influences of submerged plant collapse on diet composition
TRANSCRIPT

RESEARCH Open Access
Influences of submerged plant collapse ondiet composition breadth and overlapamong four crane species at Poyang LakeChinaJinjin Hou1 Lei Li123 Yafang Wang1 Wenjuan Wang123 Huiying Zhan4 Nianhua Dai5 and Ping Lu5
Abstract
Background Interannual variation in resource abundance has become more unpredictable and food shortageshave increasingly occurred in the recent decades However compared to seasonal fluctuations in resourceabundance the influences of interannual variation in resource abundance on the dietary niches of consumersremain poorly understood Poyang Lake China is a very important wintering ground for the globally endangeredSiberian Crane (Leucogeranus leucogeranus) White-naped Crane (Grus vipio) and Hooded Crane (G monacha) aswell as the non-endangered Eurasian Crane (G grus) Tubers of Vallisneria spp the dominant submergedmacrophytes at Poyang Lake is an important food for cranes Nevertheless submerged macrophytes haveexperienced serious degradation recently In this study we used metabarcoding technology to explore theconsequences of Vallisneria tuber collapse on the diet compositions breadths and overlaps of the four cranespecies based on fecal samples collected in winter 2017 (a year with tuber collapse) and winter 2018 (a year withhigh tuber abundance)
Results Compared to previous studies our study elucidates crane diets in an unprecedented level of detailVallisneria tubers was confirmed as an important food source of cranes Surprisingly the grassland plant Polygonumcriopolitanum was also found to be an important food source in the feces of cranes Agricultural fields wereimportant foraging sites for Siberian Cranes White-naped Cranes and Hooded Cranes providing foods that allowedthem to survive in winters with natural food shortages However the three crane species preferred natural wetlandsto agricultural fields when the abundance of natural foods was high The abundance of Vallisneria tubers andprobably P criopolitanum greatly influenced the dietary compositions breadths and overlap of cranes Duringperiods of preferred resource shortage White-naped Cranes and Hooded Cranes widened their dietary niches whileSiberian Cranes maintained a stable niche width The dietary niche overlap among crane species increasedsubstantially under conditions of plentiful preferred food resources
(Continued on next page)
copy The Author(s) 2021 Open Access This article is licensed under a Creative Commons Attribution 40 International Licensewhich permits use sharing adaptation distribution and reproduction in any medium or format as long as you giveappropriate credit to the original author(s) and the source provide a link to the Creative Commons licence and indicate ifchanges were made The images or other third party material in this article are included in the articles Creative Commonslicence unless indicated otherwise in a credit line to the material If material is not included in the articles Creative Commonslicence and your intended use is not permitted by statutory regulation or exceeds the permitted use you will need to obtainpermission directly from the copyright holder To view a copy of this licence visit httpcreativecommonsorglicensesby40The Creative Commons Public Domain Dedication waiver (httpcreativecommonsorgpublicdomainzero10) applies to thedata made available in this article unless otherwise stated in a credit line to the data
Correspondence wangwjncueducn wangwj870120163com1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China2Key Laboratory of Poyang Lake Environment and Resource UtilizationMinistry of Education Nanchang University Nanchang 330031 ChinaFull list of author information is available at the end of the article
Hou et al Frontiers in Zoology (2021) 1824 httpsdoiorg101186s12983-021-00411-2
(Continued from previous page)
Conclusions Our study emphasizes the superior quality of natural wetlands compared to agricultural fields asforaging habitats for cranes To provide safer and better foraging areas for cranes it is urgent to restore thesubmerged plants at Poyang Lake While high dietary niche overlap is often interpreted as intense interspecificcompetition our study highlights the importance of taking food abundance into account
Keywords Agricultural fields Food shortage Hooded crane Siberian crane Vallisneria tuber White-naped crane
BackgroundInterspecific competition is an important mechanism instructuring ecological communities [1 2] Competitionmay lead to resource (food habitat time) partitioningand facilitate the coexistence of sympatric species [1 34] Diet partitioning has been suggested to be the pri-mary mechanism of coexistence of many insects [5] fish[6] snakes [7] birds [8] and mammals [9]Many ecosystems are characterized by temporal varia-
tions in resource availability which may subsequently in-fluence dietary niche of birds [10ndash12] Migratory birdsmay also encounter strikingly different feeding condi-tions along their flyways [13 14] Therefore it is critic-ally important that birds are able to adjust their diets toexplore a wide range of food resources [11 15] Numer-ous examples of dietary flexibility exist Dunlins (Cali-dris alpine) [11] Semipalmated Sandpiper (C pusilla)[16] Hooded Crane (Grus monacha) [17] and Black-necked Crane (G nigricollis) [18] have all varied their di-ets with changes in prey availability In contrast strongdependence on a few prey types can make birds vulner-able to the reduction of prey abundance as has been thecase for Red Knot (C canutus) feeding on HorseshoeCrab (Limulus polyphemus) eggs [19]Optimal foraging theory is often used to predict the for-
aging decisions of animals [20 21] According to the the-ory when resources are abundant species shouldconsume the most profitable food items with high nutri-tion and accessibility while ignoring less profitable foodsWhen preferred food items become depleted species addless profitable foods to their diet [22 23] Great Knots (Ctenuirostris) for example changed their diet compositionand prey selection to include prey items that were physic-ally more difficult to digest after a severe decline in foodabundance and quality [24] Optimal foraging theory pre-dicts a reduction in dietary niche breadth during seasonsof high food abundance and an increasing trend duringseasons of low food abundance This pattern of nichewidth variation has been observed across a range of taxaincluding birds [12 25] mammals [26 27] fish [28 29]and invertebrates [30 31] The theory also predicts an in-crease in dietary niche overlap between species when re-sources are abundant [7 32 33] The increased overlapmay be related to the reduced competitive interaction be-tween species due to great resource availability For
example there was an absence of dietary niche partition-ing among shorebird species at Delaware Bay USA whenthe density of their primary prey Horseshoe Crab eggswas high [34] Extensive degree of dietary niche overlaphas also been revealed among seabird species breeding onthe Argentinean Patagonian coast with superabundance ofpelagic fish [35]Understanding how dietary niche changes for any spe-
cies as fluctuations in food abundance occurs is usefulfor designating management strategies to preserve bio-diversity and ecosystem services [32 36] However mostresearch is focused on the influences of seasonal vari-ation in food abundance on dietary niches [10 11 3237] Due to global climate change and increased humandisturbance interannual variation in resource abundancehas increased in frequency and intensity promulgatingfood scarcity at higher rates in recent decades [38ndash40]For example floods that negatively affect submergedplant growth and even lead to mass death of plants havebecome more frequent at Poyang Lake China [41ndash43]which may threaten the survival of birds that feed onthem [38 44 45] Seasonal variation in food abundanceoccurs regularly Animals have evolved strategies to copewith the predictable environmental variation [46] Inter-annual variation in food abundance occurs less fre-quently although its frequency and intensity haverecently increased [38ndash40] Without sufficient evolution-ary time animals may not adapt well to the interannualvariation thus it may pose a challenge for many taxa[19 47 48]Cranes are among the most threatened families of
birds [49] Poyang Lake which is located on the south-ern bank of the Yangtze River China is an importantwintering ground for cranes in East Asia (Fig 1) It sup-ports approximately 98 of the estimated global popula-tion of IUCN Critically Endangered Siberian Crane(Leucogeranus leucogeranus) approximately 18 of theestimated global population of Vulnerable White-napedCrane (Grus vipio) approximately 3 of the estimatedglobal population of Vulnerable Hooded Crane and ap-proximately half of the Chinese population of EurasianCrane (G grus) [50ndash52] With the degradation of lakesin the middle and lower Yangtze River floodplain Po-yang Lake is playing an increasingly important role incrane protection especially for the Siberian Crane [53
Hou et al Frontiers in Zoology (2021) 1824 Page 2 of 17
54] It has been suggested that other than Poyang Lakethere are no alternative wintering sites for SiberianCranes remaining in the Yangtze River floodplain [49]Tubers of Vallisneria spp the dominant submerged
macrophytes at Poyang Lake is an important food sourcefor cranes [38 55 56] However submerged macrophyteshave degraded seriously in recent decades [57ndash60] Theywere widely distributed throughout Poyang Lake in previ-ous years but were restricted to small areas in 2012 [58]The Vallisneria tuber density and biomass at three sub-lakes (Dahuchi Shahu and Meixihu) of Poyang Lake de-clined greatly in the winters from 1999 to 2017 [57 60]The frequency and range of Vallisneria tuber collapsehave also increased because of frequently occurring sum-mer flood and autumn drought extensive aquaculture de-clining water quality and other factors [38 57 60]Tuber collapse has led to dietary and foraging habitat
shifts of cranes For example the tuber collapses in thewinters of 2015 and 2016 drove thousands of SiberianCranes for the first time to switch from foraging inshallow waters they typically used to paddy fields and
lotus ponds [57 61] Their main foods changed fromVallisneria tubers to rice (Oryza sativa) seeds and lotus(Nelumbo nucifera) rhizomes [62] Many White-napedCranes Hooded Cranes and Eurasian Cranes also movedfrom natural wetlands to forage in paddy fields [63 64]Diet shifts may influence the dietary niche width andoverlap between crane species and subsequently thecompetition level and fitness of cranes Given the highlyendangered status of cranes and the importance of Po-yang Lake in crane protection it is important to under-stand the consequences of Vallisneria tuber collapse ondiets and competition levels among crane species so thateffective protection measures can be implementedTraditional dietary analysis methods include direct ob-
servation of foraging animals and microscopic examin-ation of feces or gut contents While these approacheshave been suggested to be useful in some cases they alsoexhibit methodological limitations [65ndash67] The maindrawbacks are that the methods can be inaccurate andlabor-intensive [65 68] Recently the emergence ofDNA metabarcoding has provided new perspectives for
Fig 1 Maps showing the three Vallisneria tuber survey sublakes (Dahuchi Shahu and Changhuchi) and the sampling sites of crane feces atPoyang Lake Circles and squares represent sampling sites in the winters of 2017 and 2018 respectively The colors of the circles and squarescorrespond to the crane species red Siberian Crane blue White-naped Crane green Hooded Crane and purple Eurasian Crane The base mapshows the inundation area of Poyang Lake in winter 2008 when the water level was similar to the historical average water level
Hou et al Frontiers in Zoology (2021) 1824 Page 3 of 17
diet analysis [65] This method is based on amplifyingand high-throughput sequencing a standardized DNAregion from stomach contents or feces and subsequentlycomparing it to a reference database for identification ofthe consumed species [65] Compared to traditionalmethods DNA metabarcoding generally provides highertaxonomic resolution identifies more food items andcan simultaneously analyze a larger number of samples[69ndash71] After its first application in assessment of thediets of Australian Fur Seals (Arctocephalus pusillus dor-iferus) [72] metabarcoding has been used successfully tostudy diets of herbivores [9 73] carnivores [69 74] andomnivores [75 76]In this study we used metabarcoding technology to
explore the consequences of Vallisneria tuber collapseon the diet compositions niche breadths and nicheoverlaps of the four crane species at Poyang Lake Fecalsamples were collected in winter 2017 (ie 20172018 ayear with Vallisneria tuber collapse) and winter 2018(ie 20182019 a year with high Vallisneria tuber abun-dance see Results for details) We first explored the di-ets of the four crane species and their interannualvariations Most previous dietary studies of cranes werebased on traditional direct observation or microscopicexamination Here we hoped to apply the metabarcod-ing method to provide a more accurate and comprehen-sive understanding of the cranesrsquo diets We thenevaluated the role of agricultural fields in crane protec-tion Vallisneria tuber collapses have driven thousandsof cranes to feed in agricultural fields [61 64] Howeverthe proportion of domesticated species in the diets ofthe four crane species has not previously been quanti-fied Here we determined the relative dietary import-ance of domesticated species in the feces of cranesFinally we assessed the consequences of Vallisneriatuber collapse on the dietary niche width and overlapamong crane species To do so we compared the nichebreadths and overlaps in winter 2017 and winter 2018with different Vallisneria tuber abundances Althoughwe focused on cranes at Poyang Lake our findings canprovide a reference for dietary shifts of consumers in de-graded wetlands worldwide
ResultsVariation in Vallisneria tuber abundanceWe investigated the density and biomass of Vallisneriatubers at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake The average tuber densityand biomass were 668 tubersm2 and 054 gm2 respect-ively in winter 2017 (Table 1) The density and biomassrapidly increased to 3457 tubersm2 and 411 gm2 inwinter 2018 The historical average tuber density andbiomass at three sublakes (Dahuchi Shahu and Meixihu)of Poyang Lake were 1033 (plusmn 1127 SD) tubersm2 and
326 (plusmn 368 SD) gm2 in the winters from 1999 to 2016[57] Tuber density in winter 2017 (t = 133 P = 0201)and tuber biomass in winter 2018 (t = minus 094 P = 0361)did not differ from the historical average values How-ever tuber biomass in winter 2017 was lower than thehistorical average values (t = 310 P = 0007) and tuberdensity in winter 2018 was higher than the historicalaverage values (t = minus 888 P = 0000) Therefore winter2017 represented a year with tuber collapse while winter2018 represented a year with high tuber abundance Co-inciding with the changes in tuber abundance a total of4014 individuals of Siberian Cranes White-napedCranes and Hooded Cranes were recorded in agricul-tural fields adjacent to Poyang Lake in winter 2017 [63]while none of the three crane species was recorded inagricultural fields in winter 2018 [77]
Collection statisticsIn total we collected 129 fecal samples with 60 samplesfrom winter 2017 and 69 samples from winter 2018 (Fig1 Additional file 1 Table S1) We collected 42 fecalsamples of Siberian Cranes from 2 sublakes 4 rice pad-dies and 1 lotus pond 20 fecal samples of White-napedCranes from 2 sublakes and 1 rice paddy 19 fecal sam-ples of Hooded Cranes from 2 sublakes and 1 ricepaddy and 48 fecal samples of Eurasian Cranes from 3sublakes and 6 rice paddies We also collected 38 plantspecies commonly seen at Poyang Lake to construct aDNA barcode reference library By downloading se-quences from GenBank and sequencing by ourselves weconstructed a DNA barcode reference library containing62 plant species
High-throughput DNA sequencingThe chloroplast trnL intron was selected as a barcodebecause of its high taxonomic coverage and resolutionespecially for wetland plants [70 78] The sequencing ofthe 129 fecal samples yielded a total of 9284819 readsAfter quality filtering 8502418 reads were obtained
Table 1 The density and biomass of Vallisneria tubers at threesublakes (Dahuchi Shahu and Changhuchi) of Poyang Lake inthe winters of 2017 and 2018
Winter Sublake Tuber density(tubersm2)
Tuber biomass(gm2)
2017 Dahuchi 980 097
Shahu 028 002
Changhuchi 997 062
Average 668 054
2018 Dahuchi 641 065
Shahu 897 060
Changhuchi 8832 1107
Average 3457 411
Hou et al Frontiers in Zoology (2021) 1824 Page 4 of 17
The average number of reads per sample was 65910 (plusmn26276 SD) and the average number of operational taxo-nomic units (OTUs) per sample was 43 (plusmn 15 SD) Toevaluate the sequencing and sampling adequacy individ-ual- species- and sample-based rarefaction curves werebuilt With the sample sequencing reads increased theindividual-based rarefaction curves reached plateau inmost cases (Additional file 1 Figure S1) and the species-based rarefaction curves reached plateau in the fourcrane species (Fig 2a) Therefore our sequencing depthwas roughly adequate to capture the numbers of OTUspresent The sample-based rarefaction curves for eachcrane species reached plateau in all cases as the samplingsizes increased (Fig 2b) suggesting that we acquired suf-ficient fecal samples After comparison to the local DNAreference database 9899 of the total reads wereassigned to specific plant taxa Most of the reads thatwere not assigned to specific plant taxa had low fre-quency In total we identified 29 plant taxa including23 species 4 genera and 2 families
Diet compositions of the four crane speciesWe used relative read abundance (ie the percentageof DNA belonging to each food item in each sampleRRA) which provides a more accurate view of speciesdiet than frequency of occurrence (ie the number ofsamples that contain a given food item) tosummarize dietary data [79] Twenty-two food itemswere identified in the feces of Siberian Cranes in thetwo winters including 17 species 3 genera and 2families (Fig 3 Additional file 1 Table S2) Rice(RRA = 7285) lotus (1201) and Polygonum criopo-litanum (1199) were the dominant food items inthe feces (ie proportion gt 10) in winter 2017 andVallisneria (5612) and P criopolitanum (4087)were the dominant food items in winter 2018Twenty-five food items were identified in the feces ofWhite-naped Cranes including 20 species 3 genera
and 2 families Tulipa edulis (3737) Potentilla lim-prichtii (1869) P criopolitanum (1652) andCarex spp (1075) were the dominant food items inthe feces in winter 2017 and P criopolitanum(8441) was the dominant food item in winter 2018Twenty-two food items were identified in the feces ofHooded Cranes including 18 species 3 genera and 1family P limprichtii (6536) and Carex (2739)were the dominant food items in the feces in winter2017 and P criopolitanum (8876) was the domin-ant food item in winter 2018 Twenty-two food itemswere identified in the feces of Eurasian Cranes in-cluding 17 species 3 genera and 2 families Rice(6120) and P limprichtii (2394) were the domin-ant food items in the feces in winter 2017 and Pcriopolitanum (5577) and rice (2740) were thedominant food items in winter 2018 Diet composi-tions of Siberian Cranes (Pearson correlation = 005P = 077) White-naped Cranes (Pearson correlation =030 P = 008) and Hooded Cranes (Pearson correl-ation = minus 004 P = 083) were not correlated betweenwinter 2017 and winter 2018 suggesting great inter-annual variations in food sources Diet composition ofEurasian Cranes was correlated (Pearson correlation =040 P = 002) between the two winters suggestinglimited interannual variation in food sourcesVallisneria tubers were an important food source for
the four crane species especially Siberian Cranes inwinter 2018 The proportion of Vallisneria was 5612512 850 and 734 in the feces of Siberian CranesWhite-naped Cranes Hooded Cranes and EurasianCranes respectively in winter 2018 (Fig 3 Additionalfile 1 Table S2) In winter 2017 when Vallisneria tuberabundance was low the proportion of Vallisneria wasless than 1 in the feces of the four crane speciesDomesticated species identified in crane feces included
rice and lotus The proportions of domesticated speciesin the feces of Siberian Cranes (8485) and White-
Fig 2 a Species-based and b sample-based rarefaction curves The species- and sample-based rarefaction curves built by randomly resamplingsequences and samples respectively at increasing levels of accumulation indicate whether the sequencing depth and sampling size weresufficient to characterize the diversity of dietary items The colors of the curves correspond to the crane species blue Siberian Crane purpleWhite-naped Crane green Hooded Crane and red Eurasian Crane
Hou et al Frontiers in Zoology (2021) 1824 Page 5 of 17
naped Cranes (1509) were high in winter 2017 but de-clined to 000 in winter 2018 (Fig 3 Additional file 1Table S2) Domesticated species occupied a small pro-portion in the feces of Hooded Cranes in winter 2017(130) but declined to 000 in winter 2018 The pro-portions of domesticated species in the feces of EurasianCranes were high in both winters with 6120 in winter2017 and 2740 in winter 2018
Dietary niche breadth and overlap among crane speciesThe numbers of food items dietary niche breadth andShannon-Wiener diversity values for Siberian Cranesand Eurasian Cranes in winter 2017 were similar tothose in winter 2018 (Table 2) Values of all three pa-rameters for White-naped Cranes and Hooded Craneswere higher in winter 2017 than in winter 2018 Dietaryniche breadth of the four crane species was lt 020 inboth winters indicating low niche breadthDietary niche overlap among crane species ranged
from 002 to 049 in winter 2017 but increased to 062to 099 in winter 2018 (Table 3) Nonmetric multidimen-sional scaling (NMDS) analysis also indicated overlap
increases in winter 2018 (Fig 4) No overlap values werelower than expected by chance in the two winters Theoverlap values were not higher than expected by chancein winter 2017 However all values showed a high de-gree of overlap in winter 2018 [P (Obs gt = null) lt 005]Overlap between White-naped Cranes and HoodedCranes was the highest among the four crane species inboth winters
DiscussionDiet compositionTo the best of our knowledge our study is the first toexplore crane diets using metabarcoding techniques Al-most all previous studies have relied on traditional directobservation or microhistological methods By usingmetabarcoding technology we were able to elucidatecrane diets at greater detail than previously known Theimproved knowledge might be due to the ability ofmetabarcoding to provide a more comprehensive per-spective and more details on food sources than trad-itional methods [66 67 80] Metabarcoding is able todetect a variety of smaller and softer bodied food items
Fig 3 Food items in the feces of Siberian Cranes White-naped Cranes Hooded Cranes and Eurasian Cranes in the winters of 2017 and 2018 atPoyang Lake
Table 2 Number of food items dietary niche breadth and Shannon-Wiener diversity values for Siberian Cranes White-napedCranes Hooded Cranes and Eurasian Cranes at Poyang Lake in the winters of 2017 and 2018
Number of foods Dietary niche breadth Shannon-Wiener diversity
2017 2018 2017 2018 2017 2018
Siberian Crane 16 17 005 007 091 085
White-naped Crane 18 15 020 003 169 059
Hooded Crane 19 13 005 002 096 043
Eurasian Crane 18 19 008 009 110 121
Hou et al Frontiers in Zoology (2021) 1824 Page 6 of 17
that are overlooked by microhistological methods [6780] It also enables the identification of food items at ahigher taxonomic resolution [69 81] However we ac-knowledge that our study has two limitations First weused fecal samples as they are non-invasive Neverthe-less fecal analysis is subjected to the biases caused bythe differential digestive rates of food items [82 83] Softfoods such as insect larvae tend to be under-representedin fecal samples because they are digested quickly andremain in gastro-intestinal tract for short periods whilehard foods such as mollusks tend to be over-representedbecause they break down slowly and remain in gastro-intestinal tracts longer Thus food proportions in the
feces may not correlate with food proportions ingestedby cranes In addition some materials identified in thefeces may be incidentally consumed by cranes and notreally be used as food Second our study only consideredthe plant component of cranesrsquo diets as plants were sug-gested to be their principal food types at Poyang LakeAnimal foods consumed by the four crane species butto a much lesser extent than plant foods [84ndash87] werenot considered in our studyThe Siberian Crane is regarded as the most aquatic of
all cranes using wetlands for nesting feeding and roost-ing [49 88] The high dependence of the species onaquatic habitats was suggested to make it vulnerable to
Table 3 The observed dietary niche overlap among the four crane species at Poyang Lake in the winters of 2017 and 2018 P(Obs lt = null) and P (Obs gt = null) indicate the tail probabilities for the observed index which were lower and higher than thehistogram of the simulated index respectively represents P lt 005 and represents P lt 001
Winter Observed niche overlap index P (Obs lt = null) P (Obs gt = null)
2017 Siberian vs White-naped 021 0819 0181
Siberian vs Hooded 002 0703 0297
Siberian vs Eurasian 016 0900 0100
White-naped vs Hooded 049 0952 0048
White-naped vs Eurasian 035 0895 0106
Hooded vs Eurasian 035 0919 0081
2018 Siberian vs White-naped 063 0952 0048
Siberian vs Hooded 066 0952 0048
Siberian vs Eurasian 062 0950 0049
White-naped vs Hooded 099 0997 0003
White-naped vs Eurasian 089 0987 0013
Hooded vs Eurasian 090 0998 0002
Fig 4 The results of nonmetric multidimensional scaling ordination (NMDS) based on Bray-Curtis dissimilarity The colors of the circlescorrespond to the crane species blue Siberian Crane purple White-naped Crane green Hooded Crane and red Eurasian Crane
Hou et al Frontiers in Zoology (2021) 1824 Page 7 of 17
impacts of habitat degradation and to lead its classifica-tion as Critically Endangered [49] At Poyang Lake Si-berian Cranes traditionally fed on tubers of Vallisneriathe dominant submerged macrophytes in shallow watersand mudflats [89 90] In winter 2010 due to a flood-induced Vallisneria tuber collapse they were observedforaging for P limprichtii taproots and T edulis bulbs ingrasslands for the first time [38 44] In the winters from2015 to 2017 the diets of Siberian Cranes changedagain with thousands of cranes feeding on rice seedsand lotus rhizomes in agricultural habitats [61 62 64]Our study identified 22 plant items in the feces of Siber-
ian Cranes These plant items were obtained from shallowwaters grasslands and agricultural fields Four plant itemswith yearly RRA gt 10 were identified in the feces Vallis-neria tuber P criopolitanum rhizomes rice seeds andlotus rhizomes (the vegetative portions consumed bycranes were based on our field observation) SiberianCranesrsquo diet compositions showed substantial interannualvariation When the abundance of Vallisneria tubers waslow (ie winter 2017) Siberian Cranes mainly fed on riceseeds lotus rhizomes and P criopolitanum rhizomes inagricultural fields and grasslands When the abundance ofVallisneria tubers was high (ie winter 2018) they mainlyfed on Vallisneria tubers and P criopolitanum rhizomesin shallow waters and grasslands Our study in combin-ation with previous studies indicates that the diets of Si-berian Cranes have changed greatly since winter 2010 [3844 62] They have gradually broadened their dietaryniches which has probably been driven by the degradationof submerged plants [38 44 57]Similar to Siberian Cranes a diet shift was also ob-
served in White-naped Cranes Historically White-naped Cranes only fed on submerged plants in shallowwaters and mudflats of Poyang Lake [91] Then drivenby the decline of submerged plants they primarily fed atgrasslands and mudflats [87 91] consuming Vallisneriatuber T edulis P limprichtii and Ranunculus polii [5587] Our results indicate that in addition to T edulisand P limprichtii that had previously been reported theproportions of P criopolitanum and Carex were alsohigh in the feces Hooded Cranes were suggested tomainly feed on the grassland plant P limprichtii throughdirect observation [86] Our study indicates that thegrassland plants P criopolitanum and Carex also occu-pied high proportions in the feces The dominant fooditems in the feces of White-naped Cranes and HoodedCranes varied greatly between the two winters The vari-ation in dietary composition indicated that the diversityand composition of the grassland plant communitymight have changed greatly between the two wintersThe dietary niche overlap between White-naped Cranesand Hooded Cranes was the highest among the fourcrane species aligning with their level of habitat niche
overlap which was also the highest among the species[92] The high dietary and habitat niche overlaps suggesthigh competition potential between the two cranespeciesThe Eurasian Crane is a habitat generalist It is known
to feed in a variety of habitats including grasslandsmudflats shallow waters and agricultural fields [88 92]Its diet at Poyang Lake has not been documented previ-ously Our study indicates that P criopolitanum P lim-prichtii and rice were the dominant food items in thefeces of Eurasian Cranes The number of plant itemsconsumed dietary niche breadth and Shannon-Wienermeasures were the highest among the four crane speciessupporting its generalist foraging strategy In contrast tothe three crane species above Eurasian Cranesrsquo dietcompositions were similar between the two winters Riceand grassland plants occupied high proportions in thefeces in both winters
Important food resources for cranesVallisneria tubers have been acknowledged as importantfood sources for cranes especially Siberian Cranes andWhite-naped Cranes [55 56] Our study confirms theimportance of Vallisneria tubers in the diet of SiberianCranes It was the most common food source in thefeces of Siberian Cranes in winter 2018 when its abun-dance was high Vallisneria tuber proportions in thefeces of the three other crane species were not as high asthat in the feces of Siberian Cranes but it still occupied5ndash9 of the total foods in winter 2018To our surprise P criopolitanum was found to be the
first dominant food in the feces of White-naped CranesHooded Cranes and Eurasian Cranes and the seconddominant food in the feces of Siberian Cranes in winter2018 In winter 2017 the proportions of P criopolita-num were lt 17 in the feces of Siberian Cranes andWhite-naped Cranes and were lt 3 in the feces ofHooded Cranes and Eurasian Cranes The higher pro-portions of P criopolitanum in winter 2018 than winter2017 indicate that the abundance and availability of Pcriopolitanum was much higher in winter 2018 P crio-politanum is an annual grassland plant It is sensitive tohydrological conditions and can only survive in a rela-tively narrow range of water depths and percent time in-undated [93] Due to the sensitivity its distributionrange and abundance showed high interannual variation[94 95] Thus it is likely that the abundance of P crio-politanum was higher in winter 2018 than in winter2017The importance of P criopolitanum as a food source
for the cranes at Poyang Lake has not been documentedpreviously It has only been suggested to be a major foodsource of Hooded Cranes at Shengjin Lake another lakein the middle and lower Yangtze River floodplain [86
Hou et al Frontiers in Zoology (2021) 1824 Page 8 of 17
96] P criopolitanum is a dominant wetland plant at Po-yang Lake [93 97] The coverage percentage of this plantis the third highest among grassland plants accountingfor 12 of the total areas of grasslands [97] This plantspecies is usually distributed at elevations of 9ndash12mabove sea level near mudflats and shallow waters [97]which are frequently used by cranes Its rhizome with ashallow burial depth is high in energy and nutritionalcontent and low in crude fibre [38 86] All these charac-teristics make P criopolitanum rhizome a potentially im-portant food source for cranes We frequently observedcranes dig and feed on rhizomes of P criopolitanum andleave aboveground parts in the fields Therefore we be-lieve that P criopolitanum was used as food not inci-dentally consumed by cranes However we need to becaution that high proportion of P criopolitanum in thefeces does not mean high proportion in the diets Morework on the importance of P criopolitanum in the dietsof cranes is needed
The roles of agricultural fields in crane protectionWith the loss and degradation of natural wetlands theroles of artificial wetlands in waterbird protection havebeen under intensive debate [98 99] Although agricul-tural fields cannot completely replace the function ofnatural wetlands as waterbird habitats [100ndash102] agri-cultural fields might provide alternative or complemen-tary habitats for waterbirds in all life stages [38 103104] The proportions of domesticated species in thefeces of Siberian Cranes varied considerably betweenyears In winter 2017 when the abundance of Vallisneriatubers was low the proportion of Vallisneria in SiberianCranesrsquo feces was only 095 Siberian Cranes may nothave found enough foods in the shallow waters andmudflats and were forced to search for rice seeds andlotus rhizomes in agricultural fields In winter 2018when the abundance of Vallisneria tubers reboundedSiberian Cranes moved back to shallow waters and mud-flats and a large proportion of their traditional food Val-lisneria tubers and also P criopolitanum were detectedin the feces A similar abandonment of tuber foragingoccurred following the tuber collapse in 2010 and thenresumed in the next winter when tubers rebounded in2011 [38 44] The diet and habitat changes indicate thatagricultural habitats were important refuges for SiberianCranes providing important food resources for them tosurvive through winters with few Vallisneria tubersThey also indicate that Siberian Cranes prefer naturalwetlands to agricultural fields [38]Similar to Siberian Cranes White-naped Cranes and
Hooded Cranes consumed rice seeds and lotus rhizomesin winter 2017 but did not consume any in winter 2018The changes in the proportions suggest that the abun-dance of food resources in natural wetlands might have
been low in winter 2017 The deficient food resourcesmight have been Vallisneria tubers but might also havebeen grassland plants the primary food sources of thesetwo crane species Under natural food shortage condi-tions (ie winter 2017) White-naped Cranes andHooded Cranes utilized agricultural fields When theabundance of Vallisneria tubers and probably grasslandplants were high (ie winter 2018) the two crane speciesreturned to their natural habitats Therefore for White-naped Cranes and Hooded Cranes agricultural fieldswere important alternative foraging habitats used asbuffers against starvation during periods of natural foodshortages The return of the two crane species to naturalwetlands in winter 2018 further emphasizes the superiorquality of natural habitats over agricultural fields as for-aging habitats for cranes [38]The proportions of rice seeds in the feces of Eur-
asian Cranes were high in the two winters thoughthe proportion in winter 2017 was twice as high asthat in winter 2018 Therefore agricultural fields ap-peared to be consistently important as foraging habi-tats for Eurasian Cranes The frequent use ofagricultural fields by Eurasian Cranes has been docu-mented widely [49 105 106]Domesticated species generally contain high energy
and nutritional content and are of superior quality com-pared to natural plants [86 104] Farmland foraging hasbeen suggested to contribute to increase in the abun-dance of some goose [107] egret [108] and wader [109]populations in Japan Korea Europe and North Amer-ica However cranes generally avoid human disturbancewhen select foraging sites [110 111] Human disturb-ance in agricultural fields around Poyang Lake is consid-erably high due to intensive agricultural activity [112]High human disturbance was suggested to be a majorreason that wintering geese were almost confined withinnatural wetlands and hesitated to exploit the riches ofthe modern agricultural habitats at Poyang Lake [112113] High human disturbance also resulted in SiberianCranes spending twice as much time alerting in agricul-tural fields than they did in natural habitats [114] Thehigh level of human disturbance in agricultural fieldsmight contribute to the preference for natural wetlandsby Siberian Cranes White-naped Cranes and HoodedCranes at Poyang Lake Moreover although agriculturefields contain energy-rich crops wetlands contain di-verse food resources especially protein-rich inverte-brates which might better satisfy dietary needs of cranes[110 115] Taken together security and diverse food re-sources might have attracted cranes to select wetlands asforaging sitesSubmerged macrophytes at Poyang Lake have de-
graded in recent decades [57ndash60] and Vallisneria tuberdisappearance is expected to occur more frequently
Hou et al Frontiers in Zoology (2021) 1824 Page 9 of 17
under future climate change [57 60] Cranes may in-creasingly depend upon agricultural fields around Po-yang Lake which may in turn cause significant cropdamage and economic losses This is especially true onagricultural fields surrounding protected wetlands wherewaterbirds concentrate in high numbers [116 117] AtPoyang Lake cranes mainly fed on post-harvest remainsof rice seeds and lotus rhizomes thus crop damage andeconomic loss were limited However we observed somefarmers scared cranes away from their agricultural fieldsThere were more than 14 million free-ranging poultryraised at agricultural fields around Poyang Lake [118]where they gleaned waste grain residues in stubble fieldsCrane feeding would reduce the food abundance of thefree-ranging domestic poultry thus farmers drovecranes away To reduce human disturbance environ-mental education is needed because farmers are morelikely to protect cranes if they understand the endan-gered degree and the ecological significance of cranes
Effects of tuber collapse on dietary niche breadth andoverlap among crane speciesVallisneria tuber is an important food source for Siber-ian Cranes (89 90 this study) thus variation in itsabundance might influence Siberian Cranesrsquo dietaryniche breadth The numbers of food items dietary nichebreadth and Shannon-Wiener measures of SiberianCranes were similar between winter 2017 and winter2018 This is inconsistent with the contention that dur-ing seasons with a low abundance of dominant food thedietary niche may broaden as consumers relying on in-sufficient preferred food items are forced to add lessprofitable resources to their diets [26 119 120] In win-ter 2017 with low Vallisneria tuber abundance SiberianCranes left shallow waters and mudflats and moved torice paddies and lotus ponds [61 62 64] Their foodschanged from Vallisneria tuber to rice seeds and lotusrhizomes almost completely As the energy protein andfat contents of crops are generally as good as or superiorto natural foods and are often present in agriculturalfields in far greater abundance [86 104] Siberian Cranesmay gain enough energy through feeding on crops anddo not need to add many other food sources Moreoverthe Siberian Crane is a dietary specialist [89 90] PoyangLake has a relatively simple plant community compos-ition with Vallisneria and Hydrilla verticillate being thedominant submerged macrophytes [59] and Carex beingthe dominant grassland plants [121] Therefore theremay be few options for Siberian Cranes which mightlead to the failure of this species to widen its dietaryniche breadthIn contrast to Siberian Cranes the numbers of food
items dietary niche breadths and Shannon-Wiener mea-sures of White-naped Cranes and Hooded Cranes were
higher in winter 2017 than in winter 2018 P criopolita-num was the predominant food item in the feces of thetwo crane species in winter 2018 and the proportions ofVallisneria tubers were also high The abundances ofVallisneria tubers and probably P criopolitanum at Po-yang Lake declined greatly in winter 2017 When thereis a short-fall in food availability animals may optimizeforaging behavior and increase their dietary breath [122123] The short-fall in the abundances of P criopolita-num and Vallisneria tubers in winter 2017 might havedriven the two crane species to add less profitable grass-land plants in their diets Although the two cranes alsoconsumed high-quality rice seeds and lotus rhizomes inwinter 2017 their proportions were low in the feces andthese foods might be unable to meet the energy require-ments of the two crane species The niche expansion ofthe two crane species during periods of preferred foodshortage was in agreement with the basic prediction ofoptimal foraging theory [22 23] It is also consistentwith a commonly observed fact that energy-maximizingmodels work better when animals food-limited [124]The dietary niche overlaps among the crane species
were low in winter 2017 with the four crane species di-verging in their principal food items However in winter2018 the niche overlaps were significantly higher thanexpected by chance They had similar diet compositionswith P criopolitanum rhizomes and Vallisneria tubersbeing the principal food items in the feces The in-creased overlap might have been caused by the highabundances of the two plant species in winter 2018which allowed the four crane species to share the samefood items with low interspecific competition Our studyis consistent with previous research that suggested thatamong-species dietary niches tended to overlap moreduring seasons of high food abundance [32 33 125126] Greater resource availability could promote an in-crease in species utilization of similar resources and re-duce competitive interactions between species [32 34126] By contrast during seasons with low food abun-dance species may partition limited food resources todecrease interspecific competition leading to a decreasein niche overlap [126 127] It has been suggested thatdiet partitioning facilitates the coexistence of manyclosely related species [7 128] The interannual variationin niche overlap observed in our study suggests that dietpartitioning may be restricted to specific temporal casesIn seasons or years with abundant food sources inter-specific competition may be relaxed and niche overlapcould be high
ConclusionsIn summary by using metabarcoding technology ourstudy gained new insights into cranesrsquo diets that are es-sential for informing habitat management to improve
Hou et al Frontiers in Zoology (2021) 1824 Page 10 of 17
the availability of foraging resources Vallisneria tuberswas confirmed as an important food source of cranesSurprisingly P criopolitanum also occupied high pro-portions in the feces of cranes and may also be import-ant Our study emphasizes the superior quality ofnatural wetlands compared to agricultural fields as for-aging habitats for cranes The recent degradation of sub-merged macrophytes at Poyang Lake potentiallythreatens the survival of cranes especially SiberianCranes Therefore it is urgent to reverse the broad de-cline of submerged macrophytes and provide importantforaging habitats for cranes during years of tuber col-lapses The abundances of Vallisneria tubers and prob-ably P criopolitanum at Poyang Lake greatly influencedthe dietary niche width and overlaps among crane spe-cies Cranes exhibited low dietary niche overlap duringperiods of preferred food shortage but significantly highoverlap during periods when preferred foods were plen-tiful While high niche overlap is often interpreted as in-tense interspecific competition our study highlights theimportance of taking food abundance into account
Materials and methodsStudy areasPoyang Lake the largest freshwater lake in China is oneof the most important wintering areas for waterbirdsalong the East Asian-Australasian Flyway (Fig 1) [51129] It is characterized by dramatic seasonal hydro-logical fluctuations with an average change of 924 mbetween the summer high and winter low water levels[130] During summer the inundation area is gt 2500km2 [131] In autumn and winter the water level dropscausing the inundation area to shrink to lt 1000 km2
[131] Poyang Lake contains 102 sublakes which are in-undated and integrated with the main body of PoyangLake during summer and are isolated during winter[132] As the water level drops the sublakes appear andlarge areas of grasslands mudflats and shallow waterareas are exposed providing winter foraging and roost-ing areas for approximately 420000 waterbirds from ap-proximately 111 species [133 134]
Vallisneria tuber surveyThe density and biomass of Vallisneria tubers were sur-veyed at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake in late October and earlyNovember of 2017 and 2018 before large numbers ofwintering waterbirds arrived We set 76 183 and 36sampling plots uniformly located at Dahuchi Shahu andChanghuchi respectively Vallisneria tubers were col-lected from two quadrats in each plot with a locallymade steel grab sampler The sampler has two long
handles At the end of the handles there are two scoopsfacing each other The sampler was inserted into thesubstrate and the scoops brought together to collectabout 15 cm long times 13 cm wide times 30 cm high sample ofsubstrate Tubers were subsequently cleaned andcounted The dry weight of tubers in each quadrat wasweighed after oven-drying at 80 degC until constant weightThe tuber density and biomass in each plot was calcu-lated by dividing total tuber number and tuber massacross two quadrats by quadrat area respectively Weused t-test to explore whether the tuber density and bio-mass in the winters of 2017 and 2018 significantly devi-ated from the historical average values in the wintersfrom 1999 to 2016 [57] The historical tuber density andbiomass data were collected by the Poyang Lake Na-tional Nature Reserve at three sublakes (Dahuchi Shahuand Meixihu) [57] R 360 was used to do the t-testanalyses
Fecal sample collectionFecal samples were collected at Poyang Lake in the win-ters of 2017 and 2018 We first investigated the distribu-tions of cranes When we saw foraging cranes we waiteduntil they left to avoid disturbing them Soon after theyleft we went to the foraging sites to collect fresh feces(ie feces with wet surface) with sterilized tweezers Tominimize the probability of multiple samples from thesame individual all collected samples were separated byat least 5 m We stored the samples in liquid nitrogen inthe field and transferred them to a minus 80 degC refrigeratorfor long-term storage in the laboratory To ensure thatthe sources of the feces were known we tried to collectsamples from monospecific flocks For samples collectedfrom mixed-species flocks we used DNA analysis toidentify the species that deposited the fecal samplesPlants were also collected in the field to construct aDNA barcoding reference database
Species identificationBecause of the similar morphologies of the feces of thefour crane species we used DNA analysis to identify thespecies that deposited the fecal samples collected frommixed-species flocks For the genetic species assignmentof each fecal sample we scraped the external surfacewhich contain DNA from the host species as a result ofthe sloughing of cells from the digestive tract DNA wasextracted with the QIAamp Power Fecal DNA Kit (Qia-gen USA) according to the manufacturerrsquos guidelinesFor each round of DNA extraction negative controls(ie extraction without feces) were included to monitorfor possible contamination We amplified the mitochon-drial DNA D-loop fragment with the primer pair DL02F
Hou et al Frontiers in Zoology (2021) 1824 Page 11 of 17
(5prime-3prime GCGGCCCGAAAAGCCGCTG) and DL02R (5prime-3prime GCCCTGACATAGGAACCAGAGGCGC) [135]PCR amplifications were carried out in a total volume of25 μl containing 125 μl PCR mix (Tiangen BeijingChina) 1 μl DNA 1 μl each primer and 95 μl H2O Thereaction conditions were as follows denaturation at94 degC for 4 min followed by 34 cycles at 94 degC for 48 s56 degC for 48 s and 72 degC for 1 min and a final 10 min at72 degC We checked for the presence of a PCR product ofsuitable length by electrophoresis on a 1 agarose gelSequencing was carried out on an ABI 3730xl analyzerby Sangon Biotech (Shanghai China) The complete se-quences were assembled using Seqman II (DNASTAR)and compared visually to the original chromatograms toavoid reading errors The taxonomic assignment wasconducted by searching against the nucleotide databaseof GenBank using Web BLAST
Construction of a barcode reference databaseWe downloaded the trnL sequences of 48 plant speciescommonly seen at Poyang Lake from GenBank For theplant species not included in GenBank we sequencedtrnL by ourselves Total DNA was extracted using thehexadecyltrimethylammonium bromide (CTAB) proto-col We used primer pair c (5prime-3prime CGAAATCGGTAGACGCTACG) and h (5prime-3prime CCATTGAGTCTCTGCACCTATC) to amplify an approximately 150 bp regionof the chloroplast trnL intron [136] This primer pairwas chosen as it can amplify DNA from a wide range ofwetland plants [70] PCR amplifications were carried outin a total volume of 25 μl containing 125 μl PCR mix(Tiangen Beijing China) 1 μl DNA 1 μl each primerand 95 μl H2O The reaction conditions were as followsdenaturation at 94 degC for 4 min followed by 35 cycles at94 degC for 30 s 56 degC for 30 s and 72 degC for 45 s and afinal 10 min at 72 degC We used electrophoresis to checkthe PCR products ABI 3730xl analyzer to perform se-quencing and Seqman II to assemble the sequences
DNA extraction and high-throughput sequencing of fecalsamplesTo uncover plant component consumed by cranes amodified CTAB protocol [70] was used to extract theDNA from approximately 200 mg of the interior of eachfecal sample DNA extraction was conducted in a dedi-cated laboratory for analyses of DNA from samples withlow DNA quality For each batch of DNA extractionnegative controls were included to monitor for possiblecontamination The primers c and h with a 6 bp tagadded to 5prime ends of each primer were used to amplifytrnL gene The PCR amplification procedure was thesame as barcode database construction For each samplea unique combination of tags in the forward and reverseprimers was used so that sample-specific data can be
recovered after sequencing Three PCR replicates wereperformed for each sample to reduce amplification biasThe three replicates were pooled and the DNA concen-tration and purity were monitored on a 1 agarose gelOnly samples with a clear band were processed furtherSamples were purified using the SanPrep Column PCRProduct Purification Kit (Sangon Biotech ShanghaiChina) The purified PCR products were pooled takinginto account of DNA concentration to obtain an ap-proximately equal amount of amplicon DNA from eachfecal sample The pool of individually tagged ampliconswas used to prepare a sequencing library using the Tru-Seq DNA PCR-Free Sample Preparation Kit (IlluminaUSA) following manufacturerrsquos recommendations Thelibrary was sequenced on an Illumina HiSeq 2500 plat-form and 250 bp paired-end reads were generated Li-brary preparation and sequencing were conducted byNovogene (Beijing China)
Sequencing data analysisAfter high-throughput sequencing the paired-end readswere demultiplexed into sample-specific files based ontheir unique tags The tags and primer sequences weresubsequently trimmed The paired-end reads weremerged using FLASH 127 [137] and then filtered usingQIIME 170 [138] Sequences were dereplicated andclustered to OTUs using USEARCH 110667 [139] atthe similarity threshold of 97 All OTUs were assignedto taxonomic units by referring to the local plant data-base using blast 2231+ with thresholds of identity gt97 and e-value lt 10 eminus 50 If an OTU matched two ormore taxa it was assigned to a higher taxonomic levelthat included all taxaTo evaluate the sufficiency of sequencing depth and
sampling size individual- species- and sample-basedrarefaction curves were built using EstimateS 82[140] The curves were computed by randomly resam-pling sequences or samples and plotting these againstthe numbers of OTUs The read abundance data wereconverted to RRA of each food item using the follow-ing equation [79]
RRA frac14 1S
XS
Kfrac141
nikPTifrac141nik
100
where nik is the number of sequences of food item i insample k T is the total number of food items and S isthe sampling size
Dietary niche breadth and overlap among crane speciesTo explore whether the cranesrsquo diets showed substantialinterannual variation we calculated Pearson correlationsbetween diet compositions of each crane species in win-ter 2017 and winter 2018 The proportion of each food
Hou et al Frontiers in Zoology (2021) 1824 Page 12 of 17
item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

(Continued from previous page)
Conclusions Our study emphasizes the superior quality of natural wetlands compared to agricultural fields asforaging habitats for cranes To provide safer and better foraging areas for cranes it is urgent to restore thesubmerged plants at Poyang Lake While high dietary niche overlap is often interpreted as intense interspecificcompetition our study highlights the importance of taking food abundance into account
Keywords Agricultural fields Food shortage Hooded crane Siberian crane Vallisneria tuber White-naped crane
BackgroundInterspecific competition is an important mechanism instructuring ecological communities [1 2] Competitionmay lead to resource (food habitat time) partitioningand facilitate the coexistence of sympatric species [1 34] Diet partitioning has been suggested to be the pri-mary mechanism of coexistence of many insects [5] fish[6] snakes [7] birds [8] and mammals [9]Many ecosystems are characterized by temporal varia-
tions in resource availability which may subsequently in-fluence dietary niche of birds [10ndash12] Migratory birdsmay also encounter strikingly different feeding condi-tions along their flyways [13 14] Therefore it is critic-ally important that birds are able to adjust their diets toexplore a wide range of food resources [11 15] Numer-ous examples of dietary flexibility exist Dunlins (Cali-dris alpine) [11] Semipalmated Sandpiper (C pusilla)[16] Hooded Crane (Grus monacha) [17] and Black-necked Crane (G nigricollis) [18] have all varied their di-ets with changes in prey availability In contrast strongdependence on a few prey types can make birds vulner-able to the reduction of prey abundance as has been thecase for Red Knot (C canutus) feeding on HorseshoeCrab (Limulus polyphemus) eggs [19]Optimal foraging theory is often used to predict the for-
aging decisions of animals [20 21] According to the the-ory when resources are abundant species shouldconsume the most profitable food items with high nutri-tion and accessibility while ignoring less profitable foodsWhen preferred food items become depleted species addless profitable foods to their diet [22 23] Great Knots (Ctenuirostris) for example changed their diet compositionand prey selection to include prey items that were physic-ally more difficult to digest after a severe decline in foodabundance and quality [24] Optimal foraging theory pre-dicts a reduction in dietary niche breadth during seasonsof high food abundance and an increasing trend duringseasons of low food abundance This pattern of nichewidth variation has been observed across a range of taxaincluding birds [12 25] mammals [26 27] fish [28 29]and invertebrates [30 31] The theory also predicts an in-crease in dietary niche overlap between species when re-sources are abundant [7 32 33] The increased overlapmay be related to the reduced competitive interaction be-tween species due to great resource availability For
example there was an absence of dietary niche partition-ing among shorebird species at Delaware Bay USA whenthe density of their primary prey Horseshoe Crab eggswas high [34] Extensive degree of dietary niche overlaphas also been revealed among seabird species breeding onthe Argentinean Patagonian coast with superabundance ofpelagic fish [35]Understanding how dietary niche changes for any spe-
cies as fluctuations in food abundance occurs is usefulfor designating management strategies to preserve bio-diversity and ecosystem services [32 36] However mostresearch is focused on the influences of seasonal vari-ation in food abundance on dietary niches [10 11 3237] Due to global climate change and increased humandisturbance interannual variation in resource abundancehas increased in frequency and intensity promulgatingfood scarcity at higher rates in recent decades [38ndash40]For example floods that negatively affect submergedplant growth and even lead to mass death of plants havebecome more frequent at Poyang Lake China [41ndash43]which may threaten the survival of birds that feed onthem [38 44 45] Seasonal variation in food abundanceoccurs regularly Animals have evolved strategies to copewith the predictable environmental variation [46] Inter-annual variation in food abundance occurs less fre-quently although its frequency and intensity haverecently increased [38ndash40] Without sufficient evolution-ary time animals may not adapt well to the interannualvariation thus it may pose a challenge for many taxa[19 47 48]Cranes are among the most threatened families of
birds [49] Poyang Lake which is located on the south-ern bank of the Yangtze River China is an importantwintering ground for cranes in East Asia (Fig 1) It sup-ports approximately 98 of the estimated global popula-tion of IUCN Critically Endangered Siberian Crane(Leucogeranus leucogeranus) approximately 18 of theestimated global population of Vulnerable White-napedCrane (Grus vipio) approximately 3 of the estimatedglobal population of Vulnerable Hooded Crane and ap-proximately half of the Chinese population of EurasianCrane (G grus) [50ndash52] With the degradation of lakesin the middle and lower Yangtze River floodplain Po-yang Lake is playing an increasingly important role incrane protection especially for the Siberian Crane [53
Hou et al Frontiers in Zoology (2021) 1824 Page 2 of 17
54] It has been suggested that other than Poyang Lakethere are no alternative wintering sites for SiberianCranes remaining in the Yangtze River floodplain [49]Tubers of Vallisneria spp the dominant submerged
macrophytes at Poyang Lake is an important food sourcefor cranes [38 55 56] However submerged macrophyteshave degraded seriously in recent decades [57ndash60] Theywere widely distributed throughout Poyang Lake in previ-ous years but were restricted to small areas in 2012 [58]The Vallisneria tuber density and biomass at three sub-lakes (Dahuchi Shahu and Meixihu) of Poyang Lake de-clined greatly in the winters from 1999 to 2017 [57 60]The frequency and range of Vallisneria tuber collapsehave also increased because of frequently occurring sum-mer flood and autumn drought extensive aquaculture de-clining water quality and other factors [38 57 60]Tuber collapse has led to dietary and foraging habitat
shifts of cranes For example the tuber collapses in thewinters of 2015 and 2016 drove thousands of SiberianCranes for the first time to switch from foraging inshallow waters they typically used to paddy fields and
lotus ponds [57 61] Their main foods changed fromVallisneria tubers to rice (Oryza sativa) seeds and lotus(Nelumbo nucifera) rhizomes [62] Many White-napedCranes Hooded Cranes and Eurasian Cranes also movedfrom natural wetlands to forage in paddy fields [63 64]Diet shifts may influence the dietary niche width andoverlap between crane species and subsequently thecompetition level and fitness of cranes Given the highlyendangered status of cranes and the importance of Po-yang Lake in crane protection it is important to under-stand the consequences of Vallisneria tuber collapse ondiets and competition levels among crane species so thateffective protection measures can be implementedTraditional dietary analysis methods include direct ob-
servation of foraging animals and microscopic examin-ation of feces or gut contents While these approacheshave been suggested to be useful in some cases they alsoexhibit methodological limitations [65ndash67] The maindrawbacks are that the methods can be inaccurate andlabor-intensive [65 68] Recently the emergence ofDNA metabarcoding has provided new perspectives for
Fig 1 Maps showing the three Vallisneria tuber survey sublakes (Dahuchi Shahu and Changhuchi) and the sampling sites of crane feces atPoyang Lake Circles and squares represent sampling sites in the winters of 2017 and 2018 respectively The colors of the circles and squarescorrespond to the crane species red Siberian Crane blue White-naped Crane green Hooded Crane and purple Eurasian Crane The base mapshows the inundation area of Poyang Lake in winter 2008 when the water level was similar to the historical average water level
Hou et al Frontiers in Zoology (2021) 1824 Page 3 of 17
diet analysis [65] This method is based on amplifyingand high-throughput sequencing a standardized DNAregion from stomach contents or feces and subsequentlycomparing it to a reference database for identification ofthe consumed species [65] Compared to traditionalmethods DNA metabarcoding generally provides highertaxonomic resolution identifies more food items andcan simultaneously analyze a larger number of samples[69ndash71] After its first application in assessment of thediets of Australian Fur Seals (Arctocephalus pusillus dor-iferus) [72] metabarcoding has been used successfully tostudy diets of herbivores [9 73] carnivores [69 74] andomnivores [75 76]In this study we used metabarcoding technology to
explore the consequences of Vallisneria tuber collapseon the diet compositions niche breadths and nicheoverlaps of the four crane species at Poyang Lake Fecalsamples were collected in winter 2017 (ie 20172018 ayear with Vallisneria tuber collapse) and winter 2018(ie 20182019 a year with high Vallisneria tuber abun-dance see Results for details) We first explored the di-ets of the four crane species and their interannualvariations Most previous dietary studies of cranes werebased on traditional direct observation or microscopicexamination Here we hoped to apply the metabarcod-ing method to provide a more accurate and comprehen-sive understanding of the cranesrsquo diets We thenevaluated the role of agricultural fields in crane protec-tion Vallisneria tuber collapses have driven thousandsof cranes to feed in agricultural fields [61 64] Howeverthe proportion of domesticated species in the diets ofthe four crane species has not previously been quanti-fied Here we determined the relative dietary import-ance of domesticated species in the feces of cranesFinally we assessed the consequences of Vallisneriatuber collapse on the dietary niche width and overlapamong crane species To do so we compared the nichebreadths and overlaps in winter 2017 and winter 2018with different Vallisneria tuber abundances Althoughwe focused on cranes at Poyang Lake our findings canprovide a reference for dietary shifts of consumers in de-graded wetlands worldwide
ResultsVariation in Vallisneria tuber abundanceWe investigated the density and biomass of Vallisneriatubers at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake The average tuber densityand biomass were 668 tubersm2 and 054 gm2 respect-ively in winter 2017 (Table 1) The density and biomassrapidly increased to 3457 tubersm2 and 411 gm2 inwinter 2018 The historical average tuber density andbiomass at three sublakes (Dahuchi Shahu and Meixihu)of Poyang Lake were 1033 (plusmn 1127 SD) tubersm2 and
326 (plusmn 368 SD) gm2 in the winters from 1999 to 2016[57] Tuber density in winter 2017 (t = 133 P = 0201)and tuber biomass in winter 2018 (t = minus 094 P = 0361)did not differ from the historical average values How-ever tuber biomass in winter 2017 was lower than thehistorical average values (t = 310 P = 0007) and tuberdensity in winter 2018 was higher than the historicalaverage values (t = minus 888 P = 0000) Therefore winter2017 represented a year with tuber collapse while winter2018 represented a year with high tuber abundance Co-inciding with the changes in tuber abundance a total of4014 individuals of Siberian Cranes White-napedCranes and Hooded Cranes were recorded in agricul-tural fields adjacent to Poyang Lake in winter 2017 [63]while none of the three crane species was recorded inagricultural fields in winter 2018 [77]
Collection statisticsIn total we collected 129 fecal samples with 60 samplesfrom winter 2017 and 69 samples from winter 2018 (Fig1 Additional file 1 Table S1) We collected 42 fecalsamples of Siberian Cranes from 2 sublakes 4 rice pad-dies and 1 lotus pond 20 fecal samples of White-napedCranes from 2 sublakes and 1 rice paddy 19 fecal sam-ples of Hooded Cranes from 2 sublakes and 1 ricepaddy and 48 fecal samples of Eurasian Cranes from 3sublakes and 6 rice paddies We also collected 38 plantspecies commonly seen at Poyang Lake to construct aDNA barcode reference library By downloading se-quences from GenBank and sequencing by ourselves weconstructed a DNA barcode reference library containing62 plant species
High-throughput DNA sequencingThe chloroplast trnL intron was selected as a barcodebecause of its high taxonomic coverage and resolutionespecially for wetland plants [70 78] The sequencing ofthe 129 fecal samples yielded a total of 9284819 readsAfter quality filtering 8502418 reads were obtained
Table 1 The density and biomass of Vallisneria tubers at threesublakes (Dahuchi Shahu and Changhuchi) of Poyang Lake inthe winters of 2017 and 2018
Winter Sublake Tuber density(tubersm2)
Tuber biomass(gm2)
2017 Dahuchi 980 097
Shahu 028 002
Changhuchi 997 062
Average 668 054
2018 Dahuchi 641 065
Shahu 897 060
Changhuchi 8832 1107
Average 3457 411
Hou et al Frontiers in Zoology (2021) 1824 Page 4 of 17
The average number of reads per sample was 65910 (plusmn26276 SD) and the average number of operational taxo-nomic units (OTUs) per sample was 43 (plusmn 15 SD) Toevaluate the sequencing and sampling adequacy individ-ual- species- and sample-based rarefaction curves werebuilt With the sample sequencing reads increased theindividual-based rarefaction curves reached plateau inmost cases (Additional file 1 Figure S1) and the species-based rarefaction curves reached plateau in the fourcrane species (Fig 2a) Therefore our sequencing depthwas roughly adequate to capture the numbers of OTUspresent The sample-based rarefaction curves for eachcrane species reached plateau in all cases as the samplingsizes increased (Fig 2b) suggesting that we acquired suf-ficient fecal samples After comparison to the local DNAreference database 9899 of the total reads wereassigned to specific plant taxa Most of the reads thatwere not assigned to specific plant taxa had low fre-quency In total we identified 29 plant taxa including23 species 4 genera and 2 families
Diet compositions of the four crane speciesWe used relative read abundance (ie the percentageof DNA belonging to each food item in each sampleRRA) which provides a more accurate view of speciesdiet than frequency of occurrence (ie the number ofsamples that contain a given food item) tosummarize dietary data [79] Twenty-two food itemswere identified in the feces of Siberian Cranes in thetwo winters including 17 species 3 genera and 2families (Fig 3 Additional file 1 Table S2) Rice(RRA = 7285) lotus (1201) and Polygonum criopo-litanum (1199) were the dominant food items inthe feces (ie proportion gt 10) in winter 2017 andVallisneria (5612) and P criopolitanum (4087)were the dominant food items in winter 2018Twenty-five food items were identified in the feces ofWhite-naped Cranes including 20 species 3 genera
and 2 families Tulipa edulis (3737) Potentilla lim-prichtii (1869) P criopolitanum (1652) andCarex spp (1075) were the dominant food items inthe feces in winter 2017 and P criopolitanum(8441) was the dominant food item in winter 2018Twenty-two food items were identified in the feces ofHooded Cranes including 18 species 3 genera and 1family P limprichtii (6536) and Carex (2739)were the dominant food items in the feces in winter2017 and P criopolitanum (8876) was the domin-ant food item in winter 2018 Twenty-two food itemswere identified in the feces of Eurasian Cranes in-cluding 17 species 3 genera and 2 families Rice(6120) and P limprichtii (2394) were the domin-ant food items in the feces in winter 2017 and Pcriopolitanum (5577) and rice (2740) were thedominant food items in winter 2018 Diet composi-tions of Siberian Cranes (Pearson correlation = 005P = 077) White-naped Cranes (Pearson correlation =030 P = 008) and Hooded Cranes (Pearson correl-ation = minus 004 P = 083) were not correlated betweenwinter 2017 and winter 2018 suggesting great inter-annual variations in food sources Diet composition ofEurasian Cranes was correlated (Pearson correlation =040 P = 002) between the two winters suggestinglimited interannual variation in food sourcesVallisneria tubers were an important food source for
the four crane species especially Siberian Cranes inwinter 2018 The proportion of Vallisneria was 5612512 850 and 734 in the feces of Siberian CranesWhite-naped Cranes Hooded Cranes and EurasianCranes respectively in winter 2018 (Fig 3 Additionalfile 1 Table S2) In winter 2017 when Vallisneria tuberabundance was low the proportion of Vallisneria wasless than 1 in the feces of the four crane speciesDomesticated species identified in crane feces included
rice and lotus The proportions of domesticated speciesin the feces of Siberian Cranes (8485) and White-
Fig 2 a Species-based and b sample-based rarefaction curves The species- and sample-based rarefaction curves built by randomly resamplingsequences and samples respectively at increasing levels of accumulation indicate whether the sequencing depth and sampling size weresufficient to characterize the diversity of dietary items The colors of the curves correspond to the crane species blue Siberian Crane purpleWhite-naped Crane green Hooded Crane and red Eurasian Crane
Hou et al Frontiers in Zoology (2021) 1824 Page 5 of 17
naped Cranes (1509) were high in winter 2017 but de-clined to 000 in winter 2018 (Fig 3 Additional file 1Table S2) Domesticated species occupied a small pro-portion in the feces of Hooded Cranes in winter 2017(130) but declined to 000 in winter 2018 The pro-portions of domesticated species in the feces of EurasianCranes were high in both winters with 6120 in winter2017 and 2740 in winter 2018
Dietary niche breadth and overlap among crane speciesThe numbers of food items dietary niche breadth andShannon-Wiener diversity values for Siberian Cranesand Eurasian Cranes in winter 2017 were similar tothose in winter 2018 (Table 2) Values of all three pa-rameters for White-naped Cranes and Hooded Craneswere higher in winter 2017 than in winter 2018 Dietaryniche breadth of the four crane species was lt 020 inboth winters indicating low niche breadthDietary niche overlap among crane species ranged
from 002 to 049 in winter 2017 but increased to 062to 099 in winter 2018 (Table 3) Nonmetric multidimen-sional scaling (NMDS) analysis also indicated overlap
increases in winter 2018 (Fig 4) No overlap values werelower than expected by chance in the two winters Theoverlap values were not higher than expected by chancein winter 2017 However all values showed a high de-gree of overlap in winter 2018 [P (Obs gt = null) lt 005]Overlap between White-naped Cranes and HoodedCranes was the highest among the four crane species inboth winters
DiscussionDiet compositionTo the best of our knowledge our study is the first toexplore crane diets using metabarcoding techniques Al-most all previous studies have relied on traditional directobservation or microhistological methods By usingmetabarcoding technology we were able to elucidatecrane diets at greater detail than previously known Theimproved knowledge might be due to the ability ofmetabarcoding to provide a more comprehensive per-spective and more details on food sources than trad-itional methods [66 67 80] Metabarcoding is able todetect a variety of smaller and softer bodied food items
Fig 3 Food items in the feces of Siberian Cranes White-naped Cranes Hooded Cranes and Eurasian Cranes in the winters of 2017 and 2018 atPoyang Lake
Table 2 Number of food items dietary niche breadth and Shannon-Wiener diversity values for Siberian Cranes White-napedCranes Hooded Cranes and Eurasian Cranes at Poyang Lake in the winters of 2017 and 2018
Number of foods Dietary niche breadth Shannon-Wiener diversity
2017 2018 2017 2018 2017 2018
Siberian Crane 16 17 005 007 091 085
White-naped Crane 18 15 020 003 169 059
Hooded Crane 19 13 005 002 096 043
Eurasian Crane 18 19 008 009 110 121
Hou et al Frontiers in Zoology (2021) 1824 Page 6 of 17
that are overlooked by microhistological methods [6780] It also enables the identification of food items at ahigher taxonomic resolution [69 81] However we ac-knowledge that our study has two limitations First weused fecal samples as they are non-invasive Neverthe-less fecal analysis is subjected to the biases caused bythe differential digestive rates of food items [82 83] Softfoods such as insect larvae tend to be under-representedin fecal samples because they are digested quickly andremain in gastro-intestinal tract for short periods whilehard foods such as mollusks tend to be over-representedbecause they break down slowly and remain in gastro-intestinal tracts longer Thus food proportions in the
feces may not correlate with food proportions ingestedby cranes In addition some materials identified in thefeces may be incidentally consumed by cranes and notreally be used as food Second our study only consideredthe plant component of cranesrsquo diets as plants were sug-gested to be their principal food types at Poyang LakeAnimal foods consumed by the four crane species butto a much lesser extent than plant foods [84ndash87] werenot considered in our studyThe Siberian Crane is regarded as the most aquatic of
all cranes using wetlands for nesting feeding and roost-ing [49 88] The high dependence of the species onaquatic habitats was suggested to make it vulnerable to
Table 3 The observed dietary niche overlap among the four crane species at Poyang Lake in the winters of 2017 and 2018 P(Obs lt = null) and P (Obs gt = null) indicate the tail probabilities for the observed index which were lower and higher than thehistogram of the simulated index respectively represents P lt 005 and represents P lt 001
Winter Observed niche overlap index P (Obs lt = null) P (Obs gt = null)
2017 Siberian vs White-naped 021 0819 0181
Siberian vs Hooded 002 0703 0297
Siberian vs Eurasian 016 0900 0100
White-naped vs Hooded 049 0952 0048
White-naped vs Eurasian 035 0895 0106
Hooded vs Eurasian 035 0919 0081
2018 Siberian vs White-naped 063 0952 0048
Siberian vs Hooded 066 0952 0048
Siberian vs Eurasian 062 0950 0049
White-naped vs Hooded 099 0997 0003
White-naped vs Eurasian 089 0987 0013
Hooded vs Eurasian 090 0998 0002
Fig 4 The results of nonmetric multidimensional scaling ordination (NMDS) based on Bray-Curtis dissimilarity The colors of the circlescorrespond to the crane species blue Siberian Crane purple White-naped Crane green Hooded Crane and red Eurasian Crane
Hou et al Frontiers in Zoology (2021) 1824 Page 7 of 17
impacts of habitat degradation and to lead its classifica-tion as Critically Endangered [49] At Poyang Lake Si-berian Cranes traditionally fed on tubers of Vallisneriathe dominant submerged macrophytes in shallow watersand mudflats [89 90] In winter 2010 due to a flood-induced Vallisneria tuber collapse they were observedforaging for P limprichtii taproots and T edulis bulbs ingrasslands for the first time [38 44] In the winters from2015 to 2017 the diets of Siberian Cranes changedagain with thousands of cranes feeding on rice seedsand lotus rhizomes in agricultural habitats [61 62 64]Our study identified 22 plant items in the feces of Siber-
ian Cranes These plant items were obtained from shallowwaters grasslands and agricultural fields Four plant itemswith yearly RRA gt 10 were identified in the feces Vallis-neria tuber P criopolitanum rhizomes rice seeds andlotus rhizomes (the vegetative portions consumed bycranes were based on our field observation) SiberianCranesrsquo diet compositions showed substantial interannualvariation When the abundance of Vallisneria tubers waslow (ie winter 2017) Siberian Cranes mainly fed on riceseeds lotus rhizomes and P criopolitanum rhizomes inagricultural fields and grasslands When the abundance ofVallisneria tubers was high (ie winter 2018) they mainlyfed on Vallisneria tubers and P criopolitanum rhizomesin shallow waters and grasslands Our study in combin-ation with previous studies indicates that the diets of Si-berian Cranes have changed greatly since winter 2010 [3844 62] They have gradually broadened their dietaryniches which has probably been driven by the degradationof submerged plants [38 44 57]Similar to Siberian Cranes a diet shift was also ob-
served in White-naped Cranes Historically White-naped Cranes only fed on submerged plants in shallowwaters and mudflats of Poyang Lake [91] Then drivenby the decline of submerged plants they primarily fed atgrasslands and mudflats [87 91] consuming Vallisneriatuber T edulis P limprichtii and Ranunculus polii [5587] Our results indicate that in addition to T edulisand P limprichtii that had previously been reported theproportions of P criopolitanum and Carex were alsohigh in the feces Hooded Cranes were suggested tomainly feed on the grassland plant P limprichtii throughdirect observation [86] Our study indicates that thegrassland plants P criopolitanum and Carex also occu-pied high proportions in the feces The dominant fooditems in the feces of White-naped Cranes and HoodedCranes varied greatly between the two winters The vari-ation in dietary composition indicated that the diversityand composition of the grassland plant communitymight have changed greatly between the two wintersThe dietary niche overlap between White-naped Cranesand Hooded Cranes was the highest among the fourcrane species aligning with their level of habitat niche
overlap which was also the highest among the species[92] The high dietary and habitat niche overlaps suggesthigh competition potential between the two cranespeciesThe Eurasian Crane is a habitat generalist It is known
to feed in a variety of habitats including grasslandsmudflats shallow waters and agricultural fields [88 92]Its diet at Poyang Lake has not been documented previ-ously Our study indicates that P criopolitanum P lim-prichtii and rice were the dominant food items in thefeces of Eurasian Cranes The number of plant itemsconsumed dietary niche breadth and Shannon-Wienermeasures were the highest among the four crane speciessupporting its generalist foraging strategy In contrast tothe three crane species above Eurasian Cranesrsquo dietcompositions were similar between the two winters Riceand grassland plants occupied high proportions in thefeces in both winters
Important food resources for cranesVallisneria tubers have been acknowledged as importantfood sources for cranes especially Siberian Cranes andWhite-naped Cranes [55 56] Our study confirms theimportance of Vallisneria tubers in the diet of SiberianCranes It was the most common food source in thefeces of Siberian Cranes in winter 2018 when its abun-dance was high Vallisneria tuber proportions in thefeces of the three other crane species were not as high asthat in the feces of Siberian Cranes but it still occupied5ndash9 of the total foods in winter 2018To our surprise P criopolitanum was found to be the
first dominant food in the feces of White-naped CranesHooded Cranes and Eurasian Cranes and the seconddominant food in the feces of Siberian Cranes in winter2018 In winter 2017 the proportions of P criopolita-num were lt 17 in the feces of Siberian Cranes andWhite-naped Cranes and were lt 3 in the feces ofHooded Cranes and Eurasian Cranes The higher pro-portions of P criopolitanum in winter 2018 than winter2017 indicate that the abundance and availability of Pcriopolitanum was much higher in winter 2018 P crio-politanum is an annual grassland plant It is sensitive tohydrological conditions and can only survive in a rela-tively narrow range of water depths and percent time in-undated [93] Due to the sensitivity its distributionrange and abundance showed high interannual variation[94 95] Thus it is likely that the abundance of P crio-politanum was higher in winter 2018 than in winter2017The importance of P criopolitanum as a food source
for the cranes at Poyang Lake has not been documentedpreviously It has only been suggested to be a major foodsource of Hooded Cranes at Shengjin Lake another lakein the middle and lower Yangtze River floodplain [86
Hou et al Frontiers in Zoology (2021) 1824 Page 8 of 17
96] P criopolitanum is a dominant wetland plant at Po-yang Lake [93 97] The coverage percentage of this plantis the third highest among grassland plants accountingfor 12 of the total areas of grasslands [97] This plantspecies is usually distributed at elevations of 9ndash12mabove sea level near mudflats and shallow waters [97]which are frequently used by cranes Its rhizome with ashallow burial depth is high in energy and nutritionalcontent and low in crude fibre [38 86] All these charac-teristics make P criopolitanum rhizome a potentially im-portant food source for cranes We frequently observedcranes dig and feed on rhizomes of P criopolitanum andleave aboveground parts in the fields Therefore we be-lieve that P criopolitanum was used as food not inci-dentally consumed by cranes However we need to becaution that high proportion of P criopolitanum in thefeces does not mean high proportion in the diets Morework on the importance of P criopolitanum in the dietsof cranes is needed
The roles of agricultural fields in crane protectionWith the loss and degradation of natural wetlands theroles of artificial wetlands in waterbird protection havebeen under intensive debate [98 99] Although agricul-tural fields cannot completely replace the function ofnatural wetlands as waterbird habitats [100ndash102] agri-cultural fields might provide alternative or complemen-tary habitats for waterbirds in all life stages [38 103104] The proportions of domesticated species in thefeces of Siberian Cranes varied considerably betweenyears In winter 2017 when the abundance of Vallisneriatubers was low the proportion of Vallisneria in SiberianCranesrsquo feces was only 095 Siberian Cranes may nothave found enough foods in the shallow waters andmudflats and were forced to search for rice seeds andlotus rhizomes in agricultural fields In winter 2018when the abundance of Vallisneria tubers reboundedSiberian Cranes moved back to shallow waters and mud-flats and a large proportion of their traditional food Val-lisneria tubers and also P criopolitanum were detectedin the feces A similar abandonment of tuber foragingoccurred following the tuber collapse in 2010 and thenresumed in the next winter when tubers rebounded in2011 [38 44] The diet and habitat changes indicate thatagricultural habitats were important refuges for SiberianCranes providing important food resources for them tosurvive through winters with few Vallisneria tubersThey also indicate that Siberian Cranes prefer naturalwetlands to agricultural fields [38]Similar to Siberian Cranes White-naped Cranes and
Hooded Cranes consumed rice seeds and lotus rhizomesin winter 2017 but did not consume any in winter 2018The changes in the proportions suggest that the abun-dance of food resources in natural wetlands might have
been low in winter 2017 The deficient food resourcesmight have been Vallisneria tubers but might also havebeen grassland plants the primary food sources of thesetwo crane species Under natural food shortage condi-tions (ie winter 2017) White-naped Cranes andHooded Cranes utilized agricultural fields When theabundance of Vallisneria tubers and probably grasslandplants were high (ie winter 2018) the two crane speciesreturned to their natural habitats Therefore for White-naped Cranes and Hooded Cranes agricultural fieldswere important alternative foraging habitats used asbuffers against starvation during periods of natural foodshortages The return of the two crane species to naturalwetlands in winter 2018 further emphasizes the superiorquality of natural habitats over agricultural fields as for-aging habitats for cranes [38]The proportions of rice seeds in the feces of Eur-
asian Cranes were high in the two winters thoughthe proportion in winter 2017 was twice as high asthat in winter 2018 Therefore agricultural fields ap-peared to be consistently important as foraging habi-tats for Eurasian Cranes The frequent use ofagricultural fields by Eurasian Cranes has been docu-mented widely [49 105 106]Domesticated species generally contain high energy
and nutritional content and are of superior quality com-pared to natural plants [86 104] Farmland foraging hasbeen suggested to contribute to increase in the abun-dance of some goose [107] egret [108] and wader [109]populations in Japan Korea Europe and North Amer-ica However cranes generally avoid human disturbancewhen select foraging sites [110 111] Human disturb-ance in agricultural fields around Poyang Lake is consid-erably high due to intensive agricultural activity [112]High human disturbance was suggested to be a majorreason that wintering geese were almost confined withinnatural wetlands and hesitated to exploit the riches ofthe modern agricultural habitats at Poyang Lake [112113] High human disturbance also resulted in SiberianCranes spending twice as much time alerting in agricul-tural fields than they did in natural habitats [114] Thehigh level of human disturbance in agricultural fieldsmight contribute to the preference for natural wetlandsby Siberian Cranes White-naped Cranes and HoodedCranes at Poyang Lake Moreover although agriculturefields contain energy-rich crops wetlands contain di-verse food resources especially protein-rich inverte-brates which might better satisfy dietary needs of cranes[110 115] Taken together security and diverse food re-sources might have attracted cranes to select wetlands asforaging sitesSubmerged macrophytes at Poyang Lake have de-
graded in recent decades [57ndash60] and Vallisneria tuberdisappearance is expected to occur more frequently
Hou et al Frontiers in Zoology (2021) 1824 Page 9 of 17
under future climate change [57 60] Cranes may in-creasingly depend upon agricultural fields around Po-yang Lake which may in turn cause significant cropdamage and economic losses This is especially true onagricultural fields surrounding protected wetlands wherewaterbirds concentrate in high numbers [116 117] AtPoyang Lake cranes mainly fed on post-harvest remainsof rice seeds and lotus rhizomes thus crop damage andeconomic loss were limited However we observed somefarmers scared cranes away from their agricultural fieldsThere were more than 14 million free-ranging poultryraised at agricultural fields around Poyang Lake [118]where they gleaned waste grain residues in stubble fieldsCrane feeding would reduce the food abundance of thefree-ranging domestic poultry thus farmers drovecranes away To reduce human disturbance environ-mental education is needed because farmers are morelikely to protect cranes if they understand the endan-gered degree and the ecological significance of cranes
Effects of tuber collapse on dietary niche breadth andoverlap among crane speciesVallisneria tuber is an important food source for Siber-ian Cranes (89 90 this study) thus variation in itsabundance might influence Siberian Cranesrsquo dietaryniche breadth The numbers of food items dietary nichebreadth and Shannon-Wiener measures of SiberianCranes were similar between winter 2017 and winter2018 This is inconsistent with the contention that dur-ing seasons with a low abundance of dominant food thedietary niche may broaden as consumers relying on in-sufficient preferred food items are forced to add lessprofitable resources to their diets [26 119 120] In win-ter 2017 with low Vallisneria tuber abundance SiberianCranes left shallow waters and mudflats and moved torice paddies and lotus ponds [61 62 64] Their foodschanged from Vallisneria tuber to rice seeds and lotusrhizomes almost completely As the energy protein andfat contents of crops are generally as good as or superiorto natural foods and are often present in agriculturalfields in far greater abundance [86 104] Siberian Cranesmay gain enough energy through feeding on crops anddo not need to add many other food sources Moreoverthe Siberian Crane is a dietary specialist [89 90] PoyangLake has a relatively simple plant community compos-ition with Vallisneria and Hydrilla verticillate being thedominant submerged macrophytes [59] and Carex beingthe dominant grassland plants [121] Therefore theremay be few options for Siberian Cranes which mightlead to the failure of this species to widen its dietaryniche breadthIn contrast to Siberian Cranes the numbers of food
items dietary niche breadths and Shannon-Wiener mea-sures of White-naped Cranes and Hooded Cranes were
higher in winter 2017 than in winter 2018 P criopolita-num was the predominant food item in the feces of thetwo crane species in winter 2018 and the proportions ofVallisneria tubers were also high The abundances ofVallisneria tubers and probably P criopolitanum at Po-yang Lake declined greatly in winter 2017 When thereis a short-fall in food availability animals may optimizeforaging behavior and increase their dietary breath [122123] The short-fall in the abundances of P criopolita-num and Vallisneria tubers in winter 2017 might havedriven the two crane species to add less profitable grass-land plants in their diets Although the two cranes alsoconsumed high-quality rice seeds and lotus rhizomes inwinter 2017 their proportions were low in the feces andthese foods might be unable to meet the energy require-ments of the two crane species The niche expansion ofthe two crane species during periods of preferred foodshortage was in agreement with the basic prediction ofoptimal foraging theory [22 23] It is also consistentwith a commonly observed fact that energy-maximizingmodels work better when animals food-limited [124]The dietary niche overlaps among the crane species
were low in winter 2017 with the four crane species di-verging in their principal food items However in winter2018 the niche overlaps were significantly higher thanexpected by chance They had similar diet compositionswith P criopolitanum rhizomes and Vallisneria tubersbeing the principal food items in the feces The in-creased overlap might have been caused by the highabundances of the two plant species in winter 2018which allowed the four crane species to share the samefood items with low interspecific competition Our studyis consistent with previous research that suggested thatamong-species dietary niches tended to overlap moreduring seasons of high food abundance [32 33 125126] Greater resource availability could promote an in-crease in species utilization of similar resources and re-duce competitive interactions between species [32 34126] By contrast during seasons with low food abun-dance species may partition limited food resources todecrease interspecific competition leading to a decreasein niche overlap [126 127] It has been suggested thatdiet partitioning facilitates the coexistence of manyclosely related species [7 128] The interannual variationin niche overlap observed in our study suggests that dietpartitioning may be restricted to specific temporal casesIn seasons or years with abundant food sources inter-specific competition may be relaxed and niche overlapcould be high
ConclusionsIn summary by using metabarcoding technology ourstudy gained new insights into cranesrsquo diets that are es-sential for informing habitat management to improve
Hou et al Frontiers in Zoology (2021) 1824 Page 10 of 17
the availability of foraging resources Vallisneria tuberswas confirmed as an important food source of cranesSurprisingly P criopolitanum also occupied high pro-portions in the feces of cranes and may also be import-ant Our study emphasizes the superior quality ofnatural wetlands compared to agricultural fields as for-aging habitats for cranes The recent degradation of sub-merged macrophytes at Poyang Lake potentiallythreatens the survival of cranes especially SiberianCranes Therefore it is urgent to reverse the broad de-cline of submerged macrophytes and provide importantforaging habitats for cranes during years of tuber col-lapses The abundances of Vallisneria tubers and prob-ably P criopolitanum at Poyang Lake greatly influencedthe dietary niche width and overlaps among crane spe-cies Cranes exhibited low dietary niche overlap duringperiods of preferred food shortage but significantly highoverlap during periods when preferred foods were plen-tiful While high niche overlap is often interpreted as in-tense interspecific competition our study highlights theimportance of taking food abundance into account
Materials and methodsStudy areasPoyang Lake the largest freshwater lake in China is oneof the most important wintering areas for waterbirdsalong the East Asian-Australasian Flyway (Fig 1) [51129] It is characterized by dramatic seasonal hydro-logical fluctuations with an average change of 924 mbetween the summer high and winter low water levels[130] During summer the inundation area is gt 2500km2 [131] In autumn and winter the water level dropscausing the inundation area to shrink to lt 1000 km2
[131] Poyang Lake contains 102 sublakes which are in-undated and integrated with the main body of PoyangLake during summer and are isolated during winter[132] As the water level drops the sublakes appear andlarge areas of grasslands mudflats and shallow waterareas are exposed providing winter foraging and roost-ing areas for approximately 420000 waterbirds from ap-proximately 111 species [133 134]
Vallisneria tuber surveyThe density and biomass of Vallisneria tubers were sur-veyed at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake in late October and earlyNovember of 2017 and 2018 before large numbers ofwintering waterbirds arrived We set 76 183 and 36sampling plots uniformly located at Dahuchi Shahu andChanghuchi respectively Vallisneria tubers were col-lected from two quadrats in each plot with a locallymade steel grab sampler The sampler has two long
handles At the end of the handles there are two scoopsfacing each other The sampler was inserted into thesubstrate and the scoops brought together to collectabout 15 cm long times 13 cm wide times 30 cm high sample ofsubstrate Tubers were subsequently cleaned andcounted The dry weight of tubers in each quadrat wasweighed after oven-drying at 80 degC until constant weightThe tuber density and biomass in each plot was calcu-lated by dividing total tuber number and tuber massacross two quadrats by quadrat area respectively Weused t-test to explore whether the tuber density and bio-mass in the winters of 2017 and 2018 significantly devi-ated from the historical average values in the wintersfrom 1999 to 2016 [57] The historical tuber density andbiomass data were collected by the Poyang Lake Na-tional Nature Reserve at three sublakes (Dahuchi Shahuand Meixihu) [57] R 360 was used to do the t-testanalyses
Fecal sample collectionFecal samples were collected at Poyang Lake in the win-ters of 2017 and 2018 We first investigated the distribu-tions of cranes When we saw foraging cranes we waiteduntil they left to avoid disturbing them Soon after theyleft we went to the foraging sites to collect fresh feces(ie feces with wet surface) with sterilized tweezers Tominimize the probability of multiple samples from thesame individual all collected samples were separated byat least 5 m We stored the samples in liquid nitrogen inthe field and transferred them to a minus 80 degC refrigeratorfor long-term storage in the laboratory To ensure thatthe sources of the feces were known we tried to collectsamples from monospecific flocks For samples collectedfrom mixed-species flocks we used DNA analysis toidentify the species that deposited the fecal samplesPlants were also collected in the field to construct aDNA barcoding reference database
Species identificationBecause of the similar morphologies of the feces of thefour crane species we used DNA analysis to identify thespecies that deposited the fecal samples collected frommixed-species flocks For the genetic species assignmentof each fecal sample we scraped the external surfacewhich contain DNA from the host species as a result ofthe sloughing of cells from the digestive tract DNA wasextracted with the QIAamp Power Fecal DNA Kit (Qia-gen USA) according to the manufacturerrsquos guidelinesFor each round of DNA extraction negative controls(ie extraction without feces) were included to monitorfor possible contamination We amplified the mitochon-drial DNA D-loop fragment with the primer pair DL02F
Hou et al Frontiers in Zoology (2021) 1824 Page 11 of 17
(5prime-3prime GCGGCCCGAAAAGCCGCTG) and DL02R (5prime-3prime GCCCTGACATAGGAACCAGAGGCGC) [135]PCR amplifications were carried out in a total volume of25 μl containing 125 μl PCR mix (Tiangen BeijingChina) 1 μl DNA 1 μl each primer and 95 μl H2O Thereaction conditions were as follows denaturation at94 degC for 4 min followed by 34 cycles at 94 degC for 48 s56 degC for 48 s and 72 degC for 1 min and a final 10 min at72 degC We checked for the presence of a PCR product ofsuitable length by electrophoresis on a 1 agarose gelSequencing was carried out on an ABI 3730xl analyzerby Sangon Biotech (Shanghai China) The complete se-quences were assembled using Seqman II (DNASTAR)and compared visually to the original chromatograms toavoid reading errors The taxonomic assignment wasconducted by searching against the nucleotide databaseof GenBank using Web BLAST
Construction of a barcode reference databaseWe downloaded the trnL sequences of 48 plant speciescommonly seen at Poyang Lake from GenBank For theplant species not included in GenBank we sequencedtrnL by ourselves Total DNA was extracted using thehexadecyltrimethylammonium bromide (CTAB) proto-col We used primer pair c (5prime-3prime CGAAATCGGTAGACGCTACG) and h (5prime-3prime CCATTGAGTCTCTGCACCTATC) to amplify an approximately 150 bp regionof the chloroplast trnL intron [136] This primer pairwas chosen as it can amplify DNA from a wide range ofwetland plants [70] PCR amplifications were carried outin a total volume of 25 μl containing 125 μl PCR mix(Tiangen Beijing China) 1 μl DNA 1 μl each primerand 95 μl H2O The reaction conditions were as followsdenaturation at 94 degC for 4 min followed by 35 cycles at94 degC for 30 s 56 degC for 30 s and 72 degC for 45 s and afinal 10 min at 72 degC We used electrophoresis to checkthe PCR products ABI 3730xl analyzer to perform se-quencing and Seqman II to assemble the sequences
DNA extraction and high-throughput sequencing of fecalsamplesTo uncover plant component consumed by cranes amodified CTAB protocol [70] was used to extract theDNA from approximately 200 mg of the interior of eachfecal sample DNA extraction was conducted in a dedi-cated laboratory for analyses of DNA from samples withlow DNA quality For each batch of DNA extractionnegative controls were included to monitor for possiblecontamination The primers c and h with a 6 bp tagadded to 5prime ends of each primer were used to amplifytrnL gene The PCR amplification procedure was thesame as barcode database construction For each samplea unique combination of tags in the forward and reverseprimers was used so that sample-specific data can be
recovered after sequencing Three PCR replicates wereperformed for each sample to reduce amplification biasThe three replicates were pooled and the DNA concen-tration and purity were monitored on a 1 agarose gelOnly samples with a clear band were processed furtherSamples were purified using the SanPrep Column PCRProduct Purification Kit (Sangon Biotech ShanghaiChina) The purified PCR products were pooled takinginto account of DNA concentration to obtain an ap-proximately equal amount of amplicon DNA from eachfecal sample The pool of individually tagged ampliconswas used to prepare a sequencing library using the Tru-Seq DNA PCR-Free Sample Preparation Kit (IlluminaUSA) following manufacturerrsquos recommendations Thelibrary was sequenced on an Illumina HiSeq 2500 plat-form and 250 bp paired-end reads were generated Li-brary preparation and sequencing were conducted byNovogene (Beijing China)
Sequencing data analysisAfter high-throughput sequencing the paired-end readswere demultiplexed into sample-specific files based ontheir unique tags The tags and primer sequences weresubsequently trimmed The paired-end reads weremerged using FLASH 127 [137] and then filtered usingQIIME 170 [138] Sequences were dereplicated andclustered to OTUs using USEARCH 110667 [139] atthe similarity threshold of 97 All OTUs were assignedto taxonomic units by referring to the local plant data-base using blast 2231+ with thresholds of identity gt97 and e-value lt 10 eminus 50 If an OTU matched two ormore taxa it was assigned to a higher taxonomic levelthat included all taxaTo evaluate the sufficiency of sequencing depth and
sampling size individual- species- and sample-basedrarefaction curves were built using EstimateS 82[140] The curves were computed by randomly resam-pling sequences or samples and plotting these againstthe numbers of OTUs The read abundance data wereconverted to RRA of each food item using the follow-ing equation [79]
RRA frac14 1S
XS
Kfrac141
nikPTifrac141nik
100
where nik is the number of sequences of food item i insample k T is the total number of food items and S isthe sampling size
Dietary niche breadth and overlap among crane speciesTo explore whether the cranesrsquo diets showed substantialinterannual variation we calculated Pearson correlationsbetween diet compositions of each crane species in win-ter 2017 and winter 2018 The proportion of each food
Hou et al Frontiers in Zoology (2021) 1824 Page 12 of 17
item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

54] It has been suggested that other than Poyang Lakethere are no alternative wintering sites for SiberianCranes remaining in the Yangtze River floodplain [49]Tubers of Vallisneria spp the dominant submerged
macrophytes at Poyang Lake is an important food sourcefor cranes [38 55 56] However submerged macrophyteshave degraded seriously in recent decades [57ndash60] Theywere widely distributed throughout Poyang Lake in previ-ous years but were restricted to small areas in 2012 [58]The Vallisneria tuber density and biomass at three sub-lakes (Dahuchi Shahu and Meixihu) of Poyang Lake de-clined greatly in the winters from 1999 to 2017 [57 60]The frequency and range of Vallisneria tuber collapsehave also increased because of frequently occurring sum-mer flood and autumn drought extensive aquaculture de-clining water quality and other factors [38 57 60]Tuber collapse has led to dietary and foraging habitat
shifts of cranes For example the tuber collapses in thewinters of 2015 and 2016 drove thousands of SiberianCranes for the first time to switch from foraging inshallow waters they typically used to paddy fields and
lotus ponds [57 61] Their main foods changed fromVallisneria tubers to rice (Oryza sativa) seeds and lotus(Nelumbo nucifera) rhizomes [62] Many White-napedCranes Hooded Cranes and Eurasian Cranes also movedfrom natural wetlands to forage in paddy fields [63 64]Diet shifts may influence the dietary niche width andoverlap between crane species and subsequently thecompetition level and fitness of cranes Given the highlyendangered status of cranes and the importance of Po-yang Lake in crane protection it is important to under-stand the consequences of Vallisneria tuber collapse ondiets and competition levels among crane species so thateffective protection measures can be implementedTraditional dietary analysis methods include direct ob-
servation of foraging animals and microscopic examin-ation of feces or gut contents While these approacheshave been suggested to be useful in some cases they alsoexhibit methodological limitations [65ndash67] The maindrawbacks are that the methods can be inaccurate andlabor-intensive [65 68] Recently the emergence ofDNA metabarcoding has provided new perspectives for
Fig 1 Maps showing the three Vallisneria tuber survey sublakes (Dahuchi Shahu and Changhuchi) and the sampling sites of crane feces atPoyang Lake Circles and squares represent sampling sites in the winters of 2017 and 2018 respectively The colors of the circles and squarescorrespond to the crane species red Siberian Crane blue White-naped Crane green Hooded Crane and purple Eurasian Crane The base mapshows the inundation area of Poyang Lake in winter 2008 when the water level was similar to the historical average water level
Hou et al Frontiers in Zoology (2021) 1824 Page 3 of 17
diet analysis [65] This method is based on amplifyingand high-throughput sequencing a standardized DNAregion from stomach contents or feces and subsequentlycomparing it to a reference database for identification ofthe consumed species [65] Compared to traditionalmethods DNA metabarcoding generally provides highertaxonomic resolution identifies more food items andcan simultaneously analyze a larger number of samples[69ndash71] After its first application in assessment of thediets of Australian Fur Seals (Arctocephalus pusillus dor-iferus) [72] metabarcoding has been used successfully tostudy diets of herbivores [9 73] carnivores [69 74] andomnivores [75 76]In this study we used metabarcoding technology to
explore the consequences of Vallisneria tuber collapseon the diet compositions niche breadths and nicheoverlaps of the four crane species at Poyang Lake Fecalsamples were collected in winter 2017 (ie 20172018 ayear with Vallisneria tuber collapse) and winter 2018(ie 20182019 a year with high Vallisneria tuber abun-dance see Results for details) We first explored the di-ets of the four crane species and their interannualvariations Most previous dietary studies of cranes werebased on traditional direct observation or microscopicexamination Here we hoped to apply the metabarcod-ing method to provide a more accurate and comprehen-sive understanding of the cranesrsquo diets We thenevaluated the role of agricultural fields in crane protec-tion Vallisneria tuber collapses have driven thousandsof cranes to feed in agricultural fields [61 64] Howeverthe proportion of domesticated species in the diets ofthe four crane species has not previously been quanti-fied Here we determined the relative dietary import-ance of domesticated species in the feces of cranesFinally we assessed the consequences of Vallisneriatuber collapse on the dietary niche width and overlapamong crane species To do so we compared the nichebreadths and overlaps in winter 2017 and winter 2018with different Vallisneria tuber abundances Althoughwe focused on cranes at Poyang Lake our findings canprovide a reference for dietary shifts of consumers in de-graded wetlands worldwide
ResultsVariation in Vallisneria tuber abundanceWe investigated the density and biomass of Vallisneriatubers at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake The average tuber densityand biomass were 668 tubersm2 and 054 gm2 respect-ively in winter 2017 (Table 1) The density and biomassrapidly increased to 3457 tubersm2 and 411 gm2 inwinter 2018 The historical average tuber density andbiomass at three sublakes (Dahuchi Shahu and Meixihu)of Poyang Lake were 1033 (plusmn 1127 SD) tubersm2 and
326 (plusmn 368 SD) gm2 in the winters from 1999 to 2016[57] Tuber density in winter 2017 (t = 133 P = 0201)and tuber biomass in winter 2018 (t = minus 094 P = 0361)did not differ from the historical average values How-ever tuber biomass in winter 2017 was lower than thehistorical average values (t = 310 P = 0007) and tuberdensity in winter 2018 was higher than the historicalaverage values (t = minus 888 P = 0000) Therefore winter2017 represented a year with tuber collapse while winter2018 represented a year with high tuber abundance Co-inciding with the changes in tuber abundance a total of4014 individuals of Siberian Cranes White-napedCranes and Hooded Cranes were recorded in agricul-tural fields adjacent to Poyang Lake in winter 2017 [63]while none of the three crane species was recorded inagricultural fields in winter 2018 [77]
Collection statisticsIn total we collected 129 fecal samples with 60 samplesfrom winter 2017 and 69 samples from winter 2018 (Fig1 Additional file 1 Table S1) We collected 42 fecalsamples of Siberian Cranes from 2 sublakes 4 rice pad-dies and 1 lotus pond 20 fecal samples of White-napedCranes from 2 sublakes and 1 rice paddy 19 fecal sam-ples of Hooded Cranes from 2 sublakes and 1 ricepaddy and 48 fecal samples of Eurasian Cranes from 3sublakes and 6 rice paddies We also collected 38 plantspecies commonly seen at Poyang Lake to construct aDNA barcode reference library By downloading se-quences from GenBank and sequencing by ourselves weconstructed a DNA barcode reference library containing62 plant species
High-throughput DNA sequencingThe chloroplast trnL intron was selected as a barcodebecause of its high taxonomic coverage and resolutionespecially for wetland plants [70 78] The sequencing ofthe 129 fecal samples yielded a total of 9284819 readsAfter quality filtering 8502418 reads were obtained
Table 1 The density and biomass of Vallisneria tubers at threesublakes (Dahuchi Shahu and Changhuchi) of Poyang Lake inthe winters of 2017 and 2018
Winter Sublake Tuber density(tubersm2)
Tuber biomass(gm2)
2017 Dahuchi 980 097
Shahu 028 002
Changhuchi 997 062
Average 668 054
2018 Dahuchi 641 065
Shahu 897 060
Changhuchi 8832 1107
Average 3457 411
Hou et al Frontiers in Zoology (2021) 1824 Page 4 of 17
The average number of reads per sample was 65910 (plusmn26276 SD) and the average number of operational taxo-nomic units (OTUs) per sample was 43 (plusmn 15 SD) Toevaluate the sequencing and sampling adequacy individ-ual- species- and sample-based rarefaction curves werebuilt With the sample sequencing reads increased theindividual-based rarefaction curves reached plateau inmost cases (Additional file 1 Figure S1) and the species-based rarefaction curves reached plateau in the fourcrane species (Fig 2a) Therefore our sequencing depthwas roughly adequate to capture the numbers of OTUspresent The sample-based rarefaction curves for eachcrane species reached plateau in all cases as the samplingsizes increased (Fig 2b) suggesting that we acquired suf-ficient fecal samples After comparison to the local DNAreference database 9899 of the total reads wereassigned to specific plant taxa Most of the reads thatwere not assigned to specific plant taxa had low fre-quency In total we identified 29 plant taxa including23 species 4 genera and 2 families
Diet compositions of the four crane speciesWe used relative read abundance (ie the percentageof DNA belonging to each food item in each sampleRRA) which provides a more accurate view of speciesdiet than frequency of occurrence (ie the number ofsamples that contain a given food item) tosummarize dietary data [79] Twenty-two food itemswere identified in the feces of Siberian Cranes in thetwo winters including 17 species 3 genera and 2families (Fig 3 Additional file 1 Table S2) Rice(RRA = 7285) lotus (1201) and Polygonum criopo-litanum (1199) were the dominant food items inthe feces (ie proportion gt 10) in winter 2017 andVallisneria (5612) and P criopolitanum (4087)were the dominant food items in winter 2018Twenty-five food items were identified in the feces ofWhite-naped Cranes including 20 species 3 genera
and 2 families Tulipa edulis (3737) Potentilla lim-prichtii (1869) P criopolitanum (1652) andCarex spp (1075) were the dominant food items inthe feces in winter 2017 and P criopolitanum(8441) was the dominant food item in winter 2018Twenty-two food items were identified in the feces ofHooded Cranes including 18 species 3 genera and 1family P limprichtii (6536) and Carex (2739)were the dominant food items in the feces in winter2017 and P criopolitanum (8876) was the domin-ant food item in winter 2018 Twenty-two food itemswere identified in the feces of Eurasian Cranes in-cluding 17 species 3 genera and 2 families Rice(6120) and P limprichtii (2394) were the domin-ant food items in the feces in winter 2017 and Pcriopolitanum (5577) and rice (2740) were thedominant food items in winter 2018 Diet composi-tions of Siberian Cranes (Pearson correlation = 005P = 077) White-naped Cranes (Pearson correlation =030 P = 008) and Hooded Cranes (Pearson correl-ation = minus 004 P = 083) were not correlated betweenwinter 2017 and winter 2018 suggesting great inter-annual variations in food sources Diet composition ofEurasian Cranes was correlated (Pearson correlation =040 P = 002) between the two winters suggestinglimited interannual variation in food sourcesVallisneria tubers were an important food source for
the four crane species especially Siberian Cranes inwinter 2018 The proportion of Vallisneria was 5612512 850 and 734 in the feces of Siberian CranesWhite-naped Cranes Hooded Cranes and EurasianCranes respectively in winter 2018 (Fig 3 Additionalfile 1 Table S2) In winter 2017 when Vallisneria tuberabundance was low the proportion of Vallisneria wasless than 1 in the feces of the four crane speciesDomesticated species identified in crane feces included
rice and lotus The proportions of domesticated speciesin the feces of Siberian Cranes (8485) and White-
Fig 2 a Species-based and b sample-based rarefaction curves The species- and sample-based rarefaction curves built by randomly resamplingsequences and samples respectively at increasing levels of accumulation indicate whether the sequencing depth and sampling size weresufficient to characterize the diversity of dietary items The colors of the curves correspond to the crane species blue Siberian Crane purpleWhite-naped Crane green Hooded Crane and red Eurasian Crane
Hou et al Frontiers in Zoology (2021) 1824 Page 5 of 17
naped Cranes (1509) were high in winter 2017 but de-clined to 000 in winter 2018 (Fig 3 Additional file 1Table S2) Domesticated species occupied a small pro-portion in the feces of Hooded Cranes in winter 2017(130) but declined to 000 in winter 2018 The pro-portions of domesticated species in the feces of EurasianCranes were high in both winters with 6120 in winter2017 and 2740 in winter 2018
Dietary niche breadth and overlap among crane speciesThe numbers of food items dietary niche breadth andShannon-Wiener diversity values for Siberian Cranesand Eurasian Cranes in winter 2017 were similar tothose in winter 2018 (Table 2) Values of all three pa-rameters for White-naped Cranes and Hooded Craneswere higher in winter 2017 than in winter 2018 Dietaryniche breadth of the four crane species was lt 020 inboth winters indicating low niche breadthDietary niche overlap among crane species ranged
from 002 to 049 in winter 2017 but increased to 062to 099 in winter 2018 (Table 3) Nonmetric multidimen-sional scaling (NMDS) analysis also indicated overlap
increases in winter 2018 (Fig 4) No overlap values werelower than expected by chance in the two winters Theoverlap values were not higher than expected by chancein winter 2017 However all values showed a high de-gree of overlap in winter 2018 [P (Obs gt = null) lt 005]Overlap between White-naped Cranes and HoodedCranes was the highest among the four crane species inboth winters
DiscussionDiet compositionTo the best of our knowledge our study is the first toexplore crane diets using metabarcoding techniques Al-most all previous studies have relied on traditional directobservation or microhistological methods By usingmetabarcoding technology we were able to elucidatecrane diets at greater detail than previously known Theimproved knowledge might be due to the ability ofmetabarcoding to provide a more comprehensive per-spective and more details on food sources than trad-itional methods [66 67 80] Metabarcoding is able todetect a variety of smaller and softer bodied food items
Fig 3 Food items in the feces of Siberian Cranes White-naped Cranes Hooded Cranes and Eurasian Cranes in the winters of 2017 and 2018 atPoyang Lake
Table 2 Number of food items dietary niche breadth and Shannon-Wiener diversity values for Siberian Cranes White-napedCranes Hooded Cranes and Eurasian Cranes at Poyang Lake in the winters of 2017 and 2018
Number of foods Dietary niche breadth Shannon-Wiener diversity
2017 2018 2017 2018 2017 2018
Siberian Crane 16 17 005 007 091 085
White-naped Crane 18 15 020 003 169 059
Hooded Crane 19 13 005 002 096 043
Eurasian Crane 18 19 008 009 110 121
Hou et al Frontiers in Zoology (2021) 1824 Page 6 of 17
that are overlooked by microhistological methods [6780] It also enables the identification of food items at ahigher taxonomic resolution [69 81] However we ac-knowledge that our study has two limitations First weused fecal samples as they are non-invasive Neverthe-less fecal analysis is subjected to the biases caused bythe differential digestive rates of food items [82 83] Softfoods such as insect larvae tend to be under-representedin fecal samples because they are digested quickly andremain in gastro-intestinal tract for short periods whilehard foods such as mollusks tend to be over-representedbecause they break down slowly and remain in gastro-intestinal tracts longer Thus food proportions in the
feces may not correlate with food proportions ingestedby cranes In addition some materials identified in thefeces may be incidentally consumed by cranes and notreally be used as food Second our study only consideredthe plant component of cranesrsquo diets as plants were sug-gested to be their principal food types at Poyang LakeAnimal foods consumed by the four crane species butto a much lesser extent than plant foods [84ndash87] werenot considered in our studyThe Siberian Crane is regarded as the most aquatic of
all cranes using wetlands for nesting feeding and roost-ing [49 88] The high dependence of the species onaquatic habitats was suggested to make it vulnerable to
Table 3 The observed dietary niche overlap among the four crane species at Poyang Lake in the winters of 2017 and 2018 P(Obs lt = null) and P (Obs gt = null) indicate the tail probabilities for the observed index which were lower and higher than thehistogram of the simulated index respectively represents P lt 005 and represents P lt 001
Winter Observed niche overlap index P (Obs lt = null) P (Obs gt = null)
2017 Siberian vs White-naped 021 0819 0181
Siberian vs Hooded 002 0703 0297
Siberian vs Eurasian 016 0900 0100
White-naped vs Hooded 049 0952 0048
White-naped vs Eurasian 035 0895 0106
Hooded vs Eurasian 035 0919 0081
2018 Siberian vs White-naped 063 0952 0048
Siberian vs Hooded 066 0952 0048
Siberian vs Eurasian 062 0950 0049
White-naped vs Hooded 099 0997 0003
White-naped vs Eurasian 089 0987 0013
Hooded vs Eurasian 090 0998 0002
Fig 4 The results of nonmetric multidimensional scaling ordination (NMDS) based on Bray-Curtis dissimilarity The colors of the circlescorrespond to the crane species blue Siberian Crane purple White-naped Crane green Hooded Crane and red Eurasian Crane
Hou et al Frontiers in Zoology (2021) 1824 Page 7 of 17
impacts of habitat degradation and to lead its classifica-tion as Critically Endangered [49] At Poyang Lake Si-berian Cranes traditionally fed on tubers of Vallisneriathe dominant submerged macrophytes in shallow watersand mudflats [89 90] In winter 2010 due to a flood-induced Vallisneria tuber collapse they were observedforaging for P limprichtii taproots and T edulis bulbs ingrasslands for the first time [38 44] In the winters from2015 to 2017 the diets of Siberian Cranes changedagain with thousands of cranes feeding on rice seedsand lotus rhizomes in agricultural habitats [61 62 64]Our study identified 22 plant items in the feces of Siber-
ian Cranes These plant items were obtained from shallowwaters grasslands and agricultural fields Four plant itemswith yearly RRA gt 10 were identified in the feces Vallis-neria tuber P criopolitanum rhizomes rice seeds andlotus rhizomes (the vegetative portions consumed bycranes were based on our field observation) SiberianCranesrsquo diet compositions showed substantial interannualvariation When the abundance of Vallisneria tubers waslow (ie winter 2017) Siberian Cranes mainly fed on riceseeds lotus rhizomes and P criopolitanum rhizomes inagricultural fields and grasslands When the abundance ofVallisneria tubers was high (ie winter 2018) they mainlyfed on Vallisneria tubers and P criopolitanum rhizomesin shallow waters and grasslands Our study in combin-ation with previous studies indicates that the diets of Si-berian Cranes have changed greatly since winter 2010 [3844 62] They have gradually broadened their dietaryniches which has probably been driven by the degradationof submerged plants [38 44 57]Similar to Siberian Cranes a diet shift was also ob-
served in White-naped Cranes Historically White-naped Cranes only fed on submerged plants in shallowwaters and mudflats of Poyang Lake [91] Then drivenby the decline of submerged plants they primarily fed atgrasslands and mudflats [87 91] consuming Vallisneriatuber T edulis P limprichtii and Ranunculus polii [5587] Our results indicate that in addition to T edulisand P limprichtii that had previously been reported theproportions of P criopolitanum and Carex were alsohigh in the feces Hooded Cranes were suggested tomainly feed on the grassland plant P limprichtii throughdirect observation [86] Our study indicates that thegrassland plants P criopolitanum and Carex also occu-pied high proportions in the feces The dominant fooditems in the feces of White-naped Cranes and HoodedCranes varied greatly between the two winters The vari-ation in dietary composition indicated that the diversityand composition of the grassland plant communitymight have changed greatly between the two wintersThe dietary niche overlap between White-naped Cranesand Hooded Cranes was the highest among the fourcrane species aligning with their level of habitat niche
overlap which was also the highest among the species[92] The high dietary and habitat niche overlaps suggesthigh competition potential between the two cranespeciesThe Eurasian Crane is a habitat generalist It is known
to feed in a variety of habitats including grasslandsmudflats shallow waters and agricultural fields [88 92]Its diet at Poyang Lake has not been documented previ-ously Our study indicates that P criopolitanum P lim-prichtii and rice were the dominant food items in thefeces of Eurasian Cranes The number of plant itemsconsumed dietary niche breadth and Shannon-Wienermeasures were the highest among the four crane speciessupporting its generalist foraging strategy In contrast tothe three crane species above Eurasian Cranesrsquo dietcompositions were similar between the two winters Riceand grassland plants occupied high proportions in thefeces in both winters
Important food resources for cranesVallisneria tubers have been acknowledged as importantfood sources for cranes especially Siberian Cranes andWhite-naped Cranes [55 56] Our study confirms theimportance of Vallisneria tubers in the diet of SiberianCranes It was the most common food source in thefeces of Siberian Cranes in winter 2018 when its abun-dance was high Vallisneria tuber proportions in thefeces of the three other crane species were not as high asthat in the feces of Siberian Cranes but it still occupied5ndash9 of the total foods in winter 2018To our surprise P criopolitanum was found to be the
first dominant food in the feces of White-naped CranesHooded Cranes and Eurasian Cranes and the seconddominant food in the feces of Siberian Cranes in winter2018 In winter 2017 the proportions of P criopolita-num were lt 17 in the feces of Siberian Cranes andWhite-naped Cranes and were lt 3 in the feces ofHooded Cranes and Eurasian Cranes The higher pro-portions of P criopolitanum in winter 2018 than winter2017 indicate that the abundance and availability of Pcriopolitanum was much higher in winter 2018 P crio-politanum is an annual grassland plant It is sensitive tohydrological conditions and can only survive in a rela-tively narrow range of water depths and percent time in-undated [93] Due to the sensitivity its distributionrange and abundance showed high interannual variation[94 95] Thus it is likely that the abundance of P crio-politanum was higher in winter 2018 than in winter2017The importance of P criopolitanum as a food source
for the cranes at Poyang Lake has not been documentedpreviously It has only been suggested to be a major foodsource of Hooded Cranes at Shengjin Lake another lakein the middle and lower Yangtze River floodplain [86
Hou et al Frontiers in Zoology (2021) 1824 Page 8 of 17
96] P criopolitanum is a dominant wetland plant at Po-yang Lake [93 97] The coverage percentage of this plantis the third highest among grassland plants accountingfor 12 of the total areas of grasslands [97] This plantspecies is usually distributed at elevations of 9ndash12mabove sea level near mudflats and shallow waters [97]which are frequently used by cranes Its rhizome with ashallow burial depth is high in energy and nutritionalcontent and low in crude fibre [38 86] All these charac-teristics make P criopolitanum rhizome a potentially im-portant food source for cranes We frequently observedcranes dig and feed on rhizomes of P criopolitanum andleave aboveground parts in the fields Therefore we be-lieve that P criopolitanum was used as food not inci-dentally consumed by cranes However we need to becaution that high proportion of P criopolitanum in thefeces does not mean high proportion in the diets Morework on the importance of P criopolitanum in the dietsof cranes is needed
The roles of agricultural fields in crane protectionWith the loss and degradation of natural wetlands theroles of artificial wetlands in waterbird protection havebeen under intensive debate [98 99] Although agricul-tural fields cannot completely replace the function ofnatural wetlands as waterbird habitats [100ndash102] agri-cultural fields might provide alternative or complemen-tary habitats for waterbirds in all life stages [38 103104] The proportions of domesticated species in thefeces of Siberian Cranes varied considerably betweenyears In winter 2017 when the abundance of Vallisneriatubers was low the proportion of Vallisneria in SiberianCranesrsquo feces was only 095 Siberian Cranes may nothave found enough foods in the shallow waters andmudflats and were forced to search for rice seeds andlotus rhizomes in agricultural fields In winter 2018when the abundance of Vallisneria tubers reboundedSiberian Cranes moved back to shallow waters and mud-flats and a large proportion of their traditional food Val-lisneria tubers and also P criopolitanum were detectedin the feces A similar abandonment of tuber foragingoccurred following the tuber collapse in 2010 and thenresumed in the next winter when tubers rebounded in2011 [38 44] The diet and habitat changes indicate thatagricultural habitats were important refuges for SiberianCranes providing important food resources for them tosurvive through winters with few Vallisneria tubersThey also indicate that Siberian Cranes prefer naturalwetlands to agricultural fields [38]Similar to Siberian Cranes White-naped Cranes and
Hooded Cranes consumed rice seeds and lotus rhizomesin winter 2017 but did not consume any in winter 2018The changes in the proportions suggest that the abun-dance of food resources in natural wetlands might have
been low in winter 2017 The deficient food resourcesmight have been Vallisneria tubers but might also havebeen grassland plants the primary food sources of thesetwo crane species Under natural food shortage condi-tions (ie winter 2017) White-naped Cranes andHooded Cranes utilized agricultural fields When theabundance of Vallisneria tubers and probably grasslandplants were high (ie winter 2018) the two crane speciesreturned to their natural habitats Therefore for White-naped Cranes and Hooded Cranes agricultural fieldswere important alternative foraging habitats used asbuffers against starvation during periods of natural foodshortages The return of the two crane species to naturalwetlands in winter 2018 further emphasizes the superiorquality of natural habitats over agricultural fields as for-aging habitats for cranes [38]The proportions of rice seeds in the feces of Eur-
asian Cranes were high in the two winters thoughthe proportion in winter 2017 was twice as high asthat in winter 2018 Therefore agricultural fields ap-peared to be consistently important as foraging habi-tats for Eurasian Cranes The frequent use ofagricultural fields by Eurasian Cranes has been docu-mented widely [49 105 106]Domesticated species generally contain high energy
and nutritional content and are of superior quality com-pared to natural plants [86 104] Farmland foraging hasbeen suggested to contribute to increase in the abun-dance of some goose [107] egret [108] and wader [109]populations in Japan Korea Europe and North Amer-ica However cranes generally avoid human disturbancewhen select foraging sites [110 111] Human disturb-ance in agricultural fields around Poyang Lake is consid-erably high due to intensive agricultural activity [112]High human disturbance was suggested to be a majorreason that wintering geese were almost confined withinnatural wetlands and hesitated to exploit the riches ofthe modern agricultural habitats at Poyang Lake [112113] High human disturbance also resulted in SiberianCranes spending twice as much time alerting in agricul-tural fields than they did in natural habitats [114] Thehigh level of human disturbance in agricultural fieldsmight contribute to the preference for natural wetlandsby Siberian Cranes White-naped Cranes and HoodedCranes at Poyang Lake Moreover although agriculturefields contain energy-rich crops wetlands contain di-verse food resources especially protein-rich inverte-brates which might better satisfy dietary needs of cranes[110 115] Taken together security and diverse food re-sources might have attracted cranes to select wetlands asforaging sitesSubmerged macrophytes at Poyang Lake have de-
graded in recent decades [57ndash60] and Vallisneria tuberdisappearance is expected to occur more frequently
Hou et al Frontiers in Zoology (2021) 1824 Page 9 of 17
under future climate change [57 60] Cranes may in-creasingly depend upon agricultural fields around Po-yang Lake which may in turn cause significant cropdamage and economic losses This is especially true onagricultural fields surrounding protected wetlands wherewaterbirds concentrate in high numbers [116 117] AtPoyang Lake cranes mainly fed on post-harvest remainsof rice seeds and lotus rhizomes thus crop damage andeconomic loss were limited However we observed somefarmers scared cranes away from their agricultural fieldsThere were more than 14 million free-ranging poultryraised at agricultural fields around Poyang Lake [118]where they gleaned waste grain residues in stubble fieldsCrane feeding would reduce the food abundance of thefree-ranging domestic poultry thus farmers drovecranes away To reduce human disturbance environ-mental education is needed because farmers are morelikely to protect cranes if they understand the endan-gered degree and the ecological significance of cranes
Effects of tuber collapse on dietary niche breadth andoverlap among crane speciesVallisneria tuber is an important food source for Siber-ian Cranes (89 90 this study) thus variation in itsabundance might influence Siberian Cranesrsquo dietaryniche breadth The numbers of food items dietary nichebreadth and Shannon-Wiener measures of SiberianCranes were similar between winter 2017 and winter2018 This is inconsistent with the contention that dur-ing seasons with a low abundance of dominant food thedietary niche may broaden as consumers relying on in-sufficient preferred food items are forced to add lessprofitable resources to their diets [26 119 120] In win-ter 2017 with low Vallisneria tuber abundance SiberianCranes left shallow waters and mudflats and moved torice paddies and lotus ponds [61 62 64] Their foodschanged from Vallisneria tuber to rice seeds and lotusrhizomes almost completely As the energy protein andfat contents of crops are generally as good as or superiorto natural foods and are often present in agriculturalfields in far greater abundance [86 104] Siberian Cranesmay gain enough energy through feeding on crops anddo not need to add many other food sources Moreoverthe Siberian Crane is a dietary specialist [89 90] PoyangLake has a relatively simple plant community compos-ition with Vallisneria and Hydrilla verticillate being thedominant submerged macrophytes [59] and Carex beingthe dominant grassland plants [121] Therefore theremay be few options for Siberian Cranes which mightlead to the failure of this species to widen its dietaryniche breadthIn contrast to Siberian Cranes the numbers of food
items dietary niche breadths and Shannon-Wiener mea-sures of White-naped Cranes and Hooded Cranes were
higher in winter 2017 than in winter 2018 P criopolita-num was the predominant food item in the feces of thetwo crane species in winter 2018 and the proportions ofVallisneria tubers were also high The abundances ofVallisneria tubers and probably P criopolitanum at Po-yang Lake declined greatly in winter 2017 When thereis a short-fall in food availability animals may optimizeforaging behavior and increase their dietary breath [122123] The short-fall in the abundances of P criopolita-num and Vallisneria tubers in winter 2017 might havedriven the two crane species to add less profitable grass-land plants in their diets Although the two cranes alsoconsumed high-quality rice seeds and lotus rhizomes inwinter 2017 their proportions were low in the feces andthese foods might be unable to meet the energy require-ments of the two crane species The niche expansion ofthe two crane species during periods of preferred foodshortage was in agreement with the basic prediction ofoptimal foraging theory [22 23] It is also consistentwith a commonly observed fact that energy-maximizingmodels work better when animals food-limited [124]The dietary niche overlaps among the crane species
were low in winter 2017 with the four crane species di-verging in their principal food items However in winter2018 the niche overlaps were significantly higher thanexpected by chance They had similar diet compositionswith P criopolitanum rhizomes and Vallisneria tubersbeing the principal food items in the feces The in-creased overlap might have been caused by the highabundances of the two plant species in winter 2018which allowed the four crane species to share the samefood items with low interspecific competition Our studyis consistent with previous research that suggested thatamong-species dietary niches tended to overlap moreduring seasons of high food abundance [32 33 125126] Greater resource availability could promote an in-crease in species utilization of similar resources and re-duce competitive interactions between species [32 34126] By contrast during seasons with low food abun-dance species may partition limited food resources todecrease interspecific competition leading to a decreasein niche overlap [126 127] It has been suggested thatdiet partitioning facilitates the coexistence of manyclosely related species [7 128] The interannual variationin niche overlap observed in our study suggests that dietpartitioning may be restricted to specific temporal casesIn seasons or years with abundant food sources inter-specific competition may be relaxed and niche overlapcould be high
ConclusionsIn summary by using metabarcoding technology ourstudy gained new insights into cranesrsquo diets that are es-sential for informing habitat management to improve
Hou et al Frontiers in Zoology (2021) 1824 Page 10 of 17
the availability of foraging resources Vallisneria tuberswas confirmed as an important food source of cranesSurprisingly P criopolitanum also occupied high pro-portions in the feces of cranes and may also be import-ant Our study emphasizes the superior quality ofnatural wetlands compared to agricultural fields as for-aging habitats for cranes The recent degradation of sub-merged macrophytes at Poyang Lake potentiallythreatens the survival of cranes especially SiberianCranes Therefore it is urgent to reverse the broad de-cline of submerged macrophytes and provide importantforaging habitats for cranes during years of tuber col-lapses The abundances of Vallisneria tubers and prob-ably P criopolitanum at Poyang Lake greatly influencedthe dietary niche width and overlaps among crane spe-cies Cranes exhibited low dietary niche overlap duringperiods of preferred food shortage but significantly highoverlap during periods when preferred foods were plen-tiful While high niche overlap is often interpreted as in-tense interspecific competition our study highlights theimportance of taking food abundance into account
Materials and methodsStudy areasPoyang Lake the largest freshwater lake in China is oneof the most important wintering areas for waterbirdsalong the East Asian-Australasian Flyway (Fig 1) [51129] It is characterized by dramatic seasonal hydro-logical fluctuations with an average change of 924 mbetween the summer high and winter low water levels[130] During summer the inundation area is gt 2500km2 [131] In autumn and winter the water level dropscausing the inundation area to shrink to lt 1000 km2
[131] Poyang Lake contains 102 sublakes which are in-undated and integrated with the main body of PoyangLake during summer and are isolated during winter[132] As the water level drops the sublakes appear andlarge areas of grasslands mudflats and shallow waterareas are exposed providing winter foraging and roost-ing areas for approximately 420000 waterbirds from ap-proximately 111 species [133 134]
Vallisneria tuber surveyThe density and biomass of Vallisneria tubers were sur-veyed at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake in late October and earlyNovember of 2017 and 2018 before large numbers ofwintering waterbirds arrived We set 76 183 and 36sampling plots uniformly located at Dahuchi Shahu andChanghuchi respectively Vallisneria tubers were col-lected from two quadrats in each plot with a locallymade steel grab sampler The sampler has two long
handles At the end of the handles there are two scoopsfacing each other The sampler was inserted into thesubstrate and the scoops brought together to collectabout 15 cm long times 13 cm wide times 30 cm high sample ofsubstrate Tubers were subsequently cleaned andcounted The dry weight of tubers in each quadrat wasweighed after oven-drying at 80 degC until constant weightThe tuber density and biomass in each plot was calcu-lated by dividing total tuber number and tuber massacross two quadrats by quadrat area respectively Weused t-test to explore whether the tuber density and bio-mass in the winters of 2017 and 2018 significantly devi-ated from the historical average values in the wintersfrom 1999 to 2016 [57] The historical tuber density andbiomass data were collected by the Poyang Lake Na-tional Nature Reserve at three sublakes (Dahuchi Shahuand Meixihu) [57] R 360 was used to do the t-testanalyses
Fecal sample collectionFecal samples were collected at Poyang Lake in the win-ters of 2017 and 2018 We first investigated the distribu-tions of cranes When we saw foraging cranes we waiteduntil they left to avoid disturbing them Soon after theyleft we went to the foraging sites to collect fresh feces(ie feces with wet surface) with sterilized tweezers Tominimize the probability of multiple samples from thesame individual all collected samples were separated byat least 5 m We stored the samples in liquid nitrogen inthe field and transferred them to a minus 80 degC refrigeratorfor long-term storage in the laboratory To ensure thatthe sources of the feces were known we tried to collectsamples from monospecific flocks For samples collectedfrom mixed-species flocks we used DNA analysis toidentify the species that deposited the fecal samplesPlants were also collected in the field to construct aDNA barcoding reference database
Species identificationBecause of the similar morphologies of the feces of thefour crane species we used DNA analysis to identify thespecies that deposited the fecal samples collected frommixed-species flocks For the genetic species assignmentof each fecal sample we scraped the external surfacewhich contain DNA from the host species as a result ofthe sloughing of cells from the digestive tract DNA wasextracted with the QIAamp Power Fecal DNA Kit (Qia-gen USA) according to the manufacturerrsquos guidelinesFor each round of DNA extraction negative controls(ie extraction without feces) were included to monitorfor possible contamination We amplified the mitochon-drial DNA D-loop fragment with the primer pair DL02F
Hou et al Frontiers in Zoology (2021) 1824 Page 11 of 17
(5prime-3prime GCGGCCCGAAAAGCCGCTG) and DL02R (5prime-3prime GCCCTGACATAGGAACCAGAGGCGC) [135]PCR amplifications were carried out in a total volume of25 μl containing 125 μl PCR mix (Tiangen BeijingChina) 1 μl DNA 1 μl each primer and 95 μl H2O Thereaction conditions were as follows denaturation at94 degC for 4 min followed by 34 cycles at 94 degC for 48 s56 degC for 48 s and 72 degC for 1 min and a final 10 min at72 degC We checked for the presence of a PCR product ofsuitable length by electrophoresis on a 1 agarose gelSequencing was carried out on an ABI 3730xl analyzerby Sangon Biotech (Shanghai China) The complete se-quences were assembled using Seqman II (DNASTAR)and compared visually to the original chromatograms toavoid reading errors The taxonomic assignment wasconducted by searching against the nucleotide databaseof GenBank using Web BLAST
Construction of a barcode reference databaseWe downloaded the trnL sequences of 48 plant speciescommonly seen at Poyang Lake from GenBank For theplant species not included in GenBank we sequencedtrnL by ourselves Total DNA was extracted using thehexadecyltrimethylammonium bromide (CTAB) proto-col We used primer pair c (5prime-3prime CGAAATCGGTAGACGCTACG) and h (5prime-3prime CCATTGAGTCTCTGCACCTATC) to amplify an approximately 150 bp regionof the chloroplast trnL intron [136] This primer pairwas chosen as it can amplify DNA from a wide range ofwetland plants [70] PCR amplifications were carried outin a total volume of 25 μl containing 125 μl PCR mix(Tiangen Beijing China) 1 μl DNA 1 μl each primerand 95 μl H2O The reaction conditions were as followsdenaturation at 94 degC for 4 min followed by 35 cycles at94 degC for 30 s 56 degC for 30 s and 72 degC for 45 s and afinal 10 min at 72 degC We used electrophoresis to checkthe PCR products ABI 3730xl analyzer to perform se-quencing and Seqman II to assemble the sequences
DNA extraction and high-throughput sequencing of fecalsamplesTo uncover plant component consumed by cranes amodified CTAB protocol [70] was used to extract theDNA from approximately 200 mg of the interior of eachfecal sample DNA extraction was conducted in a dedi-cated laboratory for analyses of DNA from samples withlow DNA quality For each batch of DNA extractionnegative controls were included to monitor for possiblecontamination The primers c and h with a 6 bp tagadded to 5prime ends of each primer were used to amplifytrnL gene The PCR amplification procedure was thesame as barcode database construction For each samplea unique combination of tags in the forward and reverseprimers was used so that sample-specific data can be
recovered after sequencing Three PCR replicates wereperformed for each sample to reduce amplification biasThe three replicates were pooled and the DNA concen-tration and purity were monitored on a 1 agarose gelOnly samples with a clear band were processed furtherSamples were purified using the SanPrep Column PCRProduct Purification Kit (Sangon Biotech ShanghaiChina) The purified PCR products were pooled takinginto account of DNA concentration to obtain an ap-proximately equal amount of amplicon DNA from eachfecal sample The pool of individually tagged ampliconswas used to prepare a sequencing library using the Tru-Seq DNA PCR-Free Sample Preparation Kit (IlluminaUSA) following manufacturerrsquos recommendations Thelibrary was sequenced on an Illumina HiSeq 2500 plat-form and 250 bp paired-end reads were generated Li-brary preparation and sequencing were conducted byNovogene (Beijing China)
Sequencing data analysisAfter high-throughput sequencing the paired-end readswere demultiplexed into sample-specific files based ontheir unique tags The tags and primer sequences weresubsequently trimmed The paired-end reads weremerged using FLASH 127 [137] and then filtered usingQIIME 170 [138] Sequences were dereplicated andclustered to OTUs using USEARCH 110667 [139] atthe similarity threshold of 97 All OTUs were assignedto taxonomic units by referring to the local plant data-base using blast 2231+ with thresholds of identity gt97 and e-value lt 10 eminus 50 If an OTU matched two ormore taxa it was assigned to a higher taxonomic levelthat included all taxaTo evaluate the sufficiency of sequencing depth and
sampling size individual- species- and sample-basedrarefaction curves were built using EstimateS 82[140] The curves were computed by randomly resam-pling sequences or samples and plotting these againstthe numbers of OTUs The read abundance data wereconverted to RRA of each food item using the follow-ing equation [79]
RRA frac14 1S
XS
Kfrac141
nikPTifrac141nik
100
where nik is the number of sequences of food item i insample k T is the total number of food items and S isthe sampling size
Dietary niche breadth and overlap among crane speciesTo explore whether the cranesrsquo diets showed substantialinterannual variation we calculated Pearson correlationsbetween diet compositions of each crane species in win-ter 2017 and winter 2018 The proportion of each food
Hou et al Frontiers in Zoology (2021) 1824 Page 12 of 17
item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-
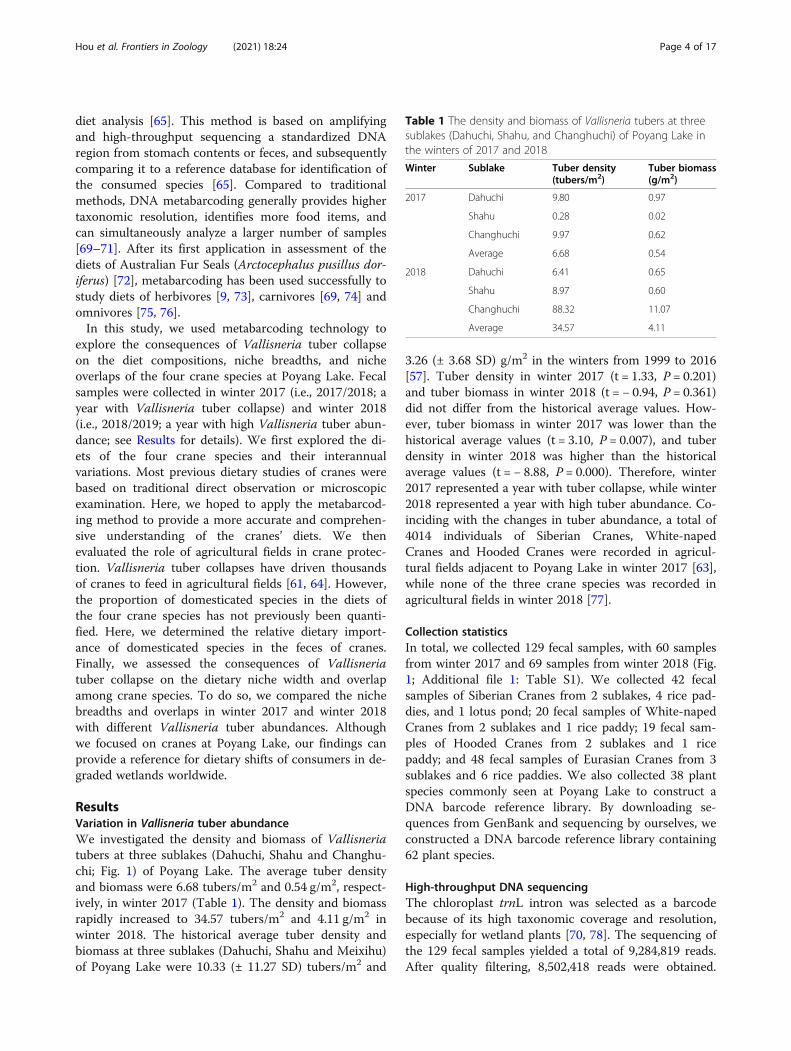
diet analysis [65] This method is based on amplifyingand high-throughput sequencing a standardized DNAregion from stomach contents or feces and subsequentlycomparing it to a reference database for identification ofthe consumed species [65] Compared to traditionalmethods DNA metabarcoding generally provides highertaxonomic resolution identifies more food items andcan simultaneously analyze a larger number of samples[69ndash71] After its first application in assessment of thediets of Australian Fur Seals (Arctocephalus pusillus dor-iferus) [72] metabarcoding has been used successfully tostudy diets of herbivores [9 73] carnivores [69 74] andomnivores [75 76]In this study we used metabarcoding technology to
explore the consequences of Vallisneria tuber collapseon the diet compositions niche breadths and nicheoverlaps of the four crane species at Poyang Lake Fecalsamples were collected in winter 2017 (ie 20172018 ayear with Vallisneria tuber collapse) and winter 2018(ie 20182019 a year with high Vallisneria tuber abun-dance see Results for details) We first explored the di-ets of the four crane species and their interannualvariations Most previous dietary studies of cranes werebased on traditional direct observation or microscopicexamination Here we hoped to apply the metabarcod-ing method to provide a more accurate and comprehen-sive understanding of the cranesrsquo diets We thenevaluated the role of agricultural fields in crane protec-tion Vallisneria tuber collapses have driven thousandsof cranes to feed in agricultural fields [61 64] Howeverthe proportion of domesticated species in the diets ofthe four crane species has not previously been quanti-fied Here we determined the relative dietary import-ance of domesticated species in the feces of cranesFinally we assessed the consequences of Vallisneriatuber collapse on the dietary niche width and overlapamong crane species To do so we compared the nichebreadths and overlaps in winter 2017 and winter 2018with different Vallisneria tuber abundances Althoughwe focused on cranes at Poyang Lake our findings canprovide a reference for dietary shifts of consumers in de-graded wetlands worldwide
ResultsVariation in Vallisneria tuber abundanceWe investigated the density and biomass of Vallisneriatubers at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake The average tuber densityand biomass were 668 tubersm2 and 054 gm2 respect-ively in winter 2017 (Table 1) The density and biomassrapidly increased to 3457 tubersm2 and 411 gm2 inwinter 2018 The historical average tuber density andbiomass at three sublakes (Dahuchi Shahu and Meixihu)of Poyang Lake were 1033 (plusmn 1127 SD) tubersm2 and
326 (plusmn 368 SD) gm2 in the winters from 1999 to 2016[57] Tuber density in winter 2017 (t = 133 P = 0201)and tuber biomass in winter 2018 (t = minus 094 P = 0361)did not differ from the historical average values How-ever tuber biomass in winter 2017 was lower than thehistorical average values (t = 310 P = 0007) and tuberdensity in winter 2018 was higher than the historicalaverage values (t = minus 888 P = 0000) Therefore winter2017 represented a year with tuber collapse while winter2018 represented a year with high tuber abundance Co-inciding with the changes in tuber abundance a total of4014 individuals of Siberian Cranes White-napedCranes and Hooded Cranes were recorded in agricul-tural fields adjacent to Poyang Lake in winter 2017 [63]while none of the three crane species was recorded inagricultural fields in winter 2018 [77]
Collection statisticsIn total we collected 129 fecal samples with 60 samplesfrom winter 2017 and 69 samples from winter 2018 (Fig1 Additional file 1 Table S1) We collected 42 fecalsamples of Siberian Cranes from 2 sublakes 4 rice pad-dies and 1 lotus pond 20 fecal samples of White-napedCranes from 2 sublakes and 1 rice paddy 19 fecal sam-ples of Hooded Cranes from 2 sublakes and 1 ricepaddy and 48 fecal samples of Eurasian Cranes from 3sublakes and 6 rice paddies We also collected 38 plantspecies commonly seen at Poyang Lake to construct aDNA barcode reference library By downloading se-quences from GenBank and sequencing by ourselves weconstructed a DNA barcode reference library containing62 plant species
High-throughput DNA sequencingThe chloroplast trnL intron was selected as a barcodebecause of its high taxonomic coverage and resolutionespecially for wetland plants [70 78] The sequencing ofthe 129 fecal samples yielded a total of 9284819 readsAfter quality filtering 8502418 reads were obtained
Table 1 The density and biomass of Vallisneria tubers at threesublakes (Dahuchi Shahu and Changhuchi) of Poyang Lake inthe winters of 2017 and 2018
Winter Sublake Tuber density(tubersm2)
Tuber biomass(gm2)
2017 Dahuchi 980 097
Shahu 028 002
Changhuchi 997 062
Average 668 054
2018 Dahuchi 641 065
Shahu 897 060
Changhuchi 8832 1107
Average 3457 411
Hou et al Frontiers in Zoology (2021) 1824 Page 4 of 17
The average number of reads per sample was 65910 (plusmn26276 SD) and the average number of operational taxo-nomic units (OTUs) per sample was 43 (plusmn 15 SD) Toevaluate the sequencing and sampling adequacy individ-ual- species- and sample-based rarefaction curves werebuilt With the sample sequencing reads increased theindividual-based rarefaction curves reached plateau inmost cases (Additional file 1 Figure S1) and the species-based rarefaction curves reached plateau in the fourcrane species (Fig 2a) Therefore our sequencing depthwas roughly adequate to capture the numbers of OTUspresent The sample-based rarefaction curves for eachcrane species reached plateau in all cases as the samplingsizes increased (Fig 2b) suggesting that we acquired suf-ficient fecal samples After comparison to the local DNAreference database 9899 of the total reads wereassigned to specific plant taxa Most of the reads thatwere not assigned to specific plant taxa had low fre-quency In total we identified 29 plant taxa including23 species 4 genera and 2 families
Diet compositions of the four crane speciesWe used relative read abundance (ie the percentageof DNA belonging to each food item in each sampleRRA) which provides a more accurate view of speciesdiet than frequency of occurrence (ie the number ofsamples that contain a given food item) tosummarize dietary data [79] Twenty-two food itemswere identified in the feces of Siberian Cranes in thetwo winters including 17 species 3 genera and 2families (Fig 3 Additional file 1 Table S2) Rice(RRA = 7285) lotus (1201) and Polygonum criopo-litanum (1199) were the dominant food items inthe feces (ie proportion gt 10) in winter 2017 andVallisneria (5612) and P criopolitanum (4087)were the dominant food items in winter 2018Twenty-five food items were identified in the feces ofWhite-naped Cranes including 20 species 3 genera
and 2 families Tulipa edulis (3737) Potentilla lim-prichtii (1869) P criopolitanum (1652) andCarex spp (1075) were the dominant food items inthe feces in winter 2017 and P criopolitanum(8441) was the dominant food item in winter 2018Twenty-two food items were identified in the feces ofHooded Cranes including 18 species 3 genera and 1family P limprichtii (6536) and Carex (2739)were the dominant food items in the feces in winter2017 and P criopolitanum (8876) was the domin-ant food item in winter 2018 Twenty-two food itemswere identified in the feces of Eurasian Cranes in-cluding 17 species 3 genera and 2 families Rice(6120) and P limprichtii (2394) were the domin-ant food items in the feces in winter 2017 and Pcriopolitanum (5577) and rice (2740) were thedominant food items in winter 2018 Diet composi-tions of Siberian Cranes (Pearson correlation = 005P = 077) White-naped Cranes (Pearson correlation =030 P = 008) and Hooded Cranes (Pearson correl-ation = minus 004 P = 083) were not correlated betweenwinter 2017 and winter 2018 suggesting great inter-annual variations in food sources Diet composition ofEurasian Cranes was correlated (Pearson correlation =040 P = 002) between the two winters suggestinglimited interannual variation in food sourcesVallisneria tubers were an important food source for
the four crane species especially Siberian Cranes inwinter 2018 The proportion of Vallisneria was 5612512 850 and 734 in the feces of Siberian CranesWhite-naped Cranes Hooded Cranes and EurasianCranes respectively in winter 2018 (Fig 3 Additionalfile 1 Table S2) In winter 2017 when Vallisneria tuberabundance was low the proportion of Vallisneria wasless than 1 in the feces of the four crane speciesDomesticated species identified in crane feces included
rice and lotus The proportions of domesticated speciesin the feces of Siberian Cranes (8485) and White-
Fig 2 a Species-based and b sample-based rarefaction curves The species- and sample-based rarefaction curves built by randomly resamplingsequences and samples respectively at increasing levels of accumulation indicate whether the sequencing depth and sampling size weresufficient to characterize the diversity of dietary items The colors of the curves correspond to the crane species blue Siberian Crane purpleWhite-naped Crane green Hooded Crane and red Eurasian Crane
Hou et al Frontiers in Zoology (2021) 1824 Page 5 of 17
naped Cranes (1509) were high in winter 2017 but de-clined to 000 in winter 2018 (Fig 3 Additional file 1Table S2) Domesticated species occupied a small pro-portion in the feces of Hooded Cranes in winter 2017(130) but declined to 000 in winter 2018 The pro-portions of domesticated species in the feces of EurasianCranes were high in both winters with 6120 in winter2017 and 2740 in winter 2018
Dietary niche breadth and overlap among crane speciesThe numbers of food items dietary niche breadth andShannon-Wiener diversity values for Siberian Cranesand Eurasian Cranes in winter 2017 were similar tothose in winter 2018 (Table 2) Values of all three pa-rameters for White-naped Cranes and Hooded Craneswere higher in winter 2017 than in winter 2018 Dietaryniche breadth of the four crane species was lt 020 inboth winters indicating low niche breadthDietary niche overlap among crane species ranged
from 002 to 049 in winter 2017 but increased to 062to 099 in winter 2018 (Table 3) Nonmetric multidimen-sional scaling (NMDS) analysis also indicated overlap
increases in winter 2018 (Fig 4) No overlap values werelower than expected by chance in the two winters Theoverlap values were not higher than expected by chancein winter 2017 However all values showed a high de-gree of overlap in winter 2018 [P (Obs gt = null) lt 005]Overlap between White-naped Cranes and HoodedCranes was the highest among the four crane species inboth winters
DiscussionDiet compositionTo the best of our knowledge our study is the first toexplore crane diets using metabarcoding techniques Al-most all previous studies have relied on traditional directobservation or microhistological methods By usingmetabarcoding technology we were able to elucidatecrane diets at greater detail than previously known Theimproved knowledge might be due to the ability ofmetabarcoding to provide a more comprehensive per-spective and more details on food sources than trad-itional methods [66 67 80] Metabarcoding is able todetect a variety of smaller and softer bodied food items
Fig 3 Food items in the feces of Siberian Cranes White-naped Cranes Hooded Cranes and Eurasian Cranes in the winters of 2017 and 2018 atPoyang Lake
Table 2 Number of food items dietary niche breadth and Shannon-Wiener diversity values for Siberian Cranes White-napedCranes Hooded Cranes and Eurasian Cranes at Poyang Lake in the winters of 2017 and 2018
Number of foods Dietary niche breadth Shannon-Wiener diversity
2017 2018 2017 2018 2017 2018
Siberian Crane 16 17 005 007 091 085
White-naped Crane 18 15 020 003 169 059
Hooded Crane 19 13 005 002 096 043
Eurasian Crane 18 19 008 009 110 121
Hou et al Frontiers in Zoology (2021) 1824 Page 6 of 17
that are overlooked by microhistological methods [6780] It also enables the identification of food items at ahigher taxonomic resolution [69 81] However we ac-knowledge that our study has two limitations First weused fecal samples as they are non-invasive Neverthe-less fecal analysis is subjected to the biases caused bythe differential digestive rates of food items [82 83] Softfoods such as insect larvae tend to be under-representedin fecal samples because they are digested quickly andremain in gastro-intestinal tract for short periods whilehard foods such as mollusks tend to be over-representedbecause they break down slowly and remain in gastro-intestinal tracts longer Thus food proportions in the
feces may not correlate with food proportions ingestedby cranes In addition some materials identified in thefeces may be incidentally consumed by cranes and notreally be used as food Second our study only consideredthe plant component of cranesrsquo diets as plants were sug-gested to be their principal food types at Poyang LakeAnimal foods consumed by the four crane species butto a much lesser extent than plant foods [84ndash87] werenot considered in our studyThe Siberian Crane is regarded as the most aquatic of
all cranes using wetlands for nesting feeding and roost-ing [49 88] The high dependence of the species onaquatic habitats was suggested to make it vulnerable to
Table 3 The observed dietary niche overlap among the four crane species at Poyang Lake in the winters of 2017 and 2018 P(Obs lt = null) and P (Obs gt = null) indicate the tail probabilities for the observed index which were lower and higher than thehistogram of the simulated index respectively represents P lt 005 and represents P lt 001
Winter Observed niche overlap index P (Obs lt = null) P (Obs gt = null)
2017 Siberian vs White-naped 021 0819 0181
Siberian vs Hooded 002 0703 0297
Siberian vs Eurasian 016 0900 0100
White-naped vs Hooded 049 0952 0048
White-naped vs Eurasian 035 0895 0106
Hooded vs Eurasian 035 0919 0081
2018 Siberian vs White-naped 063 0952 0048
Siberian vs Hooded 066 0952 0048
Siberian vs Eurasian 062 0950 0049
White-naped vs Hooded 099 0997 0003
White-naped vs Eurasian 089 0987 0013
Hooded vs Eurasian 090 0998 0002
Fig 4 The results of nonmetric multidimensional scaling ordination (NMDS) based on Bray-Curtis dissimilarity The colors of the circlescorrespond to the crane species blue Siberian Crane purple White-naped Crane green Hooded Crane and red Eurasian Crane
Hou et al Frontiers in Zoology (2021) 1824 Page 7 of 17
impacts of habitat degradation and to lead its classifica-tion as Critically Endangered [49] At Poyang Lake Si-berian Cranes traditionally fed on tubers of Vallisneriathe dominant submerged macrophytes in shallow watersand mudflats [89 90] In winter 2010 due to a flood-induced Vallisneria tuber collapse they were observedforaging for P limprichtii taproots and T edulis bulbs ingrasslands for the first time [38 44] In the winters from2015 to 2017 the diets of Siberian Cranes changedagain with thousands of cranes feeding on rice seedsand lotus rhizomes in agricultural habitats [61 62 64]Our study identified 22 plant items in the feces of Siber-
ian Cranes These plant items were obtained from shallowwaters grasslands and agricultural fields Four plant itemswith yearly RRA gt 10 were identified in the feces Vallis-neria tuber P criopolitanum rhizomes rice seeds andlotus rhizomes (the vegetative portions consumed bycranes were based on our field observation) SiberianCranesrsquo diet compositions showed substantial interannualvariation When the abundance of Vallisneria tubers waslow (ie winter 2017) Siberian Cranes mainly fed on riceseeds lotus rhizomes and P criopolitanum rhizomes inagricultural fields and grasslands When the abundance ofVallisneria tubers was high (ie winter 2018) they mainlyfed on Vallisneria tubers and P criopolitanum rhizomesin shallow waters and grasslands Our study in combin-ation with previous studies indicates that the diets of Si-berian Cranes have changed greatly since winter 2010 [3844 62] They have gradually broadened their dietaryniches which has probably been driven by the degradationof submerged plants [38 44 57]Similar to Siberian Cranes a diet shift was also ob-
served in White-naped Cranes Historically White-naped Cranes only fed on submerged plants in shallowwaters and mudflats of Poyang Lake [91] Then drivenby the decline of submerged plants they primarily fed atgrasslands and mudflats [87 91] consuming Vallisneriatuber T edulis P limprichtii and Ranunculus polii [5587] Our results indicate that in addition to T edulisand P limprichtii that had previously been reported theproportions of P criopolitanum and Carex were alsohigh in the feces Hooded Cranes were suggested tomainly feed on the grassland plant P limprichtii throughdirect observation [86] Our study indicates that thegrassland plants P criopolitanum and Carex also occu-pied high proportions in the feces The dominant fooditems in the feces of White-naped Cranes and HoodedCranes varied greatly between the two winters The vari-ation in dietary composition indicated that the diversityand composition of the grassland plant communitymight have changed greatly between the two wintersThe dietary niche overlap between White-naped Cranesand Hooded Cranes was the highest among the fourcrane species aligning with their level of habitat niche
overlap which was also the highest among the species[92] The high dietary and habitat niche overlaps suggesthigh competition potential between the two cranespeciesThe Eurasian Crane is a habitat generalist It is known
to feed in a variety of habitats including grasslandsmudflats shallow waters and agricultural fields [88 92]Its diet at Poyang Lake has not been documented previ-ously Our study indicates that P criopolitanum P lim-prichtii and rice were the dominant food items in thefeces of Eurasian Cranes The number of plant itemsconsumed dietary niche breadth and Shannon-Wienermeasures were the highest among the four crane speciessupporting its generalist foraging strategy In contrast tothe three crane species above Eurasian Cranesrsquo dietcompositions were similar between the two winters Riceand grassland plants occupied high proportions in thefeces in both winters
Important food resources for cranesVallisneria tubers have been acknowledged as importantfood sources for cranes especially Siberian Cranes andWhite-naped Cranes [55 56] Our study confirms theimportance of Vallisneria tubers in the diet of SiberianCranes It was the most common food source in thefeces of Siberian Cranes in winter 2018 when its abun-dance was high Vallisneria tuber proportions in thefeces of the three other crane species were not as high asthat in the feces of Siberian Cranes but it still occupied5ndash9 of the total foods in winter 2018To our surprise P criopolitanum was found to be the
first dominant food in the feces of White-naped CranesHooded Cranes and Eurasian Cranes and the seconddominant food in the feces of Siberian Cranes in winter2018 In winter 2017 the proportions of P criopolita-num were lt 17 in the feces of Siberian Cranes andWhite-naped Cranes and were lt 3 in the feces ofHooded Cranes and Eurasian Cranes The higher pro-portions of P criopolitanum in winter 2018 than winter2017 indicate that the abundance and availability of Pcriopolitanum was much higher in winter 2018 P crio-politanum is an annual grassland plant It is sensitive tohydrological conditions and can only survive in a rela-tively narrow range of water depths and percent time in-undated [93] Due to the sensitivity its distributionrange and abundance showed high interannual variation[94 95] Thus it is likely that the abundance of P crio-politanum was higher in winter 2018 than in winter2017The importance of P criopolitanum as a food source
for the cranes at Poyang Lake has not been documentedpreviously It has only been suggested to be a major foodsource of Hooded Cranes at Shengjin Lake another lakein the middle and lower Yangtze River floodplain [86
Hou et al Frontiers in Zoology (2021) 1824 Page 8 of 17
96] P criopolitanum is a dominant wetland plant at Po-yang Lake [93 97] The coverage percentage of this plantis the third highest among grassland plants accountingfor 12 of the total areas of grasslands [97] This plantspecies is usually distributed at elevations of 9ndash12mabove sea level near mudflats and shallow waters [97]which are frequently used by cranes Its rhizome with ashallow burial depth is high in energy and nutritionalcontent and low in crude fibre [38 86] All these charac-teristics make P criopolitanum rhizome a potentially im-portant food source for cranes We frequently observedcranes dig and feed on rhizomes of P criopolitanum andleave aboveground parts in the fields Therefore we be-lieve that P criopolitanum was used as food not inci-dentally consumed by cranes However we need to becaution that high proportion of P criopolitanum in thefeces does not mean high proportion in the diets Morework on the importance of P criopolitanum in the dietsof cranes is needed
The roles of agricultural fields in crane protectionWith the loss and degradation of natural wetlands theroles of artificial wetlands in waterbird protection havebeen under intensive debate [98 99] Although agricul-tural fields cannot completely replace the function ofnatural wetlands as waterbird habitats [100ndash102] agri-cultural fields might provide alternative or complemen-tary habitats for waterbirds in all life stages [38 103104] The proportions of domesticated species in thefeces of Siberian Cranes varied considerably betweenyears In winter 2017 when the abundance of Vallisneriatubers was low the proportion of Vallisneria in SiberianCranesrsquo feces was only 095 Siberian Cranes may nothave found enough foods in the shallow waters andmudflats and were forced to search for rice seeds andlotus rhizomes in agricultural fields In winter 2018when the abundance of Vallisneria tubers reboundedSiberian Cranes moved back to shallow waters and mud-flats and a large proportion of their traditional food Val-lisneria tubers and also P criopolitanum were detectedin the feces A similar abandonment of tuber foragingoccurred following the tuber collapse in 2010 and thenresumed in the next winter when tubers rebounded in2011 [38 44] The diet and habitat changes indicate thatagricultural habitats were important refuges for SiberianCranes providing important food resources for them tosurvive through winters with few Vallisneria tubersThey also indicate that Siberian Cranes prefer naturalwetlands to agricultural fields [38]Similar to Siberian Cranes White-naped Cranes and
Hooded Cranes consumed rice seeds and lotus rhizomesin winter 2017 but did not consume any in winter 2018The changes in the proportions suggest that the abun-dance of food resources in natural wetlands might have
been low in winter 2017 The deficient food resourcesmight have been Vallisneria tubers but might also havebeen grassland plants the primary food sources of thesetwo crane species Under natural food shortage condi-tions (ie winter 2017) White-naped Cranes andHooded Cranes utilized agricultural fields When theabundance of Vallisneria tubers and probably grasslandplants were high (ie winter 2018) the two crane speciesreturned to their natural habitats Therefore for White-naped Cranes and Hooded Cranes agricultural fieldswere important alternative foraging habitats used asbuffers against starvation during periods of natural foodshortages The return of the two crane species to naturalwetlands in winter 2018 further emphasizes the superiorquality of natural habitats over agricultural fields as for-aging habitats for cranes [38]The proportions of rice seeds in the feces of Eur-
asian Cranes were high in the two winters thoughthe proportion in winter 2017 was twice as high asthat in winter 2018 Therefore agricultural fields ap-peared to be consistently important as foraging habi-tats for Eurasian Cranes The frequent use ofagricultural fields by Eurasian Cranes has been docu-mented widely [49 105 106]Domesticated species generally contain high energy
and nutritional content and are of superior quality com-pared to natural plants [86 104] Farmland foraging hasbeen suggested to contribute to increase in the abun-dance of some goose [107] egret [108] and wader [109]populations in Japan Korea Europe and North Amer-ica However cranes generally avoid human disturbancewhen select foraging sites [110 111] Human disturb-ance in agricultural fields around Poyang Lake is consid-erably high due to intensive agricultural activity [112]High human disturbance was suggested to be a majorreason that wintering geese were almost confined withinnatural wetlands and hesitated to exploit the riches ofthe modern agricultural habitats at Poyang Lake [112113] High human disturbance also resulted in SiberianCranes spending twice as much time alerting in agricul-tural fields than they did in natural habitats [114] Thehigh level of human disturbance in agricultural fieldsmight contribute to the preference for natural wetlandsby Siberian Cranes White-naped Cranes and HoodedCranes at Poyang Lake Moreover although agriculturefields contain energy-rich crops wetlands contain di-verse food resources especially protein-rich inverte-brates which might better satisfy dietary needs of cranes[110 115] Taken together security and diverse food re-sources might have attracted cranes to select wetlands asforaging sitesSubmerged macrophytes at Poyang Lake have de-
graded in recent decades [57ndash60] and Vallisneria tuberdisappearance is expected to occur more frequently
Hou et al Frontiers in Zoology (2021) 1824 Page 9 of 17
under future climate change [57 60] Cranes may in-creasingly depend upon agricultural fields around Po-yang Lake which may in turn cause significant cropdamage and economic losses This is especially true onagricultural fields surrounding protected wetlands wherewaterbirds concentrate in high numbers [116 117] AtPoyang Lake cranes mainly fed on post-harvest remainsof rice seeds and lotus rhizomes thus crop damage andeconomic loss were limited However we observed somefarmers scared cranes away from their agricultural fieldsThere were more than 14 million free-ranging poultryraised at agricultural fields around Poyang Lake [118]where they gleaned waste grain residues in stubble fieldsCrane feeding would reduce the food abundance of thefree-ranging domestic poultry thus farmers drovecranes away To reduce human disturbance environ-mental education is needed because farmers are morelikely to protect cranes if they understand the endan-gered degree and the ecological significance of cranes
Effects of tuber collapse on dietary niche breadth andoverlap among crane speciesVallisneria tuber is an important food source for Siber-ian Cranes (89 90 this study) thus variation in itsabundance might influence Siberian Cranesrsquo dietaryniche breadth The numbers of food items dietary nichebreadth and Shannon-Wiener measures of SiberianCranes were similar between winter 2017 and winter2018 This is inconsistent with the contention that dur-ing seasons with a low abundance of dominant food thedietary niche may broaden as consumers relying on in-sufficient preferred food items are forced to add lessprofitable resources to their diets [26 119 120] In win-ter 2017 with low Vallisneria tuber abundance SiberianCranes left shallow waters and mudflats and moved torice paddies and lotus ponds [61 62 64] Their foodschanged from Vallisneria tuber to rice seeds and lotusrhizomes almost completely As the energy protein andfat contents of crops are generally as good as or superiorto natural foods and are often present in agriculturalfields in far greater abundance [86 104] Siberian Cranesmay gain enough energy through feeding on crops anddo not need to add many other food sources Moreoverthe Siberian Crane is a dietary specialist [89 90] PoyangLake has a relatively simple plant community compos-ition with Vallisneria and Hydrilla verticillate being thedominant submerged macrophytes [59] and Carex beingthe dominant grassland plants [121] Therefore theremay be few options for Siberian Cranes which mightlead to the failure of this species to widen its dietaryniche breadthIn contrast to Siberian Cranes the numbers of food
items dietary niche breadths and Shannon-Wiener mea-sures of White-naped Cranes and Hooded Cranes were
higher in winter 2017 than in winter 2018 P criopolita-num was the predominant food item in the feces of thetwo crane species in winter 2018 and the proportions ofVallisneria tubers were also high The abundances ofVallisneria tubers and probably P criopolitanum at Po-yang Lake declined greatly in winter 2017 When thereis a short-fall in food availability animals may optimizeforaging behavior and increase their dietary breath [122123] The short-fall in the abundances of P criopolita-num and Vallisneria tubers in winter 2017 might havedriven the two crane species to add less profitable grass-land plants in their diets Although the two cranes alsoconsumed high-quality rice seeds and lotus rhizomes inwinter 2017 their proportions were low in the feces andthese foods might be unable to meet the energy require-ments of the two crane species The niche expansion ofthe two crane species during periods of preferred foodshortage was in agreement with the basic prediction ofoptimal foraging theory [22 23] It is also consistentwith a commonly observed fact that energy-maximizingmodels work better when animals food-limited [124]The dietary niche overlaps among the crane species
were low in winter 2017 with the four crane species di-verging in their principal food items However in winter2018 the niche overlaps were significantly higher thanexpected by chance They had similar diet compositionswith P criopolitanum rhizomes and Vallisneria tubersbeing the principal food items in the feces The in-creased overlap might have been caused by the highabundances of the two plant species in winter 2018which allowed the four crane species to share the samefood items with low interspecific competition Our studyis consistent with previous research that suggested thatamong-species dietary niches tended to overlap moreduring seasons of high food abundance [32 33 125126] Greater resource availability could promote an in-crease in species utilization of similar resources and re-duce competitive interactions between species [32 34126] By contrast during seasons with low food abun-dance species may partition limited food resources todecrease interspecific competition leading to a decreasein niche overlap [126 127] It has been suggested thatdiet partitioning facilitates the coexistence of manyclosely related species [7 128] The interannual variationin niche overlap observed in our study suggests that dietpartitioning may be restricted to specific temporal casesIn seasons or years with abundant food sources inter-specific competition may be relaxed and niche overlapcould be high
ConclusionsIn summary by using metabarcoding technology ourstudy gained new insights into cranesrsquo diets that are es-sential for informing habitat management to improve
Hou et al Frontiers in Zoology (2021) 1824 Page 10 of 17
the availability of foraging resources Vallisneria tuberswas confirmed as an important food source of cranesSurprisingly P criopolitanum also occupied high pro-portions in the feces of cranes and may also be import-ant Our study emphasizes the superior quality ofnatural wetlands compared to agricultural fields as for-aging habitats for cranes The recent degradation of sub-merged macrophytes at Poyang Lake potentiallythreatens the survival of cranes especially SiberianCranes Therefore it is urgent to reverse the broad de-cline of submerged macrophytes and provide importantforaging habitats for cranes during years of tuber col-lapses The abundances of Vallisneria tubers and prob-ably P criopolitanum at Poyang Lake greatly influencedthe dietary niche width and overlaps among crane spe-cies Cranes exhibited low dietary niche overlap duringperiods of preferred food shortage but significantly highoverlap during periods when preferred foods were plen-tiful While high niche overlap is often interpreted as in-tense interspecific competition our study highlights theimportance of taking food abundance into account
Materials and methodsStudy areasPoyang Lake the largest freshwater lake in China is oneof the most important wintering areas for waterbirdsalong the East Asian-Australasian Flyway (Fig 1) [51129] It is characterized by dramatic seasonal hydro-logical fluctuations with an average change of 924 mbetween the summer high and winter low water levels[130] During summer the inundation area is gt 2500km2 [131] In autumn and winter the water level dropscausing the inundation area to shrink to lt 1000 km2
[131] Poyang Lake contains 102 sublakes which are in-undated and integrated with the main body of PoyangLake during summer and are isolated during winter[132] As the water level drops the sublakes appear andlarge areas of grasslands mudflats and shallow waterareas are exposed providing winter foraging and roost-ing areas for approximately 420000 waterbirds from ap-proximately 111 species [133 134]
Vallisneria tuber surveyThe density and biomass of Vallisneria tubers were sur-veyed at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake in late October and earlyNovember of 2017 and 2018 before large numbers ofwintering waterbirds arrived We set 76 183 and 36sampling plots uniformly located at Dahuchi Shahu andChanghuchi respectively Vallisneria tubers were col-lected from two quadrats in each plot with a locallymade steel grab sampler The sampler has two long
handles At the end of the handles there are two scoopsfacing each other The sampler was inserted into thesubstrate and the scoops brought together to collectabout 15 cm long times 13 cm wide times 30 cm high sample ofsubstrate Tubers were subsequently cleaned andcounted The dry weight of tubers in each quadrat wasweighed after oven-drying at 80 degC until constant weightThe tuber density and biomass in each plot was calcu-lated by dividing total tuber number and tuber massacross two quadrats by quadrat area respectively Weused t-test to explore whether the tuber density and bio-mass in the winters of 2017 and 2018 significantly devi-ated from the historical average values in the wintersfrom 1999 to 2016 [57] The historical tuber density andbiomass data were collected by the Poyang Lake Na-tional Nature Reserve at three sublakes (Dahuchi Shahuand Meixihu) [57] R 360 was used to do the t-testanalyses
Fecal sample collectionFecal samples were collected at Poyang Lake in the win-ters of 2017 and 2018 We first investigated the distribu-tions of cranes When we saw foraging cranes we waiteduntil they left to avoid disturbing them Soon after theyleft we went to the foraging sites to collect fresh feces(ie feces with wet surface) with sterilized tweezers Tominimize the probability of multiple samples from thesame individual all collected samples were separated byat least 5 m We stored the samples in liquid nitrogen inthe field and transferred them to a minus 80 degC refrigeratorfor long-term storage in the laboratory To ensure thatthe sources of the feces were known we tried to collectsamples from monospecific flocks For samples collectedfrom mixed-species flocks we used DNA analysis toidentify the species that deposited the fecal samplesPlants were also collected in the field to construct aDNA barcoding reference database
Species identificationBecause of the similar morphologies of the feces of thefour crane species we used DNA analysis to identify thespecies that deposited the fecal samples collected frommixed-species flocks For the genetic species assignmentof each fecal sample we scraped the external surfacewhich contain DNA from the host species as a result ofthe sloughing of cells from the digestive tract DNA wasextracted with the QIAamp Power Fecal DNA Kit (Qia-gen USA) according to the manufacturerrsquos guidelinesFor each round of DNA extraction negative controls(ie extraction without feces) were included to monitorfor possible contamination We amplified the mitochon-drial DNA D-loop fragment with the primer pair DL02F
Hou et al Frontiers in Zoology (2021) 1824 Page 11 of 17
(5prime-3prime GCGGCCCGAAAAGCCGCTG) and DL02R (5prime-3prime GCCCTGACATAGGAACCAGAGGCGC) [135]PCR amplifications were carried out in a total volume of25 μl containing 125 μl PCR mix (Tiangen BeijingChina) 1 μl DNA 1 μl each primer and 95 μl H2O Thereaction conditions were as follows denaturation at94 degC for 4 min followed by 34 cycles at 94 degC for 48 s56 degC for 48 s and 72 degC for 1 min and a final 10 min at72 degC We checked for the presence of a PCR product ofsuitable length by electrophoresis on a 1 agarose gelSequencing was carried out on an ABI 3730xl analyzerby Sangon Biotech (Shanghai China) The complete se-quences were assembled using Seqman II (DNASTAR)and compared visually to the original chromatograms toavoid reading errors The taxonomic assignment wasconducted by searching against the nucleotide databaseof GenBank using Web BLAST
Construction of a barcode reference databaseWe downloaded the trnL sequences of 48 plant speciescommonly seen at Poyang Lake from GenBank For theplant species not included in GenBank we sequencedtrnL by ourselves Total DNA was extracted using thehexadecyltrimethylammonium bromide (CTAB) proto-col We used primer pair c (5prime-3prime CGAAATCGGTAGACGCTACG) and h (5prime-3prime CCATTGAGTCTCTGCACCTATC) to amplify an approximately 150 bp regionof the chloroplast trnL intron [136] This primer pairwas chosen as it can amplify DNA from a wide range ofwetland plants [70] PCR amplifications were carried outin a total volume of 25 μl containing 125 μl PCR mix(Tiangen Beijing China) 1 μl DNA 1 μl each primerand 95 μl H2O The reaction conditions were as followsdenaturation at 94 degC for 4 min followed by 35 cycles at94 degC for 30 s 56 degC for 30 s and 72 degC for 45 s and afinal 10 min at 72 degC We used electrophoresis to checkthe PCR products ABI 3730xl analyzer to perform se-quencing and Seqman II to assemble the sequences
DNA extraction and high-throughput sequencing of fecalsamplesTo uncover plant component consumed by cranes amodified CTAB protocol [70] was used to extract theDNA from approximately 200 mg of the interior of eachfecal sample DNA extraction was conducted in a dedi-cated laboratory for analyses of DNA from samples withlow DNA quality For each batch of DNA extractionnegative controls were included to monitor for possiblecontamination The primers c and h with a 6 bp tagadded to 5prime ends of each primer were used to amplifytrnL gene The PCR amplification procedure was thesame as barcode database construction For each samplea unique combination of tags in the forward and reverseprimers was used so that sample-specific data can be
recovered after sequencing Three PCR replicates wereperformed for each sample to reduce amplification biasThe three replicates were pooled and the DNA concen-tration and purity were monitored on a 1 agarose gelOnly samples with a clear band were processed furtherSamples were purified using the SanPrep Column PCRProduct Purification Kit (Sangon Biotech ShanghaiChina) The purified PCR products were pooled takinginto account of DNA concentration to obtain an ap-proximately equal amount of amplicon DNA from eachfecal sample The pool of individually tagged ampliconswas used to prepare a sequencing library using the Tru-Seq DNA PCR-Free Sample Preparation Kit (IlluminaUSA) following manufacturerrsquos recommendations Thelibrary was sequenced on an Illumina HiSeq 2500 plat-form and 250 bp paired-end reads were generated Li-brary preparation and sequencing were conducted byNovogene (Beijing China)
Sequencing data analysisAfter high-throughput sequencing the paired-end readswere demultiplexed into sample-specific files based ontheir unique tags The tags and primer sequences weresubsequently trimmed The paired-end reads weremerged using FLASH 127 [137] and then filtered usingQIIME 170 [138] Sequences were dereplicated andclustered to OTUs using USEARCH 110667 [139] atthe similarity threshold of 97 All OTUs were assignedto taxonomic units by referring to the local plant data-base using blast 2231+ with thresholds of identity gt97 and e-value lt 10 eminus 50 If an OTU matched two ormore taxa it was assigned to a higher taxonomic levelthat included all taxaTo evaluate the sufficiency of sequencing depth and
sampling size individual- species- and sample-basedrarefaction curves were built using EstimateS 82[140] The curves were computed by randomly resam-pling sequences or samples and plotting these againstthe numbers of OTUs The read abundance data wereconverted to RRA of each food item using the follow-ing equation [79]
RRA frac14 1S
XS
Kfrac141
nikPTifrac141nik
100
where nik is the number of sequences of food item i insample k T is the total number of food items and S isthe sampling size
Dietary niche breadth and overlap among crane speciesTo explore whether the cranesrsquo diets showed substantialinterannual variation we calculated Pearson correlationsbetween diet compositions of each crane species in win-ter 2017 and winter 2018 The proportion of each food
Hou et al Frontiers in Zoology (2021) 1824 Page 12 of 17
item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

The average number of reads per sample was 65910 (plusmn26276 SD) and the average number of operational taxo-nomic units (OTUs) per sample was 43 (plusmn 15 SD) Toevaluate the sequencing and sampling adequacy individ-ual- species- and sample-based rarefaction curves werebuilt With the sample sequencing reads increased theindividual-based rarefaction curves reached plateau inmost cases (Additional file 1 Figure S1) and the species-based rarefaction curves reached plateau in the fourcrane species (Fig 2a) Therefore our sequencing depthwas roughly adequate to capture the numbers of OTUspresent The sample-based rarefaction curves for eachcrane species reached plateau in all cases as the samplingsizes increased (Fig 2b) suggesting that we acquired suf-ficient fecal samples After comparison to the local DNAreference database 9899 of the total reads wereassigned to specific plant taxa Most of the reads thatwere not assigned to specific plant taxa had low fre-quency In total we identified 29 plant taxa including23 species 4 genera and 2 families
Diet compositions of the four crane speciesWe used relative read abundance (ie the percentageof DNA belonging to each food item in each sampleRRA) which provides a more accurate view of speciesdiet than frequency of occurrence (ie the number ofsamples that contain a given food item) tosummarize dietary data [79] Twenty-two food itemswere identified in the feces of Siberian Cranes in thetwo winters including 17 species 3 genera and 2families (Fig 3 Additional file 1 Table S2) Rice(RRA = 7285) lotus (1201) and Polygonum criopo-litanum (1199) were the dominant food items inthe feces (ie proportion gt 10) in winter 2017 andVallisneria (5612) and P criopolitanum (4087)were the dominant food items in winter 2018Twenty-five food items were identified in the feces ofWhite-naped Cranes including 20 species 3 genera
and 2 families Tulipa edulis (3737) Potentilla lim-prichtii (1869) P criopolitanum (1652) andCarex spp (1075) were the dominant food items inthe feces in winter 2017 and P criopolitanum(8441) was the dominant food item in winter 2018Twenty-two food items were identified in the feces ofHooded Cranes including 18 species 3 genera and 1family P limprichtii (6536) and Carex (2739)were the dominant food items in the feces in winter2017 and P criopolitanum (8876) was the domin-ant food item in winter 2018 Twenty-two food itemswere identified in the feces of Eurasian Cranes in-cluding 17 species 3 genera and 2 families Rice(6120) and P limprichtii (2394) were the domin-ant food items in the feces in winter 2017 and Pcriopolitanum (5577) and rice (2740) were thedominant food items in winter 2018 Diet composi-tions of Siberian Cranes (Pearson correlation = 005P = 077) White-naped Cranes (Pearson correlation =030 P = 008) and Hooded Cranes (Pearson correl-ation = minus 004 P = 083) were not correlated betweenwinter 2017 and winter 2018 suggesting great inter-annual variations in food sources Diet composition ofEurasian Cranes was correlated (Pearson correlation =040 P = 002) between the two winters suggestinglimited interannual variation in food sourcesVallisneria tubers were an important food source for
the four crane species especially Siberian Cranes inwinter 2018 The proportion of Vallisneria was 5612512 850 and 734 in the feces of Siberian CranesWhite-naped Cranes Hooded Cranes and EurasianCranes respectively in winter 2018 (Fig 3 Additionalfile 1 Table S2) In winter 2017 when Vallisneria tuberabundance was low the proportion of Vallisneria wasless than 1 in the feces of the four crane speciesDomesticated species identified in crane feces included
rice and lotus The proportions of domesticated speciesin the feces of Siberian Cranes (8485) and White-
Fig 2 a Species-based and b sample-based rarefaction curves The species- and sample-based rarefaction curves built by randomly resamplingsequences and samples respectively at increasing levels of accumulation indicate whether the sequencing depth and sampling size weresufficient to characterize the diversity of dietary items The colors of the curves correspond to the crane species blue Siberian Crane purpleWhite-naped Crane green Hooded Crane and red Eurasian Crane
Hou et al Frontiers in Zoology (2021) 1824 Page 5 of 17
naped Cranes (1509) were high in winter 2017 but de-clined to 000 in winter 2018 (Fig 3 Additional file 1Table S2) Domesticated species occupied a small pro-portion in the feces of Hooded Cranes in winter 2017(130) but declined to 000 in winter 2018 The pro-portions of domesticated species in the feces of EurasianCranes were high in both winters with 6120 in winter2017 and 2740 in winter 2018
Dietary niche breadth and overlap among crane speciesThe numbers of food items dietary niche breadth andShannon-Wiener diversity values for Siberian Cranesand Eurasian Cranes in winter 2017 were similar tothose in winter 2018 (Table 2) Values of all three pa-rameters for White-naped Cranes and Hooded Craneswere higher in winter 2017 than in winter 2018 Dietaryniche breadth of the four crane species was lt 020 inboth winters indicating low niche breadthDietary niche overlap among crane species ranged
from 002 to 049 in winter 2017 but increased to 062to 099 in winter 2018 (Table 3) Nonmetric multidimen-sional scaling (NMDS) analysis also indicated overlap
increases in winter 2018 (Fig 4) No overlap values werelower than expected by chance in the two winters Theoverlap values were not higher than expected by chancein winter 2017 However all values showed a high de-gree of overlap in winter 2018 [P (Obs gt = null) lt 005]Overlap between White-naped Cranes and HoodedCranes was the highest among the four crane species inboth winters
DiscussionDiet compositionTo the best of our knowledge our study is the first toexplore crane diets using metabarcoding techniques Al-most all previous studies have relied on traditional directobservation or microhistological methods By usingmetabarcoding technology we were able to elucidatecrane diets at greater detail than previously known Theimproved knowledge might be due to the ability ofmetabarcoding to provide a more comprehensive per-spective and more details on food sources than trad-itional methods [66 67 80] Metabarcoding is able todetect a variety of smaller and softer bodied food items
Fig 3 Food items in the feces of Siberian Cranes White-naped Cranes Hooded Cranes and Eurasian Cranes in the winters of 2017 and 2018 atPoyang Lake
Table 2 Number of food items dietary niche breadth and Shannon-Wiener diversity values for Siberian Cranes White-napedCranes Hooded Cranes and Eurasian Cranes at Poyang Lake in the winters of 2017 and 2018
Number of foods Dietary niche breadth Shannon-Wiener diversity
2017 2018 2017 2018 2017 2018
Siberian Crane 16 17 005 007 091 085
White-naped Crane 18 15 020 003 169 059
Hooded Crane 19 13 005 002 096 043
Eurasian Crane 18 19 008 009 110 121
Hou et al Frontiers in Zoology (2021) 1824 Page 6 of 17
that are overlooked by microhistological methods [6780] It also enables the identification of food items at ahigher taxonomic resolution [69 81] However we ac-knowledge that our study has two limitations First weused fecal samples as they are non-invasive Neverthe-less fecal analysis is subjected to the biases caused bythe differential digestive rates of food items [82 83] Softfoods such as insect larvae tend to be under-representedin fecal samples because they are digested quickly andremain in gastro-intestinal tract for short periods whilehard foods such as mollusks tend to be over-representedbecause they break down slowly and remain in gastro-intestinal tracts longer Thus food proportions in the
feces may not correlate with food proportions ingestedby cranes In addition some materials identified in thefeces may be incidentally consumed by cranes and notreally be used as food Second our study only consideredthe plant component of cranesrsquo diets as plants were sug-gested to be their principal food types at Poyang LakeAnimal foods consumed by the four crane species butto a much lesser extent than plant foods [84ndash87] werenot considered in our studyThe Siberian Crane is regarded as the most aquatic of
all cranes using wetlands for nesting feeding and roost-ing [49 88] The high dependence of the species onaquatic habitats was suggested to make it vulnerable to
Table 3 The observed dietary niche overlap among the four crane species at Poyang Lake in the winters of 2017 and 2018 P(Obs lt = null) and P (Obs gt = null) indicate the tail probabilities for the observed index which were lower and higher than thehistogram of the simulated index respectively represents P lt 005 and represents P lt 001
Winter Observed niche overlap index P (Obs lt = null) P (Obs gt = null)
2017 Siberian vs White-naped 021 0819 0181
Siberian vs Hooded 002 0703 0297
Siberian vs Eurasian 016 0900 0100
White-naped vs Hooded 049 0952 0048
White-naped vs Eurasian 035 0895 0106
Hooded vs Eurasian 035 0919 0081
2018 Siberian vs White-naped 063 0952 0048
Siberian vs Hooded 066 0952 0048
Siberian vs Eurasian 062 0950 0049
White-naped vs Hooded 099 0997 0003
White-naped vs Eurasian 089 0987 0013
Hooded vs Eurasian 090 0998 0002
Fig 4 The results of nonmetric multidimensional scaling ordination (NMDS) based on Bray-Curtis dissimilarity The colors of the circlescorrespond to the crane species blue Siberian Crane purple White-naped Crane green Hooded Crane and red Eurasian Crane
Hou et al Frontiers in Zoology (2021) 1824 Page 7 of 17
impacts of habitat degradation and to lead its classifica-tion as Critically Endangered [49] At Poyang Lake Si-berian Cranes traditionally fed on tubers of Vallisneriathe dominant submerged macrophytes in shallow watersand mudflats [89 90] In winter 2010 due to a flood-induced Vallisneria tuber collapse they were observedforaging for P limprichtii taproots and T edulis bulbs ingrasslands for the first time [38 44] In the winters from2015 to 2017 the diets of Siberian Cranes changedagain with thousands of cranes feeding on rice seedsand lotus rhizomes in agricultural habitats [61 62 64]Our study identified 22 plant items in the feces of Siber-
ian Cranes These plant items were obtained from shallowwaters grasslands and agricultural fields Four plant itemswith yearly RRA gt 10 were identified in the feces Vallis-neria tuber P criopolitanum rhizomes rice seeds andlotus rhizomes (the vegetative portions consumed bycranes were based on our field observation) SiberianCranesrsquo diet compositions showed substantial interannualvariation When the abundance of Vallisneria tubers waslow (ie winter 2017) Siberian Cranes mainly fed on riceseeds lotus rhizomes and P criopolitanum rhizomes inagricultural fields and grasslands When the abundance ofVallisneria tubers was high (ie winter 2018) they mainlyfed on Vallisneria tubers and P criopolitanum rhizomesin shallow waters and grasslands Our study in combin-ation with previous studies indicates that the diets of Si-berian Cranes have changed greatly since winter 2010 [3844 62] They have gradually broadened their dietaryniches which has probably been driven by the degradationof submerged plants [38 44 57]Similar to Siberian Cranes a diet shift was also ob-
served in White-naped Cranes Historically White-naped Cranes only fed on submerged plants in shallowwaters and mudflats of Poyang Lake [91] Then drivenby the decline of submerged plants they primarily fed atgrasslands and mudflats [87 91] consuming Vallisneriatuber T edulis P limprichtii and Ranunculus polii [5587] Our results indicate that in addition to T edulisand P limprichtii that had previously been reported theproportions of P criopolitanum and Carex were alsohigh in the feces Hooded Cranes were suggested tomainly feed on the grassland plant P limprichtii throughdirect observation [86] Our study indicates that thegrassland plants P criopolitanum and Carex also occu-pied high proportions in the feces The dominant fooditems in the feces of White-naped Cranes and HoodedCranes varied greatly between the two winters The vari-ation in dietary composition indicated that the diversityand composition of the grassland plant communitymight have changed greatly between the two wintersThe dietary niche overlap between White-naped Cranesand Hooded Cranes was the highest among the fourcrane species aligning with their level of habitat niche
overlap which was also the highest among the species[92] The high dietary and habitat niche overlaps suggesthigh competition potential between the two cranespeciesThe Eurasian Crane is a habitat generalist It is known
to feed in a variety of habitats including grasslandsmudflats shallow waters and agricultural fields [88 92]Its diet at Poyang Lake has not been documented previ-ously Our study indicates that P criopolitanum P lim-prichtii and rice were the dominant food items in thefeces of Eurasian Cranes The number of plant itemsconsumed dietary niche breadth and Shannon-Wienermeasures were the highest among the four crane speciessupporting its generalist foraging strategy In contrast tothe three crane species above Eurasian Cranesrsquo dietcompositions were similar between the two winters Riceand grassland plants occupied high proportions in thefeces in both winters
Important food resources for cranesVallisneria tubers have been acknowledged as importantfood sources for cranes especially Siberian Cranes andWhite-naped Cranes [55 56] Our study confirms theimportance of Vallisneria tubers in the diet of SiberianCranes It was the most common food source in thefeces of Siberian Cranes in winter 2018 when its abun-dance was high Vallisneria tuber proportions in thefeces of the three other crane species were not as high asthat in the feces of Siberian Cranes but it still occupied5ndash9 of the total foods in winter 2018To our surprise P criopolitanum was found to be the
first dominant food in the feces of White-naped CranesHooded Cranes and Eurasian Cranes and the seconddominant food in the feces of Siberian Cranes in winter2018 In winter 2017 the proportions of P criopolita-num were lt 17 in the feces of Siberian Cranes andWhite-naped Cranes and were lt 3 in the feces ofHooded Cranes and Eurasian Cranes The higher pro-portions of P criopolitanum in winter 2018 than winter2017 indicate that the abundance and availability of Pcriopolitanum was much higher in winter 2018 P crio-politanum is an annual grassland plant It is sensitive tohydrological conditions and can only survive in a rela-tively narrow range of water depths and percent time in-undated [93] Due to the sensitivity its distributionrange and abundance showed high interannual variation[94 95] Thus it is likely that the abundance of P crio-politanum was higher in winter 2018 than in winter2017The importance of P criopolitanum as a food source
for the cranes at Poyang Lake has not been documentedpreviously It has only been suggested to be a major foodsource of Hooded Cranes at Shengjin Lake another lakein the middle and lower Yangtze River floodplain [86
Hou et al Frontiers in Zoology (2021) 1824 Page 8 of 17
96] P criopolitanum is a dominant wetland plant at Po-yang Lake [93 97] The coverage percentage of this plantis the third highest among grassland plants accountingfor 12 of the total areas of grasslands [97] This plantspecies is usually distributed at elevations of 9ndash12mabove sea level near mudflats and shallow waters [97]which are frequently used by cranes Its rhizome with ashallow burial depth is high in energy and nutritionalcontent and low in crude fibre [38 86] All these charac-teristics make P criopolitanum rhizome a potentially im-portant food source for cranes We frequently observedcranes dig and feed on rhizomes of P criopolitanum andleave aboveground parts in the fields Therefore we be-lieve that P criopolitanum was used as food not inci-dentally consumed by cranes However we need to becaution that high proportion of P criopolitanum in thefeces does not mean high proportion in the diets Morework on the importance of P criopolitanum in the dietsof cranes is needed
The roles of agricultural fields in crane protectionWith the loss and degradation of natural wetlands theroles of artificial wetlands in waterbird protection havebeen under intensive debate [98 99] Although agricul-tural fields cannot completely replace the function ofnatural wetlands as waterbird habitats [100ndash102] agri-cultural fields might provide alternative or complemen-tary habitats for waterbirds in all life stages [38 103104] The proportions of domesticated species in thefeces of Siberian Cranes varied considerably betweenyears In winter 2017 when the abundance of Vallisneriatubers was low the proportion of Vallisneria in SiberianCranesrsquo feces was only 095 Siberian Cranes may nothave found enough foods in the shallow waters andmudflats and were forced to search for rice seeds andlotus rhizomes in agricultural fields In winter 2018when the abundance of Vallisneria tubers reboundedSiberian Cranes moved back to shallow waters and mud-flats and a large proportion of their traditional food Val-lisneria tubers and also P criopolitanum were detectedin the feces A similar abandonment of tuber foragingoccurred following the tuber collapse in 2010 and thenresumed in the next winter when tubers rebounded in2011 [38 44] The diet and habitat changes indicate thatagricultural habitats were important refuges for SiberianCranes providing important food resources for them tosurvive through winters with few Vallisneria tubersThey also indicate that Siberian Cranes prefer naturalwetlands to agricultural fields [38]Similar to Siberian Cranes White-naped Cranes and
Hooded Cranes consumed rice seeds and lotus rhizomesin winter 2017 but did not consume any in winter 2018The changes in the proportions suggest that the abun-dance of food resources in natural wetlands might have
been low in winter 2017 The deficient food resourcesmight have been Vallisneria tubers but might also havebeen grassland plants the primary food sources of thesetwo crane species Under natural food shortage condi-tions (ie winter 2017) White-naped Cranes andHooded Cranes utilized agricultural fields When theabundance of Vallisneria tubers and probably grasslandplants were high (ie winter 2018) the two crane speciesreturned to their natural habitats Therefore for White-naped Cranes and Hooded Cranes agricultural fieldswere important alternative foraging habitats used asbuffers against starvation during periods of natural foodshortages The return of the two crane species to naturalwetlands in winter 2018 further emphasizes the superiorquality of natural habitats over agricultural fields as for-aging habitats for cranes [38]The proportions of rice seeds in the feces of Eur-
asian Cranes were high in the two winters thoughthe proportion in winter 2017 was twice as high asthat in winter 2018 Therefore agricultural fields ap-peared to be consistently important as foraging habi-tats for Eurasian Cranes The frequent use ofagricultural fields by Eurasian Cranes has been docu-mented widely [49 105 106]Domesticated species generally contain high energy
and nutritional content and are of superior quality com-pared to natural plants [86 104] Farmland foraging hasbeen suggested to contribute to increase in the abun-dance of some goose [107] egret [108] and wader [109]populations in Japan Korea Europe and North Amer-ica However cranes generally avoid human disturbancewhen select foraging sites [110 111] Human disturb-ance in agricultural fields around Poyang Lake is consid-erably high due to intensive agricultural activity [112]High human disturbance was suggested to be a majorreason that wintering geese were almost confined withinnatural wetlands and hesitated to exploit the riches ofthe modern agricultural habitats at Poyang Lake [112113] High human disturbance also resulted in SiberianCranes spending twice as much time alerting in agricul-tural fields than they did in natural habitats [114] Thehigh level of human disturbance in agricultural fieldsmight contribute to the preference for natural wetlandsby Siberian Cranes White-naped Cranes and HoodedCranes at Poyang Lake Moreover although agriculturefields contain energy-rich crops wetlands contain di-verse food resources especially protein-rich inverte-brates which might better satisfy dietary needs of cranes[110 115] Taken together security and diverse food re-sources might have attracted cranes to select wetlands asforaging sitesSubmerged macrophytes at Poyang Lake have de-
graded in recent decades [57ndash60] and Vallisneria tuberdisappearance is expected to occur more frequently
Hou et al Frontiers in Zoology (2021) 1824 Page 9 of 17
under future climate change [57 60] Cranes may in-creasingly depend upon agricultural fields around Po-yang Lake which may in turn cause significant cropdamage and economic losses This is especially true onagricultural fields surrounding protected wetlands wherewaterbirds concentrate in high numbers [116 117] AtPoyang Lake cranes mainly fed on post-harvest remainsof rice seeds and lotus rhizomes thus crop damage andeconomic loss were limited However we observed somefarmers scared cranes away from their agricultural fieldsThere were more than 14 million free-ranging poultryraised at agricultural fields around Poyang Lake [118]where they gleaned waste grain residues in stubble fieldsCrane feeding would reduce the food abundance of thefree-ranging domestic poultry thus farmers drovecranes away To reduce human disturbance environ-mental education is needed because farmers are morelikely to protect cranes if they understand the endan-gered degree and the ecological significance of cranes
Effects of tuber collapse on dietary niche breadth andoverlap among crane speciesVallisneria tuber is an important food source for Siber-ian Cranes (89 90 this study) thus variation in itsabundance might influence Siberian Cranesrsquo dietaryniche breadth The numbers of food items dietary nichebreadth and Shannon-Wiener measures of SiberianCranes were similar between winter 2017 and winter2018 This is inconsistent with the contention that dur-ing seasons with a low abundance of dominant food thedietary niche may broaden as consumers relying on in-sufficient preferred food items are forced to add lessprofitable resources to their diets [26 119 120] In win-ter 2017 with low Vallisneria tuber abundance SiberianCranes left shallow waters and mudflats and moved torice paddies and lotus ponds [61 62 64] Their foodschanged from Vallisneria tuber to rice seeds and lotusrhizomes almost completely As the energy protein andfat contents of crops are generally as good as or superiorto natural foods and are often present in agriculturalfields in far greater abundance [86 104] Siberian Cranesmay gain enough energy through feeding on crops anddo not need to add many other food sources Moreoverthe Siberian Crane is a dietary specialist [89 90] PoyangLake has a relatively simple plant community compos-ition with Vallisneria and Hydrilla verticillate being thedominant submerged macrophytes [59] and Carex beingthe dominant grassland plants [121] Therefore theremay be few options for Siberian Cranes which mightlead to the failure of this species to widen its dietaryniche breadthIn contrast to Siberian Cranes the numbers of food
items dietary niche breadths and Shannon-Wiener mea-sures of White-naped Cranes and Hooded Cranes were
higher in winter 2017 than in winter 2018 P criopolita-num was the predominant food item in the feces of thetwo crane species in winter 2018 and the proportions ofVallisneria tubers were also high The abundances ofVallisneria tubers and probably P criopolitanum at Po-yang Lake declined greatly in winter 2017 When thereis a short-fall in food availability animals may optimizeforaging behavior and increase their dietary breath [122123] The short-fall in the abundances of P criopolita-num and Vallisneria tubers in winter 2017 might havedriven the two crane species to add less profitable grass-land plants in their diets Although the two cranes alsoconsumed high-quality rice seeds and lotus rhizomes inwinter 2017 their proportions were low in the feces andthese foods might be unable to meet the energy require-ments of the two crane species The niche expansion ofthe two crane species during periods of preferred foodshortage was in agreement with the basic prediction ofoptimal foraging theory [22 23] It is also consistentwith a commonly observed fact that energy-maximizingmodels work better when animals food-limited [124]The dietary niche overlaps among the crane species
were low in winter 2017 with the four crane species di-verging in their principal food items However in winter2018 the niche overlaps were significantly higher thanexpected by chance They had similar diet compositionswith P criopolitanum rhizomes and Vallisneria tubersbeing the principal food items in the feces The in-creased overlap might have been caused by the highabundances of the two plant species in winter 2018which allowed the four crane species to share the samefood items with low interspecific competition Our studyis consistent with previous research that suggested thatamong-species dietary niches tended to overlap moreduring seasons of high food abundance [32 33 125126] Greater resource availability could promote an in-crease in species utilization of similar resources and re-duce competitive interactions between species [32 34126] By contrast during seasons with low food abun-dance species may partition limited food resources todecrease interspecific competition leading to a decreasein niche overlap [126 127] It has been suggested thatdiet partitioning facilitates the coexistence of manyclosely related species [7 128] The interannual variationin niche overlap observed in our study suggests that dietpartitioning may be restricted to specific temporal casesIn seasons or years with abundant food sources inter-specific competition may be relaxed and niche overlapcould be high
ConclusionsIn summary by using metabarcoding technology ourstudy gained new insights into cranesrsquo diets that are es-sential for informing habitat management to improve
Hou et al Frontiers in Zoology (2021) 1824 Page 10 of 17
the availability of foraging resources Vallisneria tuberswas confirmed as an important food source of cranesSurprisingly P criopolitanum also occupied high pro-portions in the feces of cranes and may also be import-ant Our study emphasizes the superior quality ofnatural wetlands compared to agricultural fields as for-aging habitats for cranes The recent degradation of sub-merged macrophytes at Poyang Lake potentiallythreatens the survival of cranes especially SiberianCranes Therefore it is urgent to reverse the broad de-cline of submerged macrophytes and provide importantforaging habitats for cranes during years of tuber col-lapses The abundances of Vallisneria tubers and prob-ably P criopolitanum at Poyang Lake greatly influencedthe dietary niche width and overlaps among crane spe-cies Cranes exhibited low dietary niche overlap duringperiods of preferred food shortage but significantly highoverlap during periods when preferred foods were plen-tiful While high niche overlap is often interpreted as in-tense interspecific competition our study highlights theimportance of taking food abundance into account
Materials and methodsStudy areasPoyang Lake the largest freshwater lake in China is oneof the most important wintering areas for waterbirdsalong the East Asian-Australasian Flyway (Fig 1) [51129] It is characterized by dramatic seasonal hydro-logical fluctuations with an average change of 924 mbetween the summer high and winter low water levels[130] During summer the inundation area is gt 2500km2 [131] In autumn and winter the water level dropscausing the inundation area to shrink to lt 1000 km2
[131] Poyang Lake contains 102 sublakes which are in-undated and integrated with the main body of PoyangLake during summer and are isolated during winter[132] As the water level drops the sublakes appear andlarge areas of grasslands mudflats and shallow waterareas are exposed providing winter foraging and roost-ing areas for approximately 420000 waterbirds from ap-proximately 111 species [133 134]
Vallisneria tuber surveyThe density and biomass of Vallisneria tubers were sur-veyed at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake in late October and earlyNovember of 2017 and 2018 before large numbers ofwintering waterbirds arrived We set 76 183 and 36sampling plots uniformly located at Dahuchi Shahu andChanghuchi respectively Vallisneria tubers were col-lected from two quadrats in each plot with a locallymade steel grab sampler The sampler has two long
handles At the end of the handles there are two scoopsfacing each other The sampler was inserted into thesubstrate and the scoops brought together to collectabout 15 cm long times 13 cm wide times 30 cm high sample ofsubstrate Tubers were subsequently cleaned andcounted The dry weight of tubers in each quadrat wasweighed after oven-drying at 80 degC until constant weightThe tuber density and biomass in each plot was calcu-lated by dividing total tuber number and tuber massacross two quadrats by quadrat area respectively Weused t-test to explore whether the tuber density and bio-mass in the winters of 2017 and 2018 significantly devi-ated from the historical average values in the wintersfrom 1999 to 2016 [57] The historical tuber density andbiomass data were collected by the Poyang Lake Na-tional Nature Reserve at three sublakes (Dahuchi Shahuand Meixihu) [57] R 360 was used to do the t-testanalyses
Fecal sample collectionFecal samples were collected at Poyang Lake in the win-ters of 2017 and 2018 We first investigated the distribu-tions of cranes When we saw foraging cranes we waiteduntil they left to avoid disturbing them Soon after theyleft we went to the foraging sites to collect fresh feces(ie feces with wet surface) with sterilized tweezers Tominimize the probability of multiple samples from thesame individual all collected samples were separated byat least 5 m We stored the samples in liquid nitrogen inthe field and transferred them to a minus 80 degC refrigeratorfor long-term storage in the laboratory To ensure thatthe sources of the feces were known we tried to collectsamples from monospecific flocks For samples collectedfrom mixed-species flocks we used DNA analysis toidentify the species that deposited the fecal samplesPlants were also collected in the field to construct aDNA barcoding reference database
Species identificationBecause of the similar morphologies of the feces of thefour crane species we used DNA analysis to identify thespecies that deposited the fecal samples collected frommixed-species flocks For the genetic species assignmentof each fecal sample we scraped the external surfacewhich contain DNA from the host species as a result ofthe sloughing of cells from the digestive tract DNA wasextracted with the QIAamp Power Fecal DNA Kit (Qia-gen USA) according to the manufacturerrsquos guidelinesFor each round of DNA extraction negative controls(ie extraction without feces) were included to monitorfor possible contamination We amplified the mitochon-drial DNA D-loop fragment with the primer pair DL02F
Hou et al Frontiers in Zoology (2021) 1824 Page 11 of 17
(5prime-3prime GCGGCCCGAAAAGCCGCTG) and DL02R (5prime-3prime GCCCTGACATAGGAACCAGAGGCGC) [135]PCR amplifications were carried out in a total volume of25 μl containing 125 μl PCR mix (Tiangen BeijingChina) 1 μl DNA 1 μl each primer and 95 μl H2O Thereaction conditions were as follows denaturation at94 degC for 4 min followed by 34 cycles at 94 degC for 48 s56 degC for 48 s and 72 degC for 1 min and a final 10 min at72 degC We checked for the presence of a PCR product ofsuitable length by electrophoresis on a 1 agarose gelSequencing was carried out on an ABI 3730xl analyzerby Sangon Biotech (Shanghai China) The complete se-quences were assembled using Seqman II (DNASTAR)and compared visually to the original chromatograms toavoid reading errors The taxonomic assignment wasconducted by searching against the nucleotide databaseof GenBank using Web BLAST
Construction of a barcode reference databaseWe downloaded the trnL sequences of 48 plant speciescommonly seen at Poyang Lake from GenBank For theplant species not included in GenBank we sequencedtrnL by ourselves Total DNA was extracted using thehexadecyltrimethylammonium bromide (CTAB) proto-col We used primer pair c (5prime-3prime CGAAATCGGTAGACGCTACG) and h (5prime-3prime CCATTGAGTCTCTGCACCTATC) to amplify an approximately 150 bp regionof the chloroplast trnL intron [136] This primer pairwas chosen as it can amplify DNA from a wide range ofwetland plants [70] PCR amplifications were carried outin a total volume of 25 μl containing 125 μl PCR mix(Tiangen Beijing China) 1 μl DNA 1 μl each primerand 95 μl H2O The reaction conditions were as followsdenaturation at 94 degC for 4 min followed by 35 cycles at94 degC for 30 s 56 degC for 30 s and 72 degC for 45 s and afinal 10 min at 72 degC We used electrophoresis to checkthe PCR products ABI 3730xl analyzer to perform se-quencing and Seqman II to assemble the sequences
DNA extraction and high-throughput sequencing of fecalsamplesTo uncover plant component consumed by cranes amodified CTAB protocol [70] was used to extract theDNA from approximately 200 mg of the interior of eachfecal sample DNA extraction was conducted in a dedi-cated laboratory for analyses of DNA from samples withlow DNA quality For each batch of DNA extractionnegative controls were included to monitor for possiblecontamination The primers c and h with a 6 bp tagadded to 5prime ends of each primer were used to amplifytrnL gene The PCR amplification procedure was thesame as barcode database construction For each samplea unique combination of tags in the forward and reverseprimers was used so that sample-specific data can be
recovered after sequencing Three PCR replicates wereperformed for each sample to reduce amplification biasThe three replicates were pooled and the DNA concen-tration and purity were monitored on a 1 agarose gelOnly samples with a clear band were processed furtherSamples were purified using the SanPrep Column PCRProduct Purification Kit (Sangon Biotech ShanghaiChina) The purified PCR products were pooled takinginto account of DNA concentration to obtain an ap-proximately equal amount of amplicon DNA from eachfecal sample The pool of individually tagged ampliconswas used to prepare a sequencing library using the Tru-Seq DNA PCR-Free Sample Preparation Kit (IlluminaUSA) following manufacturerrsquos recommendations Thelibrary was sequenced on an Illumina HiSeq 2500 plat-form and 250 bp paired-end reads were generated Li-brary preparation and sequencing were conducted byNovogene (Beijing China)
Sequencing data analysisAfter high-throughput sequencing the paired-end readswere demultiplexed into sample-specific files based ontheir unique tags The tags and primer sequences weresubsequently trimmed The paired-end reads weremerged using FLASH 127 [137] and then filtered usingQIIME 170 [138] Sequences were dereplicated andclustered to OTUs using USEARCH 110667 [139] atthe similarity threshold of 97 All OTUs were assignedto taxonomic units by referring to the local plant data-base using blast 2231+ with thresholds of identity gt97 and e-value lt 10 eminus 50 If an OTU matched two ormore taxa it was assigned to a higher taxonomic levelthat included all taxaTo evaluate the sufficiency of sequencing depth and
sampling size individual- species- and sample-basedrarefaction curves were built using EstimateS 82[140] The curves were computed by randomly resam-pling sequences or samples and plotting these againstthe numbers of OTUs The read abundance data wereconverted to RRA of each food item using the follow-ing equation [79]
RRA frac14 1S
XS
Kfrac141
nikPTifrac141nik
100
where nik is the number of sequences of food item i insample k T is the total number of food items and S isthe sampling size
Dietary niche breadth and overlap among crane speciesTo explore whether the cranesrsquo diets showed substantialinterannual variation we calculated Pearson correlationsbetween diet compositions of each crane species in win-ter 2017 and winter 2018 The proportion of each food
Hou et al Frontiers in Zoology (2021) 1824 Page 12 of 17
item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-
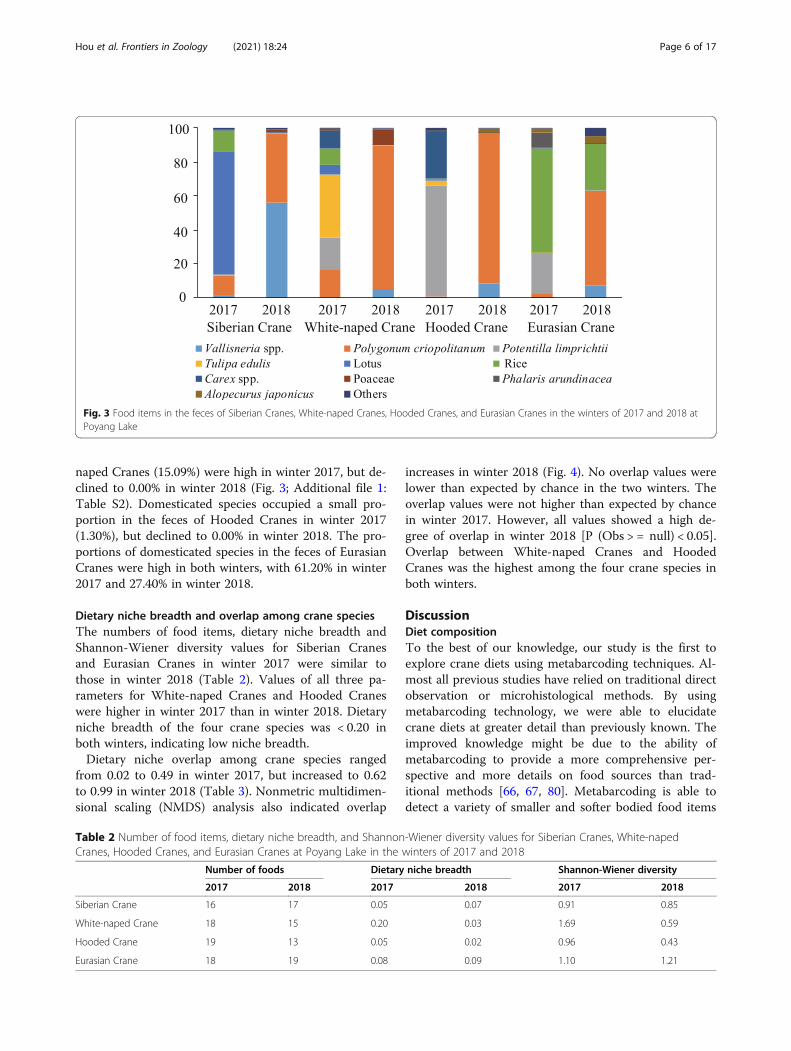
naped Cranes (1509) were high in winter 2017 but de-clined to 000 in winter 2018 (Fig 3 Additional file 1Table S2) Domesticated species occupied a small pro-portion in the feces of Hooded Cranes in winter 2017(130) but declined to 000 in winter 2018 The pro-portions of domesticated species in the feces of EurasianCranes were high in both winters with 6120 in winter2017 and 2740 in winter 2018
Dietary niche breadth and overlap among crane speciesThe numbers of food items dietary niche breadth andShannon-Wiener diversity values for Siberian Cranesand Eurasian Cranes in winter 2017 were similar tothose in winter 2018 (Table 2) Values of all three pa-rameters for White-naped Cranes and Hooded Craneswere higher in winter 2017 than in winter 2018 Dietaryniche breadth of the four crane species was lt 020 inboth winters indicating low niche breadthDietary niche overlap among crane species ranged
from 002 to 049 in winter 2017 but increased to 062to 099 in winter 2018 (Table 3) Nonmetric multidimen-sional scaling (NMDS) analysis also indicated overlap
increases in winter 2018 (Fig 4) No overlap values werelower than expected by chance in the two winters Theoverlap values were not higher than expected by chancein winter 2017 However all values showed a high de-gree of overlap in winter 2018 [P (Obs gt = null) lt 005]Overlap between White-naped Cranes and HoodedCranes was the highest among the four crane species inboth winters
DiscussionDiet compositionTo the best of our knowledge our study is the first toexplore crane diets using metabarcoding techniques Al-most all previous studies have relied on traditional directobservation or microhistological methods By usingmetabarcoding technology we were able to elucidatecrane diets at greater detail than previously known Theimproved knowledge might be due to the ability ofmetabarcoding to provide a more comprehensive per-spective and more details on food sources than trad-itional methods [66 67 80] Metabarcoding is able todetect a variety of smaller and softer bodied food items
Fig 3 Food items in the feces of Siberian Cranes White-naped Cranes Hooded Cranes and Eurasian Cranes in the winters of 2017 and 2018 atPoyang Lake
Table 2 Number of food items dietary niche breadth and Shannon-Wiener diversity values for Siberian Cranes White-napedCranes Hooded Cranes and Eurasian Cranes at Poyang Lake in the winters of 2017 and 2018
Number of foods Dietary niche breadth Shannon-Wiener diversity
2017 2018 2017 2018 2017 2018
Siberian Crane 16 17 005 007 091 085
White-naped Crane 18 15 020 003 169 059
Hooded Crane 19 13 005 002 096 043
Eurasian Crane 18 19 008 009 110 121
Hou et al Frontiers in Zoology (2021) 1824 Page 6 of 17
that are overlooked by microhistological methods [6780] It also enables the identification of food items at ahigher taxonomic resolution [69 81] However we ac-knowledge that our study has two limitations First weused fecal samples as they are non-invasive Neverthe-less fecal analysis is subjected to the biases caused bythe differential digestive rates of food items [82 83] Softfoods such as insect larvae tend to be under-representedin fecal samples because they are digested quickly andremain in gastro-intestinal tract for short periods whilehard foods such as mollusks tend to be over-representedbecause they break down slowly and remain in gastro-intestinal tracts longer Thus food proportions in the
feces may not correlate with food proportions ingestedby cranes In addition some materials identified in thefeces may be incidentally consumed by cranes and notreally be used as food Second our study only consideredthe plant component of cranesrsquo diets as plants were sug-gested to be their principal food types at Poyang LakeAnimal foods consumed by the four crane species butto a much lesser extent than plant foods [84ndash87] werenot considered in our studyThe Siberian Crane is regarded as the most aquatic of
all cranes using wetlands for nesting feeding and roost-ing [49 88] The high dependence of the species onaquatic habitats was suggested to make it vulnerable to
Table 3 The observed dietary niche overlap among the four crane species at Poyang Lake in the winters of 2017 and 2018 P(Obs lt = null) and P (Obs gt = null) indicate the tail probabilities for the observed index which were lower and higher than thehistogram of the simulated index respectively represents P lt 005 and represents P lt 001
Winter Observed niche overlap index P (Obs lt = null) P (Obs gt = null)
2017 Siberian vs White-naped 021 0819 0181
Siberian vs Hooded 002 0703 0297
Siberian vs Eurasian 016 0900 0100
White-naped vs Hooded 049 0952 0048
White-naped vs Eurasian 035 0895 0106
Hooded vs Eurasian 035 0919 0081
2018 Siberian vs White-naped 063 0952 0048
Siberian vs Hooded 066 0952 0048
Siberian vs Eurasian 062 0950 0049
White-naped vs Hooded 099 0997 0003
White-naped vs Eurasian 089 0987 0013
Hooded vs Eurasian 090 0998 0002
Fig 4 The results of nonmetric multidimensional scaling ordination (NMDS) based on Bray-Curtis dissimilarity The colors of the circlescorrespond to the crane species blue Siberian Crane purple White-naped Crane green Hooded Crane and red Eurasian Crane
Hou et al Frontiers in Zoology (2021) 1824 Page 7 of 17
impacts of habitat degradation and to lead its classifica-tion as Critically Endangered [49] At Poyang Lake Si-berian Cranes traditionally fed on tubers of Vallisneriathe dominant submerged macrophytes in shallow watersand mudflats [89 90] In winter 2010 due to a flood-induced Vallisneria tuber collapse they were observedforaging for P limprichtii taproots and T edulis bulbs ingrasslands for the first time [38 44] In the winters from2015 to 2017 the diets of Siberian Cranes changedagain with thousands of cranes feeding on rice seedsand lotus rhizomes in agricultural habitats [61 62 64]Our study identified 22 plant items in the feces of Siber-
ian Cranes These plant items were obtained from shallowwaters grasslands and agricultural fields Four plant itemswith yearly RRA gt 10 were identified in the feces Vallis-neria tuber P criopolitanum rhizomes rice seeds andlotus rhizomes (the vegetative portions consumed bycranes were based on our field observation) SiberianCranesrsquo diet compositions showed substantial interannualvariation When the abundance of Vallisneria tubers waslow (ie winter 2017) Siberian Cranes mainly fed on riceseeds lotus rhizomes and P criopolitanum rhizomes inagricultural fields and grasslands When the abundance ofVallisneria tubers was high (ie winter 2018) they mainlyfed on Vallisneria tubers and P criopolitanum rhizomesin shallow waters and grasslands Our study in combin-ation with previous studies indicates that the diets of Si-berian Cranes have changed greatly since winter 2010 [3844 62] They have gradually broadened their dietaryniches which has probably been driven by the degradationof submerged plants [38 44 57]Similar to Siberian Cranes a diet shift was also ob-
served in White-naped Cranes Historically White-naped Cranes only fed on submerged plants in shallowwaters and mudflats of Poyang Lake [91] Then drivenby the decline of submerged plants they primarily fed atgrasslands and mudflats [87 91] consuming Vallisneriatuber T edulis P limprichtii and Ranunculus polii [5587] Our results indicate that in addition to T edulisand P limprichtii that had previously been reported theproportions of P criopolitanum and Carex were alsohigh in the feces Hooded Cranes were suggested tomainly feed on the grassland plant P limprichtii throughdirect observation [86] Our study indicates that thegrassland plants P criopolitanum and Carex also occu-pied high proportions in the feces The dominant fooditems in the feces of White-naped Cranes and HoodedCranes varied greatly between the two winters The vari-ation in dietary composition indicated that the diversityand composition of the grassland plant communitymight have changed greatly between the two wintersThe dietary niche overlap between White-naped Cranesand Hooded Cranes was the highest among the fourcrane species aligning with their level of habitat niche
overlap which was also the highest among the species[92] The high dietary and habitat niche overlaps suggesthigh competition potential between the two cranespeciesThe Eurasian Crane is a habitat generalist It is known
to feed in a variety of habitats including grasslandsmudflats shallow waters and agricultural fields [88 92]Its diet at Poyang Lake has not been documented previ-ously Our study indicates that P criopolitanum P lim-prichtii and rice were the dominant food items in thefeces of Eurasian Cranes The number of plant itemsconsumed dietary niche breadth and Shannon-Wienermeasures were the highest among the four crane speciessupporting its generalist foraging strategy In contrast tothe three crane species above Eurasian Cranesrsquo dietcompositions were similar between the two winters Riceand grassland plants occupied high proportions in thefeces in both winters
Important food resources for cranesVallisneria tubers have been acknowledged as importantfood sources for cranes especially Siberian Cranes andWhite-naped Cranes [55 56] Our study confirms theimportance of Vallisneria tubers in the diet of SiberianCranes It was the most common food source in thefeces of Siberian Cranes in winter 2018 when its abun-dance was high Vallisneria tuber proportions in thefeces of the three other crane species were not as high asthat in the feces of Siberian Cranes but it still occupied5ndash9 of the total foods in winter 2018To our surprise P criopolitanum was found to be the
first dominant food in the feces of White-naped CranesHooded Cranes and Eurasian Cranes and the seconddominant food in the feces of Siberian Cranes in winter2018 In winter 2017 the proportions of P criopolita-num were lt 17 in the feces of Siberian Cranes andWhite-naped Cranes and were lt 3 in the feces ofHooded Cranes and Eurasian Cranes The higher pro-portions of P criopolitanum in winter 2018 than winter2017 indicate that the abundance and availability of Pcriopolitanum was much higher in winter 2018 P crio-politanum is an annual grassland plant It is sensitive tohydrological conditions and can only survive in a rela-tively narrow range of water depths and percent time in-undated [93] Due to the sensitivity its distributionrange and abundance showed high interannual variation[94 95] Thus it is likely that the abundance of P crio-politanum was higher in winter 2018 than in winter2017The importance of P criopolitanum as a food source
for the cranes at Poyang Lake has not been documentedpreviously It has only been suggested to be a major foodsource of Hooded Cranes at Shengjin Lake another lakein the middle and lower Yangtze River floodplain [86
Hou et al Frontiers in Zoology (2021) 1824 Page 8 of 17
96] P criopolitanum is a dominant wetland plant at Po-yang Lake [93 97] The coverage percentage of this plantis the third highest among grassland plants accountingfor 12 of the total areas of grasslands [97] This plantspecies is usually distributed at elevations of 9ndash12mabove sea level near mudflats and shallow waters [97]which are frequently used by cranes Its rhizome with ashallow burial depth is high in energy and nutritionalcontent and low in crude fibre [38 86] All these charac-teristics make P criopolitanum rhizome a potentially im-portant food source for cranes We frequently observedcranes dig and feed on rhizomes of P criopolitanum andleave aboveground parts in the fields Therefore we be-lieve that P criopolitanum was used as food not inci-dentally consumed by cranes However we need to becaution that high proportion of P criopolitanum in thefeces does not mean high proportion in the diets Morework on the importance of P criopolitanum in the dietsof cranes is needed
The roles of agricultural fields in crane protectionWith the loss and degradation of natural wetlands theroles of artificial wetlands in waterbird protection havebeen under intensive debate [98 99] Although agricul-tural fields cannot completely replace the function ofnatural wetlands as waterbird habitats [100ndash102] agri-cultural fields might provide alternative or complemen-tary habitats for waterbirds in all life stages [38 103104] The proportions of domesticated species in thefeces of Siberian Cranes varied considerably betweenyears In winter 2017 when the abundance of Vallisneriatubers was low the proportion of Vallisneria in SiberianCranesrsquo feces was only 095 Siberian Cranes may nothave found enough foods in the shallow waters andmudflats and were forced to search for rice seeds andlotus rhizomes in agricultural fields In winter 2018when the abundance of Vallisneria tubers reboundedSiberian Cranes moved back to shallow waters and mud-flats and a large proportion of their traditional food Val-lisneria tubers and also P criopolitanum were detectedin the feces A similar abandonment of tuber foragingoccurred following the tuber collapse in 2010 and thenresumed in the next winter when tubers rebounded in2011 [38 44] The diet and habitat changes indicate thatagricultural habitats were important refuges for SiberianCranes providing important food resources for them tosurvive through winters with few Vallisneria tubersThey also indicate that Siberian Cranes prefer naturalwetlands to agricultural fields [38]Similar to Siberian Cranes White-naped Cranes and
Hooded Cranes consumed rice seeds and lotus rhizomesin winter 2017 but did not consume any in winter 2018The changes in the proportions suggest that the abun-dance of food resources in natural wetlands might have
been low in winter 2017 The deficient food resourcesmight have been Vallisneria tubers but might also havebeen grassland plants the primary food sources of thesetwo crane species Under natural food shortage condi-tions (ie winter 2017) White-naped Cranes andHooded Cranes utilized agricultural fields When theabundance of Vallisneria tubers and probably grasslandplants were high (ie winter 2018) the two crane speciesreturned to their natural habitats Therefore for White-naped Cranes and Hooded Cranes agricultural fieldswere important alternative foraging habitats used asbuffers against starvation during periods of natural foodshortages The return of the two crane species to naturalwetlands in winter 2018 further emphasizes the superiorquality of natural habitats over agricultural fields as for-aging habitats for cranes [38]The proportions of rice seeds in the feces of Eur-
asian Cranes were high in the two winters thoughthe proportion in winter 2017 was twice as high asthat in winter 2018 Therefore agricultural fields ap-peared to be consistently important as foraging habi-tats for Eurasian Cranes The frequent use ofagricultural fields by Eurasian Cranes has been docu-mented widely [49 105 106]Domesticated species generally contain high energy
and nutritional content and are of superior quality com-pared to natural plants [86 104] Farmland foraging hasbeen suggested to contribute to increase in the abun-dance of some goose [107] egret [108] and wader [109]populations in Japan Korea Europe and North Amer-ica However cranes generally avoid human disturbancewhen select foraging sites [110 111] Human disturb-ance in agricultural fields around Poyang Lake is consid-erably high due to intensive agricultural activity [112]High human disturbance was suggested to be a majorreason that wintering geese were almost confined withinnatural wetlands and hesitated to exploit the riches ofthe modern agricultural habitats at Poyang Lake [112113] High human disturbance also resulted in SiberianCranes spending twice as much time alerting in agricul-tural fields than they did in natural habitats [114] Thehigh level of human disturbance in agricultural fieldsmight contribute to the preference for natural wetlandsby Siberian Cranes White-naped Cranes and HoodedCranes at Poyang Lake Moreover although agriculturefields contain energy-rich crops wetlands contain di-verse food resources especially protein-rich inverte-brates which might better satisfy dietary needs of cranes[110 115] Taken together security and diverse food re-sources might have attracted cranes to select wetlands asforaging sitesSubmerged macrophytes at Poyang Lake have de-
graded in recent decades [57ndash60] and Vallisneria tuberdisappearance is expected to occur more frequently
Hou et al Frontiers in Zoology (2021) 1824 Page 9 of 17
under future climate change [57 60] Cranes may in-creasingly depend upon agricultural fields around Po-yang Lake which may in turn cause significant cropdamage and economic losses This is especially true onagricultural fields surrounding protected wetlands wherewaterbirds concentrate in high numbers [116 117] AtPoyang Lake cranes mainly fed on post-harvest remainsof rice seeds and lotus rhizomes thus crop damage andeconomic loss were limited However we observed somefarmers scared cranes away from their agricultural fieldsThere were more than 14 million free-ranging poultryraised at agricultural fields around Poyang Lake [118]where they gleaned waste grain residues in stubble fieldsCrane feeding would reduce the food abundance of thefree-ranging domestic poultry thus farmers drovecranes away To reduce human disturbance environ-mental education is needed because farmers are morelikely to protect cranes if they understand the endan-gered degree and the ecological significance of cranes
Effects of tuber collapse on dietary niche breadth andoverlap among crane speciesVallisneria tuber is an important food source for Siber-ian Cranes (89 90 this study) thus variation in itsabundance might influence Siberian Cranesrsquo dietaryniche breadth The numbers of food items dietary nichebreadth and Shannon-Wiener measures of SiberianCranes were similar between winter 2017 and winter2018 This is inconsistent with the contention that dur-ing seasons with a low abundance of dominant food thedietary niche may broaden as consumers relying on in-sufficient preferred food items are forced to add lessprofitable resources to their diets [26 119 120] In win-ter 2017 with low Vallisneria tuber abundance SiberianCranes left shallow waters and mudflats and moved torice paddies and lotus ponds [61 62 64] Their foodschanged from Vallisneria tuber to rice seeds and lotusrhizomes almost completely As the energy protein andfat contents of crops are generally as good as or superiorto natural foods and are often present in agriculturalfields in far greater abundance [86 104] Siberian Cranesmay gain enough energy through feeding on crops anddo not need to add many other food sources Moreoverthe Siberian Crane is a dietary specialist [89 90] PoyangLake has a relatively simple plant community compos-ition with Vallisneria and Hydrilla verticillate being thedominant submerged macrophytes [59] and Carex beingthe dominant grassland plants [121] Therefore theremay be few options for Siberian Cranes which mightlead to the failure of this species to widen its dietaryniche breadthIn contrast to Siberian Cranes the numbers of food
items dietary niche breadths and Shannon-Wiener mea-sures of White-naped Cranes and Hooded Cranes were
higher in winter 2017 than in winter 2018 P criopolita-num was the predominant food item in the feces of thetwo crane species in winter 2018 and the proportions ofVallisneria tubers were also high The abundances ofVallisneria tubers and probably P criopolitanum at Po-yang Lake declined greatly in winter 2017 When thereis a short-fall in food availability animals may optimizeforaging behavior and increase their dietary breath [122123] The short-fall in the abundances of P criopolita-num and Vallisneria tubers in winter 2017 might havedriven the two crane species to add less profitable grass-land plants in their diets Although the two cranes alsoconsumed high-quality rice seeds and lotus rhizomes inwinter 2017 their proportions were low in the feces andthese foods might be unable to meet the energy require-ments of the two crane species The niche expansion ofthe two crane species during periods of preferred foodshortage was in agreement with the basic prediction ofoptimal foraging theory [22 23] It is also consistentwith a commonly observed fact that energy-maximizingmodels work better when animals food-limited [124]The dietary niche overlaps among the crane species
were low in winter 2017 with the four crane species di-verging in their principal food items However in winter2018 the niche overlaps were significantly higher thanexpected by chance They had similar diet compositionswith P criopolitanum rhizomes and Vallisneria tubersbeing the principal food items in the feces The in-creased overlap might have been caused by the highabundances of the two plant species in winter 2018which allowed the four crane species to share the samefood items with low interspecific competition Our studyis consistent with previous research that suggested thatamong-species dietary niches tended to overlap moreduring seasons of high food abundance [32 33 125126] Greater resource availability could promote an in-crease in species utilization of similar resources and re-duce competitive interactions between species [32 34126] By contrast during seasons with low food abun-dance species may partition limited food resources todecrease interspecific competition leading to a decreasein niche overlap [126 127] It has been suggested thatdiet partitioning facilitates the coexistence of manyclosely related species [7 128] The interannual variationin niche overlap observed in our study suggests that dietpartitioning may be restricted to specific temporal casesIn seasons or years with abundant food sources inter-specific competition may be relaxed and niche overlapcould be high
ConclusionsIn summary by using metabarcoding technology ourstudy gained new insights into cranesrsquo diets that are es-sential for informing habitat management to improve
Hou et al Frontiers in Zoology (2021) 1824 Page 10 of 17
the availability of foraging resources Vallisneria tuberswas confirmed as an important food source of cranesSurprisingly P criopolitanum also occupied high pro-portions in the feces of cranes and may also be import-ant Our study emphasizes the superior quality ofnatural wetlands compared to agricultural fields as for-aging habitats for cranes The recent degradation of sub-merged macrophytes at Poyang Lake potentiallythreatens the survival of cranes especially SiberianCranes Therefore it is urgent to reverse the broad de-cline of submerged macrophytes and provide importantforaging habitats for cranes during years of tuber col-lapses The abundances of Vallisneria tubers and prob-ably P criopolitanum at Poyang Lake greatly influencedthe dietary niche width and overlaps among crane spe-cies Cranes exhibited low dietary niche overlap duringperiods of preferred food shortage but significantly highoverlap during periods when preferred foods were plen-tiful While high niche overlap is often interpreted as in-tense interspecific competition our study highlights theimportance of taking food abundance into account
Materials and methodsStudy areasPoyang Lake the largest freshwater lake in China is oneof the most important wintering areas for waterbirdsalong the East Asian-Australasian Flyway (Fig 1) [51129] It is characterized by dramatic seasonal hydro-logical fluctuations with an average change of 924 mbetween the summer high and winter low water levels[130] During summer the inundation area is gt 2500km2 [131] In autumn and winter the water level dropscausing the inundation area to shrink to lt 1000 km2
[131] Poyang Lake contains 102 sublakes which are in-undated and integrated with the main body of PoyangLake during summer and are isolated during winter[132] As the water level drops the sublakes appear andlarge areas of grasslands mudflats and shallow waterareas are exposed providing winter foraging and roost-ing areas for approximately 420000 waterbirds from ap-proximately 111 species [133 134]
Vallisneria tuber surveyThe density and biomass of Vallisneria tubers were sur-veyed at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake in late October and earlyNovember of 2017 and 2018 before large numbers ofwintering waterbirds arrived We set 76 183 and 36sampling plots uniformly located at Dahuchi Shahu andChanghuchi respectively Vallisneria tubers were col-lected from two quadrats in each plot with a locallymade steel grab sampler The sampler has two long
handles At the end of the handles there are two scoopsfacing each other The sampler was inserted into thesubstrate and the scoops brought together to collectabout 15 cm long times 13 cm wide times 30 cm high sample ofsubstrate Tubers were subsequently cleaned andcounted The dry weight of tubers in each quadrat wasweighed after oven-drying at 80 degC until constant weightThe tuber density and biomass in each plot was calcu-lated by dividing total tuber number and tuber massacross two quadrats by quadrat area respectively Weused t-test to explore whether the tuber density and bio-mass in the winters of 2017 and 2018 significantly devi-ated from the historical average values in the wintersfrom 1999 to 2016 [57] The historical tuber density andbiomass data were collected by the Poyang Lake Na-tional Nature Reserve at three sublakes (Dahuchi Shahuand Meixihu) [57] R 360 was used to do the t-testanalyses
Fecal sample collectionFecal samples were collected at Poyang Lake in the win-ters of 2017 and 2018 We first investigated the distribu-tions of cranes When we saw foraging cranes we waiteduntil they left to avoid disturbing them Soon after theyleft we went to the foraging sites to collect fresh feces(ie feces with wet surface) with sterilized tweezers Tominimize the probability of multiple samples from thesame individual all collected samples were separated byat least 5 m We stored the samples in liquid nitrogen inthe field and transferred them to a minus 80 degC refrigeratorfor long-term storage in the laboratory To ensure thatthe sources of the feces were known we tried to collectsamples from monospecific flocks For samples collectedfrom mixed-species flocks we used DNA analysis toidentify the species that deposited the fecal samplesPlants were also collected in the field to construct aDNA barcoding reference database
Species identificationBecause of the similar morphologies of the feces of thefour crane species we used DNA analysis to identify thespecies that deposited the fecal samples collected frommixed-species flocks For the genetic species assignmentof each fecal sample we scraped the external surfacewhich contain DNA from the host species as a result ofthe sloughing of cells from the digestive tract DNA wasextracted with the QIAamp Power Fecal DNA Kit (Qia-gen USA) according to the manufacturerrsquos guidelinesFor each round of DNA extraction negative controls(ie extraction without feces) were included to monitorfor possible contamination We amplified the mitochon-drial DNA D-loop fragment with the primer pair DL02F
Hou et al Frontiers in Zoology (2021) 1824 Page 11 of 17
(5prime-3prime GCGGCCCGAAAAGCCGCTG) and DL02R (5prime-3prime GCCCTGACATAGGAACCAGAGGCGC) [135]PCR amplifications were carried out in a total volume of25 μl containing 125 μl PCR mix (Tiangen BeijingChina) 1 μl DNA 1 μl each primer and 95 μl H2O Thereaction conditions were as follows denaturation at94 degC for 4 min followed by 34 cycles at 94 degC for 48 s56 degC for 48 s and 72 degC for 1 min and a final 10 min at72 degC We checked for the presence of a PCR product ofsuitable length by electrophoresis on a 1 agarose gelSequencing was carried out on an ABI 3730xl analyzerby Sangon Biotech (Shanghai China) The complete se-quences were assembled using Seqman II (DNASTAR)and compared visually to the original chromatograms toavoid reading errors The taxonomic assignment wasconducted by searching against the nucleotide databaseof GenBank using Web BLAST
Construction of a barcode reference databaseWe downloaded the trnL sequences of 48 plant speciescommonly seen at Poyang Lake from GenBank For theplant species not included in GenBank we sequencedtrnL by ourselves Total DNA was extracted using thehexadecyltrimethylammonium bromide (CTAB) proto-col We used primer pair c (5prime-3prime CGAAATCGGTAGACGCTACG) and h (5prime-3prime CCATTGAGTCTCTGCACCTATC) to amplify an approximately 150 bp regionof the chloroplast trnL intron [136] This primer pairwas chosen as it can amplify DNA from a wide range ofwetland plants [70] PCR amplifications were carried outin a total volume of 25 μl containing 125 μl PCR mix(Tiangen Beijing China) 1 μl DNA 1 μl each primerand 95 μl H2O The reaction conditions were as followsdenaturation at 94 degC for 4 min followed by 35 cycles at94 degC for 30 s 56 degC for 30 s and 72 degC for 45 s and afinal 10 min at 72 degC We used electrophoresis to checkthe PCR products ABI 3730xl analyzer to perform se-quencing and Seqman II to assemble the sequences
DNA extraction and high-throughput sequencing of fecalsamplesTo uncover plant component consumed by cranes amodified CTAB protocol [70] was used to extract theDNA from approximately 200 mg of the interior of eachfecal sample DNA extraction was conducted in a dedi-cated laboratory for analyses of DNA from samples withlow DNA quality For each batch of DNA extractionnegative controls were included to monitor for possiblecontamination The primers c and h with a 6 bp tagadded to 5prime ends of each primer were used to amplifytrnL gene The PCR amplification procedure was thesame as barcode database construction For each samplea unique combination of tags in the forward and reverseprimers was used so that sample-specific data can be
recovered after sequencing Three PCR replicates wereperformed for each sample to reduce amplification biasThe three replicates were pooled and the DNA concen-tration and purity were monitored on a 1 agarose gelOnly samples with a clear band were processed furtherSamples were purified using the SanPrep Column PCRProduct Purification Kit (Sangon Biotech ShanghaiChina) The purified PCR products were pooled takinginto account of DNA concentration to obtain an ap-proximately equal amount of amplicon DNA from eachfecal sample The pool of individually tagged ampliconswas used to prepare a sequencing library using the Tru-Seq DNA PCR-Free Sample Preparation Kit (IlluminaUSA) following manufacturerrsquos recommendations Thelibrary was sequenced on an Illumina HiSeq 2500 plat-form and 250 bp paired-end reads were generated Li-brary preparation and sequencing were conducted byNovogene (Beijing China)
Sequencing data analysisAfter high-throughput sequencing the paired-end readswere demultiplexed into sample-specific files based ontheir unique tags The tags and primer sequences weresubsequently trimmed The paired-end reads weremerged using FLASH 127 [137] and then filtered usingQIIME 170 [138] Sequences were dereplicated andclustered to OTUs using USEARCH 110667 [139] atthe similarity threshold of 97 All OTUs were assignedto taxonomic units by referring to the local plant data-base using blast 2231+ with thresholds of identity gt97 and e-value lt 10 eminus 50 If an OTU matched two ormore taxa it was assigned to a higher taxonomic levelthat included all taxaTo evaluate the sufficiency of sequencing depth and
sampling size individual- species- and sample-basedrarefaction curves were built using EstimateS 82[140] The curves were computed by randomly resam-pling sequences or samples and plotting these againstthe numbers of OTUs The read abundance data wereconverted to RRA of each food item using the follow-ing equation [79]
RRA frac14 1S
XS
Kfrac141
nikPTifrac141nik
100
where nik is the number of sequences of food item i insample k T is the total number of food items and S isthe sampling size
Dietary niche breadth and overlap among crane speciesTo explore whether the cranesrsquo diets showed substantialinterannual variation we calculated Pearson correlationsbetween diet compositions of each crane species in win-ter 2017 and winter 2018 The proportion of each food
Hou et al Frontiers in Zoology (2021) 1824 Page 12 of 17
item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

that are overlooked by microhistological methods [6780] It also enables the identification of food items at ahigher taxonomic resolution [69 81] However we ac-knowledge that our study has two limitations First weused fecal samples as they are non-invasive Neverthe-less fecal analysis is subjected to the biases caused bythe differential digestive rates of food items [82 83] Softfoods such as insect larvae tend to be under-representedin fecal samples because they are digested quickly andremain in gastro-intestinal tract for short periods whilehard foods such as mollusks tend to be over-representedbecause they break down slowly and remain in gastro-intestinal tracts longer Thus food proportions in the
feces may not correlate with food proportions ingestedby cranes In addition some materials identified in thefeces may be incidentally consumed by cranes and notreally be used as food Second our study only consideredthe plant component of cranesrsquo diets as plants were sug-gested to be their principal food types at Poyang LakeAnimal foods consumed by the four crane species butto a much lesser extent than plant foods [84ndash87] werenot considered in our studyThe Siberian Crane is regarded as the most aquatic of
all cranes using wetlands for nesting feeding and roost-ing [49 88] The high dependence of the species onaquatic habitats was suggested to make it vulnerable to
Table 3 The observed dietary niche overlap among the four crane species at Poyang Lake in the winters of 2017 and 2018 P(Obs lt = null) and P (Obs gt = null) indicate the tail probabilities for the observed index which were lower and higher than thehistogram of the simulated index respectively represents P lt 005 and represents P lt 001
Winter Observed niche overlap index P (Obs lt = null) P (Obs gt = null)
2017 Siberian vs White-naped 021 0819 0181
Siberian vs Hooded 002 0703 0297
Siberian vs Eurasian 016 0900 0100
White-naped vs Hooded 049 0952 0048
White-naped vs Eurasian 035 0895 0106
Hooded vs Eurasian 035 0919 0081
2018 Siberian vs White-naped 063 0952 0048
Siberian vs Hooded 066 0952 0048
Siberian vs Eurasian 062 0950 0049
White-naped vs Hooded 099 0997 0003
White-naped vs Eurasian 089 0987 0013
Hooded vs Eurasian 090 0998 0002
Fig 4 The results of nonmetric multidimensional scaling ordination (NMDS) based on Bray-Curtis dissimilarity The colors of the circlescorrespond to the crane species blue Siberian Crane purple White-naped Crane green Hooded Crane and red Eurasian Crane
Hou et al Frontiers in Zoology (2021) 1824 Page 7 of 17
impacts of habitat degradation and to lead its classifica-tion as Critically Endangered [49] At Poyang Lake Si-berian Cranes traditionally fed on tubers of Vallisneriathe dominant submerged macrophytes in shallow watersand mudflats [89 90] In winter 2010 due to a flood-induced Vallisneria tuber collapse they were observedforaging for P limprichtii taproots and T edulis bulbs ingrasslands for the first time [38 44] In the winters from2015 to 2017 the diets of Siberian Cranes changedagain with thousands of cranes feeding on rice seedsand lotus rhizomes in agricultural habitats [61 62 64]Our study identified 22 plant items in the feces of Siber-
ian Cranes These plant items were obtained from shallowwaters grasslands and agricultural fields Four plant itemswith yearly RRA gt 10 were identified in the feces Vallis-neria tuber P criopolitanum rhizomes rice seeds andlotus rhizomes (the vegetative portions consumed bycranes were based on our field observation) SiberianCranesrsquo diet compositions showed substantial interannualvariation When the abundance of Vallisneria tubers waslow (ie winter 2017) Siberian Cranes mainly fed on riceseeds lotus rhizomes and P criopolitanum rhizomes inagricultural fields and grasslands When the abundance ofVallisneria tubers was high (ie winter 2018) they mainlyfed on Vallisneria tubers and P criopolitanum rhizomesin shallow waters and grasslands Our study in combin-ation with previous studies indicates that the diets of Si-berian Cranes have changed greatly since winter 2010 [3844 62] They have gradually broadened their dietaryniches which has probably been driven by the degradationof submerged plants [38 44 57]Similar to Siberian Cranes a diet shift was also ob-
served in White-naped Cranes Historically White-naped Cranes only fed on submerged plants in shallowwaters and mudflats of Poyang Lake [91] Then drivenby the decline of submerged plants they primarily fed atgrasslands and mudflats [87 91] consuming Vallisneriatuber T edulis P limprichtii and Ranunculus polii [5587] Our results indicate that in addition to T edulisand P limprichtii that had previously been reported theproportions of P criopolitanum and Carex were alsohigh in the feces Hooded Cranes were suggested tomainly feed on the grassland plant P limprichtii throughdirect observation [86] Our study indicates that thegrassland plants P criopolitanum and Carex also occu-pied high proportions in the feces The dominant fooditems in the feces of White-naped Cranes and HoodedCranes varied greatly between the two winters The vari-ation in dietary composition indicated that the diversityand composition of the grassland plant communitymight have changed greatly between the two wintersThe dietary niche overlap between White-naped Cranesand Hooded Cranes was the highest among the fourcrane species aligning with their level of habitat niche
overlap which was also the highest among the species[92] The high dietary and habitat niche overlaps suggesthigh competition potential between the two cranespeciesThe Eurasian Crane is a habitat generalist It is known
to feed in a variety of habitats including grasslandsmudflats shallow waters and agricultural fields [88 92]Its diet at Poyang Lake has not been documented previ-ously Our study indicates that P criopolitanum P lim-prichtii and rice were the dominant food items in thefeces of Eurasian Cranes The number of plant itemsconsumed dietary niche breadth and Shannon-Wienermeasures were the highest among the four crane speciessupporting its generalist foraging strategy In contrast tothe three crane species above Eurasian Cranesrsquo dietcompositions were similar between the two winters Riceand grassland plants occupied high proportions in thefeces in both winters
Important food resources for cranesVallisneria tubers have been acknowledged as importantfood sources for cranes especially Siberian Cranes andWhite-naped Cranes [55 56] Our study confirms theimportance of Vallisneria tubers in the diet of SiberianCranes It was the most common food source in thefeces of Siberian Cranes in winter 2018 when its abun-dance was high Vallisneria tuber proportions in thefeces of the three other crane species were not as high asthat in the feces of Siberian Cranes but it still occupied5ndash9 of the total foods in winter 2018To our surprise P criopolitanum was found to be the
first dominant food in the feces of White-naped CranesHooded Cranes and Eurasian Cranes and the seconddominant food in the feces of Siberian Cranes in winter2018 In winter 2017 the proportions of P criopolita-num were lt 17 in the feces of Siberian Cranes andWhite-naped Cranes and were lt 3 in the feces ofHooded Cranes and Eurasian Cranes The higher pro-portions of P criopolitanum in winter 2018 than winter2017 indicate that the abundance and availability of Pcriopolitanum was much higher in winter 2018 P crio-politanum is an annual grassland plant It is sensitive tohydrological conditions and can only survive in a rela-tively narrow range of water depths and percent time in-undated [93] Due to the sensitivity its distributionrange and abundance showed high interannual variation[94 95] Thus it is likely that the abundance of P crio-politanum was higher in winter 2018 than in winter2017The importance of P criopolitanum as a food source
for the cranes at Poyang Lake has not been documentedpreviously It has only been suggested to be a major foodsource of Hooded Cranes at Shengjin Lake another lakein the middle and lower Yangtze River floodplain [86
Hou et al Frontiers in Zoology (2021) 1824 Page 8 of 17
96] P criopolitanum is a dominant wetland plant at Po-yang Lake [93 97] The coverage percentage of this plantis the third highest among grassland plants accountingfor 12 of the total areas of grasslands [97] This plantspecies is usually distributed at elevations of 9ndash12mabove sea level near mudflats and shallow waters [97]which are frequently used by cranes Its rhizome with ashallow burial depth is high in energy and nutritionalcontent and low in crude fibre [38 86] All these charac-teristics make P criopolitanum rhizome a potentially im-portant food source for cranes We frequently observedcranes dig and feed on rhizomes of P criopolitanum andleave aboveground parts in the fields Therefore we be-lieve that P criopolitanum was used as food not inci-dentally consumed by cranes However we need to becaution that high proportion of P criopolitanum in thefeces does not mean high proportion in the diets Morework on the importance of P criopolitanum in the dietsof cranes is needed
The roles of agricultural fields in crane protectionWith the loss and degradation of natural wetlands theroles of artificial wetlands in waterbird protection havebeen under intensive debate [98 99] Although agricul-tural fields cannot completely replace the function ofnatural wetlands as waterbird habitats [100ndash102] agri-cultural fields might provide alternative or complemen-tary habitats for waterbirds in all life stages [38 103104] The proportions of domesticated species in thefeces of Siberian Cranes varied considerably betweenyears In winter 2017 when the abundance of Vallisneriatubers was low the proportion of Vallisneria in SiberianCranesrsquo feces was only 095 Siberian Cranes may nothave found enough foods in the shallow waters andmudflats and were forced to search for rice seeds andlotus rhizomes in agricultural fields In winter 2018when the abundance of Vallisneria tubers reboundedSiberian Cranes moved back to shallow waters and mud-flats and a large proportion of their traditional food Val-lisneria tubers and also P criopolitanum were detectedin the feces A similar abandonment of tuber foragingoccurred following the tuber collapse in 2010 and thenresumed in the next winter when tubers rebounded in2011 [38 44] The diet and habitat changes indicate thatagricultural habitats were important refuges for SiberianCranes providing important food resources for them tosurvive through winters with few Vallisneria tubersThey also indicate that Siberian Cranes prefer naturalwetlands to agricultural fields [38]Similar to Siberian Cranes White-naped Cranes and
Hooded Cranes consumed rice seeds and lotus rhizomesin winter 2017 but did not consume any in winter 2018The changes in the proportions suggest that the abun-dance of food resources in natural wetlands might have
been low in winter 2017 The deficient food resourcesmight have been Vallisneria tubers but might also havebeen grassland plants the primary food sources of thesetwo crane species Under natural food shortage condi-tions (ie winter 2017) White-naped Cranes andHooded Cranes utilized agricultural fields When theabundance of Vallisneria tubers and probably grasslandplants were high (ie winter 2018) the two crane speciesreturned to their natural habitats Therefore for White-naped Cranes and Hooded Cranes agricultural fieldswere important alternative foraging habitats used asbuffers against starvation during periods of natural foodshortages The return of the two crane species to naturalwetlands in winter 2018 further emphasizes the superiorquality of natural habitats over agricultural fields as for-aging habitats for cranes [38]The proportions of rice seeds in the feces of Eur-
asian Cranes were high in the two winters thoughthe proportion in winter 2017 was twice as high asthat in winter 2018 Therefore agricultural fields ap-peared to be consistently important as foraging habi-tats for Eurasian Cranes The frequent use ofagricultural fields by Eurasian Cranes has been docu-mented widely [49 105 106]Domesticated species generally contain high energy
and nutritional content and are of superior quality com-pared to natural plants [86 104] Farmland foraging hasbeen suggested to contribute to increase in the abun-dance of some goose [107] egret [108] and wader [109]populations in Japan Korea Europe and North Amer-ica However cranes generally avoid human disturbancewhen select foraging sites [110 111] Human disturb-ance in agricultural fields around Poyang Lake is consid-erably high due to intensive agricultural activity [112]High human disturbance was suggested to be a majorreason that wintering geese were almost confined withinnatural wetlands and hesitated to exploit the riches ofthe modern agricultural habitats at Poyang Lake [112113] High human disturbance also resulted in SiberianCranes spending twice as much time alerting in agricul-tural fields than they did in natural habitats [114] Thehigh level of human disturbance in agricultural fieldsmight contribute to the preference for natural wetlandsby Siberian Cranes White-naped Cranes and HoodedCranes at Poyang Lake Moreover although agriculturefields contain energy-rich crops wetlands contain di-verse food resources especially protein-rich inverte-brates which might better satisfy dietary needs of cranes[110 115] Taken together security and diverse food re-sources might have attracted cranes to select wetlands asforaging sitesSubmerged macrophytes at Poyang Lake have de-
graded in recent decades [57ndash60] and Vallisneria tuberdisappearance is expected to occur more frequently
Hou et al Frontiers in Zoology (2021) 1824 Page 9 of 17
under future climate change [57 60] Cranes may in-creasingly depend upon agricultural fields around Po-yang Lake which may in turn cause significant cropdamage and economic losses This is especially true onagricultural fields surrounding protected wetlands wherewaterbirds concentrate in high numbers [116 117] AtPoyang Lake cranes mainly fed on post-harvest remainsof rice seeds and lotus rhizomes thus crop damage andeconomic loss were limited However we observed somefarmers scared cranes away from their agricultural fieldsThere were more than 14 million free-ranging poultryraised at agricultural fields around Poyang Lake [118]where they gleaned waste grain residues in stubble fieldsCrane feeding would reduce the food abundance of thefree-ranging domestic poultry thus farmers drovecranes away To reduce human disturbance environ-mental education is needed because farmers are morelikely to protect cranes if they understand the endan-gered degree and the ecological significance of cranes
Effects of tuber collapse on dietary niche breadth andoverlap among crane speciesVallisneria tuber is an important food source for Siber-ian Cranes (89 90 this study) thus variation in itsabundance might influence Siberian Cranesrsquo dietaryniche breadth The numbers of food items dietary nichebreadth and Shannon-Wiener measures of SiberianCranes were similar between winter 2017 and winter2018 This is inconsistent with the contention that dur-ing seasons with a low abundance of dominant food thedietary niche may broaden as consumers relying on in-sufficient preferred food items are forced to add lessprofitable resources to their diets [26 119 120] In win-ter 2017 with low Vallisneria tuber abundance SiberianCranes left shallow waters and mudflats and moved torice paddies and lotus ponds [61 62 64] Their foodschanged from Vallisneria tuber to rice seeds and lotusrhizomes almost completely As the energy protein andfat contents of crops are generally as good as or superiorto natural foods and are often present in agriculturalfields in far greater abundance [86 104] Siberian Cranesmay gain enough energy through feeding on crops anddo not need to add many other food sources Moreoverthe Siberian Crane is a dietary specialist [89 90] PoyangLake has a relatively simple plant community compos-ition with Vallisneria and Hydrilla verticillate being thedominant submerged macrophytes [59] and Carex beingthe dominant grassland plants [121] Therefore theremay be few options for Siberian Cranes which mightlead to the failure of this species to widen its dietaryniche breadthIn contrast to Siberian Cranes the numbers of food
items dietary niche breadths and Shannon-Wiener mea-sures of White-naped Cranes and Hooded Cranes were
higher in winter 2017 than in winter 2018 P criopolita-num was the predominant food item in the feces of thetwo crane species in winter 2018 and the proportions ofVallisneria tubers were also high The abundances ofVallisneria tubers and probably P criopolitanum at Po-yang Lake declined greatly in winter 2017 When thereis a short-fall in food availability animals may optimizeforaging behavior and increase their dietary breath [122123] The short-fall in the abundances of P criopolita-num and Vallisneria tubers in winter 2017 might havedriven the two crane species to add less profitable grass-land plants in their diets Although the two cranes alsoconsumed high-quality rice seeds and lotus rhizomes inwinter 2017 their proportions were low in the feces andthese foods might be unable to meet the energy require-ments of the two crane species The niche expansion ofthe two crane species during periods of preferred foodshortage was in agreement with the basic prediction ofoptimal foraging theory [22 23] It is also consistentwith a commonly observed fact that energy-maximizingmodels work better when animals food-limited [124]The dietary niche overlaps among the crane species
were low in winter 2017 with the four crane species di-verging in their principal food items However in winter2018 the niche overlaps were significantly higher thanexpected by chance They had similar diet compositionswith P criopolitanum rhizomes and Vallisneria tubersbeing the principal food items in the feces The in-creased overlap might have been caused by the highabundances of the two plant species in winter 2018which allowed the four crane species to share the samefood items with low interspecific competition Our studyis consistent with previous research that suggested thatamong-species dietary niches tended to overlap moreduring seasons of high food abundance [32 33 125126] Greater resource availability could promote an in-crease in species utilization of similar resources and re-duce competitive interactions between species [32 34126] By contrast during seasons with low food abun-dance species may partition limited food resources todecrease interspecific competition leading to a decreasein niche overlap [126 127] It has been suggested thatdiet partitioning facilitates the coexistence of manyclosely related species [7 128] The interannual variationin niche overlap observed in our study suggests that dietpartitioning may be restricted to specific temporal casesIn seasons or years with abundant food sources inter-specific competition may be relaxed and niche overlapcould be high
ConclusionsIn summary by using metabarcoding technology ourstudy gained new insights into cranesrsquo diets that are es-sential for informing habitat management to improve
Hou et al Frontiers in Zoology (2021) 1824 Page 10 of 17
the availability of foraging resources Vallisneria tuberswas confirmed as an important food source of cranesSurprisingly P criopolitanum also occupied high pro-portions in the feces of cranes and may also be import-ant Our study emphasizes the superior quality ofnatural wetlands compared to agricultural fields as for-aging habitats for cranes The recent degradation of sub-merged macrophytes at Poyang Lake potentiallythreatens the survival of cranes especially SiberianCranes Therefore it is urgent to reverse the broad de-cline of submerged macrophytes and provide importantforaging habitats for cranes during years of tuber col-lapses The abundances of Vallisneria tubers and prob-ably P criopolitanum at Poyang Lake greatly influencedthe dietary niche width and overlaps among crane spe-cies Cranes exhibited low dietary niche overlap duringperiods of preferred food shortage but significantly highoverlap during periods when preferred foods were plen-tiful While high niche overlap is often interpreted as in-tense interspecific competition our study highlights theimportance of taking food abundance into account
Materials and methodsStudy areasPoyang Lake the largest freshwater lake in China is oneof the most important wintering areas for waterbirdsalong the East Asian-Australasian Flyway (Fig 1) [51129] It is characterized by dramatic seasonal hydro-logical fluctuations with an average change of 924 mbetween the summer high and winter low water levels[130] During summer the inundation area is gt 2500km2 [131] In autumn and winter the water level dropscausing the inundation area to shrink to lt 1000 km2
[131] Poyang Lake contains 102 sublakes which are in-undated and integrated with the main body of PoyangLake during summer and are isolated during winter[132] As the water level drops the sublakes appear andlarge areas of grasslands mudflats and shallow waterareas are exposed providing winter foraging and roost-ing areas for approximately 420000 waterbirds from ap-proximately 111 species [133 134]
Vallisneria tuber surveyThe density and biomass of Vallisneria tubers were sur-veyed at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake in late October and earlyNovember of 2017 and 2018 before large numbers ofwintering waterbirds arrived We set 76 183 and 36sampling plots uniformly located at Dahuchi Shahu andChanghuchi respectively Vallisneria tubers were col-lected from two quadrats in each plot with a locallymade steel grab sampler The sampler has two long
handles At the end of the handles there are two scoopsfacing each other The sampler was inserted into thesubstrate and the scoops brought together to collectabout 15 cm long times 13 cm wide times 30 cm high sample ofsubstrate Tubers were subsequently cleaned andcounted The dry weight of tubers in each quadrat wasweighed after oven-drying at 80 degC until constant weightThe tuber density and biomass in each plot was calcu-lated by dividing total tuber number and tuber massacross two quadrats by quadrat area respectively Weused t-test to explore whether the tuber density and bio-mass in the winters of 2017 and 2018 significantly devi-ated from the historical average values in the wintersfrom 1999 to 2016 [57] The historical tuber density andbiomass data were collected by the Poyang Lake Na-tional Nature Reserve at three sublakes (Dahuchi Shahuand Meixihu) [57] R 360 was used to do the t-testanalyses
Fecal sample collectionFecal samples were collected at Poyang Lake in the win-ters of 2017 and 2018 We first investigated the distribu-tions of cranes When we saw foraging cranes we waiteduntil they left to avoid disturbing them Soon after theyleft we went to the foraging sites to collect fresh feces(ie feces with wet surface) with sterilized tweezers Tominimize the probability of multiple samples from thesame individual all collected samples were separated byat least 5 m We stored the samples in liquid nitrogen inthe field and transferred them to a minus 80 degC refrigeratorfor long-term storage in the laboratory To ensure thatthe sources of the feces were known we tried to collectsamples from monospecific flocks For samples collectedfrom mixed-species flocks we used DNA analysis toidentify the species that deposited the fecal samplesPlants were also collected in the field to construct aDNA barcoding reference database
Species identificationBecause of the similar morphologies of the feces of thefour crane species we used DNA analysis to identify thespecies that deposited the fecal samples collected frommixed-species flocks For the genetic species assignmentof each fecal sample we scraped the external surfacewhich contain DNA from the host species as a result ofthe sloughing of cells from the digestive tract DNA wasextracted with the QIAamp Power Fecal DNA Kit (Qia-gen USA) according to the manufacturerrsquos guidelinesFor each round of DNA extraction negative controls(ie extraction without feces) were included to monitorfor possible contamination We amplified the mitochon-drial DNA D-loop fragment with the primer pair DL02F
Hou et al Frontiers in Zoology (2021) 1824 Page 11 of 17
(5prime-3prime GCGGCCCGAAAAGCCGCTG) and DL02R (5prime-3prime GCCCTGACATAGGAACCAGAGGCGC) [135]PCR amplifications were carried out in a total volume of25 μl containing 125 μl PCR mix (Tiangen BeijingChina) 1 μl DNA 1 μl each primer and 95 μl H2O Thereaction conditions were as follows denaturation at94 degC for 4 min followed by 34 cycles at 94 degC for 48 s56 degC for 48 s and 72 degC for 1 min and a final 10 min at72 degC We checked for the presence of a PCR product ofsuitable length by electrophoresis on a 1 agarose gelSequencing was carried out on an ABI 3730xl analyzerby Sangon Biotech (Shanghai China) The complete se-quences were assembled using Seqman II (DNASTAR)and compared visually to the original chromatograms toavoid reading errors The taxonomic assignment wasconducted by searching against the nucleotide databaseof GenBank using Web BLAST
Construction of a barcode reference databaseWe downloaded the trnL sequences of 48 plant speciescommonly seen at Poyang Lake from GenBank For theplant species not included in GenBank we sequencedtrnL by ourselves Total DNA was extracted using thehexadecyltrimethylammonium bromide (CTAB) proto-col We used primer pair c (5prime-3prime CGAAATCGGTAGACGCTACG) and h (5prime-3prime CCATTGAGTCTCTGCACCTATC) to amplify an approximately 150 bp regionof the chloroplast trnL intron [136] This primer pairwas chosen as it can amplify DNA from a wide range ofwetland plants [70] PCR amplifications were carried outin a total volume of 25 μl containing 125 μl PCR mix(Tiangen Beijing China) 1 μl DNA 1 μl each primerand 95 μl H2O The reaction conditions were as followsdenaturation at 94 degC for 4 min followed by 35 cycles at94 degC for 30 s 56 degC for 30 s and 72 degC for 45 s and afinal 10 min at 72 degC We used electrophoresis to checkthe PCR products ABI 3730xl analyzer to perform se-quencing and Seqman II to assemble the sequences
DNA extraction and high-throughput sequencing of fecalsamplesTo uncover plant component consumed by cranes amodified CTAB protocol [70] was used to extract theDNA from approximately 200 mg of the interior of eachfecal sample DNA extraction was conducted in a dedi-cated laboratory for analyses of DNA from samples withlow DNA quality For each batch of DNA extractionnegative controls were included to monitor for possiblecontamination The primers c and h with a 6 bp tagadded to 5prime ends of each primer were used to amplifytrnL gene The PCR amplification procedure was thesame as barcode database construction For each samplea unique combination of tags in the forward and reverseprimers was used so that sample-specific data can be
recovered after sequencing Three PCR replicates wereperformed for each sample to reduce amplification biasThe three replicates were pooled and the DNA concen-tration and purity were monitored on a 1 agarose gelOnly samples with a clear band were processed furtherSamples were purified using the SanPrep Column PCRProduct Purification Kit (Sangon Biotech ShanghaiChina) The purified PCR products were pooled takinginto account of DNA concentration to obtain an ap-proximately equal amount of amplicon DNA from eachfecal sample The pool of individually tagged ampliconswas used to prepare a sequencing library using the Tru-Seq DNA PCR-Free Sample Preparation Kit (IlluminaUSA) following manufacturerrsquos recommendations Thelibrary was sequenced on an Illumina HiSeq 2500 plat-form and 250 bp paired-end reads were generated Li-brary preparation and sequencing were conducted byNovogene (Beijing China)
Sequencing data analysisAfter high-throughput sequencing the paired-end readswere demultiplexed into sample-specific files based ontheir unique tags The tags and primer sequences weresubsequently trimmed The paired-end reads weremerged using FLASH 127 [137] and then filtered usingQIIME 170 [138] Sequences were dereplicated andclustered to OTUs using USEARCH 110667 [139] atthe similarity threshold of 97 All OTUs were assignedto taxonomic units by referring to the local plant data-base using blast 2231+ with thresholds of identity gt97 and e-value lt 10 eminus 50 If an OTU matched two ormore taxa it was assigned to a higher taxonomic levelthat included all taxaTo evaluate the sufficiency of sequencing depth and
sampling size individual- species- and sample-basedrarefaction curves were built using EstimateS 82[140] The curves were computed by randomly resam-pling sequences or samples and plotting these againstthe numbers of OTUs The read abundance data wereconverted to RRA of each food item using the follow-ing equation [79]
RRA frac14 1S
XS
Kfrac141
nikPTifrac141nik
100
where nik is the number of sequences of food item i insample k T is the total number of food items and S isthe sampling size
Dietary niche breadth and overlap among crane speciesTo explore whether the cranesrsquo diets showed substantialinterannual variation we calculated Pearson correlationsbetween diet compositions of each crane species in win-ter 2017 and winter 2018 The proportion of each food
Hou et al Frontiers in Zoology (2021) 1824 Page 12 of 17
item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

impacts of habitat degradation and to lead its classifica-tion as Critically Endangered [49] At Poyang Lake Si-berian Cranes traditionally fed on tubers of Vallisneriathe dominant submerged macrophytes in shallow watersand mudflats [89 90] In winter 2010 due to a flood-induced Vallisneria tuber collapse they were observedforaging for P limprichtii taproots and T edulis bulbs ingrasslands for the first time [38 44] In the winters from2015 to 2017 the diets of Siberian Cranes changedagain with thousands of cranes feeding on rice seedsand lotus rhizomes in agricultural habitats [61 62 64]Our study identified 22 plant items in the feces of Siber-
ian Cranes These plant items were obtained from shallowwaters grasslands and agricultural fields Four plant itemswith yearly RRA gt 10 were identified in the feces Vallis-neria tuber P criopolitanum rhizomes rice seeds andlotus rhizomes (the vegetative portions consumed bycranes were based on our field observation) SiberianCranesrsquo diet compositions showed substantial interannualvariation When the abundance of Vallisneria tubers waslow (ie winter 2017) Siberian Cranes mainly fed on riceseeds lotus rhizomes and P criopolitanum rhizomes inagricultural fields and grasslands When the abundance ofVallisneria tubers was high (ie winter 2018) they mainlyfed on Vallisneria tubers and P criopolitanum rhizomesin shallow waters and grasslands Our study in combin-ation with previous studies indicates that the diets of Si-berian Cranes have changed greatly since winter 2010 [3844 62] They have gradually broadened their dietaryniches which has probably been driven by the degradationof submerged plants [38 44 57]Similar to Siberian Cranes a diet shift was also ob-
served in White-naped Cranes Historically White-naped Cranes only fed on submerged plants in shallowwaters and mudflats of Poyang Lake [91] Then drivenby the decline of submerged plants they primarily fed atgrasslands and mudflats [87 91] consuming Vallisneriatuber T edulis P limprichtii and Ranunculus polii [5587] Our results indicate that in addition to T edulisand P limprichtii that had previously been reported theproportions of P criopolitanum and Carex were alsohigh in the feces Hooded Cranes were suggested tomainly feed on the grassland plant P limprichtii throughdirect observation [86] Our study indicates that thegrassland plants P criopolitanum and Carex also occu-pied high proportions in the feces The dominant fooditems in the feces of White-naped Cranes and HoodedCranes varied greatly between the two winters The vari-ation in dietary composition indicated that the diversityand composition of the grassland plant communitymight have changed greatly between the two wintersThe dietary niche overlap between White-naped Cranesand Hooded Cranes was the highest among the fourcrane species aligning with their level of habitat niche
overlap which was also the highest among the species[92] The high dietary and habitat niche overlaps suggesthigh competition potential between the two cranespeciesThe Eurasian Crane is a habitat generalist It is known
to feed in a variety of habitats including grasslandsmudflats shallow waters and agricultural fields [88 92]Its diet at Poyang Lake has not been documented previ-ously Our study indicates that P criopolitanum P lim-prichtii and rice were the dominant food items in thefeces of Eurasian Cranes The number of plant itemsconsumed dietary niche breadth and Shannon-Wienermeasures were the highest among the four crane speciessupporting its generalist foraging strategy In contrast tothe three crane species above Eurasian Cranesrsquo dietcompositions were similar between the two winters Riceand grassland plants occupied high proportions in thefeces in both winters
Important food resources for cranesVallisneria tubers have been acknowledged as importantfood sources for cranes especially Siberian Cranes andWhite-naped Cranes [55 56] Our study confirms theimportance of Vallisneria tubers in the diet of SiberianCranes It was the most common food source in thefeces of Siberian Cranes in winter 2018 when its abun-dance was high Vallisneria tuber proportions in thefeces of the three other crane species were not as high asthat in the feces of Siberian Cranes but it still occupied5ndash9 of the total foods in winter 2018To our surprise P criopolitanum was found to be the
first dominant food in the feces of White-naped CranesHooded Cranes and Eurasian Cranes and the seconddominant food in the feces of Siberian Cranes in winter2018 In winter 2017 the proportions of P criopolita-num were lt 17 in the feces of Siberian Cranes andWhite-naped Cranes and were lt 3 in the feces ofHooded Cranes and Eurasian Cranes The higher pro-portions of P criopolitanum in winter 2018 than winter2017 indicate that the abundance and availability of Pcriopolitanum was much higher in winter 2018 P crio-politanum is an annual grassland plant It is sensitive tohydrological conditions and can only survive in a rela-tively narrow range of water depths and percent time in-undated [93] Due to the sensitivity its distributionrange and abundance showed high interannual variation[94 95] Thus it is likely that the abundance of P crio-politanum was higher in winter 2018 than in winter2017The importance of P criopolitanum as a food source
for the cranes at Poyang Lake has not been documentedpreviously It has only been suggested to be a major foodsource of Hooded Cranes at Shengjin Lake another lakein the middle and lower Yangtze River floodplain [86
Hou et al Frontiers in Zoology (2021) 1824 Page 8 of 17
96] P criopolitanum is a dominant wetland plant at Po-yang Lake [93 97] The coverage percentage of this plantis the third highest among grassland plants accountingfor 12 of the total areas of grasslands [97] This plantspecies is usually distributed at elevations of 9ndash12mabove sea level near mudflats and shallow waters [97]which are frequently used by cranes Its rhizome with ashallow burial depth is high in energy and nutritionalcontent and low in crude fibre [38 86] All these charac-teristics make P criopolitanum rhizome a potentially im-portant food source for cranes We frequently observedcranes dig and feed on rhizomes of P criopolitanum andleave aboveground parts in the fields Therefore we be-lieve that P criopolitanum was used as food not inci-dentally consumed by cranes However we need to becaution that high proportion of P criopolitanum in thefeces does not mean high proportion in the diets Morework on the importance of P criopolitanum in the dietsof cranes is needed
The roles of agricultural fields in crane protectionWith the loss and degradation of natural wetlands theroles of artificial wetlands in waterbird protection havebeen under intensive debate [98 99] Although agricul-tural fields cannot completely replace the function ofnatural wetlands as waterbird habitats [100ndash102] agri-cultural fields might provide alternative or complemen-tary habitats for waterbirds in all life stages [38 103104] The proportions of domesticated species in thefeces of Siberian Cranes varied considerably betweenyears In winter 2017 when the abundance of Vallisneriatubers was low the proportion of Vallisneria in SiberianCranesrsquo feces was only 095 Siberian Cranes may nothave found enough foods in the shallow waters andmudflats and were forced to search for rice seeds andlotus rhizomes in agricultural fields In winter 2018when the abundance of Vallisneria tubers reboundedSiberian Cranes moved back to shallow waters and mud-flats and a large proportion of their traditional food Val-lisneria tubers and also P criopolitanum were detectedin the feces A similar abandonment of tuber foragingoccurred following the tuber collapse in 2010 and thenresumed in the next winter when tubers rebounded in2011 [38 44] The diet and habitat changes indicate thatagricultural habitats were important refuges for SiberianCranes providing important food resources for them tosurvive through winters with few Vallisneria tubersThey also indicate that Siberian Cranes prefer naturalwetlands to agricultural fields [38]Similar to Siberian Cranes White-naped Cranes and
Hooded Cranes consumed rice seeds and lotus rhizomesin winter 2017 but did not consume any in winter 2018The changes in the proportions suggest that the abun-dance of food resources in natural wetlands might have
been low in winter 2017 The deficient food resourcesmight have been Vallisneria tubers but might also havebeen grassland plants the primary food sources of thesetwo crane species Under natural food shortage condi-tions (ie winter 2017) White-naped Cranes andHooded Cranes utilized agricultural fields When theabundance of Vallisneria tubers and probably grasslandplants were high (ie winter 2018) the two crane speciesreturned to their natural habitats Therefore for White-naped Cranes and Hooded Cranes agricultural fieldswere important alternative foraging habitats used asbuffers against starvation during periods of natural foodshortages The return of the two crane species to naturalwetlands in winter 2018 further emphasizes the superiorquality of natural habitats over agricultural fields as for-aging habitats for cranes [38]The proportions of rice seeds in the feces of Eur-
asian Cranes were high in the two winters thoughthe proportion in winter 2017 was twice as high asthat in winter 2018 Therefore agricultural fields ap-peared to be consistently important as foraging habi-tats for Eurasian Cranes The frequent use ofagricultural fields by Eurasian Cranes has been docu-mented widely [49 105 106]Domesticated species generally contain high energy
and nutritional content and are of superior quality com-pared to natural plants [86 104] Farmland foraging hasbeen suggested to contribute to increase in the abun-dance of some goose [107] egret [108] and wader [109]populations in Japan Korea Europe and North Amer-ica However cranes generally avoid human disturbancewhen select foraging sites [110 111] Human disturb-ance in agricultural fields around Poyang Lake is consid-erably high due to intensive agricultural activity [112]High human disturbance was suggested to be a majorreason that wintering geese were almost confined withinnatural wetlands and hesitated to exploit the riches ofthe modern agricultural habitats at Poyang Lake [112113] High human disturbance also resulted in SiberianCranes spending twice as much time alerting in agricul-tural fields than they did in natural habitats [114] Thehigh level of human disturbance in agricultural fieldsmight contribute to the preference for natural wetlandsby Siberian Cranes White-naped Cranes and HoodedCranes at Poyang Lake Moreover although agriculturefields contain energy-rich crops wetlands contain di-verse food resources especially protein-rich inverte-brates which might better satisfy dietary needs of cranes[110 115] Taken together security and diverse food re-sources might have attracted cranes to select wetlands asforaging sitesSubmerged macrophytes at Poyang Lake have de-
graded in recent decades [57ndash60] and Vallisneria tuberdisappearance is expected to occur more frequently
Hou et al Frontiers in Zoology (2021) 1824 Page 9 of 17
under future climate change [57 60] Cranes may in-creasingly depend upon agricultural fields around Po-yang Lake which may in turn cause significant cropdamage and economic losses This is especially true onagricultural fields surrounding protected wetlands wherewaterbirds concentrate in high numbers [116 117] AtPoyang Lake cranes mainly fed on post-harvest remainsof rice seeds and lotus rhizomes thus crop damage andeconomic loss were limited However we observed somefarmers scared cranes away from their agricultural fieldsThere were more than 14 million free-ranging poultryraised at agricultural fields around Poyang Lake [118]where they gleaned waste grain residues in stubble fieldsCrane feeding would reduce the food abundance of thefree-ranging domestic poultry thus farmers drovecranes away To reduce human disturbance environ-mental education is needed because farmers are morelikely to protect cranes if they understand the endan-gered degree and the ecological significance of cranes
Effects of tuber collapse on dietary niche breadth andoverlap among crane speciesVallisneria tuber is an important food source for Siber-ian Cranes (89 90 this study) thus variation in itsabundance might influence Siberian Cranesrsquo dietaryniche breadth The numbers of food items dietary nichebreadth and Shannon-Wiener measures of SiberianCranes were similar between winter 2017 and winter2018 This is inconsistent with the contention that dur-ing seasons with a low abundance of dominant food thedietary niche may broaden as consumers relying on in-sufficient preferred food items are forced to add lessprofitable resources to their diets [26 119 120] In win-ter 2017 with low Vallisneria tuber abundance SiberianCranes left shallow waters and mudflats and moved torice paddies and lotus ponds [61 62 64] Their foodschanged from Vallisneria tuber to rice seeds and lotusrhizomes almost completely As the energy protein andfat contents of crops are generally as good as or superiorto natural foods and are often present in agriculturalfields in far greater abundance [86 104] Siberian Cranesmay gain enough energy through feeding on crops anddo not need to add many other food sources Moreoverthe Siberian Crane is a dietary specialist [89 90] PoyangLake has a relatively simple plant community compos-ition with Vallisneria and Hydrilla verticillate being thedominant submerged macrophytes [59] and Carex beingthe dominant grassland plants [121] Therefore theremay be few options for Siberian Cranes which mightlead to the failure of this species to widen its dietaryniche breadthIn contrast to Siberian Cranes the numbers of food
items dietary niche breadths and Shannon-Wiener mea-sures of White-naped Cranes and Hooded Cranes were
higher in winter 2017 than in winter 2018 P criopolita-num was the predominant food item in the feces of thetwo crane species in winter 2018 and the proportions ofVallisneria tubers were also high The abundances ofVallisneria tubers and probably P criopolitanum at Po-yang Lake declined greatly in winter 2017 When thereis a short-fall in food availability animals may optimizeforaging behavior and increase their dietary breath [122123] The short-fall in the abundances of P criopolita-num and Vallisneria tubers in winter 2017 might havedriven the two crane species to add less profitable grass-land plants in their diets Although the two cranes alsoconsumed high-quality rice seeds and lotus rhizomes inwinter 2017 their proportions were low in the feces andthese foods might be unable to meet the energy require-ments of the two crane species The niche expansion ofthe two crane species during periods of preferred foodshortage was in agreement with the basic prediction ofoptimal foraging theory [22 23] It is also consistentwith a commonly observed fact that energy-maximizingmodels work better when animals food-limited [124]The dietary niche overlaps among the crane species
were low in winter 2017 with the four crane species di-verging in their principal food items However in winter2018 the niche overlaps were significantly higher thanexpected by chance They had similar diet compositionswith P criopolitanum rhizomes and Vallisneria tubersbeing the principal food items in the feces The in-creased overlap might have been caused by the highabundances of the two plant species in winter 2018which allowed the four crane species to share the samefood items with low interspecific competition Our studyis consistent with previous research that suggested thatamong-species dietary niches tended to overlap moreduring seasons of high food abundance [32 33 125126] Greater resource availability could promote an in-crease in species utilization of similar resources and re-duce competitive interactions between species [32 34126] By contrast during seasons with low food abun-dance species may partition limited food resources todecrease interspecific competition leading to a decreasein niche overlap [126 127] It has been suggested thatdiet partitioning facilitates the coexistence of manyclosely related species [7 128] The interannual variationin niche overlap observed in our study suggests that dietpartitioning may be restricted to specific temporal casesIn seasons or years with abundant food sources inter-specific competition may be relaxed and niche overlapcould be high
ConclusionsIn summary by using metabarcoding technology ourstudy gained new insights into cranesrsquo diets that are es-sential for informing habitat management to improve
Hou et al Frontiers in Zoology (2021) 1824 Page 10 of 17
the availability of foraging resources Vallisneria tuberswas confirmed as an important food source of cranesSurprisingly P criopolitanum also occupied high pro-portions in the feces of cranes and may also be import-ant Our study emphasizes the superior quality ofnatural wetlands compared to agricultural fields as for-aging habitats for cranes The recent degradation of sub-merged macrophytes at Poyang Lake potentiallythreatens the survival of cranes especially SiberianCranes Therefore it is urgent to reverse the broad de-cline of submerged macrophytes and provide importantforaging habitats for cranes during years of tuber col-lapses The abundances of Vallisneria tubers and prob-ably P criopolitanum at Poyang Lake greatly influencedthe dietary niche width and overlaps among crane spe-cies Cranes exhibited low dietary niche overlap duringperiods of preferred food shortage but significantly highoverlap during periods when preferred foods were plen-tiful While high niche overlap is often interpreted as in-tense interspecific competition our study highlights theimportance of taking food abundance into account
Materials and methodsStudy areasPoyang Lake the largest freshwater lake in China is oneof the most important wintering areas for waterbirdsalong the East Asian-Australasian Flyway (Fig 1) [51129] It is characterized by dramatic seasonal hydro-logical fluctuations with an average change of 924 mbetween the summer high and winter low water levels[130] During summer the inundation area is gt 2500km2 [131] In autumn and winter the water level dropscausing the inundation area to shrink to lt 1000 km2
[131] Poyang Lake contains 102 sublakes which are in-undated and integrated with the main body of PoyangLake during summer and are isolated during winter[132] As the water level drops the sublakes appear andlarge areas of grasslands mudflats and shallow waterareas are exposed providing winter foraging and roost-ing areas for approximately 420000 waterbirds from ap-proximately 111 species [133 134]
Vallisneria tuber surveyThe density and biomass of Vallisneria tubers were sur-veyed at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake in late October and earlyNovember of 2017 and 2018 before large numbers ofwintering waterbirds arrived We set 76 183 and 36sampling plots uniformly located at Dahuchi Shahu andChanghuchi respectively Vallisneria tubers were col-lected from two quadrats in each plot with a locallymade steel grab sampler The sampler has two long
handles At the end of the handles there are two scoopsfacing each other The sampler was inserted into thesubstrate and the scoops brought together to collectabout 15 cm long times 13 cm wide times 30 cm high sample ofsubstrate Tubers were subsequently cleaned andcounted The dry weight of tubers in each quadrat wasweighed after oven-drying at 80 degC until constant weightThe tuber density and biomass in each plot was calcu-lated by dividing total tuber number and tuber massacross two quadrats by quadrat area respectively Weused t-test to explore whether the tuber density and bio-mass in the winters of 2017 and 2018 significantly devi-ated from the historical average values in the wintersfrom 1999 to 2016 [57] The historical tuber density andbiomass data were collected by the Poyang Lake Na-tional Nature Reserve at three sublakes (Dahuchi Shahuand Meixihu) [57] R 360 was used to do the t-testanalyses
Fecal sample collectionFecal samples were collected at Poyang Lake in the win-ters of 2017 and 2018 We first investigated the distribu-tions of cranes When we saw foraging cranes we waiteduntil they left to avoid disturbing them Soon after theyleft we went to the foraging sites to collect fresh feces(ie feces with wet surface) with sterilized tweezers Tominimize the probability of multiple samples from thesame individual all collected samples were separated byat least 5 m We stored the samples in liquid nitrogen inthe field and transferred them to a minus 80 degC refrigeratorfor long-term storage in the laboratory To ensure thatthe sources of the feces were known we tried to collectsamples from monospecific flocks For samples collectedfrom mixed-species flocks we used DNA analysis toidentify the species that deposited the fecal samplesPlants were also collected in the field to construct aDNA barcoding reference database
Species identificationBecause of the similar morphologies of the feces of thefour crane species we used DNA analysis to identify thespecies that deposited the fecal samples collected frommixed-species flocks For the genetic species assignmentof each fecal sample we scraped the external surfacewhich contain DNA from the host species as a result ofthe sloughing of cells from the digestive tract DNA wasextracted with the QIAamp Power Fecal DNA Kit (Qia-gen USA) according to the manufacturerrsquos guidelinesFor each round of DNA extraction negative controls(ie extraction without feces) were included to monitorfor possible contamination We amplified the mitochon-drial DNA D-loop fragment with the primer pair DL02F
Hou et al Frontiers in Zoology (2021) 1824 Page 11 of 17
(5prime-3prime GCGGCCCGAAAAGCCGCTG) and DL02R (5prime-3prime GCCCTGACATAGGAACCAGAGGCGC) [135]PCR amplifications were carried out in a total volume of25 μl containing 125 μl PCR mix (Tiangen BeijingChina) 1 μl DNA 1 μl each primer and 95 μl H2O Thereaction conditions were as follows denaturation at94 degC for 4 min followed by 34 cycles at 94 degC for 48 s56 degC for 48 s and 72 degC for 1 min and a final 10 min at72 degC We checked for the presence of a PCR product ofsuitable length by electrophoresis on a 1 agarose gelSequencing was carried out on an ABI 3730xl analyzerby Sangon Biotech (Shanghai China) The complete se-quences were assembled using Seqman II (DNASTAR)and compared visually to the original chromatograms toavoid reading errors The taxonomic assignment wasconducted by searching against the nucleotide databaseof GenBank using Web BLAST
Construction of a barcode reference databaseWe downloaded the trnL sequences of 48 plant speciescommonly seen at Poyang Lake from GenBank For theplant species not included in GenBank we sequencedtrnL by ourselves Total DNA was extracted using thehexadecyltrimethylammonium bromide (CTAB) proto-col We used primer pair c (5prime-3prime CGAAATCGGTAGACGCTACG) and h (5prime-3prime CCATTGAGTCTCTGCACCTATC) to amplify an approximately 150 bp regionof the chloroplast trnL intron [136] This primer pairwas chosen as it can amplify DNA from a wide range ofwetland plants [70] PCR amplifications were carried outin a total volume of 25 μl containing 125 μl PCR mix(Tiangen Beijing China) 1 μl DNA 1 μl each primerand 95 μl H2O The reaction conditions were as followsdenaturation at 94 degC for 4 min followed by 35 cycles at94 degC for 30 s 56 degC for 30 s and 72 degC for 45 s and afinal 10 min at 72 degC We used electrophoresis to checkthe PCR products ABI 3730xl analyzer to perform se-quencing and Seqman II to assemble the sequences
DNA extraction and high-throughput sequencing of fecalsamplesTo uncover plant component consumed by cranes amodified CTAB protocol [70] was used to extract theDNA from approximately 200 mg of the interior of eachfecal sample DNA extraction was conducted in a dedi-cated laboratory for analyses of DNA from samples withlow DNA quality For each batch of DNA extractionnegative controls were included to monitor for possiblecontamination The primers c and h with a 6 bp tagadded to 5prime ends of each primer were used to amplifytrnL gene The PCR amplification procedure was thesame as barcode database construction For each samplea unique combination of tags in the forward and reverseprimers was used so that sample-specific data can be
recovered after sequencing Three PCR replicates wereperformed for each sample to reduce amplification biasThe three replicates were pooled and the DNA concen-tration and purity were monitored on a 1 agarose gelOnly samples with a clear band were processed furtherSamples were purified using the SanPrep Column PCRProduct Purification Kit (Sangon Biotech ShanghaiChina) The purified PCR products were pooled takinginto account of DNA concentration to obtain an ap-proximately equal amount of amplicon DNA from eachfecal sample The pool of individually tagged ampliconswas used to prepare a sequencing library using the Tru-Seq DNA PCR-Free Sample Preparation Kit (IlluminaUSA) following manufacturerrsquos recommendations Thelibrary was sequenced on an Illumina HiSeq 2500 plat-form and 250 bp paired-end reads were generated Li-brary preparation and sequencing were conducted byNovogene (Beijing China)
Sequencing data analysisAfter high-throughput sequencing the paired-end readswere demultiplexed into sample-specific files based ontheir unique tags The tags and primer sequences weresubsequently trimmed The paired-end reads weremerged using FLASH 127 [137] and then filtered usingQIIME 170 [138] Sequences were dereplicated andclustered to OTUs using USEARCH 110667 [139] atthe similarity threshold of 97 All OTUs were assignedto taxonomic units by referring to the local plant data-base using blast 2231+ with thresholds of identity gt97 and e-value lt 10 eminus 50 If an OTU matched two ormore taxa it was assigned to a higher taxonomic levelthat included all taxaTo evaluate the sufficiency of sequencing depth and
sampling size individual- species- and sample-basedrarefaction curves were built using EstimateS 82[140] The curves were computed by randomly resam-pling sequences or samples and plotting these againstthe numbers of OTUs The read abundance data wereconverted to RRA of each food item using the follow-ing equation [79]
RRA frac14 1S
XS
Kfrac141
nikPTifrac141nik
100
where nik is the number of sequences of food item i insample k T is the total number of food items and S isthe sampling size
Dietary niche breadth and overlap among crane speciesTo explore whether the cranesrsquo diets showed substantialinterannual variation we calculated Pearson correlationsbetween diet compositions of each crane species in win-ter 2017 and winter 2018 The proportion of each food
Hou et al Frontiers in Zoology (2021) 1824 Page 12 of 17
item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

96] P criopolitanum is a dominant wetland plant at Po-yang Lake [93 97] The coverage percentage of this plantis the third highest among grassland plants accountingfor 12 of the total areas of grasslands [97] This plantspecies is usually distributed at elevations of 9ndash12mabove sea level near mudflats and shallow waters [97]which are frequently used by cranes Its rhizome with ashallow burial depth is high in energy and nutritionalcontent and low in crude fibre [38 86] All these charac-teristics make P criopolitanum rhizome a potentially im-portant food source for cranes We frequently observedcranes dig and feed on rhizomes of P criopolitanum andleave aboveground parts in the fields Therefore we be-lieve that P criopolitanum was used as food not inci-dentally consumed by cranes However we need to becaution that high proportion of P criopolitanum in thefeces does not mean high proportion in the diets Morework on the importance of P criopolitanum in the dietsof cranes is needed
The roles of agricultural fields in crane protectionWith the loss and degradation of natural wetlands theroles of artificial wetlands in waterbird protection havebeen under intensive debate [98 99] Although agricul-tural fields cannot completely replace the function ofnatural wetlands as waterbird habitats [100ndash102] agri-cultural fields might provide alternative or complemen-tary habitats for waterbirds in all life stages [38 103104] The proportions of domesticated species in thefeces of Siberian Cranes varied considerably betweenyears In winter 2017 when the abundance of Vallisneriatubers was low the proportion of Vallisneria in SiberianCranesrsquo feces was only 095 Siberian Cranes may nothave found enough foods in the shallow waters andmudflats and were forced to search for rice seeds andlotus rhizomes in agricultural fields In winter 2018when the abundance of Vallisneria tubers reboundedSiberian Cranes moved back to shallow waters and mud-flats and a large proportion of their traditional food Val-lisneria tubers and also P criopolitanum were detectedin the feces A similar abandonment of tuber foragingoccurred following the tuber collapse in 2010 and thenresumed in the next winter when tubers rebounded in2011 [38 44] The diet and habitat changes indicate thatagricultural habitats were important refuges for SiberianCranes providing important food resources for them tosurvive through winters with few Vallisneria tubersThey also indicate that Siberian Cranes prefer naturalwetlands to agricultural fields [38]Similar to Siberian Cranes White-naped Cranes and
Hooded Cranes consumed rice seeds and lotus rhizomesin winter 2017 but did not consume any in winter 2018The changes in the proportions suggest that the abun-dance of food resources in natural wetlands might have
been low in winter 2017 The deficient food resourcesmight have been Vallisneria tubers but might also havebeen grassland plants the primary food sources of thesetwo crane species Under natural food shortage condi-tions (ie winter 2017) White-naped Cranes andHooded Cranes utilized agricultural fields When theabundance of Vallisneria tubers and probably grasslandplants were high (ie winter 2018) the two crane speciesreturned to their natural habitats Therefore for White-naped Cranes and Hooded Cranes agricultural fieldswere important alternative foraging habitats used asbuffers against starvation during periods of natural foodshortages The return of the two crane species to naturalwetlands in winter 2018 further emphasizes the superiorquality of natural habitats over agricultural fields as for-aging habitats for cranes [38]The proportions of rice seeds in the feces of Eur-
asian Cranes were high in the two winters thoughthe proportion in winter 2017 was twice as high asthat in winter 2018 Therefore agricultural fields ap-peared to be consistently important as foraging habi-tats for Eurasian Cranes The frequent use ofagricultural fields by Eurasian Cranes has been docu-mented widely [49 105 106]Domesticated species generally contain high energy
and nutritional content and are of superior quality com-pared to natural plants [86 104] Farmland foraging hasbeen suggested to contribute to increase in the abun-dance of some goose [107] egret [108] and wader [109]populations in Japan Korea Europe and North Amer-ica However cranes generally avoid human disturbancewhen select foraging sites [110 111] Human disturb-ance in agricultural fields around Poyang Lake is consid-erably high due to intensive agricultural activity [112]High human disturbance was suggested to be a majorreason that wintering geese were almost confined withinnatural wetlands and hesitated to exploit the riches ofthe modern agricultural habitats at Poyang Lake [112113] High human disturbance also resulted in SiberianCranes spending twice as much time alerting in agricul-tural fields than they did in natural habitats [114] Thehigh level of human disturbance in agricultural fieldsmight contribute to the preference for natural wetlandsby Siberian Cranes White-naped Cranes and HoodedCranes at Poyang Lake Moreover although agriculturefields contain energy-rich crops wetlands contain di-verse food resources especially protein-rich inverte-brates which might better satisfy dietary needs of cranes[110 115] Taken together security and diverse food re-sources might have attracted cranes to select wetlands asforaging sitesSubmerged macrophytes at Poyang Lake have de-
graded in recent decades [57ndash60] and Vallisneria tuberdisappearance is expected to occur more frequently
Hou et al Frontiers in Zoology (2021) 1824 Page 9 of 17
under future climate change [57 60] Cranes may in-creasingly depend upon agricultural fields around Po-yang Lake which may in turn cause significant cropdamage and economic losses This is especially true onagricultural fields surrounding protected wetlands wherewaterbirds concentrate in high numbers [116 117] AtPoyang Lake cranes mainly fed on post-harvest remainsof rice seeds and lotus rhizomes thus crop damage andeconomic loss were limited However we observed somefarmers scared cranes away from their agricultural fieldsThere were more than 14 million free-ranging poultryraised at agricultural fields around Poyang Lake [118]where they gleaned waste grain residues in stubble fieldsCrane feeding would reduce the food abundance of thefree-ranging domestic poultry thus farmers drovecranes away To reduce human disturbance environ-mental education is needed because farmers are morelikely to protect cranes if they understand the endan-gered degree and the ecological significance of cranes
Effects of tuber collapse on dietary niche breadth andoverlap among crane speciesVallisneria tuber is an important food source for Siber-ian Cranes (89 90 this study) thus variation in itsabundance might influence Siberian Cranesrsquo dietaryniche breadth The numbers of food items dietary nichebreadth and Shannon-Wiener measures of SiberianCranes were similar between winter 2017 and winter2018 This is inconsistent with the contention that dur-ing seasons with a low abundance of dominant food thedietary niche may broaden as consumers relying on in-sufficient preferred food items are forced to add lessprofitable resources to their diets [26 119 120] In win-ter 2017 with low Vallisneria tuber abundance SiberianCranes left shallow waters and mudflats and moved torice paddies and lotus ponds [61 62 64] Their foodschanged from Vallisneria tuber to rice seeds and lotusrhizomes almost completely As the energy protein andfat contents of crops are generally as good as or superiorto natural foods and are often present in agriculturalfields in far greater abundance [86 104] Siberian Cranesmay gain enough energy through feeding on crops anddo not need to add many other food sources Moreoverthe Siberian Crane is a dietary specialist [89 90] PoyangLake has a relatively simple plant community compos-ition with Vallisneria and Hydrilla verticillate being thedominant submerged macrophytes [59] and Carex beingthe dominant grassland plants [121] Therefore theremay be few options for Siberian Cranes which mightlead to the failure of this species to widen its dietaryniche breadthIn contrast to Siberian Cranes the numbers of food
items dietary niche breadths and Shannon-Wiener mea-sures of White-naped Cranes and Hooded Cranes were
higher in winter 2017 than in winter 2018 P criopolita-num was the predominant food item in the feces of thetwo crane species in winter 2018 and the proportions ofVallisneria tubers were also high The abundances ofVallisneria tubers and probably P criopolitanum at Po-yang Lake declined greatly in winter 2017 When thereis a short-fall in food availability animals may optimizeforaging behavior and increase their dietary breath [122123] The short-fall in the abundances of P criopolita-num and Vallisneria tubers in winter 2017 might havedriven the two crane species to add less profitable grass-land plants in their diets Although the two cranes alsoconsumed high-quality rice seeds and lotus rhizomes inwinter 2017 their proportions were low in the feces andthese foods might be unable to meet the energy require-ments of the two crane species The niche expansion ofthe two crane species during periods of preferred foodshortage was in agreement with the basic prediction ofoptimal foraging theory [22 23] It is also consistentwith a commonly observed fact that energy-maximizingmodels work better when animals food-limited [124]The dietary niche overlaps among the crane species
were low in winter 2017 with the four crane species di-verging in their principal food items However in winter2018 the niche overlaps were significantly higher thanexpected by chance They had similar diet compositionswith P criopolitanum rhizomes and Vallisneria tubersbeing the principal food items in the feces The in-creased overlap might have been caused by the highabundances of the two plant species in winter 2018which allowed the four crane species to share the samefood items with low interspecific competition Our studyis consistent with previous research that suggested thatamong-species dietary niches tended to overlap moreduring seasons of high food abundance [32 33 125126] Greater resource availability could promote an in-crease in species utilization of similar resources and re-duce competitive interactions between species [32 34126] By contrast during seasons with low food abun-dance species may partition limited food resources todecrease interspecific competition leading to a decreasein niche overlap [126 127] It has been suggested thatdiet partitioning facilitates the coexistence of manyclosely related species [7 128] The interannual variationin niche overlap observed in our study suggests that dietpartitioning may be restricted to specific temporal casesIn seasons or years with abundant food sources inter-specific competition may be relaxed and niche overlapcould be high
ConclusionsIn summary by using metabarcoding technology ourstudy gained new insights into cranesrsquo diets that are es-sential for informing habitat management to improve
Hou et al Frontiers in Zoology (2021) 1824 Page 10 of 17
the availability of foraging resources Vallisneria tuberswas confirmed as an important food source of cranesSurprisingly P criopolitanum also occupied high pro-portions in the feces of cranes and may also be import-ant Our study emphasizes the superior quality ofnatural wetlands compared to agricultural fields as for-aging habitats for cranes The recent degradation of sub-merged macrophytes at Poyang Lake potentiallythreatens the survival of cranes especially SiberianCranes Therefore it is urgent to reverse the broad de-cline of submerged macrophytes and provide importantforaging habitats for cranes during years of tuber col-lapses The abundances of Vallisneria tubers and prob-ably P criopolitanum at Poyang Lake greatly influencedthe dietary niche width and overlaps among crane spe-cies Cranes exhibited low dietary niche overlap duringperiods of preferred food shortage but significantly highoverlap during periods when preferred foods were plen-tiful While high niche overlap is often interpreted as in-tense interspecific competition our study highlights theimportance of taking food abundance into account
Materials and methodsStudy areasPoyang Lake the largest freshwater lake in China is oneof the most important wintering areas for waterbirdsalong the East Asian-Australasian Flyway (Fig 1) [51129] It is characterized by dramatic seasonal hydro-logical fluctuations with an average change of 924 mbetween the summer high and winter low water levels[130] During summer the inundation area is gt 2500km2 [131] In autumn and winter the water level dropscausing the inundation area to shrink to lt 1000 km2
[131] Poyang Lake contains 102 sublakes which are in-undated and integrated with the main body of PoyangLake during summer and are isolated during winter[132] As the water level drops the sublakes appear andlarge areas of grasslands mudflats and shallow waterareas are exposed providing winter foraging and roost-ing areas for approximately 420000 waterbirds from ap-proximately 111 species [133 134]
Vallisneria tuber surveyThe density and biomass of Vallisneria tubers were sur-veyed at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake in late October and earlyNovember of 2017 and 2018 before large numbers ofwintering waterbirds arrived We set 76 183 and 36sampling plots uniformly located at Dahuchi Shahu andChanghuchi respectively Vallisneria tubers were col-lected from two quadrats in each plot with a locallymade steel grab sampler The sampler has two long
handles At the end of the handles there are two scoopsfacing each other The sampler was inserted into thesubstrate and the scoops brought together to collectabout 15 cm long times 13 cm wide times 30 cm high sample ofsubstrate Tubers were subsequently cleaned andcounted The dry weight of tubers in each quadrat wasweighed after oven-drying at 80 degC until constant weightThe tuber density and biomass in each plot was calcu-lated by dividing total tuber number and tuber massacross two quadrats by quadrat area respectively Weused t-test to explore whether the tuber density and bio-mass in the winters of 2017 and 2018 significantly devi-ated from the historical average values in the wintersfrom 1999 to 2016 [57] The historical tuber density andbiomass data were collected by the Poyang Lake Na-tional Nature Reserve at three sublakes (Dahuchi Shahuand Meixihu) [57] R 360 was used to do the t-testanalyses
Fecal sample collectionFecal samples were collected at Poyang Lake in the win-ters of 2017 and 2018 We first investigated the distribu-tions of cranes When we saw foraging cranes we waiteduntil they left to avoid disturbing them Soon after theyleft we went to the foraging sites to collect fresh feces(ie feces with wet surface) with sterilized tweezers Tominimize the probability of multiple samples from thesame individual all collected samples were separated byat least 5 m We stored the samples in liquid nitrogen inthe field and transferred them to a minus 80 degC refrigeratorfor long-term storage in the laboratory To ensure thatthe sources of the feces were known we tried to collectsamples from monospecific flocks For samples collectedfrom mixed-species flocks we used DNA analysis toidentify the species that deposited the fecal samplesPlants were also collected in the field to construct aDNA barcoding reference database
Species identificationBecause of the similar morphologies of the feces of thefour crane species we used DNA analysis to identify thespecies that deposited the fecal samples collected frommixed-species flocks For the genetic species assignmentof each fecal sample we scraped the external surfacewhich contain DNA from the host species as a result ofthe sloughing of cells from the digestive tract DNA wasextracted with the QIAamp Power Fecal DNA Kit (Qia-gen USA) according to the manufacturerrsquos guidelinesFor each round of DNA extraction negative controls(ie extraction without feces) were included to monitorfor possible contamination We amplified the mitochon-drial DNA D-loop fragment with the primer pair DL02F
Hou et al Frontiers in Zoology (2021) 1824 Page 11 of 17
(5prime-3prime GCGGCCCGAAAAGCCGCTG) and DL02R (5prime-3prime GCCCTGACATAGGAACCAGAGGCGC) [135]PCR amplifications were carried out in a total volume of25 μl containing 125 μl PCR mix (Tiangen BeijingChina) 1 μl DNA 1 μl each primer and 95 μl H2O Thereaction conditions were as follows denaturation at94 degC for 4 min followed by 34 cycles at 94 degC for 48 s56 degC for 48 s and 72 degC for 1 min and a final 10 min at72 degC We checked for the presence of a PCR product ofsuitable length by electrophoresis on a 1 agarose gelSequencing was carried out on an ABI 3730xl analyzerby Sangon Biotech (Shanghai China) The complete se-quences were assembled using Seqman II (DNASTAR)and compared visually to the original chromatograms toavoid reading errors The taxonomic assignment wasconducted by searching against the nucleotide databaseof GenBank using Web BLAST
Construction of a barcode reference databaseWe downloaded the trnL sequences of 48 plant speciescommonly seen at Poyang Lake from GenBank For theplant species not included in GenBank we sequencedtrnL by ourselves Total DNA was extracted using thehexadecyltrimethylammonium bromide (CTAB) proto-col We used primer pair c (5prime-3prime CGAAATCGGTAGACGCTACG) and h (5prime-3prime CCATTGAGTCTCTGCACCTATC) to amplify an approximately 150 bp regionof the chloroplast trnL intron [136] This primer pairwas chosen as it can amplify DNA from a wide range ofwetland plants [70] PCR amplifications were carried outin a total volume of 25 μl containing 125 μl PCR mix(Tiangen Beijing China) 1 μl DNA 1 μl each primerand 95 μl H2O The reaction conditions were as followsdenaturation at 94 degC for 4 min followed by 35 cycles at94 degC for 30 s 56 degC for 30 s and 72 degC for 45 s and afinal 10 min at 72 degC We used electrophoresis to checkthe PCR products ABI 3730xl analyzer to perform se-quencing and Seqman II to assemble the sequences
DNA extraction and high-throughput sequencing of fecalsamplesTo uncover plant component consumed by cranes amodified CTAB protocol [70] was used to extract theDNA from approximately 200 mg of the interior of eachfecal sample DNA extraction was conducted in a dedi-cated laboratory for analyses of DNA from samples withlow DNA quality For each batch of DNA extractionnegative controls were included to monitor for possiblecontamination The primers c and h with a 6 bp tagadded to 5prime ends of each primer were used to amplifytrnL gene The PCR amplification procedure was thesame as barcode database construction For each samplea unique combination of tags in the forward and reverseprimers was used so that sample-specific data can be
recovered after sequencing Three PCR replicates wereperformed for each sample to reduce amplification biasThe three replicates were pooled and the DNA concen-tration and purity were monitored on a 1 agarose gelOnly samples with a clear band were processed furtherSamples were purified using the SanPrep Column PCRProduct Purification Kit (Sangon Biotech ShanghaiChina) The purified PCR products were pooled takinginto account of DNA concentration to obtain an ap-proximately equal amount of amplicon DNA from eachfecal sample The pool of individually tagged ampliconswas used to prepare a sequencing library using the Tru-Seq DNA PCR-Free Sample Preparation Kit (IlluminaUSA) following manufacturerrsquos recommendations Thelibrary was sequenced on an Illumina HiSeq 2500 plat-form and 250 bp paired-end reads were generated Li-brary preparation and sequencing were conducted byNovogene (Beijing China)
Sequencing data analysisAfter high-throughput sequencing the paired-end readswere demultiplexed into sample-specific files based ontheir unique tags The tags and primer sequences weresubsequently trimmed The paired-end reads weremerged using FLASH 127 [137] and then filtered usingQIIME 170 [138] Sequences were dereplicated andclustered to OTUs using USEARCH 110667 [139] atthe similarity threshold of 97 All OTUs were assignedto taxonomic units by referring to the local plant data-base using blast 2231+ with thresholds of identity gt97 and e-value lt 10 eminus 50 If an OTU matched two ormore taxa it was assigned to a higher taxonomic levelthat included all taxaTo evaluate the sufficiency of sequencing depth and
sampling size individual- species- and sample-basedrarefaction curves were built using EstimateS 82[140] The curves were computed by randomly resam-pling sequences or samples and plotting these againstthe numbers of OTUs The read abundance data wereconverted to RRA of each food item using the follow-ing equation [79]
RRA frac14 1S
XS
Kfrac141
nikPTifrac141nik
100
where nik is the number of sequences of food item i insample k T is the total number of food items and S isthe sampling size
Dietary niche breadth and overlap among crane speciesTo explore whether the cranesrsquo diets showed substantialinterannual variation we calculated Pearson correlationsbetween diet compositions of each crane species in win-ter 2017 and winter 2018 The proportion of each food
Hou et al Frontiers in Zoology (2021) 1824 Page 12 of 17
item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

under future climate change [57 60] Cranes may in-creasingly depend upon agricultural fields around Po-yang Lake which may in turn cause significant cropdamage and economic losses This is especially true onagricultural fields surrounding protected wetlands wherewaterbirds concentrate in high numbers [116 117] AtPoyang Lake cranes mainly fed on post-harvest remainsof rice seeds and lotus rhizomes thus crop damage andeconomic loss were limited However we observed somefarmers scared cranes away from their agricultural fieldsThere were more than 14 million free-ranging poultryraised at agricultural fields around Poyang Lake [118]where they gleaned waste grain residues in stubble fieldsCrane feeding would reduce the food abundance of thefree-ranging domestic poultry thus farmers drovecranes away To reduce human disturbance environ-mental education is needed because farmers are morelikely to protect cranes if they understand the endan-gered degree and the ecological significance of cranes
Effects of tuber collapse on dietary niche breadth andoverlap among crane speciesVallisneria tuber is an important food source for Siber-ian Cranes (89 90 this study) thus variation in itsabundance might influence Siberian Cranesrsquo dietaryniche breadth The numbers of food items dietary nichebreadth and Shannon-Wiener measures of SiberianCranes were similar between winter 2017 and winter2018 This is inconsistent with the contention that dur-ing seasons with a low abundance of dominant food thedietary niche may broaden as consumers relying on in-sufficient preferred food items are forced to add lessprofitable resources to their diets [26 119 120] In win-ter 2017 with low Vallisneria tuber abundance SiberianCranes left shallow waters and mudflats and moved torice paddies and lotus ponds [61 62 64] Their foodschanged from Vallisneria tuber to rice seeds and lotusrhizomes almost completely As the energy protein andfat contents of crops are generally as good as or superiorto natural foods and are often present in agriculturalfields in far greater abundance [86 104] Siberian Cranesmay gain enough energy through feeding on crops anddo not need to add many other food sources Moreoverthe Siberian Crane is a dietary specialist [89 90] PoyangLake has a relatively simple plant community compos-ition with Vallisneria and Hydrilla verticillate being thedominant submerged macrophytes [59] and Carex beingthe dominant grassland plants [121] Therefore theremay be few options for Siberian Cranes which mightlead to the failure of this species to widen its dietaryniche breadthIn contrast to Siberian Cranes the numbers of food
items dietary niche breadths and Shannon-Wiener mea-sures of White-naped Cranes and Hooded Cranes were
higher in winter 2017 than in winter 2018 P criopolita-num was the predominant food item in the feces of thetwo crane species in winter 2018 and the proportions ofVallisneria tubers were also high The abundances ofVallisneria tubers and probably P criopolitanum at Po-yang Lake declined greatly in winter 2017 When thereis a short-fall in food availability animals may optimizeforaging behavior and increase their dietary breath [122123] The short-fall in the abundances of P criopolita-num and Vallisneria tubers in winter 2017 might havedriven the two crane species to add less profitable grass-land plants in their diets Although the two cranes alsoconsumed high-quality rice seeds and lotus rhizomes inwinter 2017 their proportions were low in the feces andthese foods might be unable to meet the energy require-ments of the two crane species The niche expansion ofthe two crane species during periods of preferred foodshortage was in agreement with the basic prediction ofoptimal foraging theory [22 23] It is also consistentwith a commonly observed fact that energy-maximizingmodels work better when animals food-limited [124]The dietary niche overlaps among the crane species
were low in winter 2017 with the four crane species di-verging in their principal food items However in winter2018 the niche overlaps were significantly higher thanexpected by chance They had similar diet compositionswith P criopolitanum rhizomes and Vallisneria tubersbeing the principal food items in the feces The in-creased overlap might have been caused by the highabundances of the two plant species in winter 2018which allowed the four crane species to share the samefood items with low interspecific competition Our studyis consistent with previous research that suggested thatamong-species dietary niches tended to overlap moreduring seasons of high food abundance [32 33 125126] Greater resource availability could promote an in-crease in species utilization of similar resources and re-duce competitive interactions between species [32 34126] By contrast during seasons with low food abun-dance species may partition limited food resources todecrease interspecific competition leading to a decreasein niche overlap [126 127] It has been suggested thatdiet partitioning facilitates the coexistence of manyclosely related species [7 128] The interannual variationin niche overlap observed in our study suggests that dietpartitioning may be restricted to specific temporal casesIn seasons or years with abundant food sources inter-specific competition may be relaxed and niche overlapcould be high
ConclusionsIn summary by using metabarcoding technology ourstudy gained new insights into cranesrsquo diets that are es-sential for informing habitat management to improve
Hou et al Frontiers in Zoology (2021) 1824 Page 10 of 17
the availability of foraging resources Vallisneria tuberswas confirmed as an important food source of cranesSurprisingly P criopolitanum also occupied high pro-portions in the feces of cranes and may also be import-ant Our study emphasizes the superior quality ofnatural wetlands compared to agricultural fields as for-aging habitats for cranes The recent degradation of sub-merged macrophytes at Poyang Lake potentiallythreatens the survival of cranes especially SiberianCranes Therefore it is urgent to reverse the broad de-cline of submerged macrophytes and provide importantforaging habitats for cranes during years of tuber col-lapses The abundances of Vallisneria tubers and prob-ably P criopolitanum at Poyang Lake greatly influencedthe dietary niche width and overlaps among crane spe-cies Cranes exhibited low dietary niche overlap duringperiods of preferred food shortage but significantly highoverlap during periods when preferred foods were plen-tiful While high niche overlap is often interpreted as in-tense interspecific competition our study highlights theimportance of taking food abundance into account
Materials and methodsStudy areasPoyang Lake the largest freshwater lake in China is oneof the most important wintering areas for waterbirdsalong the East Asian-Australasian Flyway (Fig 1) [51129] It is characterized by dramatic seasonal hydro-logical fluctuations with an average change of 924 mbetween the summer high and winter low water levels[130] During summer the inundation area is gt 2500km2 [131] In autumn and winter the water level dropscausing the inundation area to shrink to lt 1000 km2
[131] Poyang Lake contains 102 sublakes which are in-undated and integrated with the main body of PoyangLake during summer and are isolated during winter[132] As the water level drops the sublakes appear andlarge areas of grasslands mudflats and shallow waterareas are exposed providing winter foraging and roost-ing areas for approximately 420000 waterbirds from ap-proximately 111 species [133 134]
Vallisneria tuber surveyThe density and biomass of Vallisneria tubers were sur-veyed at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake in late October and earlyNovember of 2017 and 2018 before large numbers ofwintering waterbirds arrived We set 76 183 and 36sampling plots uniformly located at Dahuchi Shahu andChanghuchi respectively Vallisneria tubers were col-lected from two quadrats in each plot with a locallymade steel grab sampler The sampler has two long
handles At the end of the handles there are two scoopsfacing each other The sampler was inserted into thesubstrate and the scoops brought together to collectabout 15 cm long times 13 cm wide times 30 cm high sample ofsubstrate Tubers were subsequently cleaned andcounted The dry weight of tubers in each quadrat wasweighed after oven-drying at 80 degC until constant weightThe tuber density and biomass in each plot was calcu-lated by dividing total tuber number and tuber massacross two quadrats by quadrat area respectively Weused t-test to explore whether the tuber density and bio-mass in the winters of 2017 and 2018 significantly devi-ated from the historical average values in the wintersfrom 1999 to 2016 [57] The historical tuber density andbiomass data were collected by the Poyang Lake Na-tional Nature Reserve at three sublakes (Dahuchi Shahuand Meixihu) [57] R 360 was used to do the t-testanalyses
Fecal sample collectionFecal samples were collected at Poyang Lake in the win-ters of 2017 and 2018 We first investigated the distribu-tions of cranes When we saw foraging cranes we waiteduntil they left to avoid disturbing them Soon after theyleft we went to the foraging sites to collect fresh feces(ie feces with wet surface) with sterilized tweezers Tominimize the probability of multiple samples from thesame individual all collected samples were separated byat least 5 m We stored the samples in liquid nitrogen inthe field and transferred them to a minus 80 degC refrigeratorfor long-term storage in the laboratory To ensure thatthe sources of the feces were known we tried to collectsamples from monospecific flocks For samples collectedfrom mixed-species flocks we used DNA analysis toidentify the species that deposited the fecal samplesPlants were also collected in the field to construct aDNA barcoding reference database
Species identificationBecause of the similar morphologies of the feces of thefour crane species we used DNA analysis to identify thespecies that deposited the fecal samples collected frommixed-species flocks For the genetic species assignmentof each fecal sample we scraped the external surfacewhich contain DNA from the host species as a result ofthe sloughing of cells from the digestive tract DNA wasextracted with the QIAamp Power Fecal DNA Kit (Qia-gen USA) according to the manufacturerrsquos guidelinesFor each round of DNA extraction negative controls(ie extraction without feces) were included to monitorfor possible contamination We amplified the mitochon-drial DNA D-loop fragment with the primer pair DL02F
Hou et al Frontiers in Zoology (2021) 1824 Page 11 of 17
(5prime-3prime GCGGCCCGAAAAGCCGCTG) and DL02R (5prime-3prime GCCCTGACATAGGAACCAGAGGCGC) [135]PCR amplifications were carried out in a total volume of25 μl containing 125 μl PCR mix (Tiangen BeijingChina) 1 μl DNA 1 μl each primer and 95 μl H2O Thereaction conditions were as follows denaturation at94 degC for 4 min followed by 34 cycles at 94 degC for 48 s56 degC for 48 s and 72 degC for 1 min and a final 10 min at72 degC We checked for the presence of a PCR product ofsuitable length by electrophoresis on a 1 agarose gelSequencing was carried out on an ABI 3730xl analyzerby Sangon Biotech (Shanghai China) The complete se-quences were assembled using Seqman II (DNASTAR)and compared visually to the original chromatograms toavoid reading errors The taxonomic assignment wasconducted by searching against the nucleotide databaseof GenBank using Web BLAST
Construction of a barcode reference databaseWe downloaded the trnL sequences of 48 plant speciescommonly seen at Poyang Lake from GenBank For theplant species not included in GenBank we sequencedtrnL by ourselves Total DNA was extracted using thehexadecyltrimethylammonium bromide (CTAB) proto-col We used primer pair c (5prime-3prime CGAAATCGGTAGACGCTACG) and h (5prime-3prime CCATTGAGTCTCTGCACCTATC) to amplify an approximately 150 bp regionof the chloroplast trnL intron [136] This primer pairwas chosen as it can amplify DNA from a wide range ofwetland plants [70] PCR amplifications were carried outin a total volume of 25 μl containing 125 μl PCR mix(Tiangen Beijing China) 1 μl DNA 1 μl each primerand 95 μl H2O The reaction conditions were as followsdenaturation at 94 degC for 4 min followed by 35 cycles at94 degC for 30 s 56 degC for 30 s and 72 degC for 45 s and afinal 10 min at 72 degC We used electrophoresis to checkthe PCR products ABI 3730xl analyzer to perform se-quencing and Seqman II to assemble the sequences
DNA extraction and high-throughput sequencing of fecalsamplesTo uncover plant component consumed by cranes amodified CTAB protocol [70] was used to extract theDNA from approximately 200 mg of the interior of eachfecal sample DNA extraction was conducted in a dedi-cated laboratory for analyses of DNA from samples withlow DNA quality For each batch of DNA extractionnegative controls were included to monitor for possiblecontamination The primers c and h with a 6 bp tagadded to 5prime ends of each primer were used to amplifytrnL gene The PCR amplification procedure was thesame as barcode database construction For each samplea unique combination of tags in the forward and reverseprimers was used so that sample-specific data can be
recovered after sequencing Three PCR replicates wereperformed for each sample to reduce amplification biasThe three replicates were pooled and the DNA concen-tration and purity were monitored on a 1 agarose gelOnly samples with a clear band were processed furtherSamples were purified using the SanPrep Column PCRProduct Purification Kit (Sangon Biotech ShanghaiChina) The purified PCR products were pooled takinginto account of DNA concentration to obtain an ap-proximately equal amount of amplicon DNA from eachfecal sample The pool of individually tagged ampliconswas used to prepare a sequencing library using the Tru-Seq DNA PCR-Free Sample Preparation Kit (IlluminaUSA) following manufacturerrsquos recommendations Thelibrary was sequenced on an Illumina HiSeq 2500 plat-form and 250 bp paired-end reads were generated Li-brary preparation and sequencing were conducted byNovogene (Beijing China)
Sequencing data analysisAfter high-throughput sequencing the paired-end readswere demultiplexed into sample-specific files based ontheir unique tags The tags and primer sequences weresubsequently trimmed The paired-end reads weremerged using FLASH 127 [137] and then filtered usingQIIME 170 [138] Sequences were dereplicated andclustered to OTUs using USEARCH 110667 [139] atthe similarity threshold of 97 All OTUs were assignedto taxonomic units by referring to the local plant data-base using blast 2231+ with thresholds of identity gt97 and e-value lt 10 eminus 50 If an OTU matched two ormore taxa it was assigned to a higher taxonomic levelthat included all taxaTo evaluate the sufficiency of sequencing depth and
sampling size individual- species- and sample-basedrarefaction curves were built using EstimateS 82[140] The curves were computed by randomly resam-pling sequences or samples and plotting these againstthe numbers of OTUs The read abundance data wereconverted to RRA of each food item using the follow-ing equation [79]
RRA frac14 1S
XS
Kfrac141
nikPTifrac141nik
100
where nik is the number of sequences of food item i insample k T is the total number of food items and S isthe sampling size
Dietary niche breadth and overlap among crane speciesTo explore whether the cranesrsquo diets showed substantialinterannual variation we calculated Pearson correlationsbetween diet compositions of each crane species in win-ter 2017 and winter 2018 The proportion of each food
Hou et al Frontiers in Zoology (2021) 1824 Page 12 of 17
item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

the availability of foraging resources Vallisneria tuberswas confirmed as an important food source of cranesSurprisingly P criopolitanum also occupied high pro-portions in the feces of cranes and may also be import-ant Our study emphasizes the superior quality ofnatural wetlands compared to agricultural fields as for-aging habitats for cranes The recent degradation of sub-merged macrophytes at Poyang Lake potentiallythreatens the survival of cranes especially SiberianCranes Therefore it is urgent to reverse the broad de-cline of submerged macrophytes and provide importantforaging habitats for cranes during years of tuber col-lapses The abundances of Vallisneria tubers and prob-ably P criopolitanum at Poyang Lake greatly influencedthe dietary niche width and overlaps among crane spe-cies Cranes exhibited low dietary niche overlap duringperiods of preferred food shortage but significantly highoverlap during periods when preferred foods were plen-tiful While high niche overlap is often interpreted as in-tense interspecific competition our study highlights theimportance of taking food abundance into account
Materials and methodsStudy areasPoyang Lake the largest freshwater lake in China is oneof the most important wintering areas for waterbirdsalong the East Asian-Australasian Flyway (Fig 1) [51129] It is characterized by dramatic seasonal hydro-logical fluctuations with an average change of 924 mbetween the summer high and winter low water levels[130] During summer the inundation area is gt 2500km2 [131] In autumn and winter the water level dropscausing the inundation area to shrink to lt 1000 km2
[131] Poyang Lake contains 102 sublakes which are in-undated and integrated with the main body of PoyangLake during summer and are isolated during winter[132] As the water level drops the sublakes appear andlarge areas of grasslands mudflats and shallow waterareas are exposed providing winter foraging and roost-ing areas for approximately 420000 waterbirds from ap-proximately 111 species [133 134]
Vallisneria tuber surveyThe density and biomass of Vallisneria tubers were sur-veyed at three sublakes (Dahuchi Shahu and Changhu-chi Fig 1) of Poyang Lake in late October and earlyNovember of 2017 and 2018 before large numbers ofwintering waterbirds arrived We set 76 183 and 36sampling plots uniformly located at Dahuchi Shahu andChanghuchi respectively Vallisneria tubers were col-lected from two quadrats in each plot with a locallymade steel grab sampler The sampler has two long
handles At the end of the handles there are two scoopsfacing each other The sampler was inserted into thesubstrate and the scoops brought together to collectabout 15 cm long times 13 cm wide times 30 cm high sample ofsubstrate Tubers were subsequently cleaned andcounted The dry weight of tubers in each quadrat wasweighed after oven-drying at 80 degC until constant weightThe tuber density and biomass in each plot was calcu-lated by dividing total tuber number and tuber massacross two quadrats by quadrat area respectively Weused t-test to explore whether the tuber density and bio-mass in the winters of 2017 and 2018 significantly devi-ated from the historical average values in the wintersfrom 1999 to 2016 [57] The historical tuber density andbiomass data were collected by the Poyang Lake Na-tional Nature Reserve at three sublakes (Dahuchi Shahuand Meixihu) [57] R 360 was used to do the t-testanalyses
Fecal sample collectionFecal samples were collected at Poyang Lake in the win-ters of 2017 and 2018 We first investigated the distribu-tions of cranes When we saw foraging cranes we waiteduntil they left to avoid disturbing them Soon after theyleft we went to the foraging sites to collect fresh feces(ie feces with wet surface) with sterilized tweezers Tominimize the probability of multiple samples from thesame individual all collected samples were separated byat least 5 m We stored the samples in liquid nitrogen inthe field and transferred them to a minus 80 degC refrigeratorfor long-term storage in the laboratory To ensure thatthe sources of the feces were known we tried to collectsamples from monospecific flocks For samples collectedfrom mixed-species flocks we used DNA analysis toidentify the species that deposited the fecal samplesPlants were also collected in the field to construct aDNA barcoding reference database
Species identificationBecause of the similar morphologies of the feces of thefour crane species we used DNA analysis to identify thespecies that deposited the fecal samples collected frommixed-species flocks For the genetic species assignmentof each fecal sample we scraped the external surfacewhich contain DNA from the host species as a result ofthe sloughing of cells from the digestive tract DNA wasextracted with the QIAamp Power Fecal DNA Kit (Qia-gen USA) according to the manufacturerrsquos guidelinesFor each round of DNA extraction negative controls(ie extraction without feces) were included to monitorfor possible contamination We amplified the mitochon-drial DNA D-loop fragment with the primer pair DL02F
Hou et al Frontiers in Zoology (2021) 1824 Page 11 of 17
(5prime-3prime GCGGCCCGAAAAGCCGCTG) and DL02R (5prime-3prime GCCCTGACATAGGAACCAGAGGCGC) [135]PCR amplifications were carried out in a total volume of25 μl containing 125 μl PCR mix (Tiangen BeijingChina) 1 μl DNA 1 μl each primer and 95 μl H2O Thereaction conditions were as follows denaturation at94 degC for 4 min followed by 34 cycles at 94 degC for 48 s56 degC for 48 s and 72 degC for 1 min and a final 10 min at72 degC We checked for the presence of a PCR product ofsuitable length by electrophoresis on a 1 agarose gelSequencing was carried out on an ABI 3730xl analyzerby Sangon Biotech (Shanghai China) The complete se-quences were assembled using Seqman II (DNASTAR)and compared visually to the original chromatograms toavoid reading errors The taxonomic assignment wasconducted by searching against the nucleotide databaseof GenBank using Web BLAST
Construction of a barcode reference databaseWe downloaded the trnL sequences of 48 plant speciescommonly seen at Poyang Lake from GenBank For theplant species not included in GenBank we sequencedtrnL by ourselves Total DNA was extracted using thehexadecyltrimethylammonium bromide (CTAB) proto-col We used primer pair c (5prime-3prime CGAAATCGGTAGACGCTACG) and h (5prime-3prime CCATTGAGTCTCTGCACCTATC) to amplify an approximately 150 bp regionof the chloroplast trnL intron [136] This primer pairwas chosen as it can amplify DNA from a wide range ofwetland plants [70] PCR amplifications were carried outin a total volume of 25 μl containing 125 μl PCR mix(Tiangen Beijing China) 1 μl DNA 1 μl each primerand 95 μl H2O The reaction conditions were as followsdenaturation at 94 degC for 4 min followed by 35 cycles at94 degC for 30 s 56 degC for 30 s and 72 degC for 45 s and afinal 10 min at 72 degC We used electrophoresis to checkthe PCR products ABI 3730xl analyzer to perform se-quencing and Seqman II to assemble the sequences
DNA extraction and high-throughput sequencing of fecalsamplesTo uncover plant component consumed by cranes amodified CTAB protocol [70] was used to extract theDNA from approximately 200 mg of the interior of eachfecal sample DNA extraction was conducted in a dedi-cated laboratory for analyses of DNA from samples withlow DNA quality For each batch of DNA extractionnegative controls were included to monitor for possiblecontamination The primers c and h with a 6 bp tagadded to 5prime ends of each primer were used to amplifytrnL gene The PCR amplification procedure was thesame as barcode database construction For each samplea unique combination of tags in the forward and reverseprimers was used so that sample-specific data can be
recovered after sequencing Three PCR replicates wereperformed for each sample to reduce amplification biasThe three replicates were pooled and the DNA concen-tration and purity were monitored on a 1 agarose gelOnly samples with a clear band were processed furtherSamples were purified using the SanPrep Column PCRProduct Purification Kit (Sangon Biotech ShanghaiChina) The purified PCR products were pooled takinginto account of DNA concentration to obtain an ap-proximately equal amount of amplicon DNA from eachfecal sample The pool of individually tagged ampliconswas used to prepare a sequencing library using the Tru-Seq DNA PCR-Free Sample Preparation Kit (IlluminaUSA) following manufacturerrsquos recommendations Thelibrary was sequenced on an Illumina HiSeq 2500 plat-form and 250 bp paired-end reads were generated Li-brary preparation and sequencing were conducted byNovogene (Beijing China)
Sequencing data analysisAfter high-throughput sequencing the paired-end readswere demultiplexed into sample-specific files based ontheir unique tags The tags and primer sequences weresubsequently trimmed The paired-end reads weremerged using FLASH 127 [137] and then filtered usingQIIME 170 [138] Sequences were dereplicated andclustered to OTUs using USEARCH 110667 [139] atthe similarity threshold of 97 All OTUs were assignedto taxonomic units by referring to the local plant data-base using blast 2231+ with thresholds of identity gt97 and e-value lt 10 eminus 50 If an OTU matched two ormore taxa it was assigned to a higher taxonomic levelthat included all taxaTo evaluate the sufficiency of sequencing depth and
sampling size individual- species- and sample-basedrarefaction curves were built using EstimateS 82[140] The curves were computed by randomly resam-pling sequences or samples and plotting these againstthe numbers of OTUs The read abundance data wereconverted to RRA of each food item using the follow-ing equation [79]
RRA frac14 1S
XS
Kfrac141
nikPTifrac141nik
100
where nik is the number of sequences of food item i insample k T is the total number of food items and S isthe sampling size
Dietary niche breadth and overlap among crane speciesTo explore whether the cranesrsquo diets showed substantialinterannual variation we calculated Pearson correlationsbetween diet compositions of each crane species in win-ter 2017 and winter 2018 The proportion of each food
Hou et al Frontiers in Zoology (2021) 1824 Page 12 of 17
item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

(5prime-3prime GCGGCCCGAAAAGCCGCTG) and DL02R (5prime-3prime GCCCTGACATAGGAACCAGAGGCGC) [135]PCR amplifications were carried out in a total volume of25 μl containing 125 μl PCR mix (Tiangen BeijingChina) 1 μl DNA 1 μl each primer and 95 μl H2O Thereaction conditions were as follows denaturation at94 degC for 4 min followed by 34 cycles at 94 degC for 48 s56 degC for 48 s and 72 degC for 1 min and a final 10 min at72 degC We checked for the presence of a PCR product ofsuitable length by electrophoresis on a 1 agarose gelSequencing was carried out on an ABI 3730xl analyzerby Sangon Biotech (Shanghai China) The complete se-quences were assembled using Seqman II (DNASTAR)and compared visually to the original chromatograms toavoid reading errors The taxonomic assignment wasconducted by searching against the nucleotide databaseof GenBank using Web BLAST
Construction of a barcode reference databaseWe downloaded the trnL sequences of 48 plant speciescommonly seen at Poyang Lake from GenBank For theplant species not included in GenBank we sequencedtrnL by ourselves Total DNA was extracted using thehexadecyltrimethylammonium bromide (CTAB) proto-col We used primer pair c (5prime-3prime CGAAATCGGTAGACGCTACG) and h (5prime-3prime CCATTGAGTCTCTGCACCTATC) to amplify an approximately 150 bp regionof the chloroplast trnL intron [136] This primer pairwas chosen as it can amplify DNA from a wide range ofwetland plants [70] PCR amplifications were carried outin a total volume of 25 μl containing 125 μl PCR mix(Tiangen Beijing China) 1 μl DNA 1 μl each primerand 95 μl H2O The reaction conditions were as followsdenaturation at 94 degC for 4 min followed by 35 cycles at94 degC for 30 s 56 degC for 30 s and 72 degC for 45 s and afinal 10 min at 72 degC We used electrophoresis to checkthe PCR products ABI 3730xl analyzer to perform se-quencing and Seqman II to assemble the sequences
DNA extraction and high-throughput sequencing of fecalsamplesTo uncover plant component consumed by cranes amodified CTAB protocol [70] was used to extract theDNA from approximately 200 mg of the interior of eachfecal sample DNA extraction was conducted in a dedi-cated laboratory for analyses of DNA from samples withlow DNA quality For each batch of DNA extractionnegative controls were included to monitor for possiblecontamination The primers c and h with a 6 bp tagadded to 5prime ends of each primer were used to amplifytrnL gene The PCR amplification procedure was thesame as barcode database construction For each samplea unique combination of tags in the forward and reverseprimers was used so that sample-specific data can be
recovered after sequencing Three PCR replicates wereperformed for each sample to reduce amplification biasThe three replicates were pooled and the DNA concen-tration and purity were monitored on a 1 agarose gelOnly samples with a clear band were processed furtherSamples were purified using the SanPrep Column PCRProduct Purification Kit (Sangon Biotech ShanghaiChina) The purified PCR products were pooled takinginto account of DNA concentration to obtain an ap-proximately equal amount of amplicon DNA from eachfecal sample The pool of individually tagged ampliconswas used to prepare a sequencing library using the Tru-Seq DNA PCR-Free Sample Preparation Kit (IlluminaUSA) following manufacturerrsquos recommendations Thelibrary was sequenced on an Illumina HiSeq 2500 plat-form and 250 bp paired-end reads were generated Li-brary preparation and sequencing were conducted byNovogene (Beijing China)
Sequencing data analysisAfter high-throughput sequencing the paired-end readswere demultiplexed into sample-specific files based ontheir unique tags The tags and primer sequences weresubsequently trimmed The paired-end reads weremerged using FLASH 127 [137] and then filtered usingQIIME 170 [138] Sequences were dereplicated andclustered to OTUs using USEARCH 110667 [139] atthe similarity threshold of 97 All OTUs were assignedto taxonomic units by referring to the local plant data-base using blast 2231+ with thresholds of identity gt97 and e-value lt 10 eminus 50 If an OTU matched two ormore taxa it was assigned to a higher taxonomic levelthat included all taxaTo evaluate the sufficiency of sequencing depth and
sampling size individual- species- and sample-basedrarefaction curves were built using EstimateS 82[140] The curves were computed by randomly resam-pling sequences or samples and plotting these againstthe numbers of OTUs The read abundance data wereconverted to RRA of each food item using the follow-ing equation [79]
RRA frac14 1S
XS
Kfrac141
nikPTifrac141nik
100
where nik is the number of sequences of food item i insample k T is the total number of food items and S isthe sampling size
Dietary niche breadth and overlap among crane speciesTo explore whether the cranesrsquo diets showed substantialinterannual variation we calculated Pearson correlationsbetween diet compositions of each crane species in win-ter 2017 and winter 2018 The proportion of each food
Hou et al Frontiers in Zoology (2021) 1824 Page 12 of 17
item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

item in each sample was used as input file TheShannon-Wiener diversity index (Hprime) was calculated toexplore the diet diversity of each crane species The fol-lowing equations were used
H0 frac14 minus
Xs
ifrac141
Pi lnPi
where Pi is the proportion of food item i out of all foodsand S is the total number of food items The dietarybreadth (B) was measured using Levinsrsquo index [141] ac-cording to the following formula
B frac14 1=Xs
ifrac141P2i
Hurlbertrsquos formula [142] was applied to standardizethe trophic niche measure according to the followingformula
Ba frac14 Bminus1eth THORN= Sminus1eth THORN
where B is the Levins index of niche breadth and S isthe total number of prey categories Ba values range be-tween 0 (minimum diet breadth) and 1 (maximum dietbreadth)The dietary overlap of each species pair (Qjk) was cal-
culated using Piankarsquos index [143] according to the fol-lowing formula
Qjk frac14Ps
ifrac141PijPikffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiPsifrac141P
2ij
Psifrac141P
2ik
q
where Qjk is Piankarsquos niche overlap index between spe-cies j and species k Pij is the proportion of resource iout of all resources used by species j and Pik is the pro-portion of resource i out of all resources used by speciesk Qjk ranges from 0 where two species have no fooditems in common to 1 where there is complete overlapin resource useWe tested whether the observed niche overlap differed
from what would be expected by chance using the pro-gram EcoSim 100 [144] Null models were calculatedbased on the randomization algorithm RA3 and 1000simulated resource utilization matrices were generatedto compare with observed resource utilization dataPatterns of diet composition and overlap of the four
crane species were visualized in two-dimensional spaceusing the NMDS plots We performed NMDS analysisbased on the Bray-Curtis dissimilarity The read abun-dance of each food item in each sample was used as theinput file The R package ldquoVeganrdquo [145] was used for theNMDS analysis and ldquoggplotsrdquo [146] was used to creategraphics
AbbreviationsNMDS Nonmetric multidimensional scaling OTU Operational taxonomyunits RRA Relative read abundance
Supplementary InformationThe online version contains supplementary material available at httpsdoiorg101186s12983-021-00411-2
Additional file 1 Table S1 Sampling site information Table S2Relative read abundance (RRA ) of each food item in the diets ofSiberian Cranes White-naped Cranes Hooded Cranes and EurasianCranes in the winters of 2017 and 2018 Figure S1 Rarefaction curve ofeach crane species
AcknowledgementsWe thank Lanhua Wang Haiyan Zhou Xiaolong Wang the Poyang LakeNational Nature Reserve and the Nanji Wetland National Nature Reserve forassisting in sample collection We thank Zhichun Lan James Burnham andChunlan Zhang for helping data analyses We also thank Jeb Barzen fromPrivate Lands Conservation Andrew J Caven from Crane Trust and ananonymous reviewer for reviewing our manuscript and giving valuablecomments and suggestions
Authorsrsquo contributionsJH collected fecal samples performed the experiments and analyzed thedata LL conducted tuber surveys YW collected fecal samples and performedthe experiments WW conceived the study collected fecal samples analyzedthe data and wrote the original draft HZ ND and PL revised the manuscriptAll authors read and approved the final manuscript
FundingThis work was supported by the National Natural Science Foundation ofChina (No 31772480) the Natural Science Foundation of Jiangxi Province(No 20181BAB214007) and Funding Project of Jiangxi Academy of Sciences(No 2020-YGY-01)
Availability of data and materialsThe datasets used andor analyzed during the current study are availablefrom the corresponding authors on reasonable request
Declarations
Ethics approval and consent to participateOur study collected bird fecal samples using a non-invasive method Ourfieldwork was permitted by the Poyang Lake National Nature Reserve andthe Nanji Wetland National Nature Reserve
Consent for publicationNot applicable
Competing interestsThe authors declare they have no competing interests
Author details1Jiangxi Province Key Laboratory of Watershed Ecosystem Change andBiodiversity Center for Watershed Ecology Institute of Life Science andSchool of Life Science Nanchang University Nanchang 330031 China 2KeyLaboratory of Poyang Lake Environment and Resource Utilization Ministry ofEducation Nanchang University Nanchang 330031 China 3Jiangxi PoyangLake Wetland Conservation and Restoration National Permanent ScientificResearch Base National Ecosystem Research Station of Jiangxi Poyang LakeWetland Nanchang 330031 China 4Jiangxi Poyang Lake National NatureReserve Authority Nanchang 330038 China 5The Institute of Biology andResources Jiangxi Academy of Sciences Nanchang 330096 China
Hou et al Frontiers in Zoology (2021) 1824 Page 13 of 17
Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

Received 1 April 2020 Accepted 29 April 2021
References1 Schoener TW Resource partitioning in ecological communities Science
1974185(4145)27ndash39 httpsdoiorg101126science1854145272 Hutchinson GE Homage to Santa Rosalia or why are there so many kinds of
animals Am Nat 195993(870)145ndash59 httpsdoiorg1010862820703 Arthur W The niche in competition and evolution New York Wiley 19874 Sorensen MFL Niche shifts of coal tits Parus ater in Denmark J Avian Biol
199728(1)68ndash72 httpsdoiorg10230736770965 Aguiar CM Santos GMdM Martins CF Presley SJ Trophic niche breadth and
niche overlap in a guild of flower-visiting bees in a Brazilian dry forestApidologie 201344(2)153ndash62 httpsdoiorg101007s13592-012-0167-4
6 Leray M Alldredge AL Yang JY Meyer CP Holbrook SJ Schmitt RJ et alDietary partitioning promotes the coexistence of planktivorous species oncoral reefs Mol Ecol 201928(10)2694ndash710 httpsdoiorg101111mec15090
7 Luiselli L Resource partitioning and interspecific competition in snakes thesearch for general geographical and guild patterns Oikos 2006114(2)193ndash211 httpsdoiorg101111j20060030-129914064x
8 Herrera LG Hobson KA Rodriacuteguez M Hernandez P Trophic partitioning intropical rain forest birds insights from stable isotope analysis Oecologia2003136(3)439ndash44 httpsdoiorg101007s00442-003-1293-5
9 Kartzinel TR Chen PA Coverdale TC Erickson DL Kress WJ Kuzmina MLet al DNA metabarcoding illuminates dietary niche partitioning by Africanlarge herbivores P Nat Acad Sci USA 2015112(26)8019ndash24 httpsdoiorg101073pnas1503283112
10 Alatalo RV Seasonal dynamics of resource partitioning among foliage-gleaning passerines in northern Finland Oecologia 198045(2)190ndash6httpsdoiorg101007BF00346459
11 Martins RC Catry T Santos CD Palmeirim JM Granadeiro JP Seasonalvariations in the diet and foraging behaviour of dunlins Calidris alpina in asouth European estuary improved feeding conditions for northwardmigrants PLoS One 20138(12)e81174 httpsdoiorg101371journalpone0081174
12 Preston CR Jones RE Horton NS Golden eagle diet breadth andreproduction in relation to fluctuations in primary prey abundance inWyomings Bighorn Basin J Raptor Res 201751(3)334ndash46 httpsdoiorg103356JRR-16-391
13 Isacch JP Darrieu CA Martinez MM Food abundance and dietaryrelationships among migratory shorebirds using grasslands during the non-breeding season Waterbirds 200528(2)238ndash45 httpsdoiorg1016751524-4695(2005)028[0238FAADRA]20CO2
14 Piersma T Degoeij P Tulp I An evaluation of intertidal feeding habitatsfrom a shorebird perspective towards relevant comparisons betweentemperate and tropical mudflats Netherlands J Sea Res 199331(4)503ndash12httpsdoiorg1010160077-7579(93)90062-W
15 Quinn JT Hamilton DJ Variation in diet of Semipalmated sandpipers(Calidris pusilla) during stopover in the upper bay of Fundy Canada Can JZool 201290(9)1181ndash90 httpsdoiorg101139z2012-086
16 MacDonald EC Ginn MG Hamilton DJ Variability in foraging behavior andimplications for diet breadth among Semipalmated sandpipers staging inthe upper bay of Fundy Condor 2012114135ndash44
17 Wei Z Zheng M Zhou L Xu W Flexible foraging response of winteringhooded cranes (Grus monacha) to food availability in the lakes of theYangtze River floodplain China Animals 202010(4)568 httpsdoiorg103390ani10040568
18 Dong HY Lu GY Zhong XY Yang XJ Winter diet and food selection of theblack-necked crane Grus nigricollis in Dashanbao Yunnan China PeerJ20164e1968 httpsdoiorg107717peerj1968
19 Baker AJ Gonzalez PM Piersma T Niles LJ do Nascimento IDS Atkinson PWet al Rapid population decline in red knots fitness consequences ofdecreased refuelling rates and late arrival in Delaware Bay P Roy Soc B Bio2004271875ndash82
20 MacArthur RH Pianka ER On optimal use of a patchy environment Am Nat1966100(916)603ndash9 httpsdoiorg101086282454
21 Pyke GH Optimal foraging theory a critical review Annu Rev Ecol Syst198415(1)523ndash75 httpsdoiorg101146annureves15110184002515
22 Schoener TW Theory of feeding strategies Annu Rev Ecol Syst 19712(1)369ndash404 httpsdoiorg101146annureves02110171002101
23 Pulliam HR On the theory of optimal diets Am Nat 1974108(959)59ndash74httpsdoiorg101086282885
24 Zhang S Ma Z Choi C Peng H Melville DS Zhao T et al Morphologicaland digestive adjustments buffer performance how staging shorebirdscope with severe food declines Ecol Evol 20199(7)3868ndash78 httpsdoiorg101002ece35013
25 Baker MC Baker AE Niche relationships among six species of shorebirds ontheir wintering and breeding ranges Ecol Monogr 197343(2)193ndash212httpsdoiorg1023071942194
26 Salinas-Ramos VB Montalvo LGH Leoacuten-Regagnon V Arrizabalaga-EscuderoA Clare EL Dietary overlap and seasonality in three species of mormoopidbats from a tropical dry forest Mol Ecol 201524(20)5296ndash307 httpsdoiorg101111mec13386
27 Clare EL Symondson WO Fenton MB An inordinate fondness for beetlesVariation in seasonal dietary preferences of night-roosting big brown bats(Eptesicus fuscus) Mol Ecol 201423(15)3633ndash47 httpsdoiorg101111mec12519
28 Layman CA Quattrochi JP Peyer CM Allgeier JE Niche widthcollapse in a resilient top predator following ecosystemfragmentation Ecol Lett 200710(10)937ndash44 httpsdoiorg101111j1461-0248200701087x
29 Lesser JS James WR Stallings CD Wilson RM Nelson JA Trophic niche sizeand overlap decreases with increasing ecosystem productivity Oikos 2020129(9)1303ndash13 httpsdoiorg101111oik07026
30 Calizza E Costantini ML Careddu G Rossi L Effect of habitat degradation oncompetition carrying capacity and species assemblage stability Ecol Evol20177(15)5784ndash96 httpsdoiorg101002ece32977
31 Calizza E Careddu G Caputi SS Rossi L Costantini ML Time- and depth-wise trophic niche shifts in Antarctic benthos PLoS One 201813(3)e0194796 httpsdoiorg101371journalpone0194796
32 Pool T Holtgrieve G Elliott V McCann K McMeans B Rooney N et alSeasonal increases in fish trophic niche plasticity within a flood-pulse riverecosystem (Tonle Sap Lake Cambodia) Ecosphere 20178(7)e01881 httpsdoiorg101002ecs21881
33 Soininen EM Gauthier G Bilodeau F Berteaux D Gielly L Taberlet P et alHighly overlapping winter diet in two sympatric lemming species revealedby DNA metabarcoding PLoS One 201510(1)e0115335 httpsdoiorg101371journalpone0115335
34 Novcic I Weak niche partitioning by migrating shorebirds in a single-food-type environment Avian Biol Res 201912(3)109ndash17 httpsdoiorg1011771758155919841279
35 Forero MG Bortolotti GR Hobson KA Donazar JA Bertelloti M Blanco GHigh trophic overlap within the seabird community of ArgentineanPatagonia a multiscale approach J Anim Ecol 200473(4)789ndash801 httpsdoiorg101111j0021-8790200400852x
36 Wegge P Kastdalen L Habitat and diet of young grouse broods resourcepartitioning between Capercaillie (Tetrao urogallus) and black grouse (Tetraotetrix) in boreal forests J Ornithol 2008149(2)237 httpsdoiorg101007s10336-007-0265-7
37 Jaeger A Connan M Richard P Cherel Y Use of stable isotopes to quantifyseasonal changes of trophic niche and levels of population and individualspecialisation in seabirds Mar Ecol Prog Ser 2010401269ndash77 httpsdoiorg103354meps08380
38 Burnham J Barzen J Pidgeon AM Sun B Wu J Liu G et al Novel foragingby wintering Siberian cranes Leucogeranus leucogeranus at Chinas PoyangLake indicates broader changes in the ecosystem and raises new challengesfor a critically endangered species Bird Conserv Int 201727(2)204ndash23httpsdoiorg101017S0959270916000150
39 Chiu MC Kuo MH Sun YH Hong SY Kuo HC Effects of flooding on aviantop-predators and their invertebrate prey in a monsoonal Taiwan streamFreshw Biol 200831335ndash44
40 Hansen BB Aanes R Saeligther B-E Feeding-crater selection by high-arcticreindeer facing ice-blocked pastures Can J Zool 201088(2)170ndash7 httpsdoiorg101139Z09-130
41 Xu W Hu W Deng J Zhu J Zhou N Liu X Impacts of water depth andsubstrate type on Vallisneria natans at wave-exposed and sheltered sites ina eutrophic large lake Ecol Eng 201697344ndash54 httpsdoiorg101016jecoleng201610029
42 Li W Liu G Xiong B Pu Y The restoration of aquatic vegetation in lakes ofPoyang Lake nature reserve after catastrophic flooding 1998 J Wuhan BotRes 200422301ndash6
Hou et al Frontiers in Zoology (2021) 1824 Page 14 of 17
43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

43 Wu G de Leeuw J Barzen J Venus V Burnham J Liu Y et al In Li W editorA conceptual framework to integrate a simulation model of plant biomassfor Vallisneria spiralis L with remote sensing and a geographical informationsystem In wetland and water resource modeling and assessment awatershed perspective Boca Raton CRC Press 2008
44 Jia Y Jiao S Zhang Y Zhou Y Lei G Liu G Diet shift and its impact onforaging behavior of Siberian crane (Grus leucogeranus) in Poyang LakePLoS One 20138(6)e65843 httpsdoiorg101371journalpone0065843
45 Fox AD Cao L Zhang Y Barter M Zhao MJ Meng FJ et al Declines in thetuber-feeding waterbird guild at Shengjin Lake National Nature ReserveChinandasha barometer of submerged macrophyte collapse Aquat Conserv201121(1)82ndash91 httpsdoiorg101002aqc1154
46 Schmidt-Nielsen K Animal physiology adaptation and environmentCambridge Cambridge University Press 1997
47 Frederiksen M Daunt F Harris MP Wanless S The demographic impact ofextreme events stochastic weather drives survival and population dynamicsin a long-lived seabird J Anim Ecol 200877(5)1020ndash9 httpsdoiorg101111j1365-2656200801422x
48 Altwegg R Roulin A Kestenholz M Jenni L Demographic effects of extremewinter weather in the barn owl Oecologia 2006149(1)44ndash51 httpsdoiorg101007s00442-006-0430-3
49 Harris J Mirande C A global overview of cranes status threats andconservation priorities Chin Birds 20134(3)189ndash209 httpsdoiorg105122cbirds20130025
50 Li F Wu J Harris J Burnham J Number and distribution of cranes winteringat Poyang Lake China during 2011ndash2012 Chinese Birds 20123(3)180ndash90httpsdoiorg105122cbirds20120027
51 Barter M Cao L Chen L Lei G Results of a survey for waterbirds in thelower Yangtze floodplain China in January-February 2004 Forktail2005211
52 BirdLife International IUCN Red List for birds 2020 Available online athttpwwwbirdlifeorg Accessed 12 Jan 2020
53 Wang W Fraser JD Chen J Wintering waterbirds in the middle and lowerYangtze River floodplain changes in abundance and distribution BirdConserv Int 201727(2)167ndash86 httpsdoiorg101017S0959270915000398
54 Jiao S Zeng Q Sun G Lei G Improving conservation of cranes by modelingpotential wintering distributions in China J Res Ecol 2016744ndash50
55 Barzen J Engels M Burnham J Harris J Wu G Potential impacts of a watercontrol structure on the abundance and distribution of winteringwaterbirds at Poyang Lake Baraboo International Crane Foundation 2009
56 Chen B Cui P Liu G Li F Wu X Wu J et al Relationships betweenchanging water levels and numbers of wintering tuber-eating birds inPoyang Lake National Nature Reserve J Lake Sci 201426243ndash52
57 Hou J Liu Y Fraser JD Li L Zhao B Lan Z et al Drivers of a habitat shift bycritically endangered Siberian cranes evidence from long-term data EcolEvol 202010(20)11055ndash68 httpsdoiorg101002ece36720
58 Hu Z Lin Y Analysis of evolution process and driving factors for aquaticvegetations of Poyang Lake in 30 years Resour Environ Yangtze Basin 2019281947ndash55
59 Jian M Jian M Li L Wang S Yu H Yu G Distribution pattern of submergedplants in typical wetlands of Poyang Lake and its influencing factors ofwater environment Resour Environ Yangtze Basin 201524765ndash72
60 Li Y Zhong Y Shao R Yan C Jin J Shan J et al Modified hydrologicalregime from the three gorges dam increases the risk of food shortages forwintering waterbirds in Poyang Lake Global Ecol Conserv 202024e01286httpsdoiorg101016jgecco2020e01286
61 Wang W Wang L Hou J Man-made habitats have become importantforaging areas of Siberian cranes Chinese J Wildlife 201940133ndash7
62 Hou J Wang Y Jin B Wang L Wang W Food composition of Siberiancranes in agricultural fields in the Poyang Lake China Chinese J Zool 20195415ndash21
63 Zhong Y Li Y Numerical change of cranes in rice paddies at Poyang LakeChina Crane News 2018223ndash4
64 Li Y Rice paddy use by water birds at Poyang Lake China Crane News20182225ndash6
65 Pompanon F Deagle BE Symondson WOC Brown DS Jarman SN TaberletP Who is eating what diet assessment using next generation sequencingMol Ecol 201221(8)1931ndash50 httpsdoiorg101111j1365-294X201105403x
66 Hope PR Bohmann K Gilbert MTP Zepeda-Mendoza ML Razgour O JonesG Second generation sequencing and morphological faecal analysis reveal
unexpected foraging behaviour by Myotis nattereri (ChiropteraVespertilionidae) in winter Front Zool 201411(1)39 httpsdoiorg1011861742-9994-11-39
67 Lin X Hu S Liu S Huang H Unexpected prey of juvenile spotted scat(Scatophagus argus) near a wharf the prevalence of fouling organisms in stomachcontents Ecol Evol 20188(16)8547ndash54 httpsdoiorg101002ece34380
68 Symondson W Molecular identification of prey in predator diets Mol Ecol200211(4)627ndash41 httpsdoiorg101046j1365-294X200201471x
69 Razgour O Clare EL Zeale MR Hanmer J Schnell IB Rasmussen M et alHigh-throughput sequencing offers insight into mechanisms of resourcepartitioning in cryptic bat species Ecol Evol 20111(4)556ndash70 httpsdoiorg101002ece349
70 Yang Y Zhan A Cao L Meng F Xu W Selection of a marker gene toconstruct a reference library for wetland plants and the application ofmetabarcoding to analyze the diet of wintering herbivorous waterbirdsPeerJ 20164e2345 httpsdoiorg107717peerj2345
71 Zeale MR Butlin RK Barker GL Lees DC Jones G Taxon-specific PCR forDNA barcoding arthropod prey in bat faeces Mol Ecol Resour 201111(2)236ndash44 httpsdoiorg101111j1755-0998201002920x
72 Deagle BE Kirkwood R Jarman SN Analysis of Australian fur seal diet bypyrosequencing prey DNA in faeces Mol Ecol 200918(9)2022ndash38 httpsdoiorg101111j1365-294X200904158x
73 Srivathsan A Sha JC Vogler AP Meier R Comparing the effectiveness ofmetagenomics and metabarcoding for diet analysis of a leaf-feedingmonkey (Pygathrix nemaeus) Mol Ecol Resour 201515(2)250ndash61 httpsdoiorg1011111755-099812302
74 Gerwing TG Kim J-H Hamilton DJ Barbeau MA Addison JA Dietreconstruction using next-generation sequencing increases the knownecosystem usage by a shorebird Auk 2016133(2)168ndash77 httpsdoiorg101642AUK-15-1761
75 De Barba M Miquel C Boyer F Mercier C Rioux D Coissac E et al DNAmetabarcoding multiplexing and validation of data accuracy for dietassessment application to omnivorous diet Mol Ecol Resour 201414(2)306ndash23 httpsdoiorg1011111755-099812188
76 Xiong M Shao X Long Y Bu H Zhang D Wang D et al Molecular analysisof vertebrates and plants in scats of leopard cats (Prionailurus bengalensis) inSouthwest China J Mammal 201697(4)1054ndash64 httpsdoiorg101093jmammalgyw061
77 Zhong Y Li Y Waterbirds wintering in artificial wetland at Poyang Lake inwinter 20182019 China Crane News 2019234ndash8
78 Valentini A Miquel C Nawaz MA Bellemain E Coissac E Pompanon F et alNew perspectives in diet analysis based on DNA barcoding and parallelpyrosequencing the trnL approach Mol Ecol Resour 20099(1)51ndash60httpsdoiorg101111j1755-0998200802352x
79 Deagle BE Thomas AC McInnes JC Clarke LJ Vesterinen EJ Clare EL et alCounting with DNA in metabarcoding studies how should we convertsequence reads to dietary data Mol Ecol 201928(2)391ndash406 httpsdoiorg101111mec14734
80 Dahl KA Patterson WF Robertson A Ortmann AC DNA barcodingsignificantly improves resolution of invasive lionfish diet in the northernGulf of Mexico Biol Invasions 201719(6)1917ndash33 httpsdoiorg101007s10530-017-1407-3
81 Ando H Setsuko S Horikoshi K Suzuki H Umehara S Inoue-Murayama Met al Diet analysis by next-generation sequencing indicates the frequentconsumption of introduced plants by the critically endangered red-headedwood pigeon (Columba janthina nitens) in oceanic island habitats Ecol Evol20133(12)4057ndash69 httpsdoiorg101002ece3773
82 Swanson GA Bartonek JC Bias associated with food analysis in gizzards ofblue-winged teal J Wildlife Manage 197034(4)739ndash46 httpsdoiorg1023073799138
83 Bartonek JC Summer foods and feeding habits of diving ducks in Manitobavol 113 Madison University of Wisconsin 1968
84 Johnsgard PA Cranes of the world Lincoln Indiana University Press 198385 Zeng N Ji W Huang Z Liu Y Jia D Study on Siberian Cranes In Study on
Jiangxi Poyang Lake National Nature Reserve Wu Y Ji W Nanchang ChinaForestry Publishing House 2002135ndash137
86 Jiao S Jiang K Zuo A Wu M Lei G Zhou Y Foraging behavior and foodresources of wintering Grus monacha in China Sichuan J Zool 201736392ndash7
87 Cheng Y Home range and habitat selection of wintering white-napedcrane Grus vipio through GPS telemetry in Poyang Lake China BeijingBeijing Forestry University 2015 Master Thesis
Hou et al Frontiers in Zoology (2021) 1824 Page 15 of 17
88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

88 Meine C Archibald G The cranes status survey and conservation actionplan Gland and Cambridge IUCN 1996
89 Wu J Li F Burnham J Numerical distribution of Siberian cranes and theirrelation to food and water depth in Sha Hu at Poyang Lake China WetlandSci 201311305ndash12
90 Ding W Zhou F Research on wintering distribution of Siberian cranes InChina bird Researchs Edited by Gao W Beijing Science Press 1991
91 Harris J Liying S Higuchi H Ueta M Zhengwang Z Yanyun Z et alMigratory stopover and wintering locations in eastern China used by White-naped Cranes Grus vipio and Hooded Cranes G monacha as determined bysatellite tracking Forktail 20001693ndash100
92 Jiang J Study on abundance and distribution of wintering waterbirds andecological habits of four crane species in Poyang Lake Nanchang JiangxiNormal University 2016 Master Thesis
93 Zhang L Yin J Jiang Y Wang H Relationship between the hydrologicalconditions and the distribution of vegetation communities within thePoyang Lake National Nature Reserve China Ecol Inform 20121165ndash75httpsdoiorg101016jecoinf201205006
94 Zhang M Ni L Xu J He L Fu H Liu Z Annual dynamics of the wetlandplants community in Poyang Lake in response to water-level variations ResEnviron Sci 2013261057ndash63
95 Wang L Dronova I Gong P Yang W Li Y Liu Q A new time seriesvegetationndashwater index of phenologicalndashhydrological trait acrossspecies and functional types for Poyang Lake wetland ecosystemRemote Sens Environ 201212549ndash63 httpsdoiorg101016jrse201207003
96 Wan W Zhou L Song Y Shifts in foraging behavior of wintering hoodedcranes (Grus monacha) in three different habitats at Shengjin Lake ChinaAvian Res 20167(1)13 httpsdoiorg101186s40657-016-0047-0
97 Han Z Wang S Liu X Peng W Ge G Huang A Ecological thresholds for thedominated wetland plants of Poyang Lake along the gradient of floodingduration J Hydraul Eng 201950252ndash62
98 Ma Z Li B Zhao B Jing K Tang S Chen J Are artificial wetlands goodalternatives to natural wetlands for waterbirdsndasha case study onChongming Island China Biodivers Conserv 200413(2)333ndash50 httpsdoiorg101023BBIOC00000065029613159
99 Czech HA Parsons KC Agricultural wetlands and waterbirds a reviewWaterbirds 20022556ndash65
100 Amano T Conserving bird species in Japanese farmland past achievementsand future challenges Biol Conserv 2009142(9)1913ndash21 httpsdoiorg101016jbiocon200812025
101 Sundar KSG Subramanya S Bird use of rice fields in the Indian subcontinentWaterbirds 201033(sp1)44ndash70 httpsdoiorg101675063033s104
102 Tourenq C Bennetts RE Kowalski H Vialet E Lucchesi J-L Kayser Y et al Arericefields a good alternative to natural marshes for waterbird communitiesin the Camargue southern France Biol Conserv 2001100(3)335ndash43httpsdoiorg101016S0006-3207(01)00037-4
103 Masero JA Assessing alternative anthropogenic habitats for conservingwaterbirds Salinas as buffer areas against the impact of natural habitat lossfor shorebirds Biodivers Conserv 200312(6)1157ndash73 httpsdoiorg101023A1023021320448
104 Fox AD Abraham KF Why geese benefit from the transition from naturalvegetation to agriculture Ambio 201746(S2)188ndash97 httpsdoiorg101007s13280-016-0879-1
105 Nilsson L Bunnefeld N Persson J Mansson J Large grazing birds andagriculture-predicting field use of common cranes and implications for cropdamage prevention Agric Ecosyst Environ 2016219163ndash70 httpsdoiorg101016jagee201512021
106 Nowald G In Baraboo HJ editor Cranes and people agricultural andtourism In Cranes Agriculture and Climate Change WisconsinInternational Crane Foundation 2010 p 65ndash70
107 Fox AD Madsen J Threatened species to super-abundance the unexpectedinternational implications of successful goose conservation Ambio 201746(S2)179ndash87 httpsdoiorg101007s13280-016-0878-2
108 Fasola M Rubolini D Merli E Boncompagni E Bressan U Long-term trendsof heron and egret populations in Italy and the effects of climate human-induced mortality and habitat on population dynamics Popul Ecol 201052(1)59ndash72 httpsdoiorg101007s10144-009-0165-1
109 Toral GM Figuerola J Unraveling the importance of rice fields for waterbirdpopulations in Europe Biodivers Conserv 201019(12)3459ndash69 httpsdoiorg101007s10531-010-9907-9
110 Baasch DM Farrell PD Pearse AT Brandt DA Caven AJ Harner MJ et alDiurnal habitat selection of migrating whooping crane in the Great PlainsAvian Conserv Ecol 2019146
111 Wang Z Li Z Beauchamp G Jiang Z Flock size and human disturbance affectvigilance of endangered red-crowned cranes (Grus japonensis) Biol Conserv2011144(1)101ndash5 httpsdoiorg101016jbiocon201006025
112 Zhao Q Wang X Cao L Fox AD Why Chinese wintering geese hesitate toexploit farmland Ibis 2018160(3)703ndash5 httpsdoiorg101111ibi12605
113 Yu H Wang X Cao L Zhang L Jia Q Lee H et al Are declining populationsof wild geese in China lsquoprisonersrsquo of their natural habitats Curr Biol 201727(10)R376ndash7 httpsdoiorg101016jcub201704037
114 Shao M Gong H Dai N Zhi Y Xu N Lu P Study on time budgets andbehavioral rhythm of wintering Siberian cranes in a lotus pond reclamationarea in Poyang Lake Acta Ecol Sinica 2018385206ndash12
115 Sparling DW Krapu GL Communal roosting and foraging behavior ofstaging Sandhill cranes Wilson Bull 199410662ndash77
116 Nilsson L Persson J Bunnefeld N Maringnsson J Central place foraging in ahuman-dominated landscape how do common cranes select feeding sitesJ Avian Biol 202051e02487
117 Anteau MJ Sherfy MH Bishop AA Location and agricultural practicesinfluence spring use of harvested cornfields by cranes and geese inNebraska J Wildlife Manage 201175(5)1004ndash11 httpsdoiorg101002jwmg135
118 Choi C-Y Takekawa JY Xiong Y Liu Y Wikelski M Heine G et al Trackingdomestic ducks a novel approach for documenting poultry market chainsin the context of avian influenza transmission J Integr Agr 201615(7)1584ndash94 httpsdoiorg101016S2095-3119(15)61292-8
119 Agosta SJ Morton D Kuhn KM Feeding ecology of the bat Eptesicus fuscuslsquopreferredrsquo prey abundance as one factor influencing prey selection and dietbreadth J Zool 2003260(2)169ndash77 httpsdoiorg101017S0952836903003601
120 Khadka KK Singh N Magar KT James DA Dietary composition breadth andoverlap between seasonally sympatric Himalayan musk deer and livestockconservation implications J Nat Conser 20173830ndash6 httpsdoiorg101016jjnc201706001
121 Lei J Jia Y Wang Y Lei G Lu C Saintilan N et al Behavioural plasticity andtrophic niche shift how wintering geese respond to habitat alterationFreshw Biol 201964(6)1183ndash95 httpsdoiorg101111fwb13294
122 Krebs JR McCleery RH Optimization in behavioural ecology In Krebs JRDavies NB editors Behavioural ecology an evolutionary approachSunderland Sinauer Associates 1984
123 Stephens DW Krebs JR Foraging theory Princeton Princeton UniversityPress 1986
124 Kacelnik A Information primacy or preference for familiar foragingtechniques A critique of Inglis amp Ferguson Anim Behav 198735(3)925ndash6httpsdoiorg101016S0003-3472(87)80130-6
125 Durson AM Willson JD Winne CT Habitat influences diet overlap in aquaticsnake assemblages J Zool 2013291(3)185ndash93 httpsdoiorg101111jzo12061
126 Luiselli L Interspecific relationships between two species of sympatricAfrotropical water snake in relation to a seasonally fluctuating foodresource J Trop Ecol 200622(1)91ndash100 httpsdoiorg101017S0266467405002877
127 Steenhof K Kochert MN Dietary shifts of sympatric buteos during a preydecline Oecologia 198566(1)6ndash16 httpsdoiorg101007BF00378546
128 de Carvalho TC de Assis Montag LF dos Santos-Costa MC Dietcomposition and foraging habitat use by three species of water snakesHelicops Wagler 1830 (Serpentes Dipsadidae) in eastern BrazilianAmazonia J Herpetol 201751215ndash22
129 Wang W Fraser JD Chen J Distribution and long-term population trends ofwintering waterbirds in Poyang Lake China Wetlands 201939(Suppl 1)S125ndash35
130 Min Q On the regularites of water level fluctuations in Poyang Lake J LakeSci 19957281ndash8
131 Feng L Hu C Chen X Cai X Tian L Gan W Assessment of inundationchanges of Poyang Lake using MODIS observations between 2000 and2010 Remote Sens Environ 201212180ndash92 httpsdoiorg101016jrse201201014
132 Hu Z Zhang Z Liu Y Ji W Ge G The function and significance of theshallow-lakes in the Poyang Lake wetland ecosystem Jiangxi Hydraul SciTechn 201541317ndash23
Hou et al Frontiers in Zoology (2021) 1824 Page 16 of 17
133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-

133 Li Y Qian F Silbernagel J Larson H Community structure abundancevariation and population trends of waterbirds in relation to water levelfluctuation in Poyang Lake J Great Lakes Res 201945(5)976ndash85 httpsdoiorg101016jjglr201908002
134 Barzen J Studying and understanding wetland dynamics In Li F Liu G WuJ Zeng N Harris J Jin J editors Ecological Study of Wetlands andWaterbirds at Poyang Lake Beijing Popular Science Press 2012 p 256ndash66
135 Ponomarev A Tatarinova T Bubyakina V Smagulova F Morozov IKashentseva T et al Genetic diversity in Siberian crane (Grus leucogeranus)based on mitochondrial DNA D-loop polymorphism Doklady Biol Sci 2004397(1-6)321ndash3 httpsdoiorg101023BDOBS000003970491688ca
136 Taberlet P Coissac E Pompanon F Gielly L Miquel C Valentini A et alPower and limitations of the chloroplast trnL (UAA) intron for plant DNAbarcoding Nucleic Acids Res 200735(3)e14 httpsdoiorg101093nargkl938
137 Magoč T Salzberg SL FLASH fast length adjustment of short reads toimprove genome assemblies Bioinformatics 201127(21)2957ndash63 httpsdoiorg101093bioinformaticsbtr507
138 Caporaso JG Kuczynski J Stombaugh J Bittinger K Bushman FD CostelloEK et al QIIME allows analysis of high-throughput community sequencingdata Nat Meth 20107(5)335ndash6 httpsdoiorg101038nmethf303
139 Edgar RC Search and clustering orders of magnitude faster than BLASTBioinformatics 201026(19)2460ndash1 httpsdoiorg101093bioinformaticsbtq461
140 Colwell RK EstimateS Statistical estimation of species richness and sharedspecies from samples Retrieved from httppurloclcorgestimates 2006
141 Levins R Evolution in changing environments Princeton PrincetonUniversity Press 1968 httpsdoiorg1015159780691209418
142 Hurlbert SH The measurement of niche overlap and some relativesEcology 197859(1)67ndash77 httpsdoiorg1023071936632
143 Pianka ER The structure of lizard communities Annu Rev Ecol Syst 19734(1)53ndash74 httpsdoiorg101146annureves04110173000413
144 Gotelli NJ Ellison AM EcoSimR Version 100 Retrieved from httpwwwuvmedu~ngotelliEcoSimEcoSimhtml 2013
145 Oksanen J Blanchet FG Kindt R Legendre P Minchin PR Orsquohara R SimpsonGL Solymos P Stevens MHH Wagner H vegan Community EcologyPackage Retrieved from httpCRANR-projectorgpackage=vegan 2013
146 Wickham H ggplot2 elegant graphics for data analysis New York Springer2016 httpsdoiorg101007978-3-319-24277-4
Publisherrsquos NoteSpringer Nature remains neutral with regard to jurisdictional claims inpublished maps and institutional affiliations
Hou et al Frontiers in Zoology (2021) 1824 Page 17 of 17
- Abstract
-
- Background
- Results
- Conclusions
-
- Background
- Results
-
- Variation in Vallisneria tuber abundance
- Collection statistics
- High-throughput DNA sequencing
- Diet compositions of the four crane species
- Dietary niche breadth and overlap among crane species
-
- Discussion
-
- Diet composition
- Important food resources for cranes
- The roles of agricultural fields in crane protection
- Effects of tuber collapse on dietary niche breadth and overlap among crane species
-
- Conclusions
- Materials and methods
-
- Study areas
- Vallisneria tuber survey
- Fecal sample collection
- Species identification
- Construction of a barcode reference database
- DNA extraction and high-throughput sequencing of fecal samples
- Sequencing data analysis
- Dietary niche breadth and overlap among crane species
- Abbreviations
-
- Supplementary Information
- Acknowledgements
- Authorsrsquo contributions
- Funding
- Availability of data and materials
- Declarations
- Ethics approval and consent to participate
- Consent for publication
- Competing interests
- Author details
- References
- Publisherrsquos Note
-