1992). The temperature gradient can be reduced by increasing thermal insulation of the body, and consequently the LCT is shifted to lower Ta’s (Young, 1975). Increasing feed intake also lowers the LCT, because HP increases with increasing energy intake (Pagan & Hintz, 1986; Morgan, 1995; Ver-net et al., 1995; McDonald et al., 2002; Morgan et al., 2007). At a certain Ta below the LCT (B in Fig. 1), HP is insufficient to compensate for constantly increasing non-evaporative heat loss, and the animal be-comes hypothermic, i.e. the Tb falls (A in Fig. 1) (Mount, 1973).
Recently, this classical view of the TNZ was elaborated by Arnold et al. (2004), who showed that metabolic HP decreased in red deer (Cervus elaphus) in the night and early morning hours during late winter as a re-sponse to an energetically challenging situation. This nocturnal hypometabolism is a previously unknown thermoregulatory response in large endothermic animals to periods of food shortage and harsh climatic conditions. It contributes to lower energy expenditure by lowering an animal’s de-fended Tb, which leads to reduced BMR and lower HP, meaning that the change in defended Tb causes a corresponding change in the animal’s LCT. Therefore, the TNZ is well defined only for a given set-level of Tb.

Review of the Literature
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 25
2.3 Thermoregulation of horses in the cold 2.3.1 Lower critical temperatures for horses The measured LCT values for horses are summarized in Table 2, which is an updated version of the summary presented in the review article by Cymbaluk (1994). The values vary considerably since many bio-logical and environmental factors affect the LCT value. According to Cymbaluk (1994) and Morgan et al. (2007), age, breed, body condition, size, surface area to body mass ratio, physiological status, MR and acclima-tization of the horse, and also quality of feeds, intensity of feeding, housing system, season and climatic factors affect the LCT value. The summary indicates that the LCT values are mainly determined by age and acclimatization.
Young horses generally have higher LCT values than mature horses (Table 2). Neonatal foals, in particular, have a high LCT, about 20 °C, because their body insu-lation is low and the ability for thermoregu-lation is less developed than in mature horses (Ousey et al., 1991, 1992, 1997). Due to their high LCT, neonatal foals are susceptible to cold after birth. However, there is considerable individual variation in the thermal insulation of neonatal foals which causes noticeable variation in the LCT (range 10 to 26 °C). Sick and prema-ture foals are more susceptible to hypo-thermia than healthy foals, since they have a lower MR and minimal capacity for in-crease in metabolic HP (Ousey et al., 1997). Weanling and yearling horses are more cold-resistant than neonatal foals. The LCT values for cold-housed weanling and year-ling horses vary in studies from –11 °C (Cymbaluk & Christison, 1989a) to 0 °C
(Young & Coote, 1973; Cymbaluk et al., 1989a; Cymbaluk, 1990).
The LCT values for mature horses de-pend mainly on feed intake and the season to which the horses are acclimatized (Table 2). A cold-housed broodmare was reported to have a LCT of –1 ºC in early winter and –9 ºC in late winter (Young & Coote, 1973). The LCT was 5 °C in horses that were acclimatized to an indoor Ta of about 15 °C (Morgan, 1996, 1998; Morgan et al., 1997), and –15 °C in horses that were ac-climatized to outdoor winter weather (McBride et al., 1985). The lower energy intake of the horses in the former study may also partly explain the difference. Minor breed differences have been found, ponies having a higher LCT than Quarter Horses (QH), Standardbred (SB), Thoroughbred (TB) and Warmblood (W) horses. Competi-tion horses have a lower LCT value than maintenance horses due to their higher feed intake (Morgan et al., 2007). Clipping of the hair coat also affects the LCT value, elevating it by about 5 °C.
The LCT values in Table 2 are ap-proximations and generalizations for horses, since there are large individual vari-ations from the estimated values depending upon the factors mentioned at the beginning of this section. Moreover, the majority of these results (Young & Coote, 1973; McBride et al., 1985; Ousey et al., 1991, 1992, 1997; Morgan, 1996, 1997a, 1998; Morgan et al., 1997, 2007) only apply to dry, non-active horses in still-air conditions. Wind and wetting of the hair coat increase LCT values (McBride et al., 1985). There-fore, LCT values for horses outdoors possi-bly differ from the values given. It is also noteworthy that the methods used for as-sessing LCT in the field studies summa-rized in Table 2 varied greatly.

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 26
Cold acclimatization aims to shift the TNZ to lower Ta’s, which is achieved by increasing body insulation (Clarke, 1991) and by sustaining continuous extra HP (Cannon & Nedergaard, 2004). Usually, animals of large body size adjust body insu-lation in order to regulate heat exchange,
and animals of small body size adjust HP (Phillips & Heath, 1995; Cannon & Neder-gaard, 2004; Lovegrove, 2005). The mechanisms of acclimatization and acute responses to cold have been investigated in horses in a number of studies. Major ther-mal mechanisms are concerned with both
Table 2. Lower critical temperatures for horses (modified and updated from Cymbaluk, 1994). LCT = lower critical temperature, A = Arabian horse, BW = body weight, DE = digestible energy, MR = meta-bolic rate, P = Pony, QH = Quarter Horse, SB = Standardbred horse, TB = Thoroughbred horse, W = Warmblood horse, - = not known / studied.
LCT (°C) Age Breed / type Mean Range
Method Feed intake
Exposure type
Reference
TB, P 10, 20 - Calculateda Suckle Acute cold + heater
Ousey et al., 1991
TB, A 24 - Calculateda Suckle/ suppl./ parenteral
Acute cold + heater / rug
Ousey et al., 1997
P 22 16 to 26 MR Suckle Acute cold Ousey et al., 1992
Neonate
P 19 13 to 23.5
MR Suckle Acute cold Ousey et al., 1992
SB 0 - DE intake/ 100 kg BW
Restricted Acclimatized Cymbaluk, 1990
–11 - Gain/feed ratio
Ad libitum Acclimatized Cymbaluk & Christi-son, 1989a
Weanling
QH
0 - DE intake/ 100 kg BW
Ad libitum Acclimatized Cymbaluk et al., 1989a
Yearling and Mature
- - –9 to 0 Calculatedb - Acute cold/ acclimatized
Young & Coote, 1973
QH –15 –20 to –13
MR Restricted Acute cold / acclimatized
McBride et al., 1985
SB 5 - Total heat loss
Restricted Acute cold Morgan, 1996, 1998; Morgan et al., 1997
P - 1.4 to 6.9
TB - –2.1 to 3.4
Mature
W - –3.4 to 2.9
Calculatedc Restricted - Morgan et al., 2007
a based on body core temperature and body resistance, heat production, predicted MR and the volu-metric specific heat of air b based on heat production and rectal, skin and air temperatures c based on body core temperature, heat flow, thermal insulation in tissue, coat and air, and evapora-tive heat loss

Review of the Literature
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 27
producing sufficient body heat and retain-ing it by the means of physiological and behavioural responses. Responses may be daily, seasonal or interannual (Clarke, 1991), and seem to differ to some extent between feral and domestic horses depend-ing on nutritional status and available facili-ties. 2.3.2 Physiological responses In the horse, physiological responses to cold have been studied from many different standpoints. The observed mechanisms and
responses are summarized in Table 3, which is an updated version of the summary originally presented in the review article by Cymbaluk and Christison (1990). The mechanisms related to HP include Tb, heart rate (HR), shivering, respiratory quotient (RQ), feed/energy intake, nutrient digestion and thyroid hormones secretion. Hair coat density and length, piloerection and vaso-constriction are concerned with retaining body heat in the cold. Subcutaneous body fat and respiration rate are related both to HP and heat retention.
Table 3. Physiological responses to cold in domestic and Przewalski* horses (modified and updated from Cymbaluk and Christison, 1990). ↑ = increase, ↓ = decrease, ↔ = no effect. Mechanism Response Reference Metabolic rate ↕↔ Young & Coote, 1973; McBride et al., 1985; Ousey et al.,
1991, 1992, 1997; Arnold et al., 2006* Body temperature ↓ Arnold et al., 2006* Heart rate ↑↓ Ousey et al., 1992, Arnold et al., 2006* Shivering ↑ Irvine, 1967; McBride et al., 1985; Ousey et al., 1991,
1992, 1997; Morgan, 1997b; Morgan et al., 1997; Mejdell & Bøe, 2005
Respiratory quotient ↑ Ousey et al., 1991, 1992, 1997 Feed /energy intake ↕ Cymbaluk & Christison, 1989a; Cymbaluk et al., 1989a;
Cymbaluk, 1990; Arnold et al., 2006*; Kuntz et al., 2006* Weight gain ↓↔ Cymbaluk et al., 1989a; Cymbaluk, 1990 Body condition ↓↑↔ Dieterich & Holleman, 1973; Cymbaluk & Christison,
1989a; Berger et al., 1999*; Mejdell & Bøe, 2005; Kuntz et al., 2006*; Ingólfsdóttir & Sigurjónsdóttir, 2008
Dry matter intake ↓ Kuntz et al., 2006* Gut passage time ↔↑ Cymbaluk, 1990; Kuntz et al., 2006* Fiber digestion ↑ Cymbaluk, 1990 Phosphorus digestion ↓ Cymbaluk, 1990 Dry matter, energy, crude protein, calcium digestion
↔ Cymbaluk, 1990
Water intake ↓ Crowell-Davis et al., 1985; Cymbaluk, 1990; Kristula & McDonnell, 1994
Thyroid hormones secre-tion
↑↔ Irvine, 1967; McBride et al., 1985; Mejdell & Bøe, 2005
Hair density and length ↑↔ McBride et al., 1985; Cymbaluk, 1990; Neste, 2000; Mej-dell & Bøe, 2005
Piloerection ↑ Young & Coote, 1973; Ousey et al., 1992 Vasoconstriction ↑ Palmer, 1983; Mogg & Pollitt, 1992; Morgan, 1997a Respiration rate ↓ Dahl et al., 1987; Morgan, 1997b

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 28
Heat-producing mechanisms Most of the physiological responses to acute or chronic cold observed in the horse (Table 3) are in accordance with general theories of thermoregulation. In mature horses (Young & Coote, 1973; McBride et al., 1985), yearling horses (Young & Coote, 1973) and neonatal foals (Ousey et al., 1991, 1992), the MR was found to increase as a response to acute cold, and in the foals the increase was accompanied by increased HR (Ousey et al., 1992). In cold-acclimatized horses, the MR was the same in early and late winter, probably indicating that mild winter weather did not provide sufficient stimulus to induce a change in the MR, or that acclimatization had already occurred prior to the measurement con-ducted in early winter (McBride et al., 1985). Furthermore, sick and premature neonatal foals are not able to increase metabolic HP which makes them suscepti-ble to hypothermia (Ousey et al., 1997). It is also noteworthy that there may be large individual (Ousey et al., 1992) and breed (Ousey et al., 1991) differences in MR re-sponses to cold.
A reverse metabolic response to cold than that occurring in domestic horses has been recently demonstrated in the Przewal-ski horse (Equus ferus przewalskii), which is an ancestral wild form of the domestic horse (Arnold et al., 2006) (Table 3). As an adaptation to low Ta and to decreased avail-ability and quality of plant forage, Przewal-ski horses living under close-to-natural conditions reduced their energy expenditure during the winter by lowering Tb, which was accompanied by decreased metabolic HP and HR. Decreased Tb also lowers the temperature gradient between the body sur-face and Ta, and hence reduces heat loss. These seasonal changes were under en-dogenous control and prepared the animal in advance for predictable seasonal changes of climate and of availability and quality of
food. Whether such an adaptation mecha-nism occurs in domestic horses is not known.
An increase in metabolism by means of shivering has been observed in domestic horses at low Ta’s (Table 3). Visible shiver-ing occurred in mature horses (McBride et al., 1985; Morgan, 1997b; Morgan et al., 1997) and neonatal foals (Ousey et al., 1991, 1992, 1997) during acute cold expo-sure and in mature horses during long-term cold exposure in rainy weather (Irvine, 1967; Mejdell & Bøe, 2005). In neonatal foals, shivering was accompanied by an increased RQ, which indicated that the foals utilized muscle and liver glycogen as an energy source by shivering (Ousey et al., 1991, 1992, 1997). The results obtained by Ousey et al. (1991, 1992, 1997) on the exis-tence of shivering in neonatal foals support the general assumption that NST does not exist in the horse (see Cymbaluk, 1994). However, the presence of NST has not been studied in the horse (Cymbaluk & Christi-son, 1990; Cymbaluk, 1994). Since NST has been observed in some other large, newborn endothermic animals (calves: Al-exander et al., 1975; Vermorel et al., 1983 and reindeer: Markussen et al., 1985; Sop-pela et al., 1991, 1992), NST would be worth studying in the horse, too.
At Ta’s lower than the LCT, extra en-ergy from feed is needed in order to in-crease metabolic HP (NRC, 2007). This extra demand for feed is called climatic energy demand (CED) (MacCormack & Bruce, 1991). Horses have been reported to need about 0.2 to 2.5 % more energy for maintenance per 1 ºC decrease in Ta below the LCT (Young & Coote, 1973; McBride et al., 1985; Cymbaluk et al., 1989a; Cym-baluk, 1990). Body size is very important feature in determining LCT and CED (AAPCA, 1991; MacCormack & Bruce, 1991; Morgan, 1995). Small-sized horses have higher LCT values and need propor-

Review of the Literature
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 29
tionally more additional feed, because their heat loss in the cold is relatively greater than in larger horses. In addition, good body insulation (Young & Coote, 1973; Ousey et al., 1992; Morgan, 1997a) and high energy intake (Morgan, 1995; Morgan et al., 2007) affect cold resistance and result in a lower LCT. If CED is not taken into account in the plane of feeding, the weight gain of growing horses declines (Cymbaluk et al., 1989a; Cymbaluk, 1990). When the rate of feeding is increased or Ta rises, compensatory growth (i.e. a growth spurt) may occur (NRC, 2007). The CED of a horse can be efficiently reduced by using a rug or by providing a shelter (AAPCA, 1991; MacCormack & Bruce, 1991).
However, the results concerning volun-tary feed and energy intake of domestic horses in the cold are contradictory (Cym-baluk & Christison, 1989a; Cymbaluk et al., 1989a) (Table 3). It seems that horses do not invariably increase feed intake as Ta decreases (NRC, 2007). This may indicate that daily Ta fluctuations under cold condi-tions have little effect on energy intake by cold-acclimatized, adequately fed horses (Cymbaluk & Christison, 1989a), but may also be a consequence of different envi-ronmental or biological factors in the stud-ies, e.g. differences in body insulation, and reflect inter-individual variation in ther-moneutrality. However, feral Przewalski horses have been reported to generally vol-untarily reduce dry matter (DM) and energy intake in late winter (Kuntz et al., 2006). Reduced DM intake suggested a shift to-wards body energy reserves as the major metabolic fuel, which was supported by a remarkable decrease in body condition scores (BCS) of the horses during the win-ter. The shift towards the use of body en-ergy reserves may be an adaptive response to cold when the energetic and time costs of foraging and digestion outweigh the energy intake. Also in other studies of horses living
under close-to-natural conditions, BCS has been found to decrease during the winter (domestic horses: Dieterich & Holleman, 1973; Przewalski horses: Berger et al., 1999; Kuntz et al., 2006). This annual change in BCS does not occur in domestic, outdoor-housed horses when the horses are fed good-quality feeds in sufficient amounts or ad libitum (see Cymbaluk & Christison, 1989a; Mejdell & Bøe, 2005). However, individual differences may occur when horses are group-housed, because higher ranking individuals have been reported to gain weight in the wintertime, while subor-dinates lose weight due to limited access to food (Ingólfsdóttir & Sigurjónsdóttir, 2008).
Cold exposure may affect nutrient di-gestion (Cymbaluk, 1990) (Table 3). Przewalski horses were reported to improve nutrient absorption by increasing gut pas-sage time in late winter (Kuntz et al., 2006). Gut passage time was not related to concen-trations of nutrients in DM, but rather was related to photoperiod. Kuntz et al. (2006) concluded that this mechanism, i.e. an en-dogenous cycle in gut passage time regu-lated by photoperiod, adapts the gut func-tion and hence the digestive strategy to meet the highly predictable seasonal changes in food quality. However, in do-mestic weanling horses, cold housing envi-ronment did not affect feed retention time (Cymbaluk, 1990). Instead, fibre digestibil-ity increased and phosphorus digestibility decreased. It was speculated that increased fibre digestibility may have resulted in higher HP, but cold housing did not affect DM, energy, crude protein or calcium di-gestibility. It seems that a cold housing en-vironment generates an increased demand mainly for energy (NCR, 2007). However, it should be noted that in the study by Cymbaluk (1990), the cold effects on diet digestibility may have been confounded by

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 30
age effects and by the composition and physical characteristics of the diet.
Feed composition may also affect HP, and thus the type of feed may help the horse to cope during cold weather (NRC, 2007). The heat increment (HI) of digestion, i.e. the amount of metabolizable energy (ME) that is converted to heat during the diges-tive processes and the metabolism of ab-sorbed nutrients (Sjaastad et al., 2003), rep-resents 10 to 30 % of the feed ME in the horse (Frape, 1998). The HI increases dur-ing eating (Vernet et al., 1995), with in-creasing feed intake (Pagan & Hintz, 1986; Vernet et al., 1995) and protein content of feed (Sjaastad et al., 2003), and is greater for forages than concentrates (Vermorel et al., 1997; Frape, 1998). Therefore, forages are commonly fed ad libitum for outdoor housed horses to allow them to eat to their energy demands (NRC, 2007).
In cold-housed horses, attention should also be paid to adequate water intake, since horses have been reported to decrease water intake in the cold (Crowell-Davis et al., 1985; Cymbaluk, 1990) (Table 3). Horses also drink less cold than warm water during cold weather (Kristula & McDonnell, 1994). Insufficient water intake predisposes horses to dehydration (Houpt et al., 2000) and may lead to impaction colic (Kristula & McDonnell, 1994).
Endocrinological responses to cold are difficult to interpret, since many factors, e.g. photoperiod and age, breed, body weight (BW), diet carbohydrate content, exercise, gender and pregnancy of the horse (Irvine, 1967; Glade & Reimers, 1985; Cymbaluk, 1994; Fazio et al., 2007), affect the secretion of thyroid hormones. Accord-ingly, the thyroid hormones responses of horses to cold are contradictory (Table 3). In some studies, Ta had no effect on the thyroid hormones secretion rate (McBride et al., 1985; Mejdell & Bøe, 2005), and in some studies the secretion rate increased as
Ta decreased (Irvine, 1967; McBride et al., 1985). This signifies that horses do not in-variably respond to low Ta by increasing thyroid hormones secretion (Cymbaluk, 1994; Mejdell & Bøe, 2005), and, therefore, that thyroid hormones concentrations may not be reflective of acclimatization to cold (McBride et al., 1985) as presented in ani-mal physiology textbooks (e.g. Sjaastad et al., 2003). Accordingly, the thyroid hor-mones responses of horses to cold should be interpreted with caution.
Heat-retaining mechanisms As mentioned above, in addition to being the body’s energy reserve, subcutaneous fat is protective in cold due to its low thermal conductivity and poor blood supply (Dav-enport, 1992). The distribution of fat may be adjusted by cold acclimatization (Lan-glois, 1994). Fat is dispersed more evenly over the body surface instead of being con-centrated in particular areas as in hot condi-tions.
Body heat can also be retained with the aid of increasing hair coat density and length, which is a chronic, slow response to winter caused mainly by photoperiodism (Cymbaluk & Christison, 1990; Cymbaluk, 1990, 1994). Hair growth in horses is cyclic and the winter coat is kept for about nine months (Talukdar et al., 1972). Accord-ingly, hair coat density and length have been reported to increase during the autumn and winter (Cymbaluk, 1990; Neste, 2000; Mejdell & Bøe, 2005) (Table 3). In addition to photoperiod, Ta also seems to affect hair growth. A cold housing environment was noted to produce a higher hair coat weight than a warm housing environment (Young & Coote, 1973; Cymbaluk, 1990), and the winter coat was shed later in cold-housed horses. In cold conditions, the coat may also consist of longer, finer and more poorly medullated fibres than in warm con-ditions, which increases coat insulation

Review of the Literature
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 31
(Scott, 1988). Also hormones and horse breed affect coat growth (Scott, 1988; Cymbaluk, 1994; Cymbaluk & Christison, 1990; Neste, 2000). Therefore, there is con-siderable individual variation in the thermal insulation of the coat (Talukdar, 1972; Ousey et al., 1992), and, for example, ade-quately fed cold-housed horses do not in-evitably grow a thick winter coat (Young & Coote, 1973; McBride et al., 1985). The thermal insulation of the coat is decreased by rapid body movements during shivering (Ousey et al., 1992) and by clipping of the hair coat (Morgan, 1997b; Morgan et al., 2002). Clipping is common especially in competition horses since it facilitates heat dissipation from the body during exercise.
Piloerection of the hair and peripheral vasoconstriction, which increase body insu-lation, are rapid and effective heat-retaining mechanisms (Mogg & Pollitt, 1992; Cym-baluk, 1994). Piloerection has been studied in two studies (Table 3). Piloerection in-creased coat depth by 0.3 to 1.4 cm in neo-natal foals (Ousey et al., 1992) and 10 to 30 % in mature horses (Young & Coote, 1973). Peripheral vasoconstriction, and hence a decrease in surface temperatures, has been generally observed in horses as a response to low Ta (Palmer, 1983; Mogg & Pollitt, 1992; Morgan, 1997a). However, irregular spontaneous increases in surface temperatures were observed at low Ta (Mogg & Pollitt, 1992). This was probably attributed to periodic vasodilatator activity, which may have functioned to warm the peripheral tissues.
Decreased respiration rate has been ob-served in horses during and after exercise in response to acute cold (–25 ºC), which was possibly associated with the need to reduce heat dissipation in the cold (Dahl et al., 1987) (Table 3). Respiration rate was also lower in clipped horses than in horses with an intact hair coat in the cold, which was also interpreted as a response to reduce heat
loss from the respiratory system (Morgan, 1997b). The respiratory minute volume, which is a function of respiration rate and tidal volume, was not measured, but breath-ing of the horses appeared deep and slow in the cold.
2.3.3 Behavioural responses Behavioural changes are important ther-moregulatory mechanisms (Hetem et al., 2007), and therefore play an important role in the ability of the horse to thrive under varying climatic conditions (McDonnell, 2003). Behavioural mechanisms, too, can be divided into heat-producing and heat-retaining mechanisms, and are summarized in Table 4, which is an updated version of the summary originally presented in the review article of Cymbaluk and Christison (1990). Mechanisms related to HP include temporary activity, general activity and eating, and those related to retaining body heat include shelter- and comfort-seeking, general activity and resting. Shelter- and comfort-seeking mechanisms include the seeking of warmer microclimates (Hetem et al., 2007) and the reduction of exposed sur-face area by changing body shape or orien-tation, which encompasses standing with the hindquarters into the wind or rain or into a natural windbreak, basking (resting in warm, sunny areas during cold weather, McDonnell, 2003) and huddling (lying with bodies in contact, Sjaastad et al., 2003) (McDonnell, 2003).
Shelter- and comfort-seeking behaviour has been observed both in domestic (Gun-narsson & Ingvarsson, 1995; Michanek & Ventorp, 1996; Mejdell & Bøe, 2005; Hetem et al., 2007; Ingólfsdóttir & Sigur-jónsdóttir, 2008) and feral (Duncan, 1985) horses in winter (Table 4). In domestic horses, the main motive to use the shelter is probably the need for a comfortable lying place, whereas the need for weather protec-tion is probably a secondary motive

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 32
(Michanek & Ventorp, 1996). In addition to low Ta (Duncan, 1985; Mejdell & Bøe, 2005), also rain and/or wind and drizzle increase the use of shelter (Michanek & Ventorp, 1996; Mejdell & Bøe, 2005; Ingólfsdóttir & Sigurjónsdóttir, 2008). Huddling as a means of retaining body heat has not been extensively studied in the horse, although it has been suggested that, by standing close and exposing the surface areas to each other, the confronting surface areas of the horses have the same tempera-ture, and non-evaporative heat loss de-creases (Morgan, 1996). To my knowledge, only Gunnarsson and Ingvarsson (1995) have mentioned that loose-housed weanling horses tended to lie close to each other, and Ingólfsdóttir and Sigurjónsdóttir (2008) noted that Icelandic outdoor-housed horses formed tight groups when the weather was bad. Basking has also not been studied ex-tensively in the horse, being mentioned in only one study (see Mejdell & Bøe, 2005).
Horses may also modify their time budget in order to adjust themselves to pre-vailing climatic conditions (Langlois, 1994)
(Table 4). Feral horses have been reported to increase the time spent eating and rest-ing, and to reduce locomotor activity in autumn and winter (Duncan 1980, 1985; Berger et al., 1999; Arnold et al., 2006). In Przewalski horses, lower locomotor activity level in winter has recently been proven to be related to lower MR, and hence to lower energy expenditure (Arnold et al., 2006). However, van Dierendonck et al. (1996) reported that despite of a long, severe win-ter, the time budget of Przewalski horses did not vary between seasons. Contradic-tory results in feral horses may be due to differences in sampling procedures and to local availability and quality of food. How-ever, in domestic horses, feeding time and activity did not invariably change in the cold when the horses were acclimatized to the cold and fed sufficiently (see Mejdell & Bøe, 2005). Short-term behavioural changes, e.g restlessness and temporary activity, may also occur in response to acute cold and adverse weather (see McBride et al., 1985; Morgan, 1997b; Jørgensen & Bøe, 2007).
Table 4. Behavioural responses to cold in domestic and Przewalski* horses (modified and updated from Cymbaluk and Christison, 1990). ↑ = increase, ↓ = decrease, ↔ = no effect. Mechanism Response Reference Shelter-seeking
↑ Duncan, 1985; Gunnarsson & Ingvarsson, 1995; Michanek & Ventrop, 1996; Mejdell & Bøe, 2005; Hetem et al., 2007; Ingólfsdóttir & Sigur-jónsdóttir, 2008
Huddling ↑ Gunnarsson & Ingvarsson, 1995; Ingólfsdóttir & Sigurjónsdóttir, 2008 Basking ↑ Mejdell & Bøe, 2005 Temporary activity
↑ McBride et al., 1985; Morgan, 1997b; Jørgensen & Bøe, 2007
Activity ↓↔ Duncan, 1980, 1985; van Dierendonck et al., 1996*; Berger et al., 1999*; Mejdell & Bøe, 2005; Arnold et al., 2006*
Resting ↑↔ Duncan, 1980, 1985; van Dierendonck et al., 1996*; Berger et al., 1999*
Eating ↑↔ Duncan, 1980, 1985; van Dierendonck et al., 1996*; Berger et al., 1999*; Mejdell & Bøe, 2005; Arnold et al., 2006*

Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 33
3. OBJECTIVES
At temperatures colder than the LCT, a horse has to increase its metabolic HP in order to maintain its Tb. Therefore, a cold housing environment may have significant effects on the energy needs of a horse. Loose housing, which is expected to pro-vide a more natural housing environment for weanling horses than traditional box stabling, exposes horses to prolonged peri-ods of low Ta and changing weather condi-tions. Horses attain 60 % of their mature weight and 90 % of their mature height dur-ing the first year of life, i.e. their growth rate is very high during the first winter. Therefore, the following questions can be raised: How much does a cold housing en-vironment increase the energy needs of weanling horses? Is it possible to maintain normal growth rate and body condition in a loose housing system if CED is high? Are some horse breeds more cold resistant than others and thus probably more suitable for loose housing? Do horses acclimatize to a cold housing environment by insulation changes? Do they use behavioural ther-moregulation in the cold? In addition, there is no knowledge as to whether loose hous-ing actually provides horses with an oppor-tunity to satisfy their behavioural needs.
The general objective of this study was to examine the suitability of a loose housing system as a winter environment for wean-ling horses in a cold climate by studying the effects of a cold housing environment on weanling horses. In particular, the effects on behaviour, nutrition, growth and cold resistance were studied. In addition, the aim was to produce information for general counseling and education.
The hypotheses underlying the experi-mental scheme of this study were: 1) The loose housing system was assumed to pro-vide the horses with an opportunity to fulfil their behavioural needs, i.e. their behav-
ioural patterns and circadian rhythm were expected to resemble that of feral and pas-tured horses. In addition, it was hypothe-sized that the horses would be relatively active because of the 24-h opportunity for exercise. 2) It was expected that the horses would use behavioural adjustments to ac-climatize to the cold housing environment. It was also assumed that low Ta would af-fect the radiative heat loss and energy needs of the horses. The horses were expected to grow normally and maintain their BCS when the increased energy need is taken into account in the feeding. 3) It was ex-pected that there would be differences in cold resistance between different horse types.
The more detailed aims of the studies were: • To determine the time budget and circa-
dian rhythm of weanling horses in a loose housing system, and to investigate the effect of weather conditions on be-haviour (I).
• To observe the energy intake and growth of weanling horses in a loose housing system (II).
• To compare the rate of radiative heat loss and thermal insulation of different horse types during different seasons (III).
• To evaluate the thermoregulatory capac-ity of cold-acclimatized, loose-housed weanling horses by determining their LCT (IV).
• To measure horse activity under differ-ent management environments (U1). This examination was conducted in or-der to assess the suitability of global po-sitioning system (GPS) technology for recording horse activity, and to verify the activity results of study I.
• To complete the measurements of ther-mal insulation done in study III (U2).


Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 35
4. MATERIALS AND METHODS
This section presents concise versions of the materials and methods described more fully in the original articles (I-IV). The ma-terials and methods of the previously un-published studies (U1 and U2) are described in more detail.
4.1 Animals and housing The studies were conducted at the Voca-tional Institute of Ylä-Savo, in Kiuruvesi, Finland over the years 1998–2000 (III) and 2002–2006 (I, II, IV, U1, U2) (Table 5). The horses in the experiments were mainly weanling horses, but also foals, yearling horses and mature horses were used. The horses were classified into four types ac-cording to breed: light (L), warmblood (W), coldblood (C) and pony (P) types. The horses of type L were SB horses, type W were Danish and Polish warmblood horses,
type C were Finnish coldblood horses (FC) (i.e. Finnhorses) and type P were Conne-mara and New Forest ponies.
The horses were housed in an experi-mental loose housing/paddock system (I, II, IV, U1), in a conventional stable in single, loose boxes (III, U2), in a group box/paddock system (U1) or kept in a pas-ture (U1) (Table 5). The unheated, insulated loose house consisted of two identical sleeping halls (45 m2), entrance shelters (8.5 m2) and paddocks (0.08 ha) (described in more detail in I). The weanling horses were weaned and brought to the loose hous-ing system in October. The horses had free access to the sleeping hall and paddock. In the daytime, the horses were also able to use a large paddock (0.64 ha). There was natural ventilation in the halls and the bed-ding used was a mixture of peat and straw.
Table 5. Summary of the animals and facilities in the studies. C = coldblood horse type, L = light horse type, P = pony type, W = warmblood horse type.
Study
I, II III IV U1 U2
Year 2002–2003 1998–2000 2003–2004 2005–2006 2002
Season Winter Late summer / autumn / winter
Winter Winter / summer
Winter
n 10 18 9 4 16
Horse type (♂+♀)
L (4+3) C (1+2)
L (4+0) C (4+0) W (4+0) P (3+3)
L (3+3) C (1+3)
L (0+1) C (1+2)
L (2+2) C (2+2) W (4+0) P (2+2)
Age Weanling Mature Weanling Foal Weanling Yearling
Mature
Management conditions
Loose house + paddock
Box stable + paddock
Loose house + paddock
1) Loose house + paddock 2) Group box + paddock 3) Pasture
Box stable + paddock

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 36
Ta in the sleeping halls was on average 4 ºC higher than the outdoor Ta. The horses housed in the box stable were kept out in the paddock for a maximum of 10 h per day (III, U2). The horses in the pasture were kept out for 24 h per day (U1).
The indoor and outdoor Ta (I, III, IV, U1), RH (I, IV, U1) (HM34C, Vaisala Oyj, Helsinki, Finland) and outdoor wind speed (I, U1) (SKYWATCH® Meteos, JDC Elec-tronic SA, Yverdon-les-Bains, Switzerland) of the study site were measured on a regular basis during the experimental days (see Table 2 in I, Table 3 in IV). Cloudiness and 24-h precipitation were also determined (I, U1) (see Table 2 in I). In study II, daily Ta’s were obtained from the meteorological re-cording station at Vieremä (Finnish Me-teorological Institute), located about 30 km from the study site (see Fig. 2 in II).
In study U1, average Ta (±SD) during the experimental days in the summer was 20±2 °C during the measurements con-ducted on the foals, and 18±3 °C during the measurements conducted on the yearling horses. In the winter, Ta on the experimen-tal days was –6±3 °C during the measure-ments conducted on the weanling horses in the loose housing system, and –3±2 °C dur-ing the measurements conducted on the weanling horses in the group box/paddock system. The sky was partly clouded and the weather was mainly dry, except for a few short snowfalls during the winter examina-tions.
4.2 Animal care The weanling horses housed in the loose housing system (I, II, IV, U1) were fed timothy hay ad libitum. Hay was used as the main forage because it was easy to han-dle and did not freeze in the winter. Hay was placed in off-the-ground feeding troughs (I, II) or in feeding enclosures (IV, U1) outside the sleeping halls. Hay was supplemented individually with controlled
amounts of silage (I, II) and concentrates (protein, concentrates, and vitamin and mineral supplements) (I, II, IV, U1) in order to ensure adequate nutrient intake. Silage was fed twice a day at 06:00 and 16:00, and concentrates three times a day at 06:00, 10:30 and 16:00. The horses were tied to a wall beside feed buckets inside the halls during the feeding of concentrates and si-lage. Salt blocks and automatic water troughs were located inside the halls. The horses housed in the box stable (III, U2) were fed four times a day. Hay was fed at 07:00 and 10:00 when the horses were out-doors in the paddock, and in the stable at 16:00 and 20:00. Concentrates were fed in the stable at 06:00 and 16:00. The horses in the pasture had free access to water, salt blocks and mineral supplement (U1). The feeds used in the studies were conventional horse feeds used at the Vocational institute.
All the horses had been vaccinated against influenza and treated regularly against internal parasites. The rectal Tb of the horses was checked every morning ex-cept when the horses were in the pasture.
4.3 Behavioural measurements The time budget, circadian rhythm and ef-fect of weather conditions on horse behav-iour in the loose housing system were in-vestigated (I). The horses were housed in two groups (five horses in each group), which spent the evenings and nights (16:00–06:00) separated on their own sides of the facility. Both groups had free access to a large paddock between the hours of 06:00 and 16:00 and were then able to use both sides of the facility. In the results, the groups were combined. The behaviour of the horses was observed from December to March. Observations, a total of 23 24-h periods, were performed by video recording (evenings and nights) and by direct visual observations (daytime). The maximum number of observational days in a month

Materials and Methods
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 37
was ten days and the minimum three days. The locations of the horses in the loose housing system were noted every 15 min along with the behaviour performed at that time (instantaneous sampling, Martin & Bateson, 1993). The behaviours recorded were resting, eating and activity, which were further classified in more detail (rest-ing: standing or lying with or without hud-dling; eating: eating hay, concen-trates/silage or drinking; activity: standing, walking, running, playing, other) (for defi-nitions see Table 1 in I).
The daily distance travelled by the horses under different management condi-tions was measured using GPS technology (U1). Mobile software (MapHit© Sport, Karttaikkuna Oy, Sulkava, Finland) was installed in mobile phones (Nokia 3230, Nokia Oyj, Espoo, Finland). GPS receivers (BT GPS X-MINI, RoyalTek Company Ltd., Tao Yuan City, Taiwan) and mobile phones were placed in capsules that were attached to the horses’ halters (weanling and yearling horses) or to harnesses that were fitted on the horses’ back (foals). The horses were accustomed to carrying the devices on their halters or on their backs. The mobile software measured the distance (km) that the horses travelled as well as the average speed (km/h) in six-hour periods starting at 10:00. The measurements were conducted as follows:
1. Three-month-old foals in pasture in the summer (n=2, 4 d)
2. Yearling horses in pasture in the summer (n=2, 7 d)
3. Weanling horses in a loose housing system in the winter (n=4, 6 d)
4. Weanling horses in a group box (nights) and paddock (daytime) in the winter (n=2, 5 d)
The examinations were conducted using two horses, but altogether there were three foals in the pasture (1.4 ha) with their dams in the first examination (U1). In the second
examination, there were a total of three yearling horses and three mature horses in the pasture (1.3 ha). In the third examina-tion, there were four weanling horses in the loose housing system with 24-h access to a paddock (0.08 ha) and daytime access to a large paddock (0.64 ha). In the last exami-nation, three weanling horses were housed indoors in the group box (45 m2) during the nights (16:00–06:00) and activity was measured in the daytime (07:00–16:00), when the horses had free access to a large paddock (0.64 ha).
4.4 Energy intake and growth The effect of a cold loose housing environ-ment on the ME intake and growth of the weanling horses was studied between No-vember and March (II). The energy need was determined on the basis of the per-formance of the horses during the experi-ment. ME intake was compared to the pre-diction of energy intake for 6-12-month-old horses expected to reach a mature BW of 500 kg, which was based on the MTT 2006 and SLU 2004 requirements. The LCT of the weanling horses was determined on the basis of the ME requirements for horses with a specific thermal resistance (moderate BCS and winter hair coat).
The feeds used in study II are described in detail in Table 6. The nutrient compo-sitions of hay, silage and oats were ana-lysed by the regional laboratory of Valio Oy (Lapinlahti, Finland) by the standard NIR-method (Nousiainen et al., 2003). The nutrient compositions of protein (Racing Protein, Suomen Rehu Oy, Helsinki, Fin-land) and concentrate (Mella leseleike, Suomen Rehu Oy, Helsinki, Finland) sup-plement were analysed by the laboratory of Suomen Rehu Oy (Turku/Seinäjoki, Fin-land) by the standard methods (Ministry of Agriculture and Forestry, 2006). The la-boratories determined the feed values of the forages and concentrates by using MTT

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 38
(2006) formulas and feed tables. The nutri-ent composition of mineral and vitamin supplement (Steel Joint, Oy Steel Joint Ltd, Lahti, Finland) was analysed by Outo-kumpu Mining Services (Outokumpu Oyj, Espoo, Finland) by the standard x-ray fluo-rescence spectroscopy (XRF) method.
At the beginning of the study, the horses were given a sufficient diet to supply the nutrient requirements (MTT, 2006) (II). Group hay intake was daily determined by measuring hay refusals every morning be-fore the subsequent distribution. The amounts of silage and concentrates fed were adjusted according to voluntary hay intake; the nutrients received from hay and the amounts of other feeds given were calcu-lated so that the nutrient requirements of ME, digestible crude protein, calcium, phosphorus and lysine were met. Refusals of silage and concentrates were determined and deducted from consumption.
The BCS of the horses was assessed once a week on a subjective scale from one (poor) to nine (extremely fat) (Henneke et al., 1983) (II). Moderate BCS, i.e. a score of 5, was regarded as an indicator of ad-equate energy intake and was the target score. Thus, ME intake was adjusted indi-vidually according to BCS of the horses by
changing the amount of silage and/or con-centrates fed. Nutrient intake was calculated using Hopti software (Equine Information Centre, Kuopio, Finland). During the ex-periment, the diets were revised on average twice a month. At the same time, the in-takes of other nutrients (digestible crude protein, calcium, phosphorus and lysine) were adjusted so that at least minimum nu-trient requirements were met.
The horses were measured weekly for BW and wither height (II). In addition, the average daily gain (ADG) was determined. The wither height measurements were started on the first day of the study and the BW measurements in December due to technical problems with the weighing scale. The growth of the SB horses was compared with the growth data presented in Sandgren et al. (1993), and the growth of the FC horses with the growth data presented in Saastamoinen and Koskinen (1993), and Ahtila and Saastamoinen (2005).
4.5 Thermographic examinations In the thermographic examinations, the cold resistance of different horse types (III) and the thermoregulatory capacity of cold-acclimatized weanling horses (IV) were evaluated on the basis of radiative heat loss.
Table 6. Nutrient composition and feed values of feeds used in study II (dry matter basis).
Hay Silage Oats Protein supplement
Concentrate supplement1
Mineral and vita-min supplement
Dry matter (g/kg) 830 600 860 890 880 850
Metabolizable energy (MJ/kg)
9.0 9.8 12.3 13.1 14.6
Digestible crude protein (g/kg)
93 130 126 202 100
Calcium (g/kg) 3.0 7.6 0.8 11.2 4.6 164.7
Phosphorus (g/kg)
2.4 3.2 3.5 4.5 1.0 0.4
1 Consisting of wheat bran, sugar beet and molasses

Materials and Methods
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 39
Mature horses were used in the examination of cold resistance of different horse types instead of weanling horses in order to ex-clude the effect of growth on the results (III).
The thermographic examinations in the mature horses were carried out during dif-ferent seasons: late summer, autumn and winter (III). The horses represented four different types: L (BW 459±69 kg, mean±SD), W (604±25 kg), C (548±21 kg) and P (323±27 kg). The examinations were conducted in August (15 °C) and October (2 °C) with all horse types, in March (–12 °C) with type P, and a year later in March (–8 °C) with types L, W and C. Thermal images of the weanling horses were taken at 0 °C, –9 °C, –16 °C and –23 °C during the winter (IV). The examinations were per-formed in stationary conditions in a cold indoor riding arena (III) or storage area (IV), which protected the horses from wind, sun and rain. The Ta of the riding arena and
the storage area was the same as the out-door Ta, to which the horses were acclima-tized.
Thermal images were taken with an In-frametrics 600 camera (FLIR Systems, Inc., Portland, OR, USA) (III) and a ThermaCam PM595 (FLIR Systems, Inc., Portland, OR, USA) (IV). A general view was taken from the non-mane side of the horses at a dis-tance of 8 m. The images were analysed at Satakunta Polytechnic (Pori, Finland). The rate of radiative heat loss (W/m2) was cal-culated from the trunk and neck (Fig. 2) by the following equation:
where, P = net emitted power (W/m2), ε = emissivity 0.95, σ = Stefan-Boltzmann con-stant 5.67 x 10-8 Wm-2K-4, Ts = surface temperature (K), Ta = ambient temperature (K), ― = average of measuring points.
Thermal insulation of the body was
___ P = ε σ (Ts
4 – Ta
4)
Figure 2. A thermographic image of a weanling horse at a temperature of –16 °C. The areas of the neck and trunk where the rate of radiative heat loss was determined are outlined (III, IV).

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 40
evaluated at the same time as the thermo-graphic examinations (III, IV), because thermal insulation affects radiative heat loss. Thermal insulation was assessed on the basis of BCS (see details above), which indicates the amount of subcutaneous fat in the body, and hair coat weight of the horses. Hair samples were taken from the mid-neck about 5 cm below the base of the mane using a coat clipper (Oster Golden A5®, Oster Professional Products, McMinnville, TN, USA). The sample area was determined and the samples were oven-dried at 40 °C for 12 h and weighed. The hair weight per area unit was calculated. For the mature horses (III), complementary hair weight measurements were conducted three years later in December in order to obtain a complete hair growth curve for horses in winter (U2) (Table 5). Some of the horses used for the complementary meas-urements were the same as those used for the thermographic examinations (III). The horses that were no longer available were replaced by horses representing the same horse type.
4.6 Statistics and the presentation of results The statistical software SPSS 10.0 (III), 11.0 (IV) and 14.0 (I, II) for Windows (SPSS Inc.) were used for the statistical analyses. In addition to the descriptive be-havioural results in study I, the effects of Ta, RH, wind speed, precipitation and cloudiness on behavioural parameters were analyzed by Linear Regression.
Average weekly nutrient intakes (ME, digestible crude protein, calcium and phos-phorus) of the weanling horses were com-pared with nutrient requirements by the One-Sample t-Test (II). BCS was compared with the target BCS by the Wilcoxon Signed-Rank test. Since the comparisons were made separately for repeated meas-urements, the p-values were multiplied by
the Bonferroni correction. The effect of time and Ta on total (MJ/d) and mass-specific (MJ/100 kg BW/d) ME intake was tested using the Linear Mixed Model (LMM) (see materials & methods in II for statistical models), where Ta was included as a continuous or categorical variable. The latter analysis with Ta’s in two categories, warm (Ta > –11 °C) and cold (Ta < –11 °C), was performed to compare my own results to Cymbaluk and Christison’s (1989a) es-timation of LCT (–11 °C) for light horse weanlings. Their horses were kept under comparable climatic and management con-ditions, and the LCT was also defined on the basis of feed intake. It should be noted that, since the horses in my study were group housed, daily hay intake values were calculated from group hay intake and were uniform for all the horses. Variation in total nutrient intakes (consisting of nutrient in-takes from hay, silage and concentrates) is therefore slightly underestimated.
The differences in the rate of radiative heat loss and hair weight between different horse types during and between different seasons were tested using the Kruskall-Wallis and Mann-Whitney U tests (III). The differences in the radiative heat loss from the various body regions during and be-tween seasons were tested using the Fried-man and Wilcoxon signed-rank tests. The effect of Ta on the rate of radiative heat loss of weanling horses and the differences in radiative heat loss from the various body regions at different Ta’s were tested using the LMM (IV). The model calculated esti-mated marginal means of the radiative heat loss (±SE), where the effect of time-dependent covariates (metabolic BW [kg0.75] and hair weight) was standardized. The effects of metabolic BW, BCS and hair weight on the rate of radiative heat loss were tested using Linear Regression. In the results section of this thesis, the original values of the rate of radiative heat loss

Materials and Methods
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 41
(mean±SD) (IV) are used in Fig. 7, i.e. the effects of time-dependent covariates are not standardized, since it enables the compari-son of the radiative heat loss of the wean-ling horses (IV) with that of the mature horses (III).
The results of the activity measurements (U1) were not tested statistically because of the small number of horses in the measure-ments. The daily average distance travelled and speed were calculated.


Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 43
5. RESULTS
This section presents a summary of the main results described more fully in the original articles (I-IV). New syntheses of the results are also included. The results of the previously unpublished studies (U1 and U2) are presented in more detail.
5.1 Behavioural patterns of horses 5.1.1 Time budget and circadian rhythm The horses spent most of the day, about 37 %, on ingestive behavioural patterns (eating hay, concentrates, silage or drinking) (Fig. 3) (I). The remaining time was divided equally between resting (standing or lying) and being active (standing, walking, run-ning, playing, other). The horses rested mostly in a lying position (with or without huddling). Most of the activity was stand-ing; the percentage of locomotor behaviour was about 5 % of the observations. The frequency of major behavioural states, in
descending order, were: eating, resting, standing and moving.
Resting was most common at night, es-pecially between the hours of 01:00 and 04:00, but the horses also rested in the day-time, especially between the hours of 12:00 and 14:00 (Fig. 4) (I). Resting occurred mostly inside the loose house both in the daytime and at night, but the horses also rested to some extent outdoors in the pad-dock and in the entrance shelter. Activity (includes standing) was most common in the morning and afternoon, starting to di-minish after 14:00 and increasing again after 05:00. The horses ate hay throughout the day, but hay eating was most common in the daytime between the hours of 8:00 and 16:00, and in the evening between the hours of 18:00 and 22:00. Overall, the horses spent a total of 43 % of the observa-tions inside the loose house, 51 % in the outdoor paddock and 5 % in the entrance shelter.
Figure 3. The 24-h time budget of the weanling horses in a loose housing system from December to March (I). The behavioural classes are mutually exclusive.
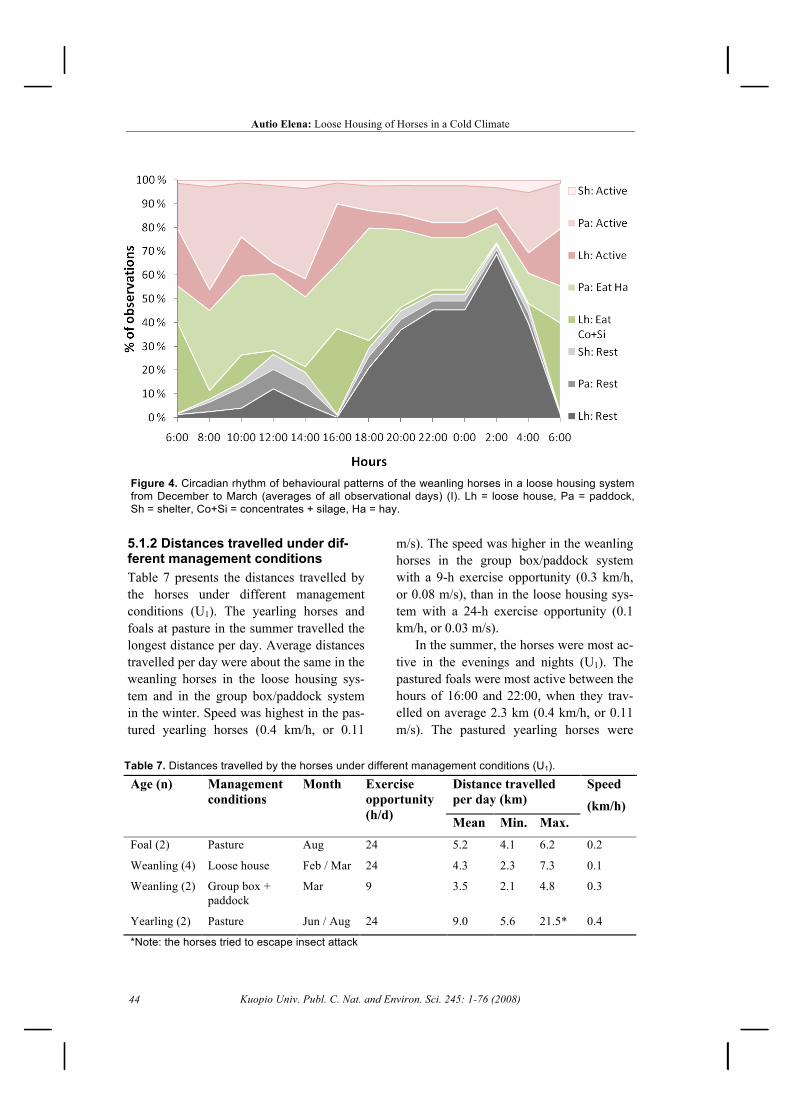
Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 44
5.1.2 Distances travelled under dif-ferent management conditions Table 7 presents the distances travelled by the horses under different management conditions (U1). The yearling horses and foals at pasture in the summer travelled the longest distance per day. Average distances travelled per day were about the same in the weanling horses in the loose housing sys-tem and in the group box/paddock system in the winter. Speed was highest in the pas-tured yearling horses (0.4 km/h, or 0.11
m/s). The speed was higher in the weanling horses in the group box/paddock system with a 9-h exercise opportunity (0.3 km/h, or 0.08 m/s), than in the loose housing sys-tem with a 24-h exercise opportunity (0.1 km/h, or 0.03 m/s).
In the summer, the horses were most ac-tive in the evenings and nights (U1). The pastured foals were most active between the hours of 16:00 and 22:00, when they trav-elled on average 2.3 km (0.4 km/h, or 0.11 m/s). The pastured yearling horses were
Table 7. Distances travelled by the horses under different management conditions (U1).
Distance travelled per day (km)
Age (n) Management conditions
Month Exercise opportunity (h/d) Mean Min. Max.
Speed (km/h)
Foal (2) Pasture Aug 24 5.2 4.1 6.2 0.2
Weanling (4) Loose house Feb / Mar 24 4.3 2.3 7.3 0.1
Weanling (2) Group box + paddock
Mar 9 3.5 2.1 4.8 0.3
Yearling (2) Pasture Jun / Aug 24 9.0 5.6 21.5* 0.4
*Note: the horses tried to escape insect attack
Figure 4. Circadian rhythm of behavioural patterns of the weanling horses in a loose housing system from December to March (averages of all observational days) (I). Lh = loose house, Pa = paddock, Sh = shelter, Co+Si = concentrates + silage, Ha = hay.

Results
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 45
most active between the hours of 22:00 and 04:00, when they travelled on average 2.9 km (0.5 km/h, or 0.14 m/s). In the winter, the horses were most active in the daytime between the hours of 10:00 and 16:00. At that time, the weanling horses in the loose housing system travelled on average 2.1 km (0.35 km/h, or 0.1 m/s), and in the group box/paddock system 2.3 km (0.4 km/h, or
0.11 m/s).
5.2 Effects of a cold housing envi-ronment on horses 5.2.1 Behaviour Ta did not correlate with the time spent ly-ing with huddling (R2=0.00, p=0.86, Linear Regression) or without huddling (R2=0.08,
Figure 5. The effects of relative humidity (RH) (a–c), precipitation (d) and wind speed (e, f) on the behaviour of the weanling horses in a loose housing system from December to March (I).

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 46
p=0.19), eating hay (R2=0.03, p=0.41) or moving (R2=0.08, p=0.19) (I). The time spent inside the loose house showed a slightly increasing trend (R2=0.16, p=0.06) and total activity a slightly decreasing trend (R2=0.16, p=0.06) with decreasing Ta. In-stead, the behaviour of the horses correlated with RH, precipitation, wind speed and cloudiness (Fig. 5). There was a decrease in the time spent eating hay with increasing RH (R2=0.30, p<0.01) (Fig. 5a) and cloudi-ness (R2=0.25, p<0.05). Correspondingly, the time spent standing increased with in-creasing RH (R2=0.22, p<0.05) (Fig. 5b) and cloudiness (R2=0.18, p<0.05). Also the time spent inside the shelter increased with increasing RH (R2=0.17, p<0.05) (Fig. 5c), cloudiness (R2=0.20, p<0.05), precipitation (R2=0.21, p<0.05) (Fig. 5d) and wind speed (R2=0.43, p=0.01) (Fig. 5e). Increasing wind speed also increased the time spent moving (R2=0.18, p<0.05) (Fig. 5f). No signs of visible shivering were observed during study I. 5.2.2 Energy intake and growth The average daily voluntary hay intake of the horses was 4.4±1.2 kg DM (mean±SD); it increased during the study being 3.7±1.1 kg DM in November, 4.0±0.8 kg DM in
December, 4.3±0.9 kg DM in January, 4.3±0.9 kg DM in February and 5.6±1.1 kg DM in March (II). Silage was fed on aver-age 1.6±0.4 kg DM, oats 1.0±0.2 kg DM, protein supplement 0.5±0.2 kg DM, con-centrate supplement 0.09±0 kg DM and mineral and vitamin supplement 0.01±0.01 kg DM per day. The average for-age:concentrate ratio was 80:20 (DM basis).
The ME intake of the weanling horses (75.5±11.8 MJ/d, mean±SD) was on aver-age 30 % above the MTT (2006) require-ments and SLU (2004) requirements for slow growth, and 15 % above the SLU (2004) requirements for rapid growth (see Fig. 1 in II). Ta increased ME intake by 0.5 % per 1 °C increase in Ta during the whole study period (p<0.001, LMM), but ME in-take (y) varied in a non-linear fashion in the course of the winter: y = 0.086x2 –0.902x + 71.5, where x is weeks from November to March (p<0.001, R2=0.63). In early winter, ME intake increased in November by 1.8 % (p<0.001), in December by 0.5 % (p<0.001) and in January by 0.2 % (p<0.05) per 1 °C decrease in Ta (Table 8). Mass-specific ME intake (MJ/100 kg BW) increased in December and January by 0.3 % (p<0.01) per 1 °C decrease in Ta. ME intake was higher, 16.1 % in November and 6.8 % in
Table 8. Mean body weight (BW), estimated surface area, metabolizable energy (ME) intake, calculated heat production (HP), the increase in ME intake per 1 ºC change in ambient temperature (Ta) and cli-matic energy demand (CED) of the weanling horses during the winter (II). Nov Dec Jan Feb Mar BW (kg) - 273.8 296.7 314.6 328.7 Surface area (m2)1 - 3.3 3.5 3.6 3.7 ME intake (MJ/d) 68.0 74.4 71.6 74.0 85.9 ME intake (W/m2) - 262.5 238.8 237.3 267.3 HP (W/m2)2 - 253.6 230.7 229.2 258.2 ME intake (MJ/d) per 1 ºC decrease or increase* in Ta
1.1 0.3 0.1 0.4* 0.8*
CED (W/m2) per 1 ºC decrease or increase* in Ta
- 1.0 0.3 1.2* 2.6*
1 Estimated by the equation A = 1.09+0.008*BW (kg) (Hodgson et al., 1993) 2 Ratio of HP to ME 0.97 was estimated by the equation HP/ME = 1.2–0.649*ME (MJ/kg0.75) (Morgan, 1996)

Results
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 47
December, at Ta’s below –11 °C than at Ta’s above –11 °C (p<0.001). Mass-specific ME intake was also higher at Ta’s below –11 °C than at Ta’s above –11 °C, about 6.3 % and 4.4 % in December and January, respectively. The calculated CED was about 1.0 W/m2 in December and 0.3 W/m2 in January per 1 ºC decrease in Ta.
In late winter, total ME intake increased in February by 0.7 % and in March by 1.3 % per 1 °C increase in Ta (Table 8) (II). Mass-specific ME intake increased in Feb-ruary by 0.3 % (p<0.01) per 1 °C increase in Ta (p<0.001). In late winter, the BCS of the horses rose permanently above the tar-get BCS (see Fig. 3 in II). The calculated CED was about 1.2 W/m2 and 2.6 W/m2 per 1 ºC increase in Ta in February and March (Table 8). The calculated rate of HP fol-lowed ME intake by the ratio 0.97.
As the feed and ME intake of the horses increased during the winter, so intakes of other nutrients such as digestible crude pro-tein, calcium and phosphorus also in-creased, and hence were highest in late win-ter (II). Digestible crude protein intake (848±128 g/d, mean±SD) exceeded the MTT (2006) requirements (500 g/d) by about 70 % (p<0.001) and the SLU (2004) requirements for slow growth (age 3-6 months: 610 g/d; age 7-12 months: 493 g/d) by about 64 % (p<0.05) and for rapid growth (610 vs. 587 g/d) by about 44 %
(p<0.05) (One-Sample t-Test). Also cal-cium intake (43±8 g/d) was on average 30 % above the MTT requirements (33 g/d) (p<0.05) and 39 % above the SLU require-ments for slow growth (37 vs. 29 g/d) (p<0.001) and 21 % above the requirements for rapid growth (37 vs. 35 g/d) differing significantly from week 8 onwards (p<0.05). Phosphorus intake (25±5 g/d) was on average 13 % above the MTT require-ments (22 g/d) (p<0.05) and 37 % above the SLU requirements for slow growth (21 vs. 17 g/d) (p<0.001) and 18 % above the re-quirements for rapid growth (21 g/d) (p<0.05). The magnitude of the excess in-take of ME (R2=0.71, p<0.001, Linear Re-gression), digestible crude protein (R2=0.71, p<0.001) and calcium (R2=0.68, p<0.001) increased with increasing volun-tary hay intake (Fig. 6).
The average BW of the SB horses dur-ing the study was about the same as that presented for SB horses in Sandgren et al. (1993) (see Fig. 4a in II). The FC horses weighed about 19 % more than the FC horses in Saastamoinen and Koskinen (1993), and Ahtila and Saastamoinen (2005) (see Fig. 4b in II). The ADG of the SB horses was 0.57 kg and FC horses 0.62 kg during the study. The ADG correlated negatively with increasing Ta (y = –0.01x+0.50, R2=0.15, p<0.05, Linear Regression).

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 48
Figure 6. Excess nutrient intakes in relation to the voluntary hay intake of the weanling horses from November to March (II). ME = metabolizable energy.
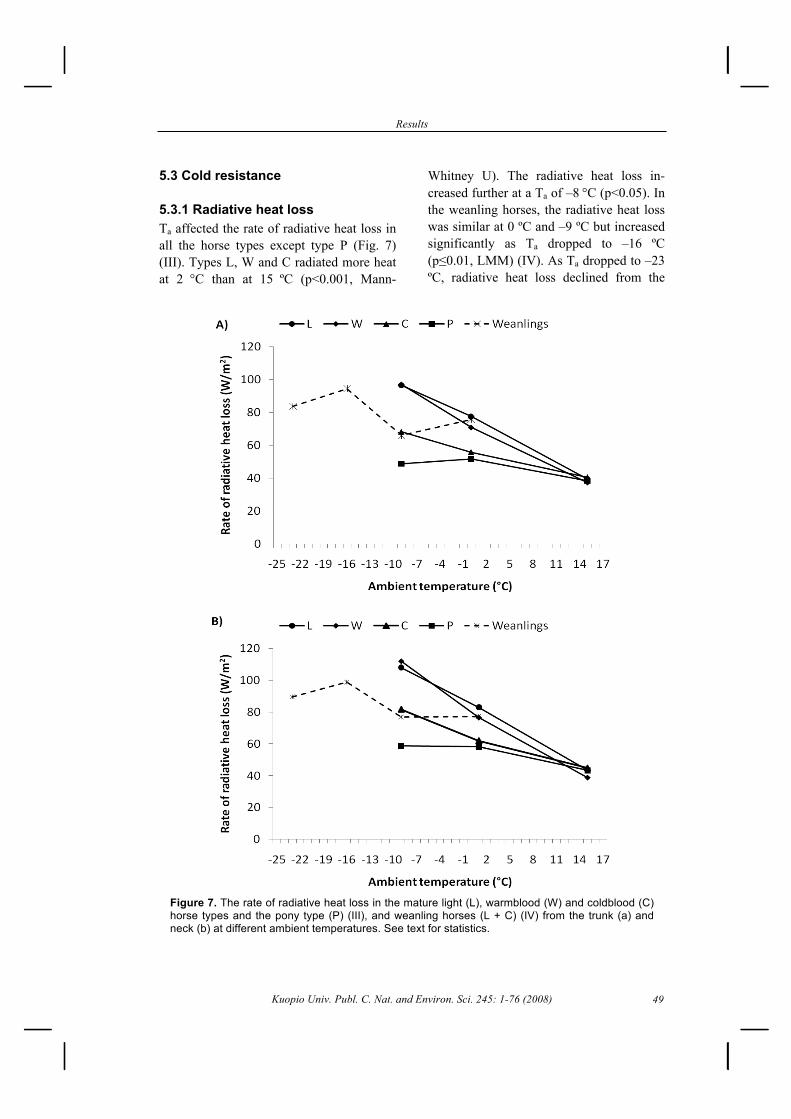
Results
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 49
5.3 Cold resistance 5.3.1 Radiative heat loss Ta affected the rate of radiative heat loss in all the horse types except type P (Fig. 7) (III). Types L, W and C radiated more heat at 2 °C than at 15 ºC (p<0.001, Mann-
Whitney U). The radiative heat loss in-creased further at a Ta of –8 °C (p<0.05). In the weanling horses, the radiative heat loss was similar at 0 ºC and –9 ºC but increased significantly as Ta dropped to –16 ºC (p≤0.01, LMM) (IV). As Ta dropped to –23 ºC, radiative heat loss declined from the
Figure 7. The rate of radiative heat loss in the mature light (L), warmblood (W) and coldblood (C) horse types and the pony type (P) (III), and weanling horses (L + C) (IV) from the trunk (a) and neck (b) at different ambient temperatures. See text for statistics.

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 50
trunk (p<0.05). There were no differences in the rate of
radiative heat loss from the trunk or neck between the horse types at 15 °C (Fig. 7) (III). Differences started to occur at a Ta of 2 ºC. Type L radiated more heat from the trunk and neck at 2 °C than types C and P (p<0.05, Mann-Whitney U), and type W radiated more heat from the neck than type C (p<0.05). At –8 °C, types L and W radi-ated more heat from the trunk and neck than type C (p<0.05). 5.3.2 Thermal insulation The hair coat weight changed in all the horse types in the course of the season (Fig. 8) (III). Hair coat weight increased between August and March (p<0.001, Mann-Whitney U). In August and October, type L had less hair than the other horse types (p<0.05, Mann-Whitney U). In October, types W and C had less hair than type P (p<0.05). In March, the hair weight of types L and W was lower than that of types C and
P (p<0.05). On the basis of complementary measurements, hair weight was highest in the mature horses in December (III, U2). In the weanling horses, hair weight was high-est in February (IV).
The rate of radiative heat loss from the neck decreased with the increasing hair weight of the mid-neck coat (R2=0.12, p<0.05, Linear Regression) (Fig. 9), and the rate of radiative heat loss from the trunk increased with increasing metabolic BW (R2=0.16, p<0.05) (Fig. 10) (IV). Hair weight (R2=0.04, p=0.27) did not correlate with the rate of radiative heat loss from the trunk, nor metabolic BW with the rate of radiative heat loss from the neck (R2=0.10, p=0.06). BCS had no effect on the rate of radiative heat loss from the trunk (R2=0.004, p=0.73) or neck (R2=0.08, p=0.10).
Figure 8. Hair coat weight development in the mature light (L), warmblood (W) and coldblood (C) horse types and the pony type (P) (III), and weanling horses (L + C) (IV) from autumn to spring. * = results originate from the complementary measurements (U2).

Results
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 51
Figure 9. Relationship between the rate of radiative heat loss (y) and hair weight from the neck (x) (IV).
Figure 10. Relationship between the rate of radiative heat loss from the trunk (y) and metabolic body weight (BW) (x) (IV).


Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 53
6. DISCUSSION
The purpose of the present thesis was to evaluate the suitability of loose housing, which is nowadays commonly used for growing horses, for a cold, northern climate by studying the effects of a cold loose hous-ing environment on weanling horses. Some welfare impacts, which are generally asso-ciated with different management envi-ronments and were discussed in the litera-ture review, are summarized in Table 9. In the present thesis, the following welfare issues related to the loose housing were examined: the opportunity to fulfil behav-ioural needs (I), the utilization of the oppor-tunity for free exercise (I, U1) and the ef-fects of a cold environment on nutrition and
growth (II) and cold resistance (III, IV, U2) (cf. Table 9). The variables used repre-sented a variety of physiological and behav-ioural mechanisms described in Tables 3 and 4 in the literature review.
With regard to the applicability of the results of the present thesis, it should be noted that the loose house provided good protection from the weather elements, since it was insulated, well-bedded and had co-vered entrance shelters. Quite often, loose houses are uninsulated buildings without entrance shelters, and thus provide less pro-tection from the weather than the facilities in the present thesis. Therefore, the results can be applied only to comparable loose
Table 9. A summary of some welfare-enhancing and -threatening factors in a box stabling and group/outdoor housing system. Factors which were examined in the present thesis are in italics. Welfare-enhancing factors Welfare-threatening factors Stabling in single, loose boxes
• Personal space and management routine • Low injury risk • Daily handling and control of diseases
and injuries • Controlled feed intake and quality • Controlled exercise
• Low environmental stimulation and lack of social contacts
• Limited opportunity to fulfil behavioural needs: high risk of aberrant or unwanted, aggressive and stereotypic behaviour
• Restricted exercise opportunity: risk of re-tardation of growth and development
• Risk of exposure to high levels of ammo-nia, organic dusts and mould spores
Group/outdoor housing
• Opportunity to fulfil behavioural needs • Free exercise opportunity: high locomo-
tor activity; supports normal growth and development
• Social contacts • Horses are less aggressive and better to
handle and train
• Social ranking order: aggressive group mates, high risk of injuries, low ranking horses may have limited access to food and shelter
• Diseases and injuries more difficult to control
• Minor daily handling: restless, shy horses • Exposure to cold and adverse weather:
increased energy need, increased risk of airway inflammations
• Feed intake difficult to control
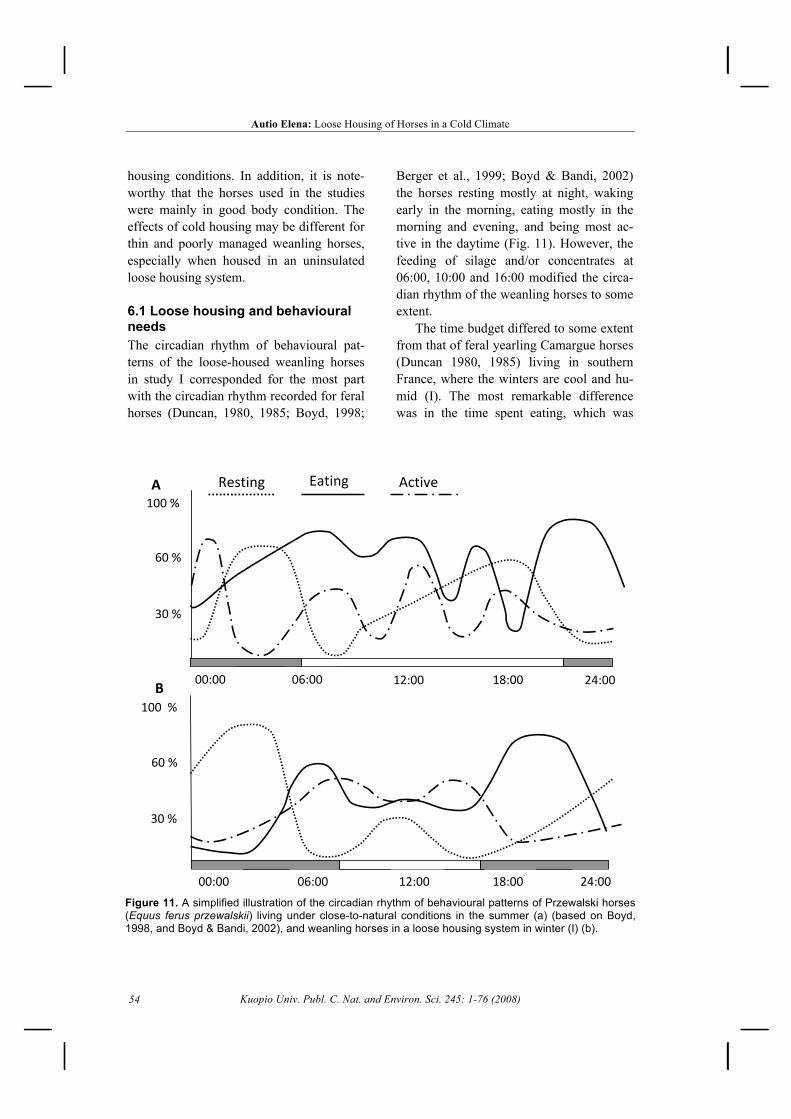
Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 54
housing conditions. In addition, it is note-worthy that the horses used in the studies were mainly in good body condition. The effects of cold housing may be different for thin and poorly managed weanling horses, especially when housed in an uninsulated loose housing system.
6.1 Loose housing and behavioural needs The circadian rhythm of behavioural pat-terns of the loose-housed weanling horses in study I corresponded for the most part with the circadian rhythm recorded for feral horses (Duncan, 1980, 1985; Boyd, 1998;
Berger et al., 1999; Boyd & Bandi, 2002) the horses resting mostly at night, waking early in the morning, eating mostly in the morning and evening, and being most ac-tive in the daytime (Fig. 11). However, the feeding of silage and/or concentrates at 06:00, 10:00 and 16:00 modified the circa-dian rhythm of the weanling horses to some extent.
The time budget differed to some extent from that of feral yearling Camargue horses (Duncan 1980, 1985) living in southern France, where the winters are cool and hu-mid (I). The most remarkable difference was in the time spent eating, which was
30%
ActiveEatingResting
60%
100%A
B00:00 24:0012:0006:00 18:00
100%
60%
30%
00:00 24:0012:0006:00 18:00
Figure 11. A simplified illustration of the circadian rhythm of behavioural patterns of Przewalski horses (Equus ferus przewalskii) living under close-to-natural conditions in the summer (a) (based on Boyd, 1998, and Boyd & Bandi, 2002), and weanling horses in a loose housing system in winter (I) (b).

Discussion
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 55
much lower in the loose-housed horses (36 vs. 63 % for loose-housed weanlings and feral yearlings, respectively). This was be-cause, in contrast to feral horses in natural environments (Duncan 1980, 1985), the feeds of the weanling horses were energy-rich and easily available, and hence less time spent eating satisfied their energy needs. Consequently, the loose-housed horses (I) replaced eating time with stand-ing, whereas feral horses (Duncan 1980, 1985) spent only a little time standing (26 vs. 6 %). The time spent on locomotor ac-tivity (5 vs. 6 %) and resting (32 vs. 25 %) was about the same, but the weanling horses rested mostly in a lying rather than a standing position (I), contrary to feral horses (Duncan, 1980, 1985) (25 vs. 9 %). This may have been because there was a comfortable, dry lying place inside the loose house. They also rested recumbent all at the same time, which shows that low-ranking horses could also use the hall. Both of these observations signify that the wean-ling horses felt safe in the loose housing environment, because, for example, horses in pasture usually reduce lying time to minimum (Dallaire, 1986) and at least one horse remains standing while the others are lying down (McDonnell, 2003), which are protective mechanisms against predation. The weanling horses used the sleeping hall for resting in the daytime, too, and spent time outdoors in the evenings and nights, which is not possible for stabled horses kept in loose boxes at night and in an outdoor paddock in the day. Therefore, the results of study I are mainly in accordance with the hypothesis that the loose housing system would provide the horses’ with an opportu-nity to fulfil their behavioural needs, which is generally considered an important cri-terion for animal welfare (Bracke & Hop-ster, 2006).
The observation of all the horses lying recumbent at the same time (I) verifies that
the sleeping hall was large enough for the number of horses. There was 9 m2 space per weanling horse in the halls, which corres-ponds to the space requirement of a loose box for a mature horse. Since Finnish legis-lation (Ministry of Agriculture and For-estry, 1998) sets a minimum space require-ment for weanling horses in a loose housing system of 3.6 m2 per horse, the weanling horses had plenty of space. However, the adequacy of this minimum requirement cannot be evaluated on the basis of this the-sis. For mature horses, on the other hand, the minimum space requirement in a loose housing system (7.2 m2 per horse) was seen to be too little, since only three mature horses could be kept in the present loose housing system, giving a space of about 15 m2 per horse. When there were four mature horses (about 11 m2 per horse), one horse was not able to use the hall at the same time as the other horses. However, because space requirements were not a topic of the present thesis, these observations are suggestive.
Another hypothesis concerning behav-iour was that the loose-housed weanling horses were expected to be quite active due to 24-h opportunity for exercise (Zeeb & Schnitzer, 1997). Contrary to this hypoth-esis and general assumption among horse breeders, the loose housing system did not encourage daily locomotor activity during the cold winter months (I). The GPS meas-urements (U1) confirmed the results of study I: the average speed of the horses in a loose housing system was 0.1 km/h (0.03 m/s), which signifies that the horses moved really slowly and stood still for most of the time, although short spurts were occasion-ally performed. The distance that the loose-housed weanling horses travelled per day in study U1 was about half of that of the year-ling horses in pasture in the summer. In winter, locomotor activity was about the same as in feral yearling horses in winter (Duncan, 1980, 1985). However, feral

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 56
horses grazed in winter, i.e. ingested food while standing or travelling slowly (search-ing for food), so their eating time (63 %) included locomotor activity. The loose-housed horses were not able to or did not have to graze due to ad libitum forage feed-ing, which may have resulted to relatively low locomotor activity. In the summer, the horses were able to graze which increased their locomotor activity in pasture. Accom-panying eating, this is a kind of obligatory activity, which could not be performed by the loose-housed horses in winter.
Feral horses have been reported to re-duce locomotor activity in winter compared to summer (Duncan, 1980, 1985; Berger et al., 1999; Arnold et al., 2006). Reduced activity in winter was an annual pattern which was related to decreased Tb, and hence to a reduction in endogenous HP and energy expenditure (Arnold et al., 2006). This adaptation mechanism helped feral horses to cope with the energetic challenge of winter. Although this adaptation mecha-nism would not exist to the same extent in domestic horses, who usually do not con-front as hard an energetic challenge in win-ter due to feeding practices, it is possible that the rather low locomotor activity may have served the same purpose: the reduction of energy expenditure in winter. It should be pointed out, however, that the thick snow cover and only daytime access to a large paddock (6,400 m2) may partly ex-plain the low activity in the present studies (I, U1). The horses had access only to smaller paddocks (800 m2) at night. How-ever, the reducing effect on activity of night-time access to smaller paddocks can be discarded, since in previous studies the sizes of paddocks which enhanced horse activity were 450 m2 (Jørgensen & Bøe, 2007) and 1,600 m2 (Johansson & Lind-ström, 2008).
The results signify that a loose housing system does not necessarily fulfil its ulti-
mate purpose as an activating housing sys-tem in areas with a cold and long winter (I, U1). Basically, a loose housing system gives horses an opportunity for high loco-motor activity, but in practice, it does not seem to serve its purpose. Furthermore, on the basis of studies that show that a lack of exercise may retard growing horses’ mus-culoskeletal development (Raub et al., 1989; Bell et al., 2001; van de Lest et al., 2002; Hiney et al., 2004), it can be hy-pothesized that if the locomotor activity is very low, the loose housing system does not necessarily bring any benefit when consid-ering musculoskeletal development. How-ever, further studies are required in order to find out whether the low locomotor activity of weanling horses in a loose housing sys-tem is adequate for normal musculoskeletal development.
Also ways to encourage locomotor ac-tivity have to be identified. For example, placing forages further from the loose house, ploughing snow in the paddocks in order to ease movement in the winter, locat-ing the loose housing system in a varied and stimulating environment (e.g. trees or other vegetation in the paddocks, varying landscape, horse traffic near the loose house), paying attention to the structure and shape of the paddocks and forming diverse loose housing groups (e.g. groups in which horses vary in age and breed) might be suit-able ways of activating horses. Agitation of loose-housed horses for exercise purposes must be avoided, since sudden, heavy exer-cise adversely affects musculoskeletal de-velopment (see van de Lest et al., 2002, 2003). However, a question remains as to whether it is even possible to increase the locomotor activity of horses in the cold, given that low locomotor activity is a nor-mal response to cold weather (Duncan 1980, 1985; Berger et al., 1999; Arnold et al., 2006) and cold adaptation is a very deep-seated mechanism (Langlois, 1994).

Discussion
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 57
The low activity level of the weanling horses also indicates that their actual need for locomotor activity was small, especially at night. This is supported by the fact that locomotor activity appeared mostly in the daytime in the loose-housed horses (I, U1), and there was a small difference in daily distances travelled between the horses kept at night in a group box and those kept 24 h in a loose housing system (U1). The small difference in total distances travelled may be a sign of compensatory locomotor be-haviour, i.e. increased locomotor activity when being turned out for part of the day, in the horses kept in a group box during the night. Compensatory locomotor behaviour is common in stabled horses in response to exercise deprivation (Houpt et al., 2001; Chaya et al., 2006; Kurvers et al., 2006) and proves that stabled horses are able to fulfil their need for locomotor activity in the daytime. However, the fact that the av-erage speed was lower in the loose housing system than in the group box/paddock sys-tem shows that locomotor activity is di-vided more equally during the day when the horses have a 24-h exercise opportunity than when the horses only have a 9-h exer-cise opportunity. On the other hand, the activity results of study U1 should be con-sidered preliminary since the number of horses involved was small.
One important purpose of the prelimi-nary study U1 was to evaluate the suitability of GPS technology for recording activity of horses. The same technology was used in a recent study, which measured the locomo-tor activity of foals kept in pastures under various management conditions (Kurvers et al., 2006). In the present study, it was pos-sible to measure the distance that the horses travelled using GPS equipment, but the results are only indicative because of sev-eral problems with GPS recordings, which were also noted by Kurvers et al. (2006). The main problem was that a lot of obser-
vational days were lost due signal breaks with satellites, especially in the loose hous-ing system, since the building sometimes interfered with reception of signals from the satellites. This is why this method cannot be recommended for use near buildings. Furthermore, the equipment recorded the activity of loose-housed horses only when they were outdoors in the paddock. There-fore, there were inaccuracies in the record-ings of the loose-housed horses and the method should have been validated. In the pasture, the number of successful meas-urements was higher because there were no disturbing trees or buildings. The horses were not distracted by the GPS devices, but the activity of the horses, e.g. rolling and biting of the harnesses or capsules, occa-sionally damaged the equipment. Accord-ingly, there is apparently still a need to de-velop better methods for automatic activity recordings of free-ranging animals.
Conclusions
According to the hypothesis, free outdoor access and ad libitum forage feeding al-lowed the horses to follow the natural cir-cadian rhythm of behavioural patterns in a loose housing system, which enhances horse welfare (I). The time budget differed more noticeably from that of feral horses, since it was greatly modified by manage-ment factors such as composition and qual-ity of diet and access to a sheltered sleeping hall. The horses spent almost half the day in the hall mostly resting recumbent, which signifies that a sheltered, comfortable loose house building that is large enough for the number of horses is very important for the welfare of outdoor-housed horses. The re-sults on activity did not support the hy-pothesis of the high locomotor activity (I, U1). Evidently, the need for locomotor ac-tivity was low and was partly a thermoregu-latory response to cold. It seems that the

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 58
opportunity to behave unrestrictedly rather than the opportunity for high locomotor activity is important for horses. Neverthe-less, the amount and type of exercise suffi-cient to ensure normal musculoskeletal de-velopment of horses in a loose housing sys-tem, and also ways to stimulate activity, should be examined in future studies.
6.2 Effects of a cold housing envi-ronment on horses The weanling horses did not modify their behaviour in response to decreasing Ta in winter (I), which does not support the hy-pothesis that the horses would use behav-ioural adjustments to acclimatize to the cold housing environment. The horses did not change the time spent resting, eating hay or moving, which accords with the results of a study of loose-housed Icelandic horses (Mejdell & Bøe, 2005). Feral horses, in contrast, increased the time spent standing resting in response to decreasing Ta, and modified their time budget between seasons (Duncan, 1980, 1985). This indicates that, in study I, the winter weather was not chal-lenging for the weanling horses, or alterna-tively, they had already acclimatized and adjusted their time budget to the cold hous-ing environment to some extent before the study began. This latter explanation is sup-ported by the fact that the horses had al-ready been in the loose housing system for over a month before the study began. In addition, an adequate plane of nutrition and an insulated loose house building may have helped them to cope with the cold.
The low level of locomotor activity and the presence of huddling, which is classi-fied as a thermoregulatory, comfort-seeking behavioural response to cold (McDonnell, 2003), at all measured Ta’s between –20 ºC and 0 ºC may also support the fact that the horses had already adjusted their behaviour to the cold, although the proportion of ac-tivity did not decrease and huddling did not
increase further at lower Ta’s (I). This would also support the hypothesis of the utilization of behavioural responses to cold. The incidence of huddling cannot be com-pared with other studies, because huddling has not apparently been previously studied in the horse. Another interesting ther-moregulatory behaviour associated with resting, which has not obviously been stud-ied in the horse, is the lying posture in rela-tion to Ta. In other domestic ungulates, e.g. dairy calves (Gonzales-Jimenez & Blaxter, 1962; Kauppinen, 2000; Hänninen et al., 2003), lying on the side decreases at low Ta’s, whereas lying with the head on the flank with the legs curled under the body increases, which may be a way of lessening the impact of the cold by reducing the ex-posed body surface.
In contrast, precipitation, RH, wind speed and cloudiness had an effect on the behaviour of the weanling horses (I). The time spent eating hay decreased and the time spent in the shelter and standing in-creased with increasing RH and cloudiness, which was naturally a consequence of the increased time spent standing in the shelter on rainy days. Wind also increased use of the shelter, but also increased locomotor activity, i.e. the horses were restless on windy days. The results of study I concern-ing use of the shelter during rainy and windy weather are in accordance with earlier studies (Michanek & Ventorp, 1996; Mejdell & Bøe, 2005), and suggest that a shelter which protects the horses from ad-verse weather conditions is important for the welfare and thermoregulation of out-door-housed horses.
Although the weanling horses did not modify the time spent eating hay as the Ta decreased (I), it was hypothesized, on the basis of previous studies, that low Ta would affect the energy needs of the horses. It was also expected that the horses would grow normally and maintain their BCS when the

Discussion
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 59
CED was taken into account in the feeding. Cold-housed growing horses have been reported to need 0.2 to 2 % more energy per 1 ºC decrease in Ta below the LCT to main-tain normal growth rate in cold (Cymbaluk et al., 1989a; Cymbaluk, 1990). The LCT of cold-housed weanling horses varies in the studies from 0 ºC in Cymbaluk (1990) and Cymbaluk et al. (1989a) to –11 ºC in Cym-baluk and Christison (1989a) depending on the housing system, environmental condi-tions and the horses’ level of acclimatiza-tion. This increase in energy need has not been taken into account in the Finnish feed-ing recommendations for horses (MTT, 2006), but the Swedish recommendations include guidelines for feeding horses during cold weather (SLU, 2004). According to the SLU, a young horse fed ad libitum may need 1.4 % more maintenance energy for each 1 ºC decrease in Ta below the LCT of –11 ºC. In study II, the results concerning CED and growth of weanling horses in a cold housing environment are in accordance with previous studies, the SLU (2004) guideline and the hypothesis of the increas-ing energy need in the cold.
Weather conditions were harsh right from the beginning of studies I and II, so the ME intake of the weanling horses had to be increased above the maintenance level in order to maintain a moderate BCS and normal growth rate (II). ME intake in-creased by 1.8 % in November, 0.5 % (CED 1.0 W/m2) in December and 0.2 % (0.3 W/m2) in January per 1 °C decrease in Ta. Accordingly, the HP of the weanling horses, as estimated from the ME intake, was relatively high, varying between 231 and 258 W/m2 during the winter. For com-parison, similar MR values have been re-corded for neonatal foals after birth (Ousey et al., 1991), and much lower values (about 120 W/m2) in few-day-old foals at Ta’s be-low their LCT (Ousey et al., 1992). The LCT of the weanling horses was about –11
°C, since ME intakes were higher at Ta’s below than above –11 °C, and were ap-proximately the same between 0 ºC and –11 ºC. This LCT result supports the estimation of LCT (–11 °C) by Cymbaluk and Christi-son (1989a) for light horse weanlings, which were kept under comparable climatic and management conditions. As in my study, Cymbaluk and Christison (1989a) also defined the LCT on the basis of feed intake, and, in both studies, the LCT was defined for animals in their natural state i.e. animals performing their normal activities (field metabolic rate, Randall et al., 2001) and with specific thermal resistance, and not in rested, fasting animals in a post-absorptive state (BMR, IUPS Thermal Commission, 2003). Although there were similarities in the horses, management con-ditions, climate and LCT definitions, it should still be remembered that the LCT values are not directly comparable since many other factors, e.g. BCS, body size, physiological status, MR and acclimatiza-tion, and also the quality of feeds and inten-sity of feeding, affect the LCT value (Cym-baluk, 1994; Morgan et al., 2007).
Low Ta increased both the total and mass-specific ME intakes of the weanling horses in early winter but not in late winter (II), which is in accordance with a previous study of cold-housed weanling horses (Cymbaluk & Christison, 1989a). A drop in Ta to –20 °C would hence increase the daily ME need by about 18 % in November, 5 % in December and 2 % in January. The de-creasing CED signifies that the horses ac-climatized progressively to the cold envi-ronment as the winter proceeded, and that the improving cold resistance was mainly caused by increasing body insulation. How-ever, it is not clear whether the improved insulation was caused by increasing coat insulation or by the decreasing ratio be-tween heat dissipating surface area and heat producing/retaining body mass as the

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 60
horses grew, or by both of these. The for-mer is supported by study IV, where the body insulation of the weanling horses, as measured by hair weight, increased during the winter. In addition, on the basis of study I, the horses probably used behavioural thermoregulation in acclimatization. It is also possible that some other physiological mechanisms, e.g. piloerection, vasocon-striction or adjustments in respiration rate, may have occurred that were not examined in the present studies.
Overall, the results indicate that the level of acclimatization has a great effect on the CED of weanling horses in a cold envi-ronment (II). Early winter, in particular, may be nutritionally very challenging be-fore weanling horses become cold-acclimatized. Conversely, in late winter when horses are older and well acclimatized to cold, the SLU (2004) guideline to in-crease energy intake by 1.4 % for each 1 ºC decrease in Ta below –11 ºC may lead to excess weight gain if followed rigidly throughout the winter. Therefore, it is im-portant that the amount of extra energy fed is adjusted according to the horse’s level of acclimatization, and that the feeding rec-ommendations take account of the reducing effect of acclimatization on the extra energy need.
In contrast to the situation in early win-ter, the energy intake of the weanling horses increased in late winter as Ta increased (II). Increasing body size (maintenance energy need increases), changes in the composition of weight gain (the protein content of gain decreases and fat content increases with age) and increasing capacity for feed intake probably explain the increased energy need (Frape, 1998; NRC, 2007) and, hence, in-creased voluntary forage intake in late win-ter. Markedly increased hay intake com-bined with slower growth rate and higher Ta in late winter caused BCS of the horses to exceed the target score. Their nutrient in-
takes were also tens of percentage points above nutrient requirements, because as the feed and ME intakes increased, also intakes of other nutrients increased.
A positive correlation between volun-tary hay intake and excessive nutrient in-take proves that overfeeding was mainly caused by ad libitum hay feeding, and that the reduction in concentrates and silage fed was not sufficient to prevent overfeeding (II). Since it has also been previously re-ported that ad libitum-fed growing horses tend to have a higher nutrient intake than their nutritional needs (Cymbaluk & Christison, 1989a, 1989b; Cymbaluk et al., 1989a) and are susceptible to overfeeding and imbalanced nutrition (Cymbaluk et al., 1989a, 1989b, 1990), ad libitum forage feeding practices in loose housing systems should be reconsidered at least in late win-ter, especially when the quality of the for-ages used is good. The excellent nutritional value of the feeds probably increased the excessive intakes in the present study. In fact, the feeding of silage should have been stopped when the horses had acclimatized to the cold. Silage was originally included in the diet in order to ensure adequate nutri-ent intake in early winter when the horses were not cold-acclimatized, had a high growth rate and had limited capacity for feed intake. The ration of mineral and vita-min supplement could not be decreased since it was important in balancing the cal-cium/phosphorus ratio in the diet.
The intakes of ME, digestible crude pro-tein, calcium and phosphorus were about 30 to 70 % above the nutrient requirements for 6-12-month-old slow growing horses, and about 15 to 45 % above the requirements for rapid growing horses (SLU, 2004; MTT, 2006) (II). Therefore, the nutrient intake of the horses was sufficient for rapid growth, their ADG and BW were the same as or higher than in horses of similar age and breed in earlier studies (Saastamoinen,

Discussion
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 61
1993; Saastamoinen & Koskinen, 1993; Sandgren et al., 1993; Ahtila & Saas-tamoinen, 2005; Andrew et al., 2006; Forsmark, 2006), and their BCS was higher than the moderate target score (Henneke et al., 1983) in late winter. The negative corre-lation between ADG and Ta signifies that, contrary to expectations, the ADG de-creased when the Ta increased. This para-dox is explained by the fact that the ADG was highest in early winter when Ta’s were lowest, and decreased during the winter while Ta increased. ADG was hence ex-plained by the age of the horses and indi-vidual differences in growth rate rather than by Ta or ME intake. Retardation of growth and subsequent periods of compensatory growth could thus not be detected. The finding that horses grow normally and maintain their BCS when CED is taken into account in the feeding, is in accordance with the hypothesis.
The results of study II indicate that ad-justing the energy intake of weanling horses
in a cold group housing system is compli-cated even under well-controlled experi-mental conditions. Thus, the risk of over-feeding and imbalanced nutrition is high in loose housing systems in late winter. This problem is illustrated in Fig. 12. The diffi-culties in regulating the nutrient intake and BCS of horses result from the changes in both the environment (fluctuating Ta) and the animals (changing level of acclimatiza-tion, growth rate and capacity for forage intake), and also from the group housing situation in which forage intake cannot be adjusted individually. It has been previ-ously reported that the balancing of nutrient intakes of growing horses is already diffi-cult for horse breeders (Gibbs & Cohen, 2001), and cold housing increases this diffi-culty. Therefore, the “nutritional welfare” of loose-housed weanling horses may be impaired. Extra energy needed for mainte-nance in early winter and excess forage intake in late winter also increase feeding costs and lower the profitability of loose
Ta Voluntaryhayintake CED ADG BCS
Nov Dec Jan Feb Mar
PROBLEMSINREGULATINGBCSANDNUTRIENTINTAKEOFTHEHORSES
Figure 12. A simplified synthesis of the fluctuation of biological and environmental variables in loose housing conditions during the winter (II). Ta = ambient temperature, CED = climatic energy demand, ADG = average daily gain, BCS = body condition score.

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 62
house rearing of horses. In addition to energy intake, the rate of
radiative heat loss, as measured by infrared thermography, was also expected to in-crease with falling Ta. In mature horses, the radiative heat loss was higher in the exam-ination conducted at 2 ºC than at 15 ºC, and higher in the examination at –8 ºC than at 2 ºC (III). In the weanling horses, radiative heat loss was similar at 0 ºC and –9 ºC, but increased as Ta dropped to –16 ºC (IV). Lower radiative heat loss from the trunk at –23 ºC than –16 ºC was probably caused by the frost on the surface of the hair coat at –23 ºC. Thus, the hypothesis that radiative heat loss increases as Ta decreases can be accepted. Increasing radiative heat loss is caused by the increasing temperature gradi-ent between the surface of the animal and Ta (Langlois, 1994; Randall et al., 2001) and may partly explain the CED result in study II.
MacCormack & Bruce (1991) reported that CED can be efficiently reduced by pro-vision of weather protection for horses in the form of a rug, shelter or both, which thus facilitates the maintenance of heat bal-ance in the cold. A shelter reduces heat loss resulting from forced convection and ev-aporation by protecting horses from wind and rain (MacCormack & Bruce, 1991). Roof and walls reduce the heat loss from horses by radiation, and conversely, give protection from solar radiation (Clark & McArthur, 1994). In addition, dry bedding reduces heat loss by conduction when the animal is lying (Sjaastad et al., 2003). These facts indicate that if the loose house building in the present study had not pro-
vided good protection from the weather elements, the CED of the loose-housed weanling horses could have been even higher (II).
Conclusions
The responses of the horses to Ta are sum-marized in Fig. 13 (I, II, III, IV, U1). Con-trary to the hypothesis, the weanling horses did not greatly change their behaviour as a response to falling Ta. This indicates that they probably had adjusted their behaviour, especially locomotor and resting behaviour, to prevailing conditions before studies I and II began. Precipitation, RH and wind had a great effect on behaviour, i.e. the weanling horses used behavioural re-sponses in order to diminish the adverse effects of rainy and windy weather on heat balance (I). According to the hypothesis, radiative heat loss increased with falling Ta
except in type P (III, IV). Similarly, energy intake increased markedly below –11 ºC signifying that the weanling horses re-sponded to cold by increasing metabolic HP (II). Low Ta did not increase the energy need in late winter, indicating that the horses acclimatized to the cold environment during the winter. Improved cold resistance was caused by increasing body insulation, as measured by hair weight (IV), and by increasing body size (II). As presented by Young (1975), horses used behavioural and metabolic thermoregulation in the cold in the short run and insulation changes in the long run.

Discussion
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 63
6.3 Cold resistance of different horse types Knowledge of the cold resistance and LCT of different types of horses is of practical use, e.g. with respect to the ventilation of stables and feeding and management prac-tices (i.e. housing system, blanketing and clipping) in a cold climate (Morgan et al., 2007). However, the cold resistance and LCT values of different horse types have not been systematically studied. Moreover, the methods used to determine the LCT and the research designs vary greatly in existing studies (see Table 2 in the literature re-view), probably because the most reliable methods for measuring the MR, an animal calorimeter and respirometer (Randall et al., 2001), demand expensive equipment, par-ticularly in the case of large animals.
In the present studies, the thermoregula-tory capacity of different horse types (III)
and weanling horses (IV) was evaluated using infrared thermography, in which the measurement of heat loss is based on the radiative temperature of an object (Dere-niak & Boreman, 1996). Therefore, ther-mography measures radiative heat loss from an object, not total heat loss, which consists of convection, conduction, radiation and evaporation (Clark & McArthur, 1994). In a thermoneutral diagram (Mount, 1973), LCT is defined on the basis of increasing non-evaporative heat loss consisting of convec-tion, conduction and radiation. It is prob-able that in a standing animal in still air conditions, this increase in non-evaporative heat loss below the LCT is mainly caused by an increase in natural convection and radiation, because, in a standing animal, conduction is small and can be neglected (Clark & McArthur, 1994). Convection and radiation follow the temperature gradient
‐Radiativeheatlossincreasedintheweanlinghorses
‐Locomotoractivitywasrelativelyhighinthefoalsandyearlinghorses
‐Radiativeheatlossdidnotchangeintheponiesbutin‐creasedinthelight,warmbloodandcoldbloodhorses
‐Locomotoractivitywaslowintheweanlinghorses
‐Totalactivitydecreasedanduseofthesleepinghallincreasedslightlyintheweanlinghorses
Ta(ºC) –25 –20 –15 –10 –5 0
5 10 15 20
IV
III
II
U1
I
‐Thetimespentlyingwithorwithouthud‐dling,movingoreatinghaydidnotchange
intheweanlinghorses
Study:
‐Energyintakeincreasedintheweanlinghorses
Figure 13. A summary of the responses of the horses to ambient temperature (Ta) in the present studies.
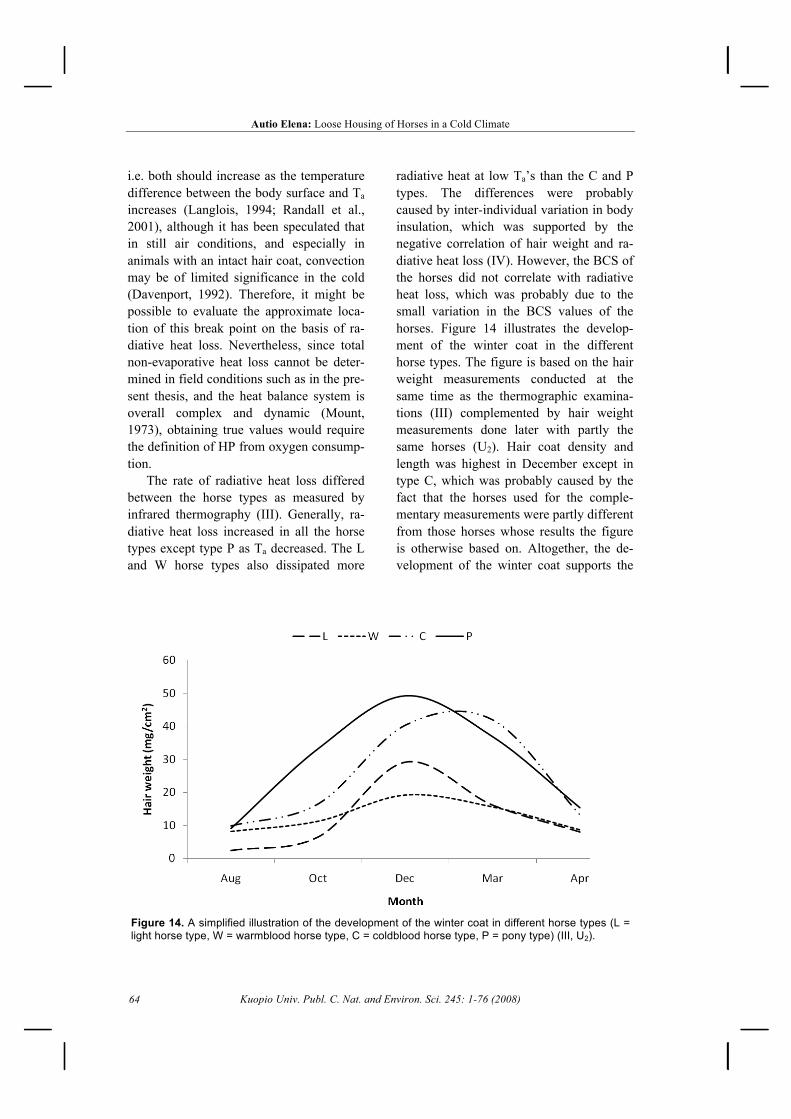
Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 64
i.e. both should increase as the temperature difference between the body surface and Ta increases (Langlois, 1994; Randall et al., 2001), although it has been speculated that in still air conditions, and especially in animals with an intact hair coat, convection may be of limited significance in the cold (Davenport, 1992). Therefore, it might be possible to evaluate the approximate loca-tion of this break point on the basis of ra-diative heat loss. Nevertheless, since total non-evaporative heat loss cannot be deter-mined in field conditions such as in the pre-sent thesis, and the heat balance system is overall complex and dynamic (Mount, 1973), obtaining true values would require the definition of HP from oxygen consump-tion.
The rate of radiative heat loss differed between the horse types as measured by infrared thermography (III). Generally, ra-diative heat loss increased in all the horse types except type P as Ta decreased. The L and W horse types also dissipated more
radiative heat at low Ta’s than the C and P types. The differences were probably caused by inter-individual variation in body insulation, which was supported by the negative correlation of hair weight and ra-diative heat loss (IV). However, the BCS of the horses did not correlate with radiative heat loss, which was probably due to the small variation in the BCS values of the horses. Figure 14 illustrates the develop-ment of the winter coat in the different horse types. The figure is based on the hair weight measurements conducted at the same time as the thermographic examina-tions (III) complemented by hair weight measurements done later with partly the same horses (U2). Hair coat density and length was highest in December except in type C, which was probably caused by the fact that the horses used for the comple-mentary measurements were partly different from those horses whose results the figure is otherwise based on. Altogether, the de-velopment of the winter coat supports the
Figure 14. A simplified illustration of the development of the winter coat in different horse types (L = light horse type, W = warmblood horse type, C = coldblood horse type, P = pony type) (III, U2).

Discussion
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 65
results of the infrared thermography, be-cause the rate of radiative heat loss was highest in horse types L and W, which had the lowest hair coat weight. The precise development and thickness of the winter coat in different horse types should be stud-ied further with more horses, with the measurements being done during one win-ter.
The results for radiative heat loss and winter coat development suggest that there might be differences in cold resistance be-tween different horse types (III). Types P and C seem to be more cold-resistant and thus better adapted to the cold than types L and W. This would be in accordance with the hypothesis and the general evolutionary theory of northern and southern horse breeds (Langlois, 1994). The evolutionary theory suggests that a spherical body form is an advantage in a cold climate, because it reduces the exposed portions of the body. Northern horse breeds are generally heavier and shorter than southern horse breeds, and have shorter extremities which are well protected by hair, mane and fetlock, whereas southern horse breeds’ extremities are longer and the coat provides less insula-tion. These facts support the hypothesis that loose housing might be better suited to coldblood horses and ponies. In addition, the preliminary results are in accordance with the study by Mejdell and Bøe (2005), who noted that Icelandic horses, which are thought to be hardy with regard to cold re-sistance, managed well in a loose housing environment with Ta’s down to –31 ºC.
In view of genetic differences in cold resistance, the loose housing of the W and L horse types may be questioned. More-over, according to the Ministry of Agricul-ture and Forestry Statute on Animal Protec-tion (396/1996), only those production animals and breeds that are adapted to out-door housing can be reared outdoors all year round. The risk of negative welfare
effects in a loose housing system may thus be higher for light and warmblood horse breeds than for coldblood and pony breeds. However, as mentioned above, for confi-dent conclusions about cold resistance, fur-ther studies are needed regarding this issue. In addition, total body insulation, which consists of the body tissue, hair coat and air acting in series (McArthur, 1991), of differ-ent horse types should also be determined.
In the weanling horses, radiative heat loss was similar at 0 ºC and –9 ºC but in-creased as Ta dropped to –16 ºC (IV). As-suming that radiative heat loss follows the heat gradient (Langlois, 1994; Randall et al., 2001), and hence increases below the LCT, the result may indicate that the LCT was probably between –9 ºC and –16 ºC. This LCT range, as measured by infrared thermography, agrees with the LCT value of –11 ºC defined on the basis of ME intake (II). It would also support the LCT result (–11 ºC) of Cymbaluk and Christison (1989a) for outdoor-housed, light horse weanlings. At the time of the thermographic examin-ations, the weanling horses had apparently acclimatized to the cold housing envi-ronment to some extent, which is supported by the fact that their hair weight per area unit was approximately the same as in ma-ture FC horses in winter in study III. So this LCT value can only be applied to weanling horses that have already acclimatized to the cold to some extent. In the autumn, when weanling horses are usually brought to loose housing systems, their LCT may be considerably higher.
According to the general model of the TNZ (Mount, 1973), increasing non-evaporative heat loss signifies that an ani-mal’s body insulation is maximal and it has to increase its metabolic HP in order to maintain a constant Tb. Although only ra-diative heat loss was measured in study IV, it can be hypothesized that convection may also have increased at very low Ta’s (–16 ºC

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 66
and –23 ºC). Therefore, the horses had to replace increasing non-evaporative heat loss by increasing metabolic HP, because they remained normothermic. This is supported by the results of study II, where the ME intake was higher at Ta’s below –11 ºC than above –11 ºC, and HP, as calculated from feed intake, was generally high. Apparently, on the basis of the literature (Pagan & Hintz, 1986; Morgan, 1995; Vernet et al., 1995; McDonald et al., 2002; Morgan et al., 2007), high ME intake and HP improved the cold resistance of the weanling horses (II). The weanling horses evidently ther-moregulated effectively even at very low Ta’s. Hence, the present (I, II, IV) and pre-vious studies (McBride et al., 1985; Cym-baluk & Christison, 1989a; Cymbaluk, 1990; Morgan et al., 1997; Mejdell & Bøe, 2005) suggest that the horse as a fairly well-insulated, large endothermic animal, could tolerate subzero temperatures rather well, provided that the CED is taken into account in the plane of feeding and the horse has access to a protective, well-bedded shelter. The present studies did not examine extensively the effect of rain and
wind on horses, but, since horses have been generally observed to shiver visibly during poor weather (Irvine, 1967; Mejdell & Bøe, 2005), it may be that rainy and windy weather is more challenging for the horse than dry, sub-zero weather.
Conclusions
Infrared thermography is not the most suit-able method for evaluating the cold resis-tance of different horse types since it meas-ures radiative heat loss, not total non-evaporative heat loss, which increases be-low the LCT. However, since there were differences in the rate of radiative heat loss between horse types and those types with the lowest hair coat weight lost most radia-tive heat, it is possible that infrared ther-mography revealed differences in cold re-sistance to some extent. Further studies, where cold resistance is evaluated, e.g. on the basis of MR, are needed in order to confirm these observations. Weanling horses are fairly cold resistant, since they remained normothermic despite of increas-ing radiative heat loss in the cold.

Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 67
7. SUMMARY AND CONCLUSIONS
The welfare of horses under different man-agement conditions has recently become a subject of debate. Loose housing, which is an alternative housing system for stabling, is expected to solve many of the welfare-threatening factors associated generally with box stabling. This thesis examined the suitability of a loose housing system as a winter environment for horses in a cold climate. The studies focused mainly on weanling horses, but the suitability for dif-ferent horse types was also considered.
Weanling horses are usually weaned from their dams just before they are brought to a loose housing system in autumn, and they originate from many different stables. Therefore, a lot of changes, such as wean-ing and changes in living and thermal envi-ronments, social relationships and feeding, occur in their lives during a short time in-terval. The time before and when arriving in a loose housing system may thus be very stressful for weanling horses. They are not acclimatized to the cold and may become ill if one of the horses is carrying, e.g. an in-fluenza virus. Accordingly, welfare prob-lems are most likely to occur in autumn and early winter.
However, the present thesis demon-strates that loose housing provides horses with the opportunity for more unrestricted behaviour than box stabling, which is un-questionably a welfare-enhancing factor. In addition, weanling horses are capable of acclimatizing to a cold housing envi-ronment, which signifies that they can be reared in cold loose housing systems in winter in a northern climate, provided that the level of feeding, care and housing facili-ties are adequate. Nevertheless, because health is an important aspect of animal wel-fare and was not examined in the present studies, further studies are needed to exam-
ine the health status of loose-housed horses (e.g. the incidence of gastrointestinal and respiratory disorders, leg injuries or bodily injuries such as wounds, bruises and hair loss).
In more detail, the results revealed that: 1. The time budget and circadian rhythm
of the weanling horses for the most part resembled those of feral horses. There-fore, the horses had a good opportunity to fulfil their behavioural needs in the loose housing system. However, loose housing did not encourage locomotor activity, which suggests that the oppor-tunity to behave unrestrictedly rather than the opportunity for high activity is important for horses.
2. The weanling horses used the sleeping hall for resting and as a shelter to a great extent, which is why the sheltered, insulated, well-bedded loose house was important for the welfare and ther-moregulation of the outdoor-housed weanling horses.
3. The cold loose housing environment increased the energy expenditure of the weanling horses in early winter. Cli-matic energy demand gradually de-creased as the horses acclimatized to the cold housing environment. The horses utilized behavioural and metabolic ad-justments when acclimatizing in early winter, and insulation adjustments in the long run. When the climatic energy demand was taken into account in the plane of feeding, the weanling horses maintained their body condition and grew at or above expected rates.
4. Changes in both the environment and weanling horses during the winter and ad libitum forage feeding made the con-trolling of nutrient intakes complicated. Thus, when the weanling horses in-

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 68
creased their voluntary forage intake in late winter, while they were rather inac-tive and their growth rate decreased, their body condition rose above a mod-erate level and they were susceptible to overfeeding.
5. Since the weanling horses maintained their thermal balance even at very low ambient temperatures despite of increas-ing radiative heat loss, they seemed to have a high thermoregulatory capacity.
6. The results for radiative heat loss and development of the winter coat indicate that there might be differences in cold resistance between different horse types, the coldblood horses and ponies being more cold-resistant than warm-blood and light horses, and thus more suitable for loose housing. Although a cold environment is not a
physiological problem in itself for the weanling horses, the stressful early loose housing period, low level of locomotor ac-tivity and complicated adjustment of nutri-ent intakes suggest that it does not necessar-ily bring significant benefits for growth and development. In fact, in order to ensure optimal growth and development, stabling in loose or group boxes with adequate, daily turnout to paddock with age-mates might be a better alternative for weanling horses than a loose housing system, because in box stabling, cold exposure is less, feed and nutrient intakes can be controlled more efficiently and, consequently, the risk for predisposition to imbalanced nutrition is lower than in loose housing systems. Loose housing might be better suited for older, yearling horses because they have nearly reached their mature size and so are not as susceptible to the effects of low Ta and ad libitum feeding. Another alternative is to regulate individually both concentrate and forage intake in loose-housed weanling horses, and to develop ways of encouraging
their locomotor activity. However, limited forage feeding makes high demands on the planning and implementation of feeding and management strategies for group-housed growing horses.
Knowing that, when given the opportu-nity, horses tend to follow the natural cir-cadian rhythm of behavioural patterns, it should be considered if this could also be allowed in box stabling by adjusting man-agement practices. For example, by taking horses to outdoor paddocks early in the morning at the time when the loose-housed horses tended to wake up and leave the hall, and by taking them into the stable at the time in the evening when the loose-housed horses tended to increase time spent resting in the sleeping hall. Providing shelter from the weather during the daytime and ensur-ing good air quality in the stable are also essential factors to take into account. There-fore, the negative effects of stabling (listed in Table 9 above) can be efficiently pre-vented by good management practices.
In practical terms, it may be said that, like every housing system, loose housing also involves both welfare-enhancing and welfare-threatening factors. Therefore, the suitability of a loose housing system as a winter environment for weanling horses depends on which welfare-related factors are emphasized. In addition, it is evident that a combination of the housing facilities, management practices and the knowledge and skills of the horse keeper determines to a great extent whether loose housing is beneficial or detrimental for weanling horses. In order to ensure the welfare of loose-housed horses, current loose housing conditions and management practices should be improved through general coun-selling and education.

Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 69
8. PRACTICAL RECOMMENDATIONS
• The need for rest is high in weanling horses. It is essential that the loose house is well-bedded and sheltered, and is large enough for the number of horses in order to enable adequate rest also for low-ranking horses.
• Insulation of the loose house and an entrance shelter are recommended, be-cause they contribute to the heat balance of the horses by reducing the adverse effects of weather, and hence reduce climatic energy demand and feeding costs.
• Weanling horses spend a large propor-tion of the day eating forages and may be rather inactive. It is recommended that forage feeds be placed outside in a covered feeder in order to increase the time that horses spend outdoors. By placing forage feeding further from the loose house building, the locomotor ac-tivity of the horses may be enhanced. Other ways to activate the horses should also be considered.
• The effect of acclimatization on cli-matic energy demand has to be taken into account in the plane of feeding of outdoor-housed weanling horses in or-
der to ensure normal growth rate and to avoid overfeeding and excess weight gain. ME intake should be increased by 1.8 % in November, 0.5 % in December and 0.2 % in January above the ME recommendations per 1 °C decrease in Ta below the LCT, whish is estimated to be –11 ºC in loose-housed weanling horses. Existing nutrient recommenda-tions should be modified accordingly.
• Forage feeding should be restricted in late winter if the nutritional value of forages is high, especially when the BCS of weanling horses rises above a moderate level. In that case, special at-tention needs to be paid to the arrange-ment of forage feeding in order to en-sure adequate forage intake also for low-ranking horses.
• Loose house rearing is more recom-mendable for yearling than weanling horses since the former are not as sus-ceptible to the effects of low Ta and ad libitum forage feeding as substantially growing weanling horses.


Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 71
9. REFERENCES
AAPCA (Aberdeen Association for the Prevention of Cruelty to Animals). (1991). The horse in win-ter – shelter and feeding. Aberdeen, UK: The Scottish Agricultural College, Centre for Rural Building.
Ahtila, L., & Saastamoinen, M. (2005). Effect of nutrition on the growth curve of weanling foals. In V. Juliand & W. Martin-Rosset (Eds.). The growing horse: nutrition and prevention of growth disorders (pp. 79-80). EAAP Publication no. 114. AE Wageningen, The Netherlands: Wageningen Academic Publishers.
Airaksinen, S. (2006). Bedding and manure man-agement in horse stables. Its effect on stable air quality, paddock hygiene and the compostability and utilization of manure. Doctoral dissertation. University of Kuopio: Kuopio, Finland.
Alexander, G., Bennett, J.W., & Gemmell, R.T. (1975). Brown adipose tissue in the new-born calf (Bos taurus). The Journal of Physiology, 244 (1), 223–234.
Andrew, J.E., Kline, K.H., & Smith, J.L. (2006). Effects of feed form on growth and blood glu-cose in weanling horses. Journal of Equine Vet-erinary Science, 26, 349–355.
Arnold, W., Ruf, T., & Kuntz, R. (2006). Seasonal adjustment of energy budget in a large wild mammal, the Przewalski horse (Equus ferus przewalskii). The Journal of Experimental Biol-ogy, 209, 4566–4573.
Arnold, W., Ruf, T., Reimoser, S., Tataruch, F., Onderscheka, K., & Schober, F. (2004). Noctur-nal hypometabolism as an overwintering strategy of red deer (Cervys elaphus). American Journal of Physiology: Regulatory, Integrative and Com-parative Physiology, 286, 174–181.
Belknap, M.A. (1997). Horsewords: the Equine dictionary. Vermont, USA: Trafalgar Square Publishing.
Bell, R.A., Nielsen, B.D., Waite, K., Rosenstein, D., & Orth, M. (2001). Daily access to pasture turn-out prevents loss of mineral in the third meta-carpus of Arabian weanlings. Journal of Animal Science, 79, 1142–1150.
Bender, I. (1992). Handbuch Offenstallhaltung. Stuttgart, Germany: Franckh-Kosmos Verlags-GmbH & Co.
Berger, A., Scheibe, K-M., Eichhorn, K., Scheibe, A., & Streich, J. (1999). Diurnal and ultradian rhythms of behaviour in a mare group of Przewalski horse (Equus ferus przewalskii), measured through one year under semi-reserve conditions. Applied Animal Behaviour Science, 64, 1–7.
Bicego, K.C., Barros, R.C.H., & Branco, L.G.S. (2007). Physiology of temperature regulation:
Comparative aspects. Comparative Biochemistry and Physiology, Part A, 147, 616–639.
Bligh, J. (1998). Mammalian homeothermy: an integrative thesis. Journal of Thermal Biology, 23, 143–258.
Bligh, J. (2006). A theoretical consideration of the means whereby the mammalian core temperature is defended at a null zone. Journal of Applied Physiology, 100, 1332–1337.
Boulant, J.A. (2000). Role of the preoptic-anterior hypothalamus in thermoregulation and fever. Clinical Infectious Diseases, 31, S157–161.
Boyd, L. (1998). The 24-h time budget of a takh harem stallion (Equus ferus przewalskii) pre- and post-reintroduction. Applied Animal Behaviour Science, 60, 291–299.
Boyd, L., & Bandi, N. (2002). Reintroduction of takhi, Equus ferus przewalskii, to Hustai National Park, Mongolia: time budget and syn-chrony of activity pre- and post-release. Applied Animal Behaviour Science, 78, 87–102.
Bracke, M.B.M., & Hopster, H. (2006). Assessing the importance of natural behaviour for animal welfare. Journal of Agricultural and Envi-ronmental Ethics, 19, 77–89.
Cannon, B., & Nedergaard, J. (2004). Brown adi-pose tissue: function and physiological signifi-cance. Physiological Reviews, 84, 277–359.
Chaya, L., Cowan, E., & McGuire, B. (2006). A note on the relationship between time spent in turnout and behaviour during turnout in horses (Equus caballus). Applied Animal Behaviour Science, 98, 155–160.
Christensen, J.W., Ladewig, J., Søndergaard, E., & Malmkvist, J. (2002). Effects of individual ver-sus group stabling on social behaviour in domes-tic stallions. Applied Animal Behaviour Science, 75, 233–248.
Clark, J.A., & McArthur, A.J. (1994). Thermal ex-changes. In C.M. Wathes & D.R. Charles (Eds.). Livestock housing (pp. 97-122). Cambridge, UK: Cambridge University Press.
Clarke, A. (1991). Cold adaptation. Journal of Zo-ology, London, 225, 691–699.
Clarke, A.F. (1994). Stables. In C.M. Wathes & D.R. Charles (Eds.). Livestock housing (pp. 379–403). Cambridge, UK: Cambridge University Press.
Cross, D.T., Threlfall, W.R., & Kline, R.C. (1991). Telemetric monitoring of body temperature in the horse mare. Theriogenology, 36 (5), 855–861.
Crowell-Davis, S.L., Houpt, K.A., & Carnevale, J. (1985). Feeding and drinking behaviour of mares and foals with free access to pasture and water. Journal of Animal Science, 60 (4), 883–889.

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 72
Cymbaluk, N.F. (1990). Cold housing effects on growth and nutrient demand of young horses. Journal of Animal Science, 68, 3152–3162.
Cymbaluk, N.F. (1994). Thermoregulation of horses in cold, winter weather: a review. Livestock Pro-duction Science, 40, 65–71.
Cymbaluk, N.F., & Christison, G.I. (1989a). Effects of diet and climate on growing horses. Journal of Animal Science, 67, 48–59.
Cymbaluk, N.F., & Christison, G.I. (1989b). Effects of dietary energy and phosphorus content on blood chemistry and development of growing horses. Journal of Animal Science, 67, 951–958.
Cymbaluk, N.F., & Christison, G.I. (1990). Envi-ronmental effects on thermoregulation and nutri-tion of horses. Veterinary Clinics of North America: Equine Practice. 6 (2), 355–372.
Cymbaluk, N.F., Christison, G.I., & Leach, D.H. (1989a). Energy uptake and utilization by limit- and ad libitum-fed growing horses. Journal of Animal Science, 67, 403–413.
Cymbaluk, N.F., Christison, G.I., & Leach, D.H. (1989b). Nutrient utilization by limit- and ad li-bitum-fed growing horses. Journal of Animal Science, 67, 414–425.
Cymbaluk, N.F., Christison, G.I., & Leach, D.H. (1990). Longitudinal growth analysis of horses following limited and ad libitum feeding. Equine Veterinary Journal, 22 (3), 198–204.
Dahl, L-G., Gillespie, J.R., Kallings, P., Persson, S.G.B., & Thornton, J.R. (1987). Effects of a cold environment on exercise tolerance in the horse. In J.R. Gillespie & N.E. Robinson (Eds.). Equine exercise physiology 2 (pp. 235–242). Davis, USA: ICEEP Publications.
Dallaire, A. (1986). Rest behaviour. Veterinary Clinics of North America: Equine Practice, 2 (3), 591–607.
Danielsson, E.W., Levin, J., & Abrams, E. (1998). Meteorology. New York, USA: McGraw-Hill, Inc.
Davenport, J. (1992). Animal life at low tempera-ture. London, UK: Chapman & Hall.
Dereniak, E.L., & Boreman, G.D. (1996). Infrared detectors and systems. New York, USA: John Wiley & Sons, Inc.
Dieterich, R.A., & Holleman, D.F. (1973). Hematol-ogy, biochemistry, and physiology of envi-ronmentally stressed horses. Canadian Journal of Zoology, 51, 867–873.
Duncan, G. (1990). Physics in the life sciences. Oxford, UK: Blackwell Scientific Publications.
Duncan, P. (1980). Time-budget of Camargue horses II. Time-budgets of adult horses and weaned sub-adults. Behaviour, 72, 26–49.
Duncan, P. (1985). Time-budget of Camargue horses III. Environmental influences. Behaviour, 92, 186–208.
Fader, C., & Sambraus, H.H. (2004). Das Ruhever-halten von Pferden in Offenlaufställen. Tierärtz-liche Umschau, 59, 320–327.
Fazio, E., Medica, P., Cravana, C., Messineo, C., & Ferlazzo, A. (2007). Total and free iodothyronine levels of growing Thoroughbred foals: Effects of weaning and gender. Livestock Science, 110, 207–213.
Forsmark, P. (2006). Fodersammansättningens betydelse för tillväxt hos häst. Examensarbete nr 223. Sveriges Lantbruksuniversitet: Uppsala, Sweden.
Frape, D. (1998). Equine nutrition & feeding. 2nd ed. Oxford, UK: Blackwell Science Ltd.
Fürst, A., Knubben, J., Kurtz, A., Auer, J., & Stauf-facher, M. (2006). Pferde in Gruppenhaltung: Eine Betrachtung aus tierärtzlicher Sicht unter besondered Berücksichtigung des Verletzungsri-sikos. Pferdeheilkunde, 22 (3), 254–258.
Gibbs, P.G., & Cohen, N.D. (2001). Early manage-ment of race-bred weanlings and yearlings on farms. Journal of Equine Veterinary Science, 21, 279–283.
Glade, M.J., & Belling, T.H. Jr. (1984). Growth plate cartilage metabolism, morphology and bio-chemical composition in over- and underfed horses. Growth, 48 (4), 473–482.
Glade, M.J., & Reimers, T.J. (1985). Effects of diet-ary energy supply on serum thyroxine, tri-iodothyronine and insulin concentrations in young horses. Journal of Endocrinology, 104, 93–98.
Glade, M.J., Gupta, S., & Reimers, T.J. (1984). Hormonal responses to high and low planes of nutrition in weanling thoroughbreds. Journal of Animal Science, 59 (3), 658–665.
Gonzales-Jimenez, E., & Blaxter, K.L. (1962). The metabolism and thermal regulation of calves in the first month of life. The British Journal of Nu-trition, 16, 199–212.
Gunnarsson, A., & Ingvarsson, H. (1995). Ettåriga travhästars utnyttjande av ligghall vid utedrift. Examensarbete 68. Sveriges Lantbruksuniversi-tet: Uppsala, Sweden.
Guyton, A.C. (1991). Textbook of medical physiol-ogy. 8th ed. Philadelphia, USA: W.B. Saunders Company.
Haim, A., Rubal, A., & Harari, J. (1993). Compara-tive thermoregulatory adaptations of field mice of the genus Apodemus to habitat challenges. Journal of Comparative Physiology. B, Bio-chemical, Systemic, and Environmental Physiol-ogy, 163 (7), 602–607.
Heiskanen, M-L., Klemola, I., Kumpulainen, M., & Kauppinen, P. (2002). Hevostalous - merkitys ja tulevaisuus Suomessa. 2nd ed. Kuopio, Finland: Kuopion Liikekirjapaino Oy.
Heleski, C.R., Shelle, A.C., Nielsen, B.D., & Zanel-la, A.J. (2002). Influence of housing on weanling

References
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 73
horse behaviour and subsequent welfare. Applied Animal Behaviour Science, 78, 291–302.
Henneke, D.R., Potter, G.D., Kreider, J.L., & Yeates, B.F. (1983). Relationship between condi-tion score, physical measurements and body fat percentage in mares. Equine Veterinary Journal, 15, 371–372.
Hetem, R.S., Maloney, S.K., Fuller, A., Meyer, L.C.R., & Mitchell, D. (2007). Validation of a biotelemetric technique, using ambulatory minia-ture black globe thermometers, to quantify ther-moregulatory behaviour in ungulates. Journal of Experimental Zoology, 307A, 342–356.
Hines, M.T. (2004). Changes in body temperature. In S.M. Reed and W.M. Bayly (Eds.). Equine in-ternal medicine (pp. 148–155). St. Louis, USA: Elsevier.
Hiney, K.M., Nielsen, B.D., & Rosenstein, D. (2004). Short-duration exercise and confinement alters bone mineral content and shape in wean-ling horses. Journal of Animal Science, 82, 2312–2320.
Hodgson, D.R., Davis, R.E., & McConaghy, F.F. (1994). Thermoregulation in the horse in re-sponse to exercise. British Veterinary Journal, 150, 219–235.
Hodgson, D.R., McCutcheon, L.J., Byrd, S.K., Brown, W.S., Bayly, W.M., Brengelmann, G.L., & Gollnick, P.D. (1993). Dissipation of meta-bolic heat in the horse during exercise. Journal of Applied Physiology, 74, 1161–1170.
Hogan, E.S., Houpt, K.A., & Sweeney, K. (1988). The effect of enclosure size on social interactions and daily activity patterns of the captive Asiatic wild horse (Equus przewalskii). Applied Animal Behaviour Science, 21, 147–168.
Holcombe, S.J., Jackson, C., Gerber, V., Jefcoat, A., Berney, C., Eberhardt, S., & Robinson, N.E. (2001). Stabling is associated with airway in-flammation in young Arabian horses. Equine Veterinary Journal, 33 (3), 244–249.
Hotchkiss, J.W., Reid, S.W.J., & Christley, R.M. (2007). A survey of horse owners in Great Brit-ain regarding horses in their care. Part 1: Horse demographic characteristics and management. Equine Veterinary Journal, 39 (4), 294–300.
Houpt, K.A., Eggleston, A., Kunkle, K., & Houpt, T.R. (2000). Effect of water restriction on equine behaviour and physiology. Equine Veterinary Journal, 32 (4), 341–344.
Houpt, K., Houpt, T.R., Johnson, J.L., Erb, H.N., & Yeon, S.C. (2001). The effect of exercise depri-vation on the behaviour and physiology of straight stall confined pregnant mares. Animal Welfare, 10 (3), 257–267.
Hänninen, L., Hepola, H., Rushen, J., de Passillé, A.M., Pursiainen, P., Tuure, V-M., Syrjälä-Qvist, L., Pyykkönen, M., & Saloniemi, H. (2003). Resting behaviour, growth and diarrhoea inci-dence rate of young dairy calves housed indi-
vidually or in groups in warm or cold buildings. Acta Acrigulturae Scandinavica, Section A, Ani-mal Science, 53, 21–28.
Ingólfsdóttir, H., & Sigurjónsdóttir, H. (2008). The benefits of high rank in the wintertime –A study of the Icelandic horse. Applied Animal Behaviour Science, 114 (3-4), 485–491.
Irvine, C.H.G. (1967). Thyroxine secretion rate in the horse in various physiological states. Journal of Endocrinology. 39, 313–320.
IUPS Thermal Commission (2003). Glossary of terms for thermal physiology. Journal of Ther-mal Physiology. 28, 75–106.
Jezierski, T., Jaworski, Z., & Górecka, A. (1999). Effects of handling on behaviour and heart rate in Konik horses: comparison of stable and forest reared youngstock. Applied Animal Behaviour Science, 62, 1–11.
Johansson, L. & Lindström, K. (2008). En pi-lotstudie av hästars rörelsemängd i stor och liten hage. Fördjupningsarbete nr 343. Sveriges Lant-bruksuniversitet: Uppsala, Sweden.
Jørgensen, G.H.M., & Bøe, K.E. (2007). A note on the effect of daily exercise and paddock size on the behaviour of domestic horses (Equus cabal-lus). Applied Animal Behaviour Science, 107 (1-2), 166–173.
Kainer, R.A., & McCracken, T.O. (1994). The color-ing atlas of horse anatomy. Loveland, CO, USA: Alpine Publications Inc.
Kauppinen, R. (2000). Acclimatization of dairy calves to a cold and variable micro-climate. Doctoral dissertation. University of Kuopio: Kuopio, Finland.
Keren, E.N., & Olson, B.E. (2007). Applying ther-mal imaging software to cattle grazing winter range. Journal of Thermal Biology, 32, 204–211.
Kiley-Worthington, M. (1987). The behavior of horses in relation to management and training. London, UK: J.A. Allen.
Klaus, S., Heldmaier, G., & Ricquier, D. (1988). Seasonal acclimation of bank voles and wood mice: nonshivering thermogenesis and ther-mogenic properties of brown adipose tissue mi-tochondria. Journal of Comparative Physiology. B, Biochemical, Systemic, and Environmental Physiology, 158 (2), 157–164.
Kohn, C.W., Hinchcliff, K.W., & McKeever, K.H. (1999). Evaluation of washing with cold water to facilitate heat dissipation in horses exercised in hot, humid conditions. American Journal of Vet-erinary Research, 60 (3), 299–305.
Kristula, M.A., & McDonnell, S.M. (1994). Drink-ing water temperature affects consumption of water during cold weather in ponies. Applied Animal Behaviour Science, 41, 155–160.
Kuntz, R., Kubalek, C., Ruf, T., Tataruch, F., & Arnold, W. (2006). Seasonal adjustment of en-ergy budget in a large wild mammal, the

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 74
Przewalski horse (Equus ferus przewalskii), The Journal of Experimental Biology, 209, 4557–4565.
Kurvers, C.M., van Weeren, P.R., Rogers, C.W., & van Dierendonck, M.C. (2006). Quantification of spontaneous locomotion activity in foals kept in pastures under various management conditions. American Journal of Veterinary Research, 67 (7), 1212–1217.
Langlois, B. (1994). Inter-breed variation in the horse with regard to cold adaptation: a review. Livestock Production Science, 40, 1–7.
Lovegrove, B.G. (2005). Seasonal thermoregulatory responses in mammals. Journal of Comparative Physiology. B, Biochemical, Systemic, and Envi-ronmental Physiology, 175, 231–247.
MacCormack, J.A.D., & Bruce, J.M. (1991). The horse in winter – shelter and feeding. Farm Building Progress, 105, 10–13.
Markussen, K.A., Rognmo, A., & Blix, A.S. (1985). Some aspects of thermoregulation in newborn reindeer calves (Rangifer tarandus tarandus). Acta Physiologica Scandinavica, 123 (2), 215–220.
Martin, P., & Bateson, P. (1993). Measuring behav-iour: an introductory guide. 2nd ed. Cambridge, UK: Cambridge University Press.
McArthur, A.J. (1991). Metabolism of homeotherms in the cold and estimation of thermal insulation. Journal of Thermal Biology, 16 (3), 149–155.
McBride, G.E., Christopherson, R.J., & Sauer, W. (1985). Metabolic rate and plasma thyroid hor-mone concentrations of mature horses in re-sponse to changes in ambient temperature. Ca-nadian Journal of Animal Science, 65, 375–382.
McDonald, P., Edwards, R.A., Greenhalgh, J.F.D., & Morgan, C.A. (2002). Animal nutrition. 6th ed. Essex, UK: Pearson Education Limited.
McDonnell, S. (2003). The Equid ethogram: a prac-tical field guide to horse behaviour. Lexington, USA: The Blood-Horse, Inc.
McGreevy, P. (2004). Equine behavior: a guide for veterinarians and Equine scientists. London, UK: Saunders Book Company.
McGreevy, P.D., French, N.P., & Nicol, C.J. (1995). The prevalence of abnormal behaviours in dres-sage, eventing and endurance horses in relation to stabling. The Veterinary Record, 137, 36–37.
Mejdell, C.M., & Bøe, K.E. (2005). Responses to climatic variables of horses housed outdoors un-der Nordic winter conditions. Canadian Journal of Animal Science, 85, 301–308.
Michanek, P., & Ventorp, M. (1996). Time spent in shelter in relation to weather by two free-ranging Thoroughbred yearlings during winter. Applied Animal Behaviour Science, 49 (1), 104.
Mills, D.S., & Clarke, A. (2002). Housing, manage-ment and welfare. In N. Waran (Ed.). The wel-fare of horses (pp. 77–97). Dordrecht, The Neth-erland: Kluwer Academic Publishers.
Ministry of Agriculture and Forestry decree on ani-mal welfare requirements for horses (14/EEO/1998). Maa- ja metsätalousministeriön päätös hevosten pidolle asetettavista eläinsuoje-luvaatimuksista (14/EEO/1998).
Ministry of Agriculture and Forestry statute on ani-mal protection (396/1996). Maa- ja metsätalous-ministeriön eläinsuojeluasetus (396/1996).
Ministry of Agriculture and Forestry statute on feed control (3/2006). Maa- ja metsätalousministeriön asetus rehujen valvonnan järjestämisestä (3/2006).
Mogg, K.C., & Pollitt, C.C. (1992). Hoof and distal limb surface temperature in the normal pony un-der constant and changing ambient temperatures. Equine Veterinary Journal, 24 (2), 134–139.
Morgan, K. (1995). Climatic energy demand of horses. Equine Veterinary Journal, Supplement, 18, 396–399.
Morgan, K. (1996). Short-term thermoregulatory responses of horses to brief changes in ambient temperature. Doctoral dissertation. Swedish University of Agricultural Sciences: Uppsala, Sweden.
Morgan, K. (1997a). Thermal insulance of periph-eral tissue and coat in sport horses. Journal of Thermal Biology, 22 (3), 169–175.
Morgan, K. (1997b). Effects of short-term changes in ambient air temperature or altered insulation in horses. Journal of Thermal Biology, 22 (3), 187–194.
Morgan, K. (1998). Thermoneutral zone and critical temperatures of horses. Journal of Thermal Biol-ogy, 23 (1), 59–61.
Morgan, K., Aspång, L., & Holmgren, S. (2007). Lower critical temperature of competition horses. In EAAP 2007 [online]. Retrieved Sep-tember 30 2007, from: http://www.eaap.org/Dublin/Sessions /session_12.htm
Morgan, K., Ehrlemark, A., & Sällvik, K. (1997). Dissipation of heat from standing horses exposed to ambient temperatures between –3 °C and 37 °C. Journal of Thermal Biology, 22 (3), 177–186.
Morgan, K., Funkquist, P., & Nyman, G. (2002). The effect of coat clipping on thermoregulation during intense exercise in trotters. Equine Vet-erinary Journal. Supplement, 34, 564–567.
Mount, L.E. (1973). The concept of thermal neutral-ity. In J.L. Monteith & L.E. Mount (Eds.). Heat loss from animals and man (pp. 425–439). Lon-don, UK: Butterworths.
MTT (2006). Rehutaulukot ja ruokintasuositukset [online]. Agrifood Research Finland, Jokioinen. Retrieved August 8, 2007, from: http://www.agronet.fi/rehutaulukot/
Neste, R. (2000). Hevosen lämmönluovutus eri vuo-denaikoina. Master’s thesis. University of Ku-opio: Kuopio, Finland.

References
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 75
Ninomiya, S., Sato, S., & Sugawara, K. (2007). Weaving in stabled horses and its relationship to other behavioural traits. Applied Animal Behav-iour Science, 106, 134–143.
Normando, S., Canali, E., Ferrante, V., & Verga, M. (2002). Behavioural problems in Italian saddle horses. Journal of Equine Veterinary Science, 22 (3), 117–120.
Nousiainen, J., Rinne, M., Hellämäki, M., & Hu-htanen, P. (2003). Prediction of the digestibility of primary growth and regrowth grass silages from chemical composition, pepsin-cellulase solubility and indigestible cell wall content. Animal Feed Science and Technology, 110, 61–74.
NRC (2007). Nutrient requirements of horses. Washington, DC, USA: The National Academies Press.
Ousey, J.C., McArthur, A.J., & Rossdale, P.D. (1991). Metabolic changes in Thoroughbred and pony foals during the first 24 h post partum. Journal of Reproduction and Fertility. Supple-ment. 44, 561–570.
Ousey, J.C., McArthur, A.J., & Rossdale, P.D. (1997). Thermoregulation in sick foals aged less than one week. The Veterinary Journal, 153, 185–196.
Ousey, J.C., McArthur, A.J., Murgatroyd, P.R., Stewart, J.H., & Rossdale, P.D. (1992). Ther-moregulation and total body insulation in the neonatal foal. Journal of Thermal Biology, 17 (1), 1–10.
Pagan, J.D., & Hintz, H.F. (1986). Equine energet-ics. I. Relationship between body weight and en-ergy requirements in horses. Journal of Animal Science, 63, 815–821.
Palmer, S.E. (1983). Effect of ambient temperature upon the surface temperature of the equine limb. American Journal of Veterinary Research, 44 (6), 1098–1101.
Parker M., Goodwin, D., & Redhead, E.S. (2008). Survey of breeders’ management of horses in Europe, North America and Australia: Compari-son of factors associated with the development of abnormal behaviour. Applied Animal Behaviour Science, 114 (1-2), 206–215.
Phillips, P.K., & Heath, J.E. (1995). Dependency of surface temperature regulation on body size in terrestrial mammals. Journal of Thermal Biol-ogy, 20 (3), 281–289.
Randall, D., Burggren, W., & French, K. (2001). Animal physiology: mechanisms and adapta-tions. New York, USA: W.H. Freeman and Company.
Raub, R.H., Jackson, S.G. & Baker, J.P. (1989). The effect of exercise on bone growth and develop-ment in weanling horses. Journal of Animal Sci-ence, 67, 2508–2514.
Reiss, M.J. (1991). The allometry of growth and reproduction. Cambridge, UK: Cambridge Uni-versity Press.
Rivera, E., Benjamin, S., Nielsen, J., Shelle, J., & Zanella, A.J. (2002). Behavioral and physiologi-cal responses of horses to initial training: the comparison between pastured versus stalled horses. Applied Animal Behaviour Science, 78, 235–252.
Robinson, N.E., Karmaus, W., Holcombe, S.J., Carr, E.A., & Derksen, F.J. (2006). Airway inflamma-tion in Michigan pleasure horses: prevalence and risk factors. Equine Veterinary Journal, 38 (4), 293–299.
Rogers, C.W., Firth, E.C., McIlwraith, C.W., Barneveld, A., Goodship, A.E., Kawcak, C.E., Smith, R.K.W., & van Weeren, P.R. (2008). Ev-aluation of a new strategy to modulate skeletal development in Thoroughbred performance horses by imposing track-based exercise during growth. Equine Veterinary Journal, 40 (2), 111–118.
Saarela, S., & Hissa, R. (1993). Metabolism, thermogenesis and daily rhythm of body tem-perature in the wood lemming, Myopus schisti-color. Journal of Comparative Physiology. B, Biochemical, Systemic, and Environmental Physiology, 163 (7), 546–555.
Saastamoinen, M.T., & Koskinen, E. (1993). Influ-ence of quality of dietary protein supplement and anabolic steroids on muscular and skeletal growth of foals. Animal Production, 56, 135–144.
Sandgren, B., Dalin, G., Carlsten, J., & Lundeheim, N. (1993). Development of osteochondrosis in the tarsocrural joint and osteochondral fragments in the fetlock joints of Standardbred trotters. II. Body measurements and clinical findings. Eq-uine Veterinary Journal. Supplement, 16, 48–53.
Scott, D.W. (1988). Large animal dermatology. Philadelphia, USA: WB Saunders.
Sjaastad, Ø.V., Hove, K., & Sand., O. (2003). Physiology of domestic animals. Oslo, Norway: Scandinavian Veterinary Press.
SLU (2004). Utfodringsrekommendationer för häst. Uppsala, Sweden: Sveriges Lantbruksuniversitet.
Søndergaard, E., & Halekoh, U. (2003). Young horses’ reactions to humans in relation to han-dling and social environment. Applied Animal Behaviour Science, 84, 265–280.
Søndergaard, E., & Ladewig, J. (2004). Group hous-ing exerts a positive effect on the behaviour of young horses during training. Applied Animal Behaviour Science, 87 (1-2), 105–118.
Soppela, P., Nieminen, M., Saarela, S., Keith, J.S., Morrison, J.N., Macfarlane, F., & Trayhurn, P. (1991). Brown-fat specific mitochondrial uncou-pling protein in adipose tissues of newborn rein-deer. The American Journal of Physiology, 260, R1229–R1234.

Autio Elena: Loose Housing of Horses in a Cold Climate
Kuopio Univ. Publ. C. Nat. and Environ. Sci. 245: 1-76 (2008) 76
Soppela, P., Sormunen, R., Saarela, S., Huttunen, P., & Nieminen, M. (1992). Localization, cellular morphology and respiratory capacity of “brown” adipose tissue in newborn reindeer. Comparative Biochemistry and Physiology. Comparative Physiology, 101 (2), 281–293.
Storer, T.I., Usinger, R.L., Stebbins, R.C., & Ny-bakke, J.W. (1979). General zoology. New York, USA: McGraw-Hill, Inc.
Suomen Hippos ry (2008). Hevoskannan kehitys maassamme 1920-2007 [online]. Retrieved June 03, 2008, from: http://www.hippos.fi/hippos/tilastot/jalostus_ja_kasvatus/hevoskannan_kehitys.php
Talukdar, A.H., Calhoun, M.L., & Stinson, A.W. (1972). Microscopic anatomy of the skin of the horse. American Journal of Veterinary Research, 33 (12), 2365–2390.
Tanaka, M., Tonouchi, M., Hosono, T., Nagashima, K., Yanase-Fujiwara, M., & Kanosue, K. (2001). Hypothalamic region facilitating shivering in rats. Japanese Journal of Physiology, 51, 625–629.
Trayhurn, P., & Jennings, G. (1988). Nonshivering thermogenesis and the thermogenic capacity of brown fat in fasted and/or refed mice. The American Journal of Physiology, 254, R11-R16.
Tremblay, G.M., Ferland, C., Lapointe, J.M., Vrins, A., Lavoie, J.P., & Cormier, Y. (1993). Effect of stabling on bronchoalveolar cells obtained from normal and COPD horses. Equine Veterinary Journal, 25 (3), 194–197.
Tuomivaara, S. (1992). Hevostallien ilmahygienia. Master’s thesis. University of Kuopio: Kuopio, Finland.
van de Lest, C.H.A, Brama, P.A.J. & van Weeren, P.R. (2002). The influence of exercise on the composition of developing equine joints. Biorhe-ology, 39, 183–191.
van de Lest, C.H.A, Brama, P.A.J., & van Weeren, P.R. (2003). The influence of exercise on bone morphogenic enzyme activity of immature eq-uine subchondral bone. Biorheology, 40, 377–382.
van Dierendonck, M.C., Bandi, N., Batdorj, D., Dügerlham, S., & Munkhtsog, B. (1996). Behav-ioural observations of reintroduced Takhi or
Przewalski horses (Equus ferus przewalskii) in Mongolia. Applied Animal Behaviour Science, 50, 95–114.
van Weeren, P.R., Firth, E.C., Brommer, H., Hytti-nen, M.M., Helminen, H.J., Rogers, C.W., De-Groot, J., & Brama, P.A. (2008). Early exercise advances the maturation of glycosaminoglycans and collagen in the extracellular matrix of artigu-lar cartilage in the horse. Equine Veterinary Journal. 40 (2), 128–135.
Ventorp, M. (1994). Grupphållna hästar ute är “inne” – hästar I lösdrift. Fakta – 2. Uppsala, Sweden: Sveriges Lantbruksuniversitet.
Vermorel, M., Vernet, J., & Martin-Rossett, W. (1997). Digestive and energy utilisation of two diets by ponies and horses. Livestock Production Science, 51, 13–19.
Vermorel, M., Dardillat, C., Vernet, J., Saido, Demigne, C. (1983). Energy metabolism and thermoregulation in the newborn calf. Annals of Veterinary Research, 14 (4), 382–389.
Vernet, J., Vermorel, M., & Martin-Rossett, W. (1995). Energy cost of eating long hay, straw and pelleted food in sport horses. Animal Science, 61, 581–588.
Verschooten, F., Desmet, P., & Verbeeck, J. (1997). Skin surface temperature measurements in horses by electronic thermometry. Equine Practice, 19 (4), 16–23.
Waters, A.J., Nicol, C.J., & French, N.P. (2002). Factors influencing the development of stereo-typic and redirected behaviours in young horses: findings of a four-year prospective epidemio-logical study. Equine Veterinary Journal, 34 (6), 572–579.
Young, B.A. (1975). Some physiological costs of cold climates. Special report 175. Missouri, USA: University of Missouri, Agricultural Ex-periment Station.
Young, B.A., & Coote, J. (1973). Some effects of cold on horses. Horse report at Feeders’ Day. Alberta, Canada: University of Alberta, Depart-ment of Animal Science.
Zeeb, K., & Schnitzer, U. (1997). Housing and train-ing of horses according to their species-specific behaviour. Livestock Production Science, 49, 181–189.

Kuopio University Publications C. Natural and Environmental Sciences C 218. Madetoja, Elina. Novel process line approach for model-based optimization in papermaking. 2007. 125 p. Acad. Diss. C 219. Hyttinen, Marko. Formation of organic compounds and subsequent emissions from ventilation filters. 2007. 80 p. Acad. Diss. C 220. Plumed-Ferrer, Carmen. Lactobacillus plantarum: from application to protein expression. 2007. 60 p. Acad. Diss. C 221. Saavalainen, Katri. Evaluation of the mechanisms of gene regulation on the chromatin level at the example of human hyaluronan synthase 2 and cyclin C genes. 2007. 102 p. Acad. Diss. C 222. Koponen, Hannu T. Production of nitrous oxide (N2O) and nitric oxide (NO) in boreal agricultural soils at low temperature. 2007. 102 p. Acad. Diss. C 223. Korkea-aho, Tiina. Epidermal papillomatosis in roach (Rutilus rutilus) as an indicator of environmental stressors. 2007. 53 p. Acad. Diss. C 224. Räisänen, Jouni. Fourier transform infrared (FTIR) spectroscopy for monitoring of solvent emission rates from industrial processes. 2007. 75 p. Acad. Diss. C 225. Nissinen, Anne. Towards ecological control of carrot psyllid (Trioza apicalis). 2008. 128 p. Acad. Diss. C 226. Huttunen, Janne. Approximation and modellingerrors in nonstationary inverse problems. 2008. 56 p. Acad. Diss. C 227. Freiwald, Vera. Does elevated ozone predispose northern deciduous tree species to abiotic and biotic stresses? 2008. 109 p. Acad. Diss. C 228. Semenov, Dmitry. Distance sensing with dynamic speckles. 2008. 63 p. Acad. Diss. C 229. Höytö, Anne. Cellular responses to mobile phone radiation: proliferation, cell death and related effects. 2008. 102 p. Acad. Diss. C 230. Hukkanen, Anne. Chemically induced resistance in strawberry (Fragaria × ananassa) and arctic bramble (Rubus arcticus): biochemical responses and efficacy against powdery mildew and downy mildew diseases. 2008. 98 p. Acad. Diss. C 231. Hanhineva, Kati. Metabolic engineering of phenolic biosynthesis pathway and metabolite profiling of strawberry (Fragaria × ananassa). 2008. 80 p. Acad. Diss. C 232. Nissi, Mikko. Magnetic resonance parameters in quantitative evaluation of articular cartilage: studies on T₁ and T₂ relaxation time. 2008. 83 p. Acad. Diss.