eyeing central neurons in vascular growth and reparative - blood
TRANSCRIPT

Review article
Eyeing central neurons in vascular growth and reparative angiogenesisPrzemyslaw Sapieha1,2
1Department of Ophthalmology & Biochemistry, Maisonneuve-Rosemont Hospital Research Centre, University of Montreal, Montreal, QC; and 2Department ofNeurology-Neurosurgery, McGill University, Montreal, QC
The generation of blood vessels is ahighly synchronized process requiringthe coordinated efforts of several vascu-lar and nonvascular cell populations aswell as a stringent orchestration by thetissue being vascularized. Stereotypedangiogenesis is vital for both developmen-tal growth and to restore tissue metabolicsupply after ischemic events. Central neu-rons such as those found in the brain,spinal cord, and retina are vast consum-ers of oxygen and nutrients and therefore
require high rates of perfusion by func-tional vascular networks to ensure propersensory transmission. During a meta-bolic mismatch, such as that occurringduring a cerebrovascular infarct or inischemic retinopathies, there is increas-ing evidence that central neurons have aninherent ability to influence the vascularresponse to injury. With a focus on theretina and retinal ischemic disorders, thisreview explores the ever-growing evi-dence suggesting that central neurons
have the propensity to impact tissue vas-cularization and reparative angiogenesis.Moreover, it addresses the paradoxicalability of severely ischemic neurons tohinder vascular regrowth and thus segre-gate the most severely injured zones ofnervous tissue. The topics covered hereare pertinent for future therapeutic strate-gies because promoting and steering vas-cular growth may be beneficial for isch-emic disorders. (Blood. 2012;120(11):2182-2194)
Introduction
The intricate wiring of nerve and vascular plexuses is arguably oneof the most monumental tasks facing developing tissue in higherorganisms. Throughout embryogenesis, nerves and blood vesselsestablish architecturally optimized networks to ensure adequatetissue perfusion and permit transmission of sensory information.The tight anatomical coupling of the 2 systems underscores theneed for honed guidance mechanisms to ensure proper targeting(Figure 1).1
Central neurons require a steady supply of nutrients and oxygento ensure appropriate function, and it is therefore key for nervousand vascular systems to be adequately paired. The ever-changingmetabolic requirements of the CNS necessitate dynamic perfusionrates because the brain only stores limited glucose. Hence, vasculardropout in the CNS and the ensuing deterioration of the neurovas-cular unit increasingly is being associated with several debilitatingneurodegenerative diseases.2 Examples include vascular demen-tia,3 which loosely encompasses syndromes that lead to vascularlesions of the white matter of the brain and are caused by generalvascular disease affecting the brain or by focal lesions such asstrokes. In Alzheimer disease, postmortem analysis reveals that upto two-thirds of patient brains show signs of stenosis resulting fromhyaline fibrosis of arterioles and smaller vessels.4 In addition,impaired vascular barrier function is a signature of multiplesclerosis5 and amyotrophic lateral sclerosis,6 where the blood-brainbarrier is compromised, resulting in inadequate cerebral and spinalperfusion.
The breakdown of vascular networks in neurodegenerative andneuroischemic conditions leads to hypoxic/anoxic events and aconstellation of biochemical changes that compromise cellularfunction.7-10 Restoring a functional and efficient vascular architec-ture is therefore key to regaining proper perfusion and neural tissuefunction. In this respect, devising strategies to accelerate therevascularization of ischemic nervous tissue with healthy vessels
may prove beneficial in salvaging the function of ischemictissue.11,12 Importantly, although blood vessels nourish the organsthrough which they course, aberrant and excessive vascularization,such as that observed in cancer13 and blinding ocular vasoprolifera-tive disease,9 contribute to the progression of disease. Therefore,therapeutic strategies aimed at regenerating vasculature will needto ensure controlled growth and accurate steering and homing ofnascent vessels.
This review addresses the emerging and ever-growing evidencesupporting a key role for central neurons in retinal vascularizationboth in health and disease. A clearer understanding of the molecularinterplay between central neurons and the vascular beds thatperfuse them will provide insight on prospective strategies totherapeutically modulate angiogenesis within the ischemic CNS.
Elucidating neurovascular interaction in theCNS: the retina as a model system
Perhaps the most accessible arena in which to study centralneurovascular interaction is the inner retina, where retinal ganglioncells (RGCs) are in close apposition with vessels of the inner retinalvascular plexus (Figure 1A). The retina is an experimentallyuser-friendly portion of the CNS (located outside the brain proper)and, given the anatomy of the ocular globe, has been poeticallydubbed the “window to the brain.”
RGCs are central neurons that have a compartmentalized layoutin that their somata lie within the eye, whereas their axons projectinto the optic nerve and synapse with subcortical regions of thebrain. From an experimental perspective, this anatomical layoutpermits for easy, minimally intrusive access to RGC cell bodiesthrough the vitreous and allows for selective labeling and manipu-lation of the optic nerve. Together, these features have made the
Submitted April 14, 2012; accepted June 12, 2012. Prepublished online asBlood First Edition paper, June 15, 2012; DOI 10.1182/blood-2012-04-396846.
© 2012 by The American Society of Hematology
2182 BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom

rodent retinocollicular pathway a system of choice to study theCNS in health and disease, starting with the pioneering work ofSantiago Ramon y Cajal on axonal regeneration of the rabbit opticnerve.14
Similarly, the retina, and more specifically the rodent retina, hasbeen instrumental in the study of angiogenesis both in physiologicand pathologic contexts.15 Comparable with the aforementionedbenefits for the study of the CNS, the retinal vascular systemspresent numerous advantages that lend themselves particularly wellto experimental manipulation. Most notably, the rodent retinalvascular plexuses develop during the first postnatal weeks of life ina highly stereotyped, temporal, and reproducible pattern (reviewedin Stahl et al15 and Dorrell and Friedlander16). Therefore, theoutcome of genetic17 or pharmacologic18-20 manipulation on vascu-lar phenotypes can be readily imaged and assessed postterm, thusavoiding the complications associated with the analysis of embry-onic vasculature. This attribute has been exploited to make severalseminal discoveries on the mechanisms governing mammalianangiogenesis, including the first identification of the tip cell,21 theelucidation of the roles of different VEGF isoforms in arteriolar andvenular patterning,22 the signaling circuits leading to endothelial
cell specialization,23-25 as well as the fundamental role of myeloidcells in angiogenesis.26,27
Physiologic vascular development in theretina
A functional circulatory system is the first of the major embryonicsystems to arise. Initially, it must provide the growing embryo withnourishment before the formation of an intestine, distribute oxygenpreceding the appearance of lungs, and evacuate waste before thepresence of kidneys. Hence, the embryonic vasculature is designedsuch that nutrients are obtained from the placenta or yolk sac andrespiration and gas exchange is provided by chorionic and allantoicmembranes.
The inner eye initially is sustained by the hyaloid vascula-ture, which consists of an arterial network residing in thevitreous. The hyaloid vasculature is perfused via the centralhyaloid artery residing in the optic nerve and drained by anannular system of collection vessels at the forefront of the eye.At approximately 16 weeks of gestation in humans, retinalvessels commence, invading the retina from the optic nerve headtoward the periphery and gradually replacing the residenthyaloid vessels. In mouse models, regression of hyaloid vesselswas shown to occur via the WNT7b short-range paracrine signalfrom macrophages.28 During ocular maturation, macrophagesinitiate a prodeath program in the vascular endothelial cells ofthe transient hyaloid vessels using the Wnt ligand to dictatevascular regression.
In several mammalian species, such as echidnas, guinea pigs,and rabbits, the retinal vasculature is absent and the oxygensupply to the full thickness of the retina is provided by diffusionfrom the choriocapillaris of the outer retina. Interestingly, thepresence of an additional blood supply from vessels located inand on the retina is directly related with retinal thickness wherevascularized retinas are approximately 60% thicker than thosewithout retinal vessels.29-31 In mammals such as mice andprimates, 3 retinal and intraretinal vascular layers form inconserved growth patterns over various time scales.32 Theformation of the primary retinal vascular plexus and subsequentdeeper layers is thought to occur largely by angiogenesis33
whereas vasculogenesis may occur in other species.16
The mouse superficial plexus forms during the first week of lifeand reaches the extremity of the retina at approximately postnatalday (P)8 (Figure 2A; retinal vascular development in C57Bl/6mice). Retinal capillaries commence to dive down at P7 and formthe deep plexus by approximately P12 and finally invade theintermediate layer from P12 to P15. Retinal vascularization is fullycompleted and interconnected by the third week of life. Severalmechanisms are in place to ensure stereotyped patterning of retinalvessels.15,16 These rely heavily on the interaction between nascentvessels and the asctrocytic template on which they grow. Astrocytegenesis precedes vascular development and begins forming atembryonic day 19 and is fully developed at birth.34 Astrocytesexpress PDGFR-� and invade the developing retina from the opticnerve head ahead of the vascular front. They travel on top ofPDGF-�–expressing RGCs,41 emphasizing the relationship be-tween neurons, astrocytes, and vessels. As vessels sprout, theyalign with astrocytic fibers and are stabilized by R-cadherin–mediated cell–cell adhesions between the endothelial filopodia andnearby astrocytes (Figure 2B).21,34,35 Although astrocytes express
Figure 1. Central neurons and blood vessels are anatomically coupled in theretina. (A) Three-dimensional reconstruction of an adult mouse retinal flatmount. Theimage depicts the inner retinal vascular plexus (red; stained with isolectin B4)interwoven with retinal ganglion neurons and their axons (green; stained with �-IIItubulin). (B) Corrosion cast of mouse retinal vasculature imaged by environmentalscanning electron microscopy. The image shows a network of microcapillaries, veins,and arteries and illustrates the complex wiring of retinal vascular beds. Scale bar:(A) 400 �m; (B) 50 �m.
NEURONAL INFLUENCE IN OCULAR VASCULARIZATION 2183BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom

angiogenic factors such as VEGF36 at birth, the indispensable rolefor astrocyte-derived growth factors in developmental vasculariza-tion recently has been challenged.37,38 It is therefore plausible thatother highly metabolic cells such as RGCs, which produceangiogenic factors throughout development, play a critical role ingoverning retinal developmental vascularization (discussed in“Neuronal influence on retinal vascularization”).19
The retina is, per weight, the most metabolically active tissue ofthe body. Regions that are poorly supplied in oxygen and nutrientsprompt the formation of angiogenic sprouts from the walls ofpreexisting vessels in response to O2-tension. This process ofadaptation is largely orchestrated by oxygen-sensing machinerysuch as the prolyl hydroxylases, which regulate the transcriptionfactor hypoxia-inducible factor (HIF).39 HIF consists of 2 distinct�- and �-subunits that are required to heterodimerize to beactive.40 HIF-1� is the constitutive unit, whereas HIF-1� andHIF-2� are continuously transcribed but only have transienthalf-lives of 5 minutes in normoxic conditions.41 The rapidturnover is mediated by the oxygen-dependent proline hydroxy-lation of HIF-1� or HIF-2� by prolyl hydroxylases. The
ultimate result is polyubiquitination and targeting for protea-somal degradation. Alternatively, in hypoxic conditions, prolinehydroxylation is inefficient, and HIF-1� or HIF-2� are stabi-lized and translocate to the nucleus and dimerize with HIF-1�.The outcome is the transcription of a cluster of target genesthat have a hypoxia-response element.42 Presently, more than70 genes have been described that encode proteins (such asVEGF43 and Epo44) that help promote the adjustment to ahypoxic state45,46 and indirectly participate in instating localtissue perfusion.
Evidence for neuronal involvement in sensing and respondingto hypoxia is substantiated by the fact that HIF-1� is mostabundantly expressed in the neuroretina.47 Correspondingly,neuroretinal-specific knockout of HIF-1� impairs vascular develop-ment by reducing tip cell filopodia as well as vessel branching47
and leading to the absence of the intermediate vascular plexus.48
Interestingly, although a pronounced retinal vascular phenotype isnoted in these animals, VEGF levels are unaffected,48 suggestingthat HIF1� may not be the main mechanism by which VEGF isregulated in response to hypoxia.
Figure 2. Physiologic retinal vascularization. (A) Schematic representing the approximate time course of retinal vascularization in a C57Bl/6 mouse. The astrocytic template(green) precedes nascent vessels and is fully laid out at birth. The superficial vascular network grows between birth and P8, the deep plexus forms approximately between P7and P12, whereas the intermediate plexus forms during the third week of life. (B) Model of vascular growth during retinal development. Astrocytes express PDGFR-� andinvade the developing retina from the optic nerve head, ahead of the vascular front. They travel on top of PDGF-A–expressing RGCs. Nascent vessels follow the astrocytictemplate and form R-cadherin junctions with proximal astrocytes.
2184 SAPIEHA BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom

Vascular growth and guidance: conservedmechanisms in neuronal and vascularpatterning
The complexity of neurovascular networks is unmistakable, giventhe elaborate webs of hundreds of billions of neurons in humans,each communicating with thousands of other neurons, in proximityand irrigated by a vessel.49 In the brain, spinal cord, and retina, aneuron must extend its axon over long distances into a maze ofchemical signals and physical barriers and ensure it arrives inproximity of the adequate target and generates a functionalsynapse. This growth process relies essentially on the axon’s abilityto read and interpret molecular growth and guidance cues. Simi-larly, albeit through a different process, vessels within the CNSundergo sprouting angiogenesis and must be adequately guided totheir destination to ensure functional perfusion of nervous tissue.During nerve growth, a single neuron extends an axon directlyfrom its cell body whereas vessel growth consists in a process ofendothelial cell delamination and assembly.50 Both neurons andvessels require honed guidance systems to ensure functionalcoupling and formation of healthy neurovascular units. Althoughthe nervous system emerged earlier in evolution, nascent vesselsshare the same arsenal of molecular guidance cues to ensure propertargeting.1
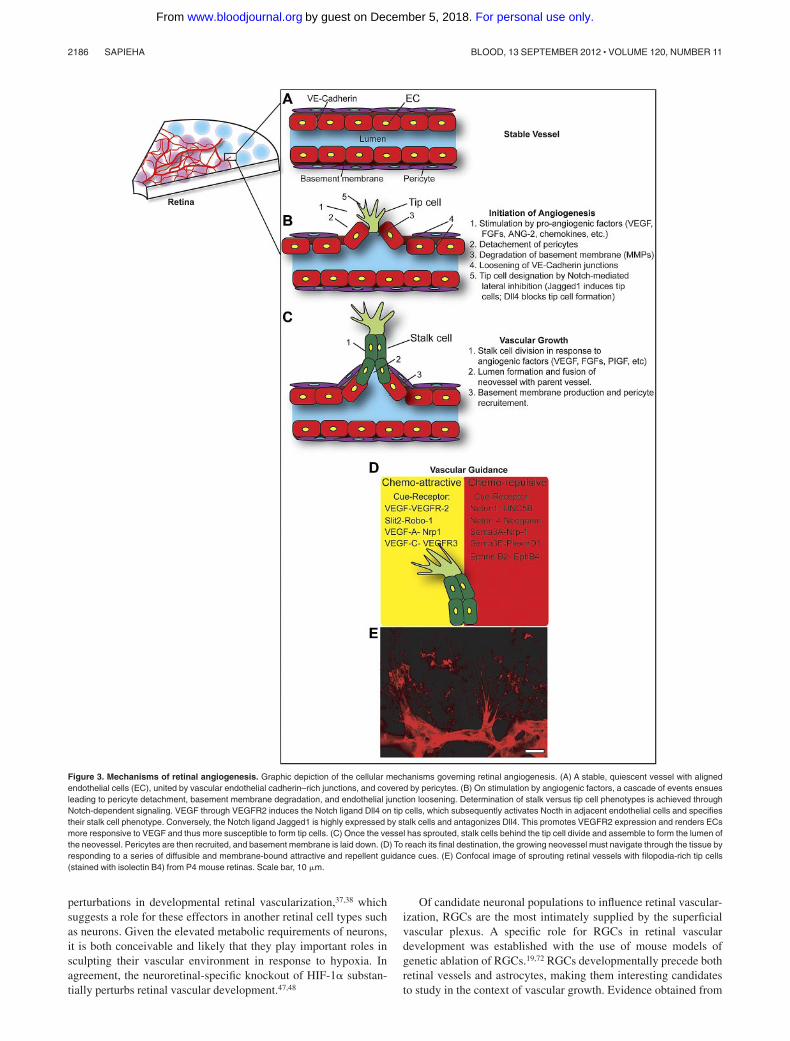
The vascular analog of the growth cone is the tip cell.21 In theadult, blood vessels are quiescent, yet endothelial cells maintainelevated plasticity and are able to transduce and respond toangiogenic signals. Angiogenesis in the retina (as elsewhere in thebody) is a concerted process involving endothelial cell prolifera-tion, migration, and assembly into tube-like structures containing alumen.16,51 Vessel growth is instigated by elevated levels ofangiogenic factors such as VEGF, fibroblast growth factors,ANG-2, and/or other chemokines. In response to angiogenicstimuli, perictyes detach from the vessel wall (ANG-2 mediated),basement membrane is degraded by matrix metaloproteinases,endothelial cells slacken their vascular endothelial cadherin andclaudin-rich junctions, and a designated leading cell (tip cell)protrudes and advances toward vaso-attractant chemotactic gradi-ents (Figure 3A-C; reviewed in Carmeliet and Jain50).
Which cell becomes the leader (tip) and which follows (stalk) isestablished by Notch-mediated lateral inhibition, and the underly-ing mechanisms have been worked out in a series of sophisticatedcell biology studies in which the mouse retina is used as a model.After stimulation of VEGFR-2 by VEGF, tip cells up-regulate theNocth ligand Dll4 (however, possibly modestly52; discussed be-low).23,25,53 The elevated levels of Dll4 in filopodia-rich endothelialtip cells activate Notch receptor in neighboring endothelial cellsand thus block generation of tip cells.23,25 This is achieved throughDll4-Notch–mediated down-regulation of VEGFR-2 on stalk cellsand simultaneous up-regulation of VEGFR-1. The stalk cells thenbecome less sensitive to VEGF-induced sprouting and becomemore responsive to VEGFR1-driven proelongation factors.
Conversely, the Notch ligand Jagged1 (which is expressed bystalk cells) antagonizes Dll4-Notch signaling and is a potentproangiogenic regulator that induces tip cell formation.24 This earlycellular specialization prevents nondiscriminant endothelial move-ment toward angiogenic signals and thus ensures the presence ofstalk cells in the wake of the advancing tip cell to form a luminaltube.54 Stalk cells respond to VEGF by proliferating21 and becomerapidly stabilized and establish adherent tight junctions to maintain
the integrity of the nascent vessel.55 The final maturation of thevascular plexus requires pruning of excess vessels,56 coalescence ofexisting vessels, and the recruitment of mural cells.57
Interestingly, beyond the canonical VEGF-A/VEGFR-2 para-digm, recent evidence points to an important role in retinalangiogenesis for VEGF-C and its main receptor, VEGFR-3, andchallenges the current views on the molecular cross-talk betweenthe VEGF and Notch pathways. The authors propose that VEGFR-3is robustly modulated by Notch, whereas expression of Dll4 inretinal tip cells is only weakly affected by VEGFR-2 signaling andNotch inhibition has minimal impact on VEGFR-2 expression.53
Importantly, VEGFR-3 kinase activity inhibitors block sproutingfrom endothelial cells that have low Notch signaling activity. Thisnovel insight suggests that effective antiangiogenic strategiestargeting these receptors and their ligands will be dependent on thestatus of endothelial Notch signaling.53 In addition, another role forVEGF-C is substantiated by new evidence suggesting thatmacrophage-derived VEGF-C activates VEGFR-3 in tip cells andaccentuates Notch signaling and partakes in the phenotypic conver-sion of endothelial cells at fusion points of vessel sprouts.58
In contrast to stalk cells, which respond to VEGF by dividing,tip cells activate Cdc42 and form motile filopodia protrusions thatprobe the environment and transduce extracellular signals (Figure3D-E).59 Vascular tip cell filopodia (as their neuronal analogous)are enriched in receptors that were classically described to respondto neuronal guidance cues.60 These include the neuropilins andplexins (for semaphorins); Unc5b, neogenin, and DCC (for ne-trins); the Eph receptors (for ephrins); and roundabouts (for slits;Figure 3D).61-63 Once the tip cells comes in contact with a givencue, it will respond by either advancing, stalling, turning, orretracting depending on which cell surface receptor predominatesand the overall intracellular environment of the tip cell.64 The roleof neuronal guidance cues in vascular growth has been comprehen-sively reviewed.1,59,64,65 Evidence for deregulation of neuronalguidance cues in neurovascular pathology is starting to emerge66-71
and will be discussed in the context of ischemic retinopathiesbelow.
Neuronal influence on retinal vascularization
A growing body of evidence is highlighting the key role of neuronsin instigating, promoting, and steering angiogenesis within nervoustissue and specifically in the retina.19,47,70,72,73 A direct role forretinal neurons in influencing vascular growth is observed inhumans during the formation of the outer vascular plexus of thehuman fetal retina in utero (25-26 weeks).74 This period coincideswith the first appearance of visually evoked potentials and thusfunctional neurons.75 A likely explanation for this concurrencestems from the increase in oxygen consumption (and thus aug-mented metabolic requirement) of newly functioning neurons andthe consequent generation of local regions of hypoxia.76
Notwithstanding the key contribution of glial cells such asastrocytes in providing a framework on which nascent vesselsadvance,21,35,77 their role as the principal source of trophic supportfor retinal vascular development may need to be revisited. Evi-dence for a retinal cell population other than astrocytes providingindispensible angiogenic factors for retinal vascularization comesfrom genetic studies in mice in which the hypoxic response wasdisrupted directly in astrocytes by cell-specific deletion of VEGF,HIF-1�, or HIF-2�. Using this approach, we noted no visible
NEURONAL INFLUENCE IN OCULAR VASCULARIZATION 2185BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom
perturbations in developmental retinal vascularization,37,38 whichsuggests a role for these effectors in another retinal cell types suchas neurons. Given the elevated metabolic requirements of neurons,it is both conceivable and likely that they play important roles insculpting their vascular environment in response to hypoxia. Inagreement, the neuroretinal-specific knockout of HIF-1� substan-tially perturbs retinal vascular development.47,48
Of candidate neuronal populations to influence retinal vascular-ization, RGCs are the most intimately supplied by the superficialvascular plexus. A specific role for RGCs in retinal vasculardevelopment was established with the use of mouse models ofgenetic ablation of RGCs.19,72 RGCs developmentally precede bothretinal vessels and astrocytes, making them interesting candidatesto study in the context of vascular growth. Evidence obtained from
Figure 3. Mechanisms of retinal angiogenesis. Graphic depiction of the cellular mechanisms governing retinal angiogenesis. (A) A stable, quiescent vessel with alignedendothelial cells (EC), united by vascular endothelial cadherin–rich junctions, and covered by pericytes. (B) On stimulation by angiogenic factors, a cascade of events ensuesleading to pericyte detachment, basement membrane degradation, and endothelial junction loosening. Determination of stalk versus tip cell phenotypes is achieved throughNotch-dependent signaling. VEGF through VEGFR2 induces the Notch ligand Dll4 on tip cells, which subsequently activates Nocth in adjacent endothelial cells and specifiestheir stalk cell phenotype. Conversely, the Notch ligand Jagged1 is highly expressed by stalk cells and antagonizes Dll4. This promotes VEGFR2 expression and renders ECsmore responsive to VEGF and thus more susceptible to form tip cells. (C) Once the vessel has sprouted, stalk cells behind the tip cell divide and assemble to form the lumen ofthe neovessel. Pericytes are then recruited, and basement membrane is laid down. (D) To reach its final destination, the growing neovessel must navigate through the tissue byresponding to a series of diffusible and membrane-bound attractive and repellent guidance cues. (E) Confocal image of sprouting retinal vessels with filopodia-rich tip cells(stained with isolectin B4) from P4 mouse retinas. Scale bar, 10 �m.
2186 SAPIEHA BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom

transgenic and knockout mice supports the importance of theseneurons in physiologic retinal vascularization. In transgenic brn3bZ-dta/�;six3-cre mice engineered to express diphtheria toxin A underthe control of a RGC-specific promoter, RGCs are ablated as theyform.78 Remarkably, brn3bZ-dta/�;six3-cre mice are completelydevoid of a retinal vascular plexus and instead show persistenthyaloid vasculature.19 Similarly, Math 5�/� mice, which lack 95%of RGCs, also do not form a retinal vascular layer.72 Of note,although the astrocytic network remains largely intact in brn3bZ-dta/�;six3-cre mice, retinal vascularization is halted. Together,these studies suggest that RGCs play a central role in instigatingretinal vascular development, whereas the astrocytic beds providethe framework on which nascent vessels advance.
Beyond RGCs, the most oxygenated and highly metabolicallyactive retinal neurons are photoreceptors.79 As RGCs, these neu-rons have also been shown to influence retinal angiogenesis.Evidence comes directly from clinical observations in whichpatients suffering from both proliferative diabetic retinopathy andretinitis pigmentosa (a group of eye conditions that lead toprogressive photoreceptors loss) show considerably less pathologicretinal angiogenesis than diabetic patients with healthy photorecep-tors.80 In addition, the retinal neovascularization that is associatedwith long-standing diabetes mellitus has been reported to spontane-ously regress with the onset of clinically evident retinitis pigmen-tosa.81 This observation held true in animal models in which micewith genetically ablated photoreceptors failed to mount reactiveretinal neovascularization in a model of oxygen-induced prolifera-tive retinopathy81 whereas diabetic mice with photoreceptor degen-eration demonstrated lower levels of retinal proangiogenic VEGF.82
The elevated energetic requirements of photoreceptor neurons83
may explain this phenomenon because the loss of photoreceptorsresults in a significantly reduced energetic burden on the tissue andthus a proportionately decreased requirement for perfusion.
Pairing neuronal energy demand to vascular supply may beefficiently achieved by directly harnessing the intermediates of theneuron’s own energy metabolism. In this respect, signaling viaenergy metabolites in response to compromised energy status hasbeen proposed as a contributor to both physiologic and pathologicretinal vascularization.19,84-86 In the context of neurovascularinterplay, this evolutionarily preserved signaling system has thusfar been described for RGCs and the dicarboxylate succinate.Succinate is a metabolite of the Krebs cycle generated duringcellular respiration. Although it plays a canonical role in energyproduction, succinate also binds and activates the formerly orphanG-protein–coupled receptor GPR91,87 inferring additional physi-ologic roles. In the retina, GPR91 is predominantly expressed inRGCs.19 Importantly, Krebs cycle intermediates, such as succinate,accumulate under conditions of hypoxic stress s because offeedback inhibition of succinate (and �-ketoglutarate) dehydroge-nase by nonoxidized flavin and nicotinamide nucleotides and byreactive oxygen species.88,89 The accumulation of succinate duringmetabolic compromise is consistent with a role in mediatingcompensatory angiogenesis to reinstate adequate blood supply andoxygen delivery. The robust retinal vascular response provoked bysuccinate via its neuronal receptor GPR91 (production of VEGF,angiopoietin 1 and 2 [Ang-1 & -2]) is suggestive of a direct role inlinking neuronal energy demand to capillary growth (Figure 4A-B).Correspondingly, succinate signaling through GPR91 can influenceboth developmental and pathologic retinal angiogenesis evenbefore HIF stabilization,19 which suggests that the succinate–GPR91 axis can act as an early sensor of hypoxic stress and work toenhance regional circulation through the release of angiogenic
factors such as VEGF and Ang-1 & -2 (Figure 3C). The HIF-independent angiogenic potential of succinate may explain the lackof variation in developmental VEGF levels in HIF-deficient mice.48
Ischemic retinopathies: microvasculardegeneration and neuronal ischemia in theretina
The breakdown of functional neurovascular circuits in the retina isobserved in sectors of nonperfused neuronal tissue in ischemicretinopathies. Given the importance of neuronal and vascularinterplay in retinal health and disease,19,70,71,73 it is important tounderstand the neuronal contribution to the progression of thesedebilitating retinal diseases.
Ischemic retinopathies encompass the initial phases of diabeticretinopathy (DR) and retinopathy of prematurity (ROP) andrepresent the leading causes of blindness in working-age adults andchildren, respectively.90,91 Both diseases are characterized by anearly phase of microvascular degeneration followed by a secondphase of pathologic neovascularization mounted by the ischemicretina attempting to reinstate metabolic equilibrium.92-94 Becauseof this secondary phase of excessive vascular regrowth, thediseases also are referred to as proliferative retinopathies.
In ROP, the ischemic stress on the neural retina is a result ofboth vascular degeneration and incomplete developmental vascular-ization. To overcome the pulmonary insufficiency of prematureinfants, artificial ventilation can be increased to a relativelyhyperoxic PaO2 of up to �100 mmHg,95 whereas normal in uteroO2 tension averages 32 mmHg in the umbilical cord, and afull-term infant will be exposed to a PaO2 of 60-100 mmHg inroom air.96 This exposure to high PaO2 induces microcapillary degen-eration by oxygen toxicity directly on endothelium via reactiveoxygen species (reviewed in Sapieha et al10 and Hardy et al97). Inaddition, the hypoxic stress resulting from oxygen-induced vascu-lar dropout is compounded by the fact that premature infants areborn with an incomplete retinal vascular plexus; the developmentof human retinal vasculature concludes at term (40 weeks).98
Ex-utero, after premature birth, the physiologic vascularizationprocess of the immature retina is perturbed because of the absenceof maternally transferred vasopotentiating and vasoprotective fac-tors such as insulin growth factor-199 and �-3 polyunsaturated fattyacids.10 In addition, the excess oxygenation resulting from ventila-tion protocols suppresses key oxygen-regulated proangiogenicgrowth factors such as VEGF and Epo,10 thus further stalling thedevelopment of a mature retinal vascular supply.
In DR, the pathomechanisms governing the loss of endothelialbarrier function and later vascular degeneration remain largelyelusive. Although often initially asymptomatic, loss of vision istriggered primarily by diabetic macular edema, vitreal hemor-rhages, and later tractional retinal detachment. Thus far, associa-tions have been made between disease progression and advancedglycation end-products,100 protein kinase C activation,101 oxidativestress,101 and initial onset of acute intensive insulin therapy.102
Moreover, several salient features of chronic inflammatory diseaseare seen in DR, and increasing evidence points to the contributionof inflammatory mediators such as IL-1�103 and TNF-�104 in bothvascular degeneration and neovascularization associated with DR(reviewed in Adamis and Berman105).
NEURONAL INFLUENCE IN OCULAR VASCULARIZATION 2187BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom

Failure of vascular regeneration in ischemicretinopathies: neuronal inhibition ofreparative angiogenesis
The limited efficacy with which blood vessels regenerate intoischemic neural tissue after vascular injury represents a noteworthychallenge in vascular biology. A direct example lies in the failure ofrevascularization of hypoxic regions of the retina in ischemic
retinopathies (Figure 5A).9,106 Both ROP and DR feature avascularpockets that are the source of hypoxic stress that instigatesderegulated retinal neovascularization.106 Therapeutically promot-ing angiogenesis into these avascular areas may aid in easing thehypoxic burden in the retina and thus alleviate the neural ischemiathat is central to disease progression.70,71
Paradoxically, although the retina mounts a robust neovascularresponse secondary to microvascular degeneration, the nascentvessels in both ROP and DR fail to effectively regenerate into the
Figure 4. Neuronal influence in retinal vascular development. (A) The superficial retinal vascular plexus forms in a stereotyped manner. As the vascular front advances, itleaves perfused RGCs in its wake. (B) The proposed model for RGC-dependent vascularization stipulates that the mildly hypoxic RGCs ahead of the vascular front accumulateenergy metabolites such as sucinnate because of the feedback inhibition of succinate dehydrogenase. The elevated levels of succinate then stimulate its RGC-specific cognatereceptor GPR91, resulting in the production of proangiogenic cues such as VEGF and Ang-2. RGCs therefore have the inherent ability to instate local microcirculation. (C) TheEC50 for succinate-mediated activation of GPR91 is � 10-fold lower than the Ki required for succinate-induced inhibition of prolyl 4-hydroxylases and consequent HIF-1�stabilization. This suggests that succinate through GPR91 acts as an early and accurate sensor of hypoxic stress.
2188 SAPIEHA BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom

most ischemic vaso-obliterated regions. The neovascularization inischemic retinopathies presents 2 prominent features that render itpathologic. First, it is initially concentrated at the avascular borderof the injured retina and hence fails to grow into and adequatelyperfuse the hypoxic tissue (Figure 5A). Second, the leaky abnormalvessels are misdirected toward the physiologically avascular vitre-ous, predisposing to traction detachment of the retina and loss of
vision. The classic interpretation of the misdirected retinal neovas-cular growth has simplistically been explained by the presence ofhigh concentrations of proangiogenic factors, such as VEGF, in thevitreous of patients with ischemic retinopathies. However, glialcells within the neural retina itself (astrocytes and Muller cells)36
and neurons (RGCs)19 produce elevated amounts of these angio-genic factors and therefore would be expected to retain nascent
Figure 5. Neuronal influence in retinal ischemic disease. In ischemic retinopathies such as that of diabetes, regional microvascular decay yields zones of avascular neuraltissue. (A) Angiogram of a human patient with DR. The red line delineates a zone of microcapillary loss with adjacent preretinal proliferation (white neovascular tuft). (Imagecourtesy of Dr Andreas Stahl, University Eye Hospital Freiburg.) (B) The schematic illustrates that in the avascular retina, hypoxic RGCs produce vasorepulsive semaphorinsthat repel nascent vessels into the vitreous and away from avascular zones of the retina. (C) Although ischemic astrocytes in the avascular retina produce VEGF via aHIF-2�–mediated mechanism, severely ischemic retinal ganglion neurons produce semaphorin 3A (D) and semaphorin 3E. (E) Astrocyte-derived VEGF stimulates vesselgrowth, whereas RGC-derived semaphorin 3A and 3E deviate vessels and thus impede reparative angiogenesis by repelling neovessels away from the avascular neural-retinatoward the vitreous. (F) Hypoxia/ischemia is a major promoter of angiogenesis through mechanisms such as metabolite signaling and HIF-1� stabilization. When hypoxiapersists and neurons are driven beyond a metabolic threshold, they revert to producing classic neuronal repulsive cues such as semaphorin 3A and 3E.
NEURONAL INFLUENCE IN OCULAR VASCULARIZATION 2189BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom

vessels on the retinal surface. Yet, neovessels continue to growaway from the hypoxic regions of the retina into the vitreous,suggesting that vasorepulsive forces are at work.70,71
The prominent clinical/pathologic features of DR and ROP havegiven rise to the notion that these diseases are entirely microvascu-lar in nature. However, evidence points to early changes in theneural retina107-109 and specifically in RGCs,110-112 which areintimately supplied by the same vascular network that is perturbedin ischemic retinopathies. Interestingly, although RGCs have ademonstrated role in governing retinal angiogenesis through produc-tion of angiogenic factors,19,72 when driven beyond a givenmetabolic threshold, severely hypoxic RGCs within vaso-obliterated retinal zones revert to producing vasorepellent cuessuch as the secreted class III semaphorins113 (semaphorin 3A71 and3E70; Figure 5B).
Consistent with a role for semaphorin 3A and 3E in blockingentry of vessels into ischemic retinal cores, extraretinal neovesselsexpress the cell surface receptors neuropilin-1 for semaphorin3A71
and plexinD1 for semaphorin3E (Figure 5B).70 Neuropilin-1 is asingle-pass transmembrane receptor with a large 860 amino acidextracellular domain subdivided into 3 subdomains (a1a2, b1b2,and c) and a short 40 amino acid intracellular domain (Figure5C).114 In neurons, the binding of semaphorin 3A to neuropilin-1recruits plexins, which transduce their intracellular signal115 andprovoke cytoskeletal collapse; the transduction mechanism inendothelial cells remains ill-defined.65 Neuropilin-1 has the uncon-ventional ability to bind 2 structurally unrelated ligands via distinctsites on its extracellular domain.116-118 It binds semaphorin 3A62,119
primarily via its a1a2 (but also b1) domains (provoking cytoskel-etal collapse) and VEGF165
62,117,118,120 via its b1b2 domain (enhanc-ing binding to VEGFR2 and thus increasing its angiogenicpotential121). Crystallographic elucidation of neuropilin-1 revealedthat VEGF165 and semaphorin 3A do not directly compete forneuropilin-1 binding but can simultaneously bind to neuropilin-1 atdistinct, nonoverlapping sites.122 Moreover, well-designed geneticstudies show that neuropilin-1 separately regulates the effects ofVEGF and semaphorin 3A on neuronal and vascular develop-ment.123 The elevated levels of semaphorin 3A in the ischemicretina may thus partake in forcing neovessels into the vitreous bycollapsing and deviating the advancing tip cells away from thesource of the repellent cue (Figure 5C-E).71
A similar pathomechanism can be observed with semaphorin3E, which signals through plexinD1 and activates the smallGTPase RhoJ in endothelial cells.70 As with semaphorin 3A, theactivation of plexinD1 by semaphorin 3E counteracts VEGF-induced filopodial projections. It is important to note that at thesame time as severely hypoxic RGCs produce vasorepulsive cues,certain RGCs in the avascular continue to produce VEGF.19,70
Moreover, astrocytes (which physically lie between RGCs and thevitreous) readily drive angiogenesis during the vasoproliferativephase of retinopathy by producing VEGF (via HIF-2� but not byHIF-1�; Figure 5C).37 Hence, in accordance with this paradigm,the neovascular sprouts are deviated from the ischemic core of theretina toward the vitreous and contribute to the pathologic prereti-nal neovascularization observed in ischemic retinopathies70,71
Understanding the dichotomy in the behavior of retinal gan-glion neurons where they are proangiogenic when subjected tomild levels of hypoxia19 and antiangiogenic when hypoxia issustained70,71 may be central to understanding the progression ofischemic retinopathies. This phenomenon may be a survivalmechanism to segregate irreversibly damaged sectors of the neural
retina and redirect metabolic supply to less affected and moresalvageable and will be discussed below.
The dichotomy of the neuronal response toischemia: the neurovascular shunt
In neuroischemic conditions such as retinopathies or stroke,vascular dropout is associated with compromised neuronal func-tion.11,12,107,108 To counter this early injury, attempts to reinstateregional microcirculation may come directly from the accumula-tion of energy metabolites such as succinate88,89 and activation ofmetabolite receptors like GPR91. Importantly, succinate triggers ahalf-maximal response on GPR91 with reported effective concen-trations of 28-56mM.87 These concentrations are approximately10-fold lower than the Ki values of 350-460mM required forsuccinate-induced inhibition of prolyl 4-hydroxylases and conse-quent HIF-1� stabilization.124,125 Succinate, through GPR91, maythus act as a more sensitive sensor of hypoxia than HIF-1� andpromote the release of finely tuned concentrations of proangiogenicfactors to enhance regional circulation and overcome metabolicimbalances in the CNS. A similar HIF-independent production ofVEGF has been reported for peroxisome proliferator–activatedreceptor-� coactivator-1�, which partakes in the regulation ofoxidative phosphorylation, mitochondrial biogenesis, and respira-tion.126 The role of peroxisome proliferator–activated receptor-�coactivator-1� in CNS angiogenesis has yet to be determined.
Although hypoxia/ischemia is undeniably a major promoter ofangiogenesis,127 when hypoxia persists and retinal neurons aredriven beyond a metabolic threshold, they revert to producingclassic neuronal repulsive cues such as semaphorin 3A71 and 3E70
(Figure 5F). The up-regulation of vasorepulsive cues such assemaphorins by severely injured cells in the CNS may be ageneralized event as levels of semaphorin 3A increase afterischemic stroke66,128 and localize to regions immediately adjacentthe zone of infarct and necrotic core.67 Similarly, semaphorin 3A isinduced after spinal cord injury in neurons near the site of lesion.69
The induction of repulsive guidance cues after CNS injuryoccurs during a period of vascular remodeling and axonal sprout-ing, suggesting that the severely damaged nervous tissue isattempting to deviate both regenerating neuronal sprouts andvessels away from the severely lesioned zones. It is thereforeconceivable that if driven beyond the threshold of recovery, heavilyhypoxic neurons mount a repulsive front in an attempt to shuntmetabolic resources away from perishing unsalvageable ischemictissue toward less affected regions of the CNS (Figure 5B and F).Reestablishing a vascular network to neurons that are unsalvage-able would be wasteful. In agreement with the theory of segregat-ing severely damaged areas of tissue, induction of semaphorin 3Ain central retinal neurons requires prolonged exposure to inflamma-tory cytokines such as IL-1�,71 a paradigm mimicking prolongedexposure to ischemia. This idea is further substantiated by the latentappearance of semaphorin 3A production at the expense of VEGFin RGCs subjected to direct hypoxic stress.71 Although early duringhypoxia, RGCs produce VEGF to reinstate vascular supply,19 whenhypoxia is sustained, RGCs reverse their signaling machinery fromVEGF production to that of the semaphorin 3A.
Although this hypothesized survival strategy may prove benefi-cial in the long term for cerebral ischemia, vascular deviation in theretina may be at the heart of pathologic extraretinal neovasculariza-tion. Given the physical properties of the eye and the lowerresistance encountered in the vitreous humor compared with the
2190 SAPIEHA BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom

retina, redirecting nascent vessels away from the ischemic retinawould deviate them toward the vitreous and result in the cardinalfeatures of proliferative retinopathies (described above).9 Hence,devising strategies to steer nascent vessels into avascular pocketsof the retina while keeping them in the retinal surface and in contactwith the underlying astrocytic bed may be therapeutic value forischemic retinopathies.
Neuronal guidance cues for therapeuticangiogenesis
Promoting angiogenesis to remedy ischemic CNS disorders maycome across as an unsubtle strategy. Yet, in the retina, it may beconsidered counterintuitive given the uncontestable involvement ofproangiogenic factors such as VEGF in the pathogenesis of ocularneovascular disease.129-131 In addition, several antiangiogenic strat-egies currently are being explored and recommended to counterproliferative ocular disease.132 However, potential complicationswith anti-VEGF therapies, including a greater incidence of strokein patients with a history of preexisting cerebrovascular disease,133
neurotoxicity,134 nonocular hemorrhage,135 and the need for re-peated injections, could limit its overall utility. An alternativestrategy may lie in directly promoting revascularization of theischemic tissue that is at the source of the angiogenic stress.Although presently still in the experimental stages, the authors of agrowing number of animal studies suggest that promoting vascularregeneration during the early ischemic phase of retinopathies maysignificantly reduce the destructive neovascularization that iscentral to disease progression.70,71,136,137 Enhancing normalizedvascular regrowth in ischemic retinopathies has previously beenachieved through the injection of myeloid progenitors,136 increas-ing dietary �-3 polyunsaturated fatty acid intake,138 or by directadministration of VEGF or FGF-2 via protection of glial cells.137
The pronounced effect of neuronal guidance cues on vascularremodeling and a role in blocking vascular regeneration70,71,139
makes them attractive candidates to modulate for reparativeangiogenesis. Evidence for the therapeutic potential of inhibitingguidance cues has been provided in models of limb ischemia indiabetic mice in whom blocking semaphorin 3E markedly im-proved the response to VEGF and promoted reparative vasculargrowth.139 Moreover, semaphorin 3E–deficient mice (or overexpres-sion of the decoy PlexinD1-Fc) showed decidedly increasedvascular regeneration and blood flow in response to ischemicinjury.139 In the retina, we demonstrated that specifically inhibitingsemaphorin 3A in RGCs using shRNAs delivered by lentiviralvectors resulted in enhanced physiologic revascularization ofischemic pockets.71 This approach directly lead to increased visualfunction as determined by electroretinograms.
In retinopathy, the position of the source of the vasorepulsiveguidance cues such as the semaphorins (RGCs beneath the vascularplexus) is speculated to repel vessels toward the vitreous. Hence,the administration of semaphorin 3E by intravitreal injection(which stimulates vessels from an extraretinal position) suppresses
preretinal vessels while preserving regeneration of normal ves-sels.70 The normalized growth is thought to occur through directstimulation of plexinD1 in pathologic retinal vessels. A similaroutcome was noted with ectopic administration of semaphorin3A.71 Both approaches showed a trend toward accelerated retinalrevascularization, although the principal noted benefit was suppres-sion of pathologic preretinal growth.
Conclusion
In the last decade, we have witnessed a surge in our understandingof the molecular mechanisms governing blood vessel50 and nerve140
growth. It is now clear that the chemotactic signals present duringembryogenesis are conserved and shared between neurons andvessels.1,64,119,141,142 Although neurovascular cross-talk shapes vas-cular development, it has received limited attention as an etiologyof disease. The tight physiologic coupling between vessels andneurons in the healthy retina suggests that therapeutic insight maybe gained through a better understanding of how the unit reactsunder stress. Notably, the identification of neuron-derived proangio-genic energy metabolites such as succinate and vasomodulatorychemotactic cues such as the semaphorins provides immediatemechanisms to modulate for reparative angiogenesis. Currently,approaches to counter aberrant ocular neovascularization (bothapproved and in human trials) rely largely on antiangiogenicstrategies. However, despite all invested efforts, effective inhibitionof pathologic ocular neovascularization remains to be achieved.Future therapies for ischemic retinal disease may therefore need togo beyond solely alleviating the angiogenic burden and incorporatestrategies to promote therapeutic angiogenesis with the aim ofreducing ischemic stress. In this respect, studying the neuronalprograms that drive retinal angiogenesis may provide attractiveavenues of research.
Acknowledgments
P.S. holds a Canada Research Chair in Retinal Cell Biology and issupported by the Canadian Institutes of Health Research (221478),the Canadian Diabetes Association (OG-3-11-3329-PS), the Natu-ral Sciences and Engineering Research Council of Canada (418637)and the Alcon Research Institute New Investigator Award.
Authorship
Contribution: P.S. conceived, assembled, and wrote the manuscript.Conflict-of-interest disclosure: The author declares no compet-
ing financial interests.Correspondence: Przemyslaw (Mike) Sapieha, PhD,
Maisonneuve-Rosemont Hospital Research Centre, 5415 Assomp-tion Boulevard, Montreal, QC, H1T 2M4, Canada; e-mail:[email protected].
References
1. Carmeliet P, Tessier-Lavigne M. Common mecha-nisms of nerve and blood vessel wiring. Nature.2005;436(7048):193-200.
2. Quaegebeur A, Lange C, Carmeliet P. The neuro-vascular link in health and disease: molecularmechanisms and therapeutic implications. Neu-ron. 2011;71(3):406-424.
3. Yamamoto Y, Craggs L, Baumann M, Kalimo H,Kalaria RN. Review: molecular genetics and pathol-ogy of hereditary small vessel diseases of the brain.Neuropathol Appl Neurobiol. 2011;37(1):94-113.
4. Brun A, Englund E. A white matter disorder in de-mentia of the Alzheimer type: a pathoanatomicalstudy. Ann Neurol. 1986;19(3):253-262.
5. Prat A, Biernacki K, Lavoie JF, Poirier J,Duquette P, Antel JP. Migration of multiple sclero-sis lymphocytes through brain endothelium. ArchNeurol. 2002;59(3):391-397.
6. Rule RR, Schuff N, Miller RG, Weiner MW. Graymatter perfusion correlates with disease severityin ALS. Neurology. 2010;74(10):821-827.
NEURONAL INFLUENCE IN OCULAR VASCULARIZATION 2191BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom

7. Lo EH. A new penumbra: transitioning from injury intorepair after stroke. Nat Med. 2008;14(5):497-500.
8. Moskowitz MA, Lo EH, Iadecola C. The scienceof stroke: mechanisms in search of treatments.Neuron. 2010;67(2):181-198.
9. Sapieha P, Hamel D, Shao Z, et al. Proliferativeretinopathies: angiogenesis that blinds. IntJ Biochem Cell Biol. 2010;42(1):5-12.
10. Sapieha P, Joyal JS, Rivera JC, et al. Retinopa-thy of prematurity: understanding ischemic retinalvasculopathies at an extreme of life. J Clin Invest.2010;120(9):3022-3032.
11. Chopp M, Zhang ZG, Jiang Q. Neurogenesis,angiogenesis, and MRI indices of functional re-covery from stroke. Stroke. 2007;38(2 Suppl):827-831.
12. Li L, Jiang Q, Zhang L, et al. Angiogenesis andimproved cerebral blood flow in the ischemicboundary area detected by MRI after administra-tion of sildenafil to rats with embolic stroke. BrainRes. 2007;1132(1):185-192.
13. Folkman J. Angiogenesis in cancer, vascular,rheumatoid and other disease. Nat Med. 1995;1(1):27-31.
14. Ramon y Cajal S. Degeneration and Regenera-tion of the Nervous System. London, United King-dom: Oxford University Press; 1928.
15. Stahl A, Connor KM, Sapieha P, et al. The mouseretina as an angiogenesis model. Invest Ophthal-mol Vis Sci. 2010;51(6):2813-2826.
16. Dorrell M, Friedlander M. Mechanisms of endo-thelial cell guidance and vascular patterning inthe developing mouse retina. Prog Retin EyeRes. 2006;25(3):277-295.
17. Pitulescu ME, Schmidt I, Benedito R, Adams RH.Inducible gene targeting in the neonatal vascula-ture and analysis of retinal angiogenesis in mice.Nat Protoc. 2010;5(9):1518-1534.
18. Hua J, Guerin KI, Chen J, et al. Resveratrol inhib-its pathologic retinal neovascularization inVldlr(�/�) mice. Invest Ophthalmol Vis Sci. 2011;52(5):2809-2816.
19. Sapieha P, Sirinyan M, Hamel D, et al. The succi-nate receptor GPR91 in neurons has a major rolein retinal angiogenesis. Nat Med. 2008;14(10):1067-1076.
20. Zaniolo K, Sapieha P, Shao Z, et al. Ghrelinmodulates physiologic and pathologic retinal an-giogenesis through GHSR-1a. Invest OphthalmolVis Sci. 2011;52(8):5376-5386.
21. Gerhardt H, Golding M, Fruttiger M, et al. VEGFguides angiogenic sprouting utilizing endothelialtip cell filopodia. J Cell Biol. 2003;161(6):1163-1177.
22. Stalmans I, Ng YS, Rohan R, et al. Arteriolar andvenular patterning in retinas of mice selectivelyexpressing VEGF isoforms. J Clin Invest. 2002;109(3):327-336.
23. Hellstrom M, Phng LK, Hofmann JJ, et al. Dll4signalling through Notch1 regulates formation oftip cells during angiogenesis. Nature. 2007;445(7129):776-780.
24. Benedito R, Roca C, Sorensen I, et al. The notchligands Dll4 and Jagged1 have opposing effectson angiogenesis. Cell. 2009;137(6):1124-1135.
25. Suchting S, Freitas C, le Noble F, et al. The Notchligand Delta-like 4 negatively regulates endothe-lial tip cell formation and vessel branching. ProcNatl Acad Sci U S A. 2007;104(9):3225-3230.
26. Fantin A, Vieira JM, Gestri G, et al. Tissue macro-phages act as cellular chaperones for vascularanastomosis downstream of VEGF-mediated en-dothelial tip cell induction. Blood. 2010;116(5):829-840.
27. Stefater Iii JA, Lewkowich I, Rao S, et al. Regula-tion of angiogenesis by a non-canonical Wnt-Flt1pathway in myeloid cells. Nature. 2011;474(7352):511-515.
28. Lobov IB, Rao S, Carroll TJ, et al. WNT7b medi-ates macrophage-induced programmed cell
death in patterning of the vasculature. Nature.2005;437(7057):417-421.
29. Chase J. The evolution of retinal vascularizationin mammals. A comparison of vascular and avas-cular retinae. Ophthalmology. 1982;89(12):1518-1525.
30. Buttery RG, Hinrichsen CF, Weller WL,Haight JR. How thick should a retina be? A com-parative study of mammalian species with andwithout intraretinal vasculature. Vision Res. 1991;31(2):169-187.
31. Dreher Z, Robinson SR, Distler C. Muller cells invascular and avascular retinae: a survey of sevenmammals. J Comp Neurol. 1992;323(1):59-80.
32. Gariano RF, Gardner TW. Retinal angiogenesis indevelopment and disease. Nature. 2005;438(7070):960-966.
33. Fruttiger M. Development of the mouse retinalvasculature: angiogenesis versus vasculogen-esis. Invest Ophthalmol Vis Sci. 2002;43(2):522-527.
34. Dorrell MI, Aguilar E, Friedlander M. Retinal vas-cular development is mediated by endothelialfilopodia, a preexisting astrocytic template andspecific R-cadherin adhesion. Invest OphthalmolVis Sci. 2002;43(11):3500-3510.
35. Fruttiger M, Calver AR, Kruger WH, et al. PDGFmediates a neuron-astrocyte interaction in thedeveloping retina. Neuron. 1996;17(6):1117-1131.
36. Stone J, Itin A, Alon T, et al. Development of reti-nal vasculature is mediated by hypoxia-inducedvascular endothelial growth factor (VEGF) ex-pression by neuroglia. J Neurosci. 1995;15(7 Pt1):4738-4747.
37. Weidemann A, Krohne TU, Aguilar E, et al. Astro-cyte hypoxic response is essential for pathologi-cal but not developmental angiogenesis of theretina. Glia. 2010;58(10):1177-1185.
38. Scott A, Powner MB, Gandhi P, et al. Astrocyte-derived vascular endothelial growth factor stabi-lizes vessels in the developing retinal vascula-ture. PLoS One. 2010;5(7):e11863.
39. Flynn JT, Bancalari E, Snyder ES, et al. A cohortstudy of transcutaneous oxygen tension and theincidence and severity of retinopathy of prematu-rity. N Engl J Med. 1992;326(16):1050-1054.
40. Wang GL, Jiang BH, Rue EA, Semenza GL.Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2tension. Proc Natl Acad Sci U S A. 1995;92(12):5510-5514.
41. Yu AY, Frid MG, Shimoda LA, Wiener CM,Stenmark K, Semenza GL. Temporal, spatial, andoxygen-regulated expression of hypoxia-induciblefactor-1 in the lung. Am J Physiol. 1998;275(4 Pt 1):L818-826.
42. Chapman-Smith A, Whitelaw ML. Novel DNAbinding by a basic helix-loop-helix protein. Therole of the dioxin receptor PAS domain. J BiolChem. 2006;281(18):12535-12545.
43. Liu Y, Cox SR, Morita T, Kourembanas S. Hyp-oxia regulates vascular endothelial growth factorgene expression in endothelial cells. Identificationof a 5 enhancer. Circ Res. 1995;77(3):638-643.
44. Wang GL, Semenza GL. General involvement ofhypoxia-inducible factor 1 in transcriptional re-sponse to hypoxia. Proc Natl Acad Sci U S A.1993;90(9):4304-4308.
45. Bruick RK. Oxygen sensing in the hypoxic re-sponse pathway: regulation of the hypoxia-inducible transcription factor. Genes Dev. 2003;17(21):2614-2623.
46. Paul SA, Simons JW, Mabjeesh NJ. HIF at thecrossroads between ischemia and carcinogen-esis. J Cell Physiol. 2004;200(1):20-30.
47. Nakamura-Ishizu A, Kurihara T, Okuno Y, et al.The formation of an angiogenic astrocyte tem-plate is regulated by the neuroretina in a HIF-1–
dependent manner. Dev Biol. 2012;363(1):106-114.
48. Caprara C, Thiersch M, Lange C, Joly S,Samardzija M, Grimm C. HIF1A is essential forthe development of the intermediate plexus of theretinal vasculature. Invest Ophthalmol Vis Sci.2011;52(5):2109-2117.
49. Kandel ER, Siegelbaum SA. In: Kandel ER,Schwartz J, Jessell TM, eds. Principals of NeuralScience. New York, NY: McGraw-Hill; 2000:280-297.
50. Carmeliet P, Jain RK. Molecular mechanisms andclinical applications of angiogenesis. Nature.2011;473(7347):298-307.
51. Provis JM, Leech J, Diaz CM, Penfold PL,Stone J, Keshet E. Development of the humanretinal vasculature: cellular relations and VEGFexpression. Exp Eye Res. 1997;65(4):555-568.
52. Benedito R, Rocha SF, Woeste M, et al. Notch-dependent VEGFR3 upregulation allows angio-genesis without VEGF-VEGFR2 signalling.Nature. 2012;484(7392):110-114.
53. Lobov IB, Renard RA, Papadopoulos N, et al.Delta-like ligand 4 (Dll4) is induced by VEGF as anegative regulator of angiogenic sprouting. ProcNatl Acad Sci U S A. 2007;104(9):3219-3224.
54. Iruela-Arispe ML, Davis GE. Cellular and molecu-lar mechanisms of vascular lumen formation. DevCell. 2009;16(2):222-231.
55. Dejana E, Tournier-Lasserve E, Weinstein BM.The control of vascular integrity by endothelialcell junctions: molecular basis and pathologicalimplications. Dev Cell. 2009;16(2):209-221.
56. Ishida S, Yamashiro K, Usui T, et al. Leukocytesmediate retinal vascular remodeling during devel-opment and vaso-obliteration in disease. NatMed. 2003;9(6):781-788.
57. Das A, McGuire PG. Retinal and choroidal angio-genesis: pathophysiology and strategies for inhi-bition. Prog Retin Eye Res. 2003;22(6):721-748.
58. Tammela T, Zarkada G, Nurmi H, et al. VEGFR-3controls tip to stalk conversion at vessel fusionsites by reinforcing Notch signalling. Nat Cell Biol.2011;13(10):1202-1213.
59. De Smet F, Segura I, De Bock K,Hohensinner PJ, Carmeliet P. Mechanisms ofvessel branching: filopodia on endothelial tip cellslead the way. Arterioscler Thromb Vasc Biol.2009;29(5):639-649.
60. Huber AB, Kolodkin AL, Ginty DD, Cloutier JF.Signaling at the growth cone: ligand-receptorcomplexes and the control of axon growth andguidance. Annu Rev Neurosci. 2003;26:509-563.
61. Adams RH, Wilkinson GA, Weiss C, et al. Rolesof ephrinB ligands and EphB receptors in cardio-vascular development: demarcation of arterial/venous domains, vascular morphogenesis, andsprouting angiogenesis. Genes Dev. 1999;13(3):295-306.
62. Klagsbrun M, Eichmann A. A role for axon guid-ance receptors and ligands in blood vessel devel-opment and tumor angiogenesis. CytokineGrowth Factor Rev. 2005;16(4-5):535-548.
63. Wilson BD, Ii M, Park KW, et al. Netrins promotedevelopmental and therapeutic angiogenesis.Science. 2006;313(5787):640-644.
64. Larrivee B, Freitas C, Suchting S, Brunet I,Eichmann A. Guidance of vascular development:lessons from the nervous system. Circ Res.2009;104(4):428-441.
65. Gelfand MV, Hong S, Gu C. Guidance fromabove: common cues direct distinct signaling out-comes in vascular and neural patterning. TrendsCell Biol. 2009;19(3):99-110.
66. Carmichael ST. Cellular and molecular mecha-nisms of neural repair after stroke: makingwaves. Ann Neurol. 2006;59(5):735-742.
67. Carmichael ST, Archibeque I, Luke L, Nolan T,Momiy J, Li S. Growth-associated gene expres-sion after stroke: evidence for a growth-promoting
2192 SAPIEHA BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom

region in peri-infarct cortex. Exp Neurol. 2005;193(2):291-311.
68. de Winter F, Cui Q, Symons N, Verhaagen J,Harvey AR. Expression of class-3 semaphorinsand their receptors in the neonatal and adult ratretina. Invest Ophthalmol Vis Sci. 2004;45(12):4554-4562.
69. De Winter F, Oudega M, Lankhorst AJ, et al.Injury-induced class 3 semaphorin expression inthe rat spinal cord. Exp Neurol. 2002;175(1):61-75.
70. Fukushima Y, Okada M, Kataoka H, et al.Sema3E-PlexinD1 signaling selectively sup-presses disoriented angiogenesis in ischemicretinopathy in mice. J Clin Invest. 2011;121(5):1974-1985.
71. Joyal J-S, Sitaras N, Binet F, et al. Ischemic neu-rons prevent vascular regeneration of neural tis-sue by secreting semaphorin 3A. Blood. 2011;117(22):6024-6035.
72. Edwards MM, McLeod DS, Li R, et al. The dele-tion of Math5 disrupts retinal blood vessel andglial development in mice. Exp Eye Res. 2012;96(1):147-156.
73. Kim J, Oh WJ, Gaiano N, Yoshida Y, Gu C.Semaphorin 3E-Plexin-D1 signaling regulatesVEGF function in developmental angiogenesisvia a feedback mechanism. Genes Dev. 2011;25(13):1399-1411.
74. Hughes S, Yang H, Chan-Ling T. Vascularizationof the human fetal retina: roles of vasculogenesisand angiogenesis. Invest Ophthalmol Vis Sci.2000;41(5):1217-1228.
75. Dreher B, Robinson SR. Development of the reti-nofugal pathway in birds and mammals: evidencefor a common ‘timetable.’ Brain Behav Evol.1988;31(6):369-390.
76. Cringle SJ, Yu PK, Su EN, Yu DY. Oxygen distri-bution and consumption in the developing ratretina. Invest Ophthalmol Vis Sci. 2006;47(9):4072-4076.
77. Uemura A, Kusuhara S, Wiegand SJ, Yu RT,Nishikawa S. Tlx acts as a proangiogenic switchby regulating extracellular assembly of fibronectinmatrices in retinal astrocytes. J Clin Invest. 2006;116(2):369-377.
78. Mu X, Fu X, Sun H, et al. Ganglion cells are re-quired for normal progenitor-cell proliferation butnot cell-fate determination or patterning in thedeveloping mouse retina. Curr Biol. 2005;15(6):525-530.
79. Wangsa-Wirawan ND, Linsenmeier RA. Retinaloxygen: fundamental and clinical aspects. ArchOphthalmol. 2003;121(4):547-557.
80. Sternberg P Jr, Landers MB 3rd, Wolbarsht M.The negative coincidence of retinitis pigmentosaand proliferative diabetic retinopathy. Am J Oph-thalmol. 1984;97(6):788-789.
81. Lahdenranta J, Pasqualini R, Schlingemann RO,et al. An anti-angiogenic state in mice and hu-mans with retinal photoreceptor cell degenera-tion. Proc Natl Acad Sci U S A. 2001;98(18):10368-10373.
82. de Gooyer TE, Stevenson KA, Humphries P,Simpson DA, Gardiner TA, Stitt AW. Retinopathyis reduced during experimental diabetes in amouse model of outer retinal degeneration. InvestOphthalmol Vis Sci. 2006;47(12):5561-5568.
83. Linsenmeier RA, Padnick-Silver L. Metabolic de-pendence of photoreceptors on the choroid in thenormal and detached retina. Invest OphthalmolVis Sci. 2000;41(10):3117-3123.
84. Grant MB, Davis MI, Caballero S, Feoktistov I,Biaggioni I, Belardinelli L. Proliferation, migration,and ERK activation in human retinal endothelialcells through A(2B) adenosine receptor stimula-tion. Invest Ophthalmol Vis Sci. 2001;42(9):2068-2073.
85. Grant MB, Tarnuzzer RW, Caballero S, et al.Adenosine receptor activation induces vascular
endothelial growth factor in human retinal endo-thelial cells. Circ Res. 1999;85(8):699-706.
86. Mino RP, Spoerri PE, Caballero S, et al. Adeno-sine receptor antagonists and retinal neovascu-larization in vivo. Invest Ophthalmol Vis Sci.2001;42(13):3320-3324.
87. He W, Miao FJ, Lin DC, et al. Citric acid cycle in-termediates as ligands for orphan G-protein-coupled receptors. Nature. 2004;429(6988):188-193.
88. Gutman M, Bonomi F, Pagani S, Cerletti P,Kroneck P. Modulation of the flavin redox poten-tial as mode of regulation of succinate dehydro-genase activity. Biochim Biophys Acta. 1980;591(2):400-408.
89. Meixner-Monori B, Kubicek CP, Habison A,Kubicek-Pranz EM, Rohr M. Presence and regu-lation of the alpha-ketoglutarate dehydrogenasemultienzyme complex in the filamentous fungusAspergillus niger. J Bacteriol. 1985;161(1):265-271.
90. Gilbert C, Rahi J, Eckstein M, O’Sullivan J,Foster A. Retinopathy of prematurity in middle-income countries. Lancet. 1997;350(9070):12-14.
91. Kempen JH, O’Colmain BJ, Leske MC, et al. Theprevalence of diabetic retinopathy among adultsin the United States. Arch Ophthalmol. 2004;122(4):552-563.
92. Chen J, Smith L. Retinopathy of prematurity. An-giogenesis. 2007;10(2):133-140.
93. Cheung N. Diabetic retinopathy and systemicvascular complications. Progr Retin Eye Res.2008;27(2):161-176.
94. Smith LE. Through the eyes of a child: under-standing retinopathy through ROP the Frieden-wald lecture. Invest Ophthalmol Vis Sci. 2008;49(12):5177-5182.
95. Weinberger B, Watorek K, Strauss R, Witz G,Hiatt M, Hegyi T. Association of lipid peroxidationwith hepatocellular injury in preterm infants. CritCare. 2002;6(6):521-525.
96. University of Iowa, Children’s Hospital, Depart-ment of Pediatrics. Comments on oxygen toxicityand retinopathy (ROP) in the premature infant.1994. http://www.uihealthcare.com/depts/med/pediatrics/iowaneonatologyhandbook/general/commentsoxygen.html. Accessed June 18, 2012.
97. Hardy P, Beauchamp M, Sennlaub F, et al. Newinsights into the retinal circulation: inflammatorylipid mediators in ischemic retinopathy. Prosta-glandins Leukot Essent Fatty Acids. 2005;72(5):301-325.
98. Roth AM. Retinal vascular development in prema-ture infants. Am J Ophthalmol. 1977;84(5):636-640.
99. Smith LE, Shen W, Perruzzi C, et al. Regulationof vascular endothelial growth factor-dependentretinal neovascularization by insulin-like growthfactor-1 receptor. Nat Med. 1999;5(12):1390-1395.
100. Brownlee M, Vlassara H, Cerami A. Nonenzy-matic glycosylation and the pathogenesis of dia-betic complications. Ann Intern Med. 1984;101(4):527-537.
101. Ways DK, Sheetz MJ. The role of protein kinaseC in the development of the complications of dia-betes. Vitam Horm. 2000;60:149-193.
102. The effect of intensive diabetes treatment on theprogression of diabetic retinopathy in insulin-dependent diabetes mellitus. The Diabetes Con-trol and Complications Trial. Arch Ophthalmol.1995;113(1):36-51.
103. Kowluru RA, Odenbach S. Role of interleukin-1beta in the pathogenesis of diabetic retinopathy.Br J Ophthalmol. 2004;88(10):1343-1347.
104. Joussen AM, Poulaki V, Mitsiades N, et al. Non-steroidal anti-inflammatory drugs prevent earlydiabetic retinopathy via TNF-alpha suppression.FASEB J. 2002;16(3):438-440.
105. Adamis AP, Berman AJ. Immunological mecha-
nisms in the pathogenesis of diabetic retinopathy.Semin Immunopathol. 2008;30(2):65-84.
106. Dorrell M, Uusitalo-Jarvinen H, Aguilar E,Friedlander M. Ocular neovascularization: basicmechanisms and therapeutic advances. SurvOphthalmol. 2007;52(Suppl 1):S3-S19.
107. Dorfman A, Dembinska O, Chemtob S,Lachapelle P. Early manifestations of postnatalhyperoxia on the retinal structure and function ofthe neonatal rat. Invest Ophthalmol Vis Sci. 2008;49(1):458-466.
108. Dorfman AL, Joly S, Hardy P, Chemtob S,Lachapelle P. The effect of oxygen and light onthe structure and function of the neonatal ratretina. Doc Ophthalmol. 2009;118(1):37-54.
109. Dorfman AL, Polosa A, Joly S, Chemtob S,Lachapelle P. Functional and structural changesresulting from strain differences in the rat modelof oxygen-induced retinopathy. Invest OphthalmolVis Sci. 2009;50(5):2436-2450.
110. Barber AJ, Antonetti DA, Kern TS, et al. TheIns2Akita mouse as a model of early retinal com-plications in diabetes. Invest Ophthalmol Vis Sci.2005;46(6):2210-2218.
111. Barber AJ, Lieth E, Khin SA, Antonetti DA,Buchanan AG, Gardner TW. Neural apoptosis inthe retina during experimental and human diabe-tes. Early onset and effect of insulin. J Clin Invest.1998;102(4):783-791.
112. Gastinger MJ, Kunselman AR, Conboy EE,Bronson SK, Barber AJ. Dendrite remodeling andother abnormalities in the retinal ganglion cells ofIns2 Akita diabetic mice. Invest Ophthalmol VisSci. 2008;49(6):2635-2642.
113. Luo Y, Raible D, Raper JA. Collapsin: a protein inbrain that induces the collapse and paralysis ofneuronal growth cones. Cell. 1993;75(2):217-227.
114. Geretti E, Shimizu A, Klagsbrun M. Neuropilinstructure governs VEGF and semaphorin bindingand regulates angiogenesis. Angiogenesis. 2008;11(1):31-39.
115. Takahashi T, Fournier A, Nakamura F, et al.Plexin-neuropilin-1 complexes form functionalsemaphorin-3A receptors. Cell. 1999;99(1):59-69.
116. Lee P, Goishi K, Davidson AJ, Mannix R, Zon L,Klagsbrun M. Neuropilin-1 is required for vasculardevelopment and is a mediator of VEGF-dependent angiogenesis in zebrafish. Proc NatlAcad Sci U S A. 2002;99(16):10470-10475.
117. Gluzman-Poltorak Z, Cohen T, Shibuya M,Neufeld G. Vascular endothelial growth factorreceptor-1 and neuropilin-2 form complexes.J Biol Chem. 2001;276(22):18688-18694.
118. Mamluk R, Gechtman Z, Kutcher ME,Gasiunas N, Gallagher J, Klagsbrun M.Neuropilin-1 binds vascular endothelial growthfactor 165, placenta growth factor-2, and heparinvia its b1b2 domain. J Biol Chem. 2002;277(27):24818-24825.
119. Miao HQ, Soker S, Feiner L, Alonso JL,Raper JA, Klagsbrun M. Neuropilin-1 mediatescollapsin-1/semaphorin III inhibition of endothelialcell motility: functional competition of collapsin-1and vascular endothelial growth factor-165. J CellBiol. 1999;146(1):233-242.
120. Klagsbrun M, Takashima S, Mamluk R. The roleof neuropilin in vascular and tumor biology. AdvExp Med Biol. 2002;515:33-48.
121. Soker S, Miao HQ, Nomi M, Takashima S,Klagsbrun M. VEGF165 mediates formation ofcomplexes containing VEGFR-2 and neuropilin-1that enhance VEGF165-receptor binding. J CellBiochem. 2002;85(2):357-368.
122. Appleton BA, Wu P, Maloney J, et al. Structuralstudies of neuropilin/antibody complexes provideinsights into semaphorin and VEGF binding.EMBO J. 2007;26(23):4902-4912.
123. Vieira JM, Schwarz Q, Ruhrberg C. Role of the
NEURONAL INFLUENCE IN OCULAR VASCULARIZATION 2193BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom

neuropilin ligands VEGF164 and SEMA3A in neu-ronal and vascular patterning in the mouse. No-vartis Found Symp. 2007;283:230-235; discus-sion 235-241.
124. Selak MA, Armour SM, MacKenzie ED, et al.Succinate links TCA cycle dysfunction to onco-genesis by inhibiting HIF-alpha prolyl hydroxy-lase. Cancer Cell. 2005;7(1):77-85.
125. Koivunen P, Hirsila M, Remes AM, Hassinen IE,Kivirikko KI, Myllyharju J. Inhibition of hypoxia-inducible factor (HIF) hydroxylases by citric acidcycle intermediates: possible links between cellmetabolism and stabilization of HIF. J Biol Chem.2007;282(7):4524-4532.
126. Arany Z, Foo SY, Ma Y, et al. HIF-independentregulation of VEGF and angiogenesis by the tran-scriptional coactivator PGC-1alpha. Nature.2008;451(7181):1008-1012.
127. Carmeliet P. Cardiovascular biology. Creatingunique blood vessels. Nature. 2001;412(6850):868-869.
128. Fujita H, Zhang B, Sato K, Tanaka J, Sakanaka M.Expressions of neuropilin-1, neuropilin-2 andsemaphorin 3A mRNA in the rat brain after middlecerebral artery occlusion. Brain Res. 2001;914(1-2):1-14.
129. Caldwell RB, Bartoli M, Behzadian MA, et al. Vas-cular endothelial growth factor and diabetic reti-
nopathy: pathophysiological mechanisms andtreatment perspectives. Diabetes Metab Res Rev.2003;19(6):442-455.
130. Aiello LP, Pierce EA, Foley ED, et al. Suppressionof retinal neovascularization in vivo by inhibitionof vascular endothelial growth factor (VEGF) us-ing soluble VEGF-receptor chimeric proteins.Proc Natl Acad Sci U S A. 1995;92(23):10457-10461.
131. Ferrara N, Kerbel RS. Angiogenesis as a thera-peutic target. Nature. 2005;438(7070):967-974.
132. Stewart MW. The expanding role of vascular en-dothelial growth factor inhibitors in ophthalmol-ogy. Mayo Clin Proc. 2012;87(1):77-88.
133. Holz FG, Amoaku W, Donate J, et al. Safety andefficacy of a flexible dosing regimen of ranibi-zumab in neovascular age-related macular de-generation: the SUSTAIN study. Ophthalmology.2011;118(4):663-671.
134. Robinson GS, Ju M, Shih SC, et al. Nonvascularrole for VEGF: VEGFR-1, 2 activity is critical forneural retinal development. FASEB J. 2001;15(7):1215-1217.
135. Gillies MC, Wong TY. Ranibizumab for neovascu-lar age-related macular degeneration. N EnglJ Med. 2007;356(7):748-749; author reply 749-750.
136. Ritter MR, Banin E, Moreno SK, Aguilar E,
Dorrell MI, Friedlander M. Myeloid progenitorsdifferentiate into microglia and promote vascularrepair in a model of ischemic retinopathy. J ClinInvest. 2006;116(12):3266-3276.
137. Dorrell MI, Aguilar E, Jacobson R, et al. Maintain-ing retinal astrocytes normalizes revasculariza-tion and prevents vascular pathology associatedwith oxygen-induced retinopathy. Glia. 2010;58(1):43-54.
138. Connor KM, SanGiovanni JP, Lofqvist C, et al.Increased dietary intake of omega-3-polyunsaturatedfatty acids reduces pathological retinal angiogen-esis. Nat Med. 2007;13(7):868-873.
139. Moriya J, Minamino T, Tateno K, et al. Inhibition ofsemaphorin as a novel strategy for therapeuticangiogenesis. Circ Res. 2010;106(2):391-398.
140. Raper J, Mason C. Cellular strategies of axonalpathfinding. Cold Spring Harb Perspect Biol.2010;2(9):a001933.
141. Erskine L, Reijntjes S, Pratt T, et al. VEGF signal-ing through neuropilin 1 guides commissural axoncrossing at the optic chiasm. Neuron. 2011;70(5):951-965.
142. Soker S, Takashima S, Miao HQ, Neufeld G,Klagsbrun M. Neuropilin-1 is expressed by endo-thelial and tumor cells as an isoform-specific re-ceptor for vascular endothelial growth factor. Cell.1998;92(6):735-745.
2194 SAPIEHA BLOOD, 13 SEPTEMBER 2012 � VOLUME 120, NUMBER 11
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom

online June 15, 2012 originally publisheddoi:10.1182/blood-2012-04-396846
2012 120: 2182-2194
Przemyslaw Sapieha Eyeing central neurons in vascular growth and reparative angiogenesis
http://www.bloodjournal.org/content/120/11/2182.full.htmlUpdated information and services can be found at:
(519 articles)Vascular Biology (800 articles)Review Articles
Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American Society
For personal use only.on December 5, 2018. by guest www.bloodjournal.orgFrom