gen selden, sigma xi 2015
TRANSCRIPT

IDENTIFICATION OF GENETIC REGIONS IN THE YUK OPERON OF BACILLUS SUBTILIS THAT ARE DIFFERENTIALLY REQUIRED FOR SECRETION OF YUKE, A HOMOLOG TO THE VIRULENCE FACTOR, ESXA, IN MYCOBACTERIUM TUBERCULOSIS
Gen Selden Pine Crest School Harvard University

Infectious Diseases
Infectious diseases are caused by pathogenic organisms such as bacteria, viruses, parasites, and fungi.
They can be spread through human interaction, through contact with animals or insects, or through contaminated food and water.
Infectious diseases kill more people worldwide than any other single cause.

Tuberculosis
Tuberculosis is a major infectious disease that affects people world wide.
According to the World Health Organization, it is the second greatest killer worldwide due to a single infectious agent (WHO, 2014). In 2012 8.6 million people were infected
with tuberculosis, and 1.3 million of those people died from the infection.
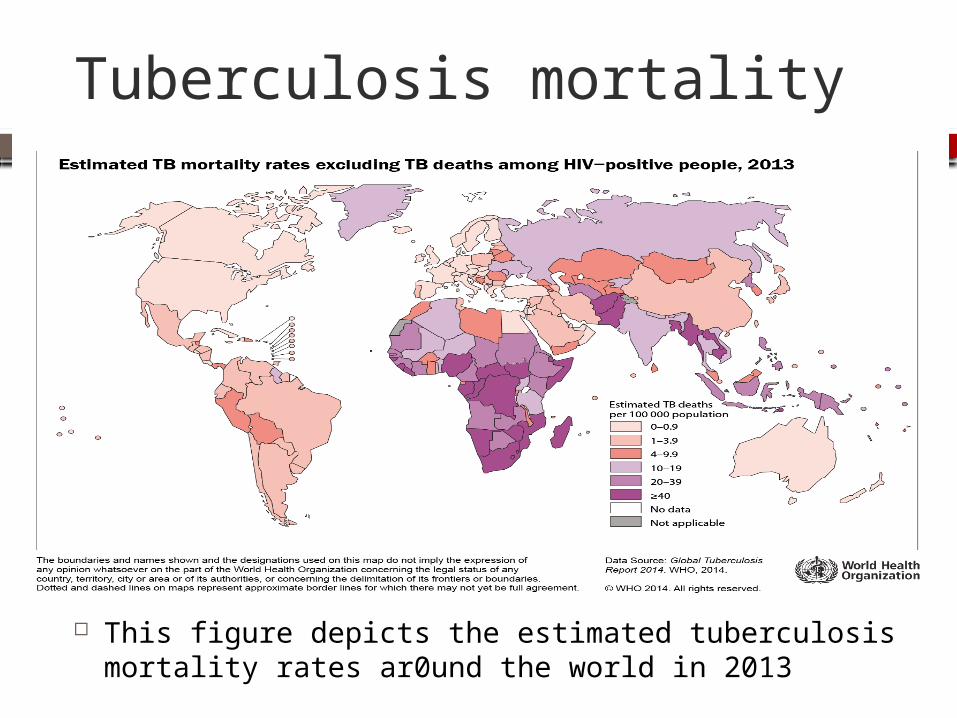
Tuberculosis mortality
This figure depicts the estimated tuberculosis mortality rates ar0und the world in 2013
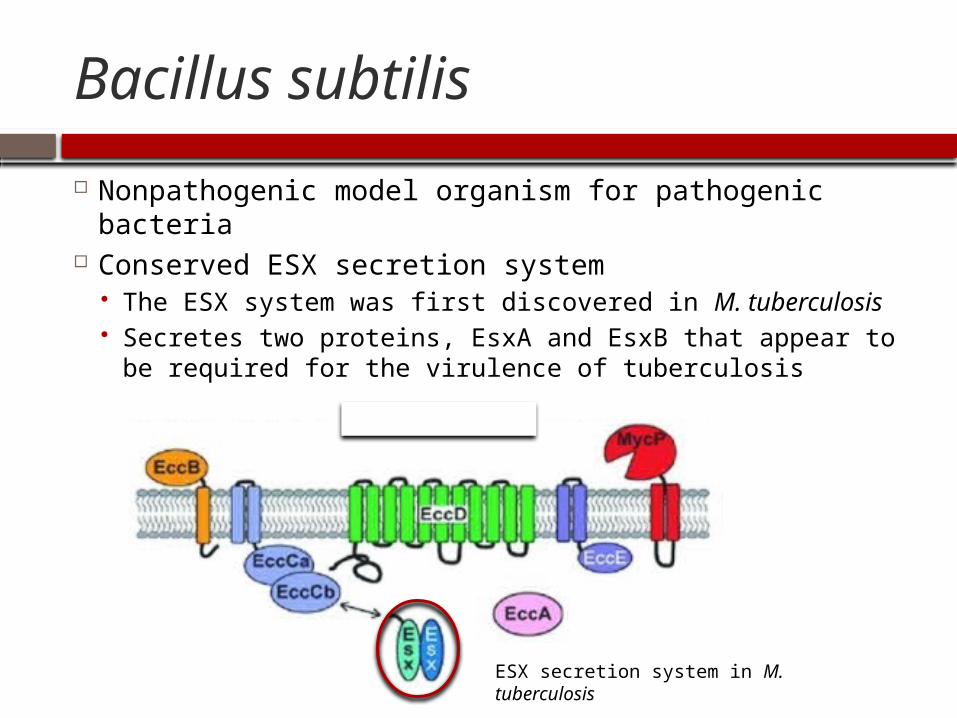
Bacillus subtilis
Nonpathogenic model organism for pathogenic bacteria
Conserved ESX secretion system The ESX system was first discovered in M. tuberculosis Secretes two proteins, EsxA and EsxB that appear to be
required for the virulence of tuberculosis
ESX secretion system in M. tuberculosis
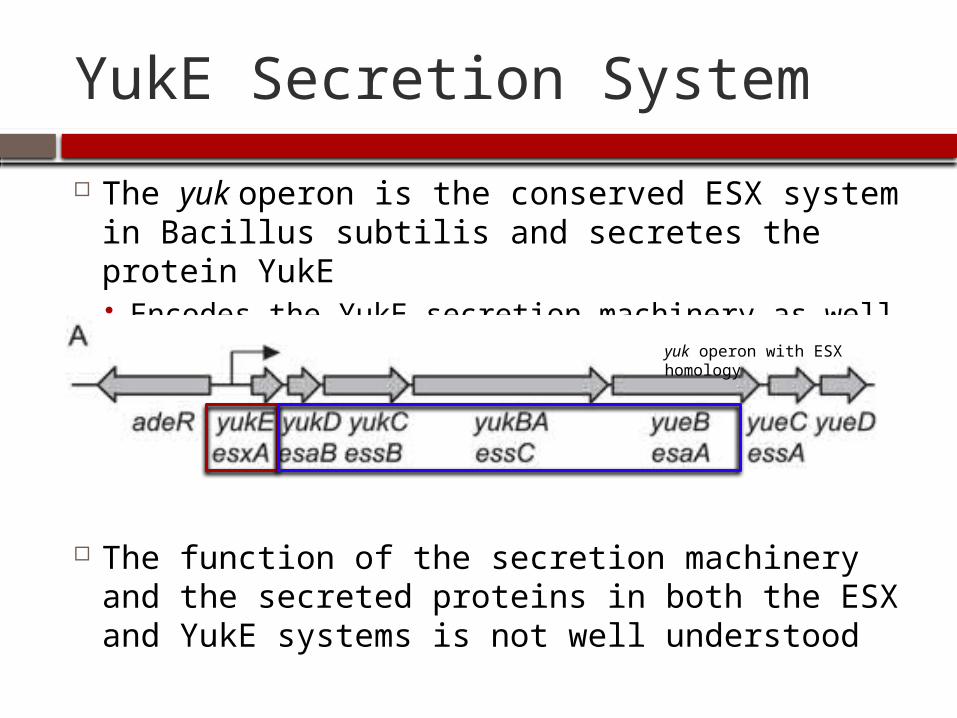
YukE Secretion System
The yuk operon is the conserved ESX system in Bacillus subtilis and secretes the protein YukE Encodes the YukE secretion machinery as well
The function of the secretion machinery and the secreted proteins in both the ESX and YukE systems is not well understood
yuk operon with ESX homology

Bacillus subtilis
4 different genetic backgrounds of B. subtilis:
3610 – “wild-type” Previous research showed that
YukE secretion was independent of the secretion machinery
PY79 – “domesticated” Previous research showed that
YukE secretion was dependent on the secretion machinery
168 and 3610 cured – intermediate Have not yet been analyzed for
differences in secretion
3610
168
PY79
3610 cured
Plasmid removal
Genetic alteration
Genetic alteration

Genetic alterations
3610 is considered “wild type” because it is genetically closest to Bacillus subtilis that would be found in the soil
3610 cured was created by removing a plasmid from the genome of 3610 because it was suspected that this plasmid encoded an alternate secretion system that could secrete YukE
Small deletions and insertions occurred in 3610 and resulted in the formation of 168
Two large insertions and four large deletions in the 168 genome resulted in the formation of PY79

Purpose
“Knowledge of MTBC virulence factors is essential for the development of new vaccines and drugs to help manage the disease toward an increasingly more tuberculosis-free world.” (Forrellad et al.)
The purpose of my research was to analyze differences in YukE secretion for variations in molecular signatures in each of the four B. subtilis backgrounds

Methods
Secretion assay Cultures grown in LB media at 37oC Cells were collected and normalized based on cell density measured at OD600nm Cell pellet and supernatant were separated
Protein precipitation Proteins in the supernatant were precipitated using trichloroacetic acid Centrifugation at 4oC at 16,000 rpm separated the proteins and the remaining
liquid Cell lysates
Frozen cell pellets were lysed with lysis buffer and heated at 80oC to release the proteins within the cell
Semi-dry method of western blotting Secretion was observed by blotting the proteins in the cell pellet and the
secreted proteins and probing for YukE Probing for the cytosolic protein, SigA, served as a lysis and loading control to
ensure that the detection of secreted YukE was not due to cell lysis Blots were exposed to chemiluminescence to view the protein bands

Bacillus subtilis
Wild type, single deletion strains, strains with the operon removed, strains with YukE inserted at an endogenous location, and strains that were a combination of the two were compared for YukE secretion
3610 wt
3610 ΔyukE
3610 ΔyukD
3610 ΔyukC
3610 ΔyukBA
3610 ΔyukEDCBAyueB
3610 amyE::yukE
3610 ΔyukEDCBAyueB; amyE::yukE
3610 cured wt
3610 cured ΔyukE
3610 cured ΔyukD
3610 cured ΔyukC
3610 cured ΔyukBA
3610 cured ΔyukEDCBAyueB
3610 cured yhDGH::yukE
3610 cured ΔyukEDCBAyueB; yhDGH::yukE
168 wt
168 ΔyukE
168 ΔyukD
168 ΔyukC
168 ΔyukBA
168 ΔyukEDCBAyueB
168 yhDGH::yukE
168 ΔyukEDCBAyueB; yhDGH::yukE
PY79 wt
PY79 ΔyukE
PY79 ΔyukD
PY79 ΔyukC
PY79 ΔyukBA
PY79 ΔyukEDCBAyueB
PY79 amyE::yukE
PY79 ΔyukEDCBAyueB; amyE::yukE
*
* *
* *
yuk
amyE
yukE
yhDGH
yukE
*
*
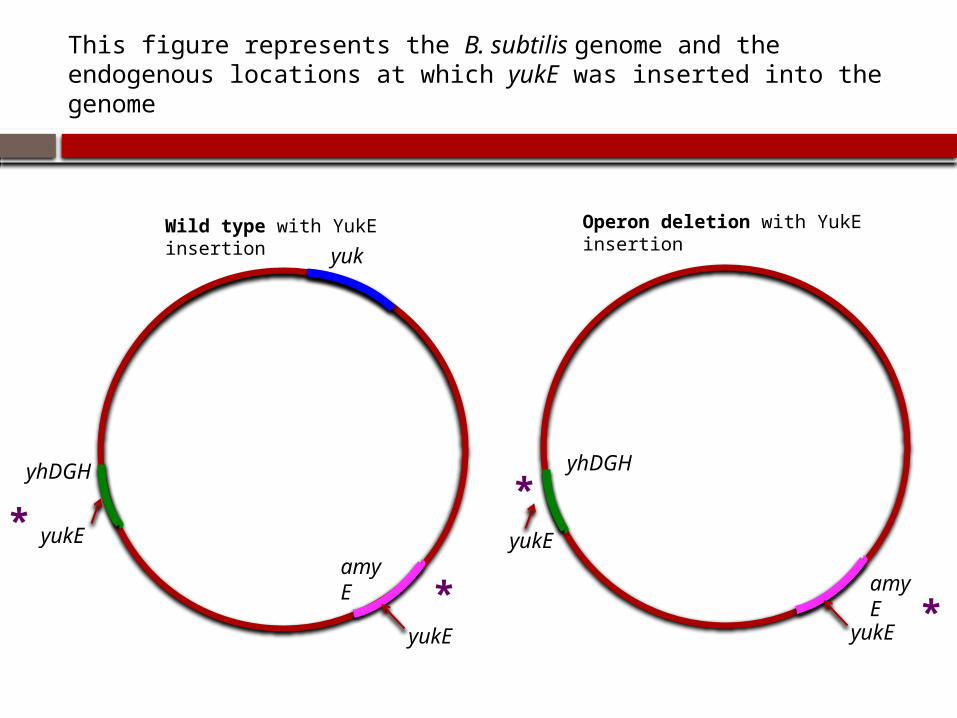
This figure represents the B. subtilis genome and the endogenous locations at which yukE was inserted into the genome
Wild type with YukE insertion
amyE
yukE
yhDGH
yukE
*
*
Operon deletion with YukE insertion
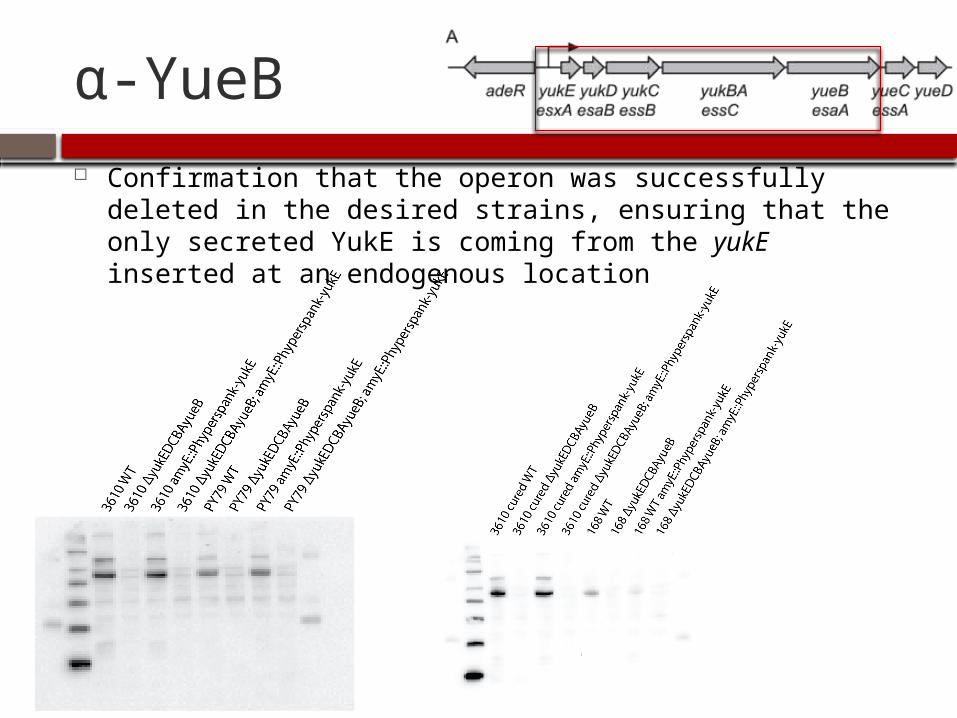
α-YueB Confirmation that the operon was successfully deleted in
the desired strains, ensuring that the only secreted YukE is coming from the yukE inserted at an endogenous location

Additional Information
SigA is a cytosolic protein and was used as a loading and lysis control to ensure secretion results are not due to cell lysis.
The pellet samples were used to observe and confirm production of YukE within the cells
However, the focus of the results is on the secreted YukE, since YukE is homologous to the secreted M. tuberculosis virulence factor, ESXA.

3610/PY79 Secretion
As previous studies indicated, 3610 deletion strains secreted YukE even with single components of the YukE secretion machinery not present, confirming its independence of the operon for YukE secretion.
However, PY79 exhibited low levels of secretion when components of the secretion machinery were deleted, suggesting a strong dependence on the presence of the operon for secretion.

After the first secretion assay The next step was to analyze the intermediate
strains, 3610 cured and 168, for YukE secretion in order to determine which two of the four B. subtilis backgrounds have the most similar secretion patterns.
In the future, the wild type genomes of the two similar B. subtilis backgrounds identified in this study can be analyzed for specific genetic differences that may be responsible for the observed differences in secretion.
Eventually, due to the homology between B. subtilis and M. tuberculosis, the goal would be to apply this knowledge of genetic differences to M. tuberculosis in order to further research on new drugs to fight the tuberculosis disease.

3610 cured/168 Secretion
I found that 3610 cured showed similar YukE secretion patterns to those of 3610, suggesting only a slight dependence on the presence of the secretion machinery for YukE secretion.
In addition, 168, which is genetically more similar to PY79 than either 3610 or 3610 cured, exhibited similar YukE secretion patterns to those of PY79, suggesting a similar, strong dependence on the secretion machinery for YukE secretion.
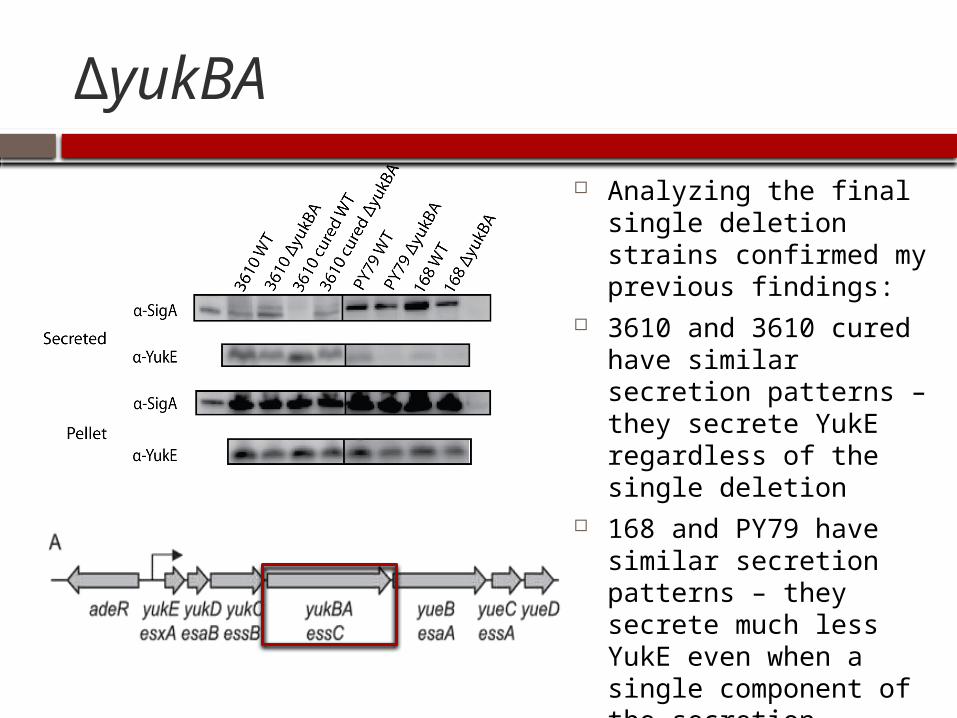
ΔyukBA
Analyzing the final single deletion strains confirmed my previous findings:
3610 and 3610 cured have similar secretion patterns – they secrete YukE regardless of the single deletion
168 and PY79 have similar secretion patterns – they secrete much less YukE even when a single component of the secretion machinery is not present

After analyzing single deletions The next step was to analyze YukE secretion
in 3 different strains: deletion of the entire yuk operon, insertion of yukE at an endogenous location, and the combination of these two strains – deletion of the yuk operon complemented with yukE insertion.
These results better represent the dependence of 3610, 3610 cured, 168, and PY79 on the yuk secretion machinery for secretion of YukE
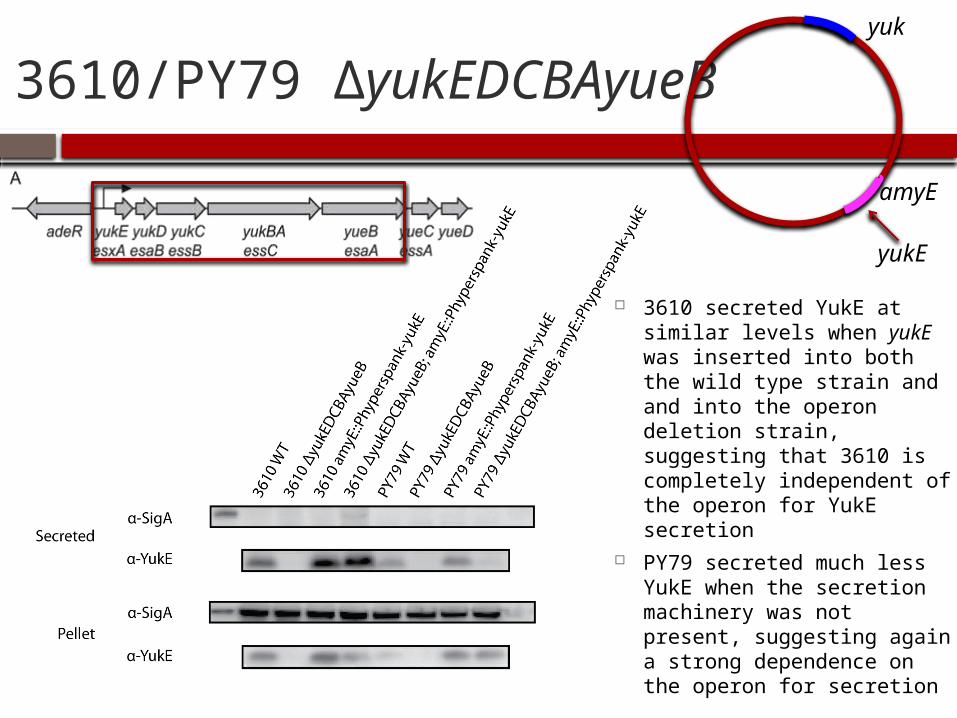
3610/PY79 ΔyukEDCBAyueB
amyEyukE
3610 secreted YukE at similar levels when yukE was inserted into both the wild type strain and and into the operon deletion strain, suggesting that 3610 is completely independent of the operon for YukE secretion
PY79 secreted much less YukE when the secretion machinery was not present, suggesting again a strong dependence on the operon for secretion
yuk
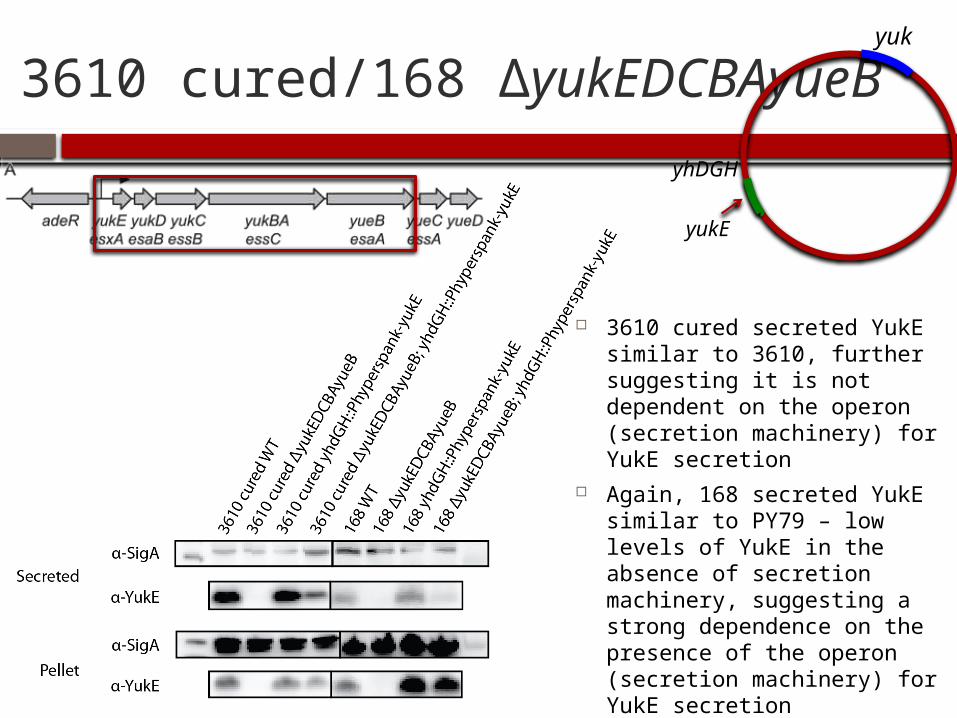
3610 cured/168 ΔyukEDCBAyueByuk
yhDGH
yukE
3610 cured secreted YukE similar to 3610, further suggesting it is not dependent on the operon (secretion machinery) for YukE secretion
Again, 168 secreted YukE similar to PY79 – low levels of YukE in the absence of secretion machinery, suggesting a strong dependence on the presence of the operon (secretion machinery) for YukE secretion

Spβ Phage
After analyzing the results and observing the similarities between YukE secretion in 168 and YukE secretion in PY79, I decided to analyze the effect of the Spβ phage on secretion
The Spβ phage is present in 3610, 3610 cured, and 168, but is absent in PY79
It was speculated that the phage might be creating holes in the cell wall, allowing YukE to leak out of the cells in the absence of the yuk secretion machinery, which could explain some of the observed secretion patterns
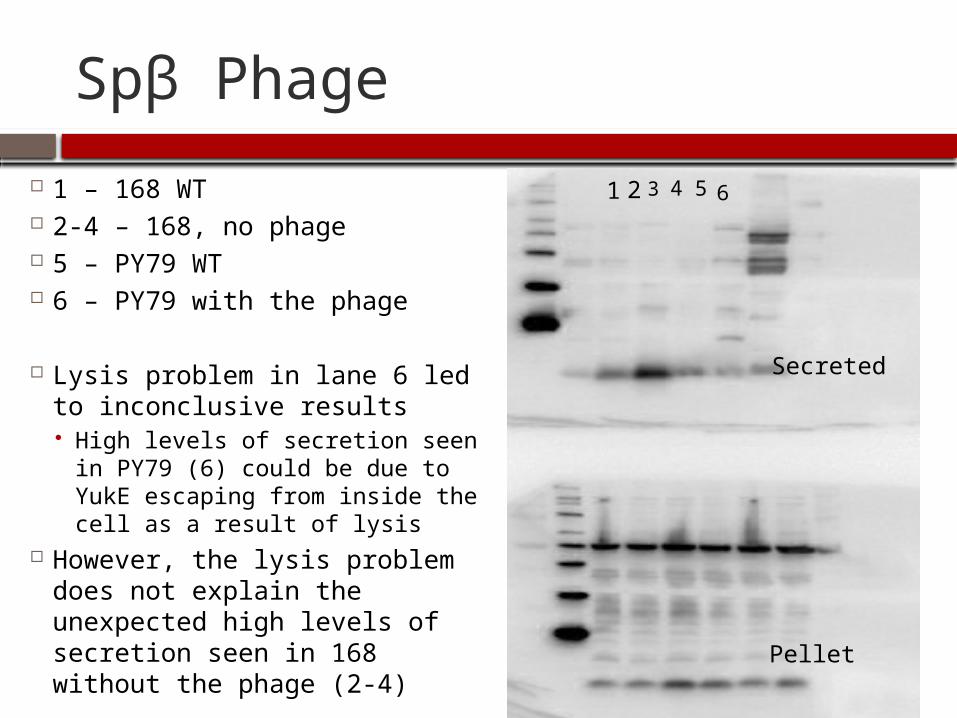
Spβ Phage
1 – 168 WT 2-4 – 168, no phage 5 – PY79 WT 6 – PY79 with the phage
Lysis problem in lane 6 led to inconclusive results High levels of secretion seen in
PY79 (6) could be due to YukE escaping from inside the cell as a result of lysis
However, the lysis problem does not explain the unexpected high levels of secretion seen in 168 without the phage (2-4)
1 2 3 4 5 6
Secreted
Pellet

Discussion
3610 secretes YukE independently of the operon
PY79 exhibits strong dependence on the presence of the operon for YukE secretion
3610
168
PY79
3610 cured
3610 cured secretes YukE independently of the operon The plasmid in 3610 is not responsible for YukE
secretion 168 secretes YukE similarly to Py79
Plasmid removalGenetic
alteration
Genetic alteration

Future research
The wild type genomes of 168 and PY79 should be analyzed for genetic differences in the future
In addition, more research should be done on possible alternate routes for YukE secretion, such as the SP phage

Future Research
Determination of the aforementioned genetic differences in 168 and PY79 may be able to help researchers target these areas in pathogenic bacteria such as M. tuberculosis in order to inhibit or reduce secretion of the virulent proteins
Specific inhibition of or reduction in virulence could contribute to the creation of new drugs to fight the disease

References1. World Health Organization (2014) Tuberculosis2. Chan, ED., Iseman MD., (2002) Current medical treatment for tuberculosis. BMJ. 325(7375):1282-12863. Cosgrove SE., Sakoulas G., Perencevich EN., Schwaber MJ., Karchmer AW., Carneli Y., (2002) Comparison of Mortality Associated with Methicillin-
Resistant and Methicillin-Susceptible Staphylococcus aureus Bacteremia: A Meta-analysis. Clinical Infectious Diseases. 36:53-94. Abdallah AM., Gey van Pittius NC., Champion PA., Cox J., Luirink J., Vandenbroucke-Grauls CM., Appelmelk BJ., Bitter W., (2007) Type VII secretion—
mycobacteria show the way. Nat Rev Microbiol. 5(11):883-915. Zoltner M., Fyfe PK., Palmer T., Huner WN., (2013) Characterization of Staphylococcus aureux EssB, an integral membrane component of the Type VII
secretion system: atomic resolution crystal structure of the cytoplasmic segment. Biochem J. 449(2):469-776. Garufi G., Butler E., Missiakas D., (2008) ESAT-6-like protein secretion in Bacillus anthracis. J Bacteriol. 190(21):7004-117. Huppert LA., Ramsdell TL., Chase MR., Sarracino DA., Fortune SM., Burtton BM., (2014) The ESX System in Bacillus subtilis Mediates Protein
Secretion. Plos One. DOI: 10.1371/journal.pone.00962678. Pallen MJ., (2002) The ESAT-6/WXG100 superfamily – and a new Gram-positive secretion system? Trends in Microbiology 10(5):209-129. Burts ML., Williams WA., DeBord K., Missiakas DM., (2005) EsxA and EsxB are secreted by an ESAT-6-like system that is required for the pathogenesis
of Staphylococcus aureus infections. Proc Natl Acad Sci USA. 102(4): 1169-117410. Champion PAD., Stanley SA., Champion MM., Brown EJ., Cox JS., (2006) C-Terminal Signal Sequence Promotes Virulence Factor Secretion in
Mycobacterium tuberculosis. Science. 313(5793): 1632-163611. Gao LY., Guo S., McLaughlin B., Morisaki H., Engel JN., Brown EJ., (2004) A mycobacterial virulence gene cluster extending RD1 is required for
cytolysis, bacterial spreading and ESAT-6 secretion. Molecular Microbiology. 53(6):1677-169312. Hsu T., Hingley-Wilson SM., Chen B., Chen M., Dai AZ., Morin PM., Marks CB., Padiyar J., Goulding C., Gingery M., Eisenberg D., Russell RG., Derrick
SC., Collins FM., Morris SL., King CH., Jacobs WR. Jr., (2003) The primary mechanism of attenuation of bacillus Calmette-Guérin is a loss of secreted lytic function required for invasion of lung interstitial tissue. Proc Natl Acad Sci USA. 100(21):12420-12425
13. Burts ML., Williams WA., Debord K., Missiakas DM., (2004) EsxA and EsxB are secreted by an ESAT-6-like system that is required for the pathogenesis of Staphylococcus aureus infections. PNAS. 102(4)1169-1174
14. Garufi G., Butler E., Missiakas D., (2008) ESAT-6-Like Protein Secretion in Bacillus anthracis. Journal of Bacteriology. 190(21):7004-701115. Barns KJ., Weisshaar JC., (2013) Real-time Attack of LL-37 on Single Bacillus subtilis Cells. Biochim Biophys Acta. 1828(6):1511-152016. Cole ST., (2002) Comparative and functional genomics of the Mycobacterium tuberculosis complex. Microbiology. 148(10):2919-2928. 17. Harwood, C., and S. M. Cutting (ed.) (1990) Molecular biological methods for Bacillus. John Wiley & Sons, Ltd., Chichester, United Kingdom.18. Forrellad MA., Klepp LI., Gioffré A., Sabio y García J., Morbidoni HR., de la Paz Santangelo M., Cataldi AA., Bigi F., (2013) Virulence factors of the
Mycobacterium tuberculosis complex. Virulence. 4(1):3-6619. Zeigler DR., Prágai Z., Rodrigues S., Chevreux B., Muffler A., Albert T., Bai R., Wyss M., Perkins JB., (2008) The Origins of 168, W23, and Other
Bacillus subtilis Legacy Strains. Journal of Bacteriology. 190(21):6983-699520. Konkol MA., Blair KM., Kearns DB., (2013) Plasmid-Encoded ComI Inhibits Competence in the Ancestral 3610 Strain of Bacillus subtilis. Journal of
Bacteriology. 195(18):4085-409321. Huppert L., (2010) Localization, Regulation, and Function of the First Type VII Protein Secretion System in Bacillus subtilis. Harvard University22. World Health Organization, (2013) Tuberculosis Control 2013. Geneva23. Kinhikar AG., Verma I., Chandra D., Singh KK., Weldingh K., Hsu T., Jacobs WR Jr., Laal S., (2010) Potential role for ESAT6 in dissemination of M.
tuberculosis via human lung epithelial cells. Microbiology. 75(1):92-10624. Chen R., Guttenplan SB., Blair KM., Kearns DB., (2009) Role of the σD-Dependent Autolysins in Bacillus subtilis Population Heterogeneity. Journal of
Bacteriology. 191(18):5775-5784
Acknowledgements
Dr. Briana Burton, Associate Professor of Molecular and Cellular Biology, Harvard University
Bram Sterling, Graduate Student, Harvard University
The Burton Lab Jennifer Gordinier, Pine Crest School