expression of the c-erbb-4/her4 protein and mrna in normal human fetal and adult tissues and in a...
TRANSCRIPT

, . 185: 236–245 (1998)
EXPRESSION OF THE c-erbB-4/HER4 PROTEIN ANDmRNA IN NORMAL HUMAN FETAL AND ADULT
TISSUES AND IN A SURVEY OF NINE SOLIDTUMOUR TYPES
1, 2, . 3 . 1*1Receptor Biology Laboratory, Imperial Cancer Research Fund Molecular Oncology Unit at the Imperial College School of
Medicine, Hammersmith Hospital, Du Cane Road, London W12 0NN, U.K.2In situ Hybridization Service, Imperial Cancer Research Fund Laboratories, 44 Lincoln’s Inn Fields, London WC2A 3PX, U.K.
3Gene Transcription Laboratory, Imperial Cancer Research Fund Molecular Oncology Unit at the Imperial College School ofMedicine, Hammersmith Hospital, Du Cane Road, London W12 0NN, U.K.
SUMMARY
The c-erbB-4/HER4 receptor belongs to the family of the type I growth factor receptors. Mouse monoclonal antibodies have beenraised to the cytoplasmic domain of the c-erbB-4 receptor and characterized; the antibody HFR-1 has been used to determine the patternof expression of the c-erbB-4 protein immunohistochemically in formalin-fixed, paraffin-embedded adult and fetal tissues. The expressionof c-erbB-4 mRNA was determined by using 35S-labelled riboprobes and tissue in situ hybridization. c-erbB-4 is widely expressed inmany adult and fetal tissues, including the lining epithelia of the gastrointestinal, urinary, reproductive, and respiratory tracts, as wellas the skin, skeletal muscle, circulatory, endocrine, and nervous systems. The developing brain and heart notably express high levels ofthis receptor. The pattern of c-erbB-4 protein expression is also reported in a survey of common solid human cancers. Loss of expressionwas noted in 40–80 per cent of adenocarcinomas and up to 100 per cent of squamous cell carcinomas, whereas overexpression wasobserved in about 10–20 per cent of adenocarcinomas and astrocytomas. In general, the pattern of c-erbB-4 expression in normal tissuesand cancers suggests that it tends to be associated with the differentiated compartment. ? 1998 John Wiley & Sons, Ltd.
J. Pathol. 185: 236–245, 1998.
KEY WORDS—c-erbB-4; HER4; growth factor receptor; tyrosine kinase; human cancer
INTRODUCTION
Growth factor receptors with tyrosine kinase activityregulate fundamental cell behaviour including cell sur-vival, cell differentiation, motility, and cell proliferation.The type I growth factor receptor family consists of theEGFR (epidermal growth factor receptor) and c-erbB-2,c-erbB-3, and c-erbB-4 (also known as HER2, 3, and 4),respectively. c-erbB-4, the fourth and most recentlydescribed member of the family, was cloned in 19931 andthe gene localized to chromosome 2q33.3–34.2 Itencodes a protein of 1283 amino acids which is post-translationally glycosylated to give a molecular weightof 180 kD.1 The protein consists of the typical extracel-lular ligand binding domain, a transmembrane region,and an intracytoplasmic domain that has ligand-activatable tyrosine kinase activity.1 The extracellulardomain of c-erbB-4 is most similar to c-erbB-3, whereasits cytoplasmic domain exhibits 79 and 77 per centidentity with EGFR and c-erbB-2, respectively.1
EGFR, c-erbB-2, and c-erbB-3 are widely expressed inepithelial, mesenchymal, and neuronal tissue, indicatingtheir importance in several cellular functions.3 Further-
*Correspondence to: William J. Gullick, Imperial Cancer ResearchFund, Molecular Oncology Unit, Imperial College School of Medi-cine, Hammersmith Hospital, Du Cane Road, London W12 0NN,U.K. E-mail: [email protected]
CCC 0022–3417/98/070236–10 $17.50? 1998 John Wiley & Sons, Ltd.
more, overexpression of the above receptors and theirligands occurs in many human malignancies and hasbeen correlated to tumour aggressiveness and poorprognosis.3–9 Recently, it has been shown that there isextensive interaction among members of the family inthe form of homo- and heterodimerization upon ligandbinding, leading to cross-activation of the receptors.10,11
In the case of c-erbB-4, the heregulins (also known asneuregulins or NDF),12,13 betacellulin14 and recentlyheparin-binding EGF (HB-EGF),15 have been shown tofunction as ligands by binding directly to the receptorand inducing its tyrosine phosphorylation. Downstream,the c-erbB-4 cytoplasmic domain interacts with SH-2-domain-containing proteins such as p120-ras GTPaseactivating protein, SHC, and the p85 subunit ofphosphatidylinositol-3*-kinase, which activate themitogen-activated protein kinase pathway.16,17
There have been a few reports describing the tissueexpression of the c-erbB-4 receptor protein. It has beenlocalized to the neuromuscular synapse,18 normal andneoplastic thyroid,19 normal and neoplastic prostate,20
and medulloblastoma.21 There is, however, no detailedanalysis of its normal pattern of expression, nor asystematic study of its expression in cancers. Wedescribe here the tissue distribution of the c-erbB-4receptor in normal human adult and fetal tissues and ina sample of common human cancers. c-erbB-4 was seen
Received 31 October 1997Accepted 2 March 1998

237c-erbB-4 EXPRESSION IN NORMAL TISSUES AND TUMOURS
to have a widespread distribution in both adult and fetaltissues. There was broad agreement between thec-erbB-4 protein and mRNA distribution as determinedby in situ hybridization (ISH). In cancers there was a lossof expression in most tumours, but a small proportion ofadenocarcinomas and astrocytomas showed evidence ofoverexpression, compared with normal controls.
MATERIAL AND METHODS
Production of anti-peptide antibodies to c-erbB-4 usingsynthetic peptides
The synthetic peptide RSTLQHPDYLQEYST (resi-dues 1249–1264) from the cytoplasmic domain ofc-erbB-4 was used to raise anti-peptide polyclonal anti-bodies in rabbits designated A.P.96.4 and monoclonalantibodies in mice designated HFR-1 and HFR-2. All ofthese antibodies were purified by affinity columnchromatography22 and characterized for their ability torecognize c-erbB-4 protein.
Cell culture and immunoprecipitation
The HER4-transfected NIH3T3 cell line (NIH3T3/HER4 cells, a gift of Dr B. Cohen, Bristol-MyersSquibb, Seattle, U.S.A.) was grown in Dulbecco’s modi-fied Eagle medium supplemented with 10 per cent fetalbovine serum. Cells were metabolically labelled for 16 hwith [35S]methionine and immunoprecipitated as pre-viously described23 using 3 ìg ml"1 each of the anti-bodies. In each case, the antibodies were pre-incubatedin parallel with 10 ìg ml"1 of the immunizing peptide.
Western blotting
This was performed as previously described24on crudecell lysates prepared from the A431, SKBR-3, c-erbB-3-transfected 293 cells (293/HER3),25 and NIH3T3/HER4cell lines known to express high levels of EGFR,c-erbB-2, c-erbB-3, and c-erbB-4 proteins respectivelyand probed with the primary antibodies HFR-1 andHFR-2 at 3 ìg ml"1, followed by peroxidase-conjugated rabbit anti-mouse antibody (Dako Ltd,U.K.) and visualized by enhanced chemiluminescence(ECL; Amersham Int., U.K.).
Immunohistochemistry
All three antibodies tested gave a good performanceon formalin-fixed, paraffin-embedded tissues. Themouse monoclonal antibody HFR-1 was used for immu-nostaining at a concentration of 1 ìg ml"1. Cytocentri-fuge preparations of NIH3T3/HER4 cells were preparedusing a Shandon cytocentrifuge II according to themanufacturer’s instructions. Formalin-fixed, paraffin-embedded tissue blocks which included 46 adult and 32fetal tissue blocks from all three trimesters representinga panel of normal tissue were retrieved from the archivesof the Histopathology Department of the HammersmithHospital. Multiple examples of each tissue type were
? 1998 John Wiley & Sons, Ltd.
examined to control for possible variations in tissuefixation and processing. A sample of ten cases of each ofthe nine solid tumour types (90 blocks in total whichincluded carcinoma of the breast, colon, ovary, prostate,cervix, endometrium, bronchus, head and neck squa-mous cell carcinomas, and astrocytomas) were retrievedfrom the archives of the Postgraduate Institute ofMedical Education and Research, India. Thestreptavidin–biotin immunoperoxidase technique wasused and the brown reaction product was scored on asemiquantitative scale as negative ("), equivocal(+/"), weakly positive (+), moderately positive (+ +),or strongly positive (+ + +). Negative controls includedomission of the primary antibody and blocking bypre-incubation with the immunizing peptide at 10 ìgml"1. The tumours were scored as previouslydescribed,9 taking into account the percentage of posi-tive cells as well as the intensity of staining comparedwith the normal control, and were assigned to thefollowing categories: lesser than normal; equal tonormal; and greater than normal.
In situ hybridization (ISH)
A 1 kb HindIII fragment (bases "47 to 1054 ofsequence L07868, Genbank acc. No.) of the c-erbB-4cDNA (kind gift from Dr G. D. Plowman, Sugen, CA,U.S.A.) was cloned into the pGEM3Z vector (Promega,U.K.) and transcribed in vitro using SP6 polymerase(Promega) and 35S-UTP (Amersham Int.) to generateradiolabelled RNA antisense probe. In situ hybridiz-ation was performed on cytocentrifuge preparationsof A431, SKBR-3, 293/HER3, and NIH3T3/HER4 celllines fixed in 4 per cent paraformaldehyde and on apanel of normal tissues which included 12 adult and tenfetal tissues from formalin-fixed, paraffin-embeddedblocks using the protocol previously described.26,27 Aparallel section in each case was hybridized to a â-actinantisense riboprobe27 to assess the quality of mRNApreservation in the tissue.
RESULTS
Properties of antibodies to c-erbB-4
The anti-peptide polyclonal antibody raised in rabbitswas designated A.P. 96.4 and the anti-peptide mono-clonal antibodies raised in mice were designated HFR-1and HFR-2, respectively. All three antibodies couldimmunoprecipitate the 35S metabolically labelledc-erbB-4 protein from the NIH3T3/HER4 cells whichcould be abolished by pre-incubation with the cognatepeptide (Fig. 1).
The mouse monoclonal antibodies HFR-1 andHFR-2 recognized a protein of 180 kD corresponding tothe c-erbB-4 protein in its denatured and reduced formin crude cell lysates of the NIH3T3/HER4 cells (Fig. 2,tracks 2 and 4). There was no crossreactivity with EGFR(Fig. 2, tracks 1 and 3) using A-431 cell lysates, or withc-erbB-2 or c-erbB-3 using cell lysates from SKBR-3 and293/HER3 cells (data not shown).
, . 185: 236–245 (1998)

238 R. SRINIVASAN ET AL.
c-erbB-4 protein and mRNA expression as determinedby immunohistochemistry and in situ hybridization
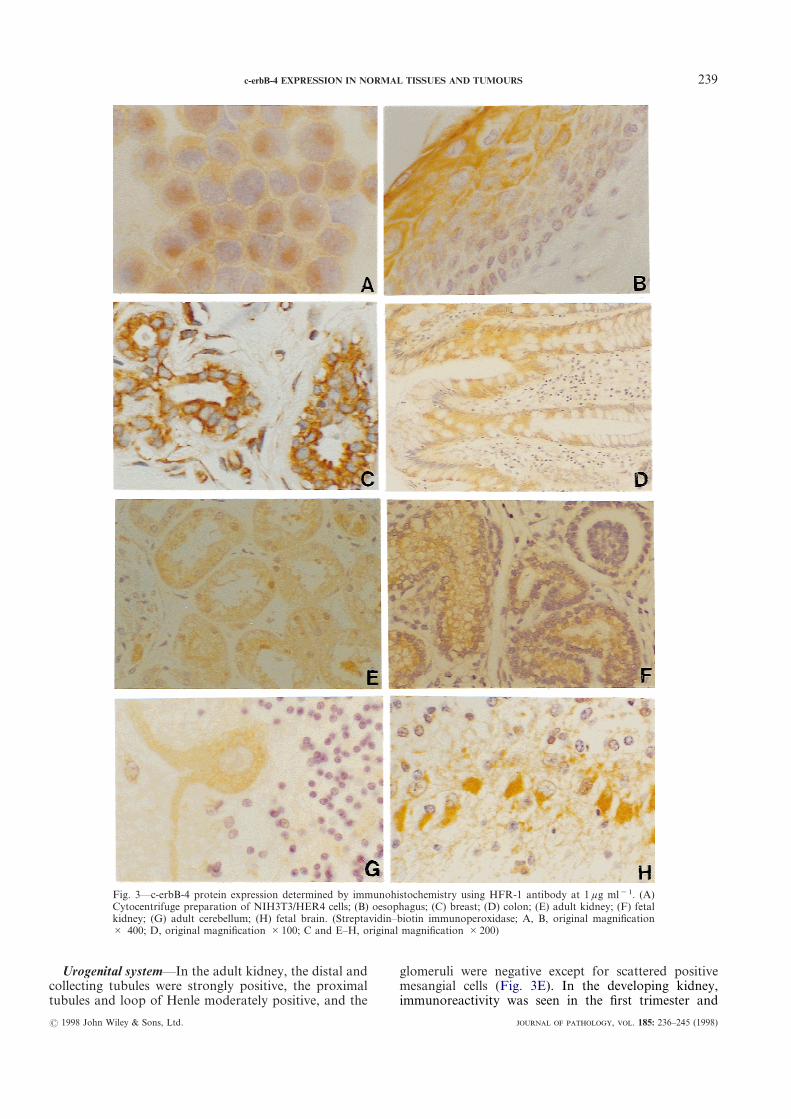
The specificity of the antibody HFR-1 to recognizethe denatured c-erbB-4 protein in cells was confirmed byimmunostaining of cytocentrifuge preparations ofNIH3T3/HER4 cells, which showed predominantlyhomogeneous cytoplasmic staining with membraneaccentuation (Fig. 3A). No immunoreactivity was seenin cytocentrifuge preparations of A431 cells. Informalin-fixed, paraffin-embedded tissues as well asfrozen sections of normal breast, the immunoreactivitywas predominantly cytoplasmic. Immunostaining couldbe blocked by competition with the cognate peptide,indicating its specificity. Occasionally, we observednuclear staining in addition to the cytoplasmic staining,as noted in the lining epithelium of the distal tubules ofthe kidney (Fig. 3E) and the ducts of the breast. Theresults of c-erbB-4 immunoreactivity in normal tissues inthis study have been tabulated and compared withsimilar studies in the literature with respect toEGFR,28,29 c-erbB-2,30 and c-erbB-3,25 respectively(Table I).
The specificity of the antisense riboprobe for c-erbB-4mRNA was also confirmed by demonstration of positivesignals by in situ hybridization on cytocentrifuge prep-arations of NIH3T3/HER4 cells and absence of anysignal in cytocentrifuge preparations of A431, SKBR-3,and 293/HER3 cell lines (data not shown).
? 1998 John Wiley & Sons, Ltd.
Overall, there was widespread expression of thec-erbB-4 receptor in various adult and developing fetaltissues, with distinctly higher levels of expression inthe differentiated compartment. The findings aresummarized as follows:
Gastrointestinal tract—The surface epithelium of thetongue, oropharynx, and oesophagus (Fig. 3B) wereimmunoreactive in their upper two-thirds. The columnarlining epithelium of the mucosa of the stomach andparietal cells were also moderately reactive. In theintestines, there was an increasing intensity of stainingtowards the surface of the villi or crypts (Fig. 3D). Thedeveloping gut showed similar reactivity. The hepato-cytes and biliary ductal epithelial cells were positive infetal and adult liver with greater intensity of staining offetal tissues. The developing pancreas also showedgreater intensity of immunoreactivity compared withadult pancreas. The pancreatic ducts were moderatelypositive and the acini were weakly positive. Tissue ISHstudies failed to reveal any c-erbB-4 mRNA in the adultstomach and intestines, despite the presence of moderatelevels of â-actin mRNA, suggesting a relatively low levelof expression in this tissue.
Fig. 1—NIH3T3/HER4 cells were metabolically labelled with [35S]me-thionine and cell lysate immunoprecipitated using 3 ìg ml"1 affinitypurified antibodies, A.P.96.4 (tracks 1, 2), HFR-2 (tracks 3, 4), andHFR-1 (tracks 5, 6). " and + indicate pre-incubation without andwith the immunizing peptide
Fig. 2—Western blotting of crude cell lysates from A431 (tracks 1, 3)and NIH3T3/HER4 (tracks 2, 4) cells with 3 ìg ml"1 affinity purifiedantibodies HFR-1 (tracks 1, 2) and HFR-2 (tracks 3, 4). Both of theantibodies specifically recognize the 180 kD protein corresponding toc-erbB-4
, . 185: 236–245 (1998)
Ucoltub
ey,
immunoreactivity was seen in the first trimester and
noes
idingin
9
? 1
rogenital system—In the adult kidney, the distal andlecting tubules were strongly positive, the proximalules and loop of Henle moderately positive, and the
Fig. 3—c-erbB-4 protein expression determined by immuCytocentrifuge preparation of NIH3T3/HER4 cells; (B) okidney; (G) adult cerebellum; (H) fetal brain. (Streptav# 400; D, original magnification #100; C and E–H, ori
998 John Wiley & Sons, Ltd.
glomeruli were negative except for scattered positivmesangial cells (Fig. 3E). In the developing kidne
histochemistry using HFR-1 antibody at 1 ìg ml"1. (A)ophagus; (C) breast; (D) colon; (E) adult kidney; (F) fetal–biotin immunoperoxidase; A, B, original magnificational magnification #200)
23c-erbB-4 EXPRESSION IN NORMAL TISSUES AND TUMOURS
, . 185: 236–245 (1998)

240 R. SRINIVASAN ET AL.
throughout development. The developing tubulesshowed moderate positivity and the undifferentiatedmesenchyme and developing glomeruli were negative orweakly positive, indicating very low levels of expression(Fig. 3F). Similarly, ISH for c-erbB-4 mRNA showedintense signals restricted to the developing tubules in thefetal kidneys (Fig. 4A and 4B) and moderate signals inthe tubules of the adult kidneys.
The adult ovarian stroma was non-reactive and thesurface epithelium was weakly positive. The epithelial
? 1998 John Wiley & Sons, Ltd.
cells lining the fallopian tube and the uterus weremoderately immunoreactive. The endometrial glands inthe early proliferative phase were weakly reactive ornegative for c-erbB-4 protein expression but towards thelate proliferative phase, patchy glandular reactivity wasseen and in the secretory phase there was uniform andstrong immunoreactivity. The endometrial stroma wasmoderately positive throughout the cycle. In situhybridization for c-erbB-4 mRNA showed low levels inthe uterine surface epithelium and patchy signals in the
Table I—Immunoreactivity of type I growth factor receptors
Organ/tissue EGFR* c-erbB-2† c-erbB-3‡c-erbB-4
adultc-erbB-4
fetal
Digestive tractOesophagus + + + + + + + NDStomach " + + + + + NDIntestines " + + + + + +Liver + + " + + + + + + +Pancreas, ducts + + + " + + + +Pancreas, acini + + " " + + +Salivary gland, ducts + + + + + + + ND
Urinary tractKidney, glomeruli " + " " "
prox. tubules + + " + + + +distal tubules + + + + + + + + + +
Transitional epithelium + + + + + + +
Respiratory tractBronchi/alveoli + +/ND + +/" + +/+ + + +/+ + +/+ +
Circulatory systemHeart ND " + + + + + +Vascular endothelium + " + + +
SkinEpidermis/adnexae + +/+ + + + +/+ + +/+ + + +/+ + + +
Endocrine systemPituitary ND ND ND + + + +Thyroid " " + +/" NDParathyroid ND " +/+ + +/+ + NDAdrenal cortex/medulla "/" "/" + + +/+ + +/+ + +/+Pancreatic islets " " + + + + +
Nervous systemBrain " " + + + + + + +Peripheral nerve "/+ ND +/+ + " "
Reproductive systemTestis " " + + + + + +Prostate + + +/+ + + + + NDBreast + + + + + + NDUterus (endometrium) + + + + + + + + NDOvary " + + + + + +
MiscellaneousSkeletal muscle ND ND ND +/+ + + +Lymphoid/haemopoietic " " " + (selectively) NDPlacenta + + " + + + + +
Tissues were scored on a semiquantitative scale based on the intensity of staining: negative ("); weakly positive (+);moderately positive (+ +); strongly positive (+ + +); not determined (ND).
EGFR, c-erbB-2, and c-erbB-3 data as reported in the literature and compared with the present study on c-erbB-4.*EGFR immunoreactivity as reported by Gusterson et al.29 and Damjanov et al.28
†c-erbB-2 immunoreactivity as reported by Press et al.30
‡c-erbB-3 immunoreactivity as reported by Prigent et al.25
, . 185: 236–245 (1998)

241c-erbB-4 EXPRESSION IN NORMAL TISSUES AND TUMOURS
glands. The cervical squamous epithelium was moder-ately positive in its upper two-thirds for the c-erbB-4protein.
The adult testicular germ and interstitial Leydig cellswere weakly positive for c-erbB-4 expression, whereasthe developing gonads showed strong immunoreactivityin the germ cells. The prostatic epithelium and smoothmuscle showed moderate and weak immunoreactivity,respectively.
Breast—The ductal epithelial and myoepithelial cellsshowed moderate positivity for c-erbB-4 protein (Fig.3C). Moderate levels of c-erbB-4 mRNA signals werepresent in the epithelium of the terminal ducts, althoughit was non-uniform.
Respiratory system—The linings of the bronchi andalveoli were moderately positive for c-erbB-4 protein in
? 1998 John Wiley & Sons, Ltd.
both adult and fetal tissues immunohistochemically. Thedeveloping fetal lungs showed moderate positivity onISH for c-erbB-4 mRNA.
Heart and circulatory system—The heart showed highlevels of c-erbB-4 protein and mRNA expressionthroughout fetal development (Fig. 4C and 4D). Mod-erate levels of c-erbB-4 protein were seen in the adultheart and vascular endothelium.
Skin—Both adult and fetal skin showed moderatelevels of c-erbB-4 protein in the epidermis and adnexae,with increasing intensity of staining towards the surface.Scattered signals were seen for c-erbB-4 mRNA by ISHin the fetal skin, but none was detected in the adult skin.
Endocrine system—Moderate immunoreactivity wasseen in the fetal and adult anterior pituitary, fetal
Fig. 4—c-erbB-4 mRNA expression determined by tissue in situ hybridization. (A, B) Fetal kidney; (C, D) fetal heart. A andC represent bright-field and B and D represent dark-field illuminated areas. (35S-labelled riboprobes and autoradiography withGiemsa counter-staining; original magnification #200)
, . 185: 236–245 (1998)

242 R. SRINIVASAN ET AL.
and adult adrenal cortex and medulla, and in theislets of Langerhans throughout development. Thefollicular cells of the thyroid were weakly reactiveand the chief cells of the parathyroid were moderatelyimmunoreactive for c-erbB-4 protein.
Nervous system—The adult cerebral and cerebellarcortex (Fig. 3G) showed weak immunoreactivity in thecell bodies of the neurones. In contrast, in the develop-ing fetal brain, moderate to strong immunoreactivitywas seen in the neurones (Fig. 3H) and ependyma forc-erbB-4 protein. The astrocytes were negative. TissueISH for c-erbB-4 mRNA revealed no signals in the adultcerebellum or brain stem. The second trimester fetalbrain showed strong regional positivity in the neuronesfor c-erbB-4 mRNA. The parasympathetic gangliashowed a moderate level of c-erbB-4 protein expression,whereas the peripheral nerves were negative.
Musculo-skeletal system—The bones and cartilagewere negative for c-erbB-4 protein in adult and fetaltissues. The adult skeletal muscle showed a hetero-geneous pattern of immunoreactivity for c-erbB-4 pro-tein ranging from very weak to moderate levels, butc-erbB-4 mRNA was undetectable in ISH studies. In thedeveloping fetal muscle, there were low to moderatelevels of expression both of the c-erbB-4 protein and ofmRNA.
Other tissues—The haemopoietic and lymphoid cellswere negative, with the exception of plasma cells andoccasional macrophages which were moderately immu-noreactive. The cyto- and syncytio-trophoblast of theplacental villi showed moderate levels of c-erbB-4 pro-tein expression in the first- and second-trimester andweak expression in the third-trimester villi. The maternaldecidua showed moderate levels of expression.
There was broad agreement in the pattern of expres-sion of the c-erbB-4 protein and mRNA in the fetalkidneys, heart, brain, and skeletal muscle, and in theadult kidneys, breast, and uterus. The adult gastrointes-tinal tract, skin, skeletal muscle, and salivary glandsfailed to reveal signals on ISH, although the protein wasreadily detected. The adult heart and pancreas specimen
? 1998 John Wiley & Sons, Ltd.
available were not suitable for ISH, due to relativelypoor preservation of mRNA.
c-erbB-4 protein expression in cancers
We looked for the trend of c-erbB-4 protein expres-sion in common human tumours by studying tensamples of each type, which included invasive carcinomaof the breast, colon, endometrium, prostate, cervix,ovary, and lung; squamous cell cancers in the head andneck region; and astrocytomas. The results were com-pared with the corresponding normal tissue and aresummarized in Table II. Overexpression was never seenin squamous cell carcinomas and occurred in 10–20 percent of adenocarcinomas (Fig. 5A). The two out of tenastrocytomas which showed greater than normal levelswere cases of anaplastic astrocytoma (Fig. 5B). Lessthan normal levels of expression were seen in 40–80 percent of various malignancies, reaching up to 100 per centin squamous cell carcinomas of the head and neckregion. An example of a squamous cell carcinoma of theoesophagus and a poorly differentiated carcinoma of thecolon showing distinctly lower levels of c-erbB-4 proteinthan normal are illustrated in Fig. 5C and 5D. In 10–40per cent of the various cancers, the levels of expressionwere equal to those seen in the adjacent morphologicallynormal tissue.
DISCUSSION
The specificity of the mouse monoclonal antibodiesHFR-1 and HFR-2 raised against the c-erbB-4 cyto-plasmic domain was demonstrated biochemically(Fig. 2) as well as immunocytochemically by positiveimmunostaining of cytocentrifuge preparations of theNIH3T3/HER4 cell line and negative staining usingthe A-431 cell line known to express high levels ofEGFR.
The nature of c-erbB-4 immunostaining was predomi-nantly cytoplasmic in both the cytocentrifuge prep-arations and the formalin-fixed, paraffin-embeddedtissues. That this pattern of staining is not an artefactwas confirmed by staining frozen sections. We observed
Table II—Expression of c-erbB-4 protein in a sample of common human malignancies
Type of tumourLess than
normalEqual tonormal
Greater thannormal
Carcinoma breast 7 1 2Carcinoma colon 5 4 1Carcinoma prostate 7 1 2Carcinoma ovary 5 4 1Carcinoma endometrium 4 4 2Carcinoma bronchus 6 4 0Carcinoma cervix 6 4 0Head and neck squamous cell carcinomas 10 0 0Astrocytoma 0 8 2Total 90 50 (55%) 30 (33%) 10 (11%)
, . 185: 236–245 (1998)

243c-erbB-4 EXPRESSION IN NORMAL TISSUES AND TUMOURS
occasional staining of nuclei (Fig. 5E) and it is interest-ing to speculate whether this receptor undergoes trans-location to the nucleus in response to certain stimuli.Heregulin-beta, a ligand of c-erbB-4, has been reportedto undergo rapid nuclear translocation in certain breastcancer cell lines.31
The pattern of c-erbB-4 expression is somewhat simi-lar to that of c-erbB-325 and quite unlike c-erbB-230 orEGFR.28,29 It should be kept in mind, while makingsuch comparisons, that the conditions for the determi-nation of receptor levels are different from study tostudy, such as, for example, the use of frozen28,30 versusparaffin-embedded sections25,30 and immunofluor-escence28 versus immunoperoxidase techniques.25,30
Notably, the heart expressed high levels of c-erbB-4 butis reported to express c-erbB-3 only weakly25 andc-erbB-2 not at all.30 This observation is consistent with
? 1998 John Wiley & Sons, Ltd.
the studies on the c-erbB-4 knockout mice, which dieduring mid-embryogenesis due to aborted developmentof cardiac muscle.32 Similar cardiac lesions in theheregulin and c-erbB-2 knockouts indicate that thesetwo receptors and the heregulins are crucial to cardiacdevelopment.31 The adult nervous system expressedc-erbB-4 only weakly in contrast to c-erbB-3,25 whereasthe developing neurones showed high levels, which isalso consistent with observations in the c-erbB-4 knock-out mice, where axonal innervation of the developinghindbrain was affected.32 In the pancreatic acinar andductal cells, c-erbB-4 is expressed similarly to EGFR.28
The weak to moderate heterogeneous expression ofc-erbB-4 in skeletal muscle is also interesting in view ofits reported importance in the formation of the neuro-muscular synapse,18 along with c-erbB-2, c-erbB-3, andtheir ligands, the neuregulins.
Fig. 5—Expression of c-erbB-4 protein in solid tumour types. (A) Invasive breast carcinoma and (B) anaplastic astrocytomashowing overexpression; (C) squamous cell carcinoma of the oesophagus showing very low levels compared with the normalcontrol shown in the inset; (D) poorly differentiated colonic carcinoma showing loss of expression in comparison with anormal crypt. (Streptavidin–biotin immunoperoxidase; A, B, original magnification #400; C, D, original magnification#200)
, . 185: 236–245 (1998)

244 R. SRINIVASAN ET AL.
In general, c-erbB-4 was seen to be associated morewith the differentiated compartment of tissues, similar toobservations for c-erbB-3 and in contrast to EGFR28,29
and c-erbB-2,30 which are seen to be associated withproliferating areas. This was especially noticeable in thelining epithelia of the gut, oesophagus, and cervix, andthe epidermis of the skin, where the intensity ofimmunoreactivity for c-erbB-4 increased progressivelytowards the surface. Similarly, in the endometrial glandsthere were progressively increasing levels of expressionfrom the early proliferative to the secretory phase. Thisobservation could also be extended to the sample ofcancers, where, among the adenocarcinomas, there wasinvariably loss of c-erbB-4 expression in the poorlydifferentiated tumours. The notable exception to thiswas the gliomas, where the only two overexpressingtumours were cases of anaplastic astrocytomas, indicat-ing that this receptor may behave differently with respectto the nervous system, compared with epithelia.Whereas overexpression of growth factor receptors inhuman malignancies is well documented,3–8 under-expression of these receptors and their significance hasonly recently been recognized. As also demonstratedhere, a recent report has shown loss of c-erbB-4 expres-sion in prostatic cancers, compared with benign lesionsand normal prostatic tissue.20 However, in a study ofthyroid tumours, overexpression of c-erbB-4 was seen in64 per cent of cases19 and in another study on medul-loblastoma, a malignant tumour of childhood, thepresence of a combination of c-erbB-4 with c-erbB-2was indicative of tumour aggressiveness and poorprognosis.21
The analysis of mRNA expression supplemented ourobservations on c-erbB-4 protein expression. They werealso consistent, with a few exceptions, with the reporteddata on northern analysis of various tissues.1 In ourstudy, tissue in situ hybridization revealed high levels ofc-erbB-4 mRNA in the fetal heart and developing kid-ney tubules, regionally intense signals in the fetal brain,and intermediate levels in the developing lung, skin, andskeletal muscle. In the intestines, skeletal muscle, andskin, c-erbB-4 mRNA was not detectable by ISH, indi-cating relatively low levels of expression. Tissue in situhybridization, while providing more specific informationon mRNA location, is sometimes less sensitive thannorthern blotting or RT-PCR for detection of mRNA,such as when the mRNA is distributed widely at lowlevels rather than clustered in a few cells.
In conclusion, this study has for the first time definedthe pattern of c-erbB-4 growth factor receptor expres-sion in normal tissues. This is an initial step towardsunderstanding its physiological role. We have alsoshown its pattern of expression in a sample of commonhuman malignancies, which we hope will form a basisfor evaluating its role in tumour pathogenesis.
ACKNOWLEDGEMENTS
We wish to thank Nicola O’Reilly of the PeptideSynthesis Laboratory; Jane Steel of the Research Mono-clonal Antibody Service; Jan Longcroft, Rosemary
? 1998 John Wiley & Sons, Ltd.
Jeffery, and Len Rogers of the In Situ HybridizationUnit; and the Biological Resources Unit of the ImperialCancer Research Fund for their assistance. We aregrateful to Professor Thomas Krausz and ProfessorGordon W. H. Stamp, Histopathology Department,Imperial College School of Medicine, HammersmithHospital, London and Professor A. K. Banerjee andProfessor S. K. Gupta, Postgraduate Institute ofMedical Education and Research, India, for the archivaltissue blocks.
Radhika Srinivasan is supported by a CommonwealthUniversities Scholarship.
REFERENCES1. Plowman GD, Culouscou J-M, Whitney GS, et al.. Ligand-specific acti-
vation of HER4/p180erbB4, a fourth member of the epidermal growth factorreceptor family. Proc Natl Acad Sci USA 1993; 90: 1746–1750.
2. Zimonjic DB, Maurizio A, Miki T, Popescu NC, Kraus MH. Localizationof the human HER4/erbB-4 gene to chromosome 2. Oncogene 1995; 10:1235–1237.
3. Prigent SA, Lemoine NR. The type I (EGFR related) family of growthfactor receptors and their ligands. Prog Growth Factor Res 1992; 4: 1–24.
4. Gullick WJ. Prevalence of aberrant expression of the epidermal growthfactor receptor in human cancers. Br Med Bull 1991; 47: 87–98.
5. Rajkumar T, Goden CSR, Lemoine NR, Gullick WJ. Expression of thec-erbB-3 protein in gastrointestinal tumours determined by monoclonalantibody RTJ1. J Pathol 1993; 170: 271–278.
6. Quinn CM, Ostrowski JL, Lane SA, Loney DP, Teasdale J, Benson FA.c-erbB-3 protein expression in human breast cancer: comparison with othertumour variables and survival. Histopathology 1994; 25: 247–252.
7. Rajkumar T, Gullick WJ. The type-I growth factor receptors in humanbreast cancer. Breast Cancer Res Treat 1994; 29: 3–9.
8. Slamon DJ, Clark GM, Wong SG, Levin WJ, Ullrich A, McGuire WL.Human breast cancer: correlation of relapse and survival with amplificationof HER-2/neu oncogene. Science 1987; 235: 177–182.
9. Rajkumar T, Stamp GWH, Pandha HS, Waxman J, Gullick WJ. Expres-sion of the type 1 tyrosine kinase growth factor receptors EGF receptor,c-erbB2 and c-erbB3 in bladder cancer. J Pathol 1996; 179: 381–385.
10. Earp HS, Dawson TL, Li X, Yu H. Heterodimerization and functionalinteraction between EGF receptor family members: a new signalling para-digm with implications for breast cancer research. Breast Cancer Res Treat1995; 35: 115–132.
11. Tzahar E, Waterman H, Chen X, et al.. A hierarchical network ofinterreceptor interactions determines signal transduction by neu differen-tiation factor/neuregulin and epidermal growth factor. Mol Cell Biol 1996;16: 5276–5287.
12. Plowman GD, Green JM, Culouscou J-M, Carlton GW, Rothwell VM,Buckley S. Heregulin induces tyrosine phosphorylation of HER4/p180erbB4.Nature 1993; 366: 473–475.
13. Tzahar E, Levkowitz G, Karunagaran D, et al.. ErbB-3 and ErbB-4function as the respective low and high affinity receptors of all neudifferentiation factor/heregulin isoforms. J Biol Chem 1994; 269: 25 226–25 233.
14. Riese DJ II, Bermingham Y, van Raaij TM, Buckley S, Plowman GD,Stern DF. Betacellulin activates the epidermal growth factor receptor anderbB-4, and induces cellular response patterns distinct from those stimulatedby epidermal growth factor or neuregulin-beta. Oncogene 1996; 12: 345–353.
15. Elenius K, Paul S, Allison G, Sun J, Klagsbrun M. Activation of HER4 byheparin-binding EGF-like growth factor stimulates chemotaxis but notproliferation. EMBO J 1997; 16: 1268–1278.
16. Cohen BD, Green JM, Foy L, Fell HP. HER4-mediated biological andbiochemical properties in NIH3T3 cells. J Biol Chem 1996; 271: 4813–4818.
17. Sepp-Lorenzino L, Eberhad I, Ma Z, et al.. Signal transduction pathwaysinduced by heregulin in MDA-MB-453 breast cancer cells. Oncogene 1996;12: 1679–1687.
18. Zhu X, Lai C, Thomas S, Burden SJ. Neuregulin receptors, erbB3 anderbB4 are localized at neuromuscular synapses. EMBO J 1995; 14: 5842–5848.
19. Haugen DR, Akslen LA, Varhaug JE, Lillehaug JR. Expression of c-erbB-3and c-erbB-4 proteins in papillary thyroid carcinomas. Cancer Res 1996; 56:1184–1188.
20. Lyne JC, Melhem MF, Finley GG, et al.. Tissue expression of neudifferentiation factor/heregulin and its receptor complex in prostate cancerand its biologic effects on prostate cancer cells in vitro. Cancer J Sci Am1997; 3: 21–30.
21. Gilbertson RJ, Perry RH, Kelly PJ, Pearson ADJ, Lunee J. Prognosticsignificance of HER2 and HER4 coexpression in childhood medullo-blastoma. Cancer Res 1997; 57: 3272–3280.
, . 185: 236–245 (1998)

245c-erbB-4 EXPRESSION IN NORMAL TISSUES AND TUMOURS
22. Rajkumar T, Gullick WJ. The production of antibodies using syntheticpeptides as immunogens. In: Jankowski JA, Polak JM, eds. Clinical GeneAnalysis and Manipulation. Cambridge: Cambridge University Press, 1996;99–110
23. Waterfield MD, Mayes ELV, Stroobant P, et al.. A monoclonal antibody tothe human epidermal growth factor receptor. J Cell Biochem 1982; 20:149–161.
24. Gullick WJ, Downward J, Foulkes JG, Waterfield MD. Antibodies to theATP-binding site of the human epidermal growth factor (EGF) receptor asspecific inhibitors of EGF-stimulated protein-kinase activity. Eur J Biochem1986; 158: 245–253.
25. Prigent SA, Lemoine NR, Hughes CM, Plowman GD, Selden C, GullickWJ. Expression of the c-erbB-3 protein in normal human adult and foetaltissues. Oncogene 1992; 7: 1273–1278.
26. Senior PV, Critchley DR, Beck F, Walker RA, Varley JM. The localizationof laminin mRNA and protein in the post-implantation embryo andplacenta of the mouse: in situ hybridization and immunocytochemical study.Development 1988; 104: 431–446.
? 1998 John Wiley & Sons, Ltd.
27. Zandvliet DW, Hanby AM, Austin CA, et al.. Analysis of foetal expressionsites of human type II DNA topoisomerase alpha and beta mRNAs by insitu hybridization. Biochim Biophys Acta 1996; 1307: 239–247.
28. Damjanov I, Mildner B, Knowles BB. Immunohistochemical localization ofthe epidermal growth factor receptor in normal human tissues. Lab Invest1986; 55: 588–592.
29. Gusterson B, Cowley G, Smith JA, Ozanne B. Cellular localization ofhuman epidermal growth factor receptor. Cell Biol Int Rep 1984; 8: 649–658.
30. Press MF, Cordon-Cardo C, Slamon DJ. Expression of the HER-2/neuproto-oncogene in normal human adult and foetal tissues. Oncogene 1990;5: 953–962.
31. Li W, Park JW, Nuijens A, Sliwkowski MX, Keller GA. Heregulin israpidly translocated to the nucleus and its transport is correlated with c-mycinduction in breast cancer cells. Oncogene 1996; 12: 2473–2477.
32. Gassmann M, Casagranda F, Orioli D, et al.. Aberrant neural and cardiacdevelopment in mice lacking the ErbB4 neuregulin receptor. Nature 1995;378: 390–394.
, . 185: 236–245 (1998)