gene relocations within chloroplast genomes of jasminum and
TRANSCRIPT

Gene Relocations within Chloroplast Genomes of Jasminum and Menodora(Oleaceae) Are Due to Multiple, Overlapping Inversions
Hae-Lim Lee,* Robert K. Jansen,� Timothy W. Chumley,� and Ki-Joong Kim**School of Life Sciences and Biotechnology, Korea University, Seoul, Korea; and �Section of Integrative Biology andInstitute of Cellular and Molecular Biology, University of Texas, Austin
The chloroplast (cp) DNA sequence of Jasminum nudiflorum (Oleaceae–Jasmineae) is completed and compared with thelarge single-copy region sequences from 6 related species. The cp genomes of the tribe Jasmineae (Jasminum and Me-nodora) show several distinctive rearrangements, including inversions, gene duplications, insertions, inverted repeat ex-pansions, and gene and intron losses. The ycf4–psaI region in Jasminum section Primulina was relocated as a result of 2overlapping inversions of 21,169 and 18,414 bp. The 1st, larger inversion is shared by all members of the Jasmineaeindicating that it occurred in the common ancestor of the tribe. Similar rearrangements were also identified in the cpgenome of Menodora. In this case, 2 fragments including ycf4 and rps4–trnS–ycf3 genes were moved by 2 additionalinversions of 14 and 59 kb that are unique to Menodora. Other rearrangements in the Oleaceae are confined to certainregions of the Jasminum and Menodora cp genomes, including the presence of highly repeated sequences and duplicationsof coding and noncoding sequences that are inserted into clpP and between rbcL and psaI. These insertions are correlatedwith the loss of 2 introns in clpP and a serial loss of segments of accD. The loss of the accD gene and clpP introns in boththe monocot family Poaceae and the eudicot family Oleaceae are clearly independent evolutionary events. However, theirgenome organization is surprisingly similar despite the distant relationship of these 2 angiosperm families.
Introduction
Chloroplast (cp) genome organization in land plants ishighly conserved (Palmer 1991; Raubeson and Jansen2005). The presence of 2 large inverted repeats (IRs), whichrange from 12 to 75 kb in length and separate the large andsmall single-copy (LSC and SSC) regions, is one of themost prominent features of these genomes (Palmer 1990,1991). Gene order and gene/intron content are typicallyconserved in the majority of land plants; rearrangementshave been documented in a number of groups, includingAsteraceae, Campanulaceae, Fabaceae, Geraniaceae, Lobe-liaceae, Onagraceae, Pinaceae, Poaceae, and Ranuncula-ceae (Palmer and Thompson 1981; Jansen and Palmer1987; Palmer, Nugent, and Herbon 1987; Palmer, Osorio,et al. 1987; Hiratsuka et al. 1989; Milligan et al. 1989;Hachtel et al. 1991; Lidholm and Gustafsson 1991; Doyleet al. 1992; Knox et al. 1993; Hoot and Palmer 1994;Cosner et al. 1997, 2004; Strauss et al. 1998; Kim et al.2005; Saski et al. 2005; Chumley et al. 2006). Several mecha-nisms have been implicated in generating changes in geneorder and content in cp genomes, including inversion, trans-position, gene duplication, gene/intron loss, insertion/deletion, and IR expansion/contraction (Palmer 1991;Raubeson and Jansen 2005). The relative frequency of eachof these changes varies with transposition being the leastcommon, and the only definitive case of cp transpositionhas been documented in the green alga Chlamydomonas(Fan et al. 1995), although this mechanism has been sug-gested in several other groups (Milligan et al. 1989; Cosneret al. 1997). Inversions are the most common mechanismfor changes in gene order, and they have been fully char-acterized in a number of cp genomes. In several cases, dis-persed repeats have been shown to promote inversions
through intermolecular recombination (Ogihara et al.1988; Palmer 1991). Several studies have demonstrateda strong correlation between the numbers and locations of re-petitive sequences and the incidence of inversions (Pombertet al. 2005, 2006). The rarity of gene-order changes in cp ge-nomes has made these events powerful phylogenetic markersin several plant groups (Jansen and Palmer 1987; Doyle et al.1992, 1996; Raubeson and Jansen 1992).
The angiosperm family Oleaceae includes approxi-mately 29 genera and 600 species that are widely distrib-uted in tropical and temperate regions of the world(Heywood 1978). The tribe Jasmineae contains only 2 gen-era, Jasminum and Menodora (Taylor 1945; Johnson 1957;Wallander and Albert 2000). Jasminum is the largest genusof the Oleaceae with approximately 200 species (Rehder1940; Green 2004) confined to the Old World. The genusis traditionally classified into 4 sections (Alternifolia, Uni-foliolata, Pinnatifolia, and Trifoliolata) based on the leafarrangement and the leaflet number (De Candolle 1844);however, several systematic studies have suggested thatmost of these sections are artificial (Rohwer 1994, 1995;Green 1997, 2001). Molecular phylogenies (Lee H-L andKim K-J, unpublished data) support the recognition of3 clades: an alternate-leaved group (section Alternifolia),section Primulina (Green 2001), and an opposite-leavedgroup that contains the majority of the species. In contrast,Menodora is a small genus with approximately 23 species(Steyermark 1932; Turner 1991) and is distributed in south-western North America, southern South America, andsouthern Africa.
In this paper, we investigate the organization and evo-lution of cp genomes in the Oleaceae. Several unusual fea-tures of the genomes of Menodora and Jasminum aredescribed, which provide valuable insights into cpDNAevolution. The comparisons identify multiple inversions,gene duplications, gene and intron losses, IR expansion,and a high incidence of dispersed repeats. Gene relocationcaused by multiple inversions is a notable feature of thecp genomes of the Jasmineae. The phylogenetic orderand timing of the major rearrangements are determined
Key words: chloroplast genome, inversions, gene duplications, inser-tions, inverted repeat expansions, gene and intron losses, Jasminum,Menodora.
E-mail: [email protected].
Mol. Biol. Evol. 24(5):1161–1180. 2007doi:10.1093/molbev/msm036Advance Access publication February 28, 2007
� 2007 The AuthorsThis is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/2.0/uk/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

by comparative sequence analysis and phylogenetic analy-sis of the major lineages within the tribe.
Materials and MethodsTaxon Sampling
The taxa used in this study are listed in table 1. The cpgenome of Jasminum nudiflorum was sequenced com-pletely, whereas inversion regions only were sequencedfrom the following 5 species: Menodora longiflora; 1 spe-cies from each of the 3 major lineages of Jasminum(Jasminum le-ratii, Jasminum abyssinicum, Jasminumsubhumile); and an outgroup species, Forsythia europaea.Seventeen species of Jasminum and 2 species of Menodorawere used for comparative gene mapping. Twenty-five ac-cessions of 24 species of Jasminum, 4 species of Menodora,and 21 species of other Oleaceae were used for the poly-merase chain reaction (PCR) diagnosis of inversions andthe comparative analysis of inversion endpoints. Forty-one ndhF and rbcL sequences representing all major cladesof Oleaceae and outgroups were also used for estimatingphylogenetic relationships and divergence times (table 1).
DNA Extraction, Enzyme Digestion, Filter Hybridization,and Restriction Site Mapping
Total DNA was isolated from the fresh leaves usingthe hexadecyltrimethylammonium bromide method (DoyleJJ and Doyle JA 1987), and DNA was further purified byultracentrifugation in cesium chloride/ethidium bromidegradients. Purified total DNA was digested with 22 restric-tion endonucleases (AvaI, AvaII, AseI, BamHI, BanI, BanII,BclI, BglII, BstNI, BstXI, ClaI, DraI, EcoRI, Eco0109I,EcoRV, HaeII, HincII, HindIII, NciI, NsiI, XbaI, andXmnI), and the resulting fragments were separated on 1–1.6% agarose gels depending on the number of expectedrestriction fragments. Bidirectional transfer of the DNAfrom agarose gels to Zetabind nylon filters (AMF Cuno,Meriden, CT), radioactive labeling of 43 cloned tobaccocpDNA probes (Olmstead and Palmer 1992), filter hybrid-ization, and autoradiography were performed as describedin Palmer (1986). In addition, DNA from Olea europaea,Menodora scabra, Jasminum humile, and J. nudiflorumwere digested with the same 22 enzymes, transferred tonylon membranes, and hybridized to 106 small cpDNAprobes from tobacco (Palmer et al. 1994) to constructdetailed gene and restriction site maps.
Genome Sequencing and Annotation
For the complete sequencing of the J. nudiflorum cpgenome, cpDNA was isolated and purified following themethods in Palmer (1986). The cpDNA was digested witha combination of 3 restriction enzymes (BamHI, SacI, andClaI), and the resulting fragments were cloned into a pBlue-script II vector. Inserted cpDNA fragments were shotgunsequenced using the BigDye3.0 terminal cycle sequenc-ing kit (Applied Biosystems, Foster City, CA) and an ABI3700 sequencer. Sequences were assembled using Se-quencher (version 4.1; Gene Codes Corporation, Ann Arbor,MI). In order to fill gap regions, primers were designed
using the complete cp sequences of Nicotiana and Panax(Shinozaki et al. 1986; Kim and Lee 2004).
A series of PCR products spanning the entire regioncontaining the inversions were generated for J. le-ratii,J. abyssinicum, J. subhumile, M. longiflora, and F. euro-paea. The PCR cycle was as follows: 94 �C for 3 min, then30 cycles of 94 �C for 30 s, 50–68 �C for 1 min, and 68–72�C for 1–8 min (depending upon the size of the target re-gion). PCR-amplified DNA was then purified using theQIAquick PCR purification kit (Qiagen, Hilden, Germany).Amplified cpDNA fragments were sequenced directly.
Gene annotations and comparative sequence analyseswere performed using the current versions of Blast (Altschulet al. 1990), ORF finder programs from National Center forBiotechnology Information, and DOGMA (Wyman et al.2004). The locations and secondary structures of tRNAgenes were determined using tRNAscan-SE version 1.21(Lowe and Eddy 1997), DOGMA (Wyman et al. 2004),and MFOLD version 3.0 (Zuker 2003). Repeated sequenceswere identified using REPuter (Kurtz et al. 2001).
PCR Diagnosis of Inversions and Determination ofInversion Endpoints
Ten PCR primers (table 1) were designed based on thesequence comparisons among 3 cp genome sequences ofJasminum (this study), Nicotiana (Shinozaki et al. 1986),and Panax (Kim and Lee 2004). Different combinationsof these primers were used to test for the presence or ab-sence of certain rearrangements. Positive PCR amplifica-tions were sequenced and aligned to determine inversionendpoints. The standard PCR-amplification reactions were30 cycles of 1 min denaturation at 94 �C, 1 min annealing at50–68 �C, and 2–8 min extension at 68–72 �C.
Phylogenetic Analyses and Estimation of DivergenceTimes of Inversions
DNA sequence data of ndhF and rbcL for 41 taxa andthe complete inversion regions (31–35 kb) of 6 taxa wereused for phylogenetic analysis. Forty-one ndhF and rbcLsequences representing all major clades of Oleaceae andoutgroups were generated by methods described elsewhere(Olmstead et al. 1992; Kim and Jansen 1995). Sequencealignment, base substitution analysis, and phylogeneticanalyses were performed using ClustalX (Higgins et al.1996), MEGA2 (Kumar et al. 2001), PAUP 4.0 (Swofford2002), and MacClade Version 4.06 (Maddison WP andMaddison DR 2003). The model of evolution was selectedusing the hierarchical likelihood ratio test as implementedin Modeltest 3.06 (Posada and Crandall 1998).
Divergence times were estimated using the combinedndhF and rbcL data. A likelihood ratio test (Felsenstein1981) was performed on the maximum likelihood (ML) treeby comparing the scores of the ML tree with and withouta molecular clock. This test rejected a molecular clockhypothesis; therefore, the penalized likelihood (PL)method (Sanderson 2002) in the program r8s version1.60 (Sanderson 2003) was used. The size of the roughnesspenalty was specified by a smoothing parameter obtainedfrom a cross-validation procedure. After cross-validation,
1162 Lee et al.
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

Table 1Taxa Used for cp DNA Sequencing, Gene Mapping, Molecular Clock Test, and PCR Diagnosis of Inversions
Groups Basedon LeafCharacters orTaxonomicGroup
Scientific Nameand AccessionInformationa
cpSequencingb
GeneMapping
ndhF/rbcLSequencesb
TribeJasmineae–Specific
1st Inversionc
SectionPrimulina–Specific
2nd InversioncMenodora-Specific 3rd and 4th
Inversionsc
P1/P2 P3/P4 P1/P3 P2/P4 P5/P6 P2/P5 P4/P6 P9/P10 P7/P8 P6/P9 P5/P7 P8/P10
Opposite lvs. Jasminum abyssinicumHochst. ex DC.(Kew 335-71-03007)
DQ673258 1 1/DQ673258 1 1 1 1 1
Opposite lvs. Jasminum angulare Vahl(Kew 367-63-36701)
1 1 1 1 1 1
Opposite lvs. Jasminum azoricum L.(Kew 000-73-12543)
1 1 1 1 1 1
Opposite lvs. Jasminum beesianum Forrest& Diels (Kew 000-69-13486)
1 1 1 1 1 1
Opposite lvs. Jasminum dallachii F. Muell.(Kew 000-73-02394)
1 1 1 1 1 1
Opposite lvs. Jasminum didymum G. Forst.(Kew 671-66-67110)
1 1 1 1 1 1
Opposite lvs. Jasminum dispermum Wall.(Kew 678-66-67818)
1 1 1 1 1 1
Opposite lvs. Jasminum floribundumR. Br. ex Fresen.(Kew 307-80-02859)
1 1 1 1 1
Opposite lvs. Jasminum gracile Andrews(UCONN 198800144)
1 1 1 1 1
Opposite lvs. Jasminum kedahense Ridl.(Kew 000-73-12545)
1 1 1 1 1
Opposite lvs. Jasminum le-ratii Schltr.(Kew 083-65-08303)
DQ673257 DQ673271/DQ673257 1 1 1 1 1
Opposite lvs. Jasminum nervosum Lour.(Kew 220-78-06124)
1 1 1 1 1 1
Opposite lvs. Jasminum officinale L.(UCONN 198502393)
1 1 1 1 1
Opposite lvs. Jasminum polyanthumFranch. (Edinburgh19697043)
1 1 1 1 1 1
Opposite lvs. Jasminum pubescens(Retzius) Willd. (FairchildPRC117)
1 1 1 1 1
Opposite lvs. Jasminum simplicifolium G.Forst. (Kew 000-73-12904)
1 DQ673270/DQ673298 1 1 1 1 1
Opposite lvs. Jasminum sinense Hemsl.(Kew 000-73-12548)
1 1 1 1 1
Opposite lvs. Jasminum tortuosum Willd.(Kew 349-67-34909)
1 1 1 1 1 1
Alternate lvs. Jasminum bignoniaceum Wall.ex G. Don (Kew000-72-12290)
1 1 1 1 1 1
Oleaceae
Ch
loro
plast
Gen
om
eR
earrang
emen
ts1
16
3
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018
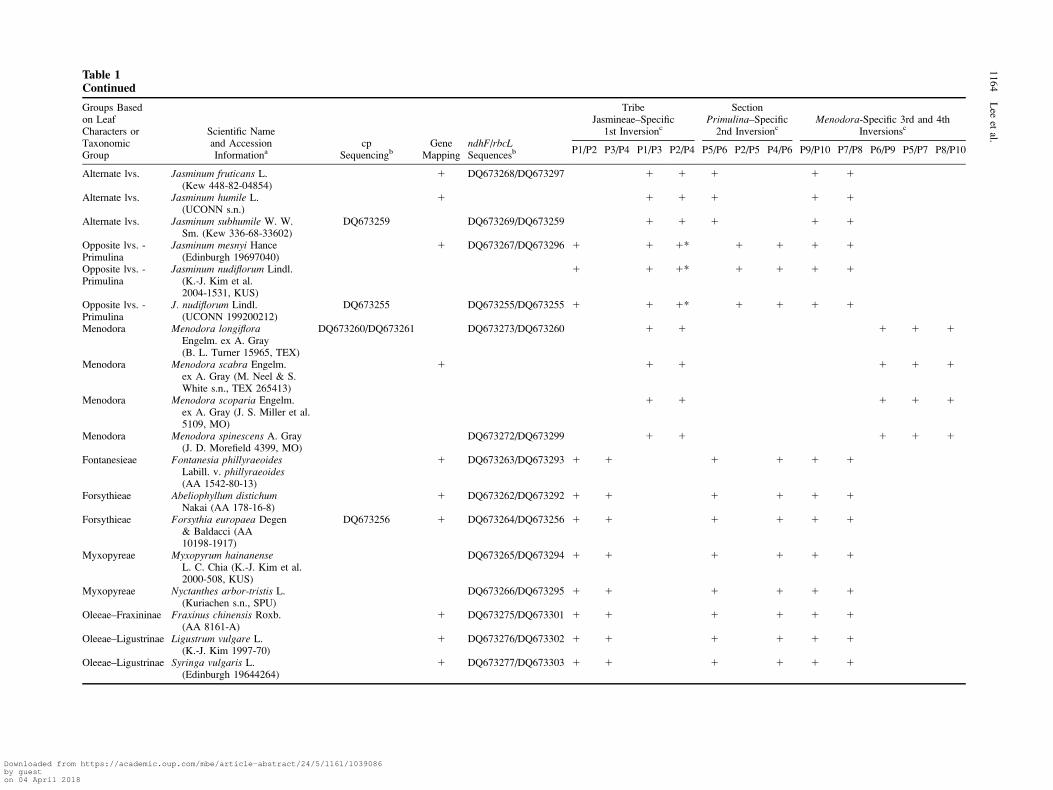
Table 1Continued
Groups Basedon LeafCharacters orTaxonomicGroup
Scientific Nameand AccessionInformationa
cpSequencingb
GeneMapping
ndhF/rbcLSequencesb
TribeJasmineae–Specific
1st Inversionc
SectionPrimulina–Specific
2nd InversioncMenodora-Specific 3rd and 4th
Inversionsc
P1/P2 P3/P4 P1/P3 P2/P4 P5/P6 P2/P5 P4/P6 P9/P10 P7/P8 P6/P9 P5/P7 P8/P10
Alternate lvs. Jasminum fruticans L.(Kew 448-82-04854)
1 DQ673268/DQ673297 1 1 1 1 1
Alternate lvs. Jasminum humile L.(UCONN s.n.)
1 1 1 1 1 1
Alternate lvs. Jasminum subhumile W. W.Sm. (Kew 336-68-33602)
DQ673259 DQ673269/DQ673259 1 1 1 1 1
Opposite lvs. -Primulina
Jasminum mesnyi Hance(Edinburgh 19697040)
1 DQ673267/DQ673296 1 1 1* 1 1 1 1
Opposite lvs. -Primulina
Jasminum nudiflorum Lindl.(K.-J. Kim et al.2004-1531, KUS)
1 1 1* 1 1 1 1
Opposite lvs. -Primulina
J. nudiflorum Lindl.(UCONN 199200212)
DQ673255 DQ673255/DQ673255 1 1 1* 1 1 1 1
Menodora Menodora longifloraEngelm. ex A. Gray(B. L. Turner 15965, TEX)
DQ673260/DQ673261 DQ673273/DQ673260 1 1 1 1 1
Menodora Menodora scabra Engelm.ex A. Gray (M. Neel & S.White s.n., TEX 265413)
1 1 1 1 1 1
Menodora Menodora scoparia Engelm.ex A. Gray (J. S. Miller et al.5109, MO)
1 1 1 1 1
Menodora Menodora spinescens A. Gray(J. D. Morefield 4399, MO)
DQ673272/DQ673299 1 1 1 1 1
Fontanesieae Fontanesia phillyraeoidesLabill. v. phillyraeoides(AA 1542-80-13)
1 DQ673263/DQ673293 1 1 1 1 1 1
Forsythieae Abeliophyllum distichumNakai (AA 178-16-8)
1 DQ673262/DQ673292 1 1 1 1 1 1
Forsythieae Forsythia europaea Degen& Baldacci (AA10198-1917)
DQ673256 1 DQ673264/DQ673256 1 1 1 1 1 1
Myxopyreae Myxopyrum hainanenseL. C. Chia (K.-J. Kim et al.2000-508, KUS)
DQ673265/DQ673294 1 1 1 1 1 1
Myxopyreae Nyctanthes arbor-tristis L.(Kuriachen s.n., SPU)
DQ673266/DQ673295 1 1 1 1 1 1
Oleeae–Fraxininae Fraxinus chinensis Roxb.(AA 8161-A)
1 DQ673275/DQ673301 1 1 1 1 1 1
Oleeae–Ligustrinae Ligustrum vulgare L.(K.-J. Kim 1997-70)
1 DQ673276/DQ673302 1 1 1 1 1 1
Oleeae–Ligustrinae Syringa vulgaris L.(Edinburgh 19644264)
1 DQ673277/DQ673303 1 1 1 1 1 1
11
64
Lee
etal.
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018
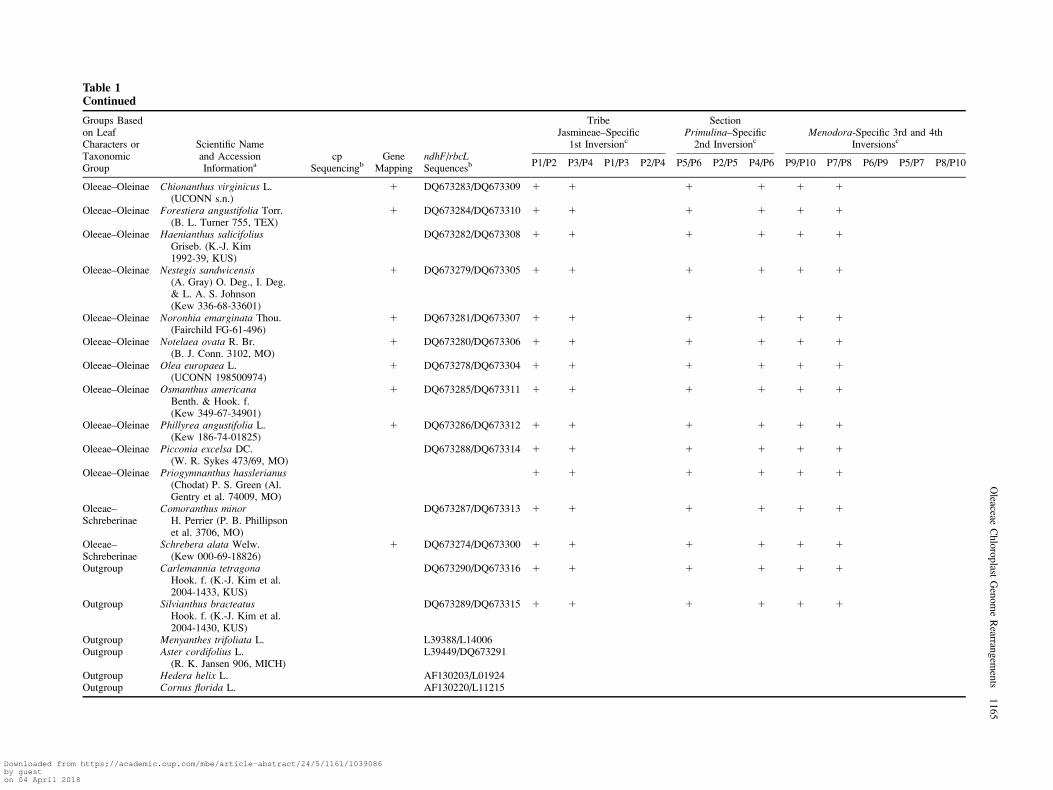
Table 1Continued
Groups Basedon LeafCharacters orTaxonomicGroup
Scientific Nameand AccessionInformationa
cpSequencingb
GeneMapping
ndhF/rbcLSequencesb
TribeJasmineae–Specific
1st Inversionc
SectionPrimulina–Specific
2nd InversioncMenodora-Specific 3rd and 4th
Inversionsc
P1/P2 P3/P4 P1/P3 P2/P4 P5/P6 P2/P5 P4/P6 P9/P10 P7/P8 P6/P9 P5/P7 P8/P10
Oleeae–Oleinae Chionanthus virginicus L.(UCONN s.n.)
1 DQ673283/DQ673309 1 1 1 1 1 1
Oleeae–Oleinae Forestiera angustifolia Torr.(B. L. Turner 755, TEX)
1 DQ673284/DQ673310 1 1 1 1 1 1
Oleeae–Oleinae Haenianthus salicifoliusGriseb. (K.-J. Kim1992-39, KUS)
DQ673282/DQ673308 1 1 1 1 1 1
Oleeae–Oleinae Nestegis sandwicensis(A. Gray) O. Deg., I. Deg.& L. A. S. Johnson(Kew 336-68-33601)
1 DQ673279/DQ673305 1 1 1 1 1 1
Oleeae–Oleinae Noronhia emarginata Thou.(Fairchild FG-61-496)
1 DQ673281/DQ673307 1 1 1 1 1 1
Oleeae–Oleinae Notelaea ovata R. Br.(B. J. Conn. 3102, MO)
1 DQ673280/DQ673306 1 1 1 1 1 1
Oleeae–Oleinae Olea europaea L.(UCONN 198500974)
1 DQ673278/DQ673304 1 1 1 1 1 1
Oleeae–Oleinae Osmanthus americanaBenth. & Hook. f.(Kew 349-67-34901)
1 DQ673285/DQ673311 1 1 1 1 1 1
Oleeae–Oleinae Phillyrea angustifolia L.(Kew 186-74-01825)
1 DQ673286/DQ673312 1 1 1 1 1 1
Oleeae–Oleinae Picconia excelsa DC.(W. R. Sykes 473/69, MO)
DQ673288/DQ673314 1 1 1 1 1 1
Oleeae–Oleinae Priogymnanthus hasslerianus(Chodat) P. S. Green (Al.Gentry et al. 74009, MO)
1 1 1 1 1 1
Oleeae–Schreberinae
Comoranthus minorH. Perrier (P. B. Phillipsonet al. 3706, MO)
DQ673287/DQ673313 1 1 1 1 1 1
Oleeae–Schreberinae
Schrebera alata Welw.(Kew 000-69-18826)
1 DQ673274/DQ673300 1 1 1 1 1 1
Outgroup Carlemannia tetragonaHook. f. (K.-J. Kim et al.2004-1433, KUS)
DQ673290/DQ673316 1 1 1 1 1 1
Outgroup Silvianthus bracteatusHook. f. (K.-J. Kim et al.2004-1430, KUS)
DQ673289/DQ673315 1 1 1 1 1 1
Outgroup Menyanthes trifoliata L. L39388/L14006Outgroup Aster cordifolius L.
(R. K. Jansen 906, MICH)L39449/DQ673291
Outgroup Hedera helix L. AF130203/L01924Outgroup Cornus florida L. AF130220/L11215
Oleaceae
Ch
loro
plast
Gen
om
eR
earrang
emen
ts1
16
5
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

the smoothing parameter was set to an absolute value of 1.0.The ML tree was converted to an ultrametric tree in PL us-ing the truncated network (TN) algorithm, and in the pro-cess, zero length branches were collapsed. Divergencetimes were calibrated using Cornaceae and Oleaceae fossils(Kavadas 1956; Muller 1981; Takahashi et al. 2002).
ResultsThe cp Genome Organization of J. nudiflorum
The cp genome of J. nudiflorum (GenBank accessionnumber NC_008407) is 165,121-bp long and is divided into4 regions by a pair of IRs (IRa and IRb) of 29,486 bp each.The LSC and SSC regions are 92,877 and 13,272 bp, re-spectively. The locations of the 133 genes in the J. nudiflo-rum cp genome are presented in figure 1. Nineteen genes (7tRNAs, 4 rRNAs, and 8 protein-coding genes) are dupli-cated in the IR. In addition, a duplicate copy of trnG-GCC is located in the middle of the LSC region. Elevenprotein-coding genes and a tRNA gene are located in theSSC region, whereas 61 protein-coding genes and 23 tRNAgenes are located in the LSC region. Nine protein-codinggenes and 6 tRNA genes each contain a single intron,and 2 protein-coding genes (rps12 and ycf3) have 2 introns(supplementary material SM 1, Supplementary Materialonline).
The J. nudiflorum cp genome consists of 57% codingsequence (50% protein and 7% RNA) and 43% noncodingsequences, including introns (12%) and intergenic spacers(31%). The overall A-T content of the J. nudiflorum cp ge-nome is 62%. The A-T content in the noncoding region(65%) is higher than in the coding region (60%). The A-T content of the IR is 58%, whereas the A-T content inthe LSC and SSC regions is 64% and 67%, respectively.The lower A-T content of the IR reflects the low A-T con-tent in the 4 rRNA (45%) genes in this region.
The gene and intron content and gene order of J. nudi-florum differ from Nicotiana tabacum in several ways. Forexample, the cp genome of J. nudiflorum is missing most ofaccD and the 2 introns of clpP. Additionally, duplicate cop-ies of trnG-GCC and ycf1 are present. The extra copy ofycf1 is due to the expansion of the IR at the IRb–SSCboundary, accounting for much of the increase in size ofthe J. nudiflorum cp genome in comparison with tobacco.In contrast, rps19 is located on the LSC region because ofthe contraction of IR. One notable rearrangement of the J.nudiflorum cp genome is the movement of a 2.8-kb regioncontaining ycf4 and psaI to the middle of the LSC region(fig. 1).
Comparative Gene Mapping of Oleaceae cp Genomes
The cp genomes were mapped for 32 representativespecies of all major genera of Oleaceae and 12 closely re-lated outgroup species (table 1). The gene maps (notshown) indicate that all genera of the Oleaceae except Jas-minum and Menodora have an identical gene order to N.tabacum. We sequenced the rearranged portions of theLSC region of the cp genomes for the representative speciesof Jasminum, Menodora, and the closely related genus For-sythia to better characterize the rearrangements in membersof the Jasmineae.T
able
1Continued
Gro
up
sB
ased
on
Lea
fC
har
acte
rso
rT
axon
om
icG
rou
p
Sci
enti
fic
Nam
ean
dA
cces
sio
nIn
form
atio
na
cpS
equ
enci
ng
bG
ene
Map
pin
gnd
hF/rbcL
Seq
uen
cesb
Tri
be
Jasm
inea
e–S
pec
ific
1st
Inver
sion
c
Sec
tio
nPrimulina–
Sp
ecifi
c2nd
Inver
sion
cMenodora-
Sp
ecifi
c3
rdan
d4
thIn
ver
sionsc
P1
/P2
P3
/P4
P1
/P3
P2
/P4
P5
/P6
P2
/P5
P4
/P6
P9
/P1
0P
7/P
8P
6/P
9P
5/P
7P
8/P
10
Ou
tgro
up
Nicotiana
taba
cum
L.
N0
01
87
9/N
001
87
9O
utg
rou
pGelsemium
sempervirens
Ait
.A
F1
30
17
0/L
14
39
7O
utg
rou
pClerodend
rum
trichotomum
Th
un
b.
exA
.M
urr
ay/
Clerodendrum
frag
rans
Wil
ld.
AF
130
14
6/L
11
68
9
Ou
tgro
up
Callicarpadichotom
aR
aeusc
h.
L3
63
95
/L1
439
3
Ou
tgro
up
Buddlejada
vidii
Fra
nch
.A
F1
30
14
3/L
14
39
2O
utg
rou
pDigitalispu
rpurea
L.
AF
130
15
0/L
01
90
2
aP
lantm
ater
ials
are
from
the
livin
gco
llec
tions
of
Kew
(Royal
Bota
nic
Gar
den
,Kew
),E
din
burg
h(R
oyal
Bota
nic
Gar
den
,Edin
burg
h),
Fai
rchild
(Fai
rchild
Tro
pic
alB
ota
nic
Gar
den
,Mia
mi)
,UC
ON
N(G
reen
house
of
the
Univ
ersi
tyof
Connec
ticu
tat
Sto
rrs)
,and
AA
(Arn
old
Arb
ore
tum
,Bost
on),
fiel
dco
llec
tions
from
Chin
a,In
dia
,and
United
Sta
tes
(all
vouch
ersp
ecim
ens
are
dep
osi
ted
atth
ein
dic
ated
her
bar
ia),
and
her
bar
ium
atM
O(M
isso
uri
Bota
nic
Gar
den
)an
dT
EX
(Univ
ersi
tyof
Tex
asat
Aust
in).
bG
enB
ank
acce
ssio
nnum
ber
sw
ith
DQ
init
ials
are
the
new
sequen
cere
port
edfr
om
this
study;
oth
erse
quen
ces
are
from
Gen
Ban
k.
cA
ster
isk
indic
ates
nonsp
ecifi
cam
pli
fica
tion.
Pri
mer
sequen
ces
are
P1:
5#-
AA
GT
TT
CC
TC
CC
GT
GA
TA
TG
AG
-3#,
P2:
5#-
CA
AG
TA
CG
GT
TC
TA
AG
GG
AA
GG
-3#,
P3
:5#-
GC
GG
GT
CT
CT
TT
AT
TA
GC
TC
CT
AT
TT
GT
-3#,
P4
:5#-
GG
GG
TG
AA
TG
GA
AT
CC
GA
TA
CA
TA
AA
-3#,
P5
:5#-
AA
TC
GA
TC
AT
AA
CC
AC
TA
CC
TA
TA
TT
CC
AG
-3#,
P6:5#-
CC
GT
AG
CT
AA
CC
GA
GT
AG
CT
CT
AG
A-3#,
P7
:5#-
CT
AT
CG
CT
AT
AT
TC
AT
CG
TT
AT
GT
GC
TA
GA
-3#,
P8:5#-
CA
T-
AC
CC
AA
TC
GA
AA
AA
GG
AT
GT
TA
TC
CA
AA
C-3#,
P9:
5#-
AA
CA
GC
AA
TC
CA
AG
GT
CG
CA
TA
CC
CA
GA
CG
GA
-3#,
and
P10:
5#-
CC
CA
TT
AG
TA
AG
CC
GG
TT
C-3#.
1166 Lee et al.
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

Identification of Inversions among Members of theOleaceae
Complete sequences for the region between psaA andpsbB in the LSC region were generated from F. europaea,J. subhumile, J. le-ratii, and J. abyssinicum (fig. 2) and areapproximately 32-kb long for each species. For M. longi-flora, 2 additional regions of the cp genome extending frompsbA to psaA (15,387 bp) and from trnK to rpl20 (18,522bp) were sequenced because mapping data (not shown) sug-gested the presence of inversions in these regions (fig. 3).
The sequenced region of the outgroup F. europaea(tribe Forsythieae) is 31,495-bp long and includes 25protein-coding and 8 tRNA genes (fig. 2). The gene contentand order are identical to the Nicotiana cp genome. Thesequence of J. subhumile (section Alternifolia) betweenpsbA and psbB is 35,515-bp long, including an inversionof 21.8 kb not present in Nicotiana or Forsythia. Relativeto the genomes of Nicotiana and Forsythia, one endpoint ofthe inversion is located between psaA and ycf3, and theother endpoint is located between ycf4 and cemA. In addi-tion, the J. subhumile cp genome lacks more than 50% ofaccD but includes trnI-CAU and trnS-GGA in this region.
Sequences between psaA and psbB genes of J. le-ratiiand J. abyssinicum (both members of the opposite-leavedgroup of Jasminum) are 31,507- and 31,044-bp long, re-spectively. These species have a similar gene order to J.subhumile, including one inversion of 19.3 kb possessingthe same endpoints. Gene and intron content of this groupare slightly different from F. europaea; they both lack mostof accD and 2 introns of clpP but include trnI-CAU down-stream from rbcL and rpl23 downstream from clpP.
The region between psaA and psbB in J. nudiflorum(section Primulina) is 36,404-bp long, and the gene orderis similar to Forsythia and Nicotiana, except for the relo-cation of ycf4 and psaI. The 2.8-kb region that includes ycf4and psaI is almost 18 kb upstream from its position in For-sythia and Nicotiana (fig. 1). One endpoint of this region islocated between psaA and ycf4, and the other is betweenpsaI and ycf3. Comparison of Jasminum, Forsythia, and Ni-cotiana cp genomes reveals that movement of this 2.8-kbregion in J. nudiflorum is caused by 2 unequal, sequentialinversions of 21 and 18 kb. The 21-kb inversion in J. nudi-florum is also present in other Jasminum species. One end-point of this inversion is located between psaA and ycf3,and the other endpoint is between ycf4 and cemA. In
FIG. 1.—Gene map of the Jasminum nudiflorum cp genome based on the complete sequence. A pair of thick lines on the inside circle represents theIRs (IRa and IRb: 29,486 bp each), which separate the LSC region (92,877 bp) from the SSC region (13,272 bp). Genes drawn inside the circle aretranscribed clockwise, whereas those outside are transcribed counterclockwise. Asterisks indicate intron-containing genes. Protein-coding gene names arein parentheses. The relocated region is located in the middle of the LSC region.
Oleaceae Chloroplast Genome Rearrangements 1167
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

addition, we hypothesize a 2nd 18-kb inversion in J. nudi-florum. One endpoint of the 2nd inversion is between ycf3and cemA and is only 134 bp from the endpoint of the 21-kbinversion. The other endpoint of the 18-kb inversion is 2.8kb from the 21-kb inversion endpoint. Therefore, the 2 in-versions represent independent evolutionary events, eventhough 1 endpoint of each inversion is found in nearlythe identical location. The incorporation of 134 bp of se-quence outside of the 1st inversion endpoint at one endand the inclusion of 42 bp of this sequence at the other end-point during the 2nd inversion clearly support the evolu-tionary ordering of these 2 inversions. The distribution
pattern of these remnant sequences indicates that the 18-kb inversion occurred after the 21-kb inversion.
The gene order in the Menodora cp genome is distinctfrom Jasminum species as well as other genera of Oleaceae.Three inversions can explain the differences in the cp ge-nome organization of Menodora (fig. 3). The 1st is 20 kband is shared with all sampled species of Jasminum. There-fore, this represents a shared derived character amongmembers of the tribe Jasmineae. The 2 endpoints of thisinversion are identical to the 1st inversion (21 kb) in Jas-minum (fig. 2). Two additional inversions (3rd and 4th in-versions in fig. 3) are unique to Menodora. The 3rd
FIG. 2.—Comparative gene maps showing the 1st and 2nd inversions in the Jasminum cp genome compared with those of the Nicotiana andForsythia genomes.
FIG. 3.—Comparative gene maps showing the 1st, 3rd, and 4th inversions in the Menodora cp genome compared with those of the Nicotiana,Forsythia, and Jasminum genomes.
1168 Lee et al.
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

inversion is approximately 14 kb, and the 2 endpoints arelocated between psaI and trnI-CAU and between trnT-UGU and rps4, and this inversion is nested within the1st inversion (fig. 3, the trnI is truncated). The4th inversion is the largest one at 59 kb. Because one end-point of the 4th inversion is located within trnK-UUU, the 2exons of trnK-UUU are located almost 59 kb apart. Theremaining endpoint is between trnI-CAU and rps4 (fig. 3).The 3rd inversion occurred prior to the 4th inversion.
Distribution of Inversions in Oleaceae
We designed 10 primers (P1–P10) to amplify the in-version endpoint regions (table 1 and fig. 4), and differentcombinations of these primers were used in PCR reactionsto determine the phylogenetic distribution of the 4 Oleaceaeinversions among 52 species (table 1).
A positive PCR amplification would be expected fromthe primer combinations of P1/P3 and P2/P4 for specieswith the 1st 21-kb inversion. In contrast, for the specieswithout the 1st inversion, a positive PCR reaction wouldresult from the primer combinations of P1/P2 and P3/P4.For the species with both the 1st and the 2nd inversions,the primer combinations of P2/P5 and P4/P6 would producepositive PCR reactions, whereas the primer combination ofP4/P6 would produce a positive PCR reaction in the specieswithout both inversions. For the 3rd and 4th inversions, a se-ries of positive amplifications would be expected from theprimer combinations of P6/P9, P5/P7, P1/P3, P8/P10, andP2/P4.
Representative positive and negative PCR results for 9species are shown in figure 5. Lanes 1–4 represent species
with only the 1st inversion (all members of Jasminum ex-cept section Primulina), lanes 5–6 represent species withthe 1st and the 2nd inversions (section Primulina), lanes7–8 are species with the 1st, 3rd, and 4th inversions(Menodora), and lane 9 represents the species withoutany inversions (Forsythia).
The PCR diagnosis (table 1 and fig. 5) indicates thatthe 1st inversion is shared by all sampled members of thetribe Jasmineae, whereas all other tribes of the Oleaceaelack this inversion. Our findings also indicate that the2nd inversion is restricted to the section Primulina of Jas-minum and that the 3rd and 4th inversions are unique toMenodora.
All PCR results agreed with our expectations with oneexception (fig. 5); an unexpected amplification was ob-served for the primer combination of P2/P4 in J. nudiflo-rum. Based on the complete cp sequence, the primercombination of P2/P4 should not generate a PCR product;however, an amplicon was produced from total DNA. Thisregion is between ycf3 and cemA, and we sequenced thefragment and compared it with sequences of this regionfrom other species. The PCR product contains many basesubstitutions and insertions/deletions, suggesting that thefragments may not be cpDNA. The amplicon apparentlycame from the primer-specific sites (see table 1), but fromcpDNA that underwent intracellular gene transfer prior tothe 2nd inversion and is no longer resident in the cp ge-nome. Filter hybridizations of J. nudiflorum in this regionalso showed weak hybridizing fragments (data not shown).These results suggest that the unexpected fragment ofJ. nudiflorum might be due to the transfer of this regionto the nucleus or mitochondrion. Transfers of cp genes
FIG. 4.—Location of 10 diagnostic PCR primers for identification of 4 inversion endpoints. Sequences for the 10 primers (P1–P10) are given intable 1.
Oleaceae Chloroplast Genome Rearrangements 1169
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018
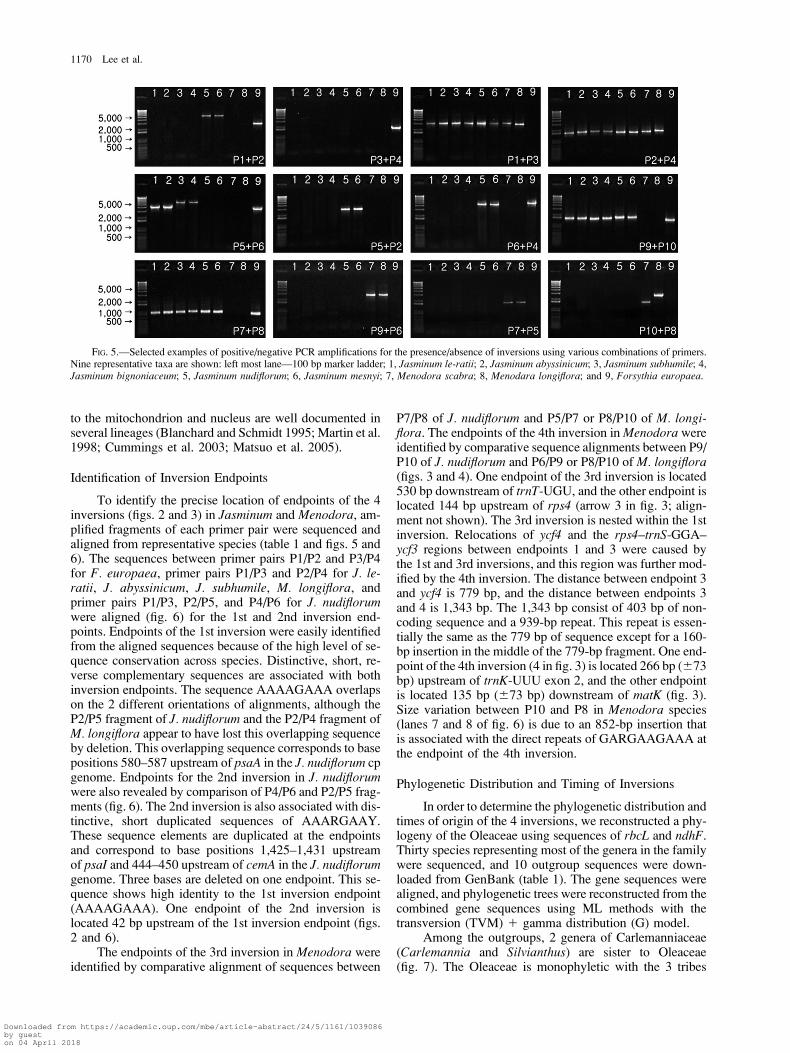
to the mitochondrion and nucleus are well documented inseveral lineages (Blanchard and Schmidt 1995; Martin et al.1998; Cummings et al. 2003; Matsuo et al. 2005).
Identification of Inversion Endpoints
To identify the precise location of endpoints of the 4inversions (figs. 2 and 3) in Jasminum and Menodora, am-plified fragments of each primer pair were sequenced andaligned from representative species (table 1 and figs. 5 and6). The sequences between primer pairs P1/P2 and P3/P4for F. europaea, primer pairs P1/P3 and P2/P4 for J. le-ratii, J. abyssinicum, J. subhumile, M. longiflora, andprimer pairs P1/P3, P2/P5, and P4/P6 for J. nudiflorumwere aligned (fig. 6) for the 1st and 2nd inversion end-points. Endpoints of the 1st inversion were easily identifiedfrom the aligned sequences because of the high level of se-quence conservation across species. Distinctive, short, re-verse complementary sequences are associated with bothinversion endpoints. The sequence AAAAGAAA overlapson the 2 different orientations of alignments, although theP2/P5 fragment of J. nudiflorum and the P2/P4 fragment ofM. longiflora appear to have lost this overlapping sequenceby deletion. This overlapping sequence corresponds to basepositions 580–587 upstream of psaA in the J. nudiflorum cpgenome. Endpoints for the 2nd inversion in J. nudiflorumwere also revealed by comparison of P4/P6 and P2/P5 frag-ments (fig. 6). The 2nd inversion is also associated with dis-tinctive, short duplicated sequences of AAARGAAY.These sequence elements are duplicated at the endpointsand correspond to base positions 1,425–1,431 upstreamof psaI and 444–450 upstream of cemA in the J. nudiflorumgenome. Three bases are deleted on one endpoint. This se-quence shows high identity to the 1st inversion endpoint(AAAAGAAA). One endpoint of the 2nd inversion islocated 42 bp upstream of the 1st inversion endpoint (figs.2 and 6).
The endpoints of the 3rd inversion in Menodora wereidentified by comparative alignment of sequences between
P7/P8 of J. nudiflorum and P5/P7 or P8/P10 of M. longi-flora. The endpoints of the 4th inversion in Menodora wereidentified by comparative sequence alignments between P9/P10 of J. nudiflorum and P6/P9 or P8/P10 of M. longiflora(figs. 3 and 4). One endpoint of the 3rd inversion is located530 bp downstream of trnT-UGU, and the other endpoint islocated 144 bp upstream of rps4 (arrow 3 in fig. 3; align-ment not shown). The 3rd inversion is nested within the 1stinversion. Relocations of ycf4 and the rps4–trnS-GGA–ycf3 regions between endpoints 1 and 3 were caused bythe 1st and 3rd inversions, and this region was further mod-ified by the 4th inversion. The distance between endpoint 3and ycf4 is 779 bp, and the distance between endpoints 3and 4 is 1,343 bp. The 1,343 bp consist of 403 bp of non-coding sequence and a 939-bp repeat. This repeat is essen-tially the same as the 779 bp of sequence except for a 160-bp insertion in the middle of the 779-bp fragment. One end-point of the 4th inversion (4 in fig. 3) is located 266 bp (673bp) upstream of trnK-UUU exon 2, and the other endpointis located 135 bp (673 bp) downstream of matK (fig. 3).Size variation between P10 and P8 in Menodora species(lanes 7 and 8 of fig. 6) is due to an 852-bp insertion thatis associated with the direct repeats of GARGAAGAAA atthe endpoint of the 4th inversion.
Phylogenetic Distribution and Timing of Inversions
In order to determine the phylogenetic distribution andtimes of origin of the 4 inversions, we reconstructed a phy-logeny of the Oleaceae using sequences of rbcL and ndhF.Thirty species representing most of the genera in the familywere sequenced, and 10 outgroup sequences were down-loaded from GenBank (table 1). The gene sequences werealigned, and phylogenetic trees were reconstructed from thecombined gene sequences using ML methods with thetransversion (TVM) 1 gamma distribution (G) model.
Among the outgroups, 2 genera of Carlemanniaceae(Carlemannia and Silvianthus) are sister to Oleaceae(fig. 7). The Oleaceae is monophyletic with the 3 tribes
FIG. 5.—Selected examples of positive/negative PCR amplifications for the presence/absence of inversions using various combinations of primers.Nine representative taxa are shown: left most lane—100 bp marker ladder; 1, Jasminum le-ratii; 2, Jasminum abyssinicum; 3, Jasminum subhumile; 4,Jasminum bignoniaceum; 5, Jasminum nudiflorum; 6, Jasminum mesnyi; 7, Menodora scabra; 8, Menodara longiflora; and 9, Forsythia europaea.
1170 Lee et al.
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

Myxopyreae, Fontanesieae, andForsythieae forming theear-liest diverging lineages. The tribe Jasmineae forms a stronglysupportedmonophyleticgroup.WithintribeJasmineae,3lin-eages are evident, and they correspond to 3 well-recognizedtaxonomic groups: the alternate-leaved Jasminum (sectionAlternifolia), the opposite-leaved Jasminum, andMenodora.
Within the opposite-leaved Jasminum, 2 subgroups are pres-ent, one of which corresponds to section Primulina (J. nudi-florum and Jasminum mesnyi) and the other includingJasminum simplicifolium and J. le-ratii.
All sampled members of the Jasmineae share the 1st21-kb inversion. The 2 species of section Primulina have
FIG. 6.—Sequence alignments showing 1 of 4 inversion endpoints (shaded area). Eight of 52 sequences are shown.
Oleaceae Chloroplast Genome Rearrangements 1171
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018
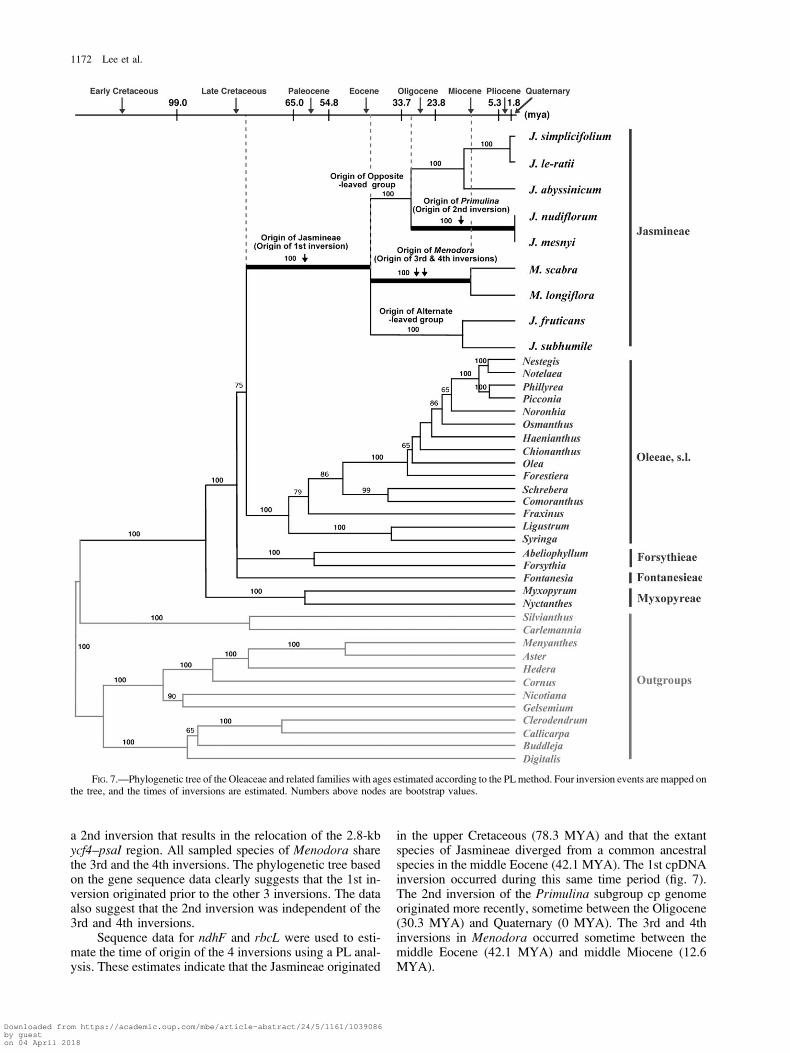
a 2nd inversion that results in the relocation of the 2.8-kbycf4–psaI region. All sampled species of Menodora sharethe 3rd and the 4th inversions. The phylogenetic tree basedon the gene sequence data clearly suggests that the 1st in-version originated prior to the other 3 inversions. The dataalso suggest that the 2nd inversion was independent of the3rd and 4th inversions.
Sequence data for ndhF and rbcL were used to esti-mate the time of origin of the 4 inversions using a PL anal-ysis. These estimates indicate that the Jasmineae originated
in the upper Cretaceous (78.3 MYA) and that the extantspecies of Jasmineae diverged from a common ancestralspecies in the middle Eocene (42.1 MYA). The 1st cpDNAinversion occurred during this same time period (fig. 7).The 2nd inversion of the Primulina subgroup cp genomeoriginated more recently, sometime between the Oligocene(30.3 MYA) and Quaternary (0 MYA). The 3rd and 4thinversions in Menodora occurred sometime between themiddle Eocene (42.1 MYA) and middle Miocene (12.6MYA).
FIG. 7.—Phylogenetic tree of the Oleaceae and related families with ages estimated according to the PL method. Four inversion events are mapped onthe tree, and the times of inversions are estimated. Numbers above nodes are bootstrap values.
1172 Lee et al.
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

In order to more accurately estimate the order and tim-ing of the 4 inversions, we performed phylogenetic analysesof the tribe Jasmineae using cpDNA sequences betweenpsaA and 5#-rps12, which includes the inversion regionsfor J. nudiflorum, J. le-ratii, J. abyssinicum, J. subhumile,M. longiflora, and F. europaea. The 6 aligned sequenceswere 34,607-bp long, and phylogenetic analyses usingML and general time reversible (GTR) 1 invariable siteproportion (I) 1 G resulted in the tree in figure 8a. We alsoperformed phylogenetic analyses of the accD gene regionafter excluding unalignable noncoding regions and all smallgaps from the data matrix; the resulting matrix included23,217 aligned base pairs, and the resulting ML tree usingGTR 1 I 1 G is shown in figure 8b.
The topologies of the 2 trees (figs. 8a and b) differ inthe position of Menodora. Menodora longifolia is sister toan alternate-leaved J. subhumile in tree A, whereas in treeB, M. longiflora is sister to the clade comprising the other 3Jasminum species. The ML values (lnL) of tree A and tree Bwith gaps are�89806.04 and�89810.90, respectively. ThelnL of the 2 trees without gaps are �48182.76 and�48177.34, respectively. The P values for the Kishino–Hasegawa test on tree A and tree B are 1.000 and 0.139,respectively. The P values for the Kishino–Hasegawa teston trees A and B without gaps are 0.320 and 1.000, respec-tively. Thus, the Kishino–Hasegawa test indicates that the 2trees are not significantly different.
The phylogenetic analyses confirm our hypotheses re-garding the evolutionary order of the inversion events. The1st inversion arose prior to the other 3 inversions (character 1in fig. 8). The 2nd inversion (character 2 in fig. 8) is indepen-dent of the3rd and 4th inversions (characters 3 and 4 infig. 8).The 3rd and the 4th inversions are restricted to theMenodora.
Other Rearrangements in Jasmineae cp GenomesDuplicated Insertions of 3 tRNA Genes
Two copies of trnG-GCC are located in the LSC re-gion of J. nudiflorum. The copy near psbZ (coordinate39,000 in fig. 1) is at the same location in tobacco, andthe copy downstream of rbcL (coordinate 63,000 in fig.1) is the duplicate. The 2 copies have 100% sequence iden-tity but are in reverse orientations. The noncoding se-quences 42 bp upstream and 54 bp downstream of thegene are also identical in both locations, suggesting thattrnG-GCC and the surrounding noncoding sequence areduplicated and inserted into a new location.
Similar duplicated insertions of tRNA genes are foundin other lineages of Jasminum and Menodora. The trnI-CAU, which is usually located in the IR, also occurs down-stream of rbcL in all species of Menodora and Jasminumexcept section Primulina. In addition, a duplicate copy oftrnS-GCU, which is normally located near psbI (near coor-dinate 10,000; fig. 1), also occurs downstream of rbcL in thealternate-leaved species of Jasminum (represented by J.subhumile in fig. 2). All 3 duplicated insertions are locateddownstream of rbcL and are correlated with the reduction orloss of accD, even though the 3 tRNA genes were originallyin several different regions of the cp genome.
Duplicated Insertion of rpl23
An extra copy of rpl23 is inserted downstream of clpPin the LSC region of J. le-ratii and J. abyssinicum. Thisgene is located between trnI-CAU and rpl2 in the IR regionin tobacco. Copies of these genes in the LSC and IR regionsshow minor differences in length and sequence identity(supplementary material SM 2, Supplementary Material
FIG. 8.—Two different phylogenetic trees illustrating the evolution of various cp genome rearrangements. Trees were generated from the alignedsequencesof31-kbregionsofcpDNAusingMLmethods.Numbersabovenodesarebootstrapvalues. (a)TreewasgeneratedbyMLmethodswithGTR1 I1G model from 34,607 bp of aligned sequences between the psaA and 5#-rps12 genes. (b) Tree was generated by ML methods with GTR 1 I 1 G modelfrom 23,217 bp excluding accD gene regions, unalignable noncoding regions, and all small gaps. Genome rearrangement characters are the following:1, inversion one; 2, inversion two; 3, inversion three; 4, inversion four; 5, accD loss; 6, intron losses in clpP; 7, rpl23 duplication and relocation; 8: trnTduplication and relocation; 9, trnG duplication; 10, trnI duplication; and 11, trnS duplication.
Oleaceae Chloroplast Genome Rearrangements 1173
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

online). The IR copy is 282-bp long, and a 3-bp deletion aswell as 4 base substitutions are present in the LSC region ofJ. le-ratii. In addition, 79 and 15 bp of noncoding sequencelocated upstream and downstream of both copies of rpl23,respectively, are duplicated. Different species of Jasminumexhibit variation between the 2 copies of rpl23. For example,the LSC region of J. abyssinicum terminates at position 228because of a 2-bp deletion, causing a frameshift mutation.The duplicated segments of rpl23 and associated noncodingregions have identical sequences in all surveyed species,suggesting that this is a shared derived character of theopposite-leaved species of Jasminum.
Duplication of ycf1 by Expansion of the IR
Detailed comparisons of the IR–SSC boundaries ofNicotiana, Panax, Arabidopsis, and J. nudiflorum (fig. 9)indicate that ycf1 is completely incorporated into the IR re-gion in J. nudiflorum. This expansion makes the IR regionof J. nudiflorum approximately 4 kb longer than the otherspecies compared, and the SSC region of Jasminum is con-sequently shorter than these other genomes.
Small Inversion of trnT-UGU
The trnT-UGU is normally located on the oppositestrand and upstream of trnL-UAA. However, in J. nudiflo-rum, it is located on the same strand as trnL-UAA (fig. 1, nearcoordinate 55,000). This small inversion is only present in J.nudiflorum and J.mesnyi. The change in strand of trnT-UGUis due to a small 219-bp inversion. In addition to the trnT-UGU gene, the inversion includes 24 bp upstream and122 bp downstreamof thegene sequence.Short IR sequencesare conserved (GAA-TTCCTAT and ATAGGAACTTC)except one base indel at both ends of the inversion.
Interruption of trnK-UUU by Inversion
The trnK-UUU is unusual because it contains matKwithin its intron in most cp genomes. Yet, the cp genomes
of Menodora have an interrupted trnK-UUU gene due to aninversion endpoint within the intron between matK and the3# exon of trnK-UUU (fig. 3, inversion 4). As a result, the5# exon is located upstream of matK, whereas the 3# exon islocated upstream of the psaA gene. The 2 exons of thetrnK-UUU gene are separated by 59 kb because of this largeinversion (fig. 3).
Serial Loss of accD
All sampled species of Jasminum and Menodora aremissing some or all of accD. The serial loss of this gene isrestricted to the members of the tribe Jasmineae, thus thischaracter supports the monophyly of the tribe Jasmineae(fig. 8, character 5). The accD is usually located betweenrbcL and psaI in cp genomes of land plants. Therefore, wesequenced the region from rbcL to psaI for 5 Jasmineae spe-cies that have lost portions of accD gene and comparedthese sequences with 6 other species in which completeaccD is present. The length of this region in species withaccD is conserved, ranging from 2,835 to 3,068 bp in 6 phy-logenetically diverse species (Nicotiana, Arabidopsis, Pan-ax, Spinacia, Atropa, and Forsythia). In contrast, the size ofthe region that corresponds to the rbcL–psaI intergenicspacer region among 5 Jasmineae species ranges from2,922 to 5,271 bp. Furthermore, average sequence diver-gence for this region among Jasmineae species is0.4428, which is far higher than the average of 0.2407 seenamong the 6 diverse outgroups.
Distinctive sequence elements are present in the regionthat corresponds to rbcL and psaI of the Jasminum species(fig. 10). Nearly half of the 3# end of accD (947–1,449 bp)remains in the alternate-leaved clade of Jasminum (repre-sented by J. subhumile). In addition, a small fragment fromthe middle of accD (822–907 bp) remains in all members ofJasminum (fig. 10). However, the middle section of theaccD sequence is completely absent in Menodora. The datasuggest that the loss of accD occurred sequentially with the
FIG. 9.—Comparison of boundaries of the SSC, LSC, and IR regions among 4 cp genomes. Length variation of ycf1 pseudogenes (5# portions) occursat the IRb–SSC boundary. The IR region extends into the ycf1 gene resulting in 2 complete copies in Jasminum nudiflorum.
1174 Lee et al.
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

5# portion first, the 3# portion next, and finally the middleportion of the gene. Complete or partial gene fragmentsfrom other parts of cp genomes are inserted into this region(fig. 10). These fragments include an atpE gene fragment(172–290 bp) in all species; a complete copy of trnI-CAU in all Jasminum except the Primulina subgroup(supplementary material SM 3, Supplementary Material on-line); complete trnS-GCU, partial trnP-UGG, and partial5#-rps12 for the alternate-leaved species of Jasminum;and complete trnG-GCC for all species of Jasminum exceptthe alternate-leaved Jasminum. The trnI-CAU and trnG-GCC genes are also inserted into this region in Menodora.In addition, a portion of the rpl23 gene is found in thisregion in Menodora and the Primulina subgroup ofJasminum. A massive duplication of a 12-bp repeated se-quence in the middle of the noncoding region between rbcLand psaI occurs in J. nudiflorum (fig. 10). The consensus12-bp sequence of MNNMAYYDGNNV is repeated 100times in the region where accD has been reduced (sup-plementary material SM 4, Supplementary Materialonline). The insertion of these repeats probably originateda long time ago because the individual repeating unitsare very divergent. Another highly repetitive element
occurs downstream of clpP in J. nudiflorum.In this case, there are 56 copies of a 27-bp repeat(YWTTTTYTTTWGDRAHHRWYAYTTHHT), and thisinsertion coincides with the loss of 2 introns in clpP(supplementary material SM 5, Supplementary Materialonline). These repeats probably originated more recentlybecause there are no base substitutions among the individ-ual copies of the repeat elements.
Intron Losses in clpP
Two introns in clpP are missing in the cp genome of J.nudiflorum. Filter hybridization and sequencing resultssuggest that the losses of 2 introns in clpP are confinedto opposite-leaved Jasminum, with both introns being wellconserved in alternate-leaved Jasminum (fig. 8, character6). The clpP introns are also absent in Menodora cpDNA.Two independent intron losses are hypothesized in figure8A, whereas single intron loss is supported in figure 8B.In addition to the clpP intron loss, further rearrangementsare located downstream of clpP. The gene rpl23 typicallyfound in the IR in other angiosperm cp genomes has beenduplicated, and one copy occurs downstream of clpP in
FIG. 10.—Detailed gene map between rbcL and psaI for 5 representative species of Jasmineae. Numbers before and after the gene name indicate thecoordinate of the gene in reference to Nicotiana tabacum. C, completed gene; p, partial gene.
Oleaceae Chloroplast Genome Rearrangements 1175
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

Menodora and the opposite-leaved species of Jasminum(figs. 2 and 3; supplementary material SM. 2, SupplementaryMaterial online). In J. nudiflorum, the above-mentioned 27-bp sequence is repeated 56 times in this region (supplemen-tary material SM 5, Supplementary Material online).
Discussion
The cp genomes of the Oleaceae tribe Jasmineae ex-hibit a number of unusual rearrangements; together thesechanges make the Jasminum and Menodora cp genomesunique among land plants. The most notable changes con-cern the relocation of genes due to multiple, overlappinginversions, and the separation of the 2 exons of trnK-UUU by 59 kb. Below, we discuss several unusual featuresof Oleaceae cp genomes and compare them with otherrearranged land plant cp genomes.
Gene Relocations Caused by Multiple Inversions
The cp genomes of the tribe Jasmineae have experi-enced multiple movements of gene segments. In Jasminumsection Primulina (J. mesnyi and J. nudiflorum), a 2.8-kbregion containing ycf4, psaI, and associated noncoding se-quences has been moved 18 kb by 2 consecutive inversionsof 21 and 18 kb. The phylogenetic distribution of these 2inversions (fig. 8, characters 1 and 2) suggests that the 21-kb inversion originated prior to the 18-kb inversion (figs. 2,3, and 7). Divergence time estimates also indicate that the1st inversion originated between the upper Cretaceous(78.3 MYA) and the middle Eocene (42.1 MYA), whereasthe 2nd inversion originated between the lower Oligocene(30.3 MYA) and Quaternary (0 MYA). In Menodora, 2 re-gions, rps4–trnS-GGA–ycf3 and ycf4, were relocated by 3different inversions (fig. 3). The 1st inversion is sharedamong all members of Jasmineae, and the other 2 (inver-sions 3 and 4 in fig. 3) are unique to Menodora. The3rd inversion likely occurred prior to the 4th inversioneven though this inversion is not present in any speciesexamined.
Relocations of genes have been documented in otherangiosperm lineages, and 2 different mechanisms havebeen proposed to cause these events. In legumes (Palmeret al. 1988; Perry et al. 2002), successive expansion andcontraction of the IR have resulted in the relocation ofa block of 6 ribosomal protein genes (rps19–rps8) fromone end of the LSC region to the other end. Gene move-ments in Pelargonium can be attributed to both IR expan-sion and contraction and multiple inversions (Chumleyet al. 2006). In grasses (Quigley and Weil 1985; Hiratsukaet al. 1989; Doyle et al. 1992), 3 inversions have occurred,which have caused the relocation of several genes, includ-ing trnG-GCC through psbD.
Short dispersed repeats have been suggested as one ofthe major factors promoting inversions (Palmer 1991; Knoxet al. 1993). Such repeats are common in cp genomes thatare highly rearranged, including algae, grasses, conifers,geranium, subclover, and Campanulaceae (Bowman andDyer 1986; Palmer, Nugent, and Herbon 1987; Bowmanet al. 1988; Milligan et al. 1989; Lidholm and Gustafsson1991; Doyle et al. 1992; Knox et al. 1993; Cosner et al.
1997; Chumley et al. 2006). Inversions have been shownto occur through intramolecular recombination between re-peat elements (Palmer, Nugent, and Herbon 1987; Ogiharaet al. 1988; Achaz et al. 2003; Rocha 2003; Pombert et al.2006). Both endpoints of the 1st inversion in Jasminum andMenodora are terminated by the short repeats of AAAA-GAAA (fig. 6). The 2nd inversion is also associated withthe similar short repeated sequences of AAARGAAY in J.nudiflorum, and 100 copies of MNNMAYYDGNNV se-quence elements occur near one endpoint of this inversion(supplementary material SM 4, Supplementary Material on-line). The endpoints of the 1st inversion in Menodora arealso associated with short repeats of AAAAGAAA. In con-trast to the short repeats at the endpoints of the 1st and 2ndinversions, the ends of the 3rd and the 4th inversions havemuch longer repeat elements of 779 and 939 bp (fig. 10).These long duplicated sequences likely facilitated the3rd and 4th inversions by intramolecular recombinationin Menodora.
Several studies have also suggested that tRNAs playa role in promoting gene-order changes by nonhomologousrecombination (Hiratsuka et al. 1989; Knox et al. 1993;Turmel et al. 2002). Duplicated copies of 2 tRNAs,trnG-GCC and trnT-UGU, are located near the endpointsof inversions 3 and 4 in Menodora (fig. 3). Thus, the wide-spread occurrence of repeats and tRNAs near the inversionendpoints both in Menodora and Jasminum cp genomes iscoincident with a series of inversions caused by intramolec-ular recombination.
Gene and Intron Loss
Gene and intron losses have been documented in anumber of land plant cp genomes (Downie and Palmer1992; Raubeson and Jansen 2005). Within the Jasmineae,accD has a series of losses in the species of Jasminum andMenodora examined, supporting the possible phylogeneticrelationship within the tribe (figs. 7, 8, and 10). This genehas been lost completely in 4 other lineages of angiosperms,including the grasses (Hiratsuka et al. 1989; Maier et al.1995; Katayama and Ogihara 1996), Campanulaceae(Cosner et al. 1997), Geraniaceae (Palmer, Nugent, andHerbon 1987; Chumley et al. 2006), and Lobeliaceae (Knoxand Palmer 1999). The loss of accD in Campanulaceae andLobeliaceae is a shared derived feature of these familiesand supports their close phylogenetic relationship (Knoxand Palmer 1999). It is notable that the loss of accD isassociated with hotspots of rearrangements in each of thefamilies in which it has been lost. In the grasses, Menodora,and thePrimulina subgroup of Jasminum in the region whereaccD normally resides, a pseudogene of rpl23 is insertedinto the region between rbcL and psaI (fig. 10). The mech-anism for insertion of different sequence elements in this re-gion is uncertain. The accD gene loss may either accelerategene relocations by unknown mechanisms, or various move-ments of genes may induce the sequential loss of accD.
Most cp genomes of land plants include 18 intron-containing genes, 15 of which have 2 exons and 3 (ycf3,rps12, and clpP) with 3 exons. A number of independentlosses of introns have been detected in various genes, in-cluding rpoC1 (Downie et al. 1996, 1998; Hansen et al.
1176 Lee et al.
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

2006), rpl16 (Downie and Palmer 1992), clpP (Maier et al.1995; Stefanovic and Olmstead 2005), rpl2 (Downie et al.1991; Doyle et al. 1995; Stefanovic and Olmstead 2005),rps12 (Hoot and Palmer 1994; Freyer et al. 1995), trnK(Wolfe et al. 1992; Turmel et al. 2005), and ycf3 (Stefanovicand Olmstead 2005). In the Oleaceae, the loss of the 2 in-trons in clpP is confined to opposite-leaved Jasminum andMenodora (figs. 3 and 8).
Gene Duplications
Most gene duplications in cp genomes are caused bythe expansion of the IR (Goulding et al. 1996). Theseexpansions tend to be relatively small and result in the du-plication of only single genes or parts of genes, which oftenproduce pseudogenes at the IR–SC boundaries (Kim andLee 2004). In a number of cases, the IR has greatly ex-panded, including a 12-kb expansion in Nicotiana acumi-nata (Solanaceae; Goulding et al. 1996), 11.5 kb inMahonia (Berberidaceae; Kim and Jansen 1994), 11 kbin Lobelia thuliana (Lobeliaceae; Knox and Palmer1999), and a remarkable 50 kb in Pelargonium (Gerania-ceae; Palmer, Nugent, and Herbon 1987; Chumley et al.2006). In J. nudiflorum, expansion of the IR has causeda duplication of the normally single-copy gene ycf1 (fig.9). The duplication ycf1 of Jasminum is analogous to therps15 duplication in the rye cp genome (Prombona andSubramanian 1989).
Duplications of cp genes that do not involve the IR areuncommon, and only a few have been documented in cpgenomes of Arabidopsis (Brassicaceae; Koch et al.2005), Pelargonium (Geraniaceae; Chumley et al. 2006),Pinus (Pinaceae; Wakasugi et al. 1994), and Trachelium(Campanulaceae; Cosner et al. 1997). These duplicationshave involved mostly tRNAs and some protein-codinggenes. The only duplication of a protein-coding gene inthe Oleaceae involves rpl23, which has resulted in a 2ndcopy of this gene in the LSC region of the opposite-leavedspecies of Jasminum. This gene is normally located be-tween trnI-CAU and rpl2 (supplementary material SM 2,Supplementary Material online) in the IR region (supple-mentary material SM 3, Supplementary Material online).A very similar duplication of rp123 has been documentedin the Poaceae, and in both the Oleaceae and Poaceae, theduplicated copy has been inserted into the intergenic regionbetween rbcL and psaI where accD normally resides. Mostcp gene duplications outside of the IR involve tRNAs, andthis is certainly the case in the Oleaceae. Several tRNA du-plications have occurred, including trnG-GCC in the LSCregion of J. nudiflorum (fig. 1), trnI-CAU in the LSC spe-cies of Menodora and Jasminum except section Primulina,and trnS-GCU in the LSC region of alternate-leaved speciesof Jasminum (represented by J. subhumile) (fig. 10). In theOleaceae, most of these tRNA duplications are associatedwith other rearrangements in the cp genomes. The mecha-nism for tRNA duplications is unknown, but they could becaused by duplicative transpositions.
Interruption of trnK-UUU
The most unusual rearrangement in the Oleaceaeinvolves the separation of the 2 exons of trnK-UUU by
59 kb. The cp genomes of Menodora have an interruptedtrnK-UUU gene because of an inversion breakpoint withinthe intron between the matK gene and the trnK-UUU 3#exon (fig. 3). The only other gene with exons that areseparated by such a large distance is rps12, which has 3exons. The 5# exon for this gene is located in the LSC re-gion within an operon that includes 2 other genes, clpP andrpl20, and the two 3# exons are in the IR, approximately 30kb from the 5# exon. In several cp genomes, transsplicinghas been demonstrated for rps12 to produce a transcript(Fukuzawa et al. 1986; Torazawa et al. 1986; Gieseet al. 1987). Separation of the 2 trnK-UUU exons suggests2 possible alternatives for the functionality of this gene inM. longiflora. Either the gene must be transspliced so that itcan function as it does in other cp genomes or the gene isnonfunctional. We have no experimental evidence demon-strating transsplicing of trnK-UUU in Menodora. However,this gene is absent in 2 cp genomes, the fern Adiantum(Wolf et al. 2003) and the holoparasite Epifagus (Wolfeet al. 1992). In both of these genomes, matK, which is en-coded in the trnK-UUU intron and is involved in intronsplicing of trnK-UUU, is present and functional (Hauseret al. 2006). The situation in Adiantum is very similar toMenodora because a large inversion in the cp genomehas one endpoint in the trnK-UUU intron, but in this casethe 2 exons were not found (Wolf et al. 2003). In Epifagus,Wolfe et al. (1992) suggested that matK may have an alter-native function, which most likely involves splicing ofother group II introns (Hauser et al. 2006). Experimentalstudies of both trnK-UUU and matK are needed to inves-tigate these 2 alternatives in Menodora.
In summary, cpDNA rearrangements in Jasminum andMenodora are confined to several regions of the genome.Gene duplications, insertions of highly repetitive DNA,and gene and intron losses are concentrated between therbcL and psaI genes and near clpP. These rearrangementsalso correlate with 4 large inversions in the LSC regions.Overall, the combination of these molecular evolutionaryevents makes the Jasmineae genomes distinct from all pre-viously examined land plant cp genomes. Duplicated inser-tions of coding or noncoding sequences into sites wheregenes or introns have been lost may compensate for theorganizational instability caused by these rearrangements.Alternatively, insertions of duplicated elements may becaused by gene or intron losses. Although the genome or-ganization of grasses and the Jasmineae is very similar, los-ses of major portions of accD and the clpP introns representparallel evolutionary events. This is also the case for theoccurrence of a large inversion that splits the trnK-UUUintron in Menodora and the fern Adiantum.
Supplementary Materials
Supplementary materials SM 1–SM 5 are availableat Molecular Biology and Evolution online (http://www.mbe.oxfordjournals.org/).
Acknowledgments
We thank several botanical gardens for providingplant material, including Kew Gardens, University of
Oleaceae Chloroplast Genome Rearrangements 1177
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

Connecticut, Fairchild Tropical Garden, The Royal BotanicGarden, Edinburgh, Arnold Arboretum, and SisanbanaTropical Garden. All DNA materials were obtained fromthe plant DNA Bank in Korea. We also thank Mary Gui-singer-Bellan, Katie Hansen, and 2 anonymous reviewersfor comments on an earlier version of the manuscript. Thisresearch was supported by a grant (C00449) from KoreaResearch Foundation to K.J.K. and a National ScienceFoundation grant (DEB-0120709) to R.K.J.
Funding to pay the Open Access publication chargesfor this article was provided by the BK21 School of LifeSciences and Biotechnology Initiative, Korea University,Seoul, Korea.
Literature Cited
Achaz G, Coissac E, Netter P, Rocha EPC. 2003. Associationsbetween inverted repeats and the structural evolution of bac-terial genomes. Genetics. 164:1279–1289.
Altschul S, Gish W, Miller W, Myers E, Lipman D. 1990. Basiclocal alignment search tool. J Mol Biol. 215:403–410.
Blanchard JL, Schmidt GW. 1995. Pervasive migration of organ-ellar DNA to the nucleus in plants. J Mol Evol. 41:397–406.
BowmanCM,BarkerRF,DyerTA.1988.InwheatctDNA,segmentsof ribosomal-protein genes are dispersed repeats, probably con-servedby nonreciprocal recombination.CurrGenet.14:127–136.
Bowman CM, Dyer TA. 1986. The location and possible evolu-tionary significance of small dispersed repeats in wheatctDNA. Curr Genet. 10:931–941.
Chumley TW, Palmer JD, Mower JP, Boore JL, Fourcade HM,Caile PJ, Jansen RK. Forthcoming 2006. The completed chlo-roplast genome sequence of Pelargonium x hortorum: organi-zation and evolution of the largest and most highly rearrangedchloroplast genome of land plants. Mol Biol Evol. 23:1–16.
Cosner ME, Jansen RK, Palmer JD, Downie SR. 1997. The highlyrearranged chloroplast genome of Trachelium caeruleum(Campanulaceae): multiple inversions, inverted repeat expan-sion and contraction, transposition, insertions/deletions, andseveral repeat families. Curr Genet. 31:419–429.
Cosner ME, Raubeson LA, Jansen RK. 2004. Chloroplast DNArearrangements in Campanulaceae: phylogenetic utility ofhighly rearranged genomes. BMC Evol Biol. 4:27.
Cummings MP, Nugent JM, Olmstead RG, Palmer JD. 2003. Phy-logenetic analysis reveals five independent transfers of thechloroplast gene rbcL to the mitochondrial genome in angio-sperms. Curr Genet. 43:131–138.
De Candolle AP. 1844. Ordo CXXVIII. Jasmineae. In: De Can-dolle A, editor. Prodromus systematis naturalis regni vegeta-bilis. Vol. 8. Paris: Fortin, Masson et sociorum. p. 300–316.
Downie SR, Katz-Downie DS, Rogers EJ, Zujewski HL, Small E.1998. Multiple independent losses of the plastid rpoC1 intronin Medicago (Fabaceae) as inferred from phylogenetic analy-ses of nuclear ribosomal DNA internal transcribed spacer se-quences. Can J Bot. 76:791–803.
Downie SR, Llanas E, Katz-Downie DS. 1996. Multiple indepen-dent losses of the rpoC1 intron in angiosperm chloroplastDNAs. Syst Bot. 21:135–151.
Downie SR, Olmstead RG, Zurawski G, Soltis DE, Soltis PS,Watson JC, Palmer JD. 1991. Six independent losses of thechloroplast DNA rpl2 intron in dicotyledons: molecular andphylogenetic implications. Evolution. 45:1245–1259.
Downie SR, Palmer JD. 1992. Use of chloroplast DNA rearrange-ments in reconstructing plant phylogeny. In: Soltis PS, SoltisDE, Doyle JJ, editors. Molecular systematics of plants. NewYork: Chapman and Hall. p. 14–35.
Doyle JJ, Davis JI, Soreng RJ, Garvin D, Anderson MJ. 1992.Chloroplast DNA inversions and the origin of the grass family(Poaceae). Proc Natl Acad Sci USA. 89:7722–7726.
Doyle JJ, Doyle JA. 1987. A rapid DNA isolation procedurefor small quantities of fresh leaf tissue. Phytochem Bull. 19:11–15.
Doyle JJ, Doyle JL, Ballenger JA, Palmer JD. 1996. The distribu-tion and phylogenetic significance of a 50-kb chloroplast DNAinversion in the flowering plant family Leguminosae. Mol Phy-logenet Evol. 5:429–438.
Doyle JJ, Doyle JL, Palmer JD. 1995. Multiple independent lossesof two genes and one intron from legume chloroplast genomes.Syst Bot. 20:272–294.
Fan WH, Woelfle MA, Mosig G. 1995. Two copies of a DNAelement, �Wendy�, in the chloroplast chromosome of Chlamy-domonas reinhardtii between rearranged gene clusters. PlantMol Biol. 29:63–80.
Felsenstein J. 1981. Evolutionary trees from DNA sequences:a maximum likelihood approach. J Mol Evol. 17:368–376.
Freyer R, Neckermann K, Maier RM, Kossel H. 1995. Structuraland functional analysis of plastid genomes from parasiticplants: loss of an intron within the genus Cuscuta. Curr Genet.27:580–586.
Fukuzawa H, Kohchi T, Ohyama K, Umesono K, Inokuchi H,Ozeki H. 1986. Coding sequences for the chloroplast ribo-somal protein S12 from the liverwort, Marchantia polymor-pha, are separated far apart on different DNA strands.FEBS Lett. 198:11–14.
Giese K, Subramanian AR, Larrinua IM, Bogorad L. 1987.Nucleotide sequence, promoter analysis, and linkage mappingof the unusually organized operon encoding proteins S7and S12 in maize chloroplasts. J Biol Chem. 262:15251–15255.
Goulding SE, Olmstead RG, Morden CW, Wolfe KH. 1996. Ebband flow of the chloroplast inverted repeat. Mol Gen Genet.252:195–206.
Green PS. 1997. A revision of the pinnate-leaved species of Jas-minum. Kew Bull. 52:933–947.
Green PS. 2001. Studies in the genus Jasminum, XVII: sectionsTrifoliolata and Primulina. Kew Bull. 56:903–915.
Green PS. 2004. Oleaceae. In: Kadereit JW, editor. Floweringplants, Dicotyledons: Lamiales (except Acanthaceae includingAvicenniaceae), Vol. 7. New York: Springer-Verlag. p. 296–306.
Hachtel W, Neuss A, Stein JV. 1991. A chloroplast DNA inver-sion marks an evolutionary split in the genus Oenothera. Evo-lution. 45:1050–1052.
Hansen AK, Gilbert LE, Simpson BB, Downie SR, Cervi AC,Jansen RK. 2006. Phylogenetic relationships and chromosomenumber evolution in Passifloria. Syst Bot. 31:138–150.
Hauser G, Olson R, Simon D, Johnson I, Sanders ER, Karol KG,McCourt RM, Zimmerly S. 2006. Origin and evolution of thechloroplast trnK (matK) intron: a model for evolution of groupII intron RNA structures. Mol Biol Evol. 23:380–391.
Heywood VH. 1978. Flowering plants of the world. New York:Mayflower Books. 335 p.
Higgins DG, Thompson JD, Gibson TJ. 1996. Using CLUSTALfor multiple sequence alignments. Methods Enzymol. 266:383–402.
Hiratsuka J, Shimada H, Whittier R, et al. (16 co-authors). 1989.The complete sequence of the rice (Oryza sativa) chloroplastgenome: intermolecular recombination between distinct tRNAgenes accounts for a major plastid DNA inversion during theevolution of the cereals. Mol Gen Genet. 217:185–194.
Hoot SB, Palmer JD. 1994. Structural rearrangements, includingparallel inversions, within the chloroplast genome of Anemoneand related genera. J Mol Evol. 38:274–281.
1178 Lee et al.
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

Jansen RK, Palmer JD. 1987. A chloroplast DNA inversion marksan ancient evolutionary split in the sunflower family (Astera-ceae). Proc Natl Acad Sci USA. 84:5818–5822.
Johnson LAS. 1957. A review of the family Oleaceae. Contr NewSouth Wales Natl Herb. 2:395–418.
Katayama H, Ogihara Y. 1996. Phylogenetic affinities of thegrasses to other monocots as revealed by molecular analysisof chloroplast DNA. Curr Genet. 29:572–581.
Kavvadas DS. 1956. Olive tree. In: Illustrated botanical-Phytological dictionary. Vol. 3. Kavvadas DS, editor. Athens(Greece): G. P. Xenos. p. 1369–1401. Greek.
Kim KJ, Choi KS, Jansen RK. 2005. Two chloroplast DNA inver-sions originated simultaneously during early evolution in thesunflower family. Mol Biol Evol. 22:1783–1792.
Kim YD, Jansen RK. 1994. Characterization and phylogeneticdistribution of a chloroplast DNA rearrangement in the Berber-idaceae. Plant Syst Evol. 193:107–114.
Kim KJ, Jansen RK. 1995. ndhF sequence evolution and the majorclades in the sunflower family. Proc Natl Acad Sci USA.92:10379–10383.
Kim KJ, Lee HL. 2004. Complete chloroplast genome sequencesfrom Korean ginseng (Panax schinseng Nees) and comparativeanalysis of sequence evolution among 17 vascular plants. DNARes. 11:247–261.
Knox EB, Downie SR, Palmer JD. 1993. Chloroplast genomerearrangements and the evolution of giant lobelias from herba-ceous ancestors. Mol Biol Evol. 10:414–430.
Knox EB, Palmer JD. 1999. The chloroplast genome arrangementof Lobelia thuliniana Lobeliaceae: expansion of the invertedrepeat in an ancestor of the Campanulales. Plant Syst Evol.214:49–64.
Koch MA, Dobes C, Matschinger M, Bleeker W, Vogel J, KieferM, Mitchell-Olds T. 2005. Evolution of the trnF(GAA) gene inArabidopsis relatives and the Brassicaceae family: monophy-letic origin and subsequent diversification of a plastidic pseu-dogene. Mol Biol Evol. 22:1032–1043.
Kumar S, Koichiro T, Jakobsen IB, Nei M. 2001. MEGA2: mo-lecular evolutionary genetics analysis software. Bioinfor-matics. 17:1244–1245.
Kurtz S, Choudhuri JV, Ohlebusch E, Schleiermacher C, Stoye J,Giegerich R. 2001. REPuter: the manifold applications of re-peat analysis on a genomic scale. Nucleic Acids Res. 29:4633–4642.
Lidholm J, Gustafsson P. 1991. A three-step model for the rear-rangement of the chloroplast trnK-psbA region of the gymno-sperm Pinus contorta. Nucleic Acids Res. 19:2881–2887.
Lowe TM, Eddy SR. 1997. tRNAscan-SE: a program for im-proved detection of transfer RNA genes in genomic sequence.Nucleic Acids Res. 25:955–964.
Maddison WP, Maddison DR. 2003. MacClade, v. 4.0 (releaseversion 4.06). Sunderland (MA): Sinauer Associates.
Maier RM, Neckermann K, Igloi GL, Kosel H. 1995. Completesequence of the maize chloroplast genome: gene content, hot-spots of divergence and fine tuning of genetic information bytranscript editing. J Mol Biol. 251:61–4628.
Martin W, Stoebe B, Goremykin V, Hansmann S, Hasegawa M,Kowallik K. 1998. Gene transfer to the nucleus and the evo-lution of chloroplasts. Nature. 393:162–165.
Matsuo M, Ito Y, Yamauchi R, Obokata J. 2005. The rice nucleargenome continuously integrates, shuffles, and eliminates thechloroplast genome to cause chloroplast-nuclear DNA flux.Plant Cell. 17:665–675.
Milligan BG, Hampton JN, Palmer JD. 1989. Dispersed repeatsand structural reorganization in subclover chloroplast DNA.Mol Biol Evol. 6:355–368.
Muller J. 1981. Fossil pollen records of extant angiosperms. BotRev. 47:1–142.
Ogihara Y, Terachi T, Sasakuma T. 1988. Intramolecular recom-bination of chloroplast genome mediated by short direct-repeatsequences in wheat species. Proc Natl Acad Sci USA.85:8573–8577.
Olmstead RG, Michaels HJ, Scott KM, Palmer JD. 1992. Mono-phyly of the Asteridae and identification of their major lineagesinferred from DNA sequences of rbcL. Ann Mo Bot Gard.79:249–265.
Olmstead RG, Palmer JD. 1992. A chloroplast DNA phylogeny ofthe Solanaceae: subfamilial relationships and character evolu-tion. Ann Mo Bot Gard. 79:346–360.
Palmer JD. 1986. Isolation and structural analysis of chloroplastDNA. In: Weissbach A, Weissbach H, editors. Methods inenzymology. Vol. 118. New York: Academic Press. p. 167–186.
Palmer JD. 1990. Contrasting modes and tempos of ge-nome evolution in land plant organelles. Trends Genet. 6:115–120.
Palmer JD. 1991. Plastid chromosomes: structure and evolution.In: Vasil IK, Bogorad L, editors. Cell culture and somatic cellgenetics in plants. Vol. 7A. The molecular biology of plastids.San Diego: Academic Press. p. 5–53.
Palmer JD, Downie SR, Nugent JM, Brandt R, Unseld M,Brennicke A, Schuster W, Borner T. 1994. The chloroplastand mitochondrial DNAs of Arabidopsis thaliana: conven-tional genomes in an unconventional plant. In: SomervilleC, Meyerowitz E, editors. Arabidopsis. Cold Spring Harbor,New York: Cold Spring Harbor Press. p. 37–67.
Palmer JD, Nugent JM, Herbon LA. 1987. Unusual structureof geranium chloroplast DNA: a triple-sized inverted repeat,extensive gene duplications, multiple inversions, and tworepeat families. Proc Natl Acad Sci USA. 84:769–773.
Palmer JD, Osorio B, Aldrich J, Thompson WF. 1987. ChloroplastDNA evolution among legumes: loss of a large inverted repeatoccurred prior to other sequence rearrangements. Curr Genet.11:275–286.
Palmer JD, Osorio B, Thompson WF. 1988. Evolutionary signif-icance of inversions in legume chloroplast DNAs. Curr Genet.14:65–74.
Palmer JD, Thompson WF. 1981. Rearrangements in the chloro-plast genomes of mung bean and pea. Proc Natl Acad Sci USA.78:5533–5537.
Perry AS, Brennan S, Murphy DJ, Wolfe KH. 2002. Evolutionaryre-organization of a large operon in Adzuki bean chloroplastDNA caused by inverted repeat movement. DNA Res.9:157–162.
Pombert JF, Lemieux C, Turmel M. 2006. The complete chloro-plast DNA sequence of the green alga Oltmannsiellopsisviridis reveals a distinctive quadripartite architecture inthe chloroplast genome of early diverging ulvophytes. BMCBiol. 4:3.
Pombert JF, Otis C, Lemieux C, Turmel M. 2005. The chloroplastgenome sequence of the green alga Pseudendoclonium akine-tum (Ulvophyceae) reveals unusual structural features and newinsights into the branching order of chlorophyte lineages. MolBiol Evol. 22:1903–1918.
Posada D, Crandall KA. 1998. Modeltest: testing the model ofDNA substitution. Bioinformatics. 14:817–818.
Prombona A, Subramanian AR. 1989. A new rearrangement ofangiosperm chloroplast DNA in rye (Secale cereale) involvingtranslocation and duplication of the ribosomal rpS15 gene.J Biol Chem. 264:19060–19065.
Quigley F, Weil JH. 1985. Organization and sequence of 5transfer-RNA genes and of an unidentified reading frame inthe wheat chloroplast genome—evidence for gene rearrange-ments during the evolution of chloroplast genomes. CurrGenet. 96:495–503.
Oleaceae Chloroplast Genome Rearrangements 1179
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018

Raubeson LA, Jansen RK. 1992. A rare chloroplast DNA struc-tural mutation is shared by all conifers. Biochem Syst Ecol.20:17–24.
Raubeson LA, Jansen RK. 2005. Chloroplast genomes of plants.In: Henry R, editor. Diversity and evolution of plants—genotypic and phenotypic variation in higher plants.Oxfordshire: CABI Publishing. p. 45–68.
Rehder A. 1940. Manual of cultivated trees and shrubs. 2nd ed.New York: McMillan. p. 452–481.
Rocha EPC. 2003. DNA repeats lead to the accelerated loss ofgene order in bacteria. Trends Genet. 19:600–603.
Rohwer JG. 1994. Seed characters in Jasminum (Oleaceae): un-expected support for De Candolles’s section. Bot Jahrb Syst.116:299–319.
Rohwer JG. 1995. Seed characters in Jasminum (Oleaceae). II.Evidence from additional species. Bot Jahrb Syst. 117:299–315.
Sanderson MJ. 2002. Estimating absolute rates of molecular evo-lution and divergence times: a penalized likelihood approach.Mol Biol Evol. 19:101–109.
Sanderson MJ. 2003. r8s: inferring absolute rates of molecularevolution and divergence times in the absence of a molecularclock. Bioinformatics. 19:301–302.
Saski C, Lee SB, Daniell H, Wood TC, Tomkins J, Kim HG,Jansen RK. 2005. Complete chloroplast genome sequenceof Glycine max and comparative analyses with other legumegenomes. Plant Mol Biol. 59:309–322.
Shinozaki K, Ohime M, Tanaka M, et al. (23 co-authors). 1986.The complete nucleotide sequence of tobacco chloroplastgenome: its gene organization and expression. EMBO J. 5:2043–2049.
Stefanovic S, Olmstead G. 2005. Down the slippery slope: plastidgenome evolution in Convolvulaceae. J Mol Evol. 61:292–305.
Steyermark JA. 1932. A revision of the genus Menodora. Ann MoBot Gard 19:87–176.
Strauss SH, Palmer JD, Howe GT, Doerksen AH. 1998. Chloro-plast genomes of two conifers lack a large inverted repeat andare extensively rearranged. Proc Natl Acad Sci USA. 85:3898–3902.
Swofford DL. 2002. PAUP: phylogenetic analysis using parsi-mony and other methods. Version 4.0. Sunderland (MA):Sinauer Associates.
Takahashi M, Crane PR, Manchester SR. 2002. Hironoia fusifor-mis gen. et sp. nov.: a cornalean fruit from the Kamikitaba
locality (Upper Cretaceous, Lower Coniacian) in northeasternJapan. J Plant Res. 115:463–473.
Taylor H. 1945. Cyto-taxonomy and phylogeny of the Oleaceae.Brittonia. 5:337–367.
Torazawa K, Hayashida N, Obokata J, Shinozaki K, Sugiura M.1986. The 5# part of the gene for ribosomal protein S12 islocated 30 kbp downstream from its 3# part in tobacco chlo-roplast genome. Nucleic Acids Res. 14:3143.
Turmel M, Otis C, Lemieux C. 2002. The chloroplast and mito-chondrial genome sequences of the charophyte Chaetosphaeri-dium globosum: insights into the timing of the events thatrestructured organelle DNAs within the green algal lineage thatled to land plants. Proc Natl Acad Sci USA. 99:11275–11280.
Turmel M, Otis C, Lemieux C. 2005. The complete chloroplastDNA sequences of the charophycean green algae Staurastrumand Zygnema reveal that the chloroplast genome underwentextensive change during the evolution of Zygnematales.BMC Biol. 3:22.
Turner BL. 1991. An overview of the North American species ofMenodora (Oleaceae). Phytologia. 71:340–356.
Wakasugi T, Tsudzuki J, Ito S, Nakashima K, Tsudzuki T, SugiuraM. 1994. Loss of all ndh genes as determined by sequencingthe entire chloroplast genome of the black pine Pinus thunber-gii. Proc Natl Acad Sci USA. 91:9794–9798.
Wallander E, Albert VA. 2000. Phylogeny and classification of theOleaceae based on rps16 and trnL-F sequence data. Am J Bot.87:1827–1841.
Wolf PG, Rowe CA, Sinclair RB, Hasebe M. 2003. Completenucleotide sequence of the chloroplast genome from a lepto-sporangiate fern, Adiantum capillus-veneris L. DNA Res. 10:59–65.
Wolfe KH, Morden CW, Palmer JD. 1992. Function and evolutionof a minimal plastid genome from a nonphotosynthetic para-sitic plant. Proc Natl Acad Sci USA. 89:10648–10652.
Wyman SK, Jansen RK, Boore JL. 2004. Automatic annotation oforganellar genomes with DOGMA. Bioinformatics. 20:3252–3255.
Zuker M. 2003. Mfold web server for nucleic acid folding andhybridization prediction. Nucleic Acids Res. 31:3406–3415.
Charles Delwiche, Associate Editor
Accepted February 18, 2007
1180 Lee et al.
Downloaded from https://academic.oup.com/mbe/article-abstract/24/5/1161/1039086by gueston 04 April 2018