gata elements are necessary for the activity and tissue specificity
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY, June 1994, p. 4286-4294 Vol. 14, No. 60270-7306/94/$04.00+0
GATA Elements Are Necessary for the Activity and TissueSpecificity of the T-Cell Receptor Beta-Chain
Transcriptional EnhancerA. J. HENDERSON,' S. McDOUGALL,2 J. LEIDEN,3 AND K. L. CALAMEl*
Department of Microbiology, College of Physicians and Surgeons, Columbia University, New York, New York 10032';Geriatric Research, Education and Clinical Center, West Los Angeles Veterans Administration Medical Center,
Los Angeles, California 90073, and Department of Medicine, UCLA School of Medicine, Los Angeles,Califomia 900242; and Department of Medicine, University of Chicago, Chicago, Illinois 606373
Received 29 December 1993/Returned for modification 31 January 1994/Accepted 8 March 1994
Three high-affinity binding sites for the GATA family of transcriptional regulators have been identifiedwithin the T-cell receptor beta-chain (TCRO) transcriptional enhancer, and their functional significance hasbeen determined in an effort to understand the T-cell specificity of the enhancer more fully. One site, TE4, isimportant for activity of the enhancer in T cells. Neither site TE1 nor site TE2 can functionally replace amutated TE4 site in T cells; however, the same protein, probably GATA-3, binds all three sites, as judged byelectrophoretic mobility shift, oligonucleotide competition, and proteolytic clipping assays. These data suggestthat additional proteins are critical for the ability of GATA-3 to activate the TCRi enhancer. In fibroblasts,the GATA sequence at site TE1 appears to bind a negative regulator. Since this is not true in B cells, B cellsand fibroblasts appear to have different mechanisms for negative regulation of the TCROi enhancer.
The ot3 T-cell receptor (TCR) is the major clonotypicantigen-specific receptor for T lymphocytes. Proper expressionof TCR ot and ,B chains is critical for normal T-cell develop-ment, since the receptor is required for positive and negativeselection of developing thymocytes and for activation of pe-ripheral T cells (5, 55). The genes encoding the ao and L chainsare similar to immunoglobulin genes with respect to organiza-tion and DNA rearrangement (5, 55). Furthermore, the samerecombination machinery appears to be used for assemblingTCR gene segments in T cells and immunoglobulin genesegments in B cells (47, 59). The T-cell specificity of rearrange-ment and expression of the TCRao and -P genes is in partregulated at the level of transcription (2, 57). TCR, genetranscription appears to be primarily controlled by a T-cell-specific transcriptional enhancer located 3' of the C,B2 codingregion (12, 21, 32, 43, 49). Protein-binding sites have beenidentified in the TCRI enhancer, and the functional signifi-cance of some sites has been established (26); however, themolecular mechanisms which confer T-cell specificity are notwell understood.GATA-3 is a transcription factor which has been shown to
be important for determining the T-cell specificity of theTCRa and -8 genes (15, 18, 20, 29). It is a member of theGATA family of transcription factors that recognize the con-sensus sequence WGATAR through a highly conserved C4zinc finger domain (39). In vertebrates, four GATA members,GATA-1 through GATA-4, have been identified, and it hasbeen shown that these proteins have somewhat overlappingbut distinctive patterns of expression (39). The best-character-ized member of this family is GATA-1, which is expressed inthe erthyroid, megakaryocytic, mast cell, and germ cell lineages(7, 8, 30, 46, 51). GATA-1 has been shown to regulate anumber of erthryoid cell-specific genes and to be required for
* Corresponding author. Mailing address: Department of Microbi-ology, College of Physicians & Surgeons, Columbia University, 701 W.168th St., New York, NY 10032. Phone: (212) 305-3504. Fax: (212)305-1468.
normal erythropoiesis (41). Another family member, GATA-3,is the predominant GATA protein present in T cells, althoughhigh levels of GATA-3 are also detected in the developingcentral nervous system (15). GATA-3 mRNA is detectableearly in thymus ontogeny, being present at day 12.5 in devel-oping embryos (26). A number of enhancers that regulateT-cell-specific genes, including CD38 and the TCRat, -P, --y,and -8 genes, contain GATA sequences (26). These observa-tions have led to the suggestion that GATA-3, like GATA-1,may be important for regulating lineage-specific gene expres-sion and restricting hematopoietic cells to a particular devel-opment fate.The role of GATA-3 in determining the T-cell specificity of
the TCRI enhancer is not known, although several regionswith good matches to the GATA consensus are present withinthe enhancer (12, 29). In this study, GATA-3-binding siteshave been identified and their functional importance has beendetermined by mutational analysis. Three GATA-containingsites, TE1, TE2, and TE4, were shown to be recognized byproteins from crude T-cell nuclear extracts and to bind recom-binant GATA-3 with high affinity. The GATA sequence at siteTE4 is critical for the activity of the enhancer in T cells. SitesTE1 and TE2 are not functionally equivalent to TE4 and areunable to compensate for a mutated TE4 site despite evidencethat indistinguishable proteins bind sites TE1 and TE4. Inaddition, site TE1, but not site TE2 or TE4, was shown tomediate negative regulation of the TCRP enhancer in fibro-blasts. Thus, the activation capability of GATA-3 for theTCRI enhancer in T cells and the repressor activity of aGATA-binding protein in fibroblasts are both context depen-dent. These findings suggest that protein-protein interactionsare critical for determining the action of GATA-bindingproteins in the TCR3 enhancer.
MATERIALS AND METHODS
EMSAs and Op-Cu footprinting. Nuclear extracts fromvarious cell lines were prepared as previously described (6, 48).
4286
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 20
Feb
ruar
y 20
22 b
y 21
1.10
8.93
.162
.

GATA ELEMENTS IN THE TCR3 ENHANCER 4287
TABLE 1. Oligonucleotides used in this studyName Sequence'
MTE1 ......... 5' GACACACACATTGTACTATACTTACAAGGCATCTG 3'MTE2 ......... 5' CAAAGAGGCCTAGGAACATCTAGAAAAA 3'MTE4 ......... 5' GGTCTGGCTGTTTGCATGTAAGTAACATCAGC 3'pUC primer 1 ......5' GTAAAACGACGGCCAG 3'pUC primer 2 ......5' AACAGCTATGACCATG 3'GATA site A.......5' GATCTATCTTACA 3'GATA site B .......5' ATAGAATGTCTAG 3'CRE site A ......... 5' ACGAGTGACGTC 3'CRE site B......... 5' CGTTGACGTACT 3'TalA ......... 5' CTCCCATTTCCATGACGTCATGGTTACCA 3'TalB......... 5' TGGTAACCATGACGTCATGGAAATGGGAG 3'
a Underlining indicates mutated bases.
Electrophoretic mobility shift assays (EMSAs) were performedwith 2 to 4 ,ug of protein from nuclear extracts or 1 ,ug ofprotein from extracts of bacteria expressing recombinant hu-man GATA-3, 3 jig of poly(dI-dC), 10 mM Tris, 80 mM NaCl,10% glycerol, and 104 cpm (1 ng) of probe. Binding reactionmixtures were incubated at room temperature for 20 min,loaded directly onto a 6% polyacrylamide gel, and electropho-resed at 120 V in 0.5x Tris-borate-EDTA. For o-phena-throline-copper (Op-Cu) nuclease footprinting, the reactionmixture described was increased 10-fold, and following elec-trophoresis, gels were treated with chemical endonucleaseOp-Cu as previously described (23). The gel was visualized byaudioradiography, and the free and bound complexes were cutout and eluted from the gel (31). The DNA was then separatedon a 6% sequencing gel with G and G+A ladders andvisualized by autoradiography.
Probes for EMSA and footprinting were generated from themurine TCRI enhancer. Probes were generated by 5' endlabeling with T4 polynucleotide kinase (31), end filling with theKlenow fragment of Escherichia coli polymerase (28), or PCRusing a kinase-treated oligonucleotide (22). All probes weregel purified.
Site-directed mutagenesis. The TCRf3 enhancer was sub-cloned into pUC19, and site-specific mutations were generatedby PCR using previously described protocols (24, 40). Briefly,the procedure requires a single mutant primer and two flank-ing primers, which in this case annealed within the polylinkerof pUC19. The oligonucleotides used in these experiments areshown in Table 1. Initially, the mutagenic primer and thedownstream flanking primer (pUC19 primer 1) are used in aPCR to generate a primary product. Conditions for this initialPCR were as follows: 20 fmol of wild-type template, 1 jiM eachprimer, 200 jiM deoxynucleoside triphosphates, 10 mM Tris,500 mM KCl, 1.5 mM MgCl2, and 0.01% gelatin. PCRs wereperformed for 30 cycles at 94°C for 1 min, 45°C for 1 min, and72°C for 2 min. The PCR product from this first reaction wasgel purified and used in a second PCR as a primer with pUC19primer 2 and the wild-type enhancer as the template. Thissecond PCR product was then gel purified and subcloned intothe appropriate vectors. All mutations were confirmed byDNA sequencing.
Transfections. For transfection experiments, the TCRI en-hancer and the mutant enhancers were subcloned intopAlOLUC. This vector was constructed by replacing the chlor-amphenicol acetyltransferase gene in pAlOCAT (11) with aluciferase reporter gene from p19LUC (54). The enhancerconstructs were cloned downstream of the luciferase reportergene. Oligonucleotides representing TE4 were multimerizedand inserted 5' of the simian virus 40 promoter of pAlOLUC
or 5' of the herpes simplex virus thymidine kinase promoter ofpBLCAT2 (27).EL-4 cells were resuspended in RPMI supplemented with
5% fetal calf serum and 5 x 10-5 M mercaptoethanol at 1 x107 to 3 x 107 cells per ml, and 0.3 ml of the cell suspensionwas electroporated by pulsing (960 ,uF, 220 V) from a Bio-RadGene Pulser apparatus (Bio-Rad, Richmond, Calif.). Cellswere then resuspended in 5 ml of medium and incubated at37°C in 5% CO2 in air for 10 to 16 h before harvesting forluciferase assays. L cells and P3XAg8.63 cells were electropo-rated at 960 ,uF and 250 V and at 960 jiF and 240 V,respectively. These cells were resuspended in Iscove modifiedDulbecco medium supplemented with 10% fetal calf serumand cultured for 16 h.For luciferase quantitation, cells were washed twice with
phosphate-buffered saline and lysed with 0.5 ml of lysis buffer(1% Triton X-100, 25 mM glycylglycine [pH 7.8], 15 mMMgSO4, 4 mM EGTA, 1 mM dithiothreitol [DTT']). Lysateswere cleared by centrifugation. To assay for luciferase activity,500 RI of reaction buffer (15 mM KPO4 buffer [pH 7.8], 2 mMATP [pH 7.0], 25 mM glycylglycine, 15 mM MgSO4, 4 mMEGTA, 2 mM DTT) and 100 RI of substrate buffer (0.4 mMluciferin, 25 mM glycylglycine, 15 mM MgSO4, 4 mM EGTA,2 mM DTT) were added to 100 RI of the supernatant. Sampleswere measured by a luminometer.
All transfections were done in at least duplicate, and trans-fection efficiencies were controlled by cotransfecting a humangrowth hormone expression vector, pXGH5 (Nichols InstituteDiagnostics, San Juan Capistrano, Calif.).
RESULTS
Three protein-binding sites within the TCR, enhancercontain a GATA consensus sequence and bind recombinantGATA-3 efliciently. Sequence analysis revealed several poten-tial GATA-binding sites within the TCR,B enhancer. To deter-mine which GATA sequences were recognized by nuclearproteins, EMSAs were performed with probes spanning the800-bp PvuII-NcoI fragment previously shown to contain en-hancer activity (32). Nuclear extracts obtained from T cells, Bcells, and fibroblasts were compared. Complexes TE1, TE2,TE4, and TE5 were unique to T-cell extracts (Fig. 1), althoughcomplexes with different mobilities bound to the same probeswith non-T-cell extracts. Two additional complexes, TE3 andTE6, were observed in all extracts (Fig. 1 and data not shown).When the six most prominent complexes were mapped bychemical nuclease footprinting, the consensus sequence for theGATA family of DNA-binding proteins, WGATAR, was ob-served in the sequences protected by the T-cell-specific com-plexes TE1, TE2, TE4, and TE5 (Fig. 1B).The possibility that sites TE1, TE2, TE4, and TE5 represent
in vivo GATA-3-binding sites was further investigated. First,the ability of recombinant GATA-3 to bind the sites was testedby EMSA. As shown in Fig. 2A, probes that included sites TEl,TE2, TE4, and TE5 gave specific gel shift complexes in thepresence of recombinant GATA-3. The lower-mobility com-plexes observed with the TE2 and TE4 probes probablyrepresent degraded GATA-3 present in the crude bacterialextracts. In addition, oligonucleotides representing each sitewere used as unlabeled competitors in an EMSA in whichrecombinant GATA-3 was bound to Ta3, an establishedGATA-3-binding site from the TCRo enhancer (15). TE1,TE2, and TE4 oligonucleotides competed very strongly forGATA-3 binding to Ta3 (Fig. 2B), indicating that these sitesbind recombinant GATA-3 with a high affinity. TE5 appears tobind recombinant GATA-3 with a lower affinity, since it was a
VOL. 14, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 20
Feb
ruar
y 20
22 b
y 21
1.10
8.93
.162
.

4288 HENDERSON ET AL.
A.
N N N
__j m _; LU Ui C.U) U)
r- r-N Nr
mIrs m X Kr 0re LLJ Cr rn CL L en c- V
.I' m
)nq < C -~~- r.m - Ln Um uw M. L/) =
_jjww-
I.~~~~~~~~~~~40
.-_:SI_'-
=bd.ihv-A _1S
_ -
B.
TE1TATATAC .osT T 1A-
TE4
TE4: TPT2A
E 1 1[3E2}-'- - r AA_CTr-rrT -A'-TAAAT 'AICA
TE2
MURINE TE4 :-G'T-TTATCTGTAAC:AACATCA
HUMAN TP2FIG. 1. Protein-binding sites within the TCR,B enhancer. (A) EMSAs using nuclear extracts prepared from NIH 3T3 cells, the plasmacytoma
cell lines P3XAg8.63 and S107, and the T-cell lines EL-4, RS4.2, and SL12.2. End-labeled probes generated from the PvuII-NcoI TCR, enhancerfragment were as follows: the TE1 probe is a 129-bp AluI-Hinfl fragment, the TE2 probe is a 139-bp Sau3A-EarI fragment, the TE4 probe is a
89-bp AluI-MboII fragment, and the TE5 probe is a 136-bp AluI-NcoI fragment. T-cell-specific complexes are indicated by solid arrows; the TE6complex is indicated by the open arrow. (B) Summary of binding sites within the TCR,3 enhancer as determined by Op-Cu footprinting.T-cell-specific complexes are represented as hatched boxes; ubiquitous complexes are shown as stippled boxes. Potential GATA sequences are
underlined. Binding sites described in previous studies (12, 49) that overlap the footprints are shown in parentheses. (C) Comparison of sequencesfrom the footprinted sites of the murine TCR,B enhancer and known human TCR,B enhancer sequences (12, 43). Differences in the murinesequence are shown below the human sequence.
weak competitor. Finally, oligonucleotides containing a
GATA-3 consensus sequence (Table 1) were used as unlabeledcompetitors to determine if the TE1, TE2, and TE4 complexesformed from T-cell nuclear extracts contained GATA-bindingproteins. GATA oligonucleotides competed very efficiently forall three of the complexes (Fig. 2C). Thus, GATA-bindingprotein is a component of the TE1, TE2, and TE4 EL-4complexes. The T-cell specificity of these complexes is consis-tent with the possibility that the protein binding the sites isGATA-3 (Fig. 1A). Taken together, the in vitro binding studiesshow that three sites, TE1, TE2, and TE4, are recognized byT-cell-specific proteins in nuclear extracts, contain GATAsequences, and bind recombinant GATA-3 efficiently. TheT-cell specificity and ability of GATA oligonucleotides to
compete for complexes TE1, TE2, and TE4 suggest thatGATA-3 in T-cell nuclear extracts binds these sites.The TE4 GATA sequence is required for TCROi enhancer
activity in T cells; sites TE1 and TE2 are not functionallyequivalent to site TE4. It is possible that the T-cell-specificactivity of the TCR,B enhancer in vivo is mediated in part byGATA-3 binding at sites TE1, TE2, and TE4. Therefore, wedesigned experiments to determine if these three sites wereimportant for TCRPi enhancer function and if they were
functionally equivalent. Site-directed mutagenesis was per-formed on each site, using PCR (24, 40) (see Materials andMethods); the oligonucleotides used to generate the variousmutations are shown in Table 1. The ability of each of themutant sites to bind protein is shown in Fig. 3. Neither
T F I TE2 T E
TE5 TE5
TE3(:ES)
TE6
lOObpII
1. 01!V/.o
.-ATA,TAT-C CA :-"r,,ir-e7-7.-A!_-,,"%t'71-_"'L-.:',--- --.. -0-
I.
MOL. CELL. BIOL.
-111.-00,
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 20
Feb
ruar
y 20
22 b
y 21
1.10
8.93
.162
.

GATA ELEMENTS IN THE TCRO ENHANCER 4289
A.COMPETITOR - +
APROBE 5
elf LU
H 2_
.. ~~~~~~~~~A
go.. W
TiE,TE2E4 TF
rT T E2t FF~ 4 N*-
Sb
,_ -;IPF7-. t; .W-'T!', TCX;
.1 x_ -!A7-_L CZt'_AL.@ <-s.fJ_
A AT A 1 TCA_-C_ t. Z.Zz. C, ._ .,.P
B _PRCEE
_ s
CC ;VGA!,'3V_.;>'A8ATC2ATA T, 7T:T,,r
A f Ae ;T A' aCT_CA--J6> ? ' 1~~~~Tq m{;-'v ' r{-' ;1.A G A A G1T
++
GAT C ATCrTC_ ACAaT- T
GP;TAv zr Tv
C.-
aoXx8 x
~ ~ ~ X8S0x 0--
Li;..:-. :F
_j: .:
-
TEI TE2 TE4
FIG. 2. GATA-containing sites within the TCRO enhancer bindGATA-3. (A) EMSAs using recombinant human GATA-3 and end-labeled probes generated from the the TCRI enhancer which spannedsite TEl, TE2, TE4, or TE5 without (-) or with (+) 100 ng of specificunlabeled competitor. (B) Summary of experiments in which oligonu-cleotides representing sites TE1, TE2, TE4, and TE5 were used asunlabeled competitors for human GATA-3 binding to Ta3, a GATA-3-binding site from the the TCRot enhancer. (C) EMSAs with EL-4nuclear extracts, using probes spanning sites TE1, TE2, and TE4 in thepresence of increasing concentrations of unlabeled oligonucleotidescontaining a GATA-binding site.
recombinant GATA-3 (Fig. 3A) nor proteins present in EL-4nuclear extracts (Fig. 3B) bound to probes containing themutated TE1, TE2, or TE4 site.These mutations were tested for enhancer activity within the
context of the 800-bp TCR,B enhancer by cloning wild-type andmutant PvuI-NcoI TCR, enhancers into a site 3' of theluciferase gene under the control of a simian virus 40 earlypromoter lacking an enhancer. Transfection into EL-4 T cellsshowed that the TE1 and the TE2 mutations had only a
N1FIG. 3. TE1, TE2, and TE4 GATA mutations abolish binding of
recombinant GATA-3 and the T-cell-specific complex. (A) EMSAsusing wild-type and mutant TE1, TE2, and TE4 probes with recombi-nant human GATA-3. (B) EMSAs with EL-4 nuclear extracts andwild-type and mutant probes.
minimal effect on enhancer activity, decreasing activity of theelement by 25% (Fig. 4). Enhancer activity was not decreasedfurther by a TE1 TE2 double mutation. However, the TE4mutation had a significant effect on the TCRf enhancerfunction, resulting in an 85% decrease in the activity of theenhancer. Combined mutations were also tested in the samevector to determine whether sites TE1 or TE2 have subtlefunctions which would be revealed in the absence of another
100-
90-
c80-
t70-
60-cu
50-
co40i.030-
o20
.-e 10-
0
LU 0 CMJLU 1: C
~~~LU LU -
FIG. 4. Site TE4 is required for full TCR,B enhancer activity in Tcells. The 800-bp PvuII-NcoI fragments of the wild-type (TCE) andmutant TCRI enhancers were cloned 3' of a luciferase reporter genein pAlOLUC. EL-4 cells were transfected by electroporation with 10pLg of the various constructs. Each datum point represents at leastthree independent experiments.
B.
+
H-
VOL. 14, 1994
x xo 8
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 20
Feb
ruar
y 20
22 b
y 21
1.10
8.93
.162
.
4290 HENDERSON ET AL.
site(s). TCRP enhancers with combinations of mutated TE1and TE4 sites or mutated TEl, TE2, and TE4 sites showedactivity comparable to that of the enhancer with mutations inTE4 alone (Fig. 4). Therefore, these data show that althoughseveral TCR3 sites can bind GATA-3, they are not functionallyequivalent. The TE4 GATA sequence is required for fullactivity of the TCRI enhancer, whereas the TE1 and TE2 sitesdo not appear to be necessary, nor can they compensatefunctionally for the TE4 site.
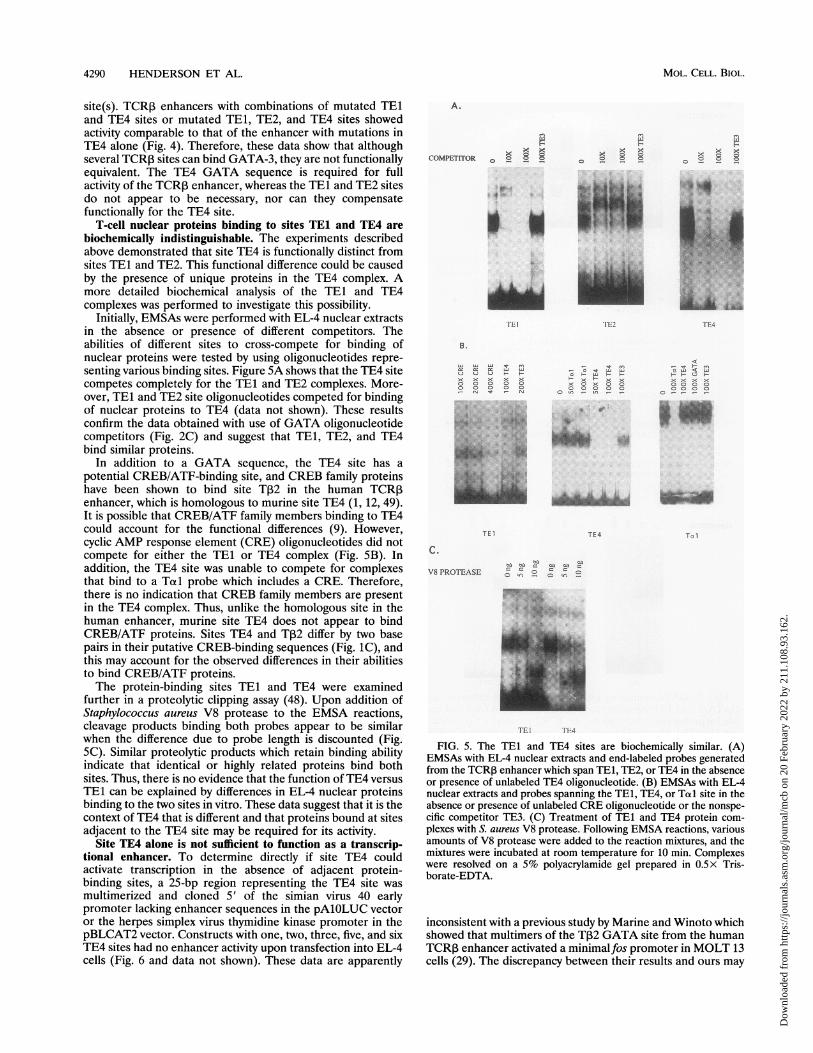
T-cell nuclear proteins binding to sites TE1 and TE4 arebiochemically indistinguishable. The experiments describedabove demonstrated that site TE4 is functionally distinct fromsites TE1 and TE2. This functional difference could be causedby the presence of unique proteins in the TE4 complex. Amore detailed biochemical analysis of the TE1 and TE4complexes was performed to investigate this possibility.
Initially, EMSAs were performed with EL-4 nuclear extractsin the absence or presence of different competitors. Theabilities of different sites to cross-compete for binding ofnuclear proteins were tested by using oligonucleotides repre-senting various binding sites. Figure 5A shows that the TE4 sitecompetes completely for the TE1 and TE2 complexes. More-over, TE1 and TE2 site oligonucleotides competed for bindingof nuclear proteins to TE4 (data not shown). These resultsconfirm the data obtained with use of GATA oligonucleotidecompetitors (Fig. 2C) and suggest that TEl, TE2, and TE4bind similar proteins.
In addition to a GATA sequence, the TE4 site has apotential CREB/ATF-binding site, and CREB family proteinshave been shown to bind site T,2 in the human TCRPenhancer, which is homologous to murine site TE4 (1, 12, 49).It is possible that CREB/ATF family members binding to TE4could account for the functional differences (9). However,cyclic AMP response element (CRE) oligonucleotides did notcompete for either the TEl or TE4 complex (Fig. 5B). Inaddition, the TE4 site was unable to compete for complexesthat bind to a Totl probe which includes a CRE. Therefore,there is no indication that CREB family members are presentin the TE4 complex. Thus, unlike the homologous site in thehuman enhancer, murine site TE4 does not appear to bindCREB/ATF proteins. Sites TE4 and T32 differ by two basepairs in their putative CREB-binding sequences (Fig. 1C), andthis may account for the observed differences in their abilitiesto bind CREB/ATF proteins.The protein-binding sites TE1 and TE4 were examined
further in a proteolytic clipping assay (48). Upon addition ofStaphylococcus aureus V8 protease to the EMSA reactions,cleavage products binding both probes appear to be similarwhen the difference due to probe length is discounted (Fig.5C). Similar proteolytic products which retain binding abilityindicate that identical or highly related proteins bind bothsites. Thus, there is no evidence that the function of TE4 versusTE1 can be explained by differences in EL-4 nuclear proteinsbinding to the two sites in vitro. These data suggest that it is thecontext of TE4 that is different and that proteins bound at sitesadjacent to the TE4 site may be required for its activity.
Site TE4 alone is not sufficient to function as a transcrip-tional enhancer. To determine directly if site TE4 couldactivate transcription in the absence of adjacent protein-binding sites, a 25-bp region representing the TE4 site wasmultimerized and cloned 5' of the simian virus 40 earlypromoter lacking enhancer sequences in the pAlOLUC vectoror the herpes simplex virus thymidine kinase promoter in thepBLCAT2 vector. Constructs with one, two, three, five, and sixTE4 sites had no enhancer activity upon transfection into EL-4cells (Fig. 6 and data not shown). These data are apparently
A.
COMPETITOR o ° 8 8
IM-...A:
T.l
TEI rE2
B.
LU U LU It rn Vcc er: 0: ui 42 _ L Lt XtJ U U F- L Fx X x x X x x x
0 0 0 0 x C, x 0 00 0 0 0 0 0 0 0o o_Ner _ rq O e _u~~~~C)n _ _
TEI
C.
V8 PROTEASE
TE4
0 8 8
TEl
st - rf}
x x x x
-
Tie1
5004 04O 0__ -
iEl LiT[4
FIG. 5. The TEI and TE4 sites are biochemically similar. (A)EMSAs with EL-4 nuclear extracts and end-labeled probes generatedfrom the TCRP enhancer which span TE1, TE2, or TE4 in the absenceor presence of unlabeled TE4 oligonucleotide. (B) EMSAs with EL-4nuclear extracts and probes spanning the TE1, TE4, or Told site in theabsence or presence of unlabeled CRE oligonucleotide or the nonspe-cific competitor TE3. (C) Treatment of TE1 and TE4 protein com-plexes with S. aureus V8 protease. Following EMSA reactions, variousamounts of V8 protease were added to the reaction mixtures, and themixtures were incubated at room temperature for 10 min. Complexeswere resolved on a 5% polyacrylamide gel prepared in 0.5x Tris-borate-EDTA.
inconsistent with a previous study by Marine and Winoto whichshowed that multimers of the T,B2 GATA site from the humanTCR,B enhancer activated a minimalfos promoter in MOLT 13cells (29). The discrepancy between their results and ours may
i.,,i
MOL. CELL. BIOL.
m
x X x
0 R 8 8
gAAAA"
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 20
Feb
ruar
y 20
22 b
y 21
1.10
8.93
.162
.

GATA ELEMENTS IN THE TCR,B ENHANCER 4291
100-
75-2:><1' 50-
25-
0-
A.
15
>
a1)cr-
TCE pAl OLUC (TE4)6
FIG. 6. The TE4 site alone does not have enhancer activity. EL-4cells were transfected by electroporation with 10 ,ug of pAlOLUC,pAlOLUC with the TCRI enhancer (TCE), or pAlOLUC with a TE4hexamer cloned 5' of the simian virus 40 promoter [(TE4)6]. Thesedata represent three independent experiments.
20X5-
4 1
2-
10
0 ~~~
0) D-J
0-
N 't (11 LU LU D
N
U)CL
B.
be due to differences in the cells used, differences in thepromoter contexts of the multimers, or sequence differencesbetween murine site TE4 and human site T12, possibly relatedto differences in their ability to bind CREB/ATF proteins. Ourdata show that although the GATA sequence within TE4 isnecessary for enhancer function, the TE4 site alone is notsufficient to activate transcription of an enhancer-dependentpromoter. These data further support the notion that tran-scription factors bound at other sites, possibly adjacent to TE4,are necessary for TE4-dependent activation.
Site TE1 functions as a negative regulator ofTCRI functionin fibroblasts but not in B cells. The PvuI-NcoI enhancerconstruct used in the experiments described above has beenpreviously shown to be T-cell specific (32). Since tissue speci-ficity of many enhancers, including T-cell-specific enhancers, isachieved by a combination of negative and positive elements(43, 57, 58), we wished to determine if site TE1, TE2, or TE4was important for keeping the enhancer activity low in fibro-blasts or B cells. Therefore, the constructs containing singlemutations were transfected into P3XAg8.63, a plasmacytomaB-cell line, and L cells, a fibroblast line. Consistent withprevious data from this laboratory, the wild-type TCR,B en-hancer was not active in the B-cell line (Fig. 7A). In addition,none of the mutants showed significantly increased activity(Fig. 7A). Negligible enhancer activity was also observed whenL-cell fibroblasts were transfected with the wild-type enhanceror with the mutant TE2 and mutant TE4 enhancer constructs(Fig. 7B). However, the TEI mutant enhancer showed mod-estly increased activity (in excess of threefold) in fibroblastsrelative to wild-type or to the other mutant enhancers (Fig.7B). These results strongly suggest that a negative regulatorpresent in fibroblasts binds to the TEI site. Since mutation ofthe GATA consensus within site TEl relieves repression, itseems likely that the negative regulator may be a member ofthe GATA family. The data further show that sites TE2 andTE4 cannot functionally replace site TEI for mediating therepressor activity. Finally, the data suggest that regulation ofthe TCRP3 enhancer is different in B cells and fibroblastsbecause the TEI mutation does not activate the enhancer in Bcells.
DISCUSSION
This study examined the functional significance of GATAelements in determining the T-cell specificity the TCR3 en-hancer; three important findings have emerged. First, only oneof three high-affinity GATA-3-binding sites is critical foractivation of the enhancer in T cells. Second, another GATA
LU 0 - N v QC5 LU LU LU
~~~~~~~~~~~~~~C\jFIG. 7. Mutations within the TCR,B enhancer do not affect the
activity of the enhancer in B cells, but the TEI mutation increases theactivity of the enhancer in fibroblasts. P3XAg8.63 cells (A) or L-cellfibroblasts (B) were transfected by electroporation with 10 jig of thevarious TCR,B enhancer constructs. Ten micrograms of pSV2LUC wasused as a positive control for transfections. Each datum point repre-sents at least three independent experiments.
site is important for repression of enhancer activity in fibro-blasts but not in B cells, demonstrating that different mecha-nisms regulate the enhancer in fibroblasts and B cells. Finally,for both activation and repression of the TCRf enhancer,GATA-binding proteins are context dependent, emphasizingthe importance of protein-protein interactions for the mecha-nism of action of GATA-binding proteins.TCR,1 activity in T cells. Identification of site TE4 as an
important functional element of the TCR3 enhancer in T cellsis consistent with previous work which defined a 350-bp core asthe minimal TCRI enhancer (12, 43, 49). TE4 is the only siteidentified in this work that is within the minimal enhancer, andit corresponds to previously described sites Pe 1 and 3e2 in themurine enhancer and T,B2 in the human enhancer (12, 49).Analysis of the TE4 GATA mutation confirms and extendsprevious deletional and mutational analyses in this region (12,21, 29, 42, 49). We examined the TE4 mutation within thecontext of an 800-bp enhancer fragment to determine whetherother sites could compensate for TE4 function. This wasparticularly important since the TCR,B enhancer includes otherGATA sequences, TEl and TE2, which bind recombinantGATA-3 with a high affinity and which are recognized byproteins from T-cell nuclear extracts. Our data show that TE4is functionally distinct and that sites TEI and TE2 are notequivalent to TE4, since they were unable to compensate forthe TE4 mutation. Although mutation of the GATA consensus
VOL. 14, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 20
Feb
ruar
y 20
22 b
y 21
1.10
8.93
.162
.

4292 HENDERSON ET AL.
within TE1 and TE2 had a negligible effect on the activity ofthe TCR,B enhancer, it is possible that these sites are importantduring T-cell development at a stage not represented by EL-4cells. In addition, these studies do not address the functionalsignificance of nonconsensus or low-affinity GATA sites withinthe TCR,B enhancer (19, 33).Our results further demonstrate that mutation of the TE4
GATA consensus sequence and loss of ability to bind recom-
binant GATA-3 correlate with loss of TE4 function, suggestingthat GATA-3 is necessary for TE4-dependent activation invivo. Four facts support this hypothesis: (i) GATA-3 is the onlyknown GATA-binding protein present in T cells (15, 18, 37,39), (ii) TE4 binds recombinant GATA-3 strongly, (iii) GATAsite oligonucleotides compete completely for the TE4 complex,and (iv) the TE4 complex is T cell specific, consistent with theT-cell-specific expression of GATA-3.TEl and TE2 also bind recombinant GATA-3 and a tissue-
specific protein complex from nuclear extracts. EMSAs withdifferent competitor oligonucleotides suggest that these sitesare also binding GATA-3. Furthermore, proteins binding toTE1 and TE4 appear to be similar, as judged from partialproteolysis. Thus, it seems likely that in vivo, GATA-3 bindssite TE1 and TE2 as well as site TE4. However, TEl and TE2do not function like TE4, strongly suggesting that a differentcontext for the sites is critical in determining their functions.According to the context model, GATA-3 bound at TE4interacts with proteins bound to nearby sites and those inter-actions permit transcriptional activation, whereas GATA-3bound at TE1 or TE2 is surrounded by different proteins thatdo not allow transcriptional activation. This model is sup-
ported by our demonstration that multimers of the TE4 siteare not able to activate transcription alone. The ability ofneighboring proteins to influence the function of the lymphoidcell-specific factor LEF-1 has been recently demonstrated (4,10).There are potentially a number of proteins that could be
required for GATA-3-dependent activation at the TE4 site. Anobvious candidate was the CREB family of activators since theTE4 footprinted region includes sequences that match theconsensus for a CRE which serves as a binding site forATF/CREB family proteins (13, 16, 36). CREs are foundwithin the V,B promoter and the enhancers for TCRot, CD38and CD2 (1, 9, 12, 25, 49). However, oligonucleotide compe-
titions and proteolytic clipping experiments show no evidenceof CREB proteins binding to TE4. This is in contrast to thehomologous sequence in the human TCRI enhancer, T,B2,which binds CREB/ATF (12). The role, if any, of CREBproteins at the TE4 site is unclear. Other protein-binding sitesthat may influence TE4 include an Ets site and a CBF sitewithin the core enhancer, a C/EBP site, a TFE-3 site, andbinding sites for the high-mobitity group family members andPOU domain proteins (14, 32, 34, 35, 38, 42, 45, 49, 50, 52, 53,56). Additional work will be necessary to test the context
model and to identify proteins which are required for GATA-3function.
Negative regulation of the TCR, enhancer in non-T cells.The TCR,B enhancer is T-cell specific, having virtually unde-tectable activity in lineages such as fibroblasts and B cells (32,49). Although some of the T-cell specificity can be explained bythe chromatin structure of the TCR3 enhancer in T cells andthe presence in T cells of T-cell-specific activators such as
GATA-3, it seems likely that negative regulators may also berequired to keep the enhancer silent in non-T cells. Silencingelements have been identified in the TCRot and -,y enhancers,and a role for negative elements in the TCRP enhancer hasbeen suggested in previous studies (17, 57, 58).
When GATA site mutations were tested for activity in Bcells and fibroblasts, the data showed that mutation of the TE1site resulted in an increase in enhancer activity in fibroblastsbut not in B cells, thus directly demonstrating negative regu-lation of the TCR,B enhancer. Mutation of sites TE2 and TE4had no effect on enhancer activity in either cell type. Theresults suggest that a negative regulator binds the TE1 site infibroblasts, although the identity of this protein is not currentlyknown. EMSAs and chemical nuclease footprinting show aprotein present in crude nuclear extracts from fibroblastswhich binds TEl and TE4, generating complexes different inmobility and footprint patterns from the T-cell complexes (Fig.1A and data not shown). Since these complexes are alsocompeted for with GATA oligonucleotides and abolished bymutation of the GATA consensus sequence (data not shown),the negative regulator may be a GATA family member.Although GATA family members have been demonstrated toact as positive factors, a recent study demonstrated that ectopicexpression of GATA-2 could inhibit erthryoid differentiation,indicating that in certain situations, GATA proteins may act asnegative regulators (3). It is also possible that the GATAelement overlaps a non-GATA-binding repressor element,similar to that proposed for the glycophorin B promoter (44).These data also reveal another functional difference betweenGATA sites, since TE4 cannot replace TE1 for negativeregulation, even though TE1 and TE4 appear to bind indistin-guishable proteins.Our data also suggest that the mechanisms for negative
regulation of the TCR,B enhancer are different in B cells andfibroblasts, since mutation of the TEl site increased enhanceractivity in fibroblasts but not in B cells. It is interesting tospeculate that this difference may be related to the need tocontrol rearrangement of T-cell receptor and immunoglobulingenes. There is evidence that enhancer-dependent transcrip-tion in the immunoglobulin and T-cell receptor loci may targetthe V(D)J recombination machinery to the correct genes in Bcells and T cells, respectively (59). Thus, it may be that specialmechanisms are necessary in B cells to silence TCR3 transcrip-tion to ensure that a-chain genes are not rearranged in B cells.
ACKNOWLEDGMENTS
We thank Gerald Siu and Hamish Young for critically reading themanuscript.
A.J.H. is supported by a postdoctoral fellowship from the AaronDiamond Foundation. S.M. was a Special Fellow of the LeukemiaSociety of America and is in part supported by VA medical researchfunds. This work was also supported by NIH grants GM29361 andGM44300 to K.C.
REFERENCES1. Anderson, S. J., S. Miyake, and D. Y. Loh. 1989. Transcription
from a murine T-cell receptor VP promoter depends on aconserved decamer motif similar to cyclic AMP response element.Mol. Cell. Biol. 9:4835-4845.
2. Blackwell, T. K., M. W. Moore, G. Yancopoulos, H. Suh, S.Lutzker, E. Selsing, and F. W. Alt. 1986. Recombination betweenimmunoglobulin variable region gene segments is enhanced bytranscription. Nature (London) 324:585-589.
3. Briegal, K., K.-C. Lin, C. Plank, H. Beug, J. D. Engel, and M.Zenke. 1993. Ectopic expression of a conditional GATA-2/estro-gen receptor chimera arrests erythroid differentiation in a hor-mone-dependent manner. Genes Dev. 7:1097-1109.
4. Carlsson, P., M. L. Waterman, and K. A. Jones. 1993. ThehLEF-1/TCF-la HMG protein contains a context-dependent tran-scriptional activation domain that induces the TCRa enhancer inT cells. Genes Dev. 7:2418-2430.
MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 20
Feb
ruar
y 20
22 b
y 21
1.10
8.93
.162
.

GATA ELEMENTS IN THE TCRI ENHANCER 4293
5. Davis, M. M., and P. J. Bjorkman. 1988. T-cell antigen receptorgenes and T-cell recognition. Nature (London) 334:395-402.
6. Dignam, J., R. Lebovitz, and R. Roeder. 1983. Accurate transcrip-tion initiation by RNA polymerase II in a soluble extract fromisolated mammalian nuclei. Nucleic Acids Res. 11:1475-89.
7. Evans, T., and G. Felsenfeld. 1989. The erythroid-specific tran-scription factor Eryfl: a new zinc finger protein. Cell 58:877-885.
8. Evans, T., M. Reitman, and G. Felsenfeld. 1988. An erythrocyte-specific DNA-binding factor recognizes a regulatory sequencecommon to all chicken globin genes. Proc. Natl. Acad. Sci. USA85:5976-5980.
9. Georgopoulos, K., B. A. Morgan, and D. D. Moore. 1992. Func-tionally distinct isoforms of the CRE-BP DNA-binding proteinmediate activity of a T-cell specific enhancer. Mol. Cell. Biol.12:747-757.
10. Giese, K., and R. Grosschedl. 1993. LEF-1 contains an activationdomain that stimulates transcription only in a specific context offactor-binding sites. EMBO J. 12:4667-4676.
11. Gorman, C. M., G. T. Merlino, M. C. Willingham, I. Pastan, andB. H. Howard. 1982. The Rous sarcoma virus long terminal repeatis a strong promoter when introduced into a variety of eukaryoticcells by DNA-mediated transfection. Proc. Natl. Acad. Sci. USA79:6777-6781.
12. Gottschalk, L. R., and J. M. Leiden. 1990. Identification andfunctional characterization of the human T cell receptor ,B genetranscriptional enhancer: common nuclear proteins interact withthe transcriptional regulatory elements of the T cell receptor cx andP3 genes. Mol. Cell. Biol. 10:5486-5495.
13. Hai, T. W., F. Liu, W. I. Coukos, and M. R. Green. 1989.Transcription factor ATF cDNA clones: an extensive family ofleucine zipper proteins able to selectively form DNA-bindingheterodimers. Genes Dev. 3:2083-2090.
14. Ho, I.-C., N. K. Bhat, L. R. Gottschalk, T. Lindstin, C. B.Thompson, T. S. Papas, and J. M. Leiden. 1990. Sequence-specificbinding of human Ets-1 to the T cell receptor a gene enhancer.Science 250:814-818.
15. Ho, I.-C., P. Voorhees, N. Marin, B. K. Oakley, S.-F. Tsai, S. H.Orkin, and J. M. Leiden. 1991. Human GATA-3: a lineagerestricted transcription factor that regulates the expression of theT cell receptor alpha gene. EMBO J. 10:1187-1192.
16. Hoeffler, J. P., T. E. Meyer, Y. Yan, J. L. Jameson, and J. F.Habener. 1988. Cyclic AMP-response DNA-binding protein:structure based on a cloned placental cDNA. Science 242:1430-1433.
17. Ishida, I., S. Verbeek, M. Bonneville, S. Itohara, A. Berns, and S.Tonegawa. 1990. T-cell receptor gamma delta and gamma trans-genic mice suggest a role of a gamma gene silencer in thegeneration of alpha beta T cells. Proc. Natl. Acad. Sci. USA87:3067-3071.
18. Joulin, V., D. Bories, J.-F. Eleouet, M.-C. Labastie, S. Chretien,M.-G. Mattei, and P. H. Romeo. 1991. A T cell specific TCR 8DNA binding protein is a member of the human GATA family.EMBO J. 10:1809-1816.
19. Ko, L. J., and J. D. Engel. 1993. DNA-binding specificities of theGATA transcription factor family. Mol. Cell. Biol. 13:4011-4022.
20. Ko, L. J., M. Yamamoto, M. W. Leonard, K. M. George, P. Ting,and J. D. Engel. 1991. Murine and human T-lymphocyte GATA-3factors mediate transcription though a cis-regulatory elementwithin the human T-cell receptor 8 gene enhancer. Mol. Cell. Biol.11:2778-2784.
21. Krimmpenfort, P., R. de Jong, Y. Uematsa, Z. Dembic, S. Ryser,H. von Boehmer, M. Steinmetz, and A. Berns. 1988. Transcriptionof T-cell receptor ,B-chain gene is controlled by downstreamregulatory element. EMBO J. 7:745-750.
22. Krummel, B. 1990. DNase 1 footprinting, p. 184-188. In M. Innis,D. Gelfand, J. Sninsky, and T. White (ed.), PCR protocols: a guideto methods and applications. Academic Press, Inc., San Diego,Calif.
23. Kuwabara, M., and D. Sigman. 1987. Footprinting DNA-proteincomplexes in situ following gel retardation assays using 1,10-phenanthroline-copper ion: Escherichia coli RNA polymerase-lacpromoter complexes. Biochemistry 26:7234-7238.
24. Landt, O., H.-P. Grunert, and U. Hahn. 1990. A general method
for rapid site-directed mutagenesis using the polymerase chainreaction. Gene 96:125-128.
25. Leiden, J. M. 1992. The T cell receptor a gene: a molecular modelof transcriptional regulation during T cell development. Immunol.Today 13:22-30.
26. Leiden, J. M. 1993. Regulation of T cell receptor genes. Annu.Rev. Immunol. 11:539-570.
27. Luckow, B., and G. Schutz. 1987. CAT constructs with multipleunique restriction sites for the functional analysis of eukaryoticpromoters and regulatory elements. Nucleic Acids Res. 15:5490.
28. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, N.Y.
29. Marine, J., and A. Winoto. 1991. The human enhancer-bindingprotein Gata-3 binds several T-cell receptor regulatory elements.Proc. Natl. Acad. Sci. USA 88:7284-7288.
30. Martin, D. I., L. I. Zon, G. Mutter, and S. H. Orkin. 1990.Expression of an erythroid transcription factor in megakaryocyticand mast cell lineages. Nature (London) 314:444-447.
31. Maxam, A., and W. Gilbert. 1980. Sequencing end-labeled DNAwith base specific chemical cleavages. Methods Enzymol. 65:499-560.
32. McDougall, S., C. L. Peterson, and K. Calame. 1988. A transcrip-tional enhancer 3' of C,B2 in the T-cell receptor 3 locus. Science241:205-208.
33. Merika, M., and S. H. Orkin. 1993. DNA-binding specificity ofGATA family transcription factors. Mol. Cell. Biol. 13:3999-4010.
34. Messier, H., H. Brickner, J. Gaikwad, and A. Fotedar. 1993. Anovel POU domain protein which binds to the T-cell beta en-hancer. Mol. Cell. Biol. 13:5450-5460.
35. Messier, H., T. Fuller, S. Mangal, H. Brickner, S. Igarashi, J.Gaiwad, R. Fotedar, and A. Fotedar. 1993. p70 lupus autoantigenbinds the enhancer of the T-cell receptor beta-chain gene. Proc.Nati. Acad. Sci. USA 90:2685-2689.
36. Montminy, M. R., K. A. Sevarino, J. A. Wagner, G. Mandel, andR. H. Goodman. 1986. Identification of a cyclic AMP-responsiveelement within the rat somatostatin gene. Proc. Natl. Acad. Sci.USA 83:6682-6686.
37. Oosterwegel, M., J. Timmerman, J. M. Leiden, A. Kruisbeek, andH. Clevers. 1992. Expression of GATA-3 during lymphocytedifferentiation and mouse embryogenesis. Dev. Immunol. 3:1-11.
38. Oosterwegel, M., M. van de Wetering, D. Dooijes, L. Klomp, A.Winoto, K. Georgopoulos, F. Meijlink, and H. Clevers. 1991.Cloning of murine TCF-1, a T cell specific transcription factorinteracting with functional motifs in the CD3-c and T cell receptorax enhancer. J. Exp. Med. 173:1133-1142.
39. Orkin, S. H. 1992. GATA-binding factors in hematopoietic cells.Blood 80:575-581.
40. Perrin, S., and G. Gilliland. 1990. Site-specific mutagenesis usingasymmetric polymerase chain reaction and a single mutant primer.Nucleic Acids Res. 18:7433-7438.
41. Pevny, L., M. C. Simon, E. Robertson, W. H. Klein, S. F. Tsai, V.D'Agati, S. H. Orkin, and F. Constantini. 1991. Erythroid differ-entiation in chimaeric mice blocked by targeted mutation in thegene for transcription factor GATA-1. Nature (London) 349:257-260.
42. Prosser, H., D. Wotton, A. Gegonne, J. Ghysdail, S. Wang, N. A.Speck, and M. J. Owen. 1992. A phorbol ester response elementwithin the human T-cell receptor ,3 chain enhancer. Proc. Natl.Acad. Sci. USA 89:9934-9938.
43. Prosser, H. M., R. A. Lake, D. Wotton, and M. J. Owen. 1991.Identification and functional analysis of the transcriptional en-hancer of the human T-cell receptor R gene. Eur. J. Immunol.21:161-166.
44. Rahuel, C., M.-V. Vimit, V. Lemar-Chandel, J.-P. Carton, andP.-H. Romeo. 1992. Erythroid-specific activity of the glycophorin Bpromoter requires GATA-1 mediated displacement of a repressor.EMBO J. 11:4095-4102.
45. Redondo, J. M., J. L. Pfol, C. Hernandez-Munain, S. Wang, N. A.Speck, and M. S. Krangel. 1992. Indistinguishable nuclear factorbinding to functional core sites of the T-cell receptor 8 and murineleukemia virus enhancer. Mol. Cell. Biol. 12:4817-4823.
46. Romeo, P. H., M. H. Prandini, V. Joulin, V. Mignotti, M. Prenart,
VOL. 14, 1994
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 20
Feb
ruar
y 20
22 b
y 21
1.10
8.93
.162
.

MOL. CELL. BIOL.
G. Marguerie, and G. Uzan. 1990. Megakaryocytic and erythroidlineages share specific transcription factors. Nature (London) 344:447-449.
47. Schatz, D., M. A. Oettinger, and D. Baltimore. 1989. The V(D)Jrecombination gene, RAG-1. Cell 59:1035-1048.
48. Scheiber, E., P. Matthias, M. M. Muller, and W. Schaffner. 1988.Identification of a novel lymphoid specific octamer binding protein(OTF-2B) by proteolytic clipping bandshift assay (PCBA). EMBOJ. 7:4221-4229.
49. Takeda, S., A. Cheng, F. Mauxim, C. A. Nelson, R. D. Newbury,W. A. Sha, R. Sen, and D. Y. Loh. 1990. Functional analysis of themurine T-cell receptor beta enhancer and characterization of itsDNA-binding proteins. Mol. Cell. Biol. 10:5027-5035.
50. Travis, A., A. Amsterdam, C. Belanger, and R. Grosschedl. 1991.LEF-1, a gene encoding a lymphoid-specific protein with an HMGdomain, regulates T-cell receptor a function. Genes Dev. 5:880-894.
51. Tsai, S. F., D. I. Martin, L. I. Zon, A. D. D'Andrea, G. G. Wong,and S. H. Orkin. 1988. Cloning of cDNA for major DNA-bindingprotein of the erythroid lineage through expression in mammaliancells. Nature (London) 339:446-451.
52. van de Wetering, M., M. Oosterwegel, D. Dooijes, and H. Clevers.1991. Identification and cloning of TCF-1, a T lymphocyte-specific
transcription factor containing a sequence-specific HMG box.EMBO J. 10:123-132.
53. van de Wetering, M., M. Oosterwegel, K. van Norren, and H.Clevers. 1993. Sox-4, an Sry-like HMG box protein, is a transcrip-tional activator in lymphocytes. EMBO J. 12:3847-3854.
54. van Zonneveld, A.-J., S. A. Curriden, and D. J. Loskutoff. 1988.Type 1 plasminogen activator inhibitor gene: functional analysisand glucocorticoid regulation of its promoter. Proc. Natl. Acad.Sci. USA 85:5525-5529.
55. von Boehmer, H. 1988. T-cell antigen receptor and T-cell recog-nition. Annu. Rev. Immunol. 6:309-326.
56. Waterman, M. L., W. H. Fisher, and K. A. Jones. 1991. Athymus-specific member of the HMG protein family regulates thehuman T cell C ao enhancer. Genes Dev. 5:656-669.
57. Wilkinson, M. F., and C. L. MacLeod. 1988. Induction of T cellreceptor a and ,B mRNA in SL12 cells can occur by transcriptionaland post-transcriptional mechanisms. EMBO J. 7:101-108.
58. Winoto, A., and D. Baltimore. 1989. caj lineage-specific expressionof the a T cell receptor gene by nearby silencers. Cell 59:649-655.
59. Yancopoulos, G., T. K. Blackwell, H. Suh, L. Hood, and F. W. Alt.1986. Introduced T-cell receptor variable region gene segmentsrecombine in pre-B cells: evidence that B and T cells use acommon recombinase. Cell 44:251-259.
4294 HENDERSON ET AL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 20
Feb
ruar
y 20
22 b
y 21
1.10
8.93
.162
.