investigating hepadnaviral capsid envelopment and …
TRANSCRIPT

INVESTIGATING HEPADNAVIRAL CAPSID
ENVELOPMENT AND VIRION PRODUCTION
Natalie J. Greco
A dissertation submitted in partial fulfillment of the requirements for
the degree of
Doctor of Philosophy
(Cellular and Molecular Pathology)
at the
UNIVERSITY OF WISCONSIN-MADISON
2015
Date of final oral examination: 12/11/2014
The dissertation is approved by the following members of the Final Oral Committee: Daniel D. Loeb, Professor, Oncology Paul Ahlquist, Professor, Oncology and Molecular Virology Shannon Kenney, Professor, Oncology and Medicine William Sugden, Professor, Oncology Marulasiddappa Suresh, Associate Professor, Pathobiological Sciences

i
Abstract
Hepadnaviruses selectively package capsids containing mature dsDNA genomes
into virions. The research presented in this dissertation provides insight into this poorly
understood aspect of viral replication. Snow goose hepatitis B virus (SGHBV) is the only
known hepadnavirus that packages capsids containing immature ssDNA into virions. I
found that cells replicating SGHBV produce virions containing ssDNA as efficiently as
virions containing dsDNA and that they support high levels of virion production,
compared to DHBV. I determined that SGHBV capsid protein (Cp) and large envelope
protein (L) independently contribute to the ability of SGHBV to produce virions
containing ssDNA with Cp making a larger contribution. Also, I found that L contributes
to the high levels of virion production characteristic of SGHBV. I conferred these
properties onto DHBV by substituting regions of the SGHBV proteins into corresponding
DHBV proteins, allowing me to identify residues within Cp and L that are responsible for
the different properties of these viruses.
I identified two amino acid residues of DHBV Cp that contribute to selective
dsDNA virion production and may interact with the envelope proteins during virion
formation. Additionally, I identified a region of DHBV L that contributes to selective
dsDNA virion production. I found that this same region of L was also responsible for
DHBV’s relatively low levels of virion production. Future studies on the role of these
residues in virion production will broaden our understanding of this aspect of virus
replication.
Finally, I found that HHBV envelope proteins cannot package DHBV or SGHBV
capsids into virions. I used this incompatibility to identify residues of Cp involved in

ii
virion formation. I substituted a small segment of HHBV Cp into DHBV Cp and this
restored the ability of HHBV envelope proteins to package DHBV capsids into virions.
Residues within this segment likely interact with envelope proteins during virion
morphogenesis. Interestingly, this segment contains the residues of Cp responsible for
selective dsDNA virion production. A similar approach can be taken to identify regions
of the envelope proteins involved in capsid packaging and virion production.

iii
Acknowledgements
First and foremost, I would like to thank Dan Loeb. I feel extremely fortunate to
have had Dan Loeb as my dissertation advisor. I am certain that I would not have gotten
as much out of this process, had I done my dissertation research anywhere but at Loeb
University. Dan’s enthusiasm for the research he does is inspiring. Always leading by
example, Dan trained me to think critically and creatively about my work, and the work
of others. He could be “harsh” at times but he was always “fair”. I appreciated this trait in
him because it helped me to identify and work on my weaknesses to become a better
scientist. Dan provided me with just the right balance of freedom to explore what
interested me and guidance to ensure that I accomplished my goals. Most importantly,
he taught me to think thoroughly about every single thing I do. Dan, I am forever
appreciative of the time and effort you put forth to help me become the scientist I am
today. I will miss you and cannot thank you enough for the lessons you taught me in lab
and in life!
I would also like to thank the current members of Loeb University, Karolyn Pionek
and Nuruddin Unchwaniwala, for always being eager to discuss interesting results and
provide feedback on my research. Thank you for making Loeb U such an awesome and
enjoyable place to be and for always being there if I needed anything, in or out of lab!
This has certainly been the best time of my life, in part because I got to come to the lab
and see you two and Dan every day! Thanks for everything, I will miss you both so
much!
I would like to thank Mike Hayes, a former research technician in our lab, for
providing his technical support when I first joined the lab and for his contributions to the

iv
early stages of my dissertation research. Mike was the first to suggest that we start
using SGHBV to study capsid maturation and selective virion production. He made
several of the initial SGHBV plasmids, from which all of the subsequent SGHBV
plasmids were derived. I would also like to thank former graduate students, Thomas
Lentz and Eric Lewellyn, who were senior graduate students when I joined the lab and
were great graduate student role models.
I would like to thank my committee members, Paul Ahlquist, Shannon Kenney,
Bill Sugden and M Suresh, for providing me with valuable feedback and advice at my
committee meetings. I am additionally grateful to Bill for taking time to share his
thoughts on choosing an appropriate post-doctoral research position.
I feel fortunate to have been able to do my dissertation research at McArdle
Laboratory for Cancer Research. I would like to acknowledge the founding fathers (and
mother) of McArdle Laboratory. These scientists deserve credit for shaping the McArdle
community into what it is today. McArdle is, and always has been, a very collaborative
and supportive community of researchers who are deeply invested in developing the
scientific abilities of their graduate students and post-docs. I would like to thank
everyone at McArdle for making this such a great place to work… from the janitors to
the administrative staff to the director of the department. I’d particularly like to mention
the ladies from Lambertville, my Beatle buddy Gaye and our dear friend Jodi, and all of
the CMP and cancer biology graduate students!
I would also like to thank all of my friends and family in Chicago for always
offering their words of encouragement throughout my time in graduate school. I am
grateful to my parents for always making my education a priority. Finally, I would like to

v
thank all of the friends that I have made in Madison; in particular Toni, Heather, Ben and
the ladies at the Lakehouse! I will miss you all and thanks for all the good times!

vi
Table of Contents
Abstract .................................................................................................................i
Acknowledgements ............................................................................................ iii
Table of Contents ............................................................................................... vi
List of Figures .................................................................................................... ix
List of Abbreviations .......................................................................................... xi
Chapter 1. Hepatitis B Virus Background and Introduction ............................1
HBV transmission, disease and treatment ..................................................2
Hepadnaviridae ..........................................................................................5
Genome organization and viral proteins .....................................................9
Viral replication ........................................................................................ 16
Selective production of virions containing mature dsDNA ........................ 24
Chapter 2: Materials and Methods ................................................................... 26
Chapter 3: Snowgoose Hepatitis B Virus (SGHBV) Capsid and
Envelope Proteins Contribute to the Ability of SGHBV to Package
Capsids Containing ssDNA in Virions ............................................................. 35
Abstract .................................................................................................... 36
Importance................................................................................................ 38
Introduction ............................................................................................... 39
Results ..................................................................................................... 43
Rationale ............................................................................................. 43
Characterizing SGHBV virion production ............................................ 43

vii
Cp contributes to SGHBV’s ability to efficiently package capsids
containing ssDNA in virions ........................................................... 45
Residues 74 and 107 of Cp contribute to DHBVs ability to
selectively package capsids containing mature dsDNA
genomes in virions ......................................................................... 47
Residues 74 and 107 of Cp contribute to SGHBVs ability to
efficiently produce virions containing ssDNA ................................. 49
Changing residues 74 and 107 of HHBV Cp does not confer the
ability to produce virions containing ssDNA onto HHBV ................ 50
SGHBV envelope proteins are sufficient to cause high levels of
virion production and the production of virions containing
ssDNA ............................................................................................ 51
A determinant within residues 61 and 120 of SGHBV L
contributes to the high levels of virion production and the
production of virions containing ssDNA characteristic of
SGHBV .......................................................................................... 52
Discussion ................................................................................................ 65
Chapter 4: Identifying Amino Acid Residues of Avihepadnaviral
Capsid and Envelope Proteins That Contribute to the Packaging of
Capsids into Virions .......................................................................................... 73
Abstract .................................................................................................... 74
Introduction ............................................................................................... 75
Results ..................................................................................................... 78

viii
HHBV envelope proteins cannot package SGHBV or DHBV
capsids into virions ........................................................................ 78
A determinant within amino acid residues 69 and 114 of Cp is
involved in virion production ........................................................... 80
Discussion ................................................................................................ 87
Chapter 5: Summary and Future Directions ................................................... 88
References ......................................................................................................... 95

ix
List of Figures
Figure 1.1. Structure of HBV viral particles ...........................................................8
Figure 1.2. Coding organization and viral transcripts .......................................... 12
Figure 1.3. Capsid protein (Cp) structure and capsid assembly .......................... 13
Figure 1.4. DHBV large (L) and small (S) surface protein topologies .................. 15
Figure 1.5. Viral replication strategy .................................................................... 21
Figure 1.6. Genome replication strategy ............................................................. 23
Figure 3.1. SGHBV supports high levels of virion production and efficiently
produces virions containing ssDNA .......................................................... 56
Figure 3.2. SGHBV Cp contributes to the efficient production of virions
containing ssDNA, while SGHBV envelope proteins contribute to the high
levels of virion production, characteristic of SGHBV ........................................... 57
Figure 3.3. Residues 74 and 107 of DHBV Cp contribute to the selective
production of virions containing dsDNA .................................................... 60
Figure 3.4. Residues 74 and 107 of SGHBV Cp contribute to the
production of virions containing ssDNA ............................................................... 61
Figure 3.5. Changing residues 74 and 107 of HHBV Cp is not sufficient to
cause the production of virions containing ssDNA .............................................. 62
Figure 3.6. A determinant within residues 61 and 120 of L contributes to
the selective production of virions containing dsDNA ............................... 63
Figure 3.7. Phylogenetic tree based on capsid protein amino acid
sequence .................................................................................................. 64

x
Figure 4.1. HHBV envelope proteins cannot package SGHBV capsids into
virions. A determinant between amino acid residues 22 and 139 of
Cp contributes to virion production ........................................................... 82
Figure 4.2. HHBV envelope proteins cannot package DHBV capsids into
virions. A determinant between amino acid residues 69 and 114 of
Cp contributes to virion production ........................................................... 84
Figure 4.3. Region of the capsid protein (Cp) found to contribute to virion
production contains the residues of Cp involved in selective production of
virions containing dsDNA .................................................................................... 86

xi
List of Abbreviations
Cp capsid protein
cccDNA covalently closed circular DNA
DHBV duck hepatitis B virus
DL DNA duplex linear DNA
DR1,2 direct repeat 1, 2
ε epsilon – cis-acting sequence
HBeAg hepatitis B virus e-antigen
HBV human hepatitis B virus
HCC hepatocellular carcinoma
HHBV heron hepatitis B virus
HIV human immunodeficiency virus
iRC DNA incomplete relaxed circular DNA
kb kilobase
kDa kilodalton
L large surface/envelope protein
M medium surface/envelope protein
nt(s) nucleotide(s)
ORF open reading frame
P viral reverse transcriptase/polymerase protein
pgRNA pregenomic RNA
RC DNA relaxed circular DNA
S small surface/envelope protein

xii
SGHBV snow goose hepatitis B virus
sgRNA subgenomic RNA
SS DNA single stranded DNA
TP terminal protein domain of P
vDNA virion associated DNA
vRI encapsidated viral replicative intermediate
WHV woodchuck hepatitis B virus
WT wild type

1
1
2
CHAPTER 1 3
4
5
HEPATITIS B VIRUS BACKGROUND AND INTRODUCTION 6
7

2
HBV Transmission, Disease and Treatment 8
As its name implies, human hepatitis B virus (HBV) causes inflammation of the liver. 9
HBV can cause an acute or a chronic infection. During a chronic infection, viral 10
replication persists and the prolonged inflammation and hepatocyte cell death caused 11
by the ongoing infection can lead to severe liver disease, such as liver cirrhosis or 12
hepatocellular carcinoma (HCC). Acute infections are typically resolved naturally without 13
treatment and only cause mild disease. HBV can be transmitted through infected blood 14
and bodily fluids. Globally, the most common route of transmission is from mother to 15
child at birth. It is estimated that 200-500 million individuals are chronically infected 16
worldwide, with around one million people dying each year from diseases associated 17
with chronic infection1–4. Prevalence of HBV infection varies greatly between geographic 18
locations. HBV is most prevalent in Sub-Saharan Africa and is less of a concern in the 19
United States (and other high income countries). It is estimated that in 2005, greater 20
than 8% of the population was chronically infected in Sub-Saharan Africa, while less 21
than 2% of the population was chronically infected in the United States5. 22
Interestingly, whether HBV causes acute versus chronic infection in an individual is 23
largely dependent on the age at which infection occurs. Around 90% of individuals 24
infected as an adult will clear the infection. In stark contrast to this, only 5-10% of those 25
infected at birth or as an infant will clear the infection; the large majority of these 26
individuals will become life-long chronic carriers of the virus and many will die of 27
diseases associated with chronic infection. The risk for chronic HBV infection decreases 28
for children between the ages 1 and 4; 30% of those infected will become chronically 29

3
infected. Hence, the risk of chronicity is inversely related to the age at which infection 30
occurs6–8. 31
Individuals chronically infected with HBV are at an increased risk for developing and 32
ultimately dying from HCC9,10. HCC is the sixth most common form of cancer worldwide 33
and the third most common cause of death by cancer11. It is thought that the 34
inflammatory immune response to infection and high rate of cell turnover contribute to 35
the development and/or maintenance of HCC. However, the mechanism by which HBV 36
contributes is not defined. Because chronic HBV infection is one of the leading causes 37
of HCC, the best way to decrease the incidence of HCC is to prevent new HBV 38
infections through the use of vaccines. 39
A safe and effective vaccine was introduced in the early 1980s which has led to a 40
dramatic decrease in HBV prevalence and a decrease in HCC incidence in many parts 41
of the world5,12. However, because vaccination rates are low in some rural and low 42
income parts of the world chronic HBV infection remains a major world health concern. 43
Further, the vaccine is prophylactic and is ineffective at treating individuals already 44
chronically infected. 45
A way to decrease HBV associated mortality is through the use of anti-viral 46
therapies. There are seven approved antiviral agents to treat chronic infection, with 47
more in preclinical and clinical trials13–15. Current therapies can be divided into two 48
types; 1) nucleos(t)ide analogues (NAs) targeted at the viral DNA polymerase, such as 49
entecavir or tenofovir, and 2) immunomodulatory/antiviral agents, such as interferon 50
(IFN) or pegylated-IFN. However, these therapies can lead to drug resistance16 and can 51
have severe adverse side effects, respectively. Further, while these treatments 52

4
suppress viral replication, none of the current treatment options eliminate the virus. 53
Because of the lack of a cure, there is a great need to develop new therapies that can 54
cure chronic HBV infection. A deeper understanding of the molecular biology underlying 55
the replication of the virus could provide insight into the design and development of 56
therapies targeted at different aspects of replication and increase likelihood of 57
eradicating the virus and the devastating diseases it causes. 58
59

5
Hepadnaviridae 60
HBV is the prototypic member of the family Hepadnaviridae, derived from the words 61
hepatic DNA virus. All share a similar replication strategy and have many characteristics 62
in common: (1) All hepadnaviruses are dsDNA viruses, which replicate their genomes 63
via an RNA intermediate, known as the pregenomic RNA (pgRNA). This is in contrast to 64
retroviruses, such as human immunodeficiency virus (HIV) which is an RNA virus and 65
replicates its genome by integrating into the genome of an infected cell. Because of this 66
difference, HBV has been classified as a para-retrovirus. (2) Genome replication occurs 67
within cytoplasmic capsids and is facilitated by the virally encoded polymerase (P) 68
protein. The P protein has several enzymatic activities, such as RNA- and DNA-69
dependent DNA polymerase activities and RNase H activity, which allow it to reverse 70
transcribe the viral genome. And (3) all hepadnaviruses are enveloped viruses. Several 71
viral envelope proteins and presumably host-derived lipids form the envelope of the 72
virion. Because virions are not produced in the absence of envelope proteins, the 73
envelope proteins are thought to play an active role in virion production17,18. 74
Viruses in the Hepadnaviridae family can be divided into ortho- and avi- 75
hepadnaviruses, found in a variety of mammals and a variety of avian species, 76
respectively. Because of the narrow host range of hepadnaviruses19–22 and the lack of 77
an effective in vivo infection model system for the human virus, non-human 78
hepadnaviruses have been invaluable models and have helped us better understand 79
how hepadnaviruses replicate. Duck hepatitis B virus (DHBV)23 has been widely studied 80
to elucidate the molecular biology of hepadnaviruses both in vitro and in vivo24. This is 81
in large part due to the availability of well-established in vitro and in vivo model systems. 82

6
Primary duck hepatocytes (PDHs), which can be harvested from either the livers of 83
Peking ducks or their embryos, have been widely used because of their availability and 84
because they can be infected in vitro. Transfection of a chicken hepatoma cell line 85
(LMH)25 has also been used extensively to study multiple aspects of replication. Virions 86
produced by LMH cells are infectious in Peking ducks and PDHs, allowing the 87
opportunity to analyze effects of mutating the virus in its natural host; making this 88
transfection system extremely useful. 89
Much of what we know about how hepadnaviruses replicate was first learned 90
through the study of duck hepatitis B virus (DHBV) and later tested in the human virus. 91
Other members of this family which have been useful model systems include 92
woodchuck hepatitis B virus (WHV)26, and of note to my dissertation, snow goose 93
hepatitis B virus (SGHBV)27 and heron hepatitis B virus (HHBV) 28. While all 94
hepadnaviruses are similar, they are not identical in their pathogenesis or in the 95
diseases they cause. Therefore, some are better suited to study certain aspects of the 96
virus than others. For example, because avihepadnaviruses do not cause liver cancer in 97
their natural hosts, DHBV is not a useful model system to study HBV’s role in HCC. 98
Instead, WHV has been used to study how HBV contributes to the development of 99
HCC. 100
Another major difference between ortho- and avi- hepadnaviruses is that 101
orthohepadnaviruses express two additional proteins, the X and the M proteins. The M 102
protein is an envelope protein present in orthohepadnaviral virions. M is thought to be 103
non-essential to virus replication, because knocking down its expression does not 104
hinder viral replication or virion production. The X protein has been shown to interact 105

7
with a large number of cellular proteins, but a consensus on its exact role in viral 106
replication has not been reached. 107
HBV virions are also called Dane particles, named after their discoverer who first 108
visualized virions in the serum of Australian patients. All hepadnaviruses share a similar 109
virion structure (Figure 1.1); Virions have a diameter of 42 nm29 and 45 nm30 for the 110
human and duck hepatitis B viruses respectively. They consist of an outer lipoprotein 111
envelope which surrounds an inner protein shell (known as the nucleocapsid). The 112
virion envelope can be removed by treatment with non-ionic detergents (such as NP40), 113
leaving the capsids intact31. Nucleocapsids from the human and duck hepatitis B virus 114
are both 34 nm in diameter and are made of 240 copies of the capsid protein (Cp). The 115
P protein, which is covalently attached to the 5’ end of the minus-strand of the dsDNA 116
genome32, resides within the nucleocapsid. Nucleocapsids can be treated with SDS and 117
proteases to release the viral genome and remove the P protein from the minus-strand, 118
making isolation of virion associated viral DNA straightforward. 119
There are two forms of the dsDNA genome that can be found within virions. The 120
predominant form is the relaxed circular genome (RC DNA). RC DNA is a partially 121
dsDNA molecule consisting of a full length minus-strand and an incomplete plus-strand. 122
RC DNA is held in a circular conformation through overlapping 5’ cohesive ends. The 123
other dsDNA genome, termed duplex linear DNA (DL DNA), is a linear dsDNA molecule 124
and is much less abundant. These two forms differ in the mechanism by which they are 125
synthesized (Figure 1.6). These two different forms are synthesized via mutually 126
exclusive pathways, which I will describe in more detail later. 127
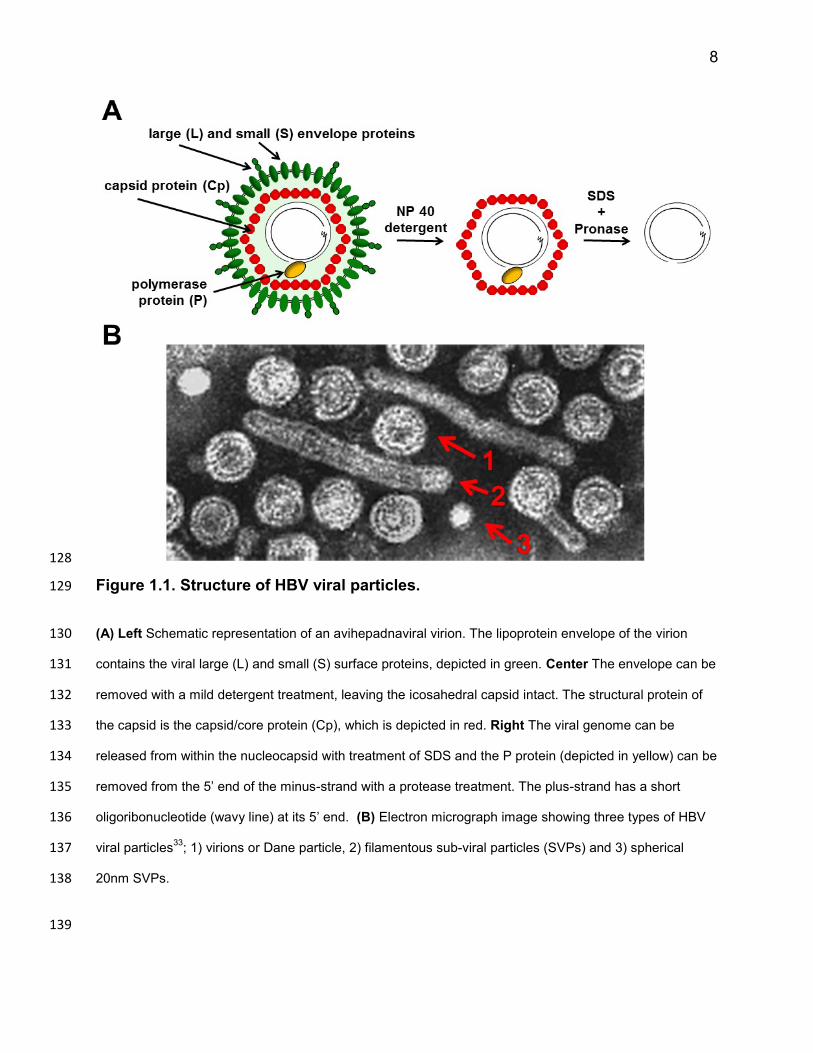
8
128
Figure 1.1. Structure of HBV viral particles. 129
(A) Left Schematic representation of an avihepadnaviral virion. The lipoprotein envelope of the virion 130
contains the viral large (L) and small (S) surface proteins, depicted in green. Center The envelope can be 131
removed with a mild detergent treatment, leaving the icosahedral capsid intact. The structural protein of 132
the capsid is the capsid/core protein (Cp), which is depicted in red. Right The viral genome can be 133
released from within the nucleocapsid with treatment of SDS and the P protein (depicted in yellow) can be 134
removed from the 5’ end of the minus-strand with a protease treatment. The plus-strand has a short 135
oligoribonucleotide (wavy line) at its 5’ end. (B) Electron micrograph image showing three types of HBV 136
viral particles33
; 1) virions or Dane particle, 2) filamentous sub-viral particles (SVPs) and 3) spherical 137
20nm SVPs. 138
139

9
Genetic Organization and Viral Proteins 140
All hepadnaviruses have a very similar genetic organization. Their genomes are 141
typically 3 to 3.2 kb in length, and code for a small number of proteins. They have 142
overlapping genes and each nucleotide codes for at least one protein. 143
Avihepadnaviruses have three genes, which code for four proteins (Figure 1.2). The P 144
gene codes for the multi-functional 90kDa P protein. P can be divided into several 145
functional domains, which are highly conserved among hepadnaviruses. These 146
domains from N-terminus to C-terminus are the terminal protein, spacer, reverse 147
transcriptase and RNase H. The reverse transcriptase and RNase H domains share 148
sequence and functional homology with other reverse transcriptases. P differs from 149
other reverse transcriptases in that P is required for selective packaging of pgRNA into 150
capsids34,35, and P serves as a protein primer for reverse transcription of the pgRNA. 151
The tyrosine residue at amino acid position 96 of DHBV P supplies the priming hydroxyl 152
group32,36. 153
The C gene codes for the capsid protein (Cp), which is the structural subunit of the 154
nucleocapsid. DHBV Cp is 262 amino acids in length and has a molecular weight of 32 155
kDa. DHBV Cp is larger than its human HBV counterpart (Figure 1.3A), which is only 156
183 amino acids in length and has a molecular weight of 21 kDa. Cp can be functionally 157
divided into two domains; the N-terminal domain and the C-terminal domain. The N-158
terminal domain is often called the assembly domain because it is sufficient for capsid 159
assembly37,38. The C-terminal domain (CTD) is highly basic, is required for pgRNA 160
encapsidation but not capsid assembly39,40 and can act as a nucleic acid chaperone41. 161
Lewellyn and Loeb have shown that the CTD contributes pleiotropically to several steps 162

10
of genome replication42. Cp can be phosphorylated at several arginine-rich regions in 163
the CTD. It is thought that the phosphorylation status of the protein correlates with 164
different stages of the replication cycle37,43–46. 165
The structure of the human HBV capsid has been determined47; however this 166
was done using a C-terminally truncated version of Cp, not full-length Cp. From 3D 167
reconstructions we see that the capsid has protruding spikes studding its surface and 168
has 2 nm pores that are large enough to allow dNTPs to freely pass in and out of the 169
capsid (Figure 1.3B). While high resolution structures of the DHBV capsid are not 170
available, cryo-EM analyses48 and structural models made using different methods 171
37,49,50 support the idea that DHBV capsids have a similar structure. 172
The “preC” portion of the C gene is used to express the e-antigen, which is similar to 173
Cp except that it is N-terminally extended and C-terminally truncated. Similar to HBV e-174
antigen (HBeAg), DHBV e-antigen (DHBeAg) can be found in the serum of infected 175
ducks. The role of e-antigen in HBV infection/replication is unknown. 176
The preS/S gene of avian hepadnaviruses consists of the PreS and S domains and 177
encodes two envelope proteins, the large (L) and the small (S) proteins. Both proteins 178
contain the C-terminal S domain. They differ in that the L protein is N-terminally 179
extended by ~163 amino acids (depending on the virus and isolate) and contains both 180
the PreS and the S domains. L and S are expressed from separate mRNAs which are 181
both transcribed from the PreS/S gene. Both proteins have complex transmembrane 182
topologies, spanning the membrane several times (Figure 1.4). The L protein is known 183
to take on at least two topologies51–55; one in which the PreS domain is cytosolically 184

11
disposed and one in which the PreS domain is disposed within the lumen of the 185
vesicular membrane it resides in. 186
The L protein’s multiple topologies allow it to perform its different functions which 187
require it to be on the interior, as well as exterior of the virion. For example, on the 188
exterior of the virion, the L protein is thought to mediate entry by binding to a cellular 189
receptor19,56–59. When cytosolically disposed, it can interact with nucleocapsids (directly 190
or indirectly) and facilitate the packaging of capsids into virions. 191
Orthohepadnaviruses have four genes which code for six proteins; the X and M 192
proteins in addition to those expressed in avihepadnaviruses. The M protein is an 193
envelope protein and is expressed from its own transcript. The X protein is coded by the 194
X gene and is expressed from its own transcript. 195
196
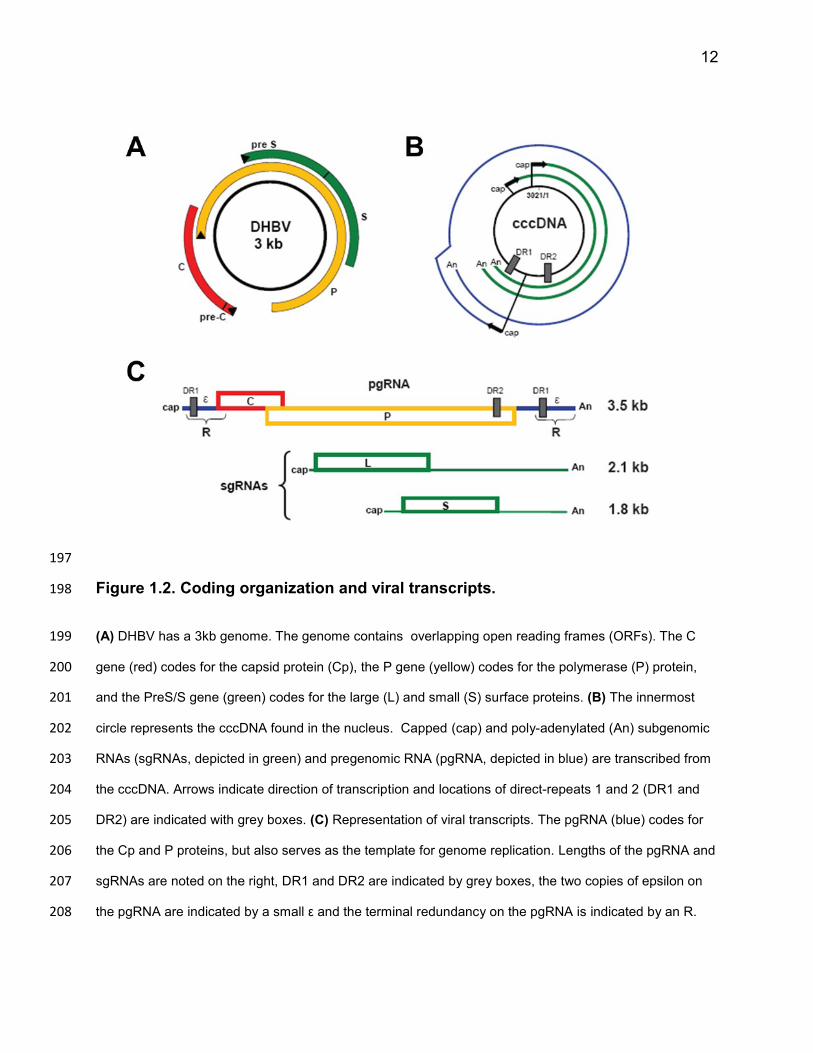
12
197
Figure 1.2. Coding organization and viral transcripts. 198
(A) DHBV has a 3kb genome. The genome contains overlapping open reading frames (ORFs). The C 199
gene (red) codes for the capsid protein (Cp), the P gene (yellow) codes for the polymerase (P) protein, 200
and the PreS/S gene (green) codes for the large (L) and small (S) surface proteins. (B) The innermost 201
circle represents the cccDNA found in the nucleus. Capped (cap) and poly-adenylated (An) subgenomic 202
RNAs (sgRNAs, depicted in green) and pregenomic RNA (pgRNA, depicted in blue) are transcribed from 203
the cccDNA. Arrows indicate direction of transcription and locations of direct-repeats 1 and 2 (DR1 and 204
DR2) are indicated with grey boxes. (C) Representation of viral transcripts. The pgRNA (blue) codes for 205
the Cp and P proteins, but also serves as the template for genome replication. Lengths of the pgRNA and 206
sgRNAs are noted on the right, DR1 and DR2 are indicated by grey boxes, the two copies of epsilon on 207
the pgRNA are indicated by a small ε and the terminal redundancy on the pgRNA is indicated by an R. 208

13
209
210
211
212

14
Figure1.3. Capsid protein (Cp) structure and capsid assembly. 213
(A) Linear representations of the DHBV and HBV capsid proteins (image is taken directly from49
). The 214
assembly and C-terminal domains (CTD) are labelled above the representations. The regions which form 215
the capsid spikes are represented by light grey, the proposed “insertion domain” within DHBV Cp is 216
indicated by the hashed pattern and the morphogeneic regions are indicated by a thick black line. (B) 3D 217
reconstruction of the crystal structure of the HBV capsid. The surface of the capsid is studded with spikes 218
and is fairly porous. (C) Ribbon representations of a HBV Cp monomer and dimer. The CTD is not 219
depicted here; these structures represent Cp which is truncated at amino acid 144. (D) Residues found to 220
be involved in HBV virion formation shown on a ribbon representation of a Cp dimer. Residues found to 221
be involved in virion formation are depicted with green spheres and are labelled in white60
. 222
223
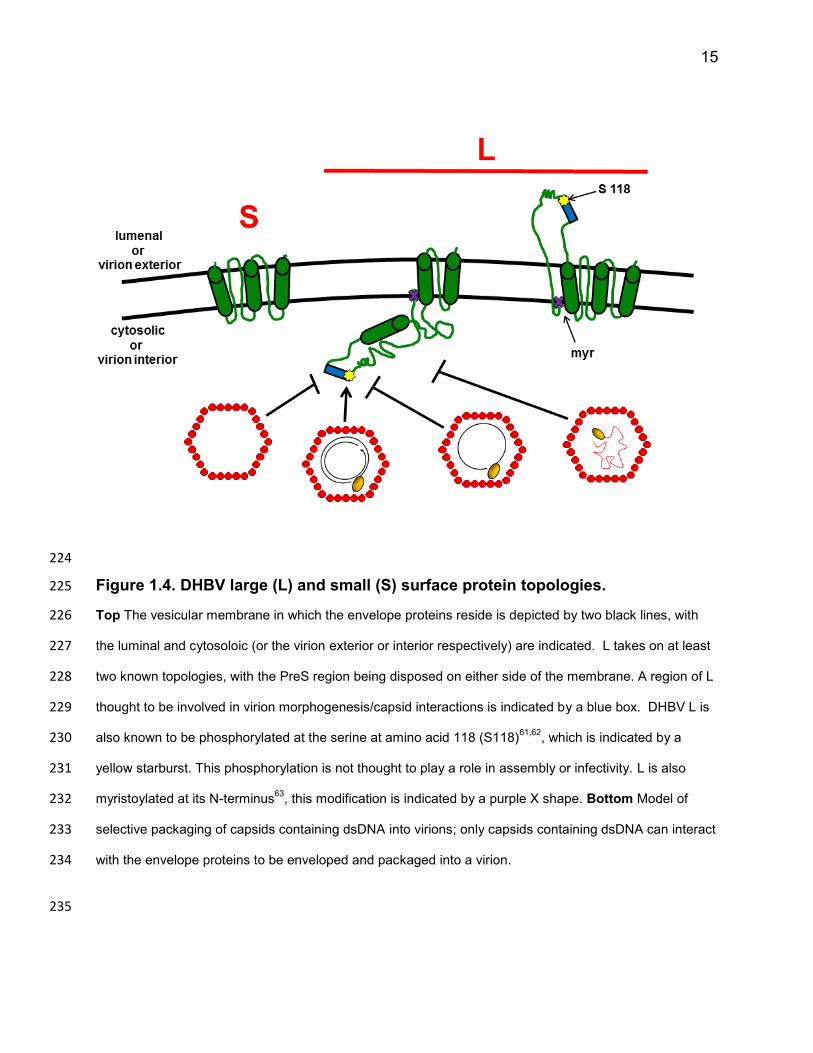
15
224
Figure 1.4. DHBV large (L) and small (S) surface protein topologies. 225
Top The vesicular membrane in which the envelope proteins reside is depicted by two black lines, with 226
the luminal and cytosoloic (or the virion exterior or interior respectively) are indicated. L takes on at least 227
two known topologies, with the PreS region being disposed on either side of the membrane. A region of L 228
thought to be involved in virion morphogenesis/capsid interactions is indicated by a blue box. DHBV L is 229
also known to be phosphorylated at the serine at amino acid 118 (S118)61,62
, which is indicated by a 230
yellow starburst. This phosphorylation is not thought to play a role in assembly or infectivity. L is also 231
myristoylated at its N-terminus63
, this modification is indicated by a purple X shape. Bottom Model of 232
selective packaging of capsids containing dsDNA into virions; only capsids containing dsDNA can interact 233
with the envelope proteins to be enveloped and packaged into a virion. 234
235

16
Viral Replication 236
Hepadnaviruses preferentially replicate in hepatocytes. They use a combination of 237
host and viral proteins to ultimately cause these cells to release infectious virions non-238
cytolytically (Figure 1.5). Upon entry, the viral nucleocapsid is trafficked to the nucleus 239
(Figure 1.5A), where its genome is deposited into the nucleus and modified to form a 240
super-coiled, covalently closed circular dsDNA (cccDNA) (Figure 1.5B). This process 241
involves several modifications, including completion of plus-strand synthesis, removal of 242
the P protein from the minus-strand DNA, removal of the RNA used to prime plus-strand 243
synthesis and ligation of the DNA strands. cccDNA plays an essential role in sustaining 244
chronic infection and viral persistence. Because of this, host and viral proteins involved 245
in the synthesis of cccDNA are attractive drug targets. Unfortunately, how HBV 246
synthesizes and maintains cccDNA in the nucleus is not well understood. 247
cccDNA is transcribed by cellular RNA polymerase II for the synthesis of pgRNA and 248
subgenomic RNAs (sgRNAs) (Figure 1.5C). The various transcripts are initiated from 249
different promoters but all use the same (and only) poly-adenylation site within the 250
genome (Figure 1.2B). The subgenomic RNAs, which are generated from differential 251
transcription of a single ORF, code for the L and S envelope proteins. L and S are 252
thought to be co-translationally inserted into the membrane of a cellular secretory 253
vesicle. The pgRNA serves as the replication template and also codes for the Cp and P 254
proteins (Figure 1.2C). 255
Cp and P proteins are expressed once pgRNA is exported from the nucleus. As Cp 256
accumulates in the cytoplasm, capsid proteins dimerize to form T-shaped structures in 257
which two alpha helices from each Cp monomer bundle together to form (what will 258

17
eventually be) the capsid spikes (Figure 1.3C). These dimers go on to form trimers of 259
dimers, which are thought to quickly coalesce to form an icosahedral capsid 260
structure64,65. Capsids can form with either a T=3 (90 dimers) or T=4 (120 dimers) 261
symmetry. Of the capsids formed from full-length Cp, approximately ~90% of the 262
capsids will have a T=4 symmetry. The T=3 symmetry becomes more favored as the 263
CTD is progressively truncated66. The role of T=3 capsids, if any, in viral replication is 264
not defined. 265
The P protein interacts with an encapsidation signal within the pgRNA, known as 266
epsilon, and this ribonucleoprotein complex becomes enclosed within the capsid, 267
forming the nucleocapsid through a process known as pgRNA encapsidation (Figure 268
1.5D). Capsids can self-assemble and it is unclear whether P and the pgRNA are 269
encapsidated before or after capsid assembly (for a review see67). 270
Genome replication is a rather complicated process, involving template switches that 271
are facilitated by a number of cis-acting sequences throughout the genome. Of 272
particular importance are the 11-12 nt long complementary sequences at either end of 273
the genome (Figure 1.2C), termed direct-repeat 1 and 2 (DR1 and DR2). Using several 274
residues in epsilon at the 5’ end of the pgRNA, P synthesizes four nucleotides of the 275
minus-strand. Serving as a protein primer in this process, P supplies the priming 276
hydroxyl from a tyrosine residue in its TP domain and becomes covalently attached to 277
the minus-strand36,68,69. P then switches templates to a complementary site near the 3’ 278
end of the pgRNA template, which overlaps with 4nt of DR170–72 (Figure 1.6 A). Here, P 279
continues to synthesize the minus-strand with its RNA-dependent DNA polymerase 280
activity as it degrades the pgRNA with its RNase H activity (Figure 1.6 B). This results in 281

18
a terminally-redundant minus-strand. P leaves a small RNA fragment (of ~18-19 nt) at 282
the 3’ end of the minus-strand which it uses as a primer to subsequently synthesize the 283
plus-strand of the genome73. This capped oligoribonucleotide is present on the final 284
dsDNA molecule found in virions (Figure 1.1A). 285
Typically, this RNA primer is transferred from DR1 to a partially complementary site 286
termed DR2, near the 5’ end of the minus strand in a process called primer 287
translocation and plus-strand synthesis begins73 (Figure 1.6 C). The plus-strand is 288
extended to the 5’ end of its template and the third and final template switch occurs. The 289
3’ end of the plus-strand to anneal to the 3’ end of the minus-strand in a process called 290
circularization (Figure 1.6 D)74; extension of the plus-strand from this site leads to the 291
formation of RC DNA. In a small number of instances, the primer does not translocate to 292
DR2 and plus-strand synthesis is initiated from DR1. This is called in situ priming 293
(Figure 1.6 E) and leads to the formation of the linear DL DNA form of the genome70. 294
Once the plus-strand is synthesized, capsids can be trafficked to the nucleus to 295
deposit the genome where the partially dsDNA genome is converted to cccDNA; 296
increasing the reservoir of cccDNA molecules in the nucleus (Figure 1.5 E). 297
Alternatively, the capsids can acquire an envelope in a process known as capsid 298
envelopment. While this step of replication is not well understood, it is thought that 299
capsids acquire an envelope by budding into the membrane of a secretory vesicle 300
(possibly a post-ER pre-golgi vesicular structure75) containing the trans-membrane 301
envelope proteins. After which, these enveloped nucleocapsids are guided through a 302
constitutive secretion pathway, non-cytolytically producing virions. Unfortunately, the 303
viral and host components involved and the mechanisms underlying this process are 304

19
not understood. Recent efforts have led to a slightly better understanding of the host 305
proteins and processes that are utilized or manipulated during virion formation, for 306
example cellular components involved in autophagy or vesicular/endosomal 307
trafficking76–83 have been proposed to contribute to virion production. However, much 308
more work needs to be done to define a canonical virion formation pathway. 309
Fortunately, more is known about the role that the viral capsid and envelope proteins 310
play in virion morphogenesis. Regions of the capsid 60,84,85 and envelope 18,86–89 proteins 311
involved in virion morphogenesis have been studied extensively (more so for HBV than 312
DHBV, for reviews see17,90). Both L and S are required for virion formation; in the 313
absence of the envelope proteins capsids are not released from the cell within a lipid 314
shell. This suggests the envelope proteins play an active role in coordinating capsid 315
envelopment and virion formation. It is hypothesized that the PreS region of the L 316
protein acts as a matrix protein, interacting with the capsid prior to capsid budding and 317
guiding it to be packaged into virions. A short sequence of the PreS domain of L has 318
been shown to be involved in virion formation87,88. For DHBV, a region between amino-319
acids 117 and 135 of DHBV L has been shown to contribute to virion morphogenesis, 320
because mutating this region decreased virion production while not affecting capsid 321
assembly. The location of this region in the proposed topological structure of DHBV L 322
(Figure 1.4) would support the idea that this region can interact with mature cytoplasmic 323
nucleocapsids; consistent with the hypothesis that L acts as a matrix protein to 324
envelope and package capsids into virions. As for Cp, several residues within the 325
assembly domain at the base of the capsid spikes are thought to play a role in virion 326
morphogenesis and potentially envelope protein interactions; when these residues were 327

20
changed in HBV, Cp capsid assembly and genome replication occurred normally but 328
virions were not produced (Figure 1.3 D). Because of this, it has been proposed that the 329
envelope interacting site is at the base of the capsid spikes. However, given the capsid 330
structure and physical proximity to the envelope, some propose the envelope-interacting 331
site is at the tip of the capsid spikes. It is possible that the envelope-capsid interaction is 332
a two-step process; for example, the first interaction may occur at the tip of the capsid 333
spikes and a second interaction occurs at the base of the spikes. 334
Interestingly, in addition to producing virions, infected cells also produce what are 335
known as sub-viral particles (SVPs) (Figures 1.1B and 1.5H). These particles are either 336
filamentous particles of various lengths or spherical; both particles are 20nm in 337
diameter. Similar to virions, SVPs contain the L and S envelope proteins. However, 338
SVPs differ from virions in that they do not contain a nucleocapsid and are therefore 339
non-infectious. DHBV virions and SVPs contain both the L and S proteins, with the S 340
protein being around four times more abundant in both54,91,92. 341
It is estimated that SVPs are produced at 1,000-10,000 fold higher levels than 342
virions. The role, if any, these particles play in viral replication is not completely 343
understood. Interestingly, SVPs enhance infection at low multiplicities of infection 93. It 344
has been proposed that these particles act as a decoy for the immune system, allowing 345
virions which would otherwise be cleared by the immune response to avoid surveillance. 346
Another idea is that these particles may bind cellular receptors of uninfected cells, 347
stimulating signaling pathways which make these cells more permissive for infection by 348
HBV. 349
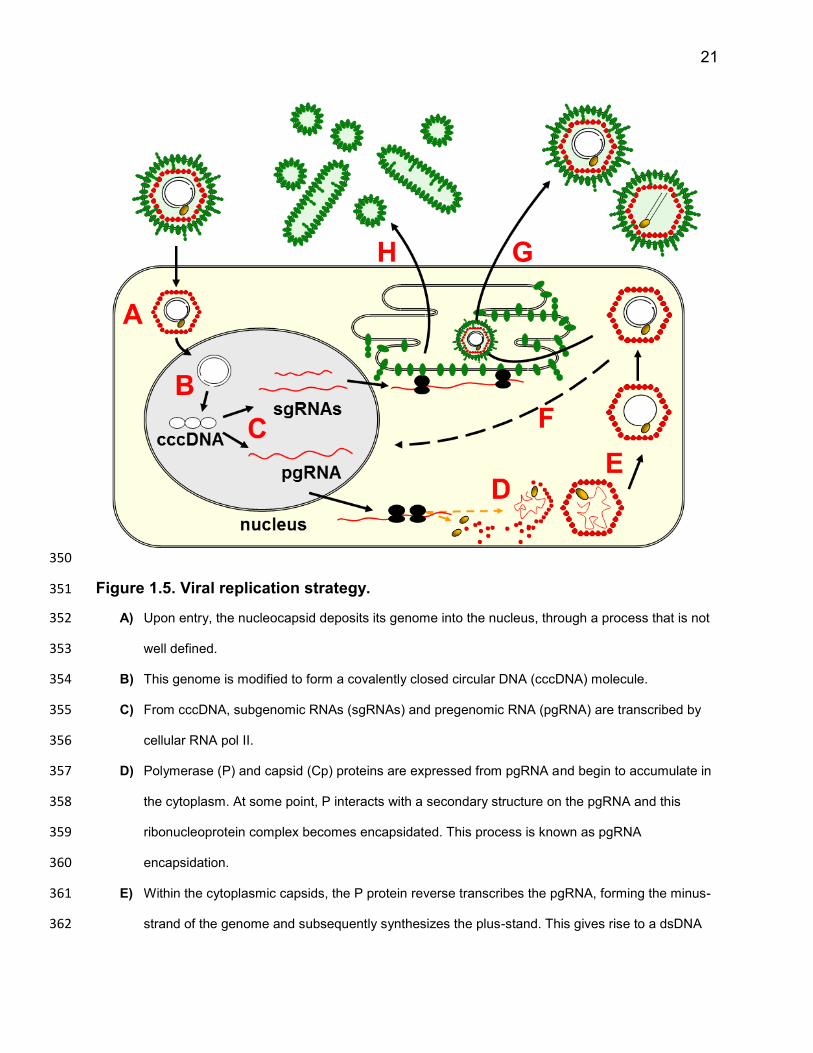
21
350
Figure 1.5. Viral replication strategy. 351
A) Upon entry, the nucleocapsid deposits its genome into the nucleus, through a process that is not 352
well defined. 353
B) This genome is modified to form a covalently closed circular DNA (cccDNA) molecule. 354
C) From cccDNA, subgenomic RNAs (sgRNAs) and pregenomic RNA (pgRNA) are transcribed by 355
cellular RNA pol II. 356
D) Polymerase (P) and capsid (Cp) proteins are expressed from pgRNA and begin to accumulate in 357
the cytoplasm. At some point, P interacts with a secondary structure on the pgRNA and this 358
ribonucleoprotein complex becomes encapsidated. This process is known as pgRNA 359
encapsidation. 360
E) Within the cytoplasmic capsids, the P protein reverse transcribes the pgRNA, forming the minus-361
strand of the genome and subsequently synthesizes the plus-stand. This gives rise to a dsDNA 362

22
viral genome. P mediated reverse transcription is discussed in further detail within the text and a 363
schematic representation of the process is depicted in Figure 1.6. 364
F) Capsids containing mature dsDNA genomes can be trafficked back to the nucleus to deposit their 365
genomes; increasing the copy number of cccDNA within the nucleus. This process is often called 366
cccDNA amplification. The L protein of DHBV and HBV have been shown to play a role in 367
regulating cccDNA amplification94,95
. 368
G) The sgRNAs code for the large (L) and small (S) envelope proteins, which are co-translationally 369
inserted into the membrane of a vesicular structure (some suggest a post-ER pre-Golgi vesicle). 370
As the envelope proteins accumulate in this membrane, they are thought to interact with each 371
other and with mature cytoplasmic nucleocapsids to facilitate capsid envelopment and the 372
packaging of capisds into virions. Support for the second function comes from the fact that both L 373
and S are required for virion formation. 374
H) The L and S proteins can also form particles which lack a nucleocapsid (and are therefore non-375
infectious), known as sub-viral particles (SVPs). Once L and S have accumulated to a certain 376
level and ratio, they are thought to aggregate and bud inward towards the lumen of the vesicle in 377
which they reside. These SVPs are thought to be released from the cell via a constitutive 378
secretion pathway, but the mechanisms underlying their formation are not defined. 379
380
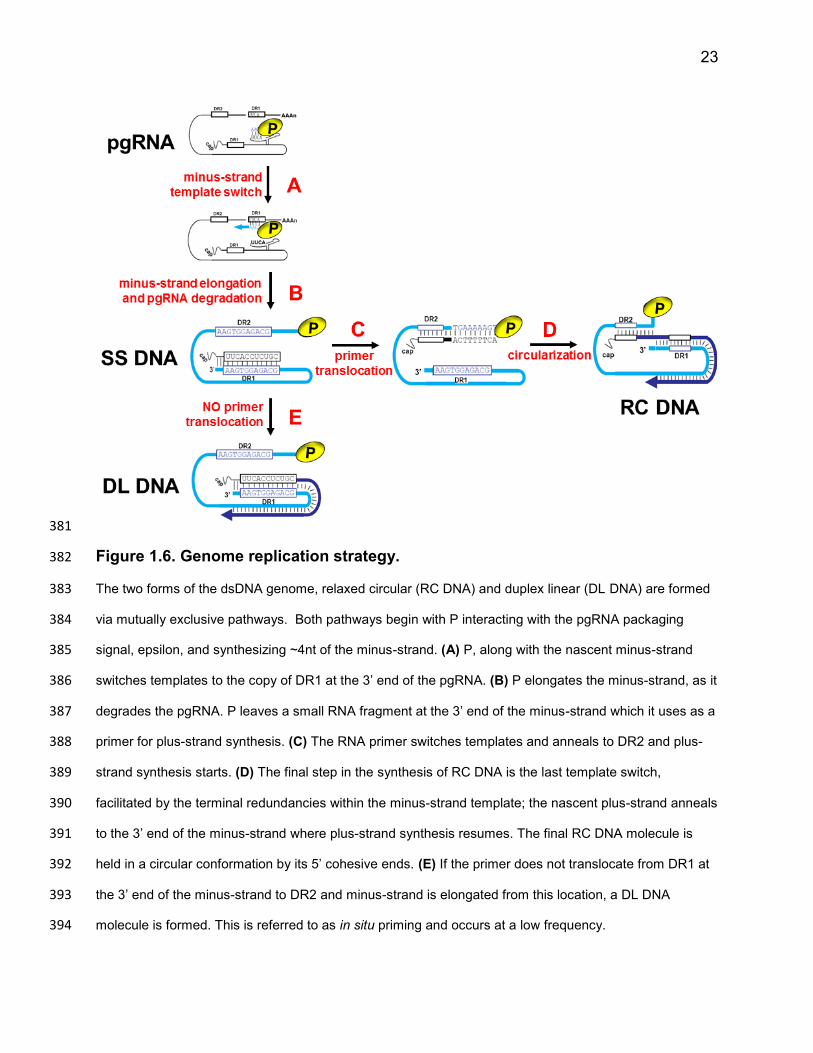
23
381
Figure 1.6. Genome replication strategy. 382
The two forms of the dsDNA genome, relaxed circular (RC DNA) and duplex linear (DL DNA) are formed 383
via mutually exclusive pathways. Both pathways begin with P interacting with the pgRNA packaging 384
signal, epsilon, and synthesizing ~4nt of the minus-strand. (A) P, along with the nascent minus-strand 385
switches templates to the copy of DR1 at the 3’ end of the pgRNA. (B) P elongates the minus-strand, as it 386
degrades the pgRNA. P leaves a small RNA fragment at the 3’ end of the minus-strand which it uses as a 387
primer for plus-strand synthesis. (C) The RNA primer switches templates and anneals to DR2 and plus-388
strand synthesis starts. (D) The final step in the synthesis of RC DNA is the last template switch, 389
facilitated by the terminal redundancies within the minus-strand template; the nascent plus-strand anneals 390
to the 3’ end of the minus-strand where plus-strand synthesis resumes. The final RC DNA molecule is 391
held in a circular conformation by its 5’ cohesive ends. (E) If the primer does not translocate from DR1 at 392
the 3’ end of the minus-strand to DR2 and minus-strand is elongated from this location, a DL DNA 393
molecule is formed. This is referred to as in situ priming and occurs at a low frequency. 394

24
Selective production of virions containing mature dsDNA 395
It has long been appreciated that hepadnaviruses selectively produce virions 396
containing mature dsDNA genomes. Capsids containing pgRNA are not packaged into 397
virions, while capsids containing ssDNA have been found to be packaged at a very low 398
efficiency96–99. Even in the absence of capsids containing dsDNA, capsids containing 399
pgRNA or ssDNA are not packaged into virions. Because of this, it is thought that 400
capsids containing mature dsDNA genomes differ from all other capsids in their ability to 401
interact with envelopment machinery and be packaged into a virion. 402
It is thought that the capsid serves as a link between genome replication and capsid 403
envelopment, relaying information about the completeness of genome replication to the 404
capsid surface. One model is that the synthesis of dsDNA triggers a physical change to 405
occur on the exterior of the capsid. This change is often referred to as the “capsid 406
maturation signal” or “capsid packaging signal” and renders the capsid competent for 407
packaging into virions and may facilitate interactions required for virion formation to 408
occur. In this way, the virus is able to prevent capsids containing incomplete/immature 409
genomes from being enveloped and packaged into virions. 410
Mutating HBV Cp at residue 97 causes the formation of virions containing ssDNA100–411
103. I similarly found that mutating a single residue of DHBV Cp causes DHBV to 412
produce virions containing ssDNA and present this work in chapter 3. Interestingly, 413
mutations in the HBV L protein can offset this secretion of virions containing immature 414
genomes, restoring preferential production of HBV virions containing mature dsDNA 415
genomes104. This implies that the envelope proteins are also involved in discriminating 416
between capsids containing mature and immature genomes. Consistent with these 417

25
findings, I present evidence which supports the idea that the envelope proteins, 418
specifically a small contiguous region of the DHBV PreS region of the L protein, 419
contribute to the ability of the virus to discriminate between capsids containing mature 420
and immature genomes. My findings suggest that L actively selects capsids which 421
contain mature dsDNA genomes for envelopment and selective packaging into virions 422
(Figure 1.4) and challenge the long-standing model that selective production of virions 423
containing dsDNA genomes is coded solely by the capsid. 424
425

26
426
427
CHAPTER 2 428
429
430
MATERIALS AND METHODS 431
432

27
Molecular Clones 433
DHBV plasmids: All DHBV molecular clones are derived from DHBV3105. The WT 434
DHBV plasmid, pD1.5G, has been described previously and contains 1.5 tandem copies 435
of DHBV3 DNA106. The DHBV L and S protein donor, DHBVEnv+, is a monomer of 436
DHBV3 in the vector pSP65. It has been previously described as pD3-SP65107. Only the 437
L and S envelope proteins are expressed from DHBVEnv+. The DHBV plasmid deficient 438
in Cp expression, DHBVpgRNA+P+Env+, expresses WT DHBV pgRNA, P, L and S proteins, 439
it contains a 4-nucleotide deletion at the NsiI site within the C gene, such that functional 440
Cp is not expressed. The DHBV plasmid deficient in envelope protein expression, 441
DHBVpgRNA+P+C+, expresses WT DHBV pgRNA, P and Cp. It contains a T1327A change, 442
which introduces a premature stop codon in the S gene. 443
The DHBV plasmid that expresses only Cp, DHBVC+, contains a deletion of nt 444
424 that creates a premature stop codon in the P gene and a deletion from nts 2549-445
2580 that inactivates the encapsidation signal. It also contains the inactivating mutation 446
in the S gene described above. 447
All DHBV Cp variants were derived from DHBVC+. Overlap extension PCR was 448
performed to create SG 74-107 D Cp. The single amino acid changes were introduced 449
using overlap extension PCR and oligonucleotide-directed mutagenesis108. PCR 450
fragments were inserted into DHBVC+. At residue 74, leucine was changed to an 451
isoleucine to create L74I DHBV Cp. At residue 87, glutamine was changed to a serine 452
to create Q87S DHBV Cp. At residue 107, histidine was changed to glutamic acid to 453
create H107E DHBV Cp. 454

28
SGHBV plasmids: All SGHBV molecular clones are derived from a plasmid 455
expressing SGHBV1-15 27. The WT SGHBV plasmid contains 1.3 tandem copies of 456
SGHBV1-15 DNA inserted into the PstI site of pBS-. The SGHBV L and S protein donor, 457
SGHBVEnv+, is a monomer of SGHBV1-15 inserted in the pBS- vector. Only L and S 458
envelope proteins are expressed from SGHBVEnv+. 459
The SGHBV plasmid deficient in Cp expression, SGHBVpgRNA+P+Env+, expresses 460
WT SGHBV pgRNA, P, L and S proteins. It contains a G2854T change within the C 461
gene, such that functional Cp is not expressed. The SGHBV plasmid deficient in 462
envelope protein expression, SGHBVpgRNA+P+C+, expresses WT SGHBV pgRNA, P and 463
Cp. It contains a TC to AA change at nt 1300, which introduces a premature stop codon 464
in the S gene. 465
The SGHBV plasmid which expresses only Cp, SGHBVC+, contains an insertion 466
at nt 426 that creates a premature stop codon in the P gene and a deletion from nts 467
2552-2582 that inactivates the encapsidation signal. It also contains the inactivating 468
mutations in the S gene described above. 469
All SGHBV Cp variants were derived from SGHBV1-15 Cp donor plasmid, 470
SGHBVC+. The amino acid changes were introduced into the SGHBV Cp gene using 471
overlap extension PCR and oligonucleotide-directed mutagenesis108. At residue 74, 472
isoleucine was changed to leucine to create I74L SGHBV Cp. At residue 107, glutamic 473
acid was changed to histidine to create E107H SGHBV Cp. These two changes were 474
combined to create the double mutant 74L 107H SGHBV Cp. 475
All L protein variants were derived from DHBVEnv+ and SGHBVEnv+. The plasmid 476
PreS-S D-SG L expresses a WT SGHBV S protein and a chimeric L protein consisting 477

29
of a DHBV PreS domain and an SGHBV S domain. PreS-S D-SG L was made by 478
inserting a KpnI-AvrII fragment from WT SGHBV into DHBVEnv+. The reciprocal plasmid 479
PreS-S SG-D L expresses a WT DHBV S protein and a chimeric L protein consisting of 480
an SGHBV PreS domain and a DHBV S domain. PreS-S SG-D L was made by inserting 481
a KpnI-NcoI fragment from WT DHBV into SGHBVEnv+. Overlap extension PCR was 482
performed to make chimeric envelope proteins SG 1-118 D L and SG 61-120 D L. PCR 483
fragments were inserted into DHBVEnv+. Both plasmids express WT DHBV proteins and 484
chimeric L proteins. The chimeric L proteins are primarily DHBV but contain SGHBV 485
sequence from amino acids 1-118 or 61-120, respectively. 486
HHBV plasmids. All molecular clones of HHBV are derived from a plasmid 487
expressing HHBV428; this plasmid is also referred to as 413-2. 413-2 contains 1.4 488
tandem copies of HHBV4 DNA inserted into an EcoRI site on the vector pIBI21106. The 489
HHBV L and S protein donor, HHBVEnv+, is a monomer of HHBV4 inserted in the pIBI21 490
vector. Only L and S envelope proteins are expressed from HHBVEnv+. 491
Details describing the HHBV Cp protein donor plasmid, HHBVC+, have been 492
previously described109. The HHBV plasmid deficient in Cp expression, 493
HHBVpgRNA+P+Env+, expresses WT HHBV pgRNA, P, L and S proteins, it contains a 494
frameshift mutation resulting from a 4nt insertion at the HindIII site at nucleotide 38, 495
such that functional Cp is not expressed. The HHBV plasmid deficient in L and 496
expression, HHBVpgRNA+P+C+, has been previously described and referred to as 497
pHSS1106. The plasmid expresses WT HHBV pgRNA, P, and Cp proteins, it contains a 498
mutation resulting in a premature stop codon in the S gene, such that functional L and S 499
are not expressed. 500

30
All HHBV Cp variants were derived from the HHBV4 Cp donor plasmid, HHBVC+. 501
PCR was performed using make chimeric Cp variants H 22-139 SG Cp and H 69-114 D 502
Cp. PCR fragments were inserted into HHBVC+. The H 22-139 SG Cp plasmid 503
expresses a chimeric Cp that is primarily HHBV, except for at 25 amino acid residues 504
between residues 22 and 139, as well as at the C-terminal residue. The H 69-114 D Cp 505
plasmid expresses a chimeric Cp that is primarily DHBV Cp, except for at eleven 506
residues within residues 69 and 114. 507
Overlap extension PCR was performed to create SG 74-107 D Cp. The three 508
HHBV Cp variants, L74I HHBV Cp, N107E HHBV Cp and 74I 107E HHBV Cp. The 509
amino acid changes were introduced into the HHBV Cp gene using overlap extension 510
PCR and oligonucleotide-directed mutagenesis108. At residue 74, leucine was changed 511
to isoleucine to create L74I HHBV Cp. At residue 107, asparagine was changed to 512
glutamic acid to create N107E HHBV Cp. These two changes were combined to create 513
the double mutant 74I 107E HHBV Cp. 514
Cell culture and transfection 515
Chicken hepatoma cell line, LMH25,110, was used in all transfections. Cells were 516
cultured and transfected as previously described111 with minor adjustments. Briefly, cells 517
were grown in Dulbecco’s Modified Eagle Medium Nutrient Mixture F-12 (Gibco) and 518
were supplemented to a final concentration of 5% fetal bovine serum and 519
penicillin/streptomycin. Cells were seeded onto 60 mm dishes 24 hours prior to 520
transfection. Plasmid DNA (10.5 ug total) was transfected into cells at 70-80% 521
confluence. In co-transfection experiments, the ratio of C protein donor plasmids to C 522
deficient plasmids was 1:1. Each transfection included 0.5 ug of a plasmid expressing 523

31
green fluorescent protein to estimate transfection efficiency. Transfections were 524
performed using the calcium phosphate method112. Media containing the calcium-525
phosphate precipitate was left on the cells for 16-18 hours. Cells were washed with 526
HBS-EGTA (2mM HEPES, 150mM NaCl, 0.5mM EGTA, pH 7.45) and fresh media was 527
replaced. Counting this time-point as 0 hours, media was changed and discarded at 24 528
hours. Subsequently, media was collected at 72 and 96 hours and pooled for virion 529
DNA isolation. After 96 hours, cells were washed with HBS-EGTA (2mM HEPES, 530
150mM NaCl, 0.5mM EGTA, pH 7.45) and stored at -700C overnight for isolation of viral 531
replicative intermediates from cytoplasmic capsids. 532
Isolation of viral replicative intermediates from cytoplasmic capsids 533
Isolation of viral replicative intermediates from cytoplasmic capsids was 534
performed as previously described113. Briefly, cells were lysed with a solution of 50mM 535
Tris pH 8.0, 1 mM EDTA, 0.2% NP-40. Nuclei were pelleted by centrifugation at 15,000 536
x g for 5 minutes at 40C and discarded. Supernatants were brought to 2mM CaCl2 and 537
treated with 44 units of micrococcal nuclease to degrade transfected plasmid DNA. After 538
1.5 hours, EDTA was added to a final concentration of 10mM to inactivate micrococcal 539
nuclease activity. Viral replicative intermediates (vRIs) were released from cytoplasmic 540
capsids and the P protein was removed from the viral DNA by the addition of Pronase to 541
a final concentration of 0.4 mg/ml (Roche) and SDS to a final concentration of 0.4%. 542
Following a 2-hour incubation at 370C, vRIs were extracted with phenol/chloroform, 543
ethanol precipitated, resuspended in TE (10mM Tris, 0.1mM EDTA. pH 8.0) and treated 544
with 2 ug of RNase A. 545
546

32
Isolation of virion DNA from LMH culture media 547
Virions were isolated as described114 with minor modifications. Briefly, pooled 548
culture media was centrifuged at 1,200 x g for 15 minutes to remove dead cells and 549
debris. To precipitate the viral particles, PEG 8000 and NaCl were added to a final 550
concentration of 10% (w/v) and 0.5M, respectively. Samples were incubated overnight 551
at 40C on a rocking platform and virions were pelleted via centrifugation at 3,200 x g for 552
15 minutes. Virions were resuspended in 400 ul of Leibovitz’s L-15 Medium (Invitrogen) 553
and buffered by the addition of Tris (pH 8.0) to a final concentration of 75mM. To 554
remove free capsids, Pronase (Roche) was added to a final concentration of 0.4 mg/ml 555
and incubated at 370C for 1-1.5 hours. This treatment was sufficient to degrade 556
unenveloped capsids, but not capsids within virions114. We demonstrated this was true 557
for SGHBV and DHBV. Viral DNA released from unenveloped capsids was degraded by 558
adding 44 units of micrococcal nuclease and CaCl2 to a final concentration of 2mM. 559
After one hour, EDTA was added to a final concentration of 10mM to inactivate 560
micrococcal nuclease activity. Virion DNA (vDNA) was released from virions by the 561
addition of SDS to a final concentration of 0.4%. The P protein was removed from the 562
viral DNA by adding Pronase (Roche) to a final concentration of 0.4 mg/ml and 563
incubating samples for 2 hours at 370C. Virion DNA was extracted with 564
phenol/chloroform, ethanol precipitated, resuspended in TE (10mM Tris, 0.1mM EDTA. 565
pH 8.0) and treated with 2 ug of RNase A for 30 minutes at 370C. 566
567

33
Southern blot analysis of viral nucleic acid 568
The method used for Southern blotting has been previously described42,115 with 569
minor alterations. Briefly, vRI and vDNA were electrophoresed through a 1.25% 570
agarose gel in Tris-borate-EDTA buffer (90mM Tris-borate, 2.5mM EDTA, pH 8.5). A 571
0.8% agarose gel was used when the packagable pgRNA in the sample being analyzed 572
was derived from HHBV. DNA was transferred to a Hybond-N membrane (Amersham) 573
and UV cross-linked to the membrane. Membranes were incubated in Church 574
hybridization solution (5mM EDTA, 1% BSA, 0.25 M Na2PO4, 7% SDS) for 15 minutes 575
and then probed overnight at 420C. The probe used was comprised of 20 576
oligonucleotides which were end-labeled with [γ-P32]ATP (Perkin-Elmer) using T4 poly-577
nucleotide kinase (New England Biolabs). All oligonucleotides used for probing are 578
complementary to the DHBV and SGHBV minus-strand. Membranes were washed in 579
Church wash solution (1mM EDTA, 20mM Na2PO4, 1% SDS). The membrane was 580
exposed in a phosphorimaging cassette, which was scanned with a Typhoon 8610 581
Variable Mode Imager (Molecular Dynamics). vRI and vDNA were quantitated using 582
ImageQuant 5.2 software (GE Healthcare). Mass of full-length minus-strand DNA was 583
determined by comparison to known masses of a linear double-stranded fragment of the 584
DHBV genome. 585
Statistical Analyses 586
All statistical analyses were done using the program MStat5.5 (provided by Norman 587
Drinkwater, UW-Madison <http://mcardle.wisc.edu/mstat/>). Statistical comparisons 588

34
between samples were made using the Wilcoxon rank sum test (two-sided). All samples 589
had n ≥ 6. We considered P < 0.05 to be statistically significant. 590
591

35
592
593
CHAPTER 3 594
595
596
SNOWGOOSE HEPATITIS B VIRUS (SGHBV) CAPSID AND 597
ENVELOPE PROTEINS CONTRIBUTE TO THE ABILITY OF 598
SGHBV TO PACKAGE CAPSIDS CONTAINING ssDNA IN 599
VIRIONS 600
601
With the exception of Figures 3.5 – 3.7, the data from this chapter has been published 602
in the Journal of Virology 603
(Greco, N., Hayes, M.H., and D.D. Loeb. 2014. J. Virol. 88(18):10705-13) 604
605
The data from figures 3.5 – 3.7 will be expanded upon and submitted as a separate 606
manuscript to the Journal of Virology. 607

36
Abstract 608
Hepadnaviruses selectively package capsids containing mature dsDNA genomes in 609
virions. Snow goose hepatitis B virus (SGHBV) is the only known hepadnavirus that 610
packages capsids containing ssDNA in virions. We found that cells replicating SGHBV 611
produce virions containing ssDNA as efficiently as virions containing mature dsDNA. 612
We determined that SGHBV capsid protein (Cp) and large envelope protein (L) 613
independently contribute to the production of virions containing ssDNA; with Cp making 614
a larger contribution. We identified that amino acid residues 74 and 107 of SGHBV Cp 615
contribute to this feature of SGHBV. When we changed these residues in DHBV Cp to 616
their SGHBV counterparts, capsids containing immature ssDNA were packaged in 617
virions. Interestingly, when we changed these residues in another avihepadnavirus, 618
heron hepatitis B virus (HHBV), capsids containing immature ssDNA were still not 619
packaged into virions. These results suggest that residues 74 and 107 contribute to the 620
appearance of the “capsid packaging signal” on the surface of capsids and interact with 621
the envelope proteins during virion formation, but that other residues of Cp and/or the 622
envelope proteins contribute as well. We also identified that a determinant within amino 623
acids 61-120 of SGHBV L contributes to its ability to produce virions containing ssDNA. 624
When we substituted this region of SGHBV L into DHBV L, capsids containing ssDNA 625
were packaged into virions. This result uncovers a new function of L and indicates that a 626
determinant between residues 61 and 120 of DHBV L contributes to its ability to 627
preferentially produce virions containing ssDNA. This, conversely, suggests that this 628
region of SGHBV L contributes to its ability to produce virions containing ssDNA. We 629
also found that cells replicating SGHBV package a larger fraction of the total RC DNA 630

37
they synthesize in virions compared to DHBV. We found that SGHBV L (and specifically 631
a determinant between amino acids 61-120) is responsible for this property of SGHBV. 632
Determining if the ability of SGHBV L to cause the formation of virions containing 633
ssDNA is related to its ability to package a large fraction of the total RC DNA they 634
synthesize in virions or if these two properties are mechanistically distinct will provide 635
insights into virion morphogenesis. 636
637

38
Importance 638
Cells replicating hepadnaviruses contain cytoplasmic capsids that contain mature and 639
immature genomes. However, only capsids containing mature dsDNA genomes are 640
packaged into virions. A mechanistic understanding of this phenomenon, which is 641
currently lacking, is critical to understanding the process of hepadnaviral virion 642
morphogenesis. In this study, we determined that the L protein (and specifically a small 643
region of the PreS of L) contributes to the ability of hepadnaviruses to selectively 644
produce virions containing mature dsDNA genomes. Our finding sheds new light on the 645
mechanisms underlying virion morphogenesis and challenges the dogma that “capsid 646
maturation”, and therefore the capsid protein (Cp), is solely responsible for the selective 647
production of virions containing mature dsDNA genomes. Further, we identified amino 648
acid residues of Cp that contribute to its ability to cause the selective production of 649
virions containing mature dsDNA genomes. Future studies on the role of these residues 650
in selective packaging of capsids containing dsDNA will broaden our understanding of 651
this poorly understood aspect of virus replication. 652
653

39
Introduction 654
Human Hepatitis B Virus (HBV) is the prototypic member of the Hepadnaviridae 655
family. Related hepadnaviruses, such as duck hepatitis B virus (DHBV)23, have been 656
invaluable in understanding HBV replication24. All hepadnaviruses are enveloped 657
viruses and preferentially replicate in hepatocytes116. All hepadnaviruses replicate their 658
genomes via reverse transcription of an RNA intermediate, the pregenomic RNA 659
(pgRNA)117. In this study, we used related avian hepadnaviruses (AHBVs), DHBV and 660
snow goose hepatitis B virus (SGHBV), to investigate capsid envelopment and virion 661
formation. 662
Avian hepadnavirus virions are composed of four viral proteins, the polymerase 663
protein (P), capsid protein (Cp) and the large (L) and small (S) envelope proteins. The 664
virion envelope consists of the L and S envelope proteins and host-cell derived 665
phospholipids118. Beneath the envelope, is the icosahedral nucleocapsid. The capsid is 666
35 nm in diameter and consists of 120 dimeric subunits of Cp. The P protein, which is 667
covalently attached to the ~3kb dsDNA genome, resides within the capsid. 668
Upon entry into a cell, the capsid delivers the viral genome to the nucleus, where 669
it is converted into covalently closed circular DNA (cccDNA). Viral RNAs, including 670
pgRNA, are transcribed from the cccDNA and exported from the nucleus. The pgRNA 671
codes for the Cp and P proteins and also serves as the template for genome replication. 672
During capsid assembly, the P protein binds to the encapsidation signal, called 673
epsilon34,35,107. Cp dimers are thought to polymerize around this ribonucleoprotein 674
complex, forming the capsid37. 675

40
Genome replication takes place within the cytoplasmic capsid and is facilitated by 676
the P protein117. Initially, P reverse transcribes the pgRNA into the minus-strand of the 677
DNA genome as it degrades the pgRNA. P then synthesizes plus-strand DNA, giving 678
rise to two forms of the dsDNA genome; relaxed circular (RC DNA) and duplex linear 679
(DL DNA). RC DNA is the predominant dsDNA form, while DL DNA is less abundant. 680
During an infection, cytoplasmic capsids contain a spectrum of replicative intermediates, 681
ranging from pgRNA to partially synthesized DNA genomes to completely synthesized 682
mature dsDNA genomes. Capsids containing mature dsDNA have two known fates; 683
they can be enveloped and secreted as a virion or they can deliver the viral DNA to the 684
nucleus to amplify and then maintain the copy number of cccDNA119. 685
The L and S proteins are coded by a single open reading frame, which consists 686
of a PreS domain and an S domain. The two proteins share the S domain. They differ in 687
that the L protein is N-terminally extended because it contains the PreS domain. The L 688
and S envelope proteins are both required for virion formation18. The envelope proteins 689
are thought to oligomerize, forming a 3D surface which interacts with cytoplasmic 690
capsids to drive capsid envelopment and virion formation. The envelope proteins are 691
localized in the membrane of a post-ER pre-Golgi secretory vesicle75. It is here where 692
the envelope proteins are hypothesized to interact with the cytoplasmic capsids during 693
virion formation, triggering budding into a secretory vesicle to acquire an envelope. 694
Both L and S have complex trans-membrane topologies. In addition, the L protein 695
is known to have multiple distinct conformations. Initially after synthesis, the PreS region 696
of the L protein is cytosolically disposed, but subsequently the PreS region of a subset 697
of the L proteins translocate across the membrane during virion formation, becoming 698

41
exposed on the exterior of the virion120. The L protein plays a role in virus entry and 699
virion formation; the multiple topologies of L allow it to perform these different functions. 700
Because cytoplasmic capsids contain pgRNA, partially synthesized DNA 701
genomes and mature dsDNA and virions only contain mature dsDNA genomes, it has 702
been hypothesized that the surface of capsids containing mature genomes differs from 703
those containing immature genomes. This difference is referred to as the “maturation” or 704
“capsid packaging signal”117. Capsids that have acquired the “capsid packaging signal” 705
are thought to productively interact with the envelope proteins to ultimately be packaged 706
in virions. Capsids containing ssDNA or pgRNA are thought to lack the “capsid 707
packaging signal”, rendering them incompetent for the interactions required for virion 708
formation. In this way hepadnaviruses selectively produce virions containing dsDNA. 709
Several studies have shown that capsids containing ssDNA or pgRNA are not packaged 710
in virions121, even in the absence of capsids containing dsDNA, supporting this 711
hypothesis96–99. Further, alternative models suggesting that mature capsids are 712
preferentially packaged resulting from an intrinsically higher affinity for interactions with 713
the envelopment machinery or due to kinetics of genome replication have already been 714
excluded through the use of a synchronized secretion system96. 715
Unlike all other hepadnaviruses characterized to date, SGHBV produces virions 716
containing ssDNA27. This property of SGHBV is surprising given its high phylogenetic 717
similarity to DHBV. Our goal was to understand how SGHBV produces virions 718
containing ssDNA, as this would provide insight into how other hepadnaviruses 719
selectively produce virions containing dsDNA. We first characterized the production of 720
virions containing mature dsDNA and immature ssDNA genomes for SGHBV and 721

42
DHBV. To do this, we measured RC DNA and ssDNA in cytoplasmic capsids and 722
extracellular virions. We found that SGHBV packages a larger fraction of the total RC 723
DNA it synthesizes in virions compared to DHBV. Further, we found that cells 724
expressing SGHBV produce virions containing immature ssDNA as efficiently as virions 725
containing mature dsDNA genomes. 726
Next, we determined which SGHBV proteins were responsible for these features 727
of SGHBV. We found that SGHBV Cp and L independently contribute to the production 728
of SGHBV virions containing ssDNA. However, Cp had a larger contribution. We 729
genetically mapped the amino acids of SGHBV Cp that contribute to the production of 730
virions containing ssDNA to residues 74 and 107. Further, we show residues 74 and 731
107 of the DHBV Cp contribute to the ability of DHBV to selectively produce virions 732
containing dsDNA. These residues are likely involved in capsid maturation and could be 733
part of a 3D surface on the exterior of the capsid that interacts with the envelope 734
proteins during capsid envelopment. We found that a determinant within amino acid 735
residues 61 and 120 of SGHBV L contributes to the production of virions containing 736
ssDNA; within this region, SGHBV and DHBV L differ at only 7 amino acid residues. In 737
addition, we found that a determinant within this same region of SGHBV L is 738
responsible for its ability to package a large fraction of the capsids containing RC DNA 739
into virions. It will be interesting to determine if the ability of SGHBV L to cause the 740
production of virions containing ssDNA is a function of its ability to package a large 741
fraction of the capsids containing RC DNA into virions or if these two properties are 742
unrelated. This will provide insight into how SGHBV L contributes to the packaging of 743
capsids containing immature ssDNA in virions. 744

43
Results 745
Rationale 746
It has long been appreciated that hepadnaviruses selectively package capsids 747
containing mature dsDNA in virions. SGHBV is unique in that it produces virions 748
containing dsDNA and ssDNA 27. By studying SGHBV virion production alongside the 749
well-characterized DHBV, we hoped to gain a better understanding of the mechanism 750
by which hepadnaviruses selectively produce virions containing mature dsDNA 751
genomes. Our goal was to confer the ability to produce virions containing ssDNA onto 752
DHBV and, conversely, confer the ability to selectively produce virions containing 753
dsDNA onto SGHBV through genetic complementation. In this way we could identify 754
which viral components were responsible for the respective behaviors of these two 755
viruses. To do this, we first analyzed the production of virions containing dsDNA and 756
ssDNA for DHBV and SGHBV. 757
Characterizing SGHBV virion production 758
LMH cells were transfected with a plasmid expressing SGHBV or DHBV. 759
Cytoplasmic capsid DNA and virion DNA were isolated and Southern blotting was 760
performed. The three major forms of vRIs (RC, DL and SS) were detected in 761
cytoplasmic DHBV capsids (Figure 3.1, lane 3). When DNA was isolated from DHBV 762
virions and analyzed similarly only RC DNA and DL DNA were detected (Figure 3.1, 763
lane 4 ). This result illustrates that DHBV capsids containing mature dsDNA genomes 764
are selectively packaged in virions. As previously described for SGHBV27, all three 765
major forms of the vRIs (RC, DL and SS) were detected in cytoplasmic capsids and 766

44
extracellular virions. This result illustrates that SGHBV produce virions containing 767
immature ssDNA (Figure 3.1, lanes 1 and 2). 768
We wanted to know if SGHBV produced virions containing ssDNA and dsDNA 769
equally well or preferentially produced one type of virion over the other. To this end, we 770
measured the levels of RC DNA and ssDNA in cytoplasmic capsids and in extracellular 771
virions. To quantitatively describe dsDNA virion production, we divided the amount of 772
RC DNA packaged in virions by the sum of RC DNA found in cytoplasmic capsids and 773
extracellular virions and multiplied this value by 100 to obtain a percentage, as shown 774
below: 775
% dsDNA packaged = (RC DNAvirions / (RC DNAcapsids + RC DNAvirions)) x 100 776
For SGHBV, this value was 58 ± 20%, and for DHBV it was 14 ± 3% (Figure 3.1). This 777
result indicated that a larger fraction of the cytoplasmic capsids containing dsDNA were 778
packaged in virions from cells expressing SGHBV compared to DHBV. 779
In the same way, we quantified ssDNA virion production by dividing the amount 780
of ssDNA packaged in virions by the sum of ssDNA found in both cytoplasmic capsids 781
and extracellular virions. We divided ssDNA virion production by RC DNA virion 782
production, to obtain a ratio of ssDNA production to RC DNA production, as shown 783
below: 784
𝑠𝑠𝐷𝑁𝐴 𝑝𝑎𝑐𝑘𝑎𝑔𝑒𝑑𝑑𝑠𝐷𝑁𝐴 𝑝𝑎𝑐𝑘𝑎𝑔𝑒𝑑 =
𝑠𝑠𝐷𝑁𝐴𝑣𝑖𝑟𝑖𝑜𝑛𝑠
𝑠𝑠𝐷𝑁𝐴𝑐𝑎𝑝𝑠𝑖𝑑𝑠 + 𝑠𝑠𝐷𝑁𝐴𝑣𝑖𝑟𝑖𝑜𝑛𝑠
𝑅𝐶 𝐷𝑁𝐴𝑣𝑖𝑟𝑖𝑜𝑛𝑠
𝑅𝐶 𝐷𝑁𝐴𝑐𝑎𝑝𝑠𝑖𝑑𝑠 + 𝑅𝐶 𝐷𝑁𝐴𝑣𝑖𝑟𝑖𝑜𝑛𝑠
785

45
For SGHBV this value was 1.01 ± 0.16, while for DHBV this value was indistinguishable 786
from zero, 0.05 ± 0.09 (Figure 3.1). This result means that SGHBV produces virions 787
containing immature ssDNA as efficiently as virions containing mature dsDNA, 788
indicating SGHBV was indiscriminate in selecting capsids containing dsDNA and 789
ssDNA for envelopment and virion formation. 790
Cp contributes to SGHBV’s ability to efficiently package capsids containing 791
ssDNA in virions 792
Given the capsid maturation hypothesis, we predicted that SGHBV Cp would be 793
sufficient to cause the production of virions containing immature ssDNA. To test this 794
prediction, we co-expressed DHBV P, pgRNA and envelope proteins with SGHBV Cp in 795
LMH cells. We did this by complementing a DHBV plasmid deficient in expressing Cp, 796
DHBVpgRNA+P+Env+, with a SGHBV Cp donor plasmid SGHBVC+. We measured the levels 797
of RC DNA and ssDNA in both cytoplasmic capsids and extracellular virions. We saw 798
no defect in cytoplasmic DNA synthesis or virion production, which allowed us to 799
measure relative ssDNA virion production. We found that relative ssDNA virion 800
production was 0.88 ± 0.11 (Figure 3.2A, lanes 3 and 4). This value was similar to the 801
SGHBV comparison; 0.99 ± 0.14 (Figure 3.2A, lanes 5 and 6). This result indicated that 802
SGHBV Cp contributes to the efficient production of virions containing ssDNA, 803
characteristic of SGHBV. 804
In addition, we found that % dsDNA virion production was low, 26 ± 14%, and 805
similar to the DHBV comparison which was 21 ± 10% (Figure 3.2A, lanes 1-4). This 806
result indicated that, despite its contribution to the efficient production of virions 807

46
containing ssDNA, SGHBV Cp does not contribute to SGHBV’s ability to package a 808
large fraction of the capsids containing RC DNA into virions. 809
To corroborate the above findings, we did complementary analyses in which we 810
co-expressed SGHBV P, pgRNA and Cp with the DHBV envelope proteins. We 811
predicted that we would see low % dsDNA virion production and high levels of relative 812
ssDNA virion production. To test this prediction, we co-transfected SGHBVpgRNA+P+C+ 813
with DHBVEnv+ and measured RC DNA and ssDNA in both cytoplasmic capsids and 814
extracellular virions. We saw no defect in cytoplasmic DNA synthesis or virion 815
production, which allowed us to measure relative ssDNA virion production. We found 816
that relative ssDNA virion production was high, 0.94 ± 0.17, which was similar to the 817
SGHBV comparison, 0.99 ± 0.32 (Figure 3.2B, lanes 5-8). Further, we found that % 818
dsDNA virion production was low, 12 ± 6% (Figure 3.2B, lanes 7 and 8), and was similar 819
to the DHBV comparison, 12 ± 6% (Figure 3.2B, lanes 1 and 2). 820
Taken together, we conclude that SGHBV Cp contributes to SGHBV’s ability to 821
produce virions containing ssDNA as efficiently as virions containing RC DNA, but not 822
its ability to package a large fraction of the capsids containing RC DNA into virions. 823
Because this was true regardless of the origin of P and pgRNA, we were able to 824
summarize these findings in Figure 3.2C. To better understand how SGHBV Cp 825
contributes to the production of virions containing ssDNA, we identified residues of 826
SGHBV Cp responsible for this property of SGHBV Cp, as well as residues of DHBV Cp 827
responsible for its ability to selectively package capsids containing mature dsDNA 828
genomes in virions. 829

47
Residues 74 and 107 of Cp contribute to DHBVs ability to selectively package 830
capsids containing mature dsDNA genomes in virions 831
After establishing that SGHBV Cp contributes to SGHBV’s ability to efficiently 832
package capsids containing ssDNA in virions, we wanted to identify the amino acid 833
residue(s) responsible for this characteristic. Because DHBV Cp and SGHBV Cp differ 834
at only 14 amino acid residues, we reasoned that one, or a few, of these different 835
residues were responsible for the behavior of the respective capsid proteins. According 836
to this logic, we should be able to change the behavior of DHBV Cp from selectively 837
packaging capsids containing mature dsDNA genomes in virions to packaging capsids 838
containing immature ssDNA in virions. We should also be able to change the behavior 839
of SGHBV Cp from packaging capsids containing immature ssDNA in virions to 840
selectively packaging capsids containing mature dsDNA genomes in virions. To identify 841
which residues of Cp were contributing to the respective behaviors of these viruses, we 842
made several chimeric SGHBV-DHBV capsid proteins, which we co-expressed with 843
DHBV P, pgRNA and envelope proteins. Of interest is a chimeric Cp in which we 844
substituted a small region, between residues 74-107, of SGHBV Cp into the DHBV Cp. 845
We called this variant SG 74-107 D Cp and it differs from DHBV Cp at only four 846
positions; 74, 83, 87 and 107 (Figure 3.3A). 847
When DHBV pgRNA, P and envelope proteins were co-expressed with SG 74-848
107 D Cp, we saw that cytoplasmic DNA synthesis and dsDNA virion production were 849
similar to DHBV. More importantly, relative ssDNA virion production was 0.80 ± 0.10 850
(Figure 3.3B, lanes 1 and 2). This value was not different, statistically, than when we 851

48
co-expressed DHBV pgRNA, P and envelope proteins with WT SGHBV Cp, where 852
relative ssDNA virion production was 0.88 ± 0.11 (Figure 3.2A, lanes 3 and 4). 853
Using a published alignment122 of the Cp amino acid sequences of ten other 854
avian hepadnaviruses known to selectively produce virions containing dsDNA and the 855
DHBV3 sequence (which was not included in the published alignment), we identified 856
three residues within this region, 74, 87 and 107, unique to SGHBV Cp. We changed 857
DHBV Cp to SGHBV individually at these three residues and named these proteins; 858
L74I DHBV Cp, Q87S DHBV Cp and H107E DHBV Cp (Figure 3.3A). As before, 859
DHBVpgRNA+P+Env+ and one of the three DHBV Cp variants were co-expressed. 860
Cytoplasmic capsid and virion DNA were isolated and analyzed by Southern blotting. In 861
all three cases, cytoplasmic DNA synthesis and dsDNA virion production were similar to 862
DHBV, which allowed us to evaluate relative ssDNA virion production. 863
When DHBV pgRNA, P and envelope proteins were co-expressed with L74I 864
DHBV Cp (Figure 3.3B, lanes 3 and 4) or H107E DHBV Cp (Figure 3.3B, lanes 7 and 8) 865
relative ssDNA virion production was 0.79 ± 0.22 and 0.34 ± 0.14, respectively. 866
However, when DHBV pgRNA, P and envelope proteins were co-expressed with Q87S 867
DHBV Cp, relative ssDNA virion production was essentially zero, 0.04 ± 0.09 (Figure 868
3.3B, lanes 5 and 6). While changing DHBV Cp to glutamic acid at residue 107, H107E 869
DHBV Cp, caused a significant increase in relative ssDNA virion production when 870
compared to the DHBV comparison, L74I DHBV Cp was the only variant which caused 871
production of ssDNA containing virions to the same relative level as WT SGHBV Cp 872
(Figure 3.2A, lanes 3 and 4). These findings indicate residue 74 plays an important role 873
in coupling genome maturation to capsid envelopment. Because it has been proposed 874

49
that capsid maturation is linked to a change on the exterior of the capsid123, structural or 875
biochemical comparisons of WT DHBV Cp and L74I DHBV Cp capsids could provide 876
insight into the nature of the “capsid packaging signal”. 877
Residues 74 and 107 of Cp contribute to SGHBVs ability to efficiently produce 878
virions containing ssDNA 879
Given the above result, we predicted that changing residues 74 and/or 107 of 880
SGHBV Cp to DHBV would convert SGHBV Cp from a producer of virions containing 881
immature ssDNA to a selective producer of virions containing mature dsDNA genomes. 882
To this end, residues 74 and 107 of SGHBV Cp were changed individually and in 883
combination; I74L SGHBV Cp, E107H SGHBV Cp and 74L 107H SGHBV Cp (Figure 884
3.4A). DHBVpgRNA+P+Env+ and each of the three SGHBV Cp variants were co-expressed, 885
viral DNA was isolated from cytoplasmic capsids and extracellular virions and were 886
analyzed via Southern blotting. We saw that cytoplasmic DNA synthesis and dsDNA 887
virion production were similar to DHBV, which allowed us to evaluate relative ssDNA 888
virion production. 889
When 74 or 107 were changed individually, relative ssDNA virion production was 890
0.26 ± 0.19 and 0.67 ± 0.30, respectively (Figure 3.4B, lanes 1-4). There was not a 891
complete loss of ssDNA virion production in either case. In fact, changing residue 107 892
had almost no effect on the ability of SGHBV Cp to produce virions containing ssDNA. 893
However, when both residues were changed, 74L 107H SGHBV Cp, relative ssDNA 894
virion production was indistinguishable from zero; 0.10 ± 0.09 (Figure 3.4B, lanes 5 and 895

50
6). These results illustrate that residues 74 and 107 of Cp contribute to 896
immature/selective production of virions. 897
Changing residues 74 and 107 of HHBV Cp does not confer the ability to produce 898
virions containing ssDNA onto HHBV 899
To extend these studies, we attempted to confer the ability to produce virions 900
containing ssDNA onto another avihepadnavirus, heron hepatitis B virus (HHBV). We 901
predicted that changing residues 74 and 107 of HHBV Cp to SGHBV would be sufficient 902
to cause HHBV to produce virions containing ssDNA, as it was for DHBV. To test this 903
prediction, we changed these residues in HHBV Cp to SGHBV and made three 904
expression plasmids; L74I H Cp, N107E H Cp and 74L 107E H Cp (Figure 3.5A). We 905
co-transfected LMH cells with HHBVpgRNA+P+Env+, which expresses pgRNA, P, L and S, 906
and a plasmid expressing HHBV Cp or one of the three HHBV Cp variants. Again, 907
cytoplasmic capsid and virion DNA were isolated and analyzed by Southern blotting. In 908
all three cases, cytoplasmic DNA synthesis and dsDNA virion production were similar to 909
HHBV, which allowed us to evaluate relative ssDNA virion production. Contrary to our 910
prediction, ssDNA was not detected in virions (Figure 3.5B). This result indicated that 911
changing residues 74 and 107 in HHBV Cp to SGHBV is not sufficient to confer the 912
ability to produce virions containing ssDNA onto HHBV, as it was for DHBV. DHBV is 913
the most closely related avihepadnavirus to SGHBV, while HHBV is the most distantly 914
related to SGHBV. Because of this, we hypothesize that additional residues in HHBV 915
Cp (or other proteins) need to be changed to convert HHBV from a preferential producer 916
of virions containing dsDNA to a producer of virions containing ssDNA. 917

51
SGHBV envelope proteins are sufficient to cause a large fraction of the capsids 918
containing RC DNA to be packaged into virions and the production of virions 919
containing ssDNA 920
We found that SGHBV packages a larger fraction of the capsids containing RC 921
DNA into virions compared to DHBV (Figure 3.1). We determined that the envelope 922
proteins are responsible for the ability of SGHBV to package a large fraction of the 923
capsids containing RC DNA into virions. When we supplied SGHBV envelope proteins 924
to an envelope-protein deficient DHBV we saw an increase in dsDNA virion production 925
when compared to the DHBV; these values were 45 ± 8% and 12 ± 6%, respectively 926
(Figure 3.2B, lanes 1-4). Conversely, when we supplied DHBV envelope proteins to an 927
envelope-protein deficient SGHBV we saw a decrease in % dsDNA virion production 928
when compared to the SGHBV comparison; these values were 12 ± 6% and 45 ± 20%, 929
respectively (Figure 3.2B, lanes 5-8). We conclude that SGHBV envelope proteins are 930
solely responsible for the ability of SGHBV to package a large fraction of the capsids 931
containing RC DNA into virions. Similarly, DHBV envelope proteins are responsible for 932
the ability of DHBV to package a small fraction of the capsids containing RC DNA into 933
virions. Unexpectedly, we found SGHBV envelope proteins were also sufficient to cause 934
production of virions containing ssDNA; relative ssDNA virion production was 0.61 ± 935
0.20 (Figure 3.2B, lanes 3 and 4). However, this was not as high as the value found 936
when SGHBV Cp was supplied to Cp deficient DHBV, 0.88 ± 0.11 (Figure 3.2A, lanes 3 937
and 4). 938
To corroborate these findings, we performed complementary analyses in which 939
we co-expressed SGHBV P, pgRNA and envelope proteins with DHBV Cp. Because 940

52
SGHBV envelope proteins were present and SGHBV Cp was absent, we expected that 941
virion production would be high and relative ssDNA virion production would be lower 942
than the SGHBV comparison. As expected, % dsDNA virion production was high, 53 ± 943
1% (Figure 3.2A, lanes 7 and 8). This value was similar to the SGHBV comparison 55 ± 944
5% (Figure 3.2A, lanes 5 and 6), supporting the idea that SGHBV’s ability to package a 945
large fraction of the capsids containing RC DNA into virions is a function of its envelope 946
proteins. Further, relative ssDNA virion production was 0.62 ± 0.13 (Figure 3.2A, lanes 947
7 and 8). This value was lower than the SGHBV comparison which was 0.99 ± 0.14 948
(Figure 3.2A, lanes 5 and 6), but higher than the DHBV comparison which was 0.07 ± 949
0.13 (Figure 3.2A, lanes 1 and 2). These results were consistent with our earlier finding 950
that SGHBV envelope proteins contribute to the production of virions containing ssDNA. 951
Taken together, we conclude that SGHBV envelope proteins are sufficient to cause 952
packaging of a large fraction of the capsids containing RC DNA into virions, and also 953
contribute to the production of virions containing ssDNA; as summarized in Figure 3.2C. 954
A determinant within residues 61 and 120 of SGHBV L contributes to the ability of 955
SGHBV to package a large fraction of the capsids containing RC DNA into virions 956
and to produce virions containing ssDNA 957
After establishing that SGHBV envelope proteins contribute to SGHBV’s ability to 958
package capsids containing ssDNA in virions and package a large fraction of the 959
capsids containing RC DNA into virions, we wanted to identify the amino acid residue(s) 960
responsible for these characteristic. It is possible these two traits could map to the same 961
or different residues. As we did with Cp, we reasoned that one, or a few, of the residues 962
which are different in SGHBV and DHBV envelope proteins were responsible for the 963

53
behavior of the respective envelope proteins. According to this logic, we should be able 964
to change the behavior of DHBV envelope proteins from selectively packaging capsids 965
containing mature dsDNA genomes in virions to packaging capsids containing immature 966
ssDNA in virions. Further, we should also be able to change the behavior of SGHBV 967
envelope proteins from packaging capsids containing immature ssDNA in virions to 968
selectively packaging capsids containing mature dsDNA genomes in virions. 969
The S proteins (and the S domain of the L protein) of SGHBV and DHBV are 970
very similar (differing at only 6 amino acid residues), while the PreS domains of their L 971
proteins are less conserved (differing at 24 amino acid residues). Because of this fact, 972
we hypothesized the determinant responsible for the respective behaviors of the 973
SGHBV and DHBV envelope proteins would be within the PreS domain. To test this, we 974
made two chimeric envelope protein constructs in which we swapped the PreS domains 975
of SGHBV and DHBV. The first, PreS-S D-SG L, expresses a WT SGHBV S protein and 976
a chimeric L protein containing a DHBV PreS domain and an SGHBV S domain. The 977
second construct, PreS-S SG-D L, expresses a WT DHBV S protein and a chimeric L 978
protein containing an SGHBV PreS domain and a DHBV S domain. We co-expressed 979
DHBV P, pgRNA and Cp with one of the two chimeric envelope protein constructs and 980
analyzed their ability to package capsids containing RC DNA into virions and to produce 981
virions containing ssDNA. We found that PreS-S D-SG L did not produce virions 982
containing ssDNA and only packaged 24 ± 11% of the RC DNA they synthesize into 983
virions, similar to DHBV envelope proteins. We also found that PreS-S SG-D L behaved 984
similar to the SGHBV envelope proteins; it packaged 42 ± 17% of the RC DNA it 985
synthesized in virions and had a relative ssDNA virion production of 0.54 ± 0.19. This 986

54
allowed us to conclude that a determinant within the PreS region of L contribute to the 987
respective behaviors of these two proteins. 988
We next wanted to identify which residues in the PreS region of L contribute to 989
the respective behaviors of these viruses. To do this, we analyzed several chimeric 990
SGHBV-DHBV L protein constructs, which we co-expressed with DHBV P, pgRNA, Cp 991
and S. Of interest are two chimeric L proteins in which we substituted a small region, 992
between residues 1 and 118 or 61 and 120, of SGHBV L into DHBV L. We called these 993
variants SG 1-118 D L and SG 61-120 D L, respectively. SG 61-120 D L differs from 994
DHBV L at only seven positions; 61, 67, 76, 79, 87, 118 and 120. 995
When DHBV pgRNA, P, Cp and S proteins were co-expressed with SG 1-118 D 996
L or SG 61-120 D L, we saw normal cytoplasmic DNA synthesis, allowing us to evaluate 997
the production of virions. We found that % dsDNA virion production was high for both 998
SG 1-118 D L and SG 61-120 D L; % dsDNA virion production was 58 ± 17% and 41± 999
21%, respectively (Figure 3.6, lanes 5-8). These values were similar to the value seen 1000
when we co-expressed DHBV pgRNA, P, and Cp proteins with WT SGHBV L and S, 1001
where 55 ± 5% of the RC DNA synthesized was packaged into virions (Figure 3.2A, 1002
lanes 5 and 6); indicating that a determinant between residues 61 and 120 of SGHBV L 1003
contributes to its ability to package a large fraction of the capsids containing RC DNA 1004
into virions. 1005
When we evaluated the ability of these L variants to produce virions containing 1006
ssDNA, we found that relative ssDNA virion production was 0.65 ± 0.17 and 0.54 ± 0.19 1007
for SG 1-118 D L and SG 61-120 D L, respectively (Figure 3.6, lanes 5-8). These 1008

55
values were not different, statistically, than when we co-expressed DHBV pgRNA, P, 1009
and Cp proteins with WT SGHBV L and S, where relative ssDNA virion production was 1010
0.61 ± 0.20 (Figure 3.2B, lanes 3 and 4). This result suggests that a determinant(s) 1011
between residues 61 and 120 of SGHBV L contributes to its ability to package capsids 1012
containing ssDNA into virions. 1013
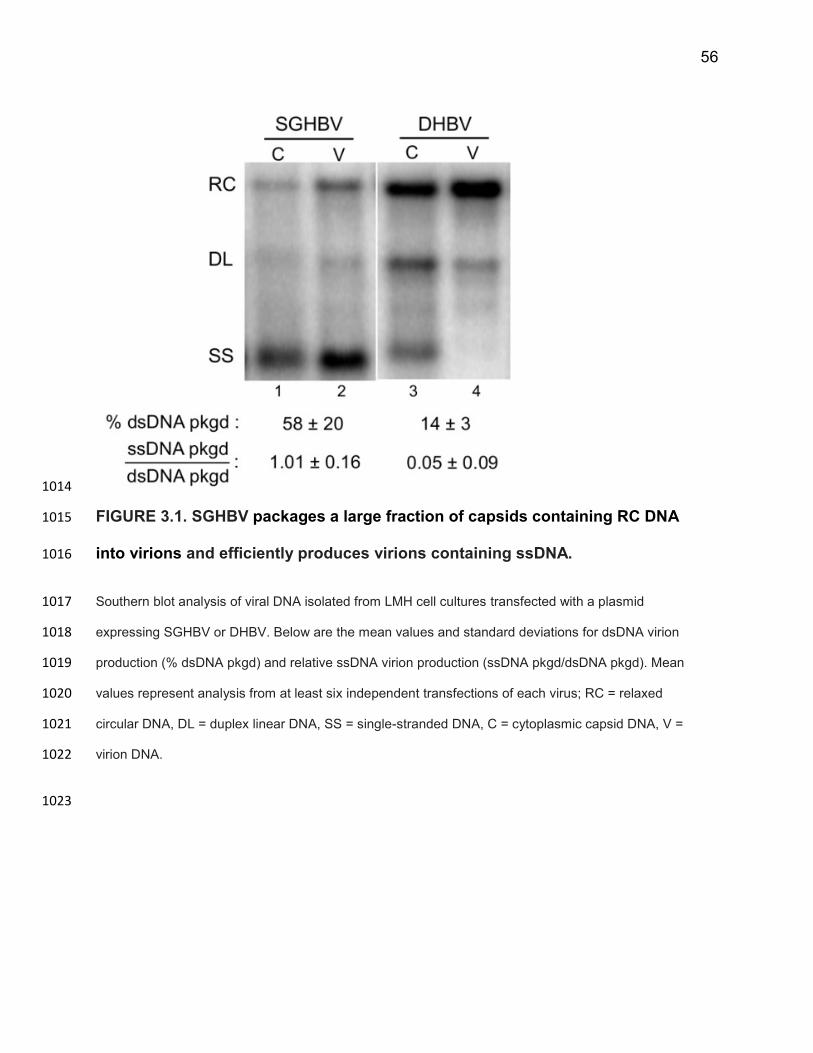
56
1014
FIGURE 3.1. SGHBV packages a large fraction of capsids containing RC DNA 1015
into virions and efficiently produces virions containing ssDNA. 1016
Southern blot analysis of viral DNA isolated from LMH cell cultures transfected with a plasmid 1017
expressing SGHBV or DHBV. Below are the mean values and standard deviations for dsDNA virion 1018
production (% dsDNA pkgd) and relative ssDNA virion production (ssDNA pkgd/dsDNA pkgd). Mean 1019
values represent analysis from at least six independent transfections of each virus; RC = relaxed 1020
circular DNA, DL = duplex linear DNA, SS = single-stranded DNA, C = cytoplasmic capsid DNA, V = 1021
virion DNA. 1022
1023

57
FIGURE 3.2. SGHBV Cp contributes to the efficient production of virions 1024
containing ssDNA, while SGHBV envelope proteins contribute to the ability of 1025
SGHBV to package a large fraction of the capsids containing RC DNA into virions, 1026
characteristic of SGHBV. 1027
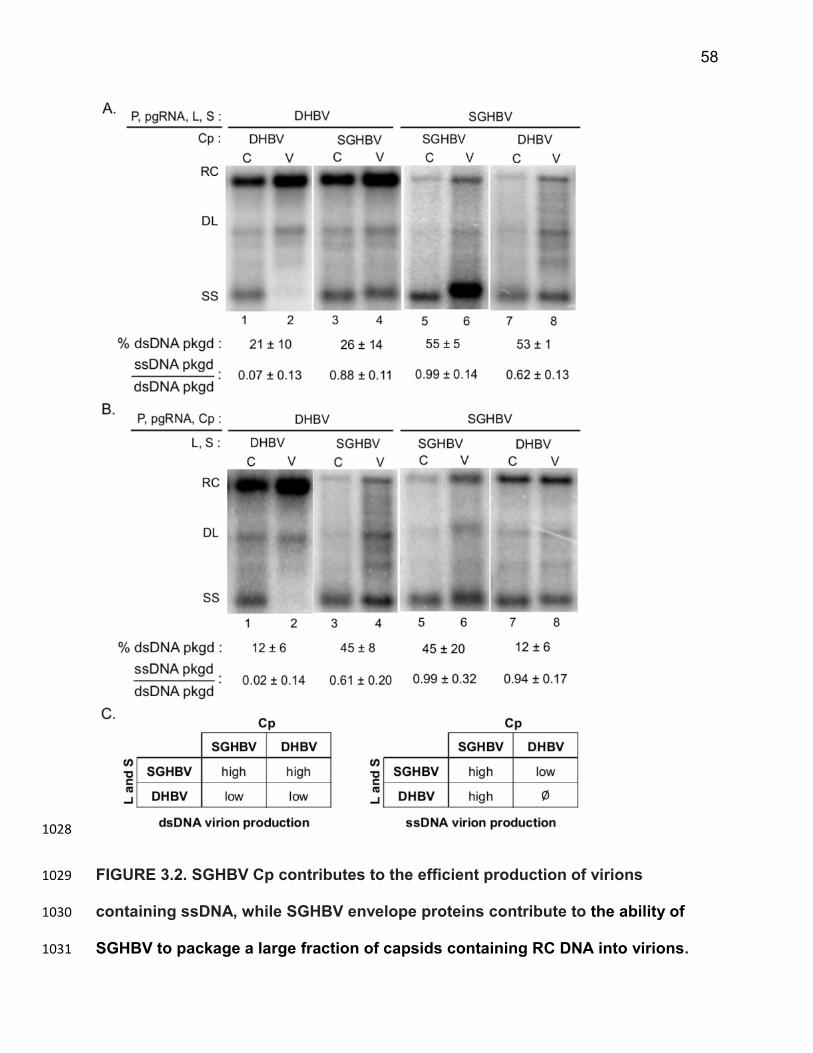
58
1028
FIGURE 3.2. SGHBV Cp contributes to the efficient production of virions 1029
containing ssDNA, while SGHBV envelope proteins contribute to the ability of 1030
SGHBV to package a large fraction of capsids containing RC DNA into virions. 1031

59
Southern blot analysis. (A.) A plasmid expressing DHBV pgRNA, P, L and S was co-1032
transfected with a plasmid expressing DHBV Cp (lanes 1 and 2) for a DHBV 1033
comparison or SGHBV Cp (lanes 3 and 4). A plasmid expressing SGHBV pgRNA, P, 1034
L and S was co-transfected with a plasmid expressing SGHBV Cp (lanes 5 and 6) for 1035
an SGHBV comparison or DHBV Cp (lanes 7 and 8). (B.) A plasmid expressing DHBV 1036
pgRNA, P and Cp was co-expressed with a plasmid expressing DHBV L and S (lanes 1037
1 and 2) for a DHBV comparison or a plasmid expressing SGHBV L and S (lanes 3 1038
and 4). A plasmid expressing SGHBV pgRNA, P and Cp was co-transfected with a 1039
plasmid expressing SGHBV L and S (lanes 5 and 6) for an SGHBV comparison or a 1040
plasmid expressing DHBV L and S (lanes 7 and 8). Below are the mean values and 1041
standard deviations for dsDNA virion production (% dsDNA pkgd) and relative ssDNA 1042
virion production (ssDNA pkgd/dsDNA pkgd). Mean values represent analysis from at 1043
least six independent transfections of each virus. (C.) Tables summarizing the effects 1044
of Cp and envelope proteins on dsDNA virion production and relative ssDNA virion 1045
production; RC = relaxed circular DNA, DL = duplex linear DNA, SS = single-stranded 1046
DNA, C = cytoplasmic capsid DNA, V = virion DNA. 1047
1048
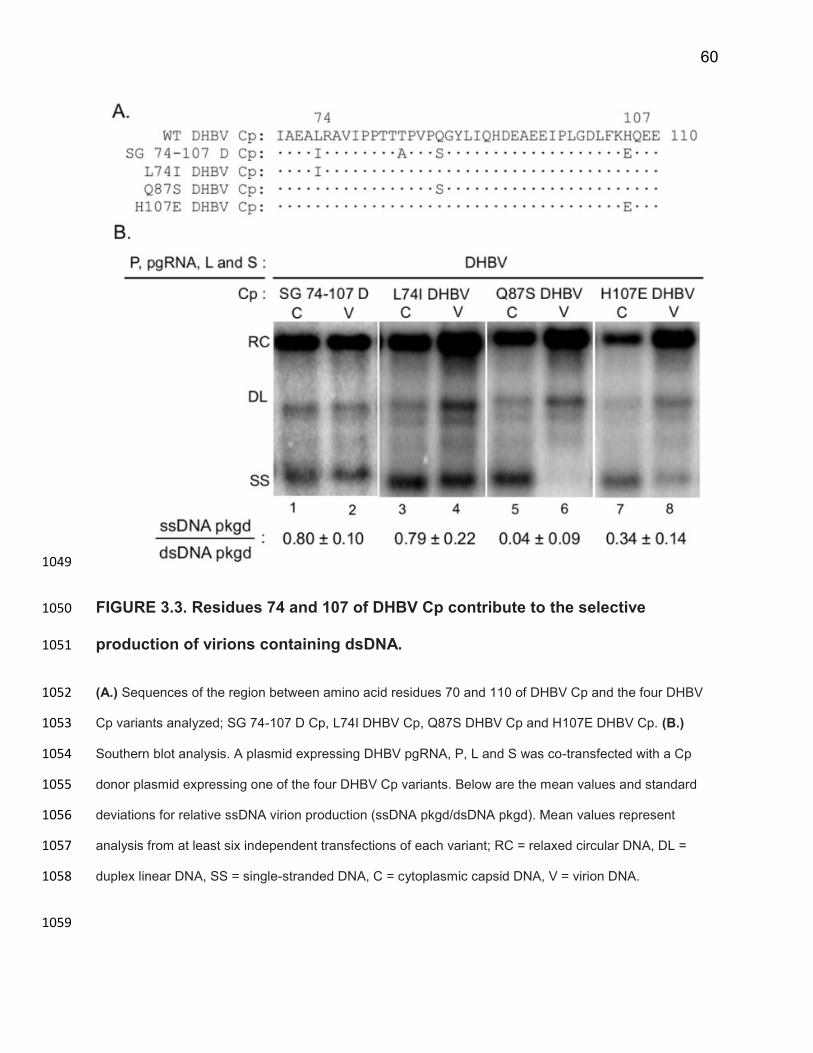
60
1049
FIGURE 3.3. Residues 74 and 107 of DHBV Cp contribute to the selective 1050
production of virions containing dsDNA. 1051
(A.) Sequences of the region between amino acid residues 70 and 110 of DHBV Cp and the four DHBV 1052
Cp variants analyzed; SG 74-107 D Cp, L74I DHBV Cp, Q87S DHBV Cp and H107E DHBV Cp. (B.) 1053
Southern blot analysis. A plasmid expressing DHBV pgRNA, P, L and S was co-transfected with a Cp 1054
donor plasmid expressing one of the four DHBV Cp variants. Below are the mean values and standard 1055
deviations for relative ssDNA virion production (ssDNA pkgd/dsDNA pkgd). Mean values represent 1056
analysis from at least six independent transfections of each variant; RC = relaxed circular DNA, DL = 1057
duplex linear DNA, SS = single-stranded DNA, C = cytoplasmic capsid DNA, V = virion DNA. 1058
1059
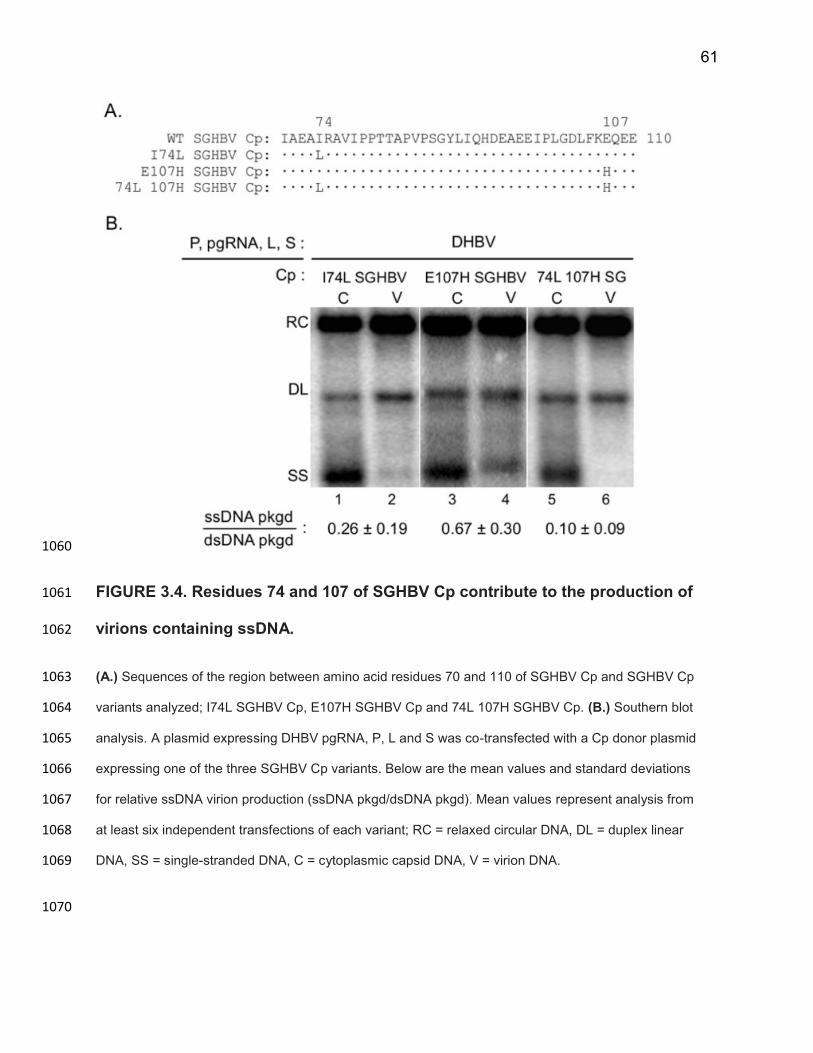
61
1060
FIGURE 3.4. Residues 74 and 107 of SGHBV Cp contribute to the production of 1061
virions containing ssDNA. 1062
(A.) Sequences of the region between amino acid residues 70 and 110 of SGHBV Cp and SGHBV Cp 1063
variants analyzed; I74L SGHBV Cp, E107H SGHBV Cp and 74L 107H SGHBV Cp. (B.) Southern blot 1064
analysis. A plasmid expressing DHBV pgRNA, P, L and S was co-transfected with a Cp donor plasmid 1065
expressing one of the three SGHBV Cp variants. Below are the mean values and standard deviations 1066
for relative ssDNA virion production (ssDNA pkgd/dsDNA pkgd). Mean values represent analysis from 1067
at least six independent transfections of each variant; RC = relaxed circular DNA, DL = duplex linear 1068
DNA, SS = single-stranded DNA, C = cytoplasmic capsid DNA, V = virion DNA. 1069
1070
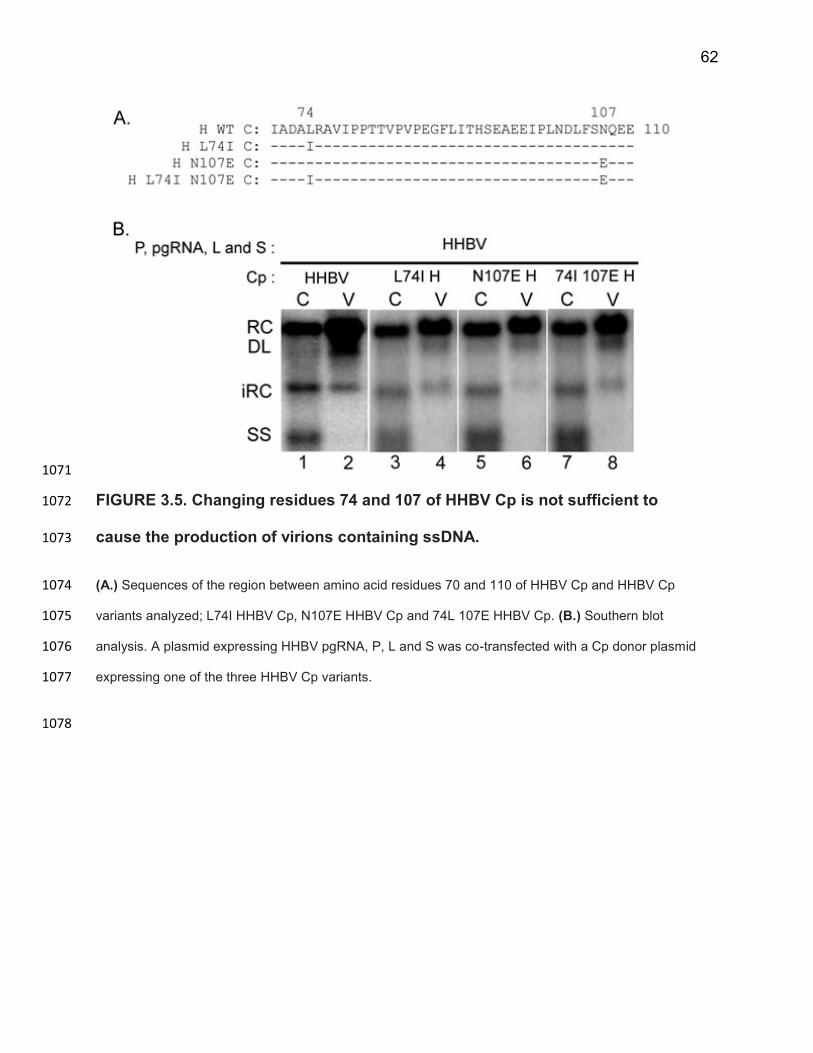
62
1071
FIGURE 3.5. Changing residues 74 and 107 of HHBV Cp is not sufficient to 1072
cause the production of virions containing ssDNA. 1073
(A.) Sequences of the region between amino acid residues 70 and 110 of HHBV Cp and HHBV Cp 1074
variants analyzed; L74I HHBV Cp, N107E HHBV Cp and 74L 107E HHBV Cp. (B.) Southern blot 1075
analysis. A plasmid expressing HHBV pgRNA, P, L and S was co-transfected with a Cp donor plasmid 1076
expressing one of the three HHBV Cp variants. 1077
1078
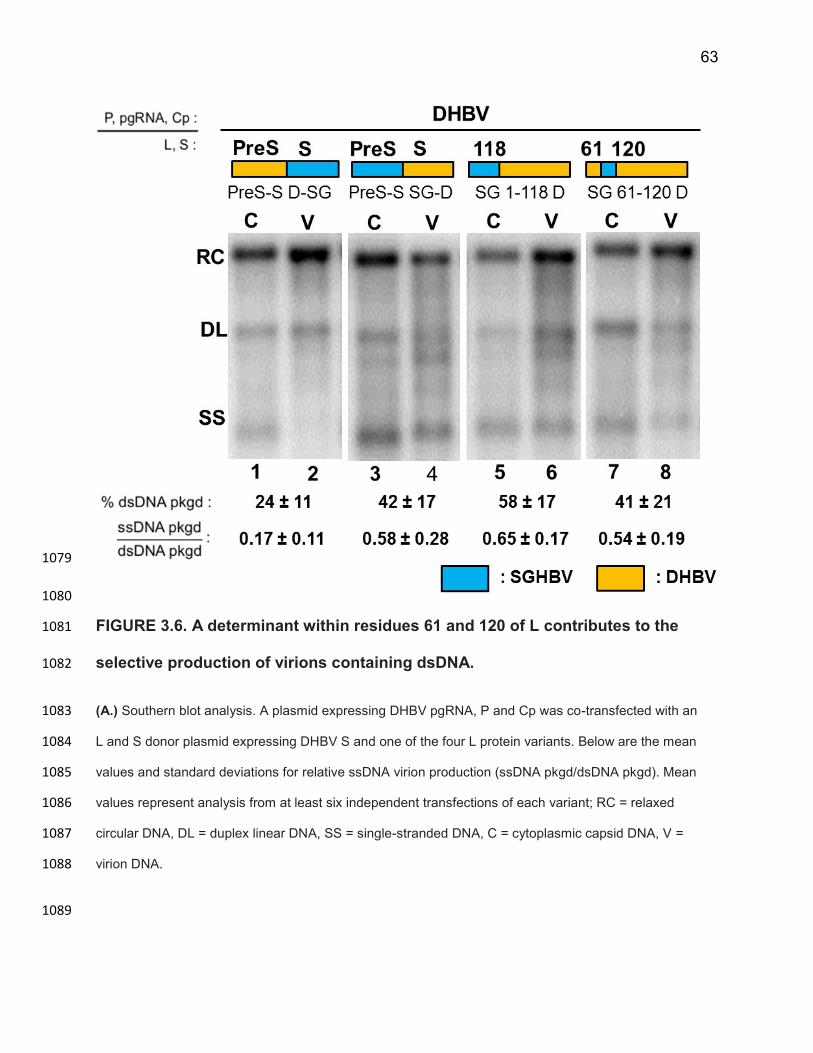
63
1079
1080
FIGURE 3.6. A determinant within residues 61 and 120 of L contributes to the 1081
selective production of virions containing dsDNA. 1082
(A.) Southern blot analysis. A plasmid expressing DHBV pgRNA, P and Cp was co-transfected with an 1083
L and S donor plasmid expressing DHBV S and one of the four L protein variants. Below are the mean 1084
values and standard deviations for relative ssDNA virion production (ssDNA pkgd/dsDNA pkgd). Mean 1085
values represent analysis from at least six independent transfections of each variant; RC = relaxed 1086
circular DNA, DL = duplex linear DNA, SS = single-stranded DNA, C = cytoplasmic capsid DNA, V = 1087
virion DNA. 1088
1089
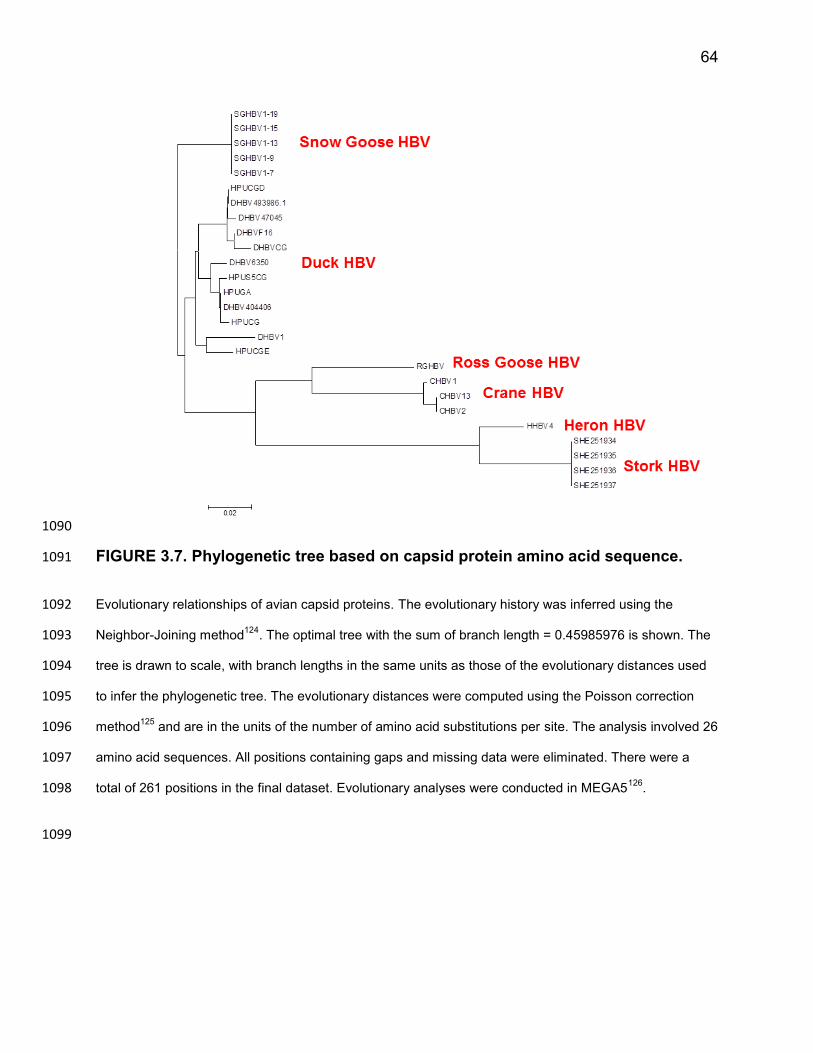
64
1090
FIGURE 3.7. Phylogenetic tree based on capsid protein amino acid sequence. 1091
Evolutionary relationships of avian capsid proteins. The evolutionary history was inferred using the 1092
Neighbor-Joining method124
. The optimal tree with the sum of branch length = 0.45985976 is shown. The 1093
tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used 1094
to infer the phylogenetic tree. The evolutionary distances were computed using the Poisson correction 1095
method125
and are in the units of the number of amino acid substitutions per site. The analysis involved 26 1096
amino acid sequences. All positions containing gaps and missing data were eliminated. There were a 1097
total of 261 positions in the final dataset. Evolutionary analyses were conducted in MEGA5126
. 1098
1099

65
Discussion 1100
SGHBV produces virions containing ssDNA as efficiently as virions containing 1101
dsDNA 1102
SGHBV is the only hepadnavirus known to produce virions containing ssDNA27. 1103
We studied SGHBV alongside DHBV to gain insight into how hepadnaviruses are able 1104
to preferentially produce virions containing mature dsDNA genomes. To measure the 1105
relative ssDNA virion production we calculated the fraction of the total ssDNA molecules 1106
synthesized that were packaged in virions and divided this by the fraction of RC DNA 1107
molecules synthesized that were packaged in virions. We found this value to be 1.01 ± 1108
0.16 for SGHBV and virtually zero for DHBV. This result means that SGHBV produces 1109
virions containing ssDNA as efficiently as virions containing dsDNA. 1110
It would be interesting to know if SGHBV produces virions containing pgRNA. 1111
Because we see such high levels of ssDNA within cytoplasmic capsids and in 1112
extracellular virions, it seems unlikely that SGHBV capsids containing pgRNA are 1113
competent for virion formation and secretion. If they were competent, we would expect 1114
to see mainly RNA-containing SGHBV virions and little, to no, SGHBV virions containing 1115
DNA. This prediction assumes synthesis of ssDNA does not occur rapidly after pgRNA 1116
encapsidation, and occurs more slowly than capsid envelopment. If reverse 1117
transcription occurs rapidly, newly formed capsids containing pgRNA would very quickly 1118
become capsids containing ssDNA. Hence, it is possible that capsids containing pgRNA 1119
would not have enough time to interact with envelope proteins and be packaged in 1120
virions before reverse transcription occurs. Therefore, the best way to determine if 1121

66
SGHBV envelope proteins can productively interact with pgRNA containing capsids to 1122
form pgRNA containing virions would be to use a variant of the SGHBV P protein which 1123
lacks the ability to reverse transcribe pgRNA. In this way, capsids containing pgRNA 1124
would accumulate within the cytoplasm and it would be straightforward to determine if 1125
SGHBV can produce virions containing pgRNA. 1126
Identifying residues of Cp involved in selective production of virions containing 1127
dsDNA 1128
Using genetic complementation between SGHBV and DHBV, we found that 1129
SGHBV Cp makes a major contribution to the ability of SGHBV to produce virions 1130
containing ssDNA. This result was not surprising given the capsid maturation 1131
hypothesis. We then identified the amino acid residues of SGHBV Cp that contribute to 1132
its ability to package capsids containing ssDNA in virions. When residues 74 or 107 of 1133
DHBV Cp were changed to their SGHBV counterparts, DHBV produced virions 1134
containing ssDNA. However, changing only residue 74 of DHBV Cp led to a larger 1135
increase in ssDNA virion production, than when only residue 107 was changed. In fact, 1136
changing only residue 74 of DHBV Cp was sufficient to cause virions containing ssDNA 1137
to be secreted as efficiently as WT SGHBV Cp. In a reciprocal analysis, we changed 1138
residues 74 and 107 in SGHBV Cp to their DHBV counterparts individually and in 1139
combination. We found that changing residues 74 and 107 in combination, reduced 1140
ssDNA virion production to almost undetectable levels. In summary, we were able to 1141
change the respective behaviors of DHBV and SGHBV by making only one or two 1142
substitutions in Cp. 1143

67
These results could mean that residues 74 and 107 contribute to the appearance 1144
of the “capsid packaging signal” on the surface of capsids. It is possible that having an 1145
isoleucine at residue 74 and a glutamic acid at residue 107 causes the constitutive or 1146
early presentation of the “capsid packaging signal”, allowing capsids containing ssDNA 1147
to be enveloped and packaged in virions by DHBV envelope proteins. Determining if 1148
SGHBV capsids containing less than full-length ssDNA or pgRNA are packaged in 1149
virions would determine whether SGHBV capsids appear constitutively mature. 1150
Similarly, we could determine if capsids formed from L74I DHBV Cp or H107E DHBV 1151
Cp appear constitutively mature. If this were the case, structural or biochemical 1152
comparisons of WT DHBV capsids and the variant DHBV capsids could provide insight 1153
into the nature of the “capsid packaging signal”. 1154
Additionally, we found that when residues 74 and/or 107 of HHBV Cp were 1155
changed to their SGHBV counterparts, HHBV did not produce virions containing ssDNA. 1156
Rather, HHBV retained its ability to preferentially produce virions containing mature 1157
dsDNA. These findings suggest that additional residues, aside from 74 and 107, also 1158
dictate whether or not these hepadnaviruses selectively produce virions containing 1159
dsDNA. This result is not surprising when you consider the phylogenetic relationship 1160
between the capsid proteins from the three viruses. DHBV and SGHBV Cp are far more 1161
similar than either is to HHBV Cp (Figure 3.7). There are likely residues which are 1162
conserved in SGHBV and DHBV Cp but are different in HHBV Cp that also contribute to 1163
the ability of these proteins to produce virions containing ssDNA. These would be fairly 1164
straight forward to identify using chimeric SGHBV-HHBV Cp proteins. It would be 1165
interesting to see if changing residues 74 and 107 in Cp of an avihepadnavirus which is 1166

68
more closely related to SGHBV Cp, for example RGHBV Cp (Figure 3.7), would be 1167
sufficient to cause virions containing ssDNA to be produced. This could provide insight 1168
into other residues which contribute to the ability of these viruses to produce virions 1169
containing ssDNA or selectively produce virions containing dsDNA. 1170
Interestingly, similar analyses have been done to determine that changing 1171
residue 97 from a phenylalanine to a leucine in the human HBV capsid protein causes 1172
the production of virions containing ssDNA in cell culture100. For HBV, the amino acid 1173
change causing these different behaviors was a very conservative change and the two 1174
residues had similar hydrophobicities. This is similar to our results; when we made a 1175
highly conservative change at residue 74 in DHBV or SGHBV Cp we saw drastically 1176
different behaviors. The conservative nature of these amino-acid substitutions makes it 1177
difficult to predict how these changes are altering the ability of these hepadnaviruses to 1178
selectively produce virions containing mature dsDNA genomes. It will be interesting to 1179
determine if the mechanism by which these changes are causing the packaging of 1180
ssDNA in virions of HBV and DHBV are similar or distinct and could provide broader 1181
insights into how hepadnaviruses are able to selectively produce virions containing 1182
mature dsDNA genomes. 1183
While evidence for a direct interaction between the capsid and envelope proteins 1184
is lacking, it is possible that we have identified a surface on the exterior of the capsid 1185
that binds to envelope proteins to initiate virion morphogenesis. By changing residues 1186
74 and 107 of DHBV Cp, we may have altered the envelope interacting site on the 1187
surface of the capsid, allowing both mature and immature capsids to productively 1188
interact with envelope proteins to be packaged into a virion. 1189

69
Unfortunately there is no high resolution crystal structure of the DHBV capsid, so 1190
it is impossible to know exactly where these residues lie on the capsid. The structure of 1191
the HBV capsid has been determined47, but DHBV Cp and human HBV Cp are only 1192
weakly phylogenetically related, making it difficult to use the structure of the HBV capsid 1193
to make predictions about the DHBV capsid structure. Using a low resolution structure 1194
of DHBV capsids that has been generated using cryo-electron microscopy and 1195
subsequent 3D image reconstruction48, as well as other models of the DHBV capsid 1196
structure, made using a different approach49,127, we predict that residues 74 and 107 of 1197
DHBV Cp are at, or near, the tip of the capsid spikes. While we cannot say with 1198
certainty whether residues 74 and 107 are buried or exposed on the surface of the 1199
capsid, it is likely that residues 74 and 107 of the DHBV Cp are located on the exterior 1200
of the capsid, rather than disposed towards the lumen of the capsid. As such, these 1201
residues would be in a position to interact with the envelope proteins and/or host factors 1202
during virion morphogenesis. 1203
The role of envelope proteins in virion formation and the selective production of 1204
virions containing mature dsDNA genomes 1205
Interestingly, we found that SGHBV a determinant between residues 61 and 120 1206
of SGHBV L contributes to the production of virions containing ssDNA independent of 1207
SGHBV Cp. This was an unexpected finding because the “capsid maturation” 1208
hypothesis posits selective production of virions containing mature dsDNA genomes is a 1209
function of the capsid and, therefore, is encoded solely by Cp. These results mean that 1210
SGHBV envelope proteins have access to cytoplasmic capsids containing both mature 1211

70
and immature genomes. This observation suggests that localization of capsids to 1212
subcellular sites of envelopment is not dependent on the state of capsid maturation. 1213
A simple explanation for our finding is that (1) the residues between 61 and 120 1214
of L form a surface which interacts with cytoplasmic capsids (2) the capsid-interacting 1215
site formed by the variant SG 61-120 D L (and WT SGHBV L) is different than the 1216
capsid interacting site formed by WT DHBV envelope proteins and (3) this difference 1217
allows SG 61-120 D L (and WT SGHBV L) to facilitate the packaging of capsids into 1218
virions, independent of capsid maturation or the genome within. 1219
Another idea is that hepadnaviruses require an “envelope protein maturation” 1220
which is required for virion formation and maturation of the envelope protein would be 1221
required for it to productively bind to and envelop capsids. One example of such a 1222
change in envelope proteins could be a structural or topological change in the envelope 1223
protein. This would not be hard to imagine given the complex and multiple topologies of 1224
the L protein. For example, this change in topology could allow for membrane curvature 1225
and subsequent budding of the capsid into the secretory vesicle. 1226
Just as genome maturation triggers capsid maturation, it is possible that capsid 1227
maturation triggers envelope protein maturation. Perhaps there are two forms of the L or 1228
S envelope proteins; a “packaging incompetent” and a “packaging competent” form. 1229
Capsid maturation could cause the envelope proteins to shift to a “packaging 1230
competent” form, thereby triggering the interactions required for capsid envelopment 1231
and subsequent packaging into a virion. SGHBV envelope proteins may constitutively 1232
be in a “packaging competent” state (or location), which is why they can envelope 1233

71
capsids independent of capsid maturation and package them in virions. Further, it is 1234
possible that a determinant within amino acids 61 and 120 of SGHBV L causes L to 1235
appear constitutively “mature” or in a “packaging competent” state. 1236
We also found that SGHBV packages a larger fraction of the capsids containing 1237
RC DNA into virions when compared to DHBV. We determined that SGHBV L protein 1238
(and specifically a determinant between residues 61 and 120) is responsible for this 1239
feature. Similarly, the PreS region of DHBV L is responsible for the ability of DHBV to 1240
package a small fraction of capsids containing RC DNA into virions and its ability to 1241
preferentially produce virions containing dsDNA. 1242
One idea is that the ability to package a large fraction of the capsids containing 1243
RC DNA into virions and the ability to produce virions containing ssDNA are related and 1244
the “envelope protein maturation” we propose is a rate-limiting step in virion 1245
morphogenesis. This would be consistent with the finding that SGHBV envelope 1246
proteins contribute to its ability to package a large fraction of the RC DNA synthesized 1247
and the ability to produce virions containing ssDNA. However, we cannot rule out the 1248
possibility that the difference we observed in ability to package capsids containing RC 1249
DNA into virions may be due to differences in expression or steady-state levels of the 1250
respective envelope proteins, in addition to or rather than intrinsic differences in the 1251
properties of these proteins. The next step to understanding the L protein’s role in the 1252
selective production of virions containing mature dsDNA genomes and in dictating the 1253
proportion of capsids that get packaged into virions would be to more precisely map the 1254
determinants in L that are responsible for the respective behaviors of SGHBV and 1255
DHBV. Determining if high/low dsDNA virion production and immature/selective 1256

72
production of virions map to the same or different residue(s) could provide insight into 1257
why SGHBV and DHBV envelope proteins exhibit these different behaviors. 1258

73
1259
1260
CHAPTER 4 1261
1262
1263
IDENTIFYING AMINO ACID RESIDUES OF 1264
AVIHEPADNAVIRAL CAPSID AND ENVELOPE PROTEINS 1265
THAT CONTRIBUTE TO THE PACKAGING OF CAPSIDS INTO 1266
VIRIONS 1267
1268
Studies from this chapter will be continued and expanded upon by 1269
Dan Loeb and Karolyn Pionek 1270

74
Abstract 1271
Hepadnaviruses are enveloped dsDNA viruses whose large (L) and small (S) envelope 1272
proteins are required for virion formation. Hepadnaviruses replicate their genomes 1273
within cytoplasmic capsids, through reverse transcription. The structural subunit of the 1274
capsid is the capsid protein (Cp). It is thought that the L protein interacts with 1275
cytoplasmic capsids to (1) regulate cccDNA amplification and (2) envelope and package 1276
capsids containing dsDNA into virions. In this chapter, I describe a serendipitously- 1277
discovered inability of HHBV envelope proteins to package DHBV and SGHBV capsids 1278
into virions and illustrate how we used this as an opportunity to identify residues of Cp 1279
that are involved in capsid packaging and virion formation. We found that substituting a 1280
region between residues 69-114 of HHBV Cp into DHBV Cp was sufficient to restore the 1281
ability of HHBV envelope proteins to package DHBV capsids into virions. This suggests 1282
that a determinant between residues 69 and 114 of Cp contributes to capsid packaging 1283
into virions and possibly interacts with envelope proteins during virion morphogenesis. 1284
Within this region, DHBV Cp and HHBV Cp differ at only eleven amino acids. A similar 1285
approach can be taken to identify regions of the envelope proteins that are involved in 1286
capsid packaging/virion production. 1287
1288

75
Introduction 1289
Human hepatitis B virus is the prototype member of the Hepadnaviridae family of 1290
viruses. This family includes viruses which can infect a variety of mammals 1291
(orthohepadnaviruses) and a variety of birds (avihepadnaviruses); all have a very 1292
narrow host range19,21. All hepadnaviruses are enveloped viruses and have similar 1293
virion architectures (Figure 1.1). The virion core contains the viral polymerase protein 1294
(P) that is covalently attached to the 5’ end of the minus-strand of its dsDNA genome. 1295
The genome and P protein are enclosed within a protein shell, known as a capsid. The 1296
capsid is made up of 240 copies of the capsid protein (Cp). Around the capsid is a 1297
lipoprotein shell, known as the envelope. The envelope of the virion consists of host-1298
derived lipids and several viral envelope proteins; for avihepadnaviruses these are the 1299
large (L) and the small (S) surface proteins (Figure 1.1A). 1300
Hepadnaviruses are dsDNA viruses which replicate their genomes through 1301
reverse transcription of an RNA intermediate known as the pregenomic RNA (pgRNA). 1302
Genome replication occurs within cytoplasmic capsids. This means cytoplasmic capsids 1303
contain an array of viral replicative intermediates, ranging from pgRNA to dsDNA. A 1304
hallmark feature of hepadnaviral virion morphogenesis is that capsids containing dsDNA 1305
are preferentially packaged into virions98,121,128. Capsids containing ssDNA or pgRNA 1306
are retained within the cell. It is thought that the surface of capsids containing mature 1307
dsDNA differ from capsids containing ssDNA. This difference is thought to arise during 1308
dsDNA synthesis and is referred to as the “capsid packaging signal” or “capsid 1309
maturation signal”. This “signal” is thought to render the capsids containing dsDNA 1310

76
competent for the interactions required to be enveloped and packaged into a virion. The 1311
nature of this “signal” is not known. 1312
During virion morphogenesis, it is thought that cytoplasmic capsids containing 1313
dsDNA interact with the envelope proteins at a post-ER pre-Golgi vesicular membrane, 1314
are enveloped as they bud into this secretory vesicle and are released from the cell via 1315
a constitutive secretion pathway (for reviews, see references17,31,90). L and S are both 1316
required for virion production18,129, which suggests they play an active role in 1317
coordinating capsid envelopment/packaging into virions. 1318
Because all hepadnaviruses share a similar replication strategy, related family 1319
members have been invaluable in understanding the replication of HBV. For example, 1320
duck hepatitis B virus (DHBV) has been studied extensively to investigate many aspects 1321
of hepadnaviral biology24. In this chapter, DHBV and other avian hepadnaviruses, heron 1322
hepatitis B virus (HHBV) and snow goose hepatitis B virus (SGHBV), were used to 1323
identify regions of the capsid protein involved in virion morphogenesis. We attempted to 1324
confer the ability to produce virions containing ssDNA onto HHBV, through genetic 1325
complementation. However, instead we serendipitously discovered that HHBV envelope 1326
proteins cannot package SGHBV or DHBV capsids into virions. 1327
We saw this incompatibility between HHBV envelope proteins and SGHBV and 1328
DHBV capsids as an opportunity to identify regions of Cp and envelope proteins that 1329
contribute to virion production. When we substituted the region between amino acids 69 1330
and 114 of HHBV Cp into DHBV Cp, the ability of HHBV envelope proteins to package 1331
these capsids into virions was restored. This result indicates that a determinant between 1332

77
amino acids 69 and 114 of Cp is involved in virion production and could possibly be part 1333
of the envelope interacting site on the surface of the capsid. Within this region, DHBV 1334
Cp and HHBV Cp differ at only eleven amino acid residues; we plan to more precisely 1335
map the residue(s) involved in capsid packaging and virion production. Taking a similar 1336
strategy, we also plan to identify regions of the envelope proteins involved in virion 1337
production. 1338

78
Results 1339
HHBV envelope proteins cannot package SGHBV or DHBV capsids into virions 1340
We were able to confer the ability to efficiently package capsids containing 1341
ssDNA into virions onto DHBV by supplying SGHBV Cp to DHBV pgRNA, P, L and S 1342
(reference 130 and chapter 3 of this dissertation). We hypothesized that we could 1343
confer the ability to produce virions containing ssDNA onto HHBV using a similar 1344
strategy. To test this prediction, we co-expressed HHBV P, pgRNA and envelope 1345
proteins with SGHBV Cp in LMH cells. We did this by complementing an HHBV plasmid 1346
deficient in expressing Cp, HHBVpgRNA+P+Env+, with an SGHBV Cp donor plasmid 1347
SGHBVC+ (Figure 4.1A lanes 3 and 4). We measured the levels of RC DNA in both 1348
cytoplasmic capsids and saw no defect in cytoplasmic DNA synthesis, allowing us to 1349
evaluate virion production. 1350
It was immediately apparent that virion production was greatly reduced compared 1351
to either the HHBV or SGHBV comparisons (Figure 4.1A), but in an effort to be 1352
objective and to allow us to see intermediate levels of virion production we quantified 1353
dsDNA virion production. To quantitatively describe dsDNA virion production, we 1354
divided the amount of RC DNA packaged in virions by the sum of RC DNA found in 1355
cytoplasmic capsids and extracellular virions and multiplied this value by 100 to obtain a 1356
percentage, as shown below: 1357
% dsDNA packaged = (RC DNAvirions / (RC DNAcapsids + RC DNAvirions)) x 100 1358
For the HHBV, this value was 34 ± 11%, and for SGHBV it was 55 ± 5% (Figure 4.1A 1359
lanes 1-2 and 7-8). When we supplied SGHBV Cp to the corresponding HHBV 1360

79
components as described above, we saw that dsDNA virion production was reduced to 1361
4 ± 1% (Figure 4.1A lanes 3-4). This suggested HHBV envelope proteins poorly 1362
facilitate the packaging of SGHBV caspids into virions. 1363
To strengthen this interpretation, we measured the ability of HHBV envelope 1364
proteins to package SGHBV capsids into virions, using a different genetic 1365
complementation strategy. We co-expressed HHBV envelope proteins with SGHBV Cp, 1366
P and pgRNA in LMH cells, using plamsids HHBVEnv+ and SGHBVpgRNA+P+C+. We 1367
measured dsDNA virion production and found that it was essentially zero (Figure 4.2B 1368
lanes 2-3). This was drastically lower than the HHBV and SGHBV comparisons (Figure 1369
4.2B lane 1-2 and 5-6), where % dsDNA packaged was 30 ± 10% and 45 ± 20%, 1370
respectively. This result strengthened our interpretation that HHBV envelope proteins 1371
are not able to interact with and package SGHBV capsids into virions. Interestingly, 1372
SGHBV envelope proteins can package HHBV capsids into virions (Figure 4.1A lanes 1373
9-10, Figure 4.1B lanes 7-8) and can even confer the ability to package capsids 1374
containing ssDNA into virions onto HHBV (Figure 4.1B lanes 7-8). Hence, the 1375
incompatibility is specifically between HHBV envelope proteins and SGHBV capsids 1376
and is not reciprocal. 1377
Given the high phylogenetic similarity between SGHBV and DHBV Cp (Figure 1378
3.7) and the fact that both are distantly related to HHBV Cp, we predicted that HHBV 1379
envelope proteins would not be able to interact with and package DHBV capsids into 1380
virions either. To test this prediction, we performed experiments similar to those 1381
described above, except in place of SGHBV components, we used DHBV components 1382
(Figure 4.2 A and B). We found that HHBV envelope proteins did not package DHBV 1383

80
capsids into virions, regardless of the pgRNA or P protein present (Figure 4.2A lanes 3-1384
4, Figure 4.2B lanes 3-4). And that the incompatibility was not reciprocal; when we 1385
supplied DHBV envelope proteins to HHBV capsids we saw virions containing dsDNA 1386
were produced as efficiently as the DHBV comparisons (Figure 4.2A lanes 7-10 and 1387
4.2B lanes 5-8). This held true regardless of the origin of the pgRNA or P protein. Taken 1388
together, we conclude that HHBV envelope proteins poorly facilitate the packaging of 1389
SGHBV or DHBV capsids into virions. 1390
A determinant within amino acid residues 69 and 114 of Cp is involved in virion 1391
production 1392
One interpretation of the above results is that HHBV envelope proteins cannot 1393
interact with SGHBV or DHBV capsids because the “envelope-interacting site” on the 1394
surface of these capsids differs from the “envelope-interacting site” on HHBV capsids. If 1395
this were true, we predicted that we could substitute the portion of HHBV Cp containing 1396
the “envelope-interacting site” into SGHBV or DHBV Cp and restore the interaction 1397
between HHBV envelope proteins and these capsids. To this end, we made chimeric 1398
SGHBV-HHBV and DHBV-HHBV capsid proteins, co-expressed them with HHBV 1399
pgRNA, P, L and S and measured dsDNA virion production. 1400
We initially substituted a region between residues 22 and 139 of HHBV Cp into 1401
SGHBV Cp; within this region, SGHBV Cp and HHBV Cp differ at seventeen amino acid 1402
residues. We called this Cp variant H 22-139 SG Cp. When we co-expressed H 22-139 1403
SG Cp with HHBV P, pgRNA, L and S, we found that 31 ± 2% of the RC DNA 1404
synthesized was packaged into virions (Figure 4.1A lanes 5-6); which was not 1405

81
statistically different than the HHBV comparison, where % dsDNA production was 34 ± 1406
11% (Figure 4.1A lanes 1-2). This result indicated that substituting the region between 1407
22 and 139 into SGHBV Cp was sufficient to restore an interaction between these 1408
capsids and HHBV envelope proteins. Suggesting a determinant between amino acid 1409
residues 22 and 139 of Cp is involved in virion production and possibly interacts with the 1410
envelope proteins during capsid envelopment/packaging into virions. 1411
We more precisely mapped the region of Cp responsible for the observed 1412
species-specific incompatibility using a chimeric DHBV-HHBV Cp variant; we 1413
substituted a region between amino acids 69 and 114 of HHBV Cp into DHBV Cp and 1414
co-expressed this chimeric Cp (H 69-114 D Cp) with HHBV envelope proteins, P and 1415
pgRNA. We saw no defect in DNA synthesis, allowing us to evaluate virion production. 1416
We found that 33 ± 16% of the RC DNA molecules produced were packaged into virions 1417
(Figure 4.1A lanes 5-6); this value was not statistically different than the HHBV 1418
comparison, 34 ± 11% (Figure 4.2A lanes 1-2). Indicating a determinant within residues 1419
69 and 114 of Cp is involved in virion production; within this region, HHBV Cp and 1420
DHBV Cp differ at only eleven residues. It is possible that a determinant within this 1421
region forms a surface on the exterior of the capsid which interacts with the envelope 1422
proteins during capsid envelopment or viral egress. 1423
1424
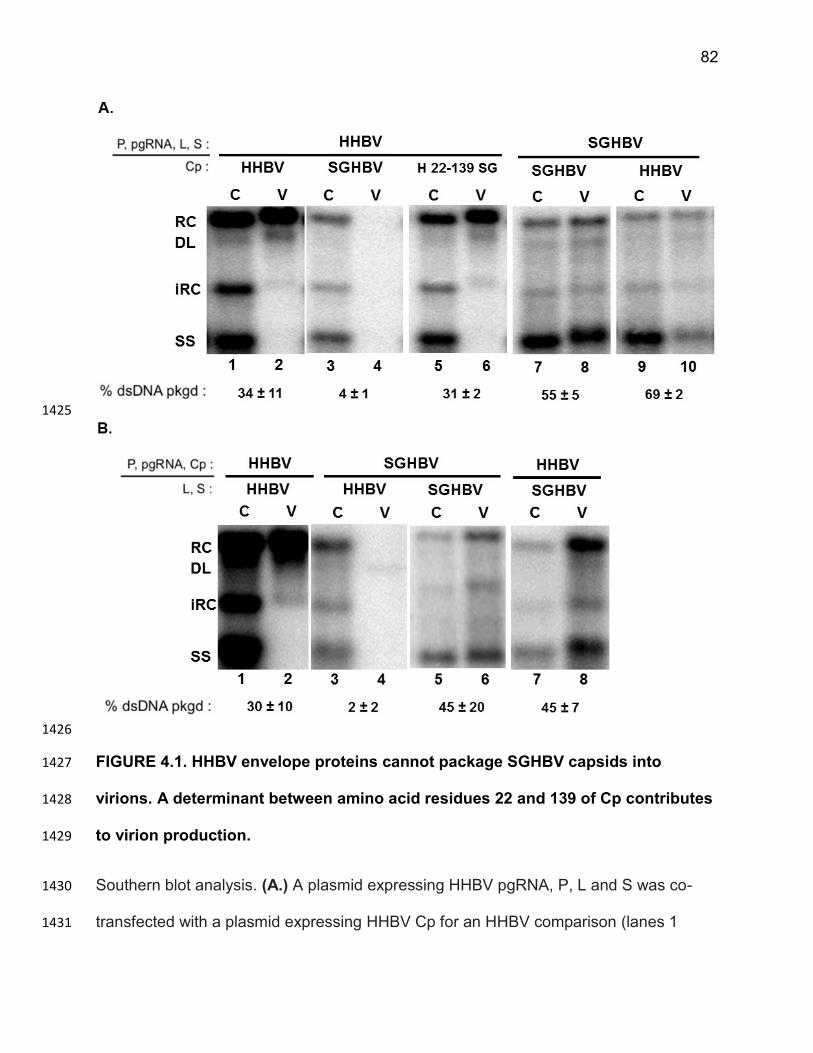
82
1425
1426
FIGURE 4.1. HHBV envelope proteins cannot package SGHBV capsids into 1427
virions. A determinant between amino acid residues 22 and 139 of Cp contributes 1428
to virion production. 1429
Southern blot analysis. (A.) A plasmid expressing HHBV pgRNA, P, L and S was co-1430
transfected with a plasmid expressing HHBV Cp for an HHBV comparison (lanes 1 1431

83
and 2), SGHBV Cp (lanes 3 and 4) or H 22-139 SG Cp (lanes 5 and 6). A plasmid 1432
expressing SGHBV pgRNA, P, L and S was co-transfected with a plasmid expressing 1433
SGHBV Cp (lanes 7 and 8) for an SGHBV comparison or HHBV Cp (lanes 9 and 10). 1434
(B.) A plasmid expressing HHBV pgRNA, P and Cp was co-expressed with a plasmid 1435
expressing HHBV L and S (lanes 1 and 2) for an HHBV comparison or a plasmid 1436
expressing SGHBV L and S (lanes 7 and 8). A plasmid expressing SGHBV pgRNA, P 1437
and Cp was co-transfected with a plasmid expressing SGHBV L and S (lanes 5 and 6) 1438
for a SGHBV comparison or a plasmid expressing HHBV L and S (lanes 3 and 4). 1439
Below are the mean values and standard deviations for dsDNA virion production (% 1440
dsDNA pkgd); RC = relaxed circular DNA, DL = duplex linear DNA, SS = single-1441
stranded DNA, C = cytoplasmic capsid DNA, V = virion DNA. 1442
1443
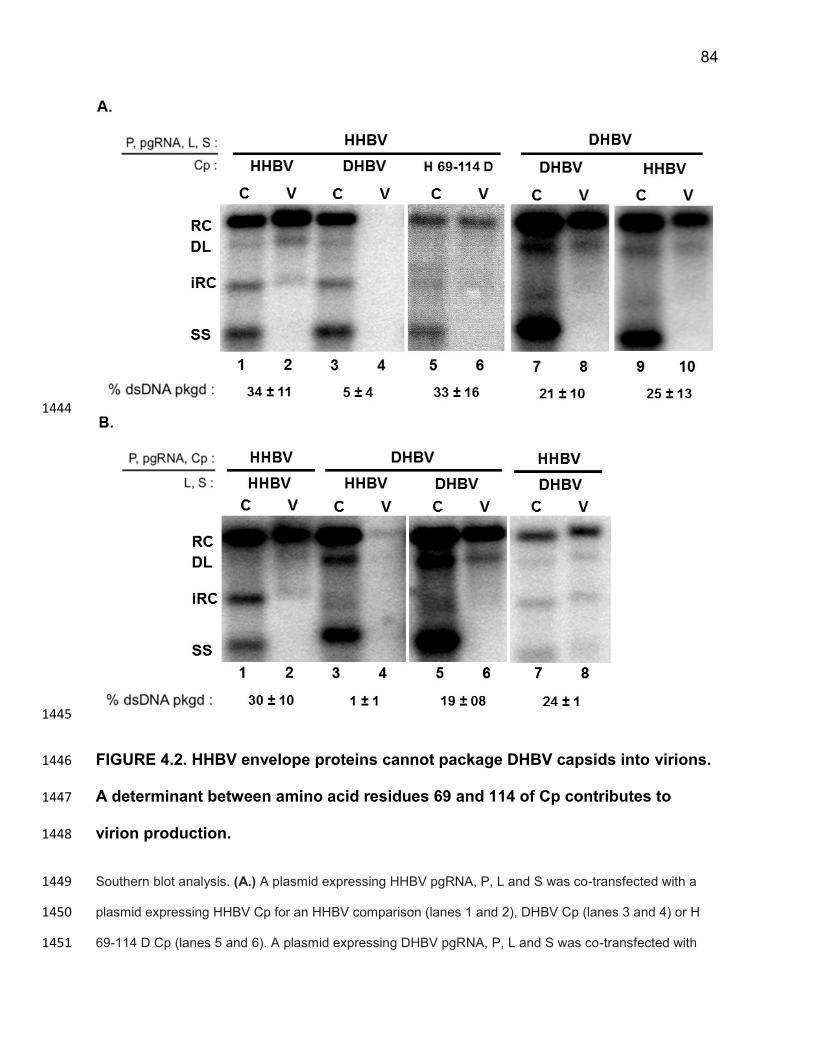
84
1444
1445
FIGURE 4.2. HHBV envelope proteins cannot package DHBV capsids into virions. 1446
A determinant between amino acid residues 69 and 114 of Cp contributes to 1447
virion production. 1448
Southern blot analysis. (A.) A plasmid expressing HHBV pgRNA, P, L and S was co-transfected with a 1449
plasmid expressing HHBV Cp for an HHBV comparison (lanes 1 and 2), DHBV Cp (lanes 3 and 4) or H 1450
69-114 D Cp (lanes 5 and 6). A plasmid expressing DHBV pgRNA, P, L and S was co-transfected with 1451

85
a plasmid expressing DHBV Cp (lanes 7 and 8) for a DHBV comparison or HHBV Cp (lanes 9 and 10). 1452
(B.) A plasmid expressing HHBV pgRNA, P and Cp was co-expressed with a plasmid expressing HHBV 1453
L and S (lanes 1 and 2) for an HHBV comparison or a plasmid expressing DHBV L and S (lanes 7 and 1454
8). A plasmid expressing DHBV pgRNA, P and Cp was co-transfected with a plasmid expressing DHBV 1455
L and S (lanes 5 and 6) for a DHBV comparison or a plasmid expressing HHBV L and S (lanes 3 and 1456
4). Below are the mean values and standard deviations for dsDNA virion production (% dsDNA pkgd); 1457
RC = relaxed circular DNA, DL = duplex linear DNA, SS = single-stranded DNA, C = cytoplasmic capsid 1458
DNA, V = virion DNA. 1459
1460
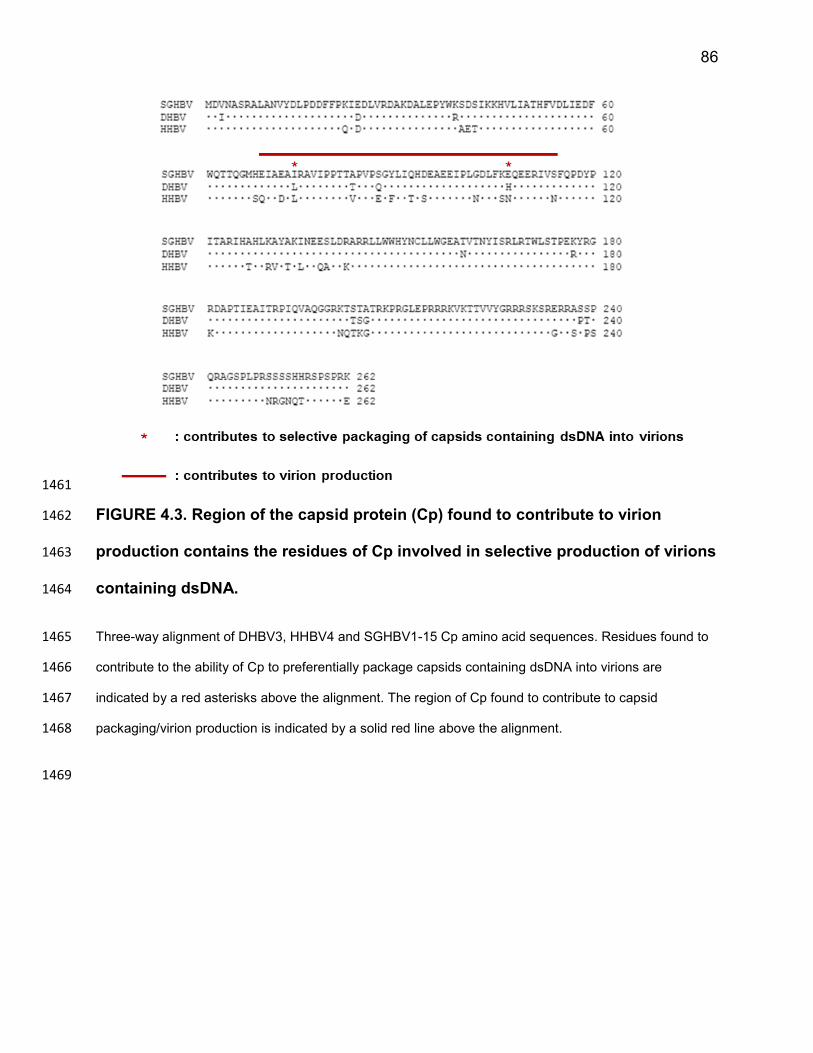
86
1461
FIGURE 4.3. Region of the capsid protein (Cp) found to contribute to virion 1462
production contains the residues of Cp involved in selective production of virions 1463
containing dsDNA. 1464
Three-way alignment of DHBV3, HHBV4 and SGHBV1-15 Cp amino acid sequences. Residues found to 1465
contribute to the ability of Cp to preferentially package capsids containing dsDNA into virions are 1466
indicated by a red asterisks above the alignment. The region of Cp found to contribute to capsid 1467
packaging/virion production is indicated by a solid red line above the alignment. 1468
1469

87
Discussion 1470
In this chapter, I provide evidence that HHBV envelope proteins cannot 1471
productively interact with DHBV or SGHBV capsids to package these capsids into 1472
virions. I found that substituting the region between residues 69 and 114 of HHBV Cp 1473
into SGHBV Cp was sufficient to restore the ability of these capsids to be packaged into 1474
virions by HHBV envelope proteins. This result indicates that a determinant within 1475
amino acid residues 69 and 114 of Cp is involved in capsid packaging/virion production 1476
and possibly forms a surface on the exterior of the capsid that interacts with the 1477
envelope proteins during virion morphogenesis. Interestingly, the residues we found to 1478
be involved in the selective production of virions containing ssDNA (74 and 107) lie 1479
within this region (Figure 4.3); suggesting residues within this region of Cp form an 1480
important surface on the exterior of the capsid. 1481
Using a similar approach, regions of the envelope proteins involved in capsid 1482
packaging/virion production can be identified. For example, regions of DHBV envelope 1483
proteins can be substituted into DHBV envelope proteins and supplied to DHBV 1484
capsids, in an attempt to restore the interaction of HHBV envelope proteins with DHBV 1485
capsids and restore capsid packaging/virion production. The residues identified in L 1486
could interact with itself and/or the S protein within the membrane in which they reside, 1487
to form an interface that cytoplasmic capsids can interact with to initiate capsid 1488
envelopment and packaging into a virion. It will be interesting to see if the residue(s) of 1489
L that we find to contribute to capsid envelopment/packaging into a virion maps to the 1490
same or different residue(s) we found to be involved in the selective production of 1491
virions containing dsDNA (in Chapter 3). 1492

88
1493
1494
CHAPTER 5 1495
1496
1497
SUMMARY AND FUTURE DIRECTIONS 1498
1499

89
Chronic infection of HBV is a major health concern because it is one of the 1500
leading causes of hepatocellular carcinoma worldwide. While therapies exist, they 1501
cannot cure a chronic HBV infection. In recent years, there has been an increased 1502
interest and effort by pharmaceutical companies to come up with a cure for chronic HBV 1503
infection. To better identify antiviral drug targets and develop new treatment options that 1504
rid infected individuals of the virus, there is a need to better understand HBV replication 1505
and biology. The goal of my dissertation research was to better understand, 1506
mechanistically, how infectious viral particles are formed, with the hope that a better 1507
understanding of this process could provide insight into how to disrupt HBV replication. 1508
HBV is a double-stranded DNA (dsDNA) virus that replicates its genome through 1509
reverse transcription. Genome replication takes place entirely within cytoplasmic 1510
capsids. HBV is also an enveloped virus, which means the capsid itself is surrounded 1511
by a lipo-protein shell, known as an envelope. How the capsid acquires an envelope 1512
and is packaged into virions is incompletely understood in HBV replication. 1513
When I started graduate school, those in the field knew that capsid envelopment 1514
and virion formation do not occur in the absence of envelope proteins. This knowledge 1515
suggested that the envelope proteins actively participate in coordinating the packaging 1516
of capsids into virions by interacting with cytoplasmic capsids to facilitate their 1517
envelopment and packaging into a virion. Hepadnavirologists also knew that only 1518
capsids containing dsDNA genomes are packaged into virions, but the mechanisms 1519
underlying the ability of hepadnaviruses to selectively package capsids containing 1520
dsDNA into virions was not defined. It was thought that at some point during DNA 1521
synthesis, capsids acquire a property and/or undergo a conformational change that 1522

90
allows them to interact with envelope proteins and packaged into virions. The 1523
acquisition of this property is referred to as “capsid maturation”. 1524
Selective packaging of capsids containing mature genomes is a general 1525
characteristic of hepadnaviruses. However, one hepadnavirus, SGHBV, does not 1526
adhere to this principle. SGHBV produces virions containing ssDNA. Because of this 1527
unique feature of SGHBV, Michael Hayes, a former research technician in our lab, 1528
proposed to use SGHBV to study “capsid maturation” and selective packaging of 1529
capsids containing dsDNA into virions. He laid the foundation for much of the work I 1530
have done by obtaining a plasmid expressing SGHBV that he used to design and test 1531
several of the initial SGHBV plasmids from which all of our SGHBV plasmids were 1532
derived. 1533
The ultimate goals of my research were to better understand (1) how cytoplasmic 1534
capsids acquire an envelope and package capsids into virions and (2) how 1535
hepadnaviruses are able to discriminate between capsids containing mature dsDNA 1536
genomes and capsids containing immature ssDNA or pgRNA to selectively produce 1537
virions containing dsDNA. However, my initial goal was to determine why SGHBV lacks 1538
a mechanism for discriminating between capsids containing mature dsDNA and 1539
immature ssDNA the different types of capsids and packages both of these types of 1540
capsids into virions. As Mike Hayes proposed, if we can determine how and why 1541
SGHBV is able to produce virions containing ssDNA, we can begin to understand how 1542
all other hepadnaviruses are able to selectively produce virions containing dsDNA. 1543

91
Early in my graduate studies I determined that when SGHBV Cp and envelope 1544
proteins were combined with either DHBV or HHBV ssDNA and P, virions containing 1545
ssDNA were produced. This result meant that SGHBV Cp and/or the envelope proteins 1546
were contributing to the ability of SGHBV to produce virions containing ssDNA. My next 1547
step was to determine if any of these proteins (SGHBV Cp, L or S proteins) were 1548
sufficient to cause the production of virions containing ssDNA. To this end, I supplied 1549
SGHBV Cp or L and S to the remaining DHBV or HHBV viral components necessary to 1550
form a virion and evaluated ssDNA virion production. From these initial experiments, I 1551
uncovered several unappreciated properties of the capsid and envelope proteins of 1552
these three avian hepadnaviruses. 1553
I found that SGHBV envelope proteins and Cp independently contribute to the 1554
ability of SGHBV to produce virions containing ssDNA, as I present in Chapter 3 and in 1555
our recent publication130. I also found that HHBV envelope proteins cannot package 1556
SGHBV or DHBV capsids into virions, which I present in Chapter 4. I devised a way to 1557
use these findings to identify amino acid residues within Cp and L that are involved in 1558
(1) preferential production of virions containing dsDNA and (2) capsid packaging into 1559
virions. 1560
Identifying amino acids residues within Cp that are involved in the preferential 1561
production of virions containing dsDNA and capsid packaging into virions 1562
I found that making a highly conservative change at residue 74 in DHBV or 1563
SGHBV Cp drastically altered the behaviors of these proteins. I was able to convert 1564
DHBV Cp from a producer of virions containing dsDNA to a producer of virions 1565

92
containing ssDNA. I was similarly able to convert SGHBV Cp from a producer of virions 1566
containing ssDNA to a preferential producer of virions containing dsDNA. Indicating that 1567
residue 74 of Cp contributes to the ability of hepadnaviruses to selectively package 1568
capsids containing dsDNA, perhaps by contributing to capsid maturation or the 1569
presentation of the “capsid packaging signal” on the surface of the capsid. The fact that 1570
these amino acid changes were so conservative, suggests that the “capsid maturation 1571
signal” could be quite subtle. 1572
I found that a determinant within amino acid residues 69 and 114 of Cp is 1573
involved in virion production. I propose that this region forms a surface on the exterior of 1574
the capsid that contributes to its subsequent packaging into a virion by interacting with 1575
the envelope proteins to initiate viral morphogenesis. This finding, along with others 1576
described in Chapter 4, suggest that the L protein-capsid interactions that occur during 1577
virion morphogenesis are direct and specific. The fact that the residue(s) I determined to 1578
contribute to the selective production of virions containing ssDNA lie within residues 69 1579
and 114 of Cp, suggests that this region of Cp forms an important surface on the 1580
exterior of the capsid. Lab mates, Dan Loeb and Karolyn Pionek, were able to take over 1581
these studies and have preliminary evidence that a determinant involved in capsid 1582
packaging and virion production is at residue 69. The proximity of this residue to the 1583
residue found to be involved in selective dsDNA virion production, residue 74, suggests 1584
that these amino acid residues contribute to the formation of a very important surface on 1585
the capsid exterior. It is possible that the envelope proteins interact with these residues, 1586
and other residues nearby, during virion morphogenesis to facilitate capsid packaging 1587
and virion production. Structural determinations of DHBV capsids would provide insight 1588

93
into the location of residues 69 and 74 on the capsid. Further, comparisons of the WT 1589
DHBV capsid structure and the capsid structure formed by the DHBV Cp variant which 1590
causes capsids containing ssDNA to be packaged into virions (L74I DHBV Cp) could 1591
provide insight into the nature of the “capsid packaging signal”. 1592
Interestingly, similar analyses have been done to determine that changing 1593
residue 97 from a phenylalanine to a leucine in the human HBV capsid protein causes 1594
the production of virions containing ssDNA in cell culture100. For HBV, the amino acid 1595
change causing these different behaviors was also a very conservative change and the 1596
two residues had similar hydrophobicities. It will be interesting to determine if the 1597
mechanism by which these changes are causing the packaging of capsids containing 1598
ssDNA into virions of HBV and DHBV are similar or distinct and could provide broader 1599
insights into how hepadnaviruses are able to selectively produce virions containing 1600
mature dsDNA genomes. 1601
Identifying amino acids residues within L that are involved in the preferential 1602
production of virions containing dsDNA and capsid packaging into virions 1603
I determined that the L protein contributes to the ability of hepadnaviruses to 1604
selectively produce virions containing mature dsDNA genomes (Chapter 3 and our 1605
recent publication, reference 130). This suggests that the L protein can actively sense 1606
and select capsids containing mature dsDNA capsids to be packaged into virions, while 1607
excluding capsids containing immature ssDNA. This finding sheds new light on the 1608
mechanisms underlying virion morphogenesis and challenges the dogma that “capsid 1609

94
maturation”, and therefore Cp, is solely responsible for the selective production of 1610
virions containing mature dsDNA genomes. 1611
Thus far, the region of L that contributes to the selective production of virions 1612
containing dsDNA has been mapped to the region between residues 61 and 120. Within 1613
this region, DHBV and SGHBV differ at only seven residues. I am currently mapping 1614
which of these seven residues contribute to the ability of hepadnaviruses to selectively 1615
package capsids containing dsDNA into virions. Ultimately, these results will be 1616
submitted for publication. 1617
As I did with Cp, I wanted to determine which residues of L are involved in capsid 1618
packaging/virion formation by exploiting the species-specific incompatibility between 1619
HHBV envelope proteins and DHBV or SGHBV capsids. Unfortunately, I will not be able 1620
to complete these analyses during my time at UW. However, I initiated collaboration 1621
with two lab mates, Dan Loeb and Karolyn Pionek, who will be continuing these studies 1622
to work on this project so that it can be completed and published in a timely manner. 1623
1624

95
References 1625
1. Maddrey, W. C. Hepatitis B--an important public health issue. Clin. Lab. 47, 51–5 1626
(2001). 1627
2. Rehermann, B. & Nascimbeni, M. Immunology of hepatitis B virus and hepatitis C 1628 virus infection. Nat. Rev. Immunol. 5, 215–29 (2005). 1629
3. Lavanchy, D. Worldwide epidemiology of HBV infection, disease burden, and 1630 vaccine prevention. J. Clin. Virol. 34 Suppl 1, S1–3 (2005). 1631
4. Lavanchy, D. Viral hepatitis: global goals for vaccination. J. Clin. Virol. 55, 296–1632
302 (2012). 1633
5. Ott, J. J., Stevens, G. A., Groeger, J. & Wiersma, S. T. Global epidemiology of 1634 hepatitis B virus infection: new estimates of age-specific HBsAg seroprevalence 1635 and endemicity. Vaccine 30, 2212–9 (2012). 1636
6. Jilbert, a R. et al. Characterization of age- and dose-related outcomes of duck 1637 hepatitis B virus infection. Virology 244, 273–82 (1998). 1638
7. McMahon, B. J. et al. Acute hepatitis B virus infection: relation of age to the 1639
clinical expression of disease and subsequent development of the carrier state. J. 1640 Infect. Dis. 151, 599–603 (1985). 1641
8. Hyams, K. C. Risks of chronicity following acute hepatitis B virus infection: a 1642 review. Clin. Infect. Dis. 20, 992–1000 (1995). 1643
9. Beasley, R. P. Rocks along the road to the control of HBV and HCC. Ann. 1644 Epidemiol. 19, 231–4 (2009). 1645
10. Beasley, R. P., Hwang, L. Y., Lin, C. C. & Chien, C. S. Hepatocellular carcinoma 1646 and hepatitis B virus. A prospective study of 22 707 men in Taiwan. Lancet 2, 1647
1129–33 (1981). 1648
11. Parkin, D. M., Bray, F., Ferlay, J. & Pisani, P. Global Cancer Statistics, 2002. CA. 1649 Cancer J. Clin. 55, 74–108 (2005). 1650
12. Chang, M. H. et al. Universal hepatitis B vaccination in Taiwan and the incidence 1651
of hepatocellular carcinoma in children. Taiwan Childhood Hepatoma Study 1652 Group. N. Engl. J. Med. 336, 1855–9 (1997). 1653
13. Wang, X.-Y. & Chen, H.-S. Emerging antivirals for the treatment of hepatitis B. 1654 World J. Gastroenterol. 20, 7707–17 (2014). 1655

96
14. Santantonio, T. A. & Fasano, M. Chronic hepatitis B: Advances in treatment. 1656 World J. Hepatol. 6, 284–92 (2014). 1657
15. Foundation, H. B. Compounds in Development for Chronic Hepatitis B. HBV Drug 1658 Watch at <http://www.hepb.org/professionals/hbf_drug_watch.htm> 1659
16. Menéndez-Arias, L., Alvarez, M. & Pacheco, B. Nucleoside/nucleotide analog 1660
inhibitors of hepatitis B virus polymerase: mechanism of action and resistance. 1661 Curr. Opin. Virol. 8C, 1–9 (2014). 1662
17. Bruss, V. Hepatitis B virus morphogenesis. World J. Gastroenterol. 13, 65–73 1663
(2007). 1664
18. Bruss, V. & Ganem, D. The role of envelope proteins in hepatitis B virus 1665 assembly. Proc. Natl. Acad. Sci. U. S. A. 88, 1059–63 (1991). 1666
19. Dallmeier, K., Schultz, U. & Nassal, M. Heterologous replacement of the 1667 supposed host determining region of avihepadnaviruses: high in vivo infectivity 1668 despite low infectivity for hepatocytes. PLoS Pathog. 4, e1000230 (2008). 1669
20. Trueba, D. et al. Transmission of ground squirrel hepatitis virus to homologous 1670 and heterologous hosts. Hepatology 5, 435–9 1671
21. Seeger, C., Marion, P. L., Ganem, D. & Varmus, H. E. In vitro recombinants of 1672
ground squirrel and woodchuck hepatitis viral DNAs produce infectious virus in 1673 squirrels. J. Virol. 61, 3241–7 (1987). 1674
22. Barker, L. F. et al. Transmission of type B viral hepatitis to chimpanzees. J. Infect. 1675
Dis. 127, 648–62 (1973). 1676
23. Mason, W. S., Seal, G. & Summers, J. Virus of Pekin ducks with structural and 1677 biological relatedness to human hepatitis B virus. J. Virol. 36, 829–36 (1980). 1678
24. Schultz, U., Grgacic, E. & Nassal, M. Duck hepatitis B virus: an invaluable model 1679 system for HBV infection. Adv. Virus Res. 63, 1–70 (2004). 1680
25. Kawaguchi, T., Nomura, K., Hirayama, Y. & Kitagawa, T. Establishment and 1681 characterization of a chicken hepatocellular carcinoma cell line, LMH. Cancer 1682
Res. 47, 4460–4 (1987). 1683
26. Summers, J., Smolec, J. M. & Snyder, R. A virus similar to human hepatitis B 1684 virus associated with hepatitis and hepatoma in woodchucks. Proc. Natl. Acad. 1685
Sci. U. S. A. 75, 4533–7 (1978). 1686

97
27. Chang, S. F. et al. A new avian hepadnavirus infecting snow geese (Anser 1687
caerulescens) produces a significant fraction of virions containing single-stranded 1688 DNA. Virology 262, 39–54 (1999). 1689
28. Sprengel, R., Kaleta, E. F. & Will, H. Isolation and characterization of a hepatitis B 1690 virus endemic in herons. J. Virol. 62, 3832–9 (1988). 1691
29. Dane, D. S., Cameron, C. H. & Briggs, M. Virus-like particles in serum of patients 1692 with Australia-antigen-associated hepatitis. Lancet 1, 695–8 (1970). 1693
30. McCaul, T. F., Tsiquaye, K. N. & Zuckerman, A. J. Studies by electron microscopy 1694 on the assembly of duck hepatitis B virus in the liver. J. Med. Virol. 16, 77–87 1695
(1985). 1696
31. Ganem, D. & Schneider, R. in Fields Virol. (Knipe, D. M. & Howley, P. M.) 2923–1697
2969 (Lippincott Williams & Wilkins, 2001). 1698
32. Bosch, V., Bartenschlager, R., Radziwill, G. & Schaller, H. The duck hepatitis B 1699
virus P-gene codes for protein strongly associated with the 5’-end of the viral DNA 1700 minus strand. Virology 166, 475–85 (1988). 1701
33. Stannard, L. M. Depiction of the three different particles of the hepatitis B virus. 1702
(1995). at <https://virus.stanford.edu/hepadna/Pictures.html> 1703
34. Bartenschlager, R., Junker-Niepmann, M. & Schaller, H. The P gene product of 1704 hepatitis B virus is required as a structural component for genomic RNA 1705 encapsidation. J. Virol. 64, 5324–32 (1990). 1706
35. Bartenschlager, R. & Schaller, H. Hepadnaviral assembly is initiated by 1707 polymerase binding to the encapsidation signal in the viral RNA genome. EMBO 1708 J. 11, 3413–20 (1992). 1709
36. Weber, M. et al. Hepadnavirus P protein utilizes a tyrosine residue in the TP 1710 domain to prime reverse transcription. J. Virol. 68, 2994–9 (1994). 1711
37. Birnbaum, F. & Nassal, M. Hepatitis B virus nucleocapsid assembly: primary 1712 structure requirements in the core protein. J. Virol. 64, 3319–30 (1990). 1713
38. Gallina, A. et al. A recombinant hepatitis B core antigen polypeptide with the 1714
protamine-like domain deleted self-assembles into capsid particles but fails to 1715 bind nucleic acids. J. Virol. 63, 4645–52 (1989). 1716
39. Schlicht, H. J., Bartenschlager, R. & Schaller, H. The duck hepatitis B virus core 1717 protein contains a highly phosphorylated C terminus that is essential for 1718 replication but not for RNA packaging. J. Virol. 63, 2995–3000 (1989). 1719

98
40. Nassal, M. The arginine-rich domain of the hepatitis B virus core protein is 1720
required for pregenome encapsidation and productive viral positive-strand DNA 1721 synthesis but not for virus assembly. J. Virol. 66, 4107–16 (1992). 1722
41. Chu, T.-H., Liou, A.-T., Su, P.-Y., Wu, H.-N. & Shih, C. Nucleic acid chaperone 1723 activity associated with the arginine-rich domain of human hepatitis B virus core 1724 protein. J. Virol. 88, 2530–43 (2014). 1725
42. Lewellyn, E. B. & Loeb, D. D. Serine phosphoacceptor sites within the core 1726 protein of hepatitis B virus contribute to genome replication pleiotropically. PLoS 1727 One 6, e17202 (2011). 1728
43. Perlman, D. H., Berg, E. a, O’connor, P. B., Costello, C. E. & Hu, J. Reverse 1729 transcription-associated dephosphorylation of hepadnavirus nucleocapsids. Proc. 1730
Natl. Acad. Sci. U. S. A. 102, 9020–5 (2005). 1731
44. Gazina, E. V, Fielding, J. E., Lin, B. & Anderson, D. A. Core protein 1732 phosphorylation modulates pregenomic RNA encapsidation to different extents in 1733 human and duck hepatitis B viruses. J. Virol. 74, 4721–8 (2000). 1734
45. Yu, M. & Summers, J. Multiple functions of capsid protein phosphorylation in duck 1735 hepatitis B virus replication. J. Virol. 68, 4341–8 (1994). 1736
46. Basagoudanavar, S. H., Perlman, D. H. & Hu, J. Regulation of hepadnavirus 1737 reverse transcription by dynamic nucleocapsid phosphorylation. J. Virol. 81, 1738
1641–9 (2007). 1739
47. Wynne, S. A., Crowther, R. A. & Leslie, A. G. The crystal structure of the human 1740 hepatitis B virus capsid. Mol. Cell 3, 771–80 (1999). 1741
48. Kenney, J. M., von Bonsdorff, C. H., Nassal, M. & Fuller, S. D. Evolutionary 1742 conservation in the hepatitis B virus core structure: comparison of human and 1743 duck cores. Structure 3, 1009–19 (1995). 1744
49. Nassal, M. et al. A structural model for duck hepatitis B virus core protein derived 1745 by extensive mutagenesis. J. Virol. 81, 13218–29 (2007). 1746
50. Vorreiter, J. et al. Monoclonal antibodies providing topological information on the 1747
duck hepatitis B virus core protein and avihepadnaviral nucleocapsid structure. J. 1748
Virol. 81, 13230–4 (2007). 1749
51. Grgacic, E. V, Kuhn, C. & Schaller, H. Hepadnavirus envelope topology: insertion 1750 of a loop region in the membrane and role of S in L protein translocation. J. Virol. 1751
74, 2455–8 (2000). 1752

99
52. Grgacic, E. V & Schaller, H. A metastable form of the large envelope protein of 1753
duck hepatitis B virus: low-pH release results in a transition to a hydrophobic, 1754 potentially fusogenic conformation. J. Virol. 74, 5116–22 (2000). 1755
53. Chojnacki, J., Anderson, D. A. & Grgacic, E. V. L. A hydrophobic domain in the 1756 large envelope protein is essential for fusion of duck hepatitis B virus at the late 1757 endosome. J. Virol. 79, 14945–55 (2005). 1758
54. Guo, J. T. & Pugh, J. C. Topology of the large envelope protein of duck hepatitis 1759 B virus suggests a mechanism for membrane translocation during particle 1760 morphogenesis. J. Virol. 71, 1107–14 (1997). 1761
55. Eble, B. E., MacRae, D. R., Lingappa, V. R. & Ganem, D. Multiple topogenic 1762
sequences determine the transmembrane orientation of the hepatitis B surface 1763 antigen. Mol. Cell. Biol. 7, 3591–601 (1987). 1764
56. Taylor, J. M. Virus entry mediated by hepatitis B virus envelope proteins. World J. 1765
Gastroenterol. 19, 6730–4 (2013). 1766
57. Urban, S., Breiner, K. M., Fehler, F., Klingmüller, U. & Schaller, H. Avian hepatitis 1767 B virus infection is initiated by the interaction of a distinct pre-S subdomain with 1768 the cellular receptor gp180. J. Virol. 72, 8089–97 (1998). 1769
58. Urban, S. & Gripon, P. Inhibition of duck hepatitis B virus infection by a 1770 myristoylated pre-S peptide of the large viral surface protein. J. Virol. 76, 1986–90 1771
(2002). 1772
59. Klingmüller, U. & Schaller, H. Hepadnavirus infection requires interaction between 1773 the viral pre-S domain and a specific hepatocellular receptor. J. Virol. 67, 7414–1774
22 (1993). 1775
60. Ponsel, D. & Bruss, V. Mapping of amino acid side chains on the surface of 1776 hepatitis B virus capsids required for envelopment and virion formation. J. Virol. 1777
77, 416–22 (2003). 1778
61. Borel, C., Sunyach, C., Hantz, O., Trepo, C. & Kay, A. Phosphorylation of DHBV 1779 pre-S: identification of the major site of phosphorylation and effects of mutations 1780 on the virus life cycle. Virology 242, 90–8 (1998). 1781
62. Grgacic, E. V, Lin, B., Gazina, E. V, Snooks, M. J. & Anderson, D. a. Normal 1782 phosphorylation of duck hepatitis B virus L protein is dispensable for infectivity. J. 1783 Gen. Virol. 79 ( Pt 11, 2743–51 (1998). 1784
63. Macrae, D. R., Bruss, V. & Ganem, D. Myristylation of a duck hepatitis B virus 1785 envelope protein is essential for infectivity but not for virus assembly. Virology 1786
181, 359–363 (1991). 1787

100
64. Wang, J. C.-Y., Dhason, M. S. & Zlotnick, A. Structural organization of 1788
pregenomic RNA and the carboxy-terminal domain of the capsid protein of 1789 hepatitis B virus. PLoS Pathog. 8, e1002919 (2012). 1790
65. Zlotnick, A., Johnson, J. M., Wingfield, P. W., Stahl, S. J. & Endres, D. A 1791 theoretical model successfully identifies features of hepatitis B virus capsid 1792 assembly. Biochemistry 38, 14644–52 (1999). 1793
66. Zlotnick, A. et al. Dimorphism of hepatitis B virus capsids is strongly influenced by 1794 the C-terminus of the capsid protein. Biochemistry 35, 7412–21 (1996). 1795
67. Bartenschlager, R. & Schaller, H. Mechanisms governing hepadnaviral 1796 nucleocapsid assembly. J. Hepatol. 17 Suppl 3, S15–9 (1993). 1797
68. Wang, G. H. & Seeger, C. The reverse transcriptase of hepatitis B virus acts as a 1798 protein primer for viral DNA synthesis. Cell 71, 663–70 (1992). 1799
69. Zoulim, F. & Seeger, C. Reverse transcription in hepatitis B viruses is primed by a 1800 tyrosine residue of the polymerase. J. Virol. 68, 6–13 (1994). 1801
70. Staprans, S., Loeb, D. D. & Ganem, D. Mutations affecting hepadnavirus plus-1802 strand DNA synthesis dissociate primer cleavage from translocation and reveal 1803 the origin of linear viral DNA. J. Virol. 65, 1255–62 (1991). 1804
71. Wang, G. H. & Seeger, C. Novel mechanism for reverse transcription in hepatitis 1805 B viruses. J. Virol. 67, 6507–12 (1993). 1806
72. Ganem, D., Pollack, J. R. & Tavis, J. Hepatitis B virus reverse transcriptase and 1807 its many roles in hepadnaviral genomic replication. Infect. Agents Dis. 3, 85–93 1808
73. Lien, J. M., Aldrich, C. E. & Mason, W. S. Evidence that a capped 1809 oligoribonucleotide is the primer for duck hepatitis B virus plus-strand DNA 1810 synthesis. J. Virol. 57, 229–36 (1986). 1811
74. Loeb, D. D., Gulya, K. J. & Tian, R. Sequence identity of the terminal 1812
redundancies on the minus-strand DNA template is necessary but not sufficient 1813 for the template switch during hepadnavirus plus-strand DNA synthesis. J. Virol. 1814
71, 152–60 (1997). 1815
75. Huovila, A. P., Eder, A. M. & Fuller, S. D. Hepatitis B surface antigen assembles 1816 in a post-ER, pre-Golgi compartment. J. Cell Biol. 118, 1305–20 (1992). 1817
76. Lambert, C., Döring, T. & Prange, R. Hepatitis B virus maturation is sensitive to 1818 functional inhibition of ESCRT-III, Vps4, and gamma 2-adaptin. J. Virol. 81, 9050–1819
60 (2007). 1820

101
77. Bardens, A., Döring, T., Stieler, J. & Prange, R. Alix regulates egress of hepatitis 1821 B virus naked capsid particles in an ESCRT-independent manner. Cell. Microbiol. 1822
13, 602–19 (2011). 1823
78. Stieler, J. T. & Prange, R. Involvement of ESCRT-II in hepatitis B virus 1824 morphogenesis. PLoS One 9, e91279 (2014). 1825
79. Prange, R. Host factors involved in hepatitis B virus maturation, assembly, and 1826 egress. Med. Microbiol. Immunol. 201, 449–61 (2012). 1827
80. Rost, M. et al. Gamma-adaptin, a novel ubiquitin-interacting adaptor, and Nedd4 1828
ubiquitin ligase control hepatitis B virus maturation. J. Biol. Chem. 281, 29297–1829
308 (2006). 1830
81. Watanabe, T. et al. Involvement of host cellular multivesicular body functions in 1831 hepatitis B virus budding. Proc. Natl. Acad. Sci. U. S. A. 104, 10205–10 (2007). 1832
82. Li, J. et al. Subversion of cellular autophagy machinery by hepatitis B virus for 1833 viral envelopment. J. Virol. 85, 6319–33 (2011). 1834
83. Sir, D. et al. The early autophagic pathway is activated by hepatitis B virus and 1835 required for viral DNA replication. Proc. Natl. Acad. Sci. U. S. A. 107, 4383–8 1836
(2010). 1837
84. Yu, M. & Summers, J. A domain of the hepadnavirus capsid protein is specifically 1838 required for DNA maturation and virus assembly. J. Virol. 65, 2511–7 (1991). 1839
85. Koschel, M., Oed, D., Gerelsaikhan, T., Thomssen, R. & Bruss, V. Hepatitis B 1840 virus core gene mutations which block nucleocapsid envelopment. J. Virol. 74, 1–1841
7 (2000). 1842
86. Summers, J., Smith, P. M., Huang, M. J. & Yu, M. S. Morphogenetic and 1843
regulatory effects of mutations in the envelope proteins of an avian hepadnavirus. 1844 J. Virol. 65, 1310–7 (1991). 1845
87. Bruss, V. A short linear sequence in the pre-S domain of the large hepatitis B 1846 virus envelope protein required for virion formation. J. Virol. 71, 9350–7 (1997). 1847
88. Bruss, V. & Vieluf, K. Functions of the internal pre-S domain of the large surface 1848 protein in hepatitis B virus particle morphogenesis. J. Virol. 69, 6652–7 (1995). 1849
89. Bruss, V. & Thomssen, R. Mapping a region of the large envelope protein 1850 required for hepatitis B virion maturation. J. Virol. 68, 1643–50 (1994). 1851

102
90. Funk, A., Mhamdi, M., Will, H. & Sirma, H. Avian hepatitis B viruses: molecular 1852 and cellular biology, phylogenesis, and host tropism. World J. Gastroenterol. 13, 1853
91–103 (2007). 1854
91. Franke, C., Matschl, U. & Bruns, M. Enzymatic treatment of duck hepatitis B virus: 1855 topology of the surface proteins for virions and noninfectious subviral particles. 1856 Virology 359, 126–36 (2007). 1857
92. Schlicht, H. J., Kuhn, C., Guhr, B., Mattaliano, R. J. & Schaller, H. Biochemical 1858 and immunological characterization of the duck hepatitis B virus envelope 1859 proteins. J. Virol. 61, 2280–5 (1987). 1860
93. Bruns, M., Miska, S., Chassot, S. & Will, H. Enhancement of hepatitis B virus 1861 infection by noninfectious subviral particles. J. Virol. 72, 1462–8 (1998). 1862
94. Lentz, T. B. & Loeb, D. D. Roles of the envelope proteins in the amplification of 1863
covalently closed circular DNA and completion of synthesis of the plus-strand 1864 DNA in hepatitis B virus. J. Virol. 85, 11916–27 (2011). 1865
95. Summers, J., Smith, P. M. & Horwich, a L. Hepadnavirus envelope proteins 1866 regulate covalently closed circular DNA amplification. J. Virol. 64, 2819–24 1867
(1990). 1868
96. Perlman, D. & Hu, J. Duck hepatitis B virus virion secretion requires a double-1869 stranded DNA genome. J. Virol. 77, 2287–94 (2003). 1870
97. Gerelsaikhan, T., Tavis, J. E. & Bruss, V. Hepatitis B virus nucleocapsid 1871 envelopment does not occur without genomic DNA synthesis. J. Virol. 70, 4269–1872
74 (1996). 1873
98. Wei, Y., Tavis, J. E. & Ganem, D. Relationship between viral DNA synthesis and 1874 virion envelopment in hepatitis B viruses. J. Virol. 70, 6455–8 (1996). 1875
99. Schormann, W., Kraft, A., Ponsel, D. & Bruss, V. Hepatitis B virus particle 1876 formation in the absence of pregenomic RNA and reverse transcriptase. J. Virol. 1877
80, 4187–90 (2006). 1878
100. Yuan, T. T., Sahu, G. K., Whitehead, W. E., Greenberg, R. & Shih, C. The 1879
mechanism of an immature secretion phenotype of a highly frequent naturally 1880 occurring missense mutation at codon 97 of human hepatitis B virus core antigen. 1881 J. Virol. 73, 5731–40 (1999). 1882
101. Chua, P. K., Wen, Y. & Shih, C. Coexistence of Two Distinct Secretion Mutations 1883 ( P5T and I97L ) in Hepatitis B Virus Core Produces a Wild-Type Pattern of 1884 Secretion. 77, 7673–7676 (2003). 1885

103
102. Yuan, T. T., Tai, P. C. & Shih, C. Subtype-independent immature secretion and 1886
subtype-dependent replication deficiency of a highly frequent, naturally occurring 1887 mutation of human hepatitis B virus core antigen. J. Virol. 73, 10122–8 (1999). 1888
103. Le Pogam, S., Yuan, T. T., Sahu, G. K., Chatterjee, S. & Shih, C. Low-level 1889 secretion of human hepatitis B virus virions caused by two independent, naturally 1890 occurring mutations (P5T and L60V) in the capsid protein. J. Virol. 74, 9099–105 1891
(2000). 1892
104. Le Pogam, S. & Shih, C. Influence of a putative intermolecular interaction 1893 between core and the pre-S1 domain of the large envelope protein on hepatitis B 1894 virus secretion. J. Virol. 76, 6510–7 (2002). 1895
105. Sprengel, R., Kuhn, C., Will, H. & Schaller, H. Comparative sequence analysis of 1896 duck and human hepatitis B virus genomes. J. Med. Virol. 15, 323–333 (1985). 1897
106. Ishikawa, T. & Ganem, D. The pre-S domain of the large viral envelope protein 1898 determines host range in avian hepatitis B viruses. Proc. Natl. Acad. Sci. U. S. A. 1899
92, 6259–63 (1995). 1900
107. Hirsch, R. C., Loeb, D. D., Pollack, J. R. & Ganem, D. cis-acting sequences 1901 required for encapsidation of duck hepatitis B virus pregenomic RNA. J. Virol. 65, 1902
3309–16 (1991). 1903
108. Kunkel, T. A., Roberts, J. D. & Zakour, R. A. [19] Rapid and efficient site-specific 1904 mutagenesis without phenotypic selection. Methods Enzymol. 154, 367–382 1905
(1987). 1906
109. Mueller-Hill, K. & Loeb, D. D. Previously unsuspected cis-acting sequences for 1907
DNA replication revealed by characterization of a chimeric heron/duck hepatitis B 1908 virus. J. Virol. 70, 8310–7 (1996). 1909
110. Condreay, L. D., Aldrich, C. E., Coates, L., Mason, W. S. & Wu, T. T. Efficient 1910 duck hepatitis B virus production by an avian liver tumor cell line. J. Virol. 64, 1911
3249–58 (1990). 1912
111. Calvert, J. & Summers, J. Two regions of an avian hepadnavirus RNA pregenome 1913 are required in cis for encapsidation. J. Virol. 68, 2084–90 (1994). 1914
112. Graham, F. L. & van der Eb, A. J. A new technique for the assay of infectivity of 1915 human adenovirus 5 DNA. Virology 52, 456–67 (1973). 1916
113. Lewellyn, E. B. & Loeb, D. D. Base pairing between cis-acting sequences 1917 contributes to template switching during plus-strand DNA synthesis in human 1918 hepatitis B virus. J. Virol. 81, 6207–15 (2007). 1919

104
114. Lenhoff, R. J. & Summers, J. Coordinate Regulation. 68, 4565–4571 (1994). 1920
115. Church, G. M. & Gilbert, W. Genomic sequencing. Proc. Natl. Acad. Sci. U. S. A. 1921
81, 1991–5 (1984). 1922
116. Ganem, D. & Varmus, H. E. The molecular biology of the hepatitis B viruses. 1923 Annu. Rev. Biochem. 56, 651–93 (1987). 1924
117. Summers, J. & Mason, W. S. Replication of the genome of a hepatitis B-like virus 1925 by reverse transcription of an RNA intermediate. Cell 29, 403–415 (1982). 1926
118. Gavilanes, F., Gonzalez-Ros, J. M. & Peterson, D. L. Structure of hepatitis B 1927 surface antigen. Characterization of the lipid components and their association 1928 with the viral proteins. J. Biol. Chem. 257, 7770–7 (1982). 1929
119. Tuttleman, J. S., Pourcel, C. & Summers, J. Formation of the pool of covalently 1930 closed circular viral DNA in hepadnavirus-infected cells. Cell 47, 451–460 (1986). 1931
120. Ostapchuk, P., Hearing, P. & Ganem, D. A dramatic shift in the transmembrane 1932 topology of a viral envelope glycoprotein accompanies hepatitis B viral 1933 morphogenesis. EMBO J. 13, 1048–57 (1994). 1934
121. Ning, X. et al. Secretion of genome-free hepatitis B virus--single strand blocking 1935 model for virion morphogenesis of para-retrovirus. PLoS Pathog. 7, e1002255 1936
(2011). 1937
122. Guo, H. et al. Identification and characterization of avihepadnaviruses isolated 1938 from exotic anseriformes maintained in captivity. J. Virol. 79, 2729–42 (2005). 1939
123. Roseman, A. M., Berriman, J. a, Wynne, S. a, Butler, P. J. G. & Crowther, R. A. A 1940 structural model for maturation of the hepatitis B virus core. Proc. Natl. Acad. Sci. 1941 U. S. A. 102, 15821–6 (2005). 1942
124. M., S. N. and N. The neighbor-joining method: A new method for reconstructing 1943 phylogenetic trees. Mol. Biol. Evol. 4, 406–425 (1987). 1944
125. L., Z. E. and P. Evolutionary divergence and convergence in proteins. Edited in 1945 Evolving Genes and Proteins. 97–166 (1965). 1946
126. Tamura, K. et al. MEGA5: molecular evolutionary genetics analysis using 1947
maximum likelihood, evolutionary distance, and maximum parsimony methods. 1948 Mol. Biol. Evol. 28, 2731–9 (2011). 1949
127. Thermet, A. et al. Identification of antigenic regions of duck hepatitis B virus core 1950 protein with antibodies elicited by DNA immunization and chronic infection. J. 1951 Virol. 78, 1945–53 (2004). 1952

105
128. Perlman, D. & Hu, J. Duck Hepatitis B Virus Virion Secretion Requires a Double-1953 Stranded DNA Genome. J. Virol. 77, 2287–2294 (2003). 1954
129. Gazina, E. V, Lin, B., Gallina, A., Milanesi, G. & Anderson, D. a. Intracellular 1955 retention of duck hepatitis B virus large surface protein is independent of preS 1956 topology. Virology 242, 266–78 (1998). 1957
130. Greco, N., Hayes, M. H. & Loeb, D. D. SGHBV Envelope and Capsid Proteins 1958 Independently Contribute to its Ability to Package Capsids Containing ssDNA in 1959 Virions. J. Virol. (2014). doi:10.1128/JVI.01694-14 1960
1961