hydrolysis of phosphoproteins and inositol phosphates by cell surface phosphatase of leishmania...
TRANSCRIPT

Molecular and Biochemical Parasitology, 20 (1986) 143-153 143 Elsevier
MBP 00689
Hydrolysis of phosphoproteins and inositol phosphates by cell surface phosphatase of Leishmania donovani
S i d d h a r t h a D a s 1, A s i s h K. S a h a 1, A l a n T. R e m a l e y 1, R o b e r t H. G l e w 1'*, J o h n N. D o w l i n g 2, M a s a o K a j i y o s h i 3 a n d M i c h a e l G o t t l i e b 4
1Department of Microbiology, Biochemistry and Molecular Biology, 2Department of Medicine, The University of Pittsburgh School of Medicine, Pittsburgh, PA 15261, 3Department of Anatomy and Cell Biology and 4Department of Immunology and
Infectious Diseases, The Johns Hopkins University School of Public Health, Baltimore, MD 21205, U.S.A.
(Received 30 January 1986; accepted 9 April 1986)
Leishmania donovani promastigotes contain intense tartrate-resistant cell surface acid phosphatase (ACP1) which blocks super- oxide anion production by activated human neutrophils [A.T. Remaley et al. (1984) J. Biol. Chem, 259, 11173-11175]. An ex- tensively purified preparation of ACP1 dephosphorylates several phosphoproteins which are phosphorylated at serine residues; these include: pyruvate kinase (Kin 1.6 p.M; Vm~, 71.4 U (mg protein)-1), phosphorylase kinase (Kin 0.076 ~M; V ~ 5.4 U (mg protein) -1) and histones (Km 4.86 p.M; Vm~ 2.2 U (mg protein)-1). However, the specific activity of the leishmanial phosphatase on these phosphoproteins is very low as compared to other phosphoprotein phosphatases. The phosphatase activity of ACP1 was also low on phosphohistone phosphorylated at tyrosine residues. Phosphatidylinositol-4,5-diphosphate (PIP2) and inositoltriphos- phate (IP3) were also tested as ACP~ substrates. PIP2 was hydrolyzed rapidly by ACP1. The rate of hydrolysis of PIP2 was higher at pH 6.8 (Kin 2.35 p,M; Vm~ 107 X 103 U (mg protein) -1) than at pH 5.5 (Kin 4.16 IzM; Vm~x 71 x 103 U (mg protein)-1). 32p. labeled IP3 was also a substrate for ACP~; the hydrolysis products consisted of a mixture of inositoldiphosphate and inositolmon- ophosphate. ACP1 and ten other phosphatases were tested for their ability to dephosphorylate proteins and to inhibit 02- pro- duction by stimulated human neutrophils. There was no correlation between the protein phosphatase activity of the acid- and al- kaline phosphatases and their ability to block neutrophil 02- production. The results indicate that ACP1 probably blocks the production of reduced oxygen intermediates by a mechanism that does not involve dephosphorylation of phosphoproteins; how- ever, the possibility that the parasite's phosphatase affects phagocyte metabolism by degrading PIP2 or IP3 should be considered.
Key words: Leishman~sis; Phosphoinositides; Phosphoproteins; Phosphatase; Phosphatidylinositoldiphosphate
Introduction
Leishmania donovani, the causative agent of Kala-azar, exists in two morphologic forms. In the digestive tract of its insect vector it exists as pro- mastigote, whereas in the phagolysosomal system of phagocytic mononuclear cells the parasite ex- ists in the amastigote form [1].
It was demonstrated using histochemical tech-
* To whom all correspondence should be addressed.
Abbreviations: ACP1, acid phosphatase; FMLP, f-Met-Leu- Phe; MES, 2-(N-morpholino)ethanesulphonic acid; MUP, 4- methylumbelliferyl phosphate; PIP2, phosphatidylinositol-4,5- diphosphate.
niques that promastigotes contain abundant acid phosphatase activity, localized on the outer sur- face of the cell [2]. The tartrate-resistant acid phosphatase (ACP1) responsible for greater than 75% of the cell surface phosphatase activity was recently purified and characterized [3]. Evidence for a possible pathophysiologic role for this en- zyme was provided when Remaley et al. [4,5] demonstrated that a pure preparation of the phosphatase inhibits superoxide anion and H202 production as well as decreases the rate of 02 consumption by FMLP (tripeptide chemoattrac- tant)-stimulated human neutrophils and macro- phages. Superoxide anions and other toxic oxi- gen metabolites derived therefrom (e.g., H202, hydroxyl radicals, and hypochlorite ions), which
0166-6851/86/$03.50 © 1986 Elsevier Science Publishers B.V. (Biomedical Division)

144
are produced by stimulated phagocytic cells, are known to contribute to the killing of microbes. Amastigotes and promastigotes of L. donovani are especially sensitive to H202 [6,7]. In another study, Saha et al. [8] demonstrated that an acid phosphatase from the facultative intracellular mi- croorganism Legionella micdadei also blocked su- peroxide anion production by neutrophils stimu- lated with FMLP.
The mechanism responsible for the acid phos- phatase-mediated suppression of 02- production by phagocytic polymorphonuclear (PMN) cells is not understood. However, the fact that the phos- phatase effect is abolished by inhibitors of the en- zyme's activity together with the report [9] that important macrophage and neutrophil functions are regulated by phosphorylation-dephos- phorylation of specific cell surface proteins sug- gested to us that the leishmanial acid phospha- tase could disarm phagocytic polymorphonuclear cells by a mechanism involving dephosphoryla- tion of critical host cell phosphoproteins and in- ositol phosphates, which are also involved in the regulation of oxidative metabolism in phagocytes [10]. We therefore investigated the ability of ACP1 to dephosphorylate known phosphopro- teins, like phosphohistones, phosphorylated forms of pyruvate kinase, and phosphorylase kinase, as well as phosphatidylinositol-4,5-diphosphate (PIP2) and inositoltriphosphate (IP3).
In addition, we compared the ability of the tar- trate-resistant leishmanial acid phosphatase and a total of 10 other acid- and alkaline phosphatases from various sources to dephosphorylate phos- phoproteins with the ability of the same phospha- tases to block superoxide anion production by neutrophils. The results of our experiments indi- cate that the phosphoprotein phosphatase activ- ity of the ACP1 is extremely low and that it is un- likely the enzyme inhibits neutrophil 02- production through a mechanism involving phos- phoproteins.
Materials and Methods
Materials. 4-Methylumbelliferyl phosphate, phos- phorylase kinase, PIP2, histone, and cyclic AMP- dependent protein kinase (bovine heart), f-Met- Leu-Phe (FMLP), various acid phosphatases (i.e.,
potato, semen, prostate and milk) and alkaline phosphatase from Escherichia coli, bovine pla- centa, and bovine intestine, sodium taurocholate, sodium taurodeoxycholate and Brij 35 were pur- chased from Sigma Chemical Co., St. Louis, MO. AG1-X8 Dowex was purchased from Bio-Rad Laboratories. Zwittergent 3-14 detergent was purchased from Calbiochem-Behring Corp., LaJolla, CA. Purified rat liver pyruvate kinase was a gift from Dr. James Blair, West Virginia University, Morgantown, WV. The polyanionic molybdenum complexes were provided by Dr. Michael T. Pope, Georgetown University, Wash- ington, D.C. [32p]Orthophosphate and [~- 32p]ATP (5-10 Ci mmo1-1) were purchased from New England Nuclear, Boston, MA.
Organism and growth conditions. A cloned strain of L. donovani (Sudan 1) promastigotes was grown and harvested as described by Gottlieb and Dwyer [2], except for the addition of streptomy- cin (50 ~g m1-1) and penicillin (50 U m1-1) to the medium.
The tartrate-resistant surface acid phosphatase (ACPI) was purified as described elsewhere [3]. The acid phosphatase secreted into the growth medium by L. donovani promastigotes was puri- fied to homogeneity as described by Gottlieb and Dwyer [11]; the specific activity of the prepara- tion used in this study was 76 units (mg pro- rein) -1. Two other acid phosphatases, designated ACP2 and ACP 3 (both tartrate-sensitive), were purified extensively from L. donovani mem- branes as described elsewhere [3].
Preparation of neutrophils. Cell suspensions con- taining 98 -+ 1% neutrophils were prepared from human blood by dextran sedimentation followed by density gradient centrifugation on Ficoll/ Hypaque gradients and isotonic NH4CI lysis of erythrocytes [12].
Measurement of superoxide anion production. The generation of superoxide anions by neutrophils was measured with a continuous assay as super- oxide dismutase-inhibitable cytochrome c reduc- tion [13]. A 1-ml suspension of neutrophils con- taining 2.5 x 106 cells was incubated for 30 min at 37°C in the presence of 5 mM glucose in Krebs-

Ringer phosphate buffer (pH 7.4) prior to stim- ulation with 100 nM FMLP. Absorbance values were converted to nanomoles of superoxide an- ion using the extinction coefficient of 0.021 cm -1 ixM -1. The phosphatase preparations used to treat the neutrophils were dialyzed exhaustively against Krebs-Ringer phosphate buffer (pH 7.4).
Preparation of phosphoproteins. Phosphorylation of the serine residues of a pure preparation of py- ruvate kinase was accomplished using the method of Blair et al. [14] with slight modification. The incubation medium contained 0.2 ml of pyruvate kinase (1.8 mg ml-1), 10 mM Tris-HC1 buffer (pH 7.4), 1 mM [~/-32p]ATP (1 x 106 cpm nmol-1), 0.05 mM cAMP, 1 mM MgC12 and 250 p~g pro- tein kinase in a final volume of 0.6 ml. The mix- ture was incubated at 37°C for 30 min. The re- action was terminated by precipitation of protein with 2.5 M ammonium sulfate and centrifugation (10000 x g, 10 min) to remove excess ATP. The preparation was then dissolved in and dialyzed against distilled water and lyophilized. The ly- ophilized powder was dissolved in the desired buffer as specified in the text. An aliquot (0.01 ml) of phosphorylated pyruvate kinase was spotted on a filter paper disc and protein-bound radioactiv- ity was measured according to Corbin and Rei- mann [15]. The phosphorylated pyruvate kinase preparation contained 1.6 mol 32p (mol pro- tein) -1 (assuming Mr = 250 000) and the specific activity was 1 x 106 cpm txmo1-1.
The phosphorylation of phosphorylase kinase was carried out according to the method of Cohen [16] with some modifications. The reaction mix- ture (0.6 ml) contained 400 txg phosphorylase ki- nase, 10 mM Tris-HCl buffer (pH 7.4), 1 mM [~/- 32p]ATP (1 x 106 cpm nmol-X), 0.05 mM cAMP, 1 mM MgC12 and 250 Ixg protein kinase. This mixture was incubated at 37°C for 1 h; the reac- tion was terminated by addition of 2 ml 2.5 M ammonium sulfate and the sample was processed as described for pyruvate kinase. The phospho- rylase kinase preparation contained 0.2 mol 32p (mol protein) -1 (assuming Mr = 318 000) and the specific activity was 1 x 105 cpm ~mo1-1.
Calf thymus histone (type II-S) was phospho- rylated using [~/-32p]ATP [17,18]. The histone preparation contained 0.2 mol 32p (mol pro-
145
tein) -1 (assuming M r = 12000) and had a specific activity of 1 x 105 cpm ~mo1-1.
Assay for protein phosphatase activity. Phospho- protein phosphatase activity was estimated at 37°C in a 0.25 ml incubation mixture containing phos- phoprotein substrate, phosphatase and buffer as specified in the text. The extent of dephosphor- ylation was determined using the filter-disc assay technique described by Corbin and Reimann [15]. One unit of protein phosphatase activity is de- fined as 1 nmol of phosphate released min -1.
The standard acid phosphatase assay. Acid phos- phatase activity was determined fluorometrically at 37°C using 7 mM 4-methylumbelliferyl phos- phate (MUP) as substrate [19]. One unit of en- zyme activity is defined as the amount of enzyme required to convert 1 nmol of substrate to prod- uct min -a at 37°C.
When PIP2 was used as the phosphatase sub- strate, the same assay as that described in the preceding paragraph was employed except that the buffers were varied (as indicated in the text) and phosphate release was estimated using the malachite green-dependent assay of Lanzetta et al. [20]. Because detergents (e.g., Triton X-100) affect the color yield, it was necessary to include detergents in the medium used to construct the phosphate standard curve.
Preparation of inositoltriphosphate. Red cells from freshly drawn human blood (30 ml) were washed three times with Hepes buffer, pH 7.4. Three ml of rejuvenation solution [10] and 20 txl of 32p i (500 ~Ci) were incubated with the red cells for 2 h at 37°C. The cells were centrifuged at 3000 x g for 10 min and then hemolysed with 2 vol of 10 mM Hepes buffer (pH 7.4) containing 1 mM EDTA. After washing three times with Hepes-EDTA buffer, the membranes were washed once more with 10 mM Hepes buffer (pH 7.4) containing 0.15 M NaC1. The pellet was suspended in 5 ml Hepes buffer (pH 7.4), supplemented with 1 mM CaCI 2 to activate phospholipase C [21]. After in- cubation at 37°C for 30 min the membranes were pelleted by centrifugation (100000 x g, 1 h); the water-soluble inositol phosphates in the super- natant were separated on a Dowex AG1-X8 col-
1 4 6
umn (1 × 3 cm) using a slight modification of the method of Emilson and Sundler [22]. The column was eluted with 12 ml 0.1 M formic acid - 0.2 M ammonium formate solution to remove inositol- monophosphate followed by 12 ml 0.1 M formic acid - 0.4 M ammonium formate to elute inosi- toldiphosphates, and finally with 12 ml 0.1 M formic acid - 1.0 M ammonium formate to elute 32p-labeled inositoltriphosphate. Fractions were pooled, desalted using a Dowex-50 column, ly- ophilized and dissolved in distilled water. The in- ositoltriphosphate-containing fractions were re- chromatographed on the same column thereby providing substrate for the leishmanial acid phos- phatase. When subjected to two-dimensional chromatography on polyethyleneimine cellulose [22] followed by autoradiography, only a single radioactive spot was detected, which corre- sponded to the inositoltriphosphate standard (data not shown).
Protein determination. Protein concentration was estimated by the method of Bradford [23] using bovine serum albumin as standard.
Results
Dephosphorylation of phosphoproteins by A CP1. ACP 1 (3.3 mU) at pH 5.5 dephosphorylated 50% or more of the radiolabeled phosphorylase ki- nase, pyruvate kinase or histone present in the assay medium in 10-15 h. The release of 32p was linear with time for up to 5 h. Heat-inactivated (5 min, 100°C) phosphatase did not release any sig- nificant amount of trichloroacetic acid-soluble 32p in 25 h. The decline in the rate after 5 h was not due to inactivation of the enzyme since activity on MUP remained more than 85% of the original activity after 15 h of incubation at 37°C,
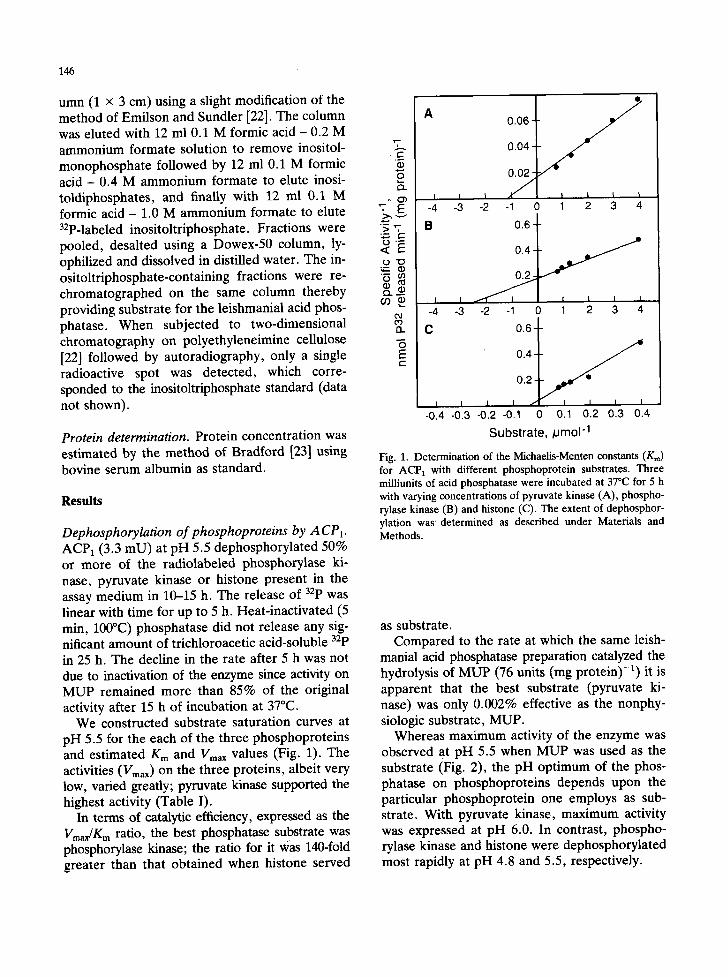
We constructed substrate saturation curves at pH 5.5 for the each of the three phosphoproteins and estimated Km and Vmax values (Fig. 1). The activities (Vm~x) on the three proteins, albeit very low, varied greatly; pyruvate kinase supported the highest activity (Table I).
In terms of catalytic efficiency, expressed as the Vmar/Km ratio, the best phosphatase substrate was phosphorylase kinase; the ratio for it ~as 140-fold greater than that obtained when histone served
> , ".~ e -
0 " ~
cO
-6 E e -
A
I I -4 -3
B
I I -4 -3
C
0.06
0.04-
-2 -1 0 1 2 3 4
0.6-
0.4-- y
I t t I
-2 -I 0 I 2 3 4
0,6-
O.4
0.2-
t I I I i I I I I
-0.4-0,3-0,2-0,1 0 0.1 0.2 0.3 0.4
Substrate, ~umol "1
Fig. 1. Determination of the Michaelis-Menten constants (Kr~) for ACP 1 with different phosphoprotein substrates. Three milliunits of acid phosphatase were incubated at 37°C for 5 h with varying concentrations of pyruvate kinase (A), phospho- rylase kinase (B) and histone (C). The extent of dephosphor- ylation was' determined as described under Materials and Methods.
as substrate. Compared to the rate at which the same leish-
martial acid phosphatase preparation catalyzed the hydrolysis of MUP (76 units (rag protein) -1) it is apparent that the best substrate (pyruvate ki- nase) was only 0.002% effective as the nonphy- siologic substrate, MUP.
Whereas maximum activity of the enzyme was observed at pH 5.5 when MUP was used as the substrate (Fig. 2), the pH optimum of the phos- phatase on phosphoproteins depends upon the particular phosphoprotein one employs as sub- strate. With pyruvate kinase, maximum activity was expressed at pH 6.0. In contrast, phospho- rylase kinase and histone were dephosphorylated most rapidly at pH 4.8 and 5.5, respectively.
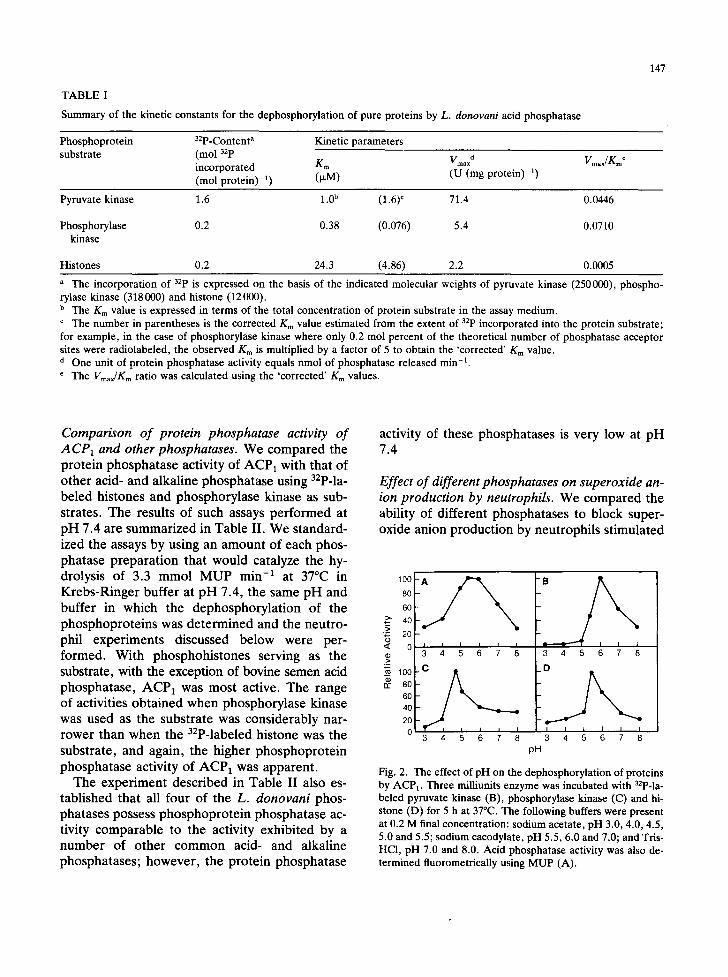
TABLE I
Summary of the kinetic constants for the dephosphorylation of pure proteins by L. donovani acid phosphatase
147
Phosphoprotein 32p-Content a Kinetic parameters substrate (mol 32p
incorporated Km Vmaxd (mol protein) -1) (v,M) (U (mg protein) -1)
Vmax/g~ o
Pyruvate kinase 1.6 1.0 b (1.6) ¢ 71.4 0.0446
Phosphorylase 0.2 0.38 (0.076) 5.4 0.0710 kinase
Histones 0.2 24.3 (4.86) 2.2 0.0005
a The incorporation of 32p is expressed on the basis of the indicated molecular weights of pyruvate kinase (250000), phospho- rylase kinase (318000) and histone (12000). b The K m value is expressed in terms of the total concentration of protein substrate in the assay medium. ¢ The number in parentheses is the corrected Km value estimated from the extent of 32p incorporated into the protein substrate; for example, in the case of phosphorylase kinase where only 0.2 mol percent of the theoretical number of phosphatase acceptor sites were radiolabeled, the observed Km is multiplied by a factor of 5 to obtain the 'corrected' K m value. d One unit of protein phosphatase activity equals nmol of phosphatase released min -1.
The Vmax/K~ ratio was calculated using the 'corrected' Km values.
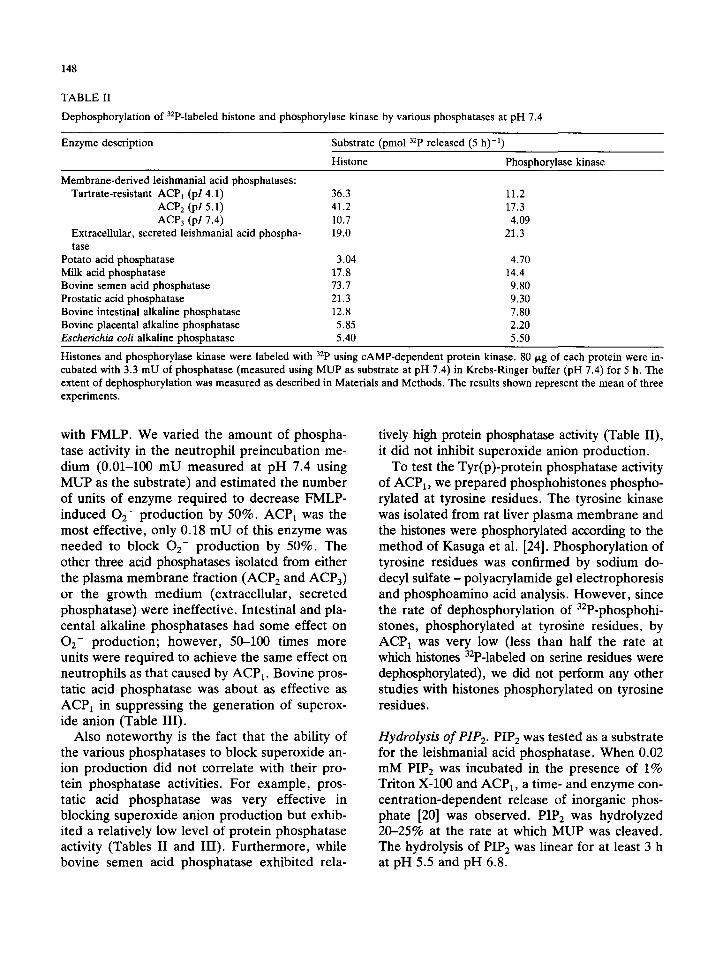
Comparison of protein phosphatase activity of ACP1 and other phosphatases. We compared the protein phosphatase activity of ACP1 with that of other acid- and alkaline phosphatase using 32p-la- beled histones and phosphorylase kinase as sub- strates. The results of such assays performed at pH 7.4 are summarized in Table II. We standard- ized the assays by using an amount of each phos- phatase preparation that would catalyze the hy- drolysis of 3.3 mmol MUP min -1 at 37°C in Krebs-Ringer buffer at pH 7.4, the same pH and buffer in which the dephosphorylation of the phosphoproteins was determined and the neutro- phil experiments discussed below were per- formed. With phosphohistones serving as the substrate, with the exception of bovine semen acid phosphatase, ACP 1 was most active. The range of activities obtained when phosphorylase kinase was used as the substrate was considerably nar- rower than when the 32p-labeled histone was the substrate, and again, the higher phosphoprotein phosphatase activity of ACP1 was apparent.
The experiment described in Table II also es- tablished that all four of the L. donovani phos- phatases possess phosphoprotein phosphatase ac- tivity comparable to the activity exhibited by a number of other common acid- and alkaline phosphatases; however, the protein phosphatase
activity of these phosphatases is very low at pH 7.4
Effect of different phosphatases on superoxide an- ion production by neutrophils. We compared the ability of different phosphatases to block super- oxide anion production by neutrophils stimulated
>" 4 0 ,_> ~6 20
~> 3 4 5 6 7 8 3 4 5 6 7 8
,'~ 100 _C or 8 0 -
6O
4 0
2 0 I I I I I I
0 3 4 5 6 7 8 3 4 ~ 6 7 8
pH
Fig. 2. The effect of pH on the dephosphorylation of proteins by ACP~. Three milliunits enzyme was incubated with 32p-la- beled pyruvate kinase (B), phosphorylase kinase (C) and hi- stone (D) for 5 h at 37°C. The following buffers were present at 0.2 M final concentration: sodium acetate, pH 3.0, 4.0, 4.5, 5.0 and 5.5; sodium cacodylate, pH 5.5, 6.0 and 7.0; and Tris- HC1, pH 7.0 and 8.0. Acid phosphatase activity was also de- termined fluorometrically using MUP (A).
148
TABLE II
Dephosphorylation of 32p-labeled histone and phosphorylase kinase by various phosphatases at pH 7.4
Enzyme description Substrate (pmol 32p released (5 h) -1) Histone Phosphorylase kinase
Membrane-derived leishmanial acid phosphatases: Tartrate-resistant ACP1 (pl 4.1) 36.3
ACP 2 (pl 5.1) 41.2 ACP 3 (pl 7.4) 10.7
Extracellular, secreted leishmanial acid phospha- 19.0 tase
Potato acid phosphatase 3.04 Milk acid phosphatase 17.8 Bovine semen acid phosphatase 73.7 Prostatic acid phosphatase 21.3 Bovine intestinal alkaline phosphatase 12.8 Bovine placental alkaline phosphatase 5.85 Escherichia coli alkaline phosphatase 5.40
11.2 17.3 4.09
21.3
4.70 14.4 9.80 9.30 7.80 2.20 5.50
Histones and phosphorylase kinase were labeled with 32p using cAMP-dependent protein kinase. 80 ~.g of each protein were in- cubated with 3.3 mU of phosphatase (measured using MUP as substrate at pH 7.4) in Krebs-Ringer buffer (pH 7.4) for 5 h. The extent of dephosphorylation was measured as described in Materials and Methods. The results shown represent the mean of three experiments.
with FMLP. We varied the amount of phospha- tase activity in the neutrophil preincubation me- dium (0.01-100 mU measured at pH 7.4 using MUP as the substrate) and estimated the number of units of enzyme required to decrease FMLP- induced OE- production by 50%. ACP1 was the most effective, only 0.18 mU of this enzyme was needed to block O2- production by 50%. The other three acid phosphatases isolated from either the plasma membrane fraction (ACP 2 and ACP3) or the growth medium (extracellular, secreted phosphatase) were ineffective. Intestinal and pla- cental alkaline phosphatases had some effect on O2- production; however, 50-100 times more units were required to achieve the same effect on neutrophils as that caused by ACP 1. Bovine pros- tatic acid phosphatase was about as effective as ACP1 in suppressing the generation of superox- ide anion (Table III).
Also noteworthy is the fact that the ability of the various phosphatases to block superoxide an- ion production did not correlate with their pro- tein phosphatase activities. For example, pros- tatic acid phosphatase was very effective in blocking superoxide anion production but exhib- ited a relatively low level of protein phosphatase activity (Tables II and III). Furthermore, while bovine semen acid phosphatase exhibited rela-
tively high protein phosphatase activity (Table II), it did not inhibit superoxide anion production.
To test the Tyr(p)-protein phosphatase activity of ACP1, we prepared phosphohistones phospho- rylated at tyrosine residues. The tyrosine kinase was isolated from rat liver plasma membrane and the histones were phosphorylated according to the method of Kasuga et al. [24]. Phosphorylation of tyrosine residues was confirmed by sodium do- decyl sulfate - polyacrylamide gel electrophoresis and phosphoamino acid analysis. However , since the rate of dephosphorylation of 32p-phosphohi- stones, phosphorylated at tyrosine residues, by ACP1 was very low (less than half the rate at which histones 32p-labeled on serine residues were dephosphorylated), we did not perform any other studies with histones phosphorylated on tyrosine residues.
Hydrolysis of PIPE. PIP2 was tested as a substrate for the leishmanial acid phosphatase. When 0.02 mM PIPE was incubated in the presence of 1% Triton X-100 and ACP1, a time- and enzyme con- centration-dependent release of inorganic phos- phate [20] was observed. PIP2 was hydrolyzed 20-25% at the rate at which MUP was cleaved. The hydrolysis of PIP 2 was linear for at least 3 h at pH 5.5 and pH 6.8.
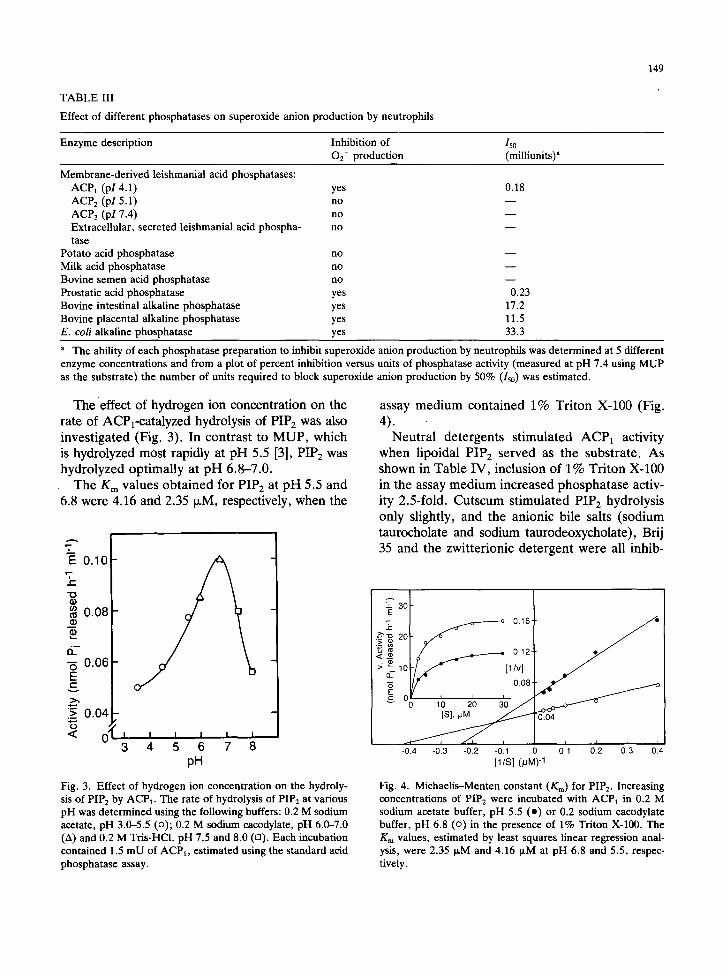
TABLE III
Effect of different phosphatases on superoxide anion production by neutrophils
149
Enzyme description Inhibition of 150 02- production (milliunits) a
Membrane-derived leishmanial acid phosphatases: ACPl (pl 4.1) yes ACP 2 (pl 5.1) no ACP3 (pI 7.4) no Extracellular, secreted leishmanial acid phospha- no tase
Potato acid phosphatase no Milk acid phosphatase no Bovine semen acid phosphatase no Prostatic acid phosphatase yes Bovine intestinal alkaline phosphatase yes Bovine placental alkaline phosphatase yes E. coli alkaline phosphatase yes
0.18
m
0.23 17.2 11.5 33.3
a The ability of each phosphatase preparation to inhibit superoxide anion production by neutrophils was determined at 5 different enzyme concentrations and from a plot of percent inhibition versus units of phosphatase activity (measured at pH 7.4 using MUP as the substrate) the number of units required to block superoxide anion production by 50% (150) was estimated.
The effect of hydrogen ion concentration on the rate of ACPx-catalyzed hydrolysis of PIPE was also investigated (Fig. 3). In contrast to MUP, which is hydrolyzed most rapidly at pH 5.5 [3], PIP 2 was hydrolyzed optimally at pH 6.8-7.0.
The K m values obtained for PIP2 at pH 5.5 and 6.8 were 4.16 and 2.35 ixM, respectively, when the
2--
E 0.10 v--
i
" '0
o.o8
-5 0.06 E t -
> 0.04 0
< O; I I I I I I 3 4 5 6 7 8
pH
assay medium contained 1% Triton X-100 (Fig. 4). Neutral detergents stimulated ACP1 activity
when lipoidal PIP 2 served as the substrate. As shown in Table IV, inclusion of 1% Triton X-100 in the assay medium increased phosphatase activ- ity 2.5-fold. Cutscum stimulated PIP 2 hydrolysis only slightly, and the anionic bile salts (sodium taurocholate and sodium taurodeoxycholate), Brij 35 and the zwitterionic detergent were all inhib-
~ ' ~ 016 .~ l / . i f - 0.12
" '~ lo [l/v] -6 0 . 0 8 -
v 0 10 , 20 30 / .
-0 .4 -0 .3 -0 .2 -0.1 0 0.1 0 .2 0 .3 0 .4
[ l /S] (pM) -1
Fig. 3. Effect of hydrogen ion concentration on the hydroly- sis of PIP2 by ACP1. The rate of hydrolysis of PIP2 at various pH was determined using the following buffers: 0.2 M sodium acetate, pH 3.0-5.5 (o); 0.2 M sodium cacodylate, pH 6.0-7.0 (A) and 0.2 M Tris-HCl. pH 7.5 and 8.0 (n). Each incubation contained 1.5 mU of ACPI, estimated using the standard acid phosphatase assay.
Fig. 4. Michaelis-Menten constant (Kin) for PIP2. Increasing concentrations of PIP 2 were incubated with ACP 1 in 0.2 M sodium acetate buffer, pH 5.5 (o) or 0.2 sodium cacodylate buffer, pH 6.8 (o) in the presence of 1% Triton X-100. The Km values, estimated by least squares linear regression anal- ysis, were 2.35 ~,M and 4.16 ~M at pH 6.8 and 5.5, respec- tively.
150
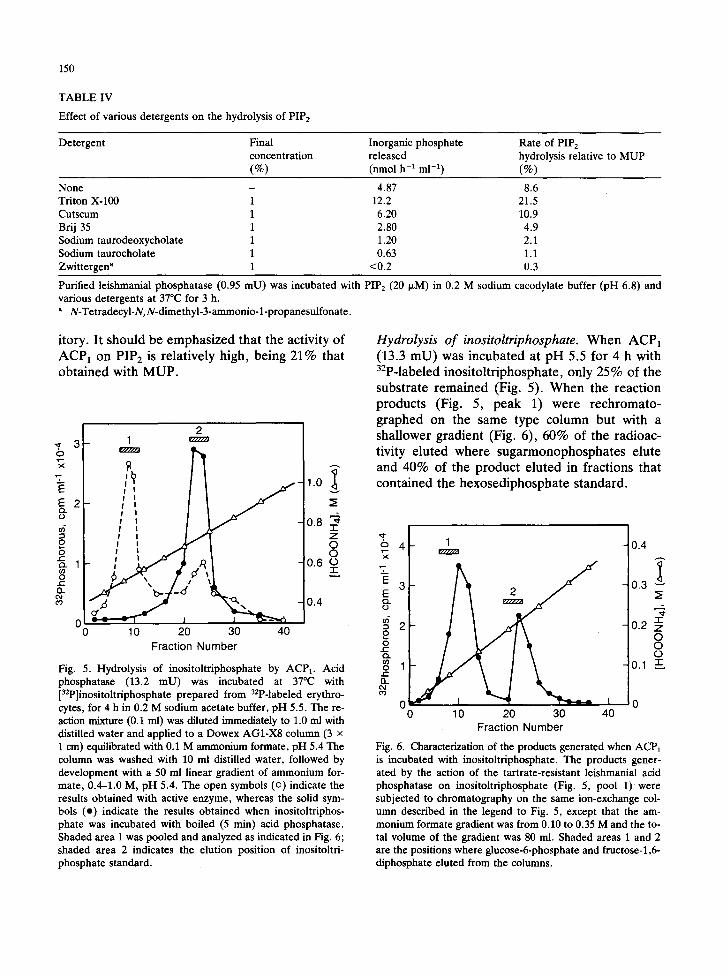
TABLE IV
Effect of various detergents on the hydrolysis of PIPs
Detergent Final Inorganic phosphate Rate of PIP 2 concentration released hydrolysis relative to MUP (%) (nmoi h -1 m1-1) (%)
None - 4.87 8.6 Triton X-100 1 12.2 21.5 Cutscum 1 6.20 10.9 Brij 35 1 2.80 4.9 Sodium taurodeoxycholate 1 1.20 2.1 Sodium taurocholate 1 0.63 1.1 Zwittergen a 1 <0.2 0.3
Purified leishmanial phosphatase (0.95 mU) was incubated with PIP 2 (20 I~M) in 0.2 M sodium cacodylate buffer (pH 6.8) and various detergents at 37°C for 3 h.
N-Tetradecyl-N,N-dimethyl-3-ammonio- l-propanesulfonate.
itory. It should be emphas ized that the activity of
A C P 1 on PIP 2 is relat ively high, being 21% that ob ta ined with M U P .
O
E E 0
£ 0 ¢ -
0 e-" n
2 3 - 1
v / / / / / ~
- 1 . 0
2 ~
0.8 :~ z 0 0
1 0.6 o =:
, 0 . 4
0 10 20 30 40 Fraction Number
Fig. 5. Hydrolysis of inositoltriphosphate by ACPI. Acid phosphatase (13.2 mU) was incubated at 37°C with [32p]inositoltriphosphate prepared from 32P-labeled erythro- cytes, for 4 h in 0.2 M sodium acetate buffer, pH 5.5. The re- action mixture (0.1 ml) was diluted immediately to 1.0 ml with distilled water and applied to a Dowex AG1-X8 column (3 x 1 cm) equilibrated with 0.1 M ammonium formate, pH 5.4 The column was washed with 10 ml distilled water, followed by development with a 50 ml linear gradient of ammonium for- mate, 0.4-1.0 M, pH 5.4. The open symbols (o) indicate the results obtained with active enzyme, whereas the solid sym- bols (o) indicate the results obtained when inositoltriphos- phate was incubated with boiled (5 min) acid phosphatase. Shaded area 1 was pooled and analyzed as indicated in Fig. 6; shaded area 2 indicates the elution position of inositoltri- phosphate standard.
Hydrolysis of inositoltriphosphate. W h e n ACP1 (13.3 m U ) was incuba ted at p H 5.5 for 4 h with 32p-labeled inos i to l t r iphosphate , only 25% of the
substrate r ema ined (Fig. 5). W h e n the react ion products (Fig. 5, peak 1) were rechromato-
graphed on the same type co lumn but with a shallower gradient (Fig. 6), 60% of the radioac- tivity e luted where suga rmonophospha te s elute and 40% of the product e lu ted in fract ions that con ta ined the hexosediphosphate s tandard .
o 4 I ~-~ r H H Z A
E 3 E Q .
0 ¢.- ,
o ] Q..
0 ~ 0 0 10 20 30 40
Fraction Number
Fig. 6. Characterization of the products generated when ACP1 is incubated with inositoltriphosphate. The products gener- ated by the action of the tartrate-resistant leishmanial acid phosphatase on inositoltriphosphate (Fig. 5, pool 1)were subjected to chromatography on the same ion-exchange col- umn described in the legend to Fig. 5, except that the am- monium formate gradient was from 0.10 to 0.35 M and the to- tal volume of the gradient was 80 ml. Shaded areas 1 and 2 are the positions where glucose-6-phosphate and fructose-l,6- diphosphate eluted from the columns,
0.4
0.3 ~
,._T,
'1- 0.2 z
0 0 0
0.1 &

These results suggest that ACP1 utilizes both inositoltriphosphate and inositoldiphosphate as substrates. However, because the specific radio- activity and concentration of the inositoltriphos- phate in the assay medium were unknown we could not express our results in terms of specific activity or compare the rate of hydrolysis of in- ositoltriphosphate with that of other substrates such as PIP2 or MUP.
Discussion
In neutrophils and macrophages phagocytosis induces a respiratory burst which leads to the production of microbicidal reactive oxygen me- tabolites (e.g. O2-, H202). Several reports [9,25] indicate that the activation of superoxide anion production by membrane-perturbing agents is ac- companied by the phosphorylation of membrane proteins. Our results argue against the involve- ment of phosphoproteins in the mechanism whereby the cell surface acid phosphatase of L. donovani inhibits the ability of phagocytes to generate superoxide anions. First, the absolute specific activity of the phosphatase on well-char- acterized phosphoproteins radiolabeled on serine residues is extremely low, being at least five or- ders of magnitude below that on low molecular weight phosphomonoester substrates such as MUP and various phosphorylated monosacchar- ides [3]. In addition, 32p-labeled histones that were radiolabeled with [~/32-p]ATP on tyrosine residues using a tyrosine-specific protein kinase [24] were not dephosphorylated by the leishman- ial phosphatase (data not shown). Although the phosphoproteins used in our study were poor substrates for ACP1 it is possible that a natural phosphoprotein on phagocytic cells might be a good substrate for the leishmanial phosphatase. To explore this possibility we radiolabeled plasma membranes from intact neutrophils with [32p]orthophosphate. Although more than two dozen radiolabeled proteins could be discerned by sodium dodecyl sulfate - polyacrylamide gel elec- trophoresis, none of these phosphoproteins was dephosphorylated by the leishmanial phospha- tase (data not shown).
The second observation which argues against dephosphorylation of phagocyte phosphoproteins
151
as playing an important role in disarming host cells is the lack of a correlation between the phospho- protein phosphatase activity of a number of acid- and alkaline phosphatases and the ability of these phosphatases to block FMLP-stimulated super- oxide anion production by neutrophils (Tables II and III). For example, of the phosphatases tested, milk acid phosphatase was about as effective as the prostatic phosphatase in dephosphorylating histones and phosphorylase kinase (Table II), yet the milk and the prostatic phosphatase had very different effects on the rate of production of su- peroxide anions by stimulated neutrophils (Table III).
NADPH oxidase is the principal plasma mem- brane enzyme responsible for the production of superoxide anions in phagocytic cells. The en- zyme is relatively inactive in resting cells and be- comes activated by various stimulants such as FMLP or phorbol myristate acetate. Although the mechanism of activation of NADPH oxidase is not well understood, it is known that calcium and the products of the phospholipase C - PIP2 reaction are involved in the process [26--28]. The very low protein phosphatase activity of ACP1 raises the possibility that phosphorylated substances other than phosphoproteins on the surface of neutro- phils may be the substrates of pathophysiologic significance in the phosphatase-mediated inhibi- tion of superoxide anions by phagocytes.
Formylated peptide-receptor-stimulated hy- drolysis of PIP 2 to generate inositoltriphosphate and diglyceride appears to be an important event in phagocyte activation. Evidence showing that inositoltriphosphate is a key mediator of intra- cellular events that lead to neutrophil and mac- rophage activation is accumulating [22]. Re- cently, Smith et al. [29] showed that the formylated peptide receptor in neutrophil mem- branes is coupled to inositoltriphosphate hydrol- ysis through a guanine nucleotide-dependent reg- ulatory protein. In the present study we demonstrate that ACP1 is capable of dephos- phorylating inositoltriphosphate (Fig. 5). Since inositoltriphosphate is a second messenger in- volved in phagocyte activation, the breakdown of this polyphosphoinositide by L. donovani acid phosphatase might compromise the host cell re- sponse to infection.

152
A l t e r n a t i v e l y , the fact tha t P IP 2 is a good sub- s t ra te for the l e i shman ia l p h o s p h a t a s e ra ises the poss ib i l i ty tha t the e n z y m e cou ld c o m p r o m i s e the host cel l ' s r e sponse to the pa ra s i t e by des t roy ing PIP2, the p r o g e n i t o r o f the s econd messenge r , in- o s i t o l t r i phospha t e . I t r e m a i n s to be seen if t he re is sufficient P IP 2 on the cell surface of phago- cytes, or , if not , w h e t h e r the p h o s p h a t a s e on the pa ra s i t e ' s sur face has access to and d e g r a d e s PIP2 conf ined to the i nne r leaf le t of the p l a s m a m e m - b rane . In t e re s t ing ly , the p H o p t i m u m of ACP1 acting on PIP2 is near neutral i ty, p resumably close
to the pH that exists at the host-parasite inter- face.
Acknowledgements
This w o r k was s u p p o r t e d by grants A I 18945 and A I 17047 f rom the N a t i o n a l Ins t i tu tes of H e a l t h , U n i t e d Sta tes Publ ic H e a l t h Service . W e are gra teful to Dr . J ames Bla i r , D e p a r t m e n t of B iochemis t ry , W e s t Vi rg in ia Unive r s i ty , M o r g a n - town, W V , for gene rous ly p rov id ing us wi th py- ruva te k inase .
References
1 Killick-Kendrick, R. (1979) Biology of Leishmania in phlebotomine sandflies. In: Biology of the Kinetoplastida (Lunsden W.H.R., and Evans D.A., eds.), Vol. 2, pp. 395--460, Academic Press, New York.
2 Gottlieb, M. and Dwyer, D.M. (1981) Leishmania dono- vani: Surface membrane acid phosphatase activity of pro- mastigotes. Exp. Parasitol. 52, 117-128.
3 Remaley, A.T., Das, S., Campbell, P.I., LaRocca, G.M., Pope, M.T. and Glew, R.H. (1985) Characterization of Leishmania donovani acid phosphatases. J. Biol. Chem. 260, 880-886.
4 Remaley, A.T., Kuhns, D.B., Basford, R.E., Glew, R.H. and Kaplan, S.S. (1984) Leishmanial phosphatase blocks neutrophil Oz- production, J. Biol. Chem. 259, 11173-11175.
5 Remaley, A.T., Glew, R.H., Kuhns, D.B., Basford, R.E., Waggoner, A.S., Ernst, L.A. and Pope, M. (1985) Leish- mania donovani: Surface membrane acid phosphatase blocks neutrophils oxidative metabolite production. Exp. Parasitol. 60, 331-341.
6 Nogueira, N. (1984) Intracellular mechanisms of killing. In: Contemporary Topics in Immunobiology: Immuno- biology of Parasites and Parasitic Infections (Marchalonis, J.J., ed.), Vol. 12, pp. 53-66, Plenum Press, New York.
7 Saha, A.K., Das., S., Glew, R.H. and Gottlieb, M. (1985) Resistance of leishmanial phosphatases to inactivation by oxygen metabolites. J. Clin. Microbiol. 22, 329-332.
8 Saha, A.K., Dowling, J.N., LaMarco, K.L., Das, S., Re- maley, A.T., Olomu, N., Pope, M.T. and Glew, R.H. (1985) Properties of an acid phosphatase from Legionella micdadei which blocks superoxide anion production by human neutrophils. Arch. Biochem. Biophys. 243, 150-160.
9 Amano, F., Kitagawa, T. and Akamatsu, Y. (1984) Pro- tein kinase activity on the cell surface of a macrophage-like line, J 774.1. Biochim. Biophys. Acta 803, 163-173.
10 Barridge, M.J. (1984) Inositol triphosphate and diacylgly- cerol as second messenger. Biochem. J. 220, 345-360.
11 Gottlieb, M. and Dwyer, D.M. (1982) Identification and partial characterization of an extracellular acid phospha- tase activity of Leishmania donovani promastigotes. Mol.
Cell. Biol. 2, 76--81. 12 Boyum, A. (1968) Isolation of mononuclear cells and
granulocytes from human blood. Scand. J. Clin. Lab. In- vest. 21 (Suppl. 97), 77-89.
13 Newburger, P.E., Chovaniec, M.E. and Cohen, H.J. (1980) Activity and activation of the granulocyte superoxide gen- erating system. Blood 55, 85-92.
14 Blair, J.B., Cimbala, M.A. and James, M.E. (1982) He- patic pyruvate kinase: Quantitative measurements of phosphorylation in vitro and in the isolated rat hepato- cyte. J. Biol. Chem. 257, 7595-7602.
15 Corbin, J.D. and Reimann, E.M. (1974) Assay of cyclic AMP-dependent protein kinases. Methods Enzymol. 38, 287-290.
16 Cohen, P. (1973) The subunit structure of rabbit-skeletal muscle phosphorylase kinase, and the molecular basis of its activation reaction. Eur. J. Biochem. 34, 1-16.
17 Robinson, D.B. and Glew, R.H. (1981) Substrate speci- ficity of Gaucher spleen phosphoprotein phosphatase. Arch. Biochem. Biophys. 210, 186-199.
18 Chou, C.K., Alfano, J. and Rosen, O.M. (1977) Purifi- cation of phosphoprotein phosphatase from bovine car- diac muscle that catalyzes dephosphorylation of cyclic AMP-binding protein component of protein kinase. J. Biol. Chem. 252, 2855-2859.
19 Robinson, D.B. and Glew, R.H. (1980) A tartrate-resist- ant acid phosphatase from Gaucher spleen. J. Biol. Chem 255, 5864-5870.
20 Lanzetta, P.A., Alvarez, L.J., Reinach, P.S. and Candia, O.A. (1979) An improved assay for nanomole amounts of inorganic phosphate. Anal. Biochem 100, 95-97.
21 Shukla, S.D., Coleman, R., Finean, J.B. and MicheU, R.H. (1978) The use of phospholipase C to detect structural changes in the membranes of human erythrocyte aged by storage. Biochim. Biophys. Acta 512, 341-349.
22 Emilson, A. and Sundler, R. (1984) Differential activa- tion of phosphatidylinositol deacylation and pathway via diphosphoinositide in macrophages responding to Zymo- san and ionophore A23187. J. Biol. Chem. 259, 3111-3116.
23 Bradford, M.M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utiliz-

ing the principle of protein-dye binding. Anal. Biochem. 72, 248--256.
24 Kasuga, M., Zick, Y., Blithe, D.L., Crettaz, M. and Kahn, R.C. (1982) Insulin stimulates tyrosine phosphorylation of the insulin receptor in a cell-free system. Nature 298, 667-669.
25 Okamura, N., Ohashi, S., Nagahisa, N. and Ishibashi, S. (1984) Changes in protein phosphorylation in guinea pig polymorpho-nuclear leukocytes by treatment with mem- brane perturbing agents which stimulate superoxide anion production. Arch. Biochem. Biophys. 288, 270-277.
26 Jones, H.P., Ghai, G., Petrone, W.F. and McCord, J.M. (1982) Calcium-dependent stimulation of the NADPH ox- idase of human neutrophils. Biochim. Biophys. Acta 714, 152-156.
153
27 Matsumoto, T., Takeshige, K. and Minakami, S. (1979) Inhibition of phagocytotic metabolic changes of leuko- cytes by an intracellular calcium antagonist 8-(N,N-dieth- ylamino)-octyl-3,4,5-trimethoxybenzoate. Biochem. Bio- phys. Res. Commun. 88, 974-979.
28 Smolen, J.E. and Weissmann, G. (1980) Effects of indo- methacin, 5,8,11,-14-eicosatetraynoic acid and p-bromo- phenacyl bromide on lysosomai enzyme release and su- peroxide anion generation by human polymorphonuclear leukocytes. Biochem. Pharmacol. 29, 533-538.
29 Smith, C.D., Land, B.C., Kusaka, I., Nerghese, M.W. and Snyderman, R.L. (1985) Chemoattractant receptor in- duced hydrolysis of phosphatidylinositol-4,5-bisphosphate in human polymorphonuclear leukocyte membranes. J. Biol. Chem. 260, 5875-5878.