improbable areas in the visual brain - vislab.ucl.ac.uk · visual brain that represent only one...
TRANSCRIPT

Improbable areas in the visual brainS. Zeki
University College London, London WC1E 6BT, UK
Recent results from anatomical, physiological and
imaging experiments cast doubt on the existence of
some areas in the primate visual brain and call for a
much overdue re-assessment of what is a conceptually
highly unsatisfactory view of how the primate visual
brain is organized, a view that has survived more or
less unscathed for at least 15 years and has been
embraced uncritically by a significant element in the
human brain imaging community. That view can be
summarized as follows: that there are areas in the
visual brain that represent only one quadrant of the
visual field, leaving the other quadrant of the hemifield
unrepresented, or represented in another area, leading
to what Jon Kaas has called ‘improbable areas’. It is
not the improbability of such a view that is surprising;
rather, it is its ready acceptance on the basis of
questionable evidence.
The right half of the visual field is represented in the leftbrain hemisphere, and vice versa, the two separaterepresentations being unified by a commissure that linksthe two cerebral hemispheres, the corpus callosum (Fig. 1).The discovery of many visual areas in the brain [1–4] withthe promise of more to come, naturally raises the questionof which criteria should be used in conferring the status ofa visual area on a cortical zone. Two obvious ones are thatan area should be activated by visual stimuli and shouldhave an independent and more or less complete map of thecontralateral visual field, to include both the upper andlower quadrants. This is so, even if a given quadrant, orpart of the retina, claims a disproportionately large spacein a cortical area, as happens even at the level of theprimary visual cortex (V1). To this can be added otherfeatures, such as a distinct set of anatomical (includingcallosal) connections, identifiable and unique functionalproperties [1] and a distinctive architecture, although thesearch for the latter has not always been fruitful. In 1986,Van Essen’s group [5] proposed a radical departure fromthis list by purporting to show that one of the areasconstituting the visual brain, area V3, does not have acomplete representation of the visual field, as had beensupposed from earlier anatomical studies [6,7]. Instead,they conceived of this area, which occupies a narrow stripanterior to area V2 (Figs 1 and 2) as consisting of twodifferent areas – a dorsal one called V3 and a ventral oneknown as ‘VP’ – each representing one quadrant only ofthe contralateral hemifield. They proposed this eventhough the retinotopic map in ‘VP’ in both the human[8–10] and the monkey [3,11] is a mirror image of that in
upper V3 (Fig. 1). The separation into two distinct areaswas based on the supposition that lower V3, unlike itsupper counterpart, not only lacks a direct anatomicalinput from V1, but also has a high proportion of colour-selective cells [5]. The implication was obvious: ‘VP’ is anarea registering activity only in the upper contralateralquadrant, without the capacity to register the sameactivity (including, above all, colour) when it occurs inthe lower contralateral quadrant – or leaving it to someother improbable area, one registering activity in lowerquadrant alone, to do so. This, in turn, leads to thesupposition that there could be other improbable areas inwhich only one quadrant of the visual field is represented(Fig. 3). This is odd: psychophysical experiments haveshown that some attributes are more readily perceivedwhen presented in one quadrant than in another [12,13],but none has ever shown that an attribute can be perceivedonly when presented in one quadrant alone.
In fact, the evidence against such improbable areas inthe visual brain is mounting. The notion that there is anasymmetrical anatomical input from V1 to upper andlower V3 was questioned years ago, when it was shownthat there is a direct input from V1 to lower V3 in Cebus
Fig. 1. Representation of the visual field in visual cortex. (a) Representation of the
right hemifield in the primary visual cortex, V1. The upper quadrant of the hemi-
field (red) is represented below the calcarine sulcus, whilst the lower quadrant
(green) is represented above the calcarine sulcus. The corpus callosum is shown
in blue. (b) The representation of the visual field in visual areas V1 to V3, shades of
red and pink indicating representation of the upper quadrant and shades of green
indicating representation of the lower quadrant. The dotted lines show the rep-
resentation of the horizontal meridian of the visual field (horizontal dotted line in
part a) in these visual areas. The solid lines between the areas show the represen-
tation of the vertical meridian.
TRENDS in Neurosciences
(a)
(b)
V1
V1
Calcarinesulcus
V2
V2
V3
V3
Corpuscallosum
Corresponding author: S. Zeki ([email protected]).
Opinion TRENDS in Neurosciences Vol.26 No.1 January 2003 23
http://tins.trends.com 0166-2236/02/$ - see front matter q 2002 Elsevier Science Ltd. All rights reserved. PII: S0166-2236(02)00008-5

monkeys [14] and when detailed recording experimentsshowed that there is a continuous representation of visualfields in V3 from lower to upper quadrants as one proceedsdorsoventrally [11]. Such findings, made in New Worldmonkeys, have been substantially reinforced by a morerecent study [15] showing that in the Old World macaquemonkey, too, there is a direct input from V1 to V3. Thatstudy, with other supporting evidence [16], has led Lyonand Kaas to conclude that V3 is one continuous area, nottwo separate areas, and that it is characteristic of allprimates. Thus, one of the main criteria for separating V3into two areas, namely an asymmetry in anatomical inputfrom V1, has lost its force.
The ready acceptance of such a subdivision in themacaque, and its uncritical translation into the humanbrain (in which the two corresponding areas have also beencalled V3 and ‘VP’) is surprising, because all humanimaging experiments have shown that upper and lowerparts of V3 are activated in the same way [17]. This speaksagainst its separation into two areas. More significantly,no human imaging study has ever shown that human ‘VP’is specifically or more vigorously activated with colour,even when the question has been directly addressed [18].The second criterion for separating V3 into two indepen-dent areas (namely, an emphasis on colour in ‘VP’) is, thus,
also weakened. The consensus of the evidence seems to bethat there is no justification for separating V3 into twoareas, on either anatomical or functional criteria. V3 is,instead, one whole area, in which both upper and lowerfields are represented – as was originally proposed.
The ‘improbable’ becomes implicitly acceptable
A consequence of the unquestioning acceptance of theseparation of V3 into two areas has been the implicitacceptance that there might be other such improbablecortical areas in which only one quarter of the visual fieldis represented. The advent of the phase-encoding method[19] for mapping visual fields in human cerebral cortexrevealed, in some hands, an area ‘V4v’ [20,21] locatedposterior to the colour centre, area V4 (Fig. 4). Theattached ‘v’ implies that the region maps the uppercontralateral quadrant only. No one seemed especiallyconcerned that the same method did not reveal ‘V4d’, thedorsal counterpart that should represent the inferiorcontralateral quadrant – presumably because of theimplicit acceptance that improbable areas might, afterall, exist. In fact, ‘V4v’ was thought to be distinct from theventrally located human V4, in which both contralateralquadrants are topographically mapped [22], and damageto which can lead to complete contralateral cerebral colourblindness (hemiachromatopsia) [23]. The confirmation byHadjikhani et al. [24], that both contralateral quadrants
Fig. 2. Areas of the visual brain. Areas V3 (dark blue), V3A (violet), V4 (red) and V5
(pink), and those of face and object recognition (blue), receive their input largely
from V1 (yellow) and V2 (green).
V1
V2
V3
V4
V5 (motion)
V3A
Face and objectrecognition areas
TRENDS in Neurosciences
Fig. 3. Two theories of how a hemifield is represented in the brain. The coloured
quadrants on the screen (a) are represented in an arbitrary region in the contralat-
eral side of the brain (b). (i) The conventional theory, which shows an orderly, con-
tinuous and more or less complete map of the contralateral hemifield. (ii) Van
Essen’s theory that such areas could be subdivided into two independent areas on
the basis of asymmetry in anatomical input and differences in function.
TRENDS in Neurosciences
(i)
(ii)
(a) (b)
Fig. 4. A ventral view of the human brain showing the representation of the upper
visual field (red) and the lower visual field (green) in V4 and V4a, the latter a newly
discovered area within the human colour centre. Blue lines around V4 show the
position of ‘V8’ according to Hadjikani et al. [24]. Purple circles caudal to V4 show
the position of area ‘V4v’ of Sereno et al. [7] and Hadjikani et al. [24], which has yet
to be shown to be a separate area [18]. Inset shows the visual field itself.
TRENDS in Neurosciences
'V4v'
V4
V4α
Opinion TRENDS in Neurosciences Vol.26 No.1 January 200324
http://tins.trends.com
are topographically mapped within human area V4 asoriginally defined [25], reveals eloquently the confusion[26] that is traceable in large measure to the blindacceptance of the concept of visual areas with only aquarter-field representation of the visual field. Hadjikhaniet al. showed, as we had before them [22], that there is acomplete representation of the contralateral hemifieldwithin the ventrally located colour centre, area V4. Butbecause of their belief in the existence of an area ‘V4v’ thatis distinct from our V4 and which represents only theupper contralateral hemifield, they imagined that theyhad discovered ‘a new retinotopic area that we call V8’.This ‘previously undifferentiated cortical area’ ‘was con-sistently located just beyond the most anterior retinotopicarea defined previously, area V4v’ (my emphasis on the v).An examination of their results shows, however, that their‘new’ retinotopic area has the same coordinates as our V4and is, thus, nothing more than the re-discovery of a
previously defined visual area (Fig. 4). Heywood andCowey [25] accepted this claim of a new area uncritically.They wrote that a ‘newlydefined [sic ] color area’ had beenfound and that it is this area, ‘V8’, rather than ‘the favoritecandidate, V4’, that, when lesioned, produces corticalcolour blindness. This led them unquestioningly to theview that ‘the human color center is distinct from area V4’(note how, in this uncritical acceptance, the small v hasbeen dropped from the equation). This is despite the factthat the ‘newlydefined color area’ has the same braincoordinates as V4 (Fig. 4).
Does ‘V4v’ exist?
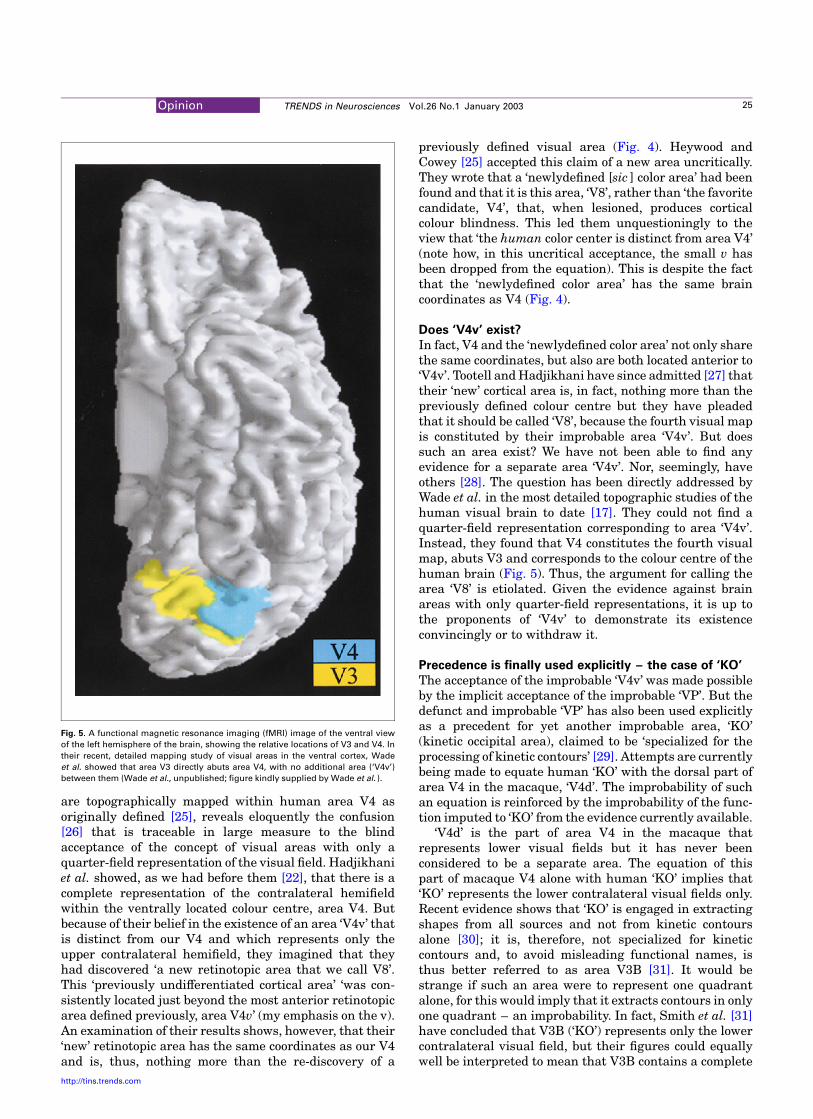
In fact, V4 and the ‘newlydefined color area’ not only sharethe same coordinates, but also are both located anterior to‘V4v’. Tootell and Hadjikhani have since admitted [27] thattheir ‘new’ cortical area is, in fact, nothing more than thepreviously defined colour centre but they have pleadedthat it should be called ‘V8’, because the fourth visual mapis constituted by their improbable area ‘V4v’. But doessuch an area exist? We have not been able to find anyevidence for a separate area ‘V4v’. Nor, seemingly, haveothers [28]. The question has been directly addressed byWade et al. in the most detailed topographic studies of thehuman visual brain to date [17]. They could not find aquarter-field representation corresponding to area ‘V4v’.Instead, they found that V4 constitutes the fourth visualmap, abuts V3 and corresponds to the colour centre of thehuman brain (Fig. 5). Thus, the argument for calling thearea ‘V8’ is etiolated. Given the evidence against brainareas with only quarter-field representations, it is up tothe proponents of ‘V4v’ to demonstrate its existenceconvincingly or to withdraw it.
Precedence is finally used explicitly – the case of ‘KO’
The acceptance of the improbable ‘V4v’ was made possibleby the implicit acceptance of the improbable ‘VP’. But thedefunct and improbable ‘VP’ has also been used explicitlyas a precedent for yet another improbable area, ‘KO’(kinetic occipital area), claimed to be ‘specialized for theprocessing of kinetic contours’ [29]. Attempts are currentlybeing made to equate human ‘KO’ with the dorsal part ofarea V4 in the macaque, ‘V4d’. The improbability of suchan equation is reinforced by the improbability of the func-tion imputed to ‘KO’ from the evidence currently available.
‘V4d’ is the part of area V4 in the macaque thatrepresents lower visual fields but it has never beenconsidered to be a separate area. The equation of thispart of macaque V4 alone with human ‘KO’ implies that‘KO’ represents the lower contralateral visual fields only.Recent evidence shows that ‘KO’ is engaged in extractingshapes from all sources and not from kinetic contoursalone [30]; it is, therefore, not specialized for kineticcontours and, to avoid misleading functional names, isthus better referred to as area V3B [31]. It would bestrange if such an area were to represent one quadrantalone, for this would imply that it extracts contours in onlyone quadrant – an improbability. In fact, Smith et al. [31]have concluded that V3B (‘KO’) represents only the lowercontralateral visual field, but their figures could equallywell be interpreted to mean that V3B contains a complete
Fig. 5. A functional magnetic resonance imaging (fMRI) image of the ventral view
of the left hemisphere of the brain, showing the relative locations of V3 and V4. In
their recent, detailed mapping study of visual areas in the ventral cortex, Wade
et al. showed that area V3 directly abuts area V4, with no additional area (‘V4v’)
between them (Wade et al., unpublished; figure kindly supplied by Wade et al. ).
Opinion TRENDS in Neurosciences Vol.26 No.1 January 2003 25
http://tins.trends.com

and independent representation of the entire contralateralhemifield, with the lower quadrant claiming more corticalspace. Moreover, other retinotopic studies of human visualcortex show that there is a complete representation of thecontralateral hemifield in V3B (‘KO’) [18].
The equation of human V3B with only a part of monkeyV4 (dorsal V4 or V4d) would mean that human V3B andmonkey V4d are improbable areas – even though theevidence suggests that both human V3B and macaque V4have a complete map of the contralateral hemifield – ‘V4d’being nothing more than the dorsal part of V4 in themacaque. In trying to understand why such claims havebeen so uncritically accepted, it becomes obvious thatprecedent, and especially that of ‘VP’, has played a big role.Tootell and Hadjikhani have written that ‘Although such‘separated’ quarter-field representations are conceptuallyunsatisfying, they are not unprecedented: the quarter-field representations in macaque ‘V3’ and ‘VP’ have longbeen considered separate areas by some investigators,based on empirical differences between V3 and VP’ [27].But there are also many who do not accept such improb-able areas, and the weight of evidence is on their side.
It is a wonder that so fragile a concept as that of visualareas in which only a quarter of the visual field isrepresented should have been so uncritically acceptedfor so many years, and that so much should have been builton such a flimsy view, even when much evidence speaksagainst it. This view was largely erected on the basis ofarea ‘VP’, but the evidence for such a separate area hasnever been convincing. Of course, when the originalconcept collapses, the rest follows, as with dominoes.This is what we are witnessing today with the demise of‘VP’. Perhaps the traditional, and even conservative, viewof what constitutes a visual area has considerable meritafter all.
References
1 Zeki, S.M. (1978) Functional specialization in the visual cortex of therhesus monkey. Nature 274, 423–428
2 Allman, J.M. et al. (1994) The functional organization of visual cortexin owl monkeys. In Aotus: The Owl Monkey (Baker, J.F. et al., eds),pp. 287–320, Academic Press
3 Baker, J.F. et al. (1981) Visual response properties of neurons in fourextrastriate areas of the owl monkey (Aotus trivirgatus ): a quantitat-ive comparison of medial, dorsomedial, dorsolateral, and middletemporal areas. J. Neurophysiol. 45, 397–416
4 Felleman, D.J. and Van Essen, D.C. (1991) Distributed hierarchicalprocessing in the primate cerebral cortex. Cereb. Cortex 1, 1–47
5 Burkhalter, A. and Van Essen, D.C. (1986) Processing of color, form anddisparity information in visual areas VP and V2 of ventral extrastriatecortex in the macaque monkey. J. Neurosci. 6, 2327–2351
6 Cragg, B.G. (1969) The topography of the afferent projections in
circumstriate visual cortex studied by the Nauta method. Vis. Res. 9,733–747
7 Zeki, S.M. (1969) Representation of central visual fields in prestriatecortex of monkey. Brain Res. 14, 271–291
8 Sereno, M.I. et al. (1995) Borders of multiple visual areas in humansrevealed by functional magnetic resonance imaging. Science 268,889–893
9 DeYoe, E.A. et al. (1996) Mapping striate and extrastriate visual areasin human cerebral cortex. Proc. Natl Acad. Sci. USA 93, 2382–2386
10 Shipp, S. et al. (1995) Retinotopic maps in human prestriate visualcortex – the demarcation of areas V2 and V3. Neuroimage 2, 125–132
11 Rosa, M.P.G. et al. (2000) Third tier ventral extrastriate cortex in NewWorld monkey, Cebus apella. Exp. Brain Res. 132, 287–305
12 Rubin, N. et al. (1996) Enhanced perception of illusory contours in thelower versus upper visual hemifields. Science 271, 651–653
13 Gordon, J. et al. (1997) The lower visual field is better than the uppervisual field at red/green recognition. Invest. Ophthalmol. Vis. Sci. 38,4199
14 Pinon, M.C. et al. (1998) Area V4 in Cebus monkey: extent andvisuotopic organization. Cereb. Cortex 8, 685–701
15 Lyon, D.C. and Kaas, J.H. (2002) Evidence for a modified V3 withdorsal and ventral halves in macaque monkeys. Neuron 33, 453–461
16 Lyon, D.C. and Kaas, J.H. (2001) Connectional and architectonicevidence for dorsal and ventral V3 and dorsomedial area in marmosetmonkeys. J. Neurosci. 21, 249–261
17 Wade, A.R. et al. (2002) Functional measurements of human ventraloccipital cortex: retinotopy and color. Phil. Trans. R. Soc. Lond. B. Biol.Sci. 357, 963–973
18 Press, W.A. et al. (2001) Visual areas and spatial summation in humanvisual cortex. Vision Res. 41, 1321–1332
19 Engel, S.A. et al. (1994) fMRI of human visual cortex. Nature 369, 52520 Sereno, M.I. et al. (1995) Borders of multiple visual areas in humans
revealed by functional magnetic resonance imaging. Science 268,889–893
21 Tootell, R.B. et al. (1996) New images from human visual cortex.Trends Neurosci. 19, 481–489
22 McKeefry, D. and Zeki, S. (1997) The position and topography of thehuman colour centre as revealed by functional magnetic resonanceimaging. Brain 120, 2229–2242
23 Verrey, L. (1888) Hemiachromatopsie droite absolute. Archs. Opthal.(Paris) 8, 289–301
24 Hadjikhani, N. et al. (1998) Retinotopy and color sensitivity in humanvisual cortical area V8. Nat. Neurosci. 1, 235–241
25 Zeki, S. et al. (1991) A direct demonstration of functional specializationin human visual cortex. J. Neurosci. 11, 641–649
26 Heywood, C. and Cowey, A. (1998) With color in mind. Nat. Neurosci. 1,171–173
27 Tootell, R.B.H. and Hadjikhani, N. (2001) Where is ‘dorsal’ V4 inhuman visual cortex? Retinotopic, topographic and functional evi-dence. Cereb. Cortex 11, 298–311
28 Kastner, S. et al. (1998) Mechanisms of directed attention in thehuman extrastriate cortex as revealed by functional MRI. Science 282,108–111
29 Van Oostende, S. et al. (1997) The kinetic occipital (KO) region in man:an fMRI study. Cereb. Cortex 7, 690–701
30 Zeki, S. et al. (2002) The processing of kinetic contours in the brain.Cereb. Cortex in press
31 Smith, A.T. et al. (1998) The processing of first- and second-ordermotion in human visual cortex assessed by functional magneticresonance imaging (fMRI). J. Neurosci. 18, 3816–3830
Do you want to reproduce material from a Trends journal?This publication and the individual contributions within it are protected by the copyright of Elsevier Science. Except as outlined inthe terms and conditions (see p. ii), no part of any Trends journal can be reproduced, either in print or electronic form, withoutwritten permission from Elsevier Science. Please address any permission requests to:
Rights and Permissions,Elsevier Science Ltd,
PO Box 800, Oxford, UK OX5 1DX.
Opinion TRENDS in Neurosciences Vol.26 No.1 January 200326
http://tins.trends.com