interaction of xenobiotics with the glucocorticoid hormone ...161635/fulltext01.pdf ·...
TRANSCRIPT

Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science and Technology 716
Interaction of Xenobiotics with the Glucocorticoid Hormone System
in vitro
BY
MARIA JOHANSSON
ACTA UNIVERSITATIS UPSALIENSIS
UPPSALA 2002

Dissertation for the Degree of Doctor of Philosophy in Environmental Toxicology presented at Uppsala University in 2002 Abstract Johansson, M. 2002. Interaction of xenobiotics with the glucocorticoid hormone system in vitro. Acta Universitatis Upsaliensis. Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science and Technology 716. 48 pp. Uppsala. ISBN 91-554-5321-X Persistent environmental pollutants were examined for their interaction with the glucocorticoid hormone system. The focus was placed on interference with the glucocorticoid synthesis and the glucocorticoid-signalling pathway in various in vitro test systems.
Several aryl methyl sulphones competitively inhibited CYP11B1 activity in mouse adrenocortical Y1 cells. The DDT metabolite, 3-methylsulphonyl-2,2’-bis(4-chlorophenyl)-1,1’-dichloroethene (3-MeSO2-DDE) had a higher affinity to the enzyme than the endogenous substrate, 11-deoxycorticosterone. In fact, 3-MeSO2-DDE (Ki 1.6 µM) was almost as potent as the drug metyrapone (Ki 0.8 µM), a well-known inhibitor of the enzyme. 3-MeSO2-DDE inhibited CYP11B1 activity in human adrenocortical H295R carcinoma cells, and at higher concentrations the CYP21 activity. The human H295R cell line seems to be a useful test system for studies of enzyme activities and could be used to screen endocrine disrupting chemicals interfering with the glucocorticoid hormone synthesis.
Several chiral PCB methyl sulphones and the fungicide tolylfluanid proved to be antagonists to the glucocorticoid receptor (GR) in rat hepatoma cells and/or Chinese hamster ovary cells stable transformed with a human GR and a responsive reporter vector. The 4-methylsulphonyl-2,3,6,2’,4’,5’-hexachlorobiphenyl (4-MeSO2-CB149) enantiomers had similar antagonistic effect on the GR. Co-exposure of substances led to additive inhibitory effects on glucocorticoid-regulated protein synthesis in rat hepatoma cells. In general, 4-substituted but not 3-substituted methylsulphonyl-PCBs interacted with the glucocorticoid hormone system.
In the environment, humans and wildlife are constantly exposed to a wide range of chemicals. Considering the effects of these substances via mechanisms of actions described in this thesis, interference of xenobiotics with the glucocorticoid hormone system deserves further attention. In conclusion, environmental pollutants can interact with the glucocorticoid hormone system in vitro, yet the effects of the tested substances on this hormone system remain to be established in vivo.
Keywords: DDT, PCB, tolylfluanid, adrenal, CYP11B1, glucocorticoid receptor, endocrine disrupter. Maria Johansson, Department of Environmental Toxicology, Uppsala University, Norbyvägen 18A, SE-752 36 Uppsala, Sweden Maria Johansson 2002 ISSN 1104-232X ISBN 91-554-5321-X Nykopia Tryck AB, Stockholm

To my family

Papers discussed This thesis is based on the Papers listed below, which will be referred to in the text by their roman numerals (I-V).
I. Johansson, M., Larsson, C., Bergman, Å. and Lund, B-O. 1998. Structure activity relationship for inhibition of CYP11B1-dependent glucocorticoid synthesis in Y1 cells by aryl methyl sulphones. Pharmacology and Toxicology, 83:225-230
II. Johansson, M., Sanderson, J.T. and Lund, B-O. 2002. Effects of 3-MeSO2-DDE and some CYP inhibitors on glucocorticoid steroidogenesis in the H295R human adrenocortical carcinoma cell line. Toxicology in vitro, 16(2):113-21
III. Johansson, M., Nilsson, S. and Lund, B-O. 1998. Interactions between PCB methyl sulphones and the glucocorticoid receptor. Environmental Health Perspectives, 106:769-772
IV. Johansson, M., Johansson, N. and Lund, B-O. Xenobiotics and the glucocorticoid receptor: Additive antagonistic effects on tyrosine aminotransferase activity in rat hepatoma cells. Manuscript
V. Johansson, M., Larsson, C., Peters, N., Ellerichmann, T. and Lund, B-O. Enantiomeric methylsulphonyl-2,2’,4’,5,5’,6-PCB; differences in biliary excretion but similar effects on the glucocorticoid-signalling pathway. Manuscript
Offprints are published with the permission of the journals concerned.

Contents Abbreviations 6 Introduction 7
Background 7 The glucocorticoid homeostasis 8 The adrenal cortex 9 The glucocorticoid receptor 10 Feedback regulation of glucocorticoid synthesis 11 Mechanism of action of glucocorticoids 12 Abnormalities of adrenocortical secretion 12 The present study 14 Aim 14 Comments on some compounds studied 15
Aryl methyl sulphones 15 Tolylfluanid 16 Metyrapone 16 Aminoglutethimide 18 Ketoconazole 18 RU486 18
Comments on some of the methods 19 Results and discussion 23
Effects of xenobiotics on the glucocorticoid hormone synthesis (paper I-II) 23 The H295R cell line as a screening system for EDCs (paper II) 27 Liver tissue slice culture as a screening system for EDCs 29 Effects of xenobiotics on the glucocorticoid-signalling pathway (paper III-V) 32 Cross-reactivity with other steroid hormone systems 35 Environmental implications 35
Concluding remarks 37 Acknowledgements 39 References 40

6
Abbreviations ACTH adrenocorticotrophic hormone ALP alkaline phosphatase AVP arginine vasopressin Bmax maximum binding capacity corticosteroids glucocorticoids; corticosterone and cortisol CRH corticotrophin releasing hormone CYP cytochrome P450 CYP11A P450scc, cholesterol side-chain cleavage enzyme CYP11B1 P450c11, 11ß-hydroxylase CYP17 17α-hydroxylase and 17/20-desmolase activity CYP21 21α-hydroxylase, converts 17-OH-progesterone to 11-
deoxycortisol DDE 2-bis(4-chlorophenyl)-1,1-dichloroethene DDT 2,2-bis(4-chlorophenyl)-1,1,1-trichloroethane EDCs endocrine disrupting chemicals GABA γ-aminobutyric acid; inhibitory neurotransmitter in the
CNS GR glucocorticoid receptor hGR human glucocorticoid receptor mGR mouse glucocorticoid receptor GRE glucocorticoid responsive element HPA axis hypothalamo-pituitary-adrenal axis HPLC high performance liquid chromatography hsp heat shock protein Kd equilibrium dissociation constant Ki inhibition constant Km Michaelis-Menten dissociation constant of the endogenous
substrate MeSO2 methyl sulphonyl MR mineralocorticoid receptor PCB polychlorinated biphenyls RU486 mifepristone; type II antagonist to GR TAT tyrosine aminotransferase For abbreviations of the PCB methyl sulphones see paper I (table 3) or paper III (table 1).

7
Introduction Background Glucocorticoids are energy mobilising catabolic hormones, which work through nuclear receptors present in virtually every cell (Munck et al., 1984; Buckingham et al., 1997; Sapolsky et al., 2000). When an animal is exposed to a stressful situation (cold, starvation, diseases, chemicals etc.), the body responds by increasing the endogenous glucocorticoid levels to promote actions, such as gluconeogenesis and lipolysis. Thus, a well-functioning glucocorticoid homeostasis is essential for survival. However, there are several field reports concerning alterations of the glucocorticoid system in mammals and fish, which show an impairment of coping with stress. The seals of the heavily polluted Baltic Sea suffered from adrenocortical hyperplasia (Bergman and Olsson, 1985), indicating an increased pressure on the adrenal gland. Fish from polluted waters show impaired cortisol stress response (Hontela et al., 1992; Hontela et al., 1995; Hontela et al., 1997). Since high body burdens of persistent chemicals are found in marine mammals (Haraguchi et al., 1992; Bergman et al., 1994; Janák et al., 1998), environmental pollutants may have interfered with the glucocorticoid system. In addition, DDT and corticosterone induced similar pathological development in amphibians, which was interpreted as a potential corticosterone-mimicking action of DDT (Hayes et al., 1997). However, the mechanisms for interference of environmental pollutants with the glucocorticoid homeostasis are still not much studied. Even though endocrine-disrupting chemicals have been defined as ”exogenous agents that interfere with the production, release, transport, metabolism, binding, action, or elimination of the natural hormones in the body responsible for the maintenance of homeostasis and the regulation of developmental processes” (Kavlock et al., 1996), research is mainly focused on xenobiotics interacting with the sex hormones. Many chemicals, including DDT, PCBs, polychlorinated dibenzo-p-dioxins/dibenzofurans, and nonylphenols are known to disturb the endocrine system by acting like oestrogens, antioestrogens or antiandrogens (Kelce et al., 1995; Soto et al., 1995; Navas and Segner, 2000; Gray et al., 2001; Hoekstra et al., 2001; Meerts et al., 2001; Sanderson et al., 2001). Reports concerning effects of environmental pollutants on the glucocorticoid system are scarcer. This thesis deals with the interaction

8
of xenobiotics with the glucocorticoid system. To study these interactions I have chosen different in vitro test systems. The focus is placed on effects on enzyme activities in the glucocorticoid synthesis and regulation of protein synthesis via the glucocorticoid receptor. The glucocorticoid homeostasis Glucocorticoids exert numerous actions in the body and play an essential role in maintaining the basal and stress-induced homeostasis. Depending on species, the most important glucocorticoids are corticosterone and cortisol. At normal conditions, the secretion of glucocorticoids exhibits a diurnal rhythm with a maximal cortisol peak at the end of the sleep phase. Thereafter, the levels are decreasing to a nadir just before or after falling asleep. The glucocorticoid production is under the influence of the hypothalamo-pituitary-adrenal (HPA) axis. Whenever the HPA-axis is activated by a stimulus, it responds by releasing the neuropeptids corticotrophin-releasing hormone (CRH) and arginine vasopressin (AVP) from the hypothalamus. CRH and AVP bind to membrane receptors in the anterior pituitary gland, which results in the release of adrenocorticotropic hormones (ACTH) and related peptides. A long-term treatment with CRH, but not with AVP, stimulates the biosynthesis of ACTH and promotes corticotrophic mitogenesis (Buckingham et al., 1997). ACTH is synthesised from proopiomelanocortin (POMC), which is a pro-hormone to ACTH. Following gene transcription and translation, newly formed pre-POMC is packaged in secretory vesicles together with proteolytic enzymes needed for the generation of ACTH. These polypeptide hormones stimulate the cortex of the adrenal gland to synthesise and release glucocorticoids according to the intensity and duration of the stimulus. To control glucocorticoid synthesis, the release of neuropeptids by the hypothalamus are negatively regulated by feed-back systems, responding to increased levels of glucocorticoids in the blood, input of neurotransmitters like noradrenaline and GABA and, to a lesser extent, increased levels of ACTH.

9
The adrenal cortex The adrenal glands are paired organs located close to the cephalic pole of the kidneys. The adrenals consist of two endocrine organs, the adrenal cortex and the adrenal medulla, which are embryologically and functionally distinct. The medulla produces chatecolamines from tyrosine and the cortex secretes steroid hormones derived from cholesterol. The adrenal cortex consists of three anatomically distinct zones: zona glomerulosa, zona fasciculata and zona reticularis. Zona glomerulosa is mainly regulated by the renin-angiotensin system and is responsible for the synthesis of the mineralocorticoid aldosterone in all mammals. The synthesis of glucocorticoids is regulated by ACTH and occurs in zona fasciculata/reticularis depending on species. A part from glucocorticoids, zona reticularis also synthesises various sex hormones in humans. Glucocorticoids cannot be stored in the adrenal cortex. Therefore, all adrenocortical steroids are instantly synthesised from cholesterol as a response to ACTH stimulation. During basal conditions, the cholesterol used in the steroidogenesis is either extracted from plasma lipoproteins by receptor-mediated endocytosis or endogenously synthesised from acetate. There is also a depot of esterified cholesterol stored in lipid droplets in the adrenal cortex. This storage is the major source of cholesterol, which is used following stimulation of steroidogenesis (Hinson and Raven, 1996). Mainly CYP enzymes participate in the glucocorticoid synthesis (fig. 1). The first and rate-limiting step in steroidogenesis is the mobilisation of cholesterol for subsequent conversion to pregnenolone by the mitochondrial CYP11A (P450scc or 20,22-desmolase). Pregnenolone enters the endoplasmatic reticulum, where the main product of the corticosteroid synthesis is either 11-deoxycorticosterone or 11-deoxycortisol, depending on species. The final step in humans is the conversion of 11-deoxycortisol to cortisol (in mice and rats 11-deoxycorticosterone is converted to corticosterone) by CYP11B1 (11ß-hydroxylase) in the inner membrane of the mitochondria. The glucocorticoids are secreted from the adrenal cortex, transported by a corticosteroid-binding globulin (CBG; transcortin) and distributed by the blood circulation to their target cells.

10
CYP11A CYP17
CYP21CYP21
CYP11B1
CYP17
CYP17
CYP11B1
CYP17
Cholesterol Pregnenolone Progesterone 17α-Hydroxyprogesterone
Androstenedione
11-Deoxycortisol11-Deoxycorticosterone
Cortisol
3βHSD
17α-Hydroxypregnenolone
Dehydroepiandrosterone
3βHSD
3βHSD
Corticosterone
Figure 1. The glucocorticoid synthesis pathway in the human adrenal cortex, focusing on steroids of interest of this study. Steroids detected by HPLC are shown in bold. The glucocorticoid receptor The glucocorticoid receptor (GR) belongs to the nuclear receptor family, a distinct class of transcription factors that are activated by the binding of a signalling molecule, i.e., the ligand. Other members of this gene superfamily are the mineralocorticoid, androgen, oestrogen, thyroid hormone, retinoid and vitamin D3 receptors and a growing number of orphan receptors for which no specific ligands have yet been found (Bamberger et al., 1996). The GR is most often associated with a multiprotein complex containing heat shock proteins, i.e., two molecules of hsp90 and one molecule each of hsp70 and hsp56 (Muller and Renkawitz, 1991; Bamberger et al., 1996). The main function of the multiprotein complex is to keep the GR inactivated. GR is present in virtually every mammalian cell type. The glucocorticoid response of a single cell is dependent on the intracellular availability of the hormone, the GR expression level, and the ability of the cell transduce the hormone signal. The endogenous glucocorticoid is lipophilic and crosses the cellular membrane by diffusion. However, corticosterone uptake in rat hepatocytes may also be mediated by active transport (Spindler et al., 1991). Ligand binding, which is cell cycle-dependent, induces hyperphosphorylation and a conformational change in the GR molecule by the dissociation of the

11
multiprotein complex (Bodwell et al., 1998). This results in exposure of the DNA-binding domain. The DNA-binding domain contains two motifs, zinc fingers, which help the activated receptor to distinguish between the glucocorticoid responsive element (GRE) and other steroid responsive elements. The GR controls transcription by two major mechanisms of action. GR can either dimerise to bind positively and negatively regulated GREs (nGRE) in the promoter region of target genes, or work as a monomer by modulating the activity of other transcription factors such as AP-1 and Stat5, via protein-protein interactions. Unlike most other steroid receptors, GR is refractory to ligand-independent activation of both GRE and nGRE (Wiegel and Zhang, 1998; Turney and Kovacs, 2001). The ligand-dependent activity of GR can be stimulated by activation of the protein kinase A and C pathway (Rangarajan et al., 1992; Nordeen et al., 1994). The GR is thought to be recycled in a process where reassociation with the multiprotein complex demands ATP (Bodwell et al., 1993; Hu et al., 1994). Synergism between GRE and liver-specific transcription factors is necessary for full activity of the tyrosine aminotransferase (TAT) gene in hepatoma cells, where binding sites for e.g. hepatocyte nuclear factor 3 is required (Nitsch et al., 1993; Christoffels et al., 1998). Feedback regulation of glucocorticoid synthesis The feedback regulation of glucocorticoid secretion involves at least three distinct mechanisms: rapid, early delayed, and late delayed feedback (Keller-Wood and Dallman, 1985). The rapid feedback develops within seconds of an increase in glucocorticoids and has a short duration (< 10 min) even though plasma levels remain elevated. The rapid feedback action takes place in the hypothalamus and to some extent in the pituitary gland. This feedback is more sensitive to an alteration of glucocorticoids, than the actual concentration of glucocorticoids in plasma. The mechanisms are thought to be non-genomic, resulting in an inhibition of exocytosis of the neuropeptids (Buckingham et al., 1997). The early delayed feedback develops within hours after an increase in concentration of glucocorticoids. Depending on the intensity of the stimulus, it may persist for up to 24 h. The early delayed feedback is suggested to inhibit the release of ACTH and CRH. It is mediated by the intracellular mineralocorticoid (MR) and glucocorticoid receptors (GR) in the anterior pituitary gland and hypothalamus, but also in the hippocampus and elsewhere

12
in the brain. Glucocorticoids have a higher affinity to the MR (1-2 nM) than to the GR (10-20 nM) (Buckingham et al., 1997). During non-stressed conditions the MR is almost fully occupied by glucocorticoids in the brain. At increased glucocorticoid levels following stress, the GR also becomes occupied. When the glucocorticoids bind to GRs, the HPA axis is enabled to restore the glucocorticoid release to basal levels. The late delayed feedback has a latency of 24 h and is the response of repeated or continuos administration of high doses of corticosteroids. The suppression of the HPA axis may persist for weeks after withdrawal of the steroid treatment and can be life threatening (Mann, 1996; Buckingham et al., 1997). Late delayed feedback is also mediated by MR and GR and inhibits both synthesis and release of CRH/AVP and ACTH. Mechanism of action of glucocorticoids Glucocorticoids exert many metabolic, cardiovascular, anti-inflammatory/immunosupressive and developmental functions in the body. Their catabolic properties promote the breakdown of carbohydrate, protein and lipid. The mobilisation of energy reserves makes glucocorticoids slow but effective antagonists to insulin (Munck et al., 1984; Buckingham et al., 1997). Glucocorticoids have a similar affinity to the MR as aldosterone and may therefore modulate the salt and water balance. An excess of glucocorticoids affects bone and mineral metabolism by decreasing calcium absorption from the gut and increasing renal calcium excretion, which can lead to secondary hyperparathyroidism and osteoporosis (Ziegler and Kasperk, 1998). At normal foetal development, glucocorticoids are responsible for the surfactant secretion in the lungs and the regulation and differentiation of neural crest epithelial cells (Raven and Hinson, 1996). Glucocorticoids modulate neuroendocrine functions with complex effects on mood, behaviour and appetite. Abnormalities of adrenocortical secretion Addison’s disease results from failure of the adrenocortical cells to produce corticosteroids, which is most frequently caused by atrophy. Atrophy may be caused by autoimmunity against adrenocortical cells. Adrenotoxic substances may also give rise to atrophy. Cancer can cause adrenocortical dysfunction. Loss of cortisol secretion reduces the mobilisation of protein and fat from the tissues, thereby depressing many other metabolic functions in the body. A

13
depressed cortisol secretion results in a compensatory elevation of the ACTH levels. In situations of severe stress, however, the adrenal cortex cannot respond to increased demands of cortisol production. This malfunction of the adrenal cortex may lead to death. Symptoms of Addison’s disease are characterised by e.g. muscle weakness, pigmentation, anorexia, and fatigue. Hypersecretion by the adrenal cortex may result in Cushing’s syndrome and is characterised by abnormal cortisol secretion, but also increased androgen secretion can cause effects. Hypercortisolism can result from multiple causes including adenomas in the anterior pituitary or adrenocortical cancer. Cushing’s syndrome may also occur during long-term treatment with glucocorticoids to combat inflammatory diseases such as asthma. Cushing’s syndrome may be characterised by e.g. protein catabolism in peripheral tissues, a suppressed immune system, and osteoporosis.

14
The present study Aims The purpose of this thesis was to shed light on how xenobiotics can interfere with the glucocorticoid homeostasis. As tools, several in vitro systems were used to study some possible mechanisms whereby xenobiotics may interact with the glucocorticoid system. As model compounds, 3-MeSO2-DDE and drugs with well-known and specific mechanisms of action on the glucocorticoid homeostasis were used. Several PCB methyl sulphones were chosen based on their structural similarities with 3-MeSO2-DDE (all these compounds are referred to as aryl methyl sulphones) and prevalence in biota. Some fungicides, e.g. tolylfluanid and penconazole, were chosen based on structural resemblance with the methyl sulphonyl group or the possession of an azole group.
The specific aims were:
• to examine if environmental pollutants affect the glucocorticoid synthesis in mouse Y1 cells and in human H295R cells (paper I, II).
• to examine the response of some drugs with specific inhibitory effects on catalytic CYP enzyme activities in the glucocorticoid synthesis, with the purpose to apply the H295R cell line as a test system for screening potential endocrine disrupters (paper II).
• to examine if environmental pollutants can affect the glucocorticoid-signalling pathway via direct interaction with the human, mouse and rat glucocorticoid receptors (paper III-V).
• to examine if environmental pollutants can exert additive and/or synergistic effects on the glucocorticoid-signalling pathway in a rat hepatoma cell line (paper IV).

15
Comments on some compounds studied The chemical structures of some of the substances are shown in figure 2. Aryl methyl sulphones Methyl sulphones of DDE and PCBs belong to the group of aryl methyl sulphones. These environmental metabolites have a lipophilicity close to that of the parent compounds (Bergman et al., 1994). Because of their chemical stability and resistance to UV radiation, aryl methyl sulphones are as persistent and widespread as their parent compounds (Letcher, 1996). Aryl methyl sulphones were first discovered in the blubber of Baltic grey seals (Jensen and Jansson, 1976). Since then, they have been widely identified in the ecosystem, including fish, birds, mammals, and humans (Letcher et al., 2000). PCB methyl sulphones have been reported to be the third most abundant pollutant in Scandinavian and Canadian biota (Bergman et al., 1994). Aryl methyl sulphones are present in human breast milk, blood and adipose tissue (Newsome and Davies, 1996; Norén et al., 1996; Weistrand and Norén, 1997). Some of the PCB methyl sulphones (e.g. 3- and 4-MeSO2 of CB91, CB132, CB149 and CB174) as well as their parent congeners exist as chiral compounds. In grey seal blubber and in polar bear fat, there is an enantiomeric dominance of one of the atropisomeric congeners of 3-MeSO2-CB149, 4-MeSO2-CB149, 3-MeSO2-CB132, and 4-MeSO2-CB132 (Larsson, 2002; Wiberg et al., 1998). Also human liver samples show excess of one enantiomer of 3-MeSO2-CB132 and 3-MeSO2-CB149 (Ellerichmann et al., 1998). Aryl methyl sulphones are formed in entero-hepatic circulation, which involves both hepatic metabolism via the mercapturic acid pathway and metabolism of the intestinal microflora (Bakke et al., 1982; Brandt et al., 1982). Some PCB methyl sulphones showed an accumulation in lungs, liver, kidney cortex and adipose tissues in human, rat and mouse (Brandt and Bergman, 1981; Haraguchi et al., 1984; Brandt et al., 1985; Klasson-Wehler et al., 1987), whereas 3-MeSO2-DDE is reported to bind irreversibly in the adrenal cortex of mice (Lund et al., 1988). The biological effects are generally not much studied, however, some toxicological features are known. 3-MeSO2-DDE is a potent adrenocorticolytic

16
agent following a local metabolic activation by the adrenocortical CYP11B1 (Jönsson et al., 1992; Lund, 1994; Lund and Lund, 1995). Several 3-MeSO2-PCBs are potent inducers of the hepatic microsomal drug-metabolising enzymes CYP2B1, 2B2, 3A2 and 2C6 (Kato et al., 1995a; Kato et al., 1995b; Kato et al., 1997). Furthermore, 3-MeSO2-PCBs induce the conjugating enzyme UDP-glucuronosyltransferase (Kato et al., 2000b). Methylsulphonyl-PCBs, such as 4-MeSO2-CB52, 4-MeSO2-CB70, 4-MeSO2-CB101 have shown a high affinity to an uteroglobin-like protein residing in pulmonary Clara cells, and are proposed to have caused the respiratory distress of Yusho patients in Japan (Brandt et al., 1985; Lund et al., 1985). An increased progesterone metabolism was seen in minks fed with aryl methyl sulphones (Lund et al., 1999). In the thyroid hormone system, 3-MeSO2-CB132, 3-MeSO2-CB141, 3-MeSO2-CB149 and 4-MeSO2-CB149 reduced serum thyroxin (T4) level and increased serum thyroid stimulating hormone level in rats (Kato et al., 1998b; Kato et al., 1999; Kato et al., 2000a). In addition, 4-MeSO2-CB101 and 4-MeSO2-CB149 inhibited the cell-cell communication in vitro (Kato et al., 1998a). Tolylfluanid Little is reported in the open literature on the toxicity of tolylfluanid. However, this fungicide affects the liver, adrenal and thymus in rat during chronic exposure and the fluoride is incorporated in teeth and bone (Kemikalieinspektionen, 1998). Metyrapone The pyridine derivative metyrapone is a selective inhibitor of CYP11B1 at low concentrations (Tobes et al., 1985). However, at higher concentrations this compound also inhibits CYP11A and most of the phenobarbital-inducible microsomal CYP enzymes (Carballeira et al., 1976; Tobes et al., 1985; Lamberts et al., 1987; Raven and Hinson, 1996). Metyrapone induces the human drug-metabolising CYP3A4 via activation of the pregnane X receptor (Harvey et al., 2000). Metyrapone has been used for evaluation of the function of HPA axis and is still used as a treatment for Cushing’s syndrome and depression (Engelhardt and Weber, 1994; Raven and Hinson, 1996; Mitchell and O'Keane, 1998).

17
O
OH
O
O
OH
OHO
11-deoxycorticosterone corticosterone
Cl Cl
ClCl
SCH3
O
O
Cl
Cl
Cl
Cl
SCH3
OO
3-MeSO2-DDE 4-MeSO2-CB64
Cl
Cl
Cl
ClCl
SCH3
OO
Cl
CH3
NSSN
H3CH3C
O
OCFCl2
4-MeSO2-CB149 tolylfluanid
N
NH2C
O O
ClCl
O N N C 3
OCH
O
NOH
C CH
H3CH3C
ketoconazole RU486
Figure 2. Structures of some compounds studied. The endogenous substrate of CYP11B1, 11-deoxycorticosterone, and the endogenous ligand of GR, corticosterone. The competitive CYP11B1 inhibitors 3-MeSO2-DDE and 4-MeSO2-CB64; the glucocorticoid receptor (GR) antagonists 4-MeSO2-CB149 and tolylfluanid; the drugs ketoconazole (CYP11B1 inhibitor, GR antagonist) and RU486 (mifepristone; GR antagonist).

18
Aminoglutethimide Aminoglutethimide was originally developed as an anticonvulsant, but has also been used as an aromatase inhibitor in breast cancer therapy and in the treatment of ACTH-dependent Cushing’s syndrome (Nicholls et al., 1986). It reversibly inhibits several CYP enzymes in the corticosteroid synthesis such as CYP11A, CYP21 and CYP11B1 (Shaw et al., 1988; Raven and Hinson, 1996). Ketoconazole Ketoconazole is an antimycotic imidazole derivative that is used in the treatment of Cushing’s syndrome (Venkatakrishnan et al., 2000). This compound has endocrinological effects such as suppression of the testosterone and corticosteroids synthesis caused by inhibition of several CYP enzymes (Kan et al., 1985; Nagai et al., 1987; Albertson et al., 1988; Engelhardt and Weber, 1994; Ideyama et al., 1999). Ketoconazole is also a potent inhibitor of hepatic drug-metabolising CYP enzymes (Venkatakrishnan et al., 2000). The azole group is known to co-ordinate its heterocyclic nitrogen with the haem iron of the CYP enzymes, which prevents an activation of molecular oxygen (Feldman, 1986). In addition, Loose and co-workers (Loose et al., 1983) have reported that ketoconazole is a competitive GR antagonist. RU486 RU486 (mifepristone) has several clinical applications including birth control and treatment of Cushing’s syndrome (Sarkar, 2002). This compound is an antagonist both to the glucocorticoid and progesterone receptor (PR), to which it binds with an equally high affinity (Kd≤ 1 nM) (Cadepond et al., 1997). RU486 has a relatively low affinity to the androgen receptor (AR), but no affinity to the MR and the oestrogen receptor (ER). Complexes of GR or PR with RU486 can bind to GRE/PRE but cannot initiate transcription (Nordeen et al., 1995). Hence, RU486 is defined as a type II antagonist.

19
Comments on some of the methods used The glucocorticoid hormone synthesis (paper I, II) The well-established mouse adrenocortical Y1 cell line was used for studies of effects on the CYP11B1 activity (paper I). This cell line has a high and stable activity of CYP11B1, but lacks receptors on the cell surface and can therefore not respond to ACTH. Instead, forskolin is used to stimulate CYP11B1 activity via a direct release of cAMP, which in turn initiates the transcription. The Y1 cell line expresses only two CYP enzymes, CYP11A and CYP11B1, with high activity of the latter enzyme (Es-Souni et al., 1992). The Y1 cell line is a simple cell system that can be used as a, rapid, effective and cheap in vitro model for research on how environmental pollutants affect CYP11B1-dependent formation of corticosterone. The human adrenocortical H295R carcinoma cell line was used for studies of effects of xenobiotics on apparent glucocorticoid CYP enzyme activities (paper II). The presence or absence of ACTH receptors on the surface of these cells have been debated (Rainey et al., 1993; Zenkert et al., 2000), therefore forskolin was used to stimulate enzyme activities. The H295R cells express mRNA of all CYP enzymes that participate in the glucocorticoid synthesis, i.e. CYP11A, CYP11B1, CYP17, and CYP21 (Gazdar et al., 1990; Staels et al., 1993; Rainey et al., 1994). In addition, we examined the response of this cell line to some well-known CYP inhibitors, i.e. ketoconazole, metyrapone and aminoglutethimide, with the purpose to apply this in vitro system to screen potential endocrine disrupting chemicals (EDCs) interfering with the glucocorticoid synthesis. Exposure regimen and steroid analysis (paper I, II) The effects of xenobiotics on glucocorticoid CYP enzyme activities were determined by measuring the ability of the enzyme(s) to transform the substrate(s) to their steroid product (s) (see figure 1 for glucocorticoid synthesis). Y1 and H295R cells were stimulated by forskolin for 24 h to elevate the CYP enzyme activity/ies. Simultaneously, the cells were exposed to the test substance. Three hours before terminating the experiment, the enzyme substrate (20 µM) was added to the medium. However, for studies of CYP11B1 activity in the H295R cell line it was necessary with 24h presence of the endogenous substrate to obtain measurable amounts of cortisol in the medium.

20
An aliquot of the medium was extracted in chloroform:methanol. The steroids were analysed by HPLC (Lichrosorb column, RP18 of 5 µm diameter; 4 mm x 20 cm) at their UV absorbency peak of 241 nm as described in paper I, II (fig. 3). Pregnenolone, which was not detectable with HPLC, was measured with radioimmuno assay (paper II). The protein content of the Y1 and H295R cells was determined according to Lowry et al. (1951). The enzyme activities were expressed as the amount of steroid formed per mg protein and per hour (nmol/mg/h).
Figure 3. HPLC chromatogram showing steroids secreted into the culture medium by human adrenocortical H295R carcinoma cells (1= cortisol, 2= 11-deoxycortisol, 3= androstenedione, 4= 11-deoxycorticosterone, 5= 17α-hydroxyprogesterone, 6= progesterone). Enzyme kinetics (paper I) A Lineweaver-Burk double-reciprocal plot (see paper I, figure 2 and 3) was used to assess the type of inhibition and the inhibition constant (Ki) of CYP11B1 activity in Y1 cells for the most potent aryl methyl sulphones. The assessment was based on the exposure regimen described above. However, five to six different concentrations of the substrate deoxycorticosterone (ranging from 2 to 20 µM) and a constant concentration of the test substance were added to the medium.

21
Competitive binding to the glucocorticoid receptor (paper III, IV) Affinity studies of the test substances to GR were performed on mouse GR (mGR) extracted from liver cytosol. Some aryl methyl sulphones were also tested for affinity to the human GR (hGR), which was extracted from insect cells infected with recombinant baculovirus containing full-length hGR. [3H]-dexamethasone, a specific ligand to the GR, was used as the radioligand. First, the protein content of the mouse liver cytosol was determined using the BCA Protein Assay Reagent. Then, a saturation binding study was performed to find out the receptor density (Bmax; maximum binding capacity), the receptor dissociation constant (Kd; dissociation equilibrium constant for binding of any ligand to the receptor) and, the non-specific binding of [3H]-dexamethasone to other cytosolic macromolecules. The ligand-receptor complex was separated from the unbound radioligand by elution through a Sephadex column and the radioligand-receptor complex was quantified by liquid scintillation. Bmax and protein content were used to calculate the concentration of GR in the cytosol. The obtained Kd value of [3H]-dexamethasone showed that the receptor found in the cytosol was the GR. Following the competitive binding studies, increasing concentrations of xenobiotics were added to a constant concentration of radioligand (10-12 nM [3H]-dexamethasone) and one nanomole of GR. The ligand-receptor complex was separated and analysed as described above. The binding of a test substance to the GR was indicated by decreasing amounts of [3H]-dexamethasone bound to the receptor at increasing concentration of test substance. The glucocorticoid-signalling pathway (paper III-V) Glucocorticoid signal transduction includes receptor activation through ligand binding, configuration of the receptor, translocation to the nucleus, binding to the DNA (i.e. GRE), and induction of protein synthesis, e.g. tyrosine aminotransferase (TAT). To determine the nature and type of binding of xenobiotics to GR, a GRAF reporter cell line (paper III) and the Reuber rat hepatoma cell line (H4-II-E-C3) cell line (paper IV, V) were used. The GRAF cell line is generated by stable transformation of Chinese Hamster Ovary cells and has a human placental alkaline phosphatase (ALP) reporter element in connection to the glucocorticoid responsive element (GRE). Whenever the human GR (hGR) is activated by a ligand, the ligand-hGR complex binds to the GRE, i.e. an agonistic response is achieved, and placental

22
ALP reporter proteins are expressed and secreted from the cells into the medium. The ALP in the medium was analysed by a chemiluminescence assay and measured in a luminometer. The ALP activity was expressed in light units (LU), which are directly proportional to the amount of ALP secreted from the cells. The Reuber rat hepatoma cell line (H4-II-E-C3) contains a physiologically relevant cytosolic enzyme, TAT, the transcription of which is regulated by glucocorticoids. Dexamethasone was used to stimulate TAT synthesis. The cells were harvested and sonicated to release the TAT enzyme. The TAT specific activity was analysed in an enzyme assay and measured in a spectrophotometer (Diamondstone, 1966; Heiman and Tonner, 1995). The TAT specific activity is directly proportional to the formation of 4-hydroxyphenylpyruvate at 37 °C and expressed as nmol/mg protein/min. TAT assay (paper IV, V) The cell sonicate containing TAT was added to phosphate buffer (0.2 M; pH 7.3) containing 6.85 mM tyrosine and 1.33 mM pyridoxal-5’-phosphate. The reaction was started by addition of 0.3 M α-ketoglutarate. The reaction was stopped by addition of 10 M NaOH and read spectrophotometrically at 331 nm against a zero time blank prepared by adding the NaOH to the reaction components before addition of cell sonicate (Diamondstone, 1966; Heiman and Tonner, 1995). A fluorescence-based protein assay was applied to analyse the protein content, using bovine serum albumin as a standard (Lorenzen and Kennedy, 1993). TAT in mouse liver slices (only in thesis-summary) Female mice were sacrificed by cervical dislocation. The livers were rapidly excised and kept in cold PBS until embedded in 3% agarose. Precision-cut slices (200 µm) were prepared in a Krumdieck tissue slicer (Alabama Research and Development, Munford, Alabama, USA) and cultured according to Lindhe et al. (2001). The liver slices were exposed to different concentrations of either dexamethasone or corticosterone in order to stimulate the TAT activity. The incubation time ranged from 5-48h. After exposure, the slices were removed from the medium and rinsed three times in cold PBS. The slices were transferred to a test tube containing PBS and sonicated. An aliquot of the cell-

23
sonicate was taken for TAT assay and analysis of the protein content as described above. Toxicity assays (paper I-V) Toxicity was assessed by microscopic evaluation of cell morphology (paper I-V), protein content (Lowry et al., 1951; paper I-V) and mitochondrial damage (MTT, Mosmann, 1983; paper III). Results and Discussion Effects of xenobiotics on the glucocorticoid hormone synthesis 3-MeSO2-DDE 3-MeSO2-DDE was found to inhibit the adrenocortical mitochondrial CYP11B1 activity in mouse Y1 and human H295R cells, and thus, to decrease the synthesis of glucocorticoids (paper I, II). In paper I, 3-MeSO2-DDE was tested for type of inhibition, where double-reciprocal plots showed that this substance exerted competitive inhibition of CYP11B1. To exclude the possibility that toxicity or effects on enzyme synthesis affected corticosterone formation at 24h of exposure, the enzyme activity was measured also at a shorter exposure time (3 h). In all cases, 3-MeSO2-DDE displayed a competitive inhibition of CYP11B1 with an apparent Ki of 1.6 µM. This Ki value is significantly lower than the apparent Km for the endogenous substrate, 11-deoxycorticosterone (Km 3.5 µM). In mouse Y1 cells, 3-MeSO2-DDE almost completely inhibited the corticosterone secretion (paper I). A similar decrease in corticosterone secretion by 3-MeSO2-DDE has been reported in mouse adrenal tissue slices (Lindhe et al., 2001). Mice given an injection of 3-MeSO2-DDE, and suckling pups exposed to 3-MeSO2-DDE via the milk, had decreased ACTH-induced plasma corticosterone levels (Jönsson, 1994). These decreased corticosterone levels were long-lasting. Taken together, 3-MeSO2-DDE decreased the corticosteroid production both in vitro, ex vivo and in vivo in mice.

24
3-MeSO2-DDE inhibited the CYP11B1 activity in human H295R cells in a dose-dependent manner with a potency only somewhat lower than that reported in mouse Y1 cells (paper II; Lund and Lund, 1995). In human H295R cells, the highest concentration of 3-MeSO2-DDE (10 µM) decreased the cortisol formation to 66% of control (paper II). In contrast, a 2.5-fold higher concentration of 3-MeSO2-DDE did not affect cortisol formation in human adrenal tissue slices, although an accumulation of other steroids such as 11-deoxycortisol, 11-deoxycorticosterone and androstenedione was seen in the medium (Lindhe et al., 2002). An accumulation of CYP11B1 precursors gives an indication of a CYP11B1 inhibition, yet the accumulation of androstenedione is puzzling. Increased androstenedione levels may be caused by either an enzymatic conversion of 11-deoxycortisol to androstenedione (Brown and Fishman, 2000), or an inhibition of CYP21 activity. In H295R cells, 3-MeSO2-DDE displayed inhibitory effects also on CYP21 activity albeit at higher concentrations than those needed for inhibition of CYP11B1 (fig. 4). This indicates that the increased androstenedione levels may be a result of CYP21 inhibition. The effects of 3-MeSO2-DDE on androstenedione production merits further attention. In the Y1 cells, the inhibition of CYP11B1 activity was correlated with a lowered cellular protein content at a concentration of 10 µM 3-MeSO2-DDE, as compared to controls (Lund and Lund, 1995). As stated above, it is not likely that the inhibition of CYP11B1 activity was caused by toxicity. Rather, the toxicity seemed to be caused by CYP11B1-dependent activation of 3-MeSO2-DDE. A corresponding toxicity was not seen in human H295R cells (paper II). This finding does not necessarily imply species-differences in adrenocorticolytical effects of 3-MeSO2-DDE. On the contrary, 3-MeSO2-DDE has been reported to be bioactivated and irreversibly bound to the specific zones in the adrenal cortex that express CYP11B1 in both humans and mice ( Lund et al., 1988; Jönsson and Lund, 1994; Lindhe, 2001; Lindhe et al., 2002). In mice, this binding was followed by mitochondrial damage at lower doses, whereas higher doses necrotised the zona fasciculata (Lund et al., 1988; Jönsson et al., 1991). Further, the irreversible binding and adrenocorticolytic activity of MeSO2-DDE was reported to be dependent on the concentration of the ACTH-regulated enzyme CYP11B1 in mice (Lindhe, 2001). This suggests that the lack of CYP11B1-mediated toxicity by 3-MeSO2-DDE in human H295R cells was due to low catalytic enzyme activity.

25
-7 -6 -5 -40
20
40
60
80
100
120
concentration of3-MeSO2-DDE
(log M)
% o
f con
trol
Figure 4. Effects of 3-MeSO2-DDE on CYP21 activity as measured by the conversion of progesterone to deoxycorticosterone in human H295R cells. 10-7 in the graph represents absence of 3-MeSO2-DDE. There were no signs of toxicity at the concentrations used. Each point represents mean ± standard deviation of quadruplicates. PCB methyl sulphones Several PCB methyl sulphones decreased corticosterone formation to varying degrees in mouse Y1 cells (paper I). The two most potent methylsulphonyl-PCBs, 4-MeSO2-2,3,6,4'-tetrachlorobiphenyl (4-MeSO2-CB64) and 4-MeSO2-2,3,6,3’,4’-pentachlorobiphenyl (4-MeSO2-CB110) in the screening test were also tested for type of inhibition. Double-reciprocal plots revealed that these PCB methyl sulphones competitively inhibited CYP11B1, with apparent Ki values of 4.6 and 6.7 µM for 4-MeSO2-CB64 and 4-MeSO2-CB110, respectively. Several MeSO2-PCBs (20 µM) decreased the cellular protein content following a 24 h exposure period. In contrast to what was stated in paper I, additional statistical linear regression analysis revealed a correlation between a lowered protein content in the cells and the degree of enzyme inhibition caused by MeSO2-PCBs (fig. 5). However, it seems unlikely that the inhibitory effects of MeSO2-PCBs in mouse Y1 cells reflected a toxic response. This interpretation is supported by the fact that two of the most potent methyl sulphones (10 µM) exhibited competitive enzyme kinetics after 24h of exposure. In addition, 3-MeSO2-DDE displayed the same Ki independent of concentration or time of exposure, which further supports that the inhibition of CYP11B1 activity was
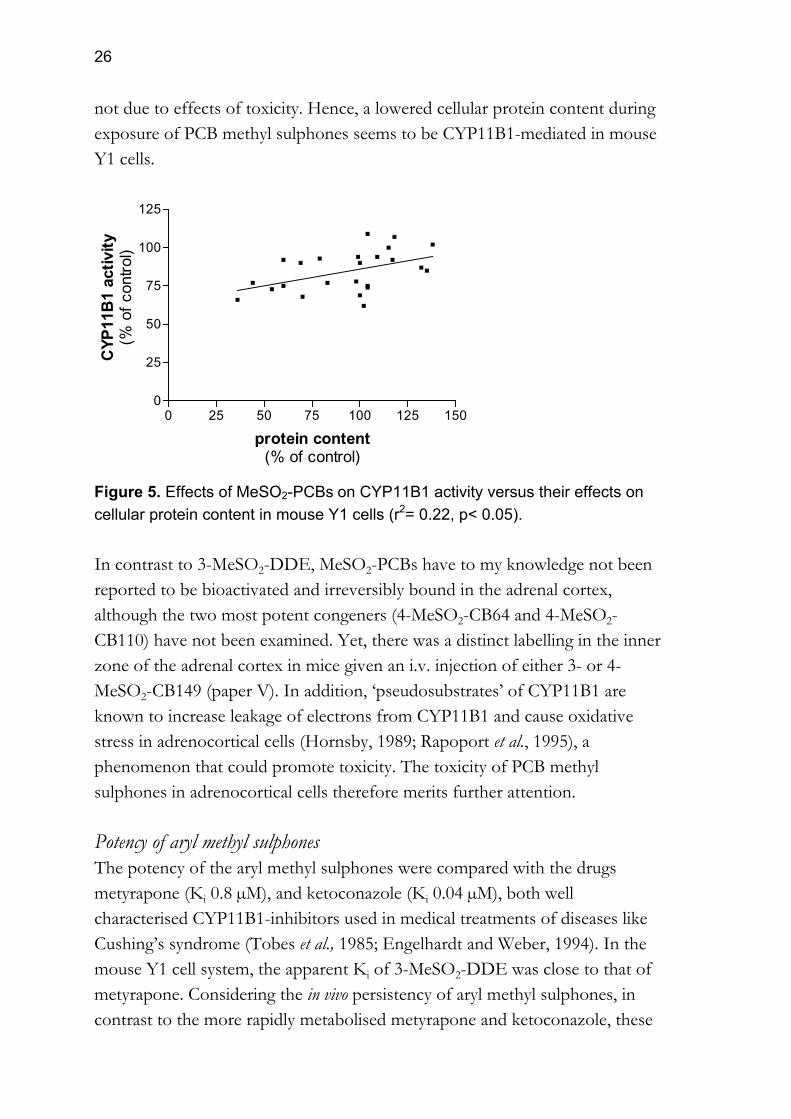
26
not due to effects of toxicity. Hence, a lowered cellular protein content during exposure of PCB methyl sulphones seems to be CYP11B1-mediated in mouse Y1 cells.
0 25 50 75 100 125 1500
25
50
75
100
125
protein content(% of control)
CYP
11B
1 ac
tivity
(% o
f con
trol)
Figure 5. Effects of MeSO2-PCBs on CYP11B1 activity versus their effects on cellular protein content in mouse Y1 cells (r2= 0.22, p< 0.05). In contrast to 3-MeSO2-DDE, MeSO2-PCBs have to my knowledge not been reported to be bioactivated and irreversibly bound in the adrenal cortex, although the two most potent congeners (4-MeSO2-CB64 and 4-MeSO2-CB110) have not been examined. Yet, there was a distinct labelling in the inner zone of the adrenal cortex in mice given an i.v. injection of either 3- or 4-MeSO2-CB149 (paper V). In addition, ‘pseudosubstrates’ of CYP11B1 are known to increase leakage of electrons from CYP11B1 and cause oxidative stress in adrenocortical cells (Hornsby, 1989; Rapoport et al., 1995), a phenomenon that could promote toxicity. The toxicity of PCB methyl sulphones in adrenocortical cells therefore merits further attention. Potency of aryl methyl sulphones The potency of the aryl methyl sulphones were compared with the drugs metyrapone (Ki 0.8 µM), and ketoconazole (Ki 0.04 µM), both well characterised CYP11B1-inhibitors used in medical treatments of diseases like Cushing’s syndrome (Tobes et al., 1985; Engelhardt and Weber, 1994). In the mouse Y1 cell system, the apparent Ki of 3-MeSO2-DDE was close to that of metyrapone. Considering the in vivo persistency of aryl methyl sulphones, in contrast to the more rapidly metabolised metyrapone and ketoconazole, these

27
environmental pollutants may pose a threat to the glucocorticoid system in heavily exposed populations. Structure-Activity Relationships Several environmental pollutants such as DDT metabolites, Clophen A50, polyaromatic hydrocarbons, and polybrominated diphenylethers, and some prototype PCBs were examined for effects on the CYP11B1 activity in mouse Y1 cells (paper I). p,p’-DDT and o,p’-DDD seemed to increase the conversion of DOC to corticosterone, whereas none of the other compounds inhibited the CYP11B1 activity. However, the increased corticosterone formation was correlated with a decreased formation of another metabolite, presumed to be 20α-deoxycorticosterone, leaving higher amounts of substrate for CYP11B1 conversion. The aryl methyl sulphones are novel CYP11B1 inhibitors that do not possess a heterocyclic nitrogen, such as the azole compounds. A methyl group next to a sulphonyl or carbonyl group is present in aryl methyl sulphones and in ketoconazole. These findings suggest that CYP11B1 contains an active site that attracts such a structural feature. However, the screening study of the PCB methyl sulphones showed that also the position of the methyl sulphone moiety was important. A methyl sulphone group in 4-position relative to 3-position increased the inhibitory effect on CYP11B1 activity considerably. In addition, a chlorine in 4’-position, and two ortho-chlorines in 2,6-position seemed optimal for the inhibitory activity. H295R cell line as a screening system for EDCs There is a lack of in vitro test systems to screen suspected endocrine disrupting chemicals and pharmaceuticals for effects on the corticosteroid synthesis. In paper II, the human adrenocortical H295R carcinoma cell line was evaluated as a test system for screening potential glucocorticoid disrupting substances. In order to do so, some well-characterised CYP inhibitors (ketoconazole, metyrapone, and aminoglutethimide) were examined for their effects on catalytic CYP enzyme activities in the hormone synthesis.

28
In H295R cells, ketoconazole inhibited all examined CYP enzyme activities (CYP11A, CYP11B1, CYP17, and CYP21), whereas metyrapone inhibited the mitochondrial CYP11A and CYP11B1 enzyme activities. Both compounds inhibited CYP11B1 with a similar and high potency. Aminoglutethimide was only evaluated for effects on CYP11A activity and was found to inhibit this enzyme with a low potency. Taken together, the overall inhibition profile of these drugs on the CYP enzyme activities in the human H295R carcinoma cell line resembled that previously described in the literature (Carballeira et al., 1976; Tobes et al., 1985; Lamberts et al., 1987; Albertson et al., 1988; Ayub and Levell, 1989; Engelhardt and Weber, 1994; Raven and Hinson, 1996; Ideyama et al., 1999). The human adrenocortical H295R cell line has a functioning protein kinase A signalling pathway, expresses mRNA and catalytic enzyme activities of all enzymes participating in the glucocorticoid synthesis pathway (fig. 1 for glucocorticoid synthesis; Gazdar et al., 1990; Rainey et al., 1993; Staels et al., 1993; Rainey et al., 1994). Furthermore, this cell line has been used for studies of chemical interference with CYP19 (aromatase) expression and catalytic activity (Sanderson et al., 2000). Thus, the H295R cell line may provide a test system for studies of altered steroid hormone production caused by chemicals interfering with a single enzyme activity or with the transcription and expression of enzymes. A change in a single enzyme activity can cause different types and relative proportions of steroids to be formed. In addition, these cells secrete low concentrations of endogenous steroids, which enable studies of specific enzymes by addition of substrate for that particular enzyme. However, the high activities of the microsomal CYP17 and CYP21 in H295R cells call for short exposure times to keep a linear conversion rate to the steroid product. One should also be aware of the difficulties in studying the CYP17 activity in whole cell systems, since in addition to the rapid conversion to the steroid product, there is further metabolism of this product by other CYP enzymes such as CYP21. Therefore, the CYP17 activity in H295R cells was referred to as apparent CYP17 activity (paper II). The steroid secretion pattern of the H295R carcinoma cells resembled that of NCI-H295 cells, where the mitochondrial enzyme activities of CYP11A and CYP11B1 were low (Gazdar et al., 1990). These low activities are in accordance with reports of patients with adrenocortical carcinomas and a patient with non-functional adrenocortical hyperplasia, which also showed very low mitochondrial enzyme activities (Brown and Fishman, 2000; Lindhe et al., 2002). In fact, studies of effects

29
on CYP11B1 activity in H295R cells required an optimised system to get detectable amounts of cortisol, which discourage studies of enzyme kinetics such as the one performed in paper I. However, Rainey and co-workers (1993) reported that forskolin-treated H295R cells primarily secreted cortisol. Thus, there are differences in mitochondrial enzyme activities between at least some clones of H295R cells and normal adrenal cells, although the presence of all CYP enzymes in H295R cells still makes this cell line a suitable tool for studying effects of EDCs. Liver tissue slice culture as a screening system for EDCs Liver tissue slice culture was examined as an ex vivo system to screen substances that may affect the glucocorticoid-signalling pathway. The aim was to set up a test system resembling a normal organ, which enables studies of not only experimental animal but also wildlife tissue.
cont
rol
dexa
met
haso
ne
corti
cost
eron
e0
5
10
150% FCS2% FCS
TAT
activ
ity(µ
mol
/mg/
min
)
Figure 6. Induction of TAT activity in mouse liver tissue slices after a 24h incubation with either dexamethasone (50 µM) or corticosterone (50 µM) in the absence or presence of foetal calf serum (FCS; 2%) in the medium. Each bar represents mean ± standard deviation of triplicate samples.
When removing the liver, it can be assumed that a homogenous and fairly high level of TAT is present in the tissue slice. As shown in figure 6, a 24h-incubation with a high concentration of either corticosterone or

30
dexamethasone induced TAT activity in liver slices from mice. Serum-containing medium seemed to increase both basal and induced TAT activity, indicating an influence of glucocorticoids present in the serum. Hence, the following studies were performed in the absence of serum in the medium. After a 24h exposure, there was a considerable elevation of TAT activity in the presence of either 10 µM of dexamethasone or corticosterone (fig. 7). This could be compared with a concentration of 1-5 nM of dexamethasone needed for a similar induction in H4-II-E-C3 cells and GRAF cells (paper III-V).
-8 -7 -6 -5 -40
5
10
15
20
dexamethasonecorticosterone
concentration of xenobiotic (log M)
TAT
activ
ity(µ
mol
/mg/
min
)
Figure 7. Effects of dexamethasone and corticosterone on TAT induction. 10-8M in the graph represents absence of dexamethasone and corticosterone. Each point represents mean ± standard deviation of triplicate samples.
0 10 20 30 40 500
10
20
30controldexamethasone
time (h)
TAT
activ
ity(µ
mol
/mg/
min
)
Figure 8. TAT activity in liver tissue slices at different times of incubation. Each point represents mean ± standard deviation of triplicate samples.

31
The glucocorticoid-induced TAT activity was examined in liver tissue slice cultures with or without dexamethasone (10 µM). The result show that the TAT activity decreased with time in both control slices and slices treated with dexamethasone (fig. 8). In fact, directly sonicated liver slices that were not put into tissue culture expressed a higher activity than liver slices in culture at any time point. The contribution of the central part of the liver slice is of importance since the whole liver slice was analysed for TAT activity. A decreased TAT activity over time may therefore indicate that the diffusion of dexamethasone into the central parts of the liver slice was low and/or that there was progressive degeneration of the inner parts. However, no viability parameters were used during the experiment. Hart and co-workers (1983) showed that the cell nuclei stained paler in the one third of adult rat liver slices (300 µm) that were not in contact with the gas phase after 24 h of culturing. However, that experimental design is not directly comparable with the design used in the present study. In either case, the findings suggest that only the peripheral layers of the liver slice contributed to the observed glucocorticoid-mediated induction of TAT activity. Possibly the diffusion of dexamethasone into the slice can be described as a concentration gradient of dexamethasone. Thus, higher concentrations of dexamethasone will lead to higher cellular concentrations and deeper layers of the slice will be reached by dexamethasone. This may explain the higher concentration of dexamethasone needed for TAT induction in rodent liver slice cultures as compared to primary rat hepatocytes and rat hepatoma cells (Gayda and Pariza, 1983; Sultan et al., 1996; Niimi et al., 1998; paper III-V). However, it has been reported that a simultaneous presence of cortisol and dBcAMP amplifies the induction of TAT in rat liver tissue slices (Hart et al., 1983), due to stimulation of GR action by the protein kinase A (PKA) pathway. Further studies are needed to examine if the sensitivity of slices can be increased by a simultaneous use of a PKA stimulator and dexamethasone. Taken together, mouse liver tissue slice culture seems too insensitive to dexamethasone to serve as a useful test system for detection of chemicals affecting the glucocorticoid signal transduction. As a result of this finding, the designed GRAF cell line and the Reuber rat hepatoma (H4-II-E-C3) cell line were taken into use.

32
Effects of xenobiotics on the glucocorticoid-signalling pathway Several test substances were examined for their affinity to mGR and hGR by competition with [3H]-dexamethasone for binding to the receptor (paper III, IV). Only PCB methyl sulphones with three ortho chlorines had affinity to mGR. A methyl sulphone group in 4-position gave a greater affinity than one in 3-position. In addition, tolylfluanid, penconazole and ketoconazole bound to the mGR, whereas 3-MeSO2-DDE and triadimefon failed to do so (paper IV). As a comparison, the affinity of RU486 was 1000-fold higher than that of 3-MeSO2-CB149 (IC50 1 µM) to hGR. Considering the in vivo persistency of methyl sulphones compared to the rapidly metabolised RU486 (t1/2= 20 h in man) (Chrousos et al., 1988), one may assume a smaller difference in biological potency in vivo than that seen in vitro. To assess the biological significance of GR affinity, the test substances were examined for their effects in GRAF and Reuber rat hepatoma cells (paper III-IV). In GRAF cells, 3-MeSO2-CB149 appeared to compete with the endogenous glucocorticoid in an antagonistic manner, similarly to that of RU486 (paper III), whereas 3-MeSO2-CB101 (possessing only two ortho chlorines, instead of three required for GR affinity) and 3-MeSO2-DDE were inactive. This is in agreement with the affinity studies with mGR (see above). In contrast, 3-MeSO2-CB149 lacked antagonistic effect in Reuber rat hepatoma cells (paper IV). Whether this difference is a result of species-specific hormone-binding domains of GR or of a different sensitivity of the GRE promoter sequences in the designed GRAF cell line as compared to that of the rat hepatoma cell line remains unknown. In the Reuber rat hepatoma cell line, 4-substituted MeSO2-PCBs displayed antagonistic effects on dexamethasone-induced TAT activity, whereas 3-substituted PCB methyl sulphones, with the exception of 3-MeSO2-CB174, were inactive (paper IV). Furthermore, all of the PCB methyl sulphones that bound to GR are chiral congeners. The biological activity of one enantiomer could be very different from its mirror image. Since most biochemical processes are stereospecific, 4-MeSO2-CB149 enantiomers were studied for antagonistic effects in Reuber rat hepatoma cells (paper V). The enantiomers had antagonistic effects on the glucocorticoid-signalling pathway in rat

33
hepatoma cells. This is in accordance with the behaviour of racemic 4-MeSO2-CB149 (1:1 enantiomeric ratio; paper IV). The concentrations needed for inhibitory effects (IC50) were almost the same for the enantiomers and the racemate of 4-MeSO2-CB149, although there was a 2-fold difference in IC100 (paper V). In mammals there is a high enantiomeric dominance of one enantiomer (Ellerichmann et al., 1998; Wiberg et al., 1998; Larsson, 2002; Larsson et al., 2002), which was proposed to be due to an enantioselective body clearance (paper V). Taken together, the antagonistic effect on the GR seems to be independent of enantiomeric stereochemistry. Therefore, the enantiomer of 4-MeSO2-CB149 found in biota seems to be biologically active in vitro. The fungicide tolylfluanid exerted an inhibitory effect on TAT specific activity in Reuber rat hepatoma cells (paper IV). The antifungal properties of tolylfluanid are believed to be mediated by inhibition of certain thiol-containing enzymes, thereby affecting the formation of amino acid in the fungus. The rat liver GR contains several thiol groups in the hormone-binding domain (Grippo et al., 1985; Miesfeld et al., 1986; Simons et al., 1987), which tolylfluanid may affect. In addition, the dimethyl-N-tolylsulphide structure of tolylfluanid resembles that of a methyl sulphone group, suggesting that this structure may affect the affinity to the hormone-binding domain of the receptor. This fungicide is known to affect several organs during sub-chronic and chronic exposure, among them the adrenal in the rat (Kemikalieinspektionen, 1998). Considering the antagonistic properties of tolylfluanid to GR, and the chronic adrenal effects observed, this implies that this substance may affect more than one function in the glucocorticoid system. Wildlife and man is constantly exposed to numerous chemicals in the environment. Therefore, it is of great importance to study the consequences of simultaneous exposure to chemicals affecting the glucocorticoid-signalling pathway. Two PCB methyl sulphones and the fungicide tolylfluanid were incubated together at various combinations of inhibitory concentrations corresponding to their individual IC10, IC50 and IC90 values in Reuber rat hepatoma cells (paper IV). The results showed that the antagonistic effects on glucocorticoid-regulated gene transcription in the cells were close to additive (fig. 9).
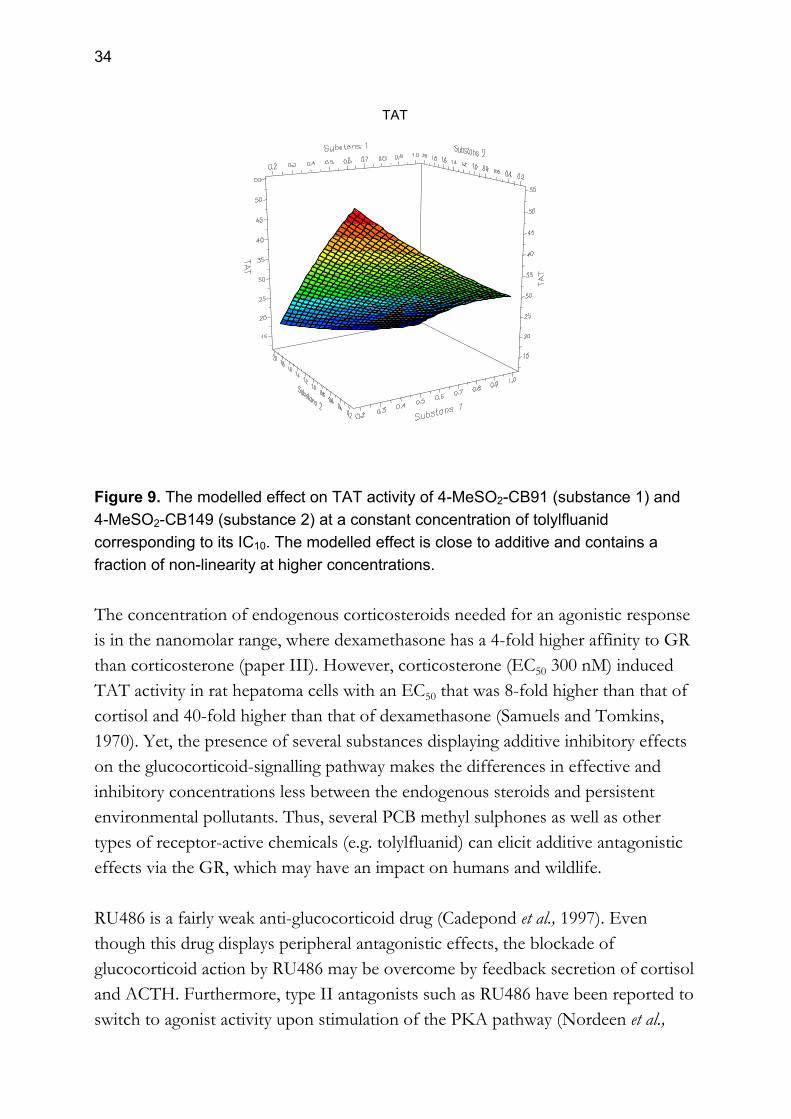
34
TAT
Figure 9. The modelled effect on TAT activity of 4-MeSO2-CB91 (substance 1) and 4-MeSO2-CB149 (substance 2) at a constant concentration of tolylfluanid corresponding to its IC10. The modelled effect is close to additive and contains a fraction of non-linearity at higher concentrations. The concentration of endogenous corticosteroids needed for an agonistic response is in the nanomolar range, where dexamethasone has a 4-fold higher affinity to GR than corticosterone (paper III). However, corticosterone (EC50 300 nM) induced TAT activity in rat hepatoma cells with an EC50 that was 8-fold higher than that of cortisol and 40-fold higher than that of dexamethasone (Samuels and Tomkins, 1970). Yet, the presence of several substances displaying additive inhibitory effects on the glucocorticoid-signalling pathway makes the differences in effective and inhibitory concentrations less between the endogenous steroids and persistent environmental pollutants. Thus, several PCB methyl sulphones as well as other types of receptor-active chemicals (e.g. tolylfluanid) can elicit additive antagonistic effects via the GR, which may have an impact on humans and wildlife. RU486 is a fairly weak anti-glucocorticoid drug (Cadepond et al., 1997). Even though this drug displays peripheral antagonistic effects, the blockade of glucocorticoid action by RU486 may be overcome by feedback secretion of cortisol and ACTH. Furthermore, type II antagonists such as RU486 have been reported to switch to agonist activity upon stimulation of the PKA pathway (Nordeen et al.,

35
1995), which may explain the absence of cortisol deficiency during treatment. In contrast, type I antagonists (that fail to promote dissociation of the multiprotein complex) still act as antagonists upon activation of PKA. Therefore, further studies are needed to assess if the PCB methyl sulphones and tolylfluanid are type I or type II antagonists, and to better understand the consequences of their possible antagonism on GR in vivo. Cross-reactivity with other steroid hormone systems All steroids are derived from cholesterol, which explains their structural similarity. Cross binding of steroids to enzymes and nuclear receptors are likely to occur, as seen with the endogenous glucocorticoid, cortisol, which has affinity to both the MR and the GR in vivo (Buckingham et al., 1997). The drugs RU486, and ketoconazole affect other steroid systems than the glucocorticoid system (Feldman, 1986; Cadepond et al., 1997). In a recent study, designed quinoline derivatives with an O-methyl group displayed affinity to both the GR and the PR (Coghlan et al., 2001). In addition, high concentrations of some 4-substituted PCB methyl sulphones were reported to accumulate in pulmonary Clara cells as a cause of their high affinity to an uteroglobin-like protein (Brandt and Bergman, 1981; Brandt et al., 1985; Lund et al., 1985; Klasson-Wehler et al., 1987). Considering that the endogenous ligand to this protein is proposed to be progesterone, this may imply a structural resemblance of MeSO2-PCBs also with progesterone. These examples of cross-reactivity suggest that a substance that interacts with one hormonal system may affect also other endocrine systems. Environmental implications An inhibition of CYP11B1 activity will decrease the formation and release of glucocorticoids in the blood circulation and thus, due to the feedback system of the HPA axis, increase the secretion of ACTH from the pituitary to restore the glucocorticoid levels. In addition, an antagonistic block of the GR results in an inhibition of glucocorticoid gene transcription. Since the feedback system in most cases works through the GR (Buckingham et al., 1997), a rise in ACTH levels will increase the pressure on the adrenal cortex to produce more

36
glucocorticoids and restore the function. Thus, one may speculate that a disruption of these two functions may cause adrenocortical hyperplasia. This condition was reported in Baltic seals, where the influence of PCBs and other organochlorines were proposed to explain the disease complex (Bergman and Olsson, 1985). Grey seals have been reported to bioactivate 3-MeSO2-DDE to a lesser degree, whereas several species such as mink and otter, are insensitive to this metabolite (Jönsson et al., 1993; Lund, 1994). Technical PCB (Aroclor 1254) has been reported to elevate basal levels of cortisol in seal adrenal tissue slices (Freeman and Sangalang, 1977). Similarly, Aroclor 1254 was reported to elevate basal levels of corticosterone in rats, but did not affect stress-induced levels (Miller et al., 1993). However, no effect on basal cortisol levels was seen in harbour seals fed with herring from the polluted Baltic Sea, whereas stress, subsequent to short fasting, induced a slightly higher elevation of cortisol than in the control animals fed North Atlantic herring (Swart, 1995). In contrast, fish from polluted waters had a reduced ability to elevate blood cortisol levels in response to an acute stress (Hontela et al., 1992; Hontela et al., 1995; Hontela et al., 1997). Negative correlations were observed between the amount of total PCDDs/PCDFs, total PCBs, or non-ortho PCBs in individual yolk sac and basal plasma corticosterone levels in herring gull embryos (Lorenzen et al., 1999). Thus, there are examples of adrenocortical impairment in wildlife that perhaps could be related to the mechanisms of actions described in this thesis.

37
Concluding remarks Environmental pollutants such as aryl methyl sulphones may affect the glucocorticoid system by inhibition of CYP enzyme(s) involved in the glucocorticoid synthesis pathway. A decrease in corticosteroid production caused by competitive inhibition of CYP11B1 would probably, due to the feedback control of the HPA axis, increase the secretion of ACTH. Furthermore, some aryl methyl sulphones may exert adrenocorticolytic effects in the zones of the adrenal cortex that express CYP11B1. This may lead to insufficient glucocorticoid production in vivo. Several substances affected the glucocorticoid-signalling pathway by antagonistic effects on the GR. The structural resemblance of a methyl sulphonyl group and an azole group may be important for the affinity to the GR. In addition, all PCB methyl sulphones with affinity to the GR are chiral congeners. However, even though there were enantiomeric differences in biliary/intestinal excretion, which may explain the enantiomeric ratios in biota, their inhibitory effects on dexamethasone-induced TAT activity were similar. Effects of chiral substances, such as atropisomeric MeSO2-PCBs, are mainly observed for the racemic mixture. Considering the differences in persistency of MeSO2-PCBs in biota, there is a need to separately study the effects of the enantiomers to support a proper risk assessment of these chiral substances. Co-exposure of substances revealed additive antagonistic effects on glucocorticoid regulated genes, indicating that the difference in effective and inhibitory concentration may be less between the endogenous steroid and the persistent environmental pollutants. Hence, co-exposure of chemicals, such as PCB methyl sulphones and tolylfluanid, may have an impact on humans and wildlife due to additive effects. Further studies on their possible effects on the glucocorticoid-regulation in vivo are needed. This thesis shows that several environmental pollutants can interact with the glucocorticoid hormone system. There is thus a need for rapid in vitro test systems as screening tools to predict chemicals that may be potential glucocorticoid disrupters. My studies showed that the human H295R cell line is a useful in vitro test system for studies of chemical interference with the glucocorticoid production. However, liver tissue slice culture was found to be

38
too insensitive to serve as a useful test system on glucocorticoid-regulated TAT activity. Instead, cell lines that express glucocorticoid-regulated genes such as the GRAF and/or H4-II-E-C3 cell lines are proposed as useful tools.

39
Acknowledgements This work was performed at the Dept. of Environmental Toxicology, Uppsala University, and at the Dept. of Pharmacology and Toxicology, Swedish University of Agricultural Sciences, Sweden. I wish to express my sincere gratitude to all those who in any way have contributed to this thesis, and especially to:
Associate Professor Bert-Ove Lund, the one and only Supervisor. You always had a moment to spare and scientific thoughts to share.
Professor Ingvar Brandt, for providing good working facilities and a friendly atmosphere, as well as pep-talks…
Professor Björn Brunström, for statistical advice
My co-authors: Prof. Åke Bergman, Dr. Stefan Nilsson, Dr. Thomas Sanderson, Dr. Niklas Johansson for valuable scientific discussions.
/ Dr. Christina Larsson, co-author, for synthesis of all PCB methyl sulphones, valuable discussions about science and other matters in life, and for your friendship. ☺ Dr. Örjan Lindhe, for endless talks about the adrenal cortex and one specific methyl sulphone. For technical assistance with Krumdieck. ☼ All the present and past friends and colleagues at Environmental Toxicology, Pharmacology and Toxicology, and Toxicology in Uppsala. Especially Carina, Maria, Fariba, Ulrika and Maggan. ☯ Lena for your friendship, baby-walks and talks, support, strategies, and above all giving me perspective of life.
To my family, Mats and Hulda, for your love, encouragement, patience and support. You are the sunshine of my life. Kjell, Karin, Emma and Jonas- for so many reasons... Anders and Kersti for care and support in so many different ways. I’d never had come this far without my family. I love you all!
This work was funded by the Oscar and Lili Lamm’s Foundation, the Swedish Environmental Protection Agency, the Central Agency of Laboratory Animals and the Helge A:xson Johnson Foundation.

40
References Albertson B.D., Maronian N.C., Frederick K.L., DiMattina M., Feuillan P., Dunn J.F. and
Loriaux D.L. (1988) The effect of ketoconazole on steroidogenesis II. Adrenocortical enzyme activity in vitro. Research Communications in Chemical Pathology and Pharmacology 61, 27-34.
Ayub M. and Levell M.J. (1989) Inhibition of human adrenal steroidogenic enzymes in vitro by imidazole drugs including ketoconazole. Journal of Steroid Biochemistry 32, 515-524.
Bakke J.E., Bergman Å. and Larsen G.L. (1982) Metabolism of 2,4',5-trichlorobiphenyl in the mercapturic pathway. Science 217, 645-647.
Bamberger C.M., Schulte H.M. and Chrousos G.P. (1996) Molecular determinants of glucocorticoid receptor function and tissue sensitivity to glucocorticoids. Endocrine Reviews 17, 245-261.
Bergman A. and Olsson M. (1985) Pathology of Baltic grey seal and ringed seal females with special reference to adrenocortical hyperplasia: is environmental pollution the cause of a widely distributed disease syndrome? Finnish Game Research, 47-62.
Bergman Å., Norstrom R.J., Haraguchi K., Kuroki H. and Béland P. (1994) PCB and DDE methyl sulfones in mammals from Canada and Sweden. Environmental Toxicology and Chemistry 13, 121-128.
Bodwell J.E., Hu L.-M., Hu J.-M., Ortí E. and Munck A. (1993) Glucocorticoid receptors: ATP-dependent cycling and hormone-dependent hyperphosphorylation. Journal of Steroid Biochemistry and Molecular Biology 47, 31-38.
Bodwell J.E., Webster J.C., Jewell C.M., Cidlowski J.A., Hu J.-M. and Munck A. (1998) Glucocorticoid receptor phosphorylation: Overview, function and cell cycle-dependence. Journal of Steroid Biochemistry and Molecular Biology 65, 91-99.
Brandt I. and Bergman Å. (1981) Bronchial mucosal and kidney cortex affinity of 4- and 4,4'-substituted sulphur-containing derivatives of 2,2',5,5'-tetrachlorobiphenyl in mice. Chemico-Biological Interactions 34, 47-55.
Brandt I. and Bergman Å. (1987) PCB methyl sulphones and related compounds: identification of target cells and tissues in different species. Chemosphere 16, 1671-1676.
Brandt I., Klasson-Wehler E., Rafter J. and Bergman Å. (1982) Metabolism of 2,4',5-trichlorobiphenyl: tissue concentrations of methylsulphonyl-2,4',5-trichlorobiphenyl in germfree and conventional mice. Toxicology Letters 12, 273-280.
Brandt I., Lund J., Bergman Å., Klasson-Wehler E., Poellinger L. and Gustafsson J.Å. (1985) Target cells for the polychlorinated biphenyl metabolite 4,4'-bis(methylsulfonyl)-2,2',5,5'-tetrachlorobiphenyl in lung and kidney. Drug Metabolism and Disposition 13, 490-496.
Brown J.W. and Fishman L.M. (2000) Biosynthesis and metabolism of steroid hormones by human adrenal carcinomas. Brazilian Journal of Medical and Biological Research 33, 1235-1244.

41
Buckingham J.C., Cowell A.-M., Gillies G.E., Herbison A.E. and Steel J.H. (1997) The neuroendocrine system: Anatomy, physiology and responses to stress. In Stress, stress hormones and the immune system. Edited by J.C. Buckingham, G.E. Gillies & A.-M. Cowell. Wiley J. & Son Ltd, Chichester.
Cadepond F., Ulmann A. and Baulieu E.-E. (1997) RU486 (mifepristone): Mechanisms of action and clinical uses. Annual Reviews of Medicine 48, 129-156.
Carballeira A., Fishman L.M. and Jacobi J.D. (1976) Dual sites of inhibition by metyrapone of human adrenal steroidogenesis: Correlation of in vivo and in vitro studies. Journal of Clinical Endocrinology and Metabolism 42, 687-695.
Christoffels V.M., Grange T., Kaestner K.H., Cole T.J., Darlington G.J., Croniger C.M. and Lamers W.H. (1998) Glucocorticoid receptor, C/EBP, HNF3, and protein kinase A coordinately activate the glucocorticoid response unit of the carbamoylphosphate synthetase I gene. Molecular and Cellular Biology 18, 6305-6315.
Chrousos G.P., Laue L., Nieman L.K., Kawai S., Udelsman R.U., Brandon D.D. and Loriaux D.L. (1988) Glucocorticoids and glucocorticoid antagonists: Lessons from RU486. Kidney Int 34, S18-S23.
Coghlan M.J., Kym P.R., Elmore S.W., Wang A.X., Luly J.R., Wilcox D., Stashko M., Lin C.-W., Miner J., Tyree C., Nakane M., Jacobson P. and Lane B.C. (2001) Synthesis and characterization of non-steroidal ligands for the glucocorticoid receptor: Selective Quinoline derivatives with prednisolone-equivalent functional activity. Journal of Medical Chemistry 44, 2879-2885.
Diamondstone T.I. (1966) Assay of tyrosine transaminase activity by conversion of p-hydroxyphenylpiruvate to p-hydroxybenzaldehyde. Analytical Biochemistry 16, 395-401.
Ellerichmann T., Bergman Å., Franke S., Huhnerfuss H., Jakobsson E., König W.A. and Larsson C. (1998) Gas chromatographic enantiomer separations of chiral PCB methyl sulfones and identification of selectively retained enantiomers in human liver. Fresenius Environmental Bulletin 7, 244-257.
Engelhardt D. and Weber M.M. (1994) Therapy of Cushing's syndrome with steroid biosynthesis inhibitors. Journal of Steroid Biochemistry and Molecular Biology 49, 261-267.
Es-Souni M., Ramirez L.C. and Bournot P. (1992) 18-hydroxylation in the Y-1 adrenal cell line: response to ACTH and to culture conditions. Journal of Steroid Biochemistry and Molecular Biology 43, 535-541.
Feldman D. (1986) Ketoconazole and other imidazole derivatives as inhibitors of steroidogenesis. Endocrine Reviews 7, 409-420.
Freeman H.C. and Sangalang G.B. (1977) A study of the effects of methyl mercury, cadmium, arsenic, selenium, and a PCB, (Aroclor 1254) on adrenal and testicular steroidogeneses in vitro, by the gray seal Halichoerus grypus. Archives of Environmental Contamination and Toxicology 5, 369-383.
Gayda D.P. and Pariza M.W. (1983) Effects of carcinogens on hormonal regulation of gene expression in primary cultures of adult rat hepatocytes. Carcinogenesis 4, 1127-1131.

42
Gazdar A.F., Oie H.K., Shackelton C.H., Chen T.R., Triche T.J., Myers C.E., Chrousos G.P., Brennan M.F., Stein C.A. and La Rocca R.V. (1990) Establishment and characterization of a human adrenocortical carcinoma cell line that expresses multiple pathways of steroid biosynthesis. Cancer Research 50, 5488-5496.
Gray L.E., Ostby J., Furr J., Wolf C.J., Lambright C., Parks L., Veeramachaneni D.N., Wilson V., Price M., Hotchkiss A., Orlando E. and Guilette L. (2001) Effects of environmental antiandrogens on reproductive development in experimental animals. Human Reproduction Update 7, 248-264.
Grippo J.F., Holmgren A. and Pratt W.B. (1985) Proof that the endogenous heat-stable glucocorticoid receptor activating factor is thioredoxin. Journal of Biological Chemistry 258, 13658-13664.
Haraguchi H., Kuroki H., Masuda Y. and Shigematsu N. (1984) Determination of methylthio and methylsulphone polychlorinated biphenyls in tissues of patients with 'yusho'. Food Chem Toxicology 22, 283-288.
Haraguchi K., Athanasiadou M. and Bergman Å. (1992) PCB and PCB methyl sulfones in selected groups of seals from Swedish waters. Ambio 21, 546-549.
Hart A., Mattheyse F.J. and Balinsky J.B. (1983) An organ culture of postnatal rat liver slices. In Vitro 19, 841-852.
Harvey J., Paine A.J., Maurel P. and Wright M.C. (2000) Effect of the adrenal 11-b-hydroxylase inhibitor metyrapone on human hepatic cytochrome P-450 expression: Induction of cytochrome P-450 3A4. Drug Metabolism and Disposition 28, 96-101.
Hayes T.B., Wu T.H. and Gill T.N. (1997) DDT-like effects as a result of corticosterone treatment in an anuran amphibian: is DDT a corticoid mimic or a stressor? Environmental Toxicology and Chemistry 16, 1948-1953.
Heiman A.S. and Tonner L.E. (1995) The acute effect of lead acetate on glucocorticoid regulation of tyrosine aminotransferase in hepatoma cells. Toxicology 1995 Jun 26;100(1-3):57-68 100, 57-68.
Hinson J.P. and Raven P.W. (1996) Adrenal morphology and hormone synthesis and regulation. In The adrenal in toxicology: Target organ and modulator of toxicity. Edited by P.W. Harvey. pp 23-52. Taylor & Francis Ltd, England, London.
Hoekstra P.F., Burnison B.K., Neheli T. and Muir D.C.G. (2001) Enantiomer-specific activity of o,p'-DDT with the human estrogen receptor. Toxicology Letters 125, 75-81.
Hontela A., Daniel C. and Rasmussen J.B. (1997) Structural and functional impairment of the hypothalamo-pituitary-interrenal axis in fish exposed to bleached kraft mill effluent in the St Maurice River, Québec. Ecotoxicology, 1-12.
Hontela A., Dumont P., Duclos D. and Fortin R. (1995) Endocrine and metabolic dysfunction in yellow perch, Perca flavescens, exposed to organic contaminants andheavy metals in the St. Lawrence River. Environmental Toxicology and Chemistry 14, 725-731.
Hontela A., Rasmussen J.B., Audet C. and Chevalier G. (1992) Impaired cortisol stress response in fish from environments polluted by PAHs, PCBs, and mercury. Archives of Environmental Contamination and Toxicology 22, 278-283.
Hornsby P. (1989) Steroid and xenobiotic effects on the adrenal cortex: Mediated by oxidative and other mechanisms. Free Radical Biology and Medicine 6, 103-115.

43
Hu L.-M., Bodwell J.E., Hu J.-M., Orti E. and Munck A. (1994) Glucocorticoid receptors in
ATP-depleted cells. Journal of Biological Chemistry 269, 6571-6577. Ideyama Y., Kudoh M., Tanimoto K., Susaki Y., Nanya T., Nakahara T., Ishikawa H.,
Fujikura T., Akaza H. and Shikama H. (1999) YM116, 2-(1H-imidazol-4-ylmethyl)-9H-carbazole, decreases adrenal androgen synthesis by inhibiting C17-20 lyase activity in NCI-H295 human adrenocortical carcinoma cells. Japanese Journal of Pharmacology 79, 213-220.
Janák K., Becker G., Colmsjö A., Östman C., Athanasiadou M., Valters K. and Bergman Å. (1998) Methyl sulfonyl polychlorinated biphenyls and 2,2-bis(4-chlorophenyl)-1,1-dichloroethane in gray seal tissue determined by gas chromatography with electron capture detection and atomic emission detection. Environmental Toxicology and Chemistry 17, 1046-1055.
Jensen S. and Jansson B. (1976) Antropogenic substances in seal from the Baltic: Methyl sulfone metabolites. Ambio 5, 257-260.
Jönsson C.-J. (1994) Decreased plasma corticosterone levels in suckling mice following injection of the adrenal toxicant, MeSO2-DDE, to the lactating dam. Pharmacology and Toxicology 74, 58-60.
Jönsson C.-J. and Lund B.-O. (1994) In vitro bioactivation of the environmental pollutant 3-methylsulphonyl-2, 2-bis(4-chlorophenyl)-1,1-dichloroethene in the human adrenal gland. Toxicology Letters 71, 169-175.
Jönsson C.-J., Lund B.-O., Bergman Å. and Brandt I. (1992) Adrenocortical toxicity of 3-methylsulphonyl-DDE; 3: Studies in fetal and suckling mice. Reproductive Toxicology 6, 233-240.
Jönsson C.-J., Lund B.-O. and Brandt I. (1993) Adrenocorticolytic DDT-metabolites: Studies in mink, Mustela vision and otter, Lutra lutra. Ecotoxicology 2, 41-53.
Jönsson C.-J., Rodriguez-Martinez H., Lund B.-O., Bergman Å. and Brandt I. (1991) Adrenocortical toxicity of 3-methylsulfonyl-DDE in mice. II. Mitochondrial changes following ecologically relevant doses. Fundamental and Applied Toxicology 16, 365-374.
Kan P.B., Hirst M.A. and Feldman D. (1985) Inhibition of steroidogenic cytochrome P-450 enzymes in rat testis by ketoconazole and related imidazole anti-fungal drugs. Journal of Steroid Biochemistry 23, 1023-1029.
Kato Y., Haraguchi K., Kawashima M., Yamada S., Masuda Y. and Kimura R. (1995a) Characterization of hepatic microsomal cytochrome P-450 from rats treated with methylsulphonyl metabolites of polychlorinated biphenyl congeners. Chemico-Biological Interactions 95, 269-278.
Kato Y., Haraguchi K., Kawashima M., Yamada S., Masuda Y. and Kimura R. (1995b) Induction of hepatic microsomal drug-metabolizing enzymes by methylsulphonyl metabolites of polychlorinated biphenyl congeners in rats. Chemico-Biological Interactions 95, 257-268.
Kato Y., Haraguchi K., Masuda Y., Kimura R. and Wärngård L. (1998a) Inhibition of cell-cell communication by methylsulfonyl metabolites of polychlorinated biphenyl congeners. Archives of Toxicology 72, 178-182.

44
Kato Y., Haraguchi K., Shibahara T., Masuda Y. and Kimura R. (1998b) Reduction of thyroid hormone levels by methylsulfonyl metabolites of polychlorinated biphenyl congeners in rats. Archives of Toxicology 72, 541-544.
Kato Y., Haraguchi K., Shibahara T., Shinmura Y., Masuda Y. and Kimura R. (2000b) The induction of hepatic microsomal UDP-glucuronosyltransferase by the methylsulfonyl metabolites of polychlorinated biphenyl congeners in rats. Chemico-Biological Interactions 125, 107-115.
Kato Y., Haraguchi K., Shibahara T., Yumoto S., Masuda Y. and Kimura R. (1999) Reduction of thyroid hormone levels by methylsulfonyl metabolites of tetra- and pentachlorinated biphenyls in male Sprague-Dawley rats. Toxicological Sciences 48, 51-54.
Kato Y., Haraguchi K., Shibahara T., Yumoto S., Masuda Y. and Kimura R. (2000a) Reduction of serum thyroxine concentrations by methylsulfonyl metabolites of tetra-, penta- and hexachlorinated biphenyls in male Sprague-Dawley rats. Chemosphere 40, 1233-1240.
Kato Y., Haraguchi K., Tomiyasu K., Saito H., Isogai M., Masuda Y. and Kimura R. (1997) Structure-dependent induction of CYP2B1/2 by 3-methylsulfonyl metabolites of polychlorinated biphenyl congeners in rats. Environmental Toxicology and Pharmacology 3, 137-144.
Kavlock R.J., Daston G.P., DeRosa C., Fenner-Crisp P., Gray L.E., Kaattari S., Lucier G., Luster M., Mac M.J., Maczka C., Miller R., Moore J., Rolland R., Scott G., Sheehan D.M., Sinks T. and Tilson H.A. (1996) Research needs for the risk assessment of health and environmental effects of endocrine disruptors: a report of the U.S. EPA-sponsored workshop. Environmental Health Perspectives 104 Suppl, 715-740.
Kelce W.R., Stone C.R., Laws S.C., Gray L.E., Kemppainen J.a. and Wilson E.M. (1995) Persistent DDT metabolite p,p'-DDE is a potent androgen receptor antagonist. Nature 375, 581-585.
Keller-Wood M.E. and Dallman M.F. (1985) Corticosteroid inhibition of ACTH secretion. Endocrine Reviews 5, 1-24.
Kemikalieinspektionen. (1998) PM Nr. 6/98. Ämnen mot svampangrepp. Solna Klasson-Wehler E., Bergman Å., Kowalski B. and Brandt I. (1987) Metabolism of 2,3,4',6-
tetrachlorobiphenyl: formation and tissue localization of mercapturic acid pathway metabolites in mice. Xenobiotica 17, 477-486.
Lamberts S.W., Bons E.G., Bruining H.A. and de Jong F.H. (1987) Differential effects of the imidazole derivatives etomidate, ketoconazole and miconazole and of metyrapone on the secretion of cortisol and its precursors by human adrenocortical cells. Journal of Pharmacology and Experimental Therapeutics 240, 259-264.
Larsson C. (2002) Atropisomers of PCB methyl sulfone metabolites: Radiosynthesis, analysis and distribution. Thesis. Stockholm University, Stockholm
Larsson C., Ellerichmann T., Hühnerfuss H. and Bergman Å. (2002) Chiral PCB methyl sulfones in rat tissues after exposure to technical PCBs. Environmental Science and Technology, in press,

45
Letcher R.J. (1996) The ecological and analytical chemistry of chlorinated hydrocarbon contaminants and methyl sulfonyl-containing metabolites in the polar bear (Ursus maritimus) food chain. Thesis. Carleton University, Ottawa
Letcher R.J., Klasson-Wehler E. and Bergman Å. (2000) Methyl sulfone and hydroxylated metabolites of polychlorinated biphenyls. In The Handbook of Environmental Chemistry. Edited by J. Paasivirta. pp 315-359. Springer-Verlag, Berlin Heidelberg.
Lindhe Ö. (2001) Adrenal Bioactivation and toxicity of 3-MeSO2-DDE, o,p'-DDD and DMBA investigated in tissue slice culture. Thesis. Uppsala University, Uppsala.
Lindhe Ö., Lund B.-O., Bergman Å. and Brandt I. (2001) Irreversible binding and adrenocorticolytic activity of the DDT metabolite 3-methylsulfonyl-DDE examined in tissue slice culture. Environmental Health Perspectives 109, 105-110.
Lindhe Ö., Skogseid B. and Brandt I. (2002) Cytochrome P450-catalysed binding of 3-methylsulfonyl-DDE and o,p'-DDDin human adrenal zona fasciculata/reticularis. The Journal of Clinical Endocrinology and Metabolism 87, 1319-1326.
Loose D.S., Stover E.P. and Feldman D. (1983) Ketoconazole binds to glucocorticoid receptors and exhibits glucocorticoid antagonist activity in cultured cells. Journal of Clinical Investigation 72, 404-408.
Lorenzen A. and Kennedy S.W. (1993) A fluorescence-based protein assay for use with a microplate reader. Analytical Biochemistry 214, 346-348.
Lorenzen A., Moon T.W., Kennedy S.W. and Fox G.A. (1999) Relationships between environmental organochlorine contaminant residues, plasma corticosterone concentrations, and intermediary metabolic enzyme activities in Great Lakes herring gull embryos. Environmental Health Perspectives 107, 179-186.
Lowry O.H., Rosebrough N.J., Farr A.L. and Randall R.J. (1951) Protein measurement with the folin reagent. The Journal of Biological Chemistry 193, 265-275.
Lund B.-O. (1994) In vitro adrenal bioactivation and effects on steroid metabolism of DDT and their metabolites in the gray seal (Halichoerus grypus). Environmental Toxicology and Chemistry 13, 911-917.
Lund B.-O., Bergman A. and Brandt I. (1988) Metabolic activation and toxicity of a DDT-metabolite, 3-methylsulphonyl-DDE, in the adrenal zona fasciculata in mice. Chemico-Biological Interactions 65, 25-40.
Lund B.-O. and Lund J. (1995) Novel involvement of a mitochondrial steroid hydroxylase (P450c11) in xenobiotic metabolism. The Journal of Biological Chemistry 270, 20895-20897.
Lund B.-O., Örberg J., Bergman Å., Larsson C., Bergman A., Bäcklin B.-M., Håkansson H., Madej A., Brouwer A. and Brunström B. (1999) Chronic and reproductive toxicity of a mixture of 15 methylsulfonyl-polychlorinated biphenyls and 3-methylsulfonyl-2,2-bis(4-chlorophenyl)-1,1-dichloroethene in mink (Mustela vision). Environmental Toxicology and Chemistry 18, 292-298.
Lund J., Brandt I., Poellinger L., Bergman Å., Klasson-Wehler E. and Gustafsson J.Å. (1985) Target cells for the polychlorinated biphenyl metabolite 4,4'-bis(methylsulfonyl)-2,2',5,5'-tetrachlorobiphenyl. Characterization of high affinity binding in rat and mouse lung cytosol. Molecular Pharmacology 27, 314-323.

46
Mann R.D. (1996) Adverse drug reactions and the adrenal glands. In The adrenal in toxicology: Target organ and modulator of toxicity. Edited by P.W. Harvey. pp 279-300. Taylor & Francis, London.
Meerts I.A.T.M., Letcher R.J., Hoving S., Marsh G., Bergman Å., Lemmen J.G., van den Burg B. and Brouwer A. (2001) In vitro estrogenicity of polybrominated diphenyl ethers, hydroxylated PBDEs, and polybrominated bisphenol A compounds. Environmental Health Perspectives 109, 399-407.
Miesfeld R., Rusconi S., Godowski P.J., Maler B.A., Okret S., Wikström A.-C., Gustafsson J.Å. and Yamamoto K.R. (1986) Genetic complementation of a glucocorticoid receptor deficiency by expression of cloned receptor cDNA. Cell 46, 389-399.
Miller D.B., Gray L.E.J., Andrews J.E., Luebke R.W. and Smialowicz R.J. (1993) Repeated exposure to the polychlorinated biphenyl (Aroclor 1254) elevates the basal serum levels of corticosterone but does not affect the stress-induced rise. Toxicology 81, 217-222.
Mitchell A. and O'Keane V. (1998) Steroids and depression: Glucocorticoid steroids affect behaviour and mood. British Medical Journal 316, 244-245.
Mosmann T. (1983) Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. Journal of Immunological Methods 65, 55-63.
Muller M. and Renkawitz R. (1991) The glucocorticoid receptor. Biochemica et Biophysica Acta 1088, 171-182.
Munck A., Guyre P.M. and Holbrook N.J. (1984) Physiological functions of glucocorticoids in stress and their relation to pharmacological actions. Endocrine Reviews 5, 25-44.
Nagai K., Miyamori I., Takeda R., Suhara K. and Katagiri M. (1987) Effect of ketoconazole, etomidate and other inhibitors of steroidogenesis on cytochrome P-450sccII-catalyzed reactions. Journal of Steroid Biochemistry 28, 333-336.
Navas J.M. and Segner H. (2000) Antiestrogenicity of beta-nphtoflavone and PAHs in cultured rainbow trout hepatocytes: evidence for a role of the arylhydrocarbon receptor. Aquatic Toxicology 51, 79-92.
Newsome W.H. and Davies D. (1996) Determination of PCB metabolites in Canadian human milk. Chemosphere 33, 559-565.
Nicholls P.J., Daly M.J. and Smith H.J. (1986) Pharmacology of aminoglutethimide: Structure/activity relationships and receptor interactions. Breast Cancer Research and Treatment 7, S55-S67.
Niimi S., Yamaguchi T. and Hayakawa T. (1998) Effect of dexamethasone pretreatment on the dexamethasone-dependent induction of tyrosine aminotransferase activity in primary cultured rat hepatocytes. Biological and Pharmacological Bulletin 21, 1009-1012.
Nitsch D., Boshart M. and Schutz G. (1993) Activation of the tyrosine aminotransferase gene is dependent on synergy between liver-specific and hormone-responsive elements. Proceedings of National Academic Scienes USA 90, 5479-5483.
Nordeen S.K., Bona B.J., Beck C.A., Edwards D.P., Borror K.C. and DeFranco D.B. (1995) The two faces of a steroid antagonist: When an antagonist isn't. Steroids 60, 97-104.

47
Nordeen S.K., Moyer M.L. and Bona B.J. (1994) The coupling of multiple signal transduction pathways with steroid response mechanisms. Endocrinology 134, 1723-1732.
Norén K., Lundén Å., Pettersson E. and Bergman Å. (1996) Methylsulfonyl metabolites of PCBs and DDE in human milk in Sweden, 1972-1992. Environmental Health Perspectives 104, 766-772.
Rainey W.E., Bird I.M. and Mason J.I. (1994) The NCI-H295 cell line: a pluripotent model for human adrenocortical studies. Molecular and Cellular Endocrinology 100, 45-50.
Rainey W.E., Bird I.M., Sawetawan C., Hanley N.A., McCarthy J.L., McGee E.A., Wester R. and Mason J.I. (1993) Regulation of human adrenal carcinoma cell (NCI-H295) production of C19 steroids. Journal of Clinical Endocrinology and Metabolism 77, 731-737.
Rangarajan P.N., Umesono K. and Evans R.M. (1992) Modulation of glucocorticoid receptor function by protein kinase A. Molecular Endocrinology 6, 1451-1457.
Rapoport R., Sklan D. and Hanukoglu I. (1995) Electron leakage from the adrenal cortex mitochondrial P450scc and P450c11 systems: NADPH and steroid dependence. Archives of Biochemistry and Biophysics 317, 412-416.
Raven P.W. and Hinson J.P. (1996) Transport, actions and metabolism of adrenal hormones and pathology and pharmacology of the adrenal gland. In The adrenal in toxicology: Target organ and modulator of toxicity. Edited by P.W. Harvey. pp 53-79. Taylor & Francis Ltd, England, London.
Samuels H.H. and Tomkins G.M. (1970) Relation of steroid structure to enzyme induction in hepatoma tissue culture cells. Journal of Molecular Biology 52, 57-74.
Sanderson J.T., Letcher R.J., Heneweer M., Giesy J.P. and van den Berg M. (2001) Effects of chloro-s-triazine herbicides and metabolites on aromatase activity in various human cell lines and on vitellogenin production in male carp hepatocytes. Environmental Health Perspectives 109, 1027-1031.
Sanderson J.T., Seinen W., Giesy J.P. and van den Berg M. (2000) 2-Chloro-s-triazine herbicides induce aromatase (CYP19) activity in the H295R human adrenocortical carcinoma cells: A novel mechanism for estrogenicity? Toxicological Sciences 54, 121-127.
Sapolsky R.M., Romero L.M. and Munck A. (2000) How do glucocorticoids influence stress response? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocrine Reviews 21, 55-89.
Sarkar N.N. (2002) Mifepristone: bioavailability, pharmacokinetics and use-effectiveness. European Journal of Obstetrics & Gynecology and Reproductive Biology 101, 113-120.
Shaw M.A., Nicholls P.J. and Smith H.J. (1988) Aminoglutethimide and ketoconazole: Historical perspectives and future prospects. Journal of Steroid Biochemistry 31, 137-146.
Simons S.S.Jr., Pumphrey J.G., Rudikoff S. and Eisen H.J. (1987) Identification of cysteine 656 as the amino acid of hepatoma tissue culture cell glucocorticoid receptors that is covalently labeled by dexamethasone 21-mesylate. The Journal of Biological Chemistry 262, 9676-9680.

48
Soto A.M., Sonnenschein C., Chung K.L., Fernandez M.F., Olea N. and Serrano F.O. (1995) The E-SCREEN assay as a tool to identify estrogens: an update on estrogenic environmental pollutants. Environmental Health Perspectives 103 Suppl, 113-122.
Spindler K.D., Kanuma K. and Grossman D. (1991) Uptake of corticosterone into isolated rat liver cells: possible involvement of Na+/K+-ATPase. Journal of Steroid Biochemistry and Molecular Biology 38, 721-725.
Staels B., Hum D.W. and Miller W.L. (1993) Regulation of steroidogenesis in NCI-H295 cells: a cellular model of the human fetal adrenal. Molecular Endocrinology 7, 423-433.
Sultan K., Hartung J. and Bade E.G. (1996) Hormone- and endotoxin-modulated gene expression of a long-term organ culture system of adult rat liver. Fresenius Environmental Bulletin Letters 394, 51-54.
Swart d.R.L. (1995) Impaired immunity in seals exposed to bioaccumulated environmental contaminants. Thesis. Erasmus Universiteit Rotterdam, The Netherlands, Rotterdam
Tobes M.C., Hays S.J., Gildersleeve D.L., Wieland D.M. and Beierwaltes W.H. (1985) Adrenal cortical 11 beta-hydroxylase and side-chain cleavage enzymes. Requirement for the A- or B-pyridyl ring in metyrapone for inhibition. Journal of Steroid Biochemistry 22, 103-110.
Turney M.K. and Kovacs W.J. (2001) Function of a truncated glucocorticoid receptor form at a negative glucocorticoid response element in the proopiomelanocortin gene. Journal of Molecular Endocrinology 26, 43-49.
Weistrand C. and Norén K. (1997) Methylsulfonyl metabolites of PCBs and DDE in human tissue. Environmental Health Perspectives 105, 644-649.
Venkatakrishnan K., von Moltke L.L. and Greenblatt D.J. (2000) Effects of the antifungal agentson oxidative drug metabolism. Clinical Pharmacokinetics 38, 111-180.
Wiberg K., Letcher R.J., Sandau C., Duffe J., Norstrom R.J., Haglund P. and Bidleman T. (1998) Enantioselective gas chromatography/mass spectrometry of methylsulfonyl PCBs with application to arctic marine mammals. Analytical Chemistry 70, 3845-3852.
Wiegel N.L. and Zhang Y. (1998) Ligand-independent activation of steroid hormone receptors. Journal of Molecular Medicine 76, 469-479.
Zenkert S., Schubert B., Fassnacht M., Beuschlein F., Allolio B. and Reincke M. (2000) Steroidogenic acute regulatory protein mRNA expression in adrenal tumours. European Journal of Endocrinology 142, 294-299.
Ziegler R. and Kasperk C. (1998) Glucocorticoid-induced osteoporosis: prevention and treatment. Steroids 63, 344-348.
‘Things can only get better’ Howard Jones